
Geographic variation in resin canal defenses in seedlings from the Sitka spruce × white spruce...
11
Geographic variation in resin canal defenses in seedlings from the Sitka spruce × white spruce introgression zone G.A. O’Neill, S.N. Aitken, J.N. King, and R.I. Alfaro Abstract: Seedlings from 18 provenances along a coast–interior transect in the Picea sitchensis (Bong.) Carrière × Picea glauca (Moench) Voss introgression zone in northwestern British Columbia were mechanically wounded at the beginning of their third growing season to simulate natural attack by the white pine weevil, Pissodes strobi (Peck). Constitutive resin canals (CRC) in the cortex and traumatic resin canals (TRC) in the xylem of terminal shoots were characterized microscopically 4 months after wounding. Wounding resulted in a large increase in CRC size and in TRC number and density. Provenances differed significantly in TRC number and in CRC number, size, total area, and the proportion of total bark area occupied by CRC. CRC number and size, TRC number, and provenance weevil resistance (obtained from previously published data) increased with increasing latitude, elevation, and distance from the Pacific Ocean (i.e., towards the P. glauca end of the introgression zone) and decreased with increasing longitude (i.e., towards P. sitchensis). These traits also increased with aridity and continentality and decreased with most temperature, precipita- tion, and growing season length variables. Statistically significant multiple regression models related variation in some resin canal traits to geographic (r 2 = 0.71) and climatic (r 2 = 0.62) variables. Provenance mean values for weevil resis- tance were positively associated with predicted values for TRC number and CRC size. These results indicate that it is possible to predict locations in the introgression zone containing trees that possess desirable resin canal traits using geographic or climatic variables. Résumé : Des semis de 18 provenances récoltées le long d’un transect allant de la région côtière vers l’intérieur des terres, dans la zone d’introgression de Picea sitchensis (Bong.) Carrière × Picea glauca (Moench) Voss située dans le nord-ouest de la Colombie-Britannique, ont été blessés mécaniquement au début de la troisième saison de croissance pour simuler une attaque par le charançon du pin blanc, Pissodes strobi (Peck). Quatre mois plus tard, les caractéristi- ques microscopiques des canaux résinifères constitutifs (CRC) présents dans le cortex et les canaux résinifères d’origine traumatique (CRT) formés dans le xylème des pousses terminales ont été examinées. La dimension des CRC ainsi que le nombre et la densité des CRT ont beaucoup augmenté suite aux blessures. Le nombre de CRT ainsi que le nombre, la dimension, la surface totale et la proportion de la surface totale d’écorce occupée par les CRC diffèrent si- gnificativement selon les provenances. Le nombre et la dimension des CRC, le nombre de CRT et la résistance des provenances au charançon (obtenue à partir de données déjà publiées) augmentent avec la latitude, l’altitude et la dis- tance de l’océan Pacifique (i.e., vers le côté de P. glauca dans la zone d’introgression) et diminuent avec l’augmentation de la longitude (i.e., vers P. sitchensis). Ces caractères augmentent également avec le degré d’aridité et de continentalité mais diminuent avec la plupart des variables reliées à la température, à la précipitation et à la lon- gueur de la saison de croissance. Des équations de régression multiple statistiquement significatives relient la variation de certains caractères des canaux résinifères aux variables géographiques (r 2 = 0,71) et climatiques (r 2 = 0,62). Les va- leurs moyennes de résistance au charançon des provenances sont positivement reliées aux valeurs prédites du nombre de CRT et de la dimension des CRC. Ces résultats indiquent qu’il est possible de prédire à l’aide de variables géogra- phiques ou climatiques les endroits dans la zone d’introgression où sont situés les arbres dont les canaux résinifères possèdent les caractéristiques souhaitées. [Traduit par la Rédaction] O’Neill et al. 400 Can. J. For. Res. 32: 390–400 (2002) DOI: 10.1139/X01-206 © 2002 NRC Canada 390 Received 6 April 2001. Accepted 12 October 2001. Published on the NRC Research Press Web site at http://cjfr.nrc.ca on 16 February 2002. G.A. O’Neill 1,2 and S.N. Aitken. Department of Forest Science, Faculty of Forestry, University of British Columbia, 3041-2424 Main Mall, Vancouver, BC V6T 1Z4, Canada. J.N. King. British Columbia Ministry of Forests, P.O. Box 9519, Stn. Prov. Govt., Victoria, BC V8W 9C2, Canada. R.I. Alfaro. Pacific Forestry Centre, Canadian Forest Service, 506 West Burnside Road, Victoria, BC V8Z 1M5, Canada. 1 Corresponding author. 2 Present address: Kalamalka Forestry Centre, Research Branch, B.C. Ministry of Forests, 3401 Reservoir Road, Vernon, BC V1B 2C7, Canada (e-mail: [email protected]).
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Geographic variation in resin canal defenses in seedlings from the Sitka spruce × white spruce...

Geographic variation in resin canal defenses inseedlings from the Sitka spruce × white spruceintrogression zone
G.A. O’Neill, S.N. Aitken, J.N. King, and R.I. Alfaro
Abstract: Seedlings from 18 provenances along a coast–interior transect in thePicea sitchensis(Bong.) Carrière ×Picea glauca(Moench) Voss introgression zone in northwestern British Columbia were mechanically wounded at thebeginning of their third growing season to simulate natural attack by the white pine weevil,Pissodes strobi(Peck).Constitutive resin canals (CRC) in the cortex and traumatic resin canals (TRC) in the xylem of terminal shoots werecharacterized microscopically 4 months after wounding. Wounding resulted in a large increase in CRC size and in TRCnumber and density. Provenances differed significantly in TRC number and in CRC number, size, total area, and theproportion of total bark area occupied by CRC. CRC number and size, TRC number, and provenance weevil resistance(obtained from previously published data) increased with increasing latitude, elevation, and distance from the PacificOcean (i.e., towards theP. glaucaend of the introgression zone) and decreased with increasing longitude (i.e., towardsP. sitchensis). These traits also increased with aridity and continentality and decreased with most temperature, precipita-tion, and growing season length variables. Statistically significant multiple regression models related variation in someresin canal traits to geographic (r2 = 0.71) and climatic (r2 = 0.62) variables. Provenance mean values for weevil resis-tance were positively associated with predicted values for TRC number and CRC size. These results indicate that it ispossible to predict locations in the introgression zone containing trees that possess desirable resin canal traits usinggeographic or climatic variables.
Résumé: Des semis de 18 provenances récoltées le long d’un transect allant de la région côtière vers l’intérieur desterres, dans la zone d’introgression dePicea sitchensis(Bong.) Carrière ×Picea glauca(Moench) Voss située dans lenord-ouest de la Colombie-Britannique, ont été blessés mécaniquement au début de la troisième saison de croissancepour simuler une attaque par le charançon du pin blanc,Pissodes strobi(Peck). Quatre mois plus tard, les caractéristi-ques microscopiques des canaux résinifères constitutifs (CRC) présents dans le cortex et les canaux résinifèresd’origine traumatique (CRT) formés dans le xylème des pousses terminales ont été examinées. La dimension des CRCainsi que le nombre et la densité des CRT ont beaucoup augmenté suite aux blessures. Le nombre de CRT ainsi que lenombre, la dimension, la surface totale et la proportion de la surface totale d’écorce occupée par les CRC diffèrent si-gnificativement selon les provenances. Le nombre et la dimension des CRC, le nombre de CRT et la résistance desprovenances au charançon (obtenue à partir de données déjà publiées) augmentent avec la latitude, l’altitude et la dis-tance de l’océan Pacifique (i.e., vers le côté deP. glaucadans la zone d’introgression) et diminuent avecl’augmentation de la longitude (i.e., versP. sitchensis). Ces caractères augmentent également avec le degré d’aridité etde continentalité mais diminuent avec la plupart des variables reliées à la température, à la précipitation et à la lon-gueur de la saison de croissance. Des équations de régression multiple statistiquement significatives relient la variationde certains caractères des canaux résinifères aux variables géographiques (r2 = 0,71) et climatiques (r2 = 0,62). Les va-leurs moyennes de résistance au charançon des provenances sont positivement reliées aux valeurs prédites du nombrede CRT et de la dimension des CRC. Ces résultats indiquent qu’il est possible de prédire à l’aide de variables géogra-phiques ou climatiques les endroits dans la zone d’introgression où sont situés les arbres dont les canaux résinifèrespossèdent les caractéristiques souhaitées.
