
Inheritance of chloroplast and mitochondrial DNA in Picea and composition of hybrids from...
7
Theor Appl Genet (1991) 82:242 248 Springer-Verlag 1991 Inheritance of chloroplast and mitochondrial DNA in Picea and composition of hybrids from introgression zones B. C. S. Sutton 1,2,*, D.J. Flanagan 1, j.R. Gawley 1, C.H. Newton 1, D.T. Lester 2 and Y.A. EI-Kassaby 2,3 Forest Biotechnology Centre, British Columbia Research Corporation, 3650 Wesbrook Mall, Vancouver, B.C., Canada V6S 2L2 2 Forest Science Department, 2357 Main Mall, University of British Columbia, Vancouver, B.C., Canada V6T IW5 3 Saanich Forestry Centre, Canadian Pacific Forest Products Ltd, 8067 East Saanich Rd, RRI, Saanichton, B.C., Canada V0S IM0 Received December 19, 1990; Accepted January 8, 1991 Communicated by P. M.A. Tigerstedt Summary. The cloning of white spruce (Picea glauca) mitochondrial DNA homologous to the cytochrome ox- idase II and ATPasec~ genes of maize is described. These probes were used to define restriction fragment length polymorphisms which distinguish the white, Engelmann (P. engelmannii) and Sitka spruce (P. sitchensis) popula- tions that occur in British Columbia. Analysis of progeny from crosses between the species revealed that mitochon- drial DNA was maternally inherited in all cases (32 prog- eny from five independent crosses). The inheritance of chloroplast DNA was determined using a probe de- scribed previously; in this case, all progeny exhibited paternal inheritance (27 progeny from four crosses). Mi- tochondrial and chloroplast probes were used to test trees from zones of introgression between coastal (Sitka) and interior spruces (white and Engelmann). In most cases mitochondria and chloroplasts within individuals were contributed by different species. The data shows that there is a significant Sitka spruce component in trees east of the coastal watershed in British Columbia. Key words: Mitochondria - Chloroplast - Polymorphism - Hybrids - Picea Introduction In British Columbia (B.C.) introgression occurs between three spruce species, white (P. glauca), Sitka (P. sitchen- sis) and Engelmann spruce (P. engelmannii) (Daubenmire 1968; Roche 1969; Krajina et al. 1982). Two main groups of hybrids occur naturally: interior hybrids which com- prise mixes of white and Engelmann spruce and interior/ * To whom correspondence should be addressed coastal hybrids which comprise mixes of the interior spe- cies with Sitka spruce. Recent work has demonstrated that chloroplast DNA probes can be used to determine the composition of Sitka/interior hybrid seedlots (Sutton et al. 1991). This research also showed that, in some cases, seedlings derived from these seedlots contained chloroplast DNA fragments of both species groups. This result suggested that biparental inheritance may occur. A similar observation of heteroplasmy has been reported for Pinus monticola (White 1990). The objective of this study was to determine the inheritance of both chloro- plasts and mitochondria in defined crosses between the three spruce species which occur in B.C. and to apply this information to the analysis of trees in introgression zones. In general, organelles are inherited maternally in an- giosperms (Palmer 1987; Sederoff 1987); however, recent work with gymnosperms has revealed a different pattern of inheritance. All conifer genera examined to date have exhibited paternal inheritance of chloroplasts, including Pinus (Wagner et al. 1987; Neale and Sederoff 1989), Larix (Szmidt et al. 1987) and Pseudotsuga (Neale et al. 1986). Recently, mitochondria were shown to be inherit- ed maternally in Pinus and paternally in Sequoia (Neal et al. 1989). To date there has been no comprehensive study of organelle inheritance in Picea. One study was carried out to examine chloroplast inheritance in which one Sitka x white individual and one (Sitka x white) x white individual were examined by purification and re- striction digestion of chloroplast DNA. In these cases paternal inheritance was observed (Szmidt et al. 1988). The Nass Skeena Transition (Fig. 1) is an area of B.C. where introgression has been determined by previous studies (Daubenmire 1968). Plans for the future refor- estation of this region are to develop seed orchards from selected trees that are located adjacent to this area (PR,
Transcript of Inheritance of chloroplast and mitochondrial DNA in Picea and composition of hybrids from...