[Traduit par la Rédaction] O’Neill et al. 400
Can. J. For. Res.32: 390–400 (2002) DOI: 10.1139/X01-206 © 2002 NRC Canada
390
Received 6 April 2001. Accepted 12 October 2001. Published on the NRC Research Press Web site at http://cjfr.nrc.ca on16 February 2002.
G.A. O’Neill 1,2 and S.N. Aitken. Department of Forest Science, Faculty of Forestry, University of British Columbia,3041-2424 Main Mall, Vancouver, BC V6T 1Z4, Canada.J.N. King. British Columbia Ministry of Forests, P.O. Box 9519, Stn. Prov. Govt., Victoria, BC V8W 9C2, Canada.R.I. Alfaro. Pacific Forestry Centre, Canadian Forest Service, 506 West Burnside Road, Victoria, BC V8Z 1M5, Canada.
1Corresponding author.2Present address: Kalamalka Forestry Centre, Research Branch, B.C. Ministry of Forests, 3401 Reservoir Road, Vernon,BC V1B 2C7, Canada (e-mail: [email protected]).
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:58:56 PM
Color profile: Generic CMYK printer profileComposite Default screen

Introduction
The growth rate of Sitka spruce (Picea sitchensis(Bong.)Carrière) in plantations is among the highest of all tree spe-cies in Canada and the Pacific Northwest U.S.A. (Mitchelland Grout 1995), and its wood quality commands one of thehighest log prices in the region (http://www.for.gov.bc.ca/revenue/timberp/amv.htm). It is also planted extensively inwestern Europe for timber production. However, on the Pa-cific coast of British Columbia, where Sitka spruce is nativeand widespread, it is planted on only about 2% of reforesta-tion sites because repeated annual infestation by the whitepine weevil (Pissodes strobi(Peck)) (= terminal weevil,Sitka spruce weevil) severely reduces its growth rate andwood quality in plantations. Other susceptible species in-clude Engelmann spruce (Picea engelmanniiParry exEngelm.), white spruce (Picea glauca (Moench) Voss),white × Engelmann and white × Sitka hybrids, eastern whitepine (Pinus strobusL.), jack pine (Pinus banksianaLamb.),and Norway spruce (Picea abies(L.) Karst.) (Humble et al.1994).
Adults feed and oviposit in the spring in the bark of theprevious year’s leader, immediately below the elongatingterminal bud. Damage is caused by larvae that mine down-ward, consuming the phloem and cambium, and girdling theleader. If the leader dies and apical dominance is lost,branches of the uppermost whorl turn vertically, resulting incrooks and forks in the stem (Alfaro and Ying 1990). Re-peated attacks may render trees unmerchantable for lumberproduction. In high-hazard areas of coastal British Colum-bia, attacks usually begin at about age 5 years, as weevilpopulations begin to increase in plantations, peaking at ages15–20 years. Attack rates decline to endemic levels at ages25–45 years (Mitchell et al. 1990; Alfaro 1994; Ying 1997).
Genetic variation in weevil resistance has been detected inboth Sitka (Alfaro and Ying 1990; Ying 1991) and whitespruce (King et al. 1997). Variation in resistance has alsobeen observed in natural hybrids of these two species (Ying1991, 1997), which in British Columbia, are found only inthe midreaches of the Nass and Skeena watersheds in therugged northwest of the province (Roche 1969). Both weevilresistance and cold hardiness are greater, but growth isslower, in white spruce than in Sitka spruce (Mitchell et al.1990; Ying 1997). White spruce grows in the interior of theprovince, where climates are continental and weevil damageis less severe. Silvicultural techniques, including overstoreyshading, terminal clipping, insecticides, and biological con-trols have not proved consistently effective and practical onhigh weevil-hazard sites. These findings have led to the initi-ation of a genetic improvement program for Sitka spruce asthe main approach for obtaining healthy Sitka spruce planta-tions in British Columbia (King 1994).
The durability of host resistance against rapidly evolvingpests is a major concern in plant breeding (Namkoong1994). Several putative weevil resistance mechanisms havebeen identified, including chemical attraction or deterency(Alfaro 1988; Alfaro et al. 1996; Tomlin and Borden 1996),leader morphology (Wilkinson 1983; Tomlin and Borden1994b) and growth rate (King et al. 1997), the physical andchemical properties of resin (van Buijtenen and Santamour
1972; Santamour and Zinkel 1977; Tomlin and Borden1996), the resin duct system (Wilkinson 1983; Alfaro 1995;Alfaro et al. 1996; Tomlin and Borden 1997b; Tomlin et al.1998), foliar chemistry (Tomlin et al. 1997), bud burstphenology (Hulme 1995; Alfaro et al. 2000), antibiosis(Sahota et al. 1998), and the ability to recover from weevilattack (i.e., tolerance) (Alfaro and Ying 1990). Resistance ofa provenance, family, or individual may result from the si-multaneous contributions of several mechanisms (Tomlinand Borden 1994a, 1997a), and the relative importance ofeach mechanism may vary among provenances (Tomlin andBorden 1996) and individuals (Tomlin and Borden 1997a,1997b). Consequently, if resistance mechanisms were betterunderstood and could be readily evaluated for a large num-ber of genotypes, several mechanisms could be included inselection programs for the development of durable resistance(Brooks and Borden 1992; Namkoong 1994; Tomlin andBorden 1997a).
Several approaches to screening trees for traits related tospecific resistance mechanisms are being investigated.Ideally, such traits should be identifiable at an early age,strongly genetically correlated with resistance in plantations,and highly heritable (i.e., under strong genetic control). Me-chanical wounding of seedlings in common garden tests toassess defense responses may provide earlier and more accu-rate estimates of resistance than assessing natural attackrates in field-progeny tests where attack is often spatiallyand temporally heterogeneous and cannot be accurately as-sessed until age 10 or 15 years (Alfaro and Ying 1990).
One potentially exploitable set of defenses involves theresin canal system, which plays an important role in resis-tance of many conifer genera to insects (Thomson and Sifton1926; Berryman 1988; Panda and Khush 1995). On attackby herbivores, resin from severed canals inundates egg cavi-ties and larval galleries, creating a physical and chemical de-terrent to invading organisms (Alfaro 1995). Resin canalsthat are present in the absence of injury are referred to as“constitutive” or “preformed”, whereas those that develop inresponse to wounding are termed “traumatic” or “induced”resin canals (Berryman 1988). Constitutive resin canals arisefrom the apical meristem and, therefore, are not expected toappear in response to trauma. They occur in two forms: large“inner” canals in the middle of the cortex ring, and smaller“outer” canals at the periphery of the cortex, in the ridgesformed by the sterigmata. Traumatic resin canals arise fromthe cambium shortly after attack, form a ring in the develop-ing xylem, and are usually about 1/10 the diameter of innerconstitutive resin canals.
Genetic variation in resin canal defenses and associationsbetween resin canal traits and weevil resistance have beenwell documented. For example, the number of weevil attacksin eastern white pine were significantly and positively corre-lated with the depth (distance from bark surface) of both in-side and outside resin ducts (Wilkinson 1983). Traumaticresin canals formed faster and more numerously in responseto mechanical wounding in weevil-resistant than in weevil-susceptible white spruce from central British Columbia(Tomlin et al. 1998). In Sitka spruce, the density of outerconstitutive resin canals and bark thickness (Tomlin andBorden 1994b), and the size of constitutive resin canals and
© 2002 NRC Canada
O’Neill et al. 391
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:58:57 PM
Color profile: Generic CMYK printer profileComposite Default screen

number of traumatic resin canals (Brescia 2000) weregreater in resistant than in susceptible provenances. Simi-larly, in interior spruce the numbers and densities of both in-ner and outer constitutive resin canals were greater inresistant than in susceptible families (Alfaro et al. 1997).However, genetic clines (i.e., geographic or climatic patternsof the distribution of genetic variation) for resin canal de-fenses have not been previously characterized. Geographicor climatic models of genetic clines for resin canal defenseswould facilitate the identification of areas likely to containtrees possessing strong weevil resistance.
Introgression zones are typically high in genetic variation(Whitham et al. 1999), so they may serve well in the searchfor fast-growing, weevil-resistant genotypes. Weevil-resistant spruce genotypes have been found in the Sitka–white spruce introgression zone in the Nass–Skeena water-sheds (Alfaro and Ying 1990; Ying 1991, 1997; Ying andEbata 1994). However, substantial variation in resistance ex-ists among relatively closely situated provenances (Ying1991). This has led to the need to take a more systematic,genecological approach to investigating genetic variation inresistance-related traits within this zone.
The primary goal of this study was to determine how resincanal traits previously shown to be related to weevil resis-tance vary across the introgression zone between Sitka andwhite spruce. Knowledge of these patterns of genetic varia-tion would aid in locating regions likely to possess weevil-resistant trees. We hypothesized that (i) significant variationexists among provenances in resin canal traits; (ii ) variationamong provenances in resin canal traits is related to varia-tion in geographic and climatic variables of the provenances;and (iii ) resin canal defenses are related to weevil resistance.
These hypotheses were investigated by examining the ef-fect of mechanical wounding on the resin canal system in 3-year-old spruce seedlings from 18 provenances in the Sitka–white spruce introgression zone in the Nass and Skeena rivervalleys. Weevil resistance data from a previous study (Ying1991) of similar provenances were combined with data fromthe present study to indirectly examine relationships be-tween resin canal and resistance traits.
Materials and methods
Materials and experimental designSeeds for the resin canal study were collected from 60
randomly selected open-pollinated trees (families) growingin naturally regenerated stands from 18 provenances in theNass and Skeena river valleys in northwestern British Co-lumbia (Fig. 1). The collection region was approximately175 × 175 km and spanned 960 m elevation. The mostcoastal provenances were located in regions of pure Sitkaspruce, and the most eastern provenances approached thepure white spruce region. Within each provenance, seedswere collected from dominant and codominant trees sepa-rated by at least 50 m.
Seeds were sown into containers in a greenhouse in thespring of 1997, grown for one growing season, then trans-ferred to a shelterhouse at the University of British Colum-bia in Vancouver, Canada. Containers were arranged in arandomized complete block design consisting of 4 blocks,each containing 60 rows (i.e., 60 families) × 4 columns (i.e.,
four trees in a family-row plot). Survival after two growingseasons was approximately 95%.
Weevil resistance for 13 provenances in the Nass andSkeena watersheds was assessed in an earlier study of anopen-pollinated provenance trial at Sayward, on the eastcoast of Vancouver Island, British Columbia (Ying 1991).Latitudinal and longitudinal ranges sampled by the prove-nance trial (Ying 1991) are similar to those of the 18 prove-nances sampled in the present resin canal study (Fig. 1),although the elevational range of the provenances in theresin canal study (20–980 m) is slightly wider than that ofthe provenance trial (0–660 m).
Wounding and sample preparationTraumatic response was elicited by mechanical wounding,
following a procedure developed for white spruce (Tomlin etal. 1998). Eighteen holes, 1 mm in diameter and 1–2 mmdeep, were drilled into the 1998 leaders with a portable elec-tric drill in the first week of May 1999 (beginning of thethird growing season), coinciding with the naturaloviposition period of the weevils. The holes, 0.5–1.0 cmapart, were made along three equally spaced vertical lines,with the top hole 5 cm below the elongating apical bud. Halfof the seedlings in each family were wounded (two seedlingsin each row-plot of each block), and the remaining seedlingsused as controls.
A 5-cm stem section spanning the wound sites of each1998 leader, or the comparable section in nonwounded seed-lings, was removed after bud set, 20 weeks after wounding,
© 2002 NRC Canada
392 Can. J. For. Res. Vol. 32, 2002
Alaska
Queen
Charlotte
Islands
# Smithers
#Vancouver
British
Columbia
Alberta
N
U.S.A.
Fig. 1. Map of British Columbia showing the location of studyarea.
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:58:58 PM
Color profile: Generic CMYK printer profileComposite Default screen

during the third week of September 1999. Samples werefixed in a formalin – acetic acid – alcohol solution(Johansen 1940) for 1 week, then stored in 70% alcohol.Cross sections 35µm in thickness were cut from the middleof the samples with a sliding microtome, stained for 20 s in0.01% aqueous safranin, and mounted permanently ontoglass slides.
MeasurementsThree traits describing the traumatic resin canal response
and five traits describing the constitutive resin canal systemwere measured or derived (Table 1). The number of trau-matic and constitutive resin canals (TRCn and CRCn) ofeach section were counted under a 4× dissecting microscope.An image-analysis system (Panasonic WV BP310 videocamera mounted on the microscope and connected toSigmaScan version 3 software) allowed precise distance andarea measurement of the remaining resin canal traits. TRCand CRC density (TRCd and CRCd) were expressed as thenumber of traumatic and constitutive resin canals per centi-metre of 1999 xylem circumference, the circumference de-termined from distance measurements from the pith to theouter edge of the 1999 xylem. The relative rapidity of the re-sponse of the TRC system to wounding (TRCrr) was ex-pressed as the relative position of the TRC ring within the1999 xylem ring (i.e., as a proportion of the distance acrossthe xylem ring). Trees that responded rapidly to mechanicalwounding had a TRC ring located closer to the inside of the1999 xylem ring, and, therefore, a smaller TRCrr value, thantrees that responded more slowly to wounding, assumingthat all seedlings initiated cambial growth at the same time(see Statistical analyses section below). Additional CRCtraits included mean size (CRCs) of an individual canal, esti-mated from the surface area of a randomly chosen sample ofthree constitutive resin canals; total area (CRCa), the prod-uct of CRCn and CRCs; and the proportion of bark area oc-cupied by constitutive resin canals (CRCb), where bark areawas determined from measurements of bark ring size. Alldistance measurements are the mean of two observationsmade in opposing quadrants, where possible.
Bud burst phenology, seedling height, and 1998 leader di-ameter were recorded at the time of wounding. Phenologywas recorded as the stage of bud burst on a 0–8 scale (Alfaroet al. 2000). In the provenance trial (Ying 1991, Appen-dix 3), the percentage of trees of each provenance attackedat least once (% attack) was measured or inferred in each
tree every 2–3 years to age 15 from the presence of one ormore stem defects (crooks and forks). Susceptibility (% at-tack) values were converted to resistance values in the pres-ent study, i.e., 100 – % attack = % attack free.
Statistical analysesPreliminary analyses showed negligible differences be-
tween wounded and nonwounded (control) treatments forheight and leader diameter at the time of wounding.Nonwounded trees had progressed significantly (p = 0.0066)further into the annual developmental cycle than woundedtrees at the time of wounding; however, differences weresmall (bud burst stage = 6.24 for nonwounded trees and 6.05for wounded trees). Therefore, bud burst stage was not usedas a covariate in subsequent analyses of variance. However,strong edge effects were observed in the blocks. That is,trees on the outer edges of each 60-row × 4-column block(i.e., trees in columns 1 and 4) were substantially smallerthan those in the two inner columns (i.e., trees in columns 2and 3). Consequently, “column” was included as a fixed fac-tor in all analyses of variance.
Preliminary analyses also showed that all interactions in-volving wounding (i.e., block × wounding, provenance ×wounding, family within provenance × wounding, andblock × family within provenance × wounding) were non-significant for all traits. Consequently, all interactions in-volving wounding were excluded from the model. As a finalsimplification, correlations of provenance means amongCRCs, CRCa, and CRCb approached unity (r ≥ 0.95).Therefore, only CRCs was included in the subsequent gen-ecological analyses.
Analyses of variance were performed on all traits usingthe following linear model:
[1] yjkln = µ + Wi + Bj + Pk + BPjk + F(P)l(k)
+ BF(P)jl (k) + Cm + en(jkl)
whereyjkln is the observation of seedlingn in family l withinprovenancek in block j; µ is the overall mean;Bj andPk arethe effects of blockj and provenancek, respectively;BPjk isthe effect of provenancek in block j; F(P)l(k) is the effect offamily l within provenancek; BF(P)jl (k) is the effect of fam-ily l within provenancek in block j; Cm is the effect of col-umn m; Wi is the effect of wounding treatmenti; anden(jkl) isthe residual, composed of the interactions of (i) seedlingswith provenances and blocks and (ii ) wounding treatment
© 2002 NRC Canada
O’Neill et al. 393
Abbreviation Units Description
TRCn Count Number of traumatic resin canals observed in second-year xylemTRCd Count/mm Density of traumatic resin canals, i.e., number per millimetre of 1999 ring circumferenceTRCrr Proportion Relative rapidity of traumatic resin canal response (location of TRC ring within 1999
xylem ring as a proportion of 1999 xylem ring width)CRCn Count Number of constitutive resin canals observed in barkCRCd Count/mm Density of constitutive resin canals, i.e., number per millimetre of 1999 ring circumferenceCRCs mm2 Mean size of a single constitutive resin canalCRCa mm2 Total area occupied by all constitutive resin canals (CRCs × CRCn)CRCb Proportion Proportion of bark ring area occupied by constitutive resin canals = CRCa/bark area
Table 1. Abbreviations, units of measure, and descriptions of traumatic and constitutive resin canal (TRC and CRC) traitsused in the study.
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:58:59 PM
Color profile: Generic CMYK printer profileComposite Default screen

and columns with all of the above effects. Column andwounding treatment were treated as fixed effects.
Type III sums of squares were calculated using the SASGLM procedure (SAS Institute Inc. 1996). Nonwoundedseedlings were omitted from analyses involving TRC traits,except in the estimation of the effect of the wounding treat-ment. Examination of residuals indicated that errors werenormally distributed for all traits, andF tests were con-ducted with the appropriate mean squares to assess the sig-nificance (p ≤ 0.05) of effects.
Variance component estimates were derived using the re-stricted maximum likelihood (REML) estimator in theMIXED procedure of SAS (SAS Institute Inc. 1996). Prove-nance and family coeffients of variation were calculated as100 times the ratio of the square root of the provenance andfamily variance components (respectively) to the mean ofthe provenance means.
To assess geographic patterns of genetic variation in resincanal traits and weevil resistance, Pearson’s correlation coef-ficients were estimated between provenance trait means andgeographic and climatic variables. Only those resin canaltraits that displayed significant provenance variation wereevaluated. Geographic variables included elevation (EL) inmetres; latitude in degree-decimals north minus 48.0 (LAT);longitude in degree-decimals west minus 110.0 (LONG);their second-order terms (EL2, LAT2, LONG2); drainage dis-tance (DD) in kilometres, i.e., the distance along valley bot-toms from the provenance to the coast or nearest inlet; andinteractions: NW (LONG × LAT), SW (LONG/LAT), LTEL(LAT × EL), LGEL (LONG × EL), NWEL (NW × EL),SWEL (SW × EL), NWLAT (NW × LAT), NWLONG(NW × LONG), SWLAT (LONG/LAT2), and SWLONG(SW × LONG).
For each provenance, seven climate variables were derivedfrom its geographic variables using equations provided inRehfeldt et al. (1999): mean annual temperature (MAT);mean temperature of the coldest month (MCMT); mean tem-perature of the warmest month (MWMT); mean annual pre-cipitation (MAP); mean summer precipitation (MSP); meannumber of frost-free days (MNFFD); and frost-free period,the number of days between the last spring frost and first fallfrost (FFP). From these seven variables, three additional cli-mate variables were derived: annual aridity index(ARID_ANN) = (MAT + 10)/(MAP/1000); summer aridityindex (ARID_SUM) = MWMT/(MSP/1000); and contin-entality (CONT) = MWMT – MCMT.
Geographic patterns of genetic variation in resin canaltraits and observed weevil resistance were also analyzedwith multiple regression. For each trait, geographic and cli-matic variables for the provenances were used as independ-ent variables in two separate multiple regressions (one withgeographic independent variables and one with climatic in-dependent variables). Firstly, all models containing three orless independent variables were identified (to avoid over-parameterization) and ranked by adjustedR2. Secondly, tominimize model error and bias, three to five candidate mod-els were selected from the models identified in the first step,based on plots of Mallow’s Cp statistic versus the number ofindependent variables for each model (Neter et al. 1996). Afinal model for each trait in each regression (geographic andclimatic) was selected from among the candidate models
based onR2, Cp, model significance, and significance ofmodel parameters. All final models were significant atp ≤0.05, and all parameters were significant atp ≤ 0.10.
Pearson’s correlation coefficients were estimated based onprovenance means for pairs of resin canal traits and betweenprovenance means for resin canal traits and resistance (% at-tack free). Resin canal trait values used in the latter correla-tions were predicted at each of the 13 provenances sampledin the resistance study (Ying 1991) using geographic vari-ables in the multiple regression equations developed in thepresent study (see Table 5A).
Results
Mechanical woundingThe wounding procedure was highly effective (p <
0.0001) in eliciting a clear traumatic response. Traumaticresin canals were frequent in almost all wounded seedlings(mean TRCn = 16.9/seedling) and absent in almost allnonwounded seedlings (mean TRCn = 0.2/seedling) (Ta-ble 2). The effect on the density of traumatic resin canals(TRCd) was equally strong: mean TRCd was 1.55/mm forwounded and 0.02/mm for nonwounded. A typical woundresponse is shown in Fig. 2. Only 11% (47 of 446) of thenonwounded seedlings produced≥1 TRC, whereas 97%(428 of 439) of the wounded seedlings produced≥1 TRC.
All CRC traits responded to wounding. CRCs and CRCawere larger, and CRCn, CRCd, and CRCb were smaller, inthe wounded than in the nonwounded treatment (Table 2).Although statistically significant, differences betweenwounded and nonwounded treatments were relatively smallfor CRCn and CRCb.
Genetic and geographic variation in resin canal traitsand weevil resistance
Significant variation among provenances was observed forTRCn and for all CRC traits except CRCd (Table 3). For ex-ample, the provenance with the strongest TRCn response(TRCn = 27.4/cm) formed almost three times as many trau-matic resin canals as the provenance with the weakest TRCnresponse (TRCn = 10.0/cm). Coefficients of variation amongprovenances were moderately large for TRCn, TRCd, andCRCb (CVp = 14.8, 11.5, and 9.3%, respectively).
Provenance mean resin canal traits and observed weevilresistance varied across the study area. In general, TRCn,CRCn, CRCs, and weevil resistance increased with geo-graphic variables latitude, elevation, and distance from thePacific Ocean and decreased with longitude. These traitsalso increased with climatic variables aridity andcontinentality and decreased with most temperature, precipi-tation, and growing season length variables (Table 4).
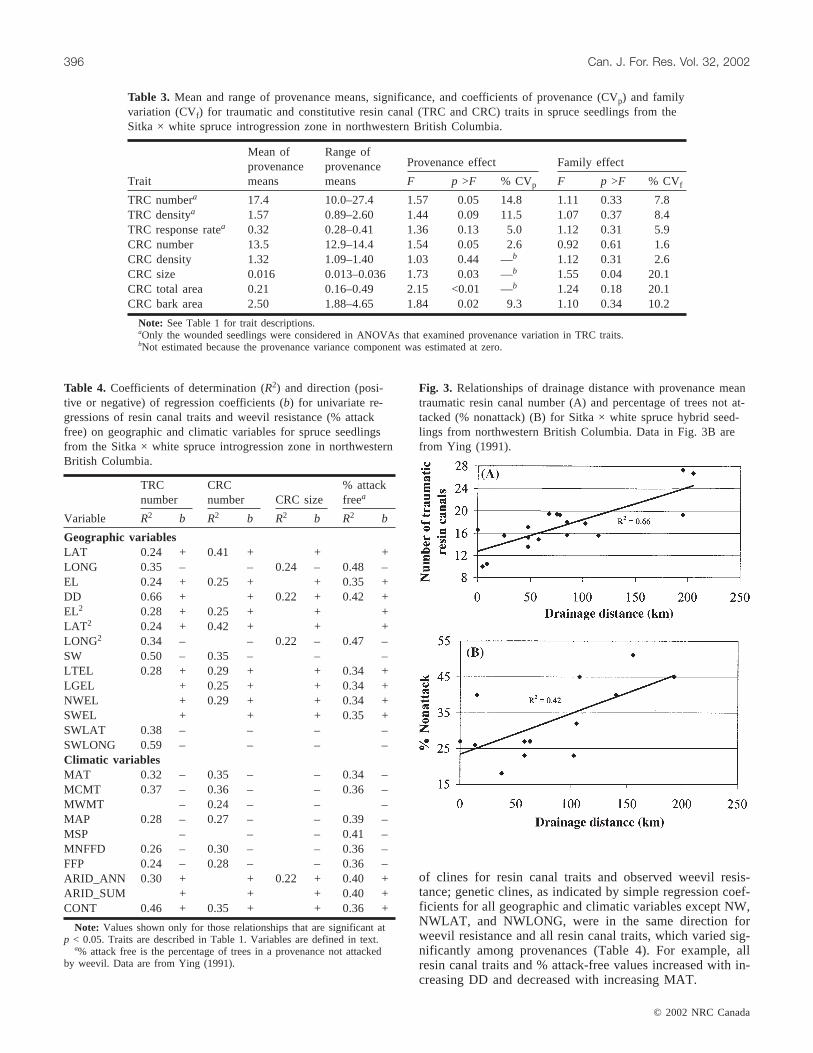
Simple regression also related significant proportions ofvariation among populations in TRC and CRC number andobserved weevil resistance to geographic and climatic vari-ables. For example, in the strongest relationship, 66% of thevariation among provenances in TRC number could be ac-counted for by drainage distance (DD) (Table 4, Fig. 3A).Longitude and drainage distance, accounted for 48 and 42%,respectively, of the variation in observed weevil resistance(% attack free) (Table 4, Fig. 3B). Not all relationships werelinear; the addition of the quadratic terms LAT2 and LONG2
© 2002 NRC Canada
394 Can. J. For. Res. Vol. 32, 2002
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:59:00 PM
Color profile: Generic CMYK printer profileComposite Default screen

accounted for significant improvement in the models de-scribing CRCn and CRCs, respectively (Table 5A). In the“best” multiple-regression model of resin canal traits, 71%of the variation in TRCn was accounted for by elevation andlongitude × elevation. Multiple regression models involvinggeographic variables accounted for more variation in resincanal traits than models involving climatic variables (Ta-ble 5).
Correlations among resin canal traitsProvenance mean correlations were strong and significant
(r ≥ 0.70) among TRC traits but highly variable among CRC
traits (Table 6). Provenance mean correlations between TRCand CRC traits, except those involving CRCd, were moder-ate and positive.
Correlations between resin canal traits and weevilresistance
Provenance mean correlations were moderately strong be-tween weevil resistance and predicted values of TRCn (r =0.61) and between weevil resistance and predicted values ofCRCs (r = 0.67) (Table 4). The association of geographicand climatic variables with resin canal traits and weevil re-sistance is also evident in the consistently similar orientation
© 2002 NRC Canada
O’Neill et al. 395
Mean
Trait Nonwounded Wounded F p >F
TRC number 0.22 16.91 149.3 0.0001TRC density (no./mm) 0.02 1.55 138.9 0.0001TRC response rate (proportion) —a 0.32 —a —a
CRC number 13.80 13.10 17.2 0.0001CRC density (no./mm) 1.43 1.22 97.6 0.0001CRC size (mm2) 0.013 0.018 50.2 0.0001CRC total area (mm2) 0.17 0.23 42.7 0.0001CRC bark area (proportion) 2.51 2.30 5.3 0.0213
Note: See Table 1 for trait descriptions.aValues were not calculated, because traumatic resin canals were too infrequent and randomly
located to identify the position of the TRC ring in individual nonwounded seedlings.
Table 2. Effect of mechanical wounding on traumatic and constitutive resin canal(TRC and CRC) traits in spruce seedlings from the Sitka × white spruceintrogression zone in northwestern British Columbia.
Fig. 2. Photomicrograph of a partial transverse section of a 1998 leader of a 3-year-old Sitka × white spruce seedling that was me-chanically wounded in spring 1999 and excised in fall 1999. Shown are the 1998 xylem ring (X98); a ring of traumatic resin canals(TRC); the 1999 xylem ring (X99); wound location (W); and a constitutive resin canal (CRC).
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:59:04 PM
Color profile: Generic CMYK printer profileComposite Default screen
of clines for resin canal traits and observed weevil resis-tance; genetic clines, as indicated by simple regression coef-ficients for all geographic and climatic variables except NW,NWLAT, and NWLONG, were in the same direction forweevil resistance and all resin canal traits, which varied sig-nificantly among provenances (Table 4). For example, allresin canal traits and % attack-free values increased with in-creasing DD and decreased with increasing MAT.
© 2002 NRC Canada
396 Can. J. For. Res. Vol. 32, 2002
Mean ofprovenancemeans
Range ofprovenancemeans
Provenance effect Family effect
Trait F p >F % CVp F p >F % CVf
TRC numbera 17.4 10.0–27.4 1.57 0.05 14.8 1.11 0.33 7.8TRC densitya 1.57 0.89–2.60 1.44 0.09 11.5 1.07 0.37 8.4TRC response ratea 0.32 0.28–0.41 1.36 0.13 5.0 1.12 0.31 5.9CRC number 13.5 12.9–14.4 1.54 0.05 2.6 0.92 0.61 1.6CRC density 1.32 1.09–1.40 1.03 0.44 —b 1.12 0.31 2.6CRC size 0.016 0.013–0.036 1.73 0.03 —b 1.55 0.04 20.1CRC total area 0.21 0.16–0.49 2.15 <0.01 —b 1.24 0.18 20.1CRC bark area 2.50 1.88–4.65 1.84 0.02 9.3 1.10 0.34 10.2
Note: See Table 1 for trait descriptions.aOnly the wounded seedlings were considered in ANOVAs that examined provenance variation in TRC traits.bNot estimated because the provenance variance component was estimated at zero.
Table 3. Mean and range of provenance means, significance, and coefficients of provenance (CVp) and familyvariation (CVf) for traumatic and constitutive resin canal (TRC and CRC) traits in spruce seedlings from theSitka × white spruce introgression zone in northwestern British Columbia.
TRCnumber
CRCnumber CRC size
% attackfreea
Variable R2 b R2 b R2 b R2 b
Geographic variablesLAT 0.24 + 0.41 + + +LONG 0.35 – – 0.24 – 0.48 –EL 0.24 + 0.25 + + 0.35 +DD 0.66 + + 0.22 + 0.42 +EL2 0.28 + 0.25 + + +LAT 2 0.24 + 0.42 + + +LONG2 0.34 – – 0.22 – 0.47 –SW 0.50 – 0.35 – – –LTEL 0.28 + 0.29 + + 0.34 +LGEL + 0.25 + + 0.34 +NWEL + 0.29 + + 0.34 +SWEL + + + 0.35 +SWLAT 0.38 – – – –SWLONG 0.59 – – – –Climatic variablesMAT 0.32 – 0.35 – – 0.34 –MCMT 0.37 – 0.36 – – 0.36 –MWMT – 0.24 – – –MAP 0.28 – 0.27 – – 0.39 –MSP – – – 0.41 –MNFFD 0.26 – 0.30 – – 0.36 –FFP 0.24 – 0.28 – – 0.36 –ARID_ANN 0.30 + + 0.22 + 0.40 +ARID_SUM + + + 0.40 +CONT 0.46 + 0.35 + + 0.36 +
Note: Values shown only for those relationships that are significant atp < 0.05. Traits are described in Table 1. Variables are defined in text.
a% attack free is the percentage of trees in a provenance not attackedby weevil. Data are from Ying (1991).
Table 4. Coefficients of determination (R2) and direction (posi-tive or negative) of regression coefficients (b) for univariate re-gressions of resin canal traits and weevil resistance (% attackfree) on geographic and climatic variables for spruce seedlingsfrom the Sitka × white spruce introgression zone in northwesternBritish Columbia.
Fig. 3. Relationships of drainage distance with provenance meantraumatic resin canal number (A) and percentage of trees not at-tacked (% nonattack) (B) for Sitka × white spruce hybrid seed-lings from northwestern British Columbia. Data in Fig. 3B arefrom Ying (1991).
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:59:07 PM
Color profile: Generic CMYK printer profileComposite Default screen
Discussion
Mechanical woundingThe strong TRC response to mechanical wounding ob-
served in this study corroborates previous studies in whichresinosis was observed following natural weevil attack(Alfaro 1995) and mechanical wounding (Tomlin et al. 1998;Brescia 2000). This finding suggests that assessment of trau-matic resin canal traits following mechanical woundingshould be suitable for studies of weevil resistance. The fewtraumatic resin canals observed in nonwounded seedlingsmay have arisen in response to inadvertent mechanical dam-age or attack by insects, including weevils during seedlinggrowth.
Mechanical wounding also affected the constitutive resincanal system. Constitutive resin canals were slightly less nu-merous, but substantially larger, in wounded seedlings thanin nonwounded seedlings, consistent with previous studiesof white spruce (Tomlin et al. 1998) and Sitka spruce(Brescia 2000). Whether the decrease in CRC number in thepresent study was a result of fusion of adjacent canals (Jou
1971), CRC occlusion (Overhaulser and Gara 1981; Tomlinet al. 1998) (observed in about 5% of constitutive resin ca-nals in the present study), or an unidentified cause is un-clear. The small reduction in CRC number, however, canaccount for only a fraction of the observed large increase inCRC size following wounding. Induced resinosis (Ruel et al.1998), possibly augmented by reactivation of quiescent epi-thelial secretory cells lining constitutive resin canals (Walteret al. 1989), may be responsible for the engorgement ofthese constitutive resin canals.
Less than 1% of the constitutive resin canals observed inthe present study were of the outer form (i.e., located in thesterigmatic ridges). The paucity of outer constitutive resincanals may be related to the young age of the seedlings, asouter resin canals were abundant in studies involving olderSitka spruce (Jou 1971) and white spruce (Alfaro et al.1997; Tomlin et al. 1998).
It has been speculated that high vapour pressure deficits,which occur during summer drought and which result in pro-longed periods of reduced oleoresin exudation pressure dur-ing the day, may reduce the ability of the constitutive resin
© 2002 NRC Canada
O’Neill et al. 397
(A) Geographic variables.
Trait and independentvariables
PartialR2
ModelR2 Intercept
Regressioncoefficient
TRC number 0.71 14.37EL 0.24 0.181LGEL 0.47 –0.009 349
CRC number 0.63 59.68LAT 2 0.42 1.100LAT 0.11 –14.36LGEL 0.10 0.000 034 8
CRC size 0.70 2.181LONG 0.24 –0.226LONG2 0.46 0.005 89
% attack freea 0.74 1096SWLONG 0.29 14.57SWLAT 0.18 2628SW 0.26 –1048
(B) Climatic variables.
TRC number 0.62 –95.77CONT 0.4632 4.202MAP 0.1534 0.016 2
CRC number 0.46 12.98MAT 0.3518 –0.808FFP 0.1106 0.031 2
CRC size 0.22 0.002 58ARID_ANN 0.2187 0.001 09
% attack freea 0.41 57.483 41MSP 0.41 –0.059 51
Note: All models are significant atp < 0.05. All independent variables are significantat p < 0.10. Geographic and climatic variables are defined in the text.
a% attack free is the percentage of trees in a provenance that were not attacked byweevils. Data are from Ying (1991).
Table 5. Multiple regressions of (A) geographic variables and (B) climaticvariables on traumatic and constitutive resin canal (TRC and CRC) traits for18 provenances and on weevil resistance (% attack free) values for 13 prove-nances in the Sitka × white spruce introgression zone in northwestern BritishColumbia.
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:59:07 PM
Color profile: Generic CMYK printer profileComposite Default screen
canal system to defend against weevil attack (Warkentin etal. 1992). However, summer aridity and weevil resistance(% attack free) were positively correlated in the presentstudy (Table 4), suggesting that genotypes adapted to high-aridity areas have a greater capacity for weevil resistance ina common environment than genotypes from low aridity ar-eas. In the present study, high-aridity areas are predomi-nantly inland, where summer heat sums required to completethe weevil life cycle are lower than in coastal areas(McMullen 1976), and summer heat sum accumulation isgreater. Consequently, weevil pressures and natural selectionfor resistance to the weevil may be greater in high-aridity ar-eas, despite possible concomitant lower exudation pressure.
Tree-improvement implicationsBased on the magnitude of coefficients of variation, esti-
mated heritability, and the significance of provenance andfamily effects, it appears that genetic improvement throughartificial selection of provenances or families within prove-nances may be most feasible for TRCn and CRCs or for thetraits that are strongly related to CRCs (i.e., CRCa andCRCb). Lack of significant family variation may be relatedto the small mean number of open-pollinated families perprovenance (3.3).
Higher weevil attack rates in Sitka spruce and Sitka ×white spruce hybrids than in white spruce may be due to dif-ferences between the two species in resin canal defences. In-ner resin canal density in nonwounded saplings wasapproximately 2.20 (canals per millimetre of circumference)in white spruce from the central interior of British Columbia(Tomlin et al. 1998) and 1.43 in Sitka × white spruce hy-brids from the Nass–Skeena region in the present study.While comparisons of responses across studies are tenuous,traumatic defences also appear to be stronger in whitespruce, where the number of resin canals in a typical trau-matic resin canal ring of a wounded sapling (Tomlin et al.1998) greatly exceeds the mean of 16.9 traumatic resin ca-nals in a wounded Sitka × white spruce seedling in the pres-ent study.
The size of constitutive resin canals was more than twiceas large in the Smithers provenance than in the provenancewith the next largest CRCs. This observation may be relatedto the location of the Smithers provenance; it is the most in-land and also has the largest indices for summer and annualaridity and for continentality.
Strong natural selection for TRCn and CRCn is suggestedby the large proportion of provenance variation for thesetraits, which is explained by a few geographic or climaticvariables. However, the genetic clines observed could be duesimply to the introgression of white spruce with Sitkaspruce, with selection playing a minor role. Why pure Sitkaand white spruce differ so greatly in resin canal traits is aninteresting question. Historically, white spruce may havebeen subject to greater pest pressure generally or to strongeror more prolonged exposure toPissodes strobiin particular,because of the high incidence of even-aged, fire-regeneratedwhite spruce stands in the more continental interior regions,which favour weevil reproduction (Kimoto et al. 2000). Theintrogression zone may have served as a hybrid bridge(Pilson 1999) to facilitate the spread ofPissodes strobiwest-ward from white spruce to Sitka spruce in the relatively re-cent past.
If TRCn or CRCn are ultimately chosen for indirect artifi-cial selection of weevil resistance, multiple-regression equa-tions, such as those calculated in the present study, shouldfacilitate the identification of provenances likely to possessthe desired resin canal traits. The stronger predictive powerof geographic-based models versus climate-based models inthis study may be related to weaknesses in the climate equa-tions in northern latitudes because of the paucity of climatestations in northern British Columbia (Rehfeldt et al. 1999).
In general, weevil resistance, TRCn and CRCn, all in-crease with distance inland (i.e., drainage distance), as doesthe proportion of the genome represented by white spruce(Yeh and Arnott 1986). However, strong population differen-tiation within Sitka spruce has been reported (Ying 1997),and caution is recommended for seed transfer, even forshort-distance transfers at a constant elevation for popula-
© 2002 NRC Canada
398 Can. J. For. Res. Vol. 32, 2002
TRCnumber
TRCdensity
TRCresponserate
CRCnumber
CRCdensity
CRCsize
% attackfree
TRC number 0.96* 0.74* 0.31 –0.14 0.53* 0.61*a
TRC density 0.70* 0.26 0.06 0.39 —b
TRC response rate 0.54* 0.05 0.36 —b
CRC number 0.32 –0.06 0.41a
CRC density –0.70* —b
CRC size 0.67*a
Note: Resin canal traits are defined in Table 1. Only wounded trees considered in the estimation of correlationsinvolving TRC traits. Correlations with asterisks are significant atp < 0.05 (N = 18 for all correlations, except thoseinvolving % attack free, whereN = 13).
aPopulation mean resin canal trait values used in these correlations were predicted for the 13 provenances examinedin the resistance study of Ying (1991). Expected resin canal values were predicted using geographic parameters of the13 provenances in the multiple regression equations for resin canal traits in Table 5A.
bNot estimated because multiple regression equations for predicting these traits were not determined. See Statisticalanalysis section in text.
Table 6. Pearson’s correlation coefficients (r) for correlations between provenance means for pairs ofresin canal traits and between resin canal traits and weevil resistance (% attack free) for spruce seed-lings from the Sitka × white spruce introgression zone in northwestern British Columbia.
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:59:08 PM
Color profile: Generic CMYK printer profileComposite Default screen

tions near the Nass–Skeena region (Campbell et al. 1989).Consequently, identification of provenances for parent treeselection in a Sitka spruce breeding program should considerboth resistance and adaptation to the abiotic environment ofthe deployment site.
Correlations between resin canal traits and weevilresistance
While the provenance mean correlations between resin ca-nal traits (TRCn and CRCs) and weevil resistance do notconfirm a common genetic basis for these traits, they do sup-port earlier evidence of their genetic association (Wilkinson1983; Tomlin and Borden 1994b; Alfaro et al. 1997; Tomlinet al. 1998; Brescia 2000). However, any estimable geneticcorrelation may be more indicative of linkage disequilibriumthan of pleiotropy in this region. Linkage disequilibrium is acharacteristic of introgression zones, which results from mi-gration of the parental species into the zone or from reducedhybrid fitness (although hybrids are not always less fit thanparental genotypes) (Arnold 1997, Ch. 5). Therefore, geneticcorrelations between resin canal traits and resistance shouldbe estimated within breeding populations before final selec-tion of a resin canal trait is made for use in a selection indexfor indirect, early selection for weevil resistance.
Correlations among resin canal traitsThe hypothesis that trees have evolved defense strategies
involving primarily one or the other resin canal defense sys-tem (i.e., CRC or TRC) (Tomlin et al. 1998) would requirethe existence of negative correlations between TRC andCRC traits. However, induced and constitutive resin canaldefenses were unrelated at the individual phenotypic level inwhite spruce (Tomlin et al. 1998) and were positively corre-lated at the provenance mean level in the present study.
Positive provenance mean correlations were also foundbetween TRCrr and the two other TRC traits. If these corre-lations or the correlations between TRC and CRC traits arepartly the result of pleiotropy, genetic selection for one TRCtrait may indirectly improve other TRC and CRC traits, fur-thering the potential for genetic selection of resin canal traitsto provide weevil resistance in Sitka and Sitka × whitespruce hybrids.
Acknowledgements
Funding was provided by the B.C. Forest Genetics Coun-cil. Seedlings were grown by the late Yaw Bennuah. DaveBrescia, Lara van Akker, Sonya Budge, and Joanne Tuytelprovided technical advice and assistance. Jerry Maedel pre-pared Fig. 1.
References
Alfaro, R.I. 1988. Laboratory feeding and colonization of non-hostlodgepole pine by two populations ofPissodes strobi(Peck)(Coleoptera: Curculionidae). Can. Entomol.120: 167–173.
Alfaro, R.I. 1994. White pine weevil in British Columbia: biologyand damage.In Proceedings, The White Pine Weevil: Biology,Damage and Management, Jan. 1994, Richmond, B.C.Edited byR.I. Alfaro, G. Kiss, and R.G. Fraser. Canadian Forestry Serviceand B.C. Ministry of Forests, Victoria, B.C. pp. 7–22.
Alfaro, R.I. 1995. An induced defence reaction in white spruce toattack by the white pine weevil,Pissodes strobi. Can. J. For.Res.25: 1725–1730.
Alfaro, R.I., and Ying, C.C. 1990. Levels of Sitka spruce weevil,Pissodes strobi(Peck), damage among Sitka spruce provenancesand families near Sayward, British Columbia. Can. Entomol.122: 7–8.
Alfaro, R.I., Kiss, G.K., and Yanchuk, A. 1996. Variation in the in-duced resin response of white spruce,Picea glauca, to attack byPissodes strobi. Can. J. For. Res.26: 967–972.
Alfaro, R.I., He, F., Tomlin, E., and Kiss, G. 1997. White spruceresistance to white pine weevil related to bark resin canal den-sity. Can. J. Bot.75: 568–573.
Alfaro, R.I., Lewis, K.G., King, J.N., El-Kassaby, Y.A., Brown, G.,and Smith, L.D. 2000. Budburst phenology of Sitka spruce andits relationship to white pine weevil attack. For. Ecol. Manage.127: 19–29.
Arnold, M.L. 1997. Natural hybridization and evolution. OxfordUniversity Press, Oxford, U.K.
Berryman, A.A. 1988. Towards a unified theory of plant defense.In Mechanisms of woody plant defenses against insectsEditedby W.J. Mattson, J. Levieux, and C. Bernard-Dagan. Springer-Verlag, New York. pp. 39–56.
Brescia, D. 2000. Seasonal variation in constitutive and induceddefenses of spruce (Picea spp.) hosts of the white pine weevil(Pissodes strobiPeck). Department of Forest Sciences, Univer-sity of British Columbia, Vancouver, B.C.
Brooks, J.E., and Borden, J.H. 1992. Development of a resistanceindex for Sitka spruce against the white pine weevilPissodesstrobi Peck. Forestry Canada and B.C. Ministry of Forests, Vic-toria, B.C. For. Resour. Dev. Agree. Rep. 180. pp. 18.
Campbell, R.K., Pawuk, W.A., and Harris, A.S. 1989. Microgeo-graphic genetic variation of Sitka spruce in southeastern Alaska.Can. J. For. Res.19: 1004–1013.
Hulme, M. 1995. Resistance of translocated Sitka spruce to dam-age by Pissodes strobi(Coleoptera: Curculionidae) related totree phenology. Econ. Entomol.88: 1525–1530.
Humble, L.M., Humphreys, N., and Van Sickle, G.A. 1994. Distri-bution and hosts of the white pine weevil,Pissodes strobi(Peck),in Canada.In The Proceedings, The White Pine Weevil: Biology,Damage and Management, Jan. 1994, Richmond, B.C.Edited byR.I. Alfaro, G. Kiss, and R.G. Fraser. Canadian Forestry Serviceand B.C. Ministry of Forests, Victoria, B.C. pp. 68–75.
Johansen, D.A. 1940. Plant microtechnique. McGraw-Hill, NewYork.
Jou, S.-M. 1971. The resin canal system in Sitka spruce,Piceasitchensis(Bong.) Carr. M.Sc. thesis, Department of Forest Re-sources, University of Washington, Seattle, Wash.
Kimoto, T.T.M., Alfaro, R.I., and Borden, J.H. 2000. Past infesta-tions of the white pine weevil in naturally regenerated sprucestands. For. Chron.76: 627–632.
King, J.N. 1994. Delivering durable resistant Sitka spruce for plan-tations.In Proceedings, The White Pine Weevil: Biology, Dam-age and Management, Jan. 1994, Richmond, B.C.Edited byR.I.Alfaro, G. Kiss, and R.G. Fraser. Canadian Forestry Service andB.C. Ministry of Forests, Victoria, B.C. pp. 134–149.
King, J.N., Yanchuk, A.D., Kiss, G.K., and Alfaro, R.I. 1997. Ge-netic and phenotypic relationships between weevil (Pissodesstrobi) resistance and height growth in spruce populations ofBritish Columbia. Can. J. For. Res.27: 732–739.
McMullen, L.H. 1976. Effect of temperature on oviposition andbrood development of Pissodes strobi (Coleoptera:Curculionidae). Can. Entomol.108: 1167–1172.
© 2002 NRC Canada
O’Neill et al. 399
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:59:09 PM
Color profile: Generic CMYK printer profileComposite Default screen

© 2002 NRC Canada
400 Can. J. For. Res. Vol. 32, 2002
Mitchell, K.J., and Grout, S.E. 1995. User’s guide for producingmanaged stand yield tables with WinTIPSY, version 1.3 underMicrosoft Windows. Research Branch, B.C. Ministry of Forests,Victoria, B.C.
Mitchell, R.G., Wright, K.H., and Johnson, N.E. 1990. Damage bythe Sitka spruce weevil (Pissodes strobi) and growth patterns for10 spruce species and hybrids over 26 years in the PacificNorthwest. USDA For. Serv. Res. Pap. PNW-RP-434.
Namkoong, G. 1994. Breeding strategies of resistance.In Proceed-ings, The White Pine Weevil: Biology, Damage and Manage-ment, Jan. 1994, Richmond, B.C.Edited byR.I. Alfaro, G. Kiss,and R.G. Fraser. Forestry Canada and B.C. Ministry of Forests,Victoria, B.C. pp. 218–221.
Neter, J., Kutner, M.H., and Wasserman, W. 1996. Applied linearstatistical models. McGraw-Hill, Chicago, Ill.
Overhulser, D.L., and Gara, R.I. 1981. Occluded resin canals asso-ciated with egg cavities made by shoot infestingPissodes. For.Sci. 27: 297–298.
Panda, N., and Khush, G.A. 1995. Host plant resistance to insects.CAB International, Wallingford, U.K.
Pilson, D. 1999. Plant hybrid zones and insect host range expan-sion. Ecology,80: 407–415.
Rehfeldt, G.E., Ying, C.C., Spittlehouse, D.L., and Hamilton, D.A.,Jr. 1999. Genetic responses to climate inPinus contorta: nichebreadth, climate change, and reforestation. Ecol. Monogr.69:375–407.
Roche, L. 1969. A genecological study of the genusPicea in Brit-ish Columbia. New Phytol.68: 505–554.
Ruel, J.J., Ayres, M.P., and Lorio, P.L., Jr. 1998. Loblolly pine re-sponds to mechanical wounding with increased resin flow. Can.J. For. Res.28: 596–602.
Sahota, T.S., Manville, J.H., Peet, F.G., Ibaraki, A., and White, E.1998. Weevil physiology controls the feeding rates ofPissodesstrobi on Picea sitchensis. Can. Entomol.130: 305–314.
Santamour, F.S., and Zinkel, D.F. 1977. Resin acids, resin crystalli-zation and weeviling in Balkan × eastern white pine hybrids.Proc. Northeast. For. Tree Improv. Conf.25: 164–175.
SAS Institute Inc. 1996. SAS/STAT software: changes and en-hancements through release 6.11. SAS Institute Inc., Cary, N.C.
Thomson, R.B., and Sifton, H.B. 1926. Resin canals in the Cana-dian spruce (Picea canadensis(Mill.) B.S.P.)—an anatomicalstudy, especially in relation to traumatic effects and their bear-ing on phylogeny. Philos. Trans. R. Soc. London B,214: 111.
Tomlin, E.S., and Borden, J.H. 1994a. Development of a multi-component index for Sitka spruce resistant to the white pineweevil. In Proceedings, The White Pine Weevil: Biology, Dam-age and Management, Jan. 1994, Richmond, B.C.Edited byR.I.Alfaro, G. Kiss, and R.G. Fraser. Forestry Canada and B.C.Ministry of Forests, Victoria, B.C. pp. 117–133.
Tomlin, E.S., and Borden, J.H. 1994b. Relationship between leadermorphology and resistance or susceptibility of Sitka spruce tothe white pine weevil. Can. J. For. Res.24: 810–816.
Tomlin, E.S., and Borden, J.H. 1996. Feeding responses of thewhite pine weevil, Pissodes strobi (Peck) (Coleoptera:Curculionidae), in relation to host resistance in British Colum-bia. Can. Entomol.128: 539–549.
Tomlin, E.S., and Borden, J.H. 1997a. Multicomponent index forevaluating resistance by Sitka spruce to the white pine weevil(Coleoptera: Curculionidae). Econ. Entomol.90: 704–714.
Tomlin, E.S., and Borden, J.H. 1997b. Thin bark and high densityof outer resin ducts: interrelated resistance traits in Sitka spruceagainst the white pine weevil (Coleoptera: Curculionidae). J.Econ. Entomol.90: 235–239.
Tomlin, E.S., Borden, J.H., and Pierce, H.D.J. 1997. Relationshipbetween volatile foliar terpenes and resistance of Sitka spruce tothe white pine weevil. For. Sci.43: 501–508.
Tomlin, E.S., Alfaro, R.I., Borden, J.H., and He, F. 1998.Histological response of resistant and susceptible white spruce tosimulated white pine weevil damage. Tree Physiol.18: 21–28.
van Buijtenen, J.P., and Santamour, F.S. 1972. Resin crystalizationrelated to weevil resistance in white pine (Pinus strobus). Can.Entomol.104: 215–219.
Walter, J., Charon, J., Marpeau, A., and Launay, J. 1989. Effects ofwounding on the terpene content of twigs of maritime pine(Pinus pinaster Ait.). I. Changes in the concentration ofditerpene resin acids and ultrastructural modifications of theresin duct epithelial cells following injury. Trees,3: 210–219.
Warkentin, D.L., Overhaulser, D.L., Gara, R.I., and Hinckley, T.M.1992. Relationships between weather patterns, Sitka spruce(Picea sitchensis) stress, and possible tip weevil (Pissodesstrobi) infestation levels. Can. J. For. Res.22: 667–673.
Whitham, T.G., Martinsen, G.D., Floate, K.D., Dungey, H.S.,Potts, B.M., and Keim, P. 1999. Plant hybrid zones affectbiodiversity: tools for a genetic-based understanding of commu-nity structure. Ecology,80: 416–428.
Wilkinson, R.C. 1983. Leader and growth characteristics of easternwhite pine associated with white pine weevil attack susceptibil-ity. Can. J. For. Res.13: 78–84.
Yeh, F.C., and Arnott, J.T. 1986. Electrophoretic and morphologi-cal differentiation ofPicea sitchensis, Picea glauca, and theirhybrids. Can. J. For. Res.16: 791–798.
Ying, C.C. 1991. Genetic resistance to the white pine weevil in Sitkaspruce. B.C. Ministry of Forests, Victoria, B.C. Res. Note 106.
Ying, C.C. 1997. Effects of site, provenance, and provenance andsite interaction in Sitka spruce in coastal British Columbia. For.Genet.4: 99–112.
Ying, C.C., and Ebata, T. 1994. Provenance variation in weevil at-tack in Sitka spruce.In Proceedings, The White Pine Weevil:Biology, Damage and Management, Jan. 1994, Richmond, B.C.Edited byR.I. Alfaro, G. Kiss, and R.G. Fraser. Canadian For-estry Service and B.C. Ministry of Forests, Victoria, B.C.pp. 98–109.
I:\cjfr\cjfr32\cjfr-03\X01-206.vpTuesday, February 12, 2002 4:59:10 PM
Color profile: Generic CMYK printer profileComposite Default screen
![genetic variation of norway spruce (picea abies [l.] karst ...](https://static.fdokumen.com/doc/165x107/633d6582b988f9753c06267d/genetic-variation-of-norway-spruce-picea-abies-l-karst-.jpg)



