Theor Appl Genet (1991) 82:242 248
�9 Springer-Verlag 1991
Inheritance of chloroplast and mitochondrial DNA in Picea and composition of hybrids from introgression zones
B. C. S. Sutton 1,2,*, D.J. Flanagan 1, j .R. Gawley 1, C.H. Newton 1, D.T. Lester 2 and Y.A. EI-Kassaby 2,3
Forest Biotechnology Centre, British Columbia Research Corporation, 3650 Wesbrook Mall, Vancouver, B.C., Canada V6S 2L2 2 Forest Science Department, 2357 Main Mall, University of British Columbia, Vancouver, B.C., Canada V6T IW5 3 Saanich Forestry Centre, Canadian Pacific Forest Products Ltd, 8067 East Saanich Rd, RRI, Saanichton, B.C., Canada V0S IM0
Received December 19, 1990; Accepted January 8, 1991 Communicated by P. M.A. Tigerstedt
Summary. The cloning of white spruce (Picea glauca) mitochondrial DNA homologous to the cytochrome ox- idase II and ATPasec~ genes of maize is described. These probes were used to define restriction fragment length polymorphisms which distinguish the white, Engelmann (P. engelmannii) and Sitka spruce (P. sitchensis) popula- tions that occur in British Columbia. Analysis of progeny from crosses between the species revealed that mitochon- drial DNA was maternally inherited in all cases (32 prog- eny from five independent crosses). The inheritance of chloroplast DNA was determined using a probe de- scribed previously; in this case, all progeny exhibited paternal inheritance (27 progeny from four crosses). Mi- tochondrial and chloroplast probes were used to test trees from zones of introgression between coastal (Sitka) and interior spruces (white and Engelmann). In most cases mitochondria and chloroplasts within individuals were contributed by different species. The data shows that there is a significant Sitka spruce component in trees east of the coastal watershed in British Columbia.
Key words: Mitochondria - Chloroplast - Polymorphism - Hybrids - Picea
Introduction
In British Columbia (B.C.) introgression occurs between three spruce species, white (P. glauca), Sitka (P. sitchen- sis) and Engelmann spruce (P. engelmannii) (Daubenmire 1968; Roche 1969; Krajina et al. 1982). Two main groups of hybrids occur naturally: interior hybrids which com- prise mixes of white and Engelmann spruce and interior/
* To whom correspondence should be addressed
coastal hybrids which comprise mixes of the interior spe- cies with Sitka spruce. Recent work has demonstrated that chloroplast DNA probes can be used to determine the composition of Sitka/interior hybrid seedlots (Sutton et al. 1991). This research also showed that, in some cases, seedlings derived from these seedlots contained chloroplast DNA fragments of both species groups. This result suggested that biparental inheritance may occur. A similar observation of heteroplasmy has been reported for Pinus monticola (White 1990). The objective of this study was to determine the inheritance of both chloro- plasts and mitochondria in defined crosses between the three spruce species which occur in B.C. and to apply this information to the analysis of trees in introgression zones.
In general, organelles are inherited maternally in an- giosperms (Palmer 1987; Sederoff 1987); however, recent work with gymnosperms has revealed a different pattern of inheritance. All conifer genera examined to date have exhibited paternal inheritance of chloroplasts, including Pinus (Wagner et al. 1987; Neale and Sederoff 1989), Larix (Szmidt et al. 1987) and Pseudotsuga (Neale et al. 1986). Recently, mitochondria were shown to be inherit- ed maternally in Pinus and paternally in Sequoia (Neal et al. 1989). To date there has been no comprehensive study of organelle inheritance in Picea. One study was carried out to examine chloroplast inheritance in which one Sitka x white individual and one (Sitka x white) x white individual were examined by purification and re- striction digestion of chloroplast DNA. In these cases paternal inheritance was observed (Szmidt et al. 1988).
The Nass Skeena Transition (Fig. 1) is an area of B.C. where introgression has been determined by previous studies (Daubenmire 1968). Plans for the future refor- estation of this region are to develop seed orchards from selected trees that are located adjacent to this area (PR,

243
NST
Prince Rupert mlthers !Q~C ~ ~ ~J) iL~, R ~Prince George
iVo.nr
~Victoria
Fig. t. Map of British Columbia showing the locations of trees tested in this study. The populations used for the analysis of individual trees are shown as regions: EK East Kootenay, PG Prince George, PR Prince Rupert, Smithers selection unit, VI Vancouver Island, QC Queen Charlotte Islands. The Nass Skeena Transition (NST) is a seed planning zone defined on the basis of previous studies of spruce introgression. The line marked with dots indicates the divide between the eastern and western watersheds of the coastal mountains
Fig. 1). We chose to analyze a small number of these individuals to determine whether they were hybrids.
This type of research requires probes that can reliably distinguish the chloroplasts and mi tochondr ia of these species and do not exhibit intraspecies polymorphism. We repor t the use of the chloroplast probe previously characterized by Sutton et al. (1991) and the isolation of two homologous mi tochondr ia l probes that can be used to distinguish the three species. These probes have been used to demonstra te the inheritance of mi tochondr ia and chloroplasts in interspecific crosses and to analyze the individual selections described above.
Materials and methods
Plant material
For the identification and characterization of species-specific polymorphisms, trees of four spruce populations, representing three species, were used. These included three B. C. populations: white spruce from low to moderate elevations in the Prince George (PG) region, Engelmann spruce from high elevations in the East Kootenay region (EK) and Sitka spruce from low eleva- tions of Vancouver Island (VI) and the Queen Charlotte Islands (QC). In addition, white spruce, referred to as eastern white spruce (EW), from Ontario was analyzed. Trees from all four populations are referred to as 'type' trees that are considered representative of the pure species (not from areas of introgres- sion). For the analysis of naturally occurring hybrids, trees from a selection unit near Smithers B.C. were analyzed. This area is within the Prince Rupert (PR) forest region, and the tree num- bers are prefixed thus. The locations of these populations are shown in Fig. t.
Purification of DNA and recombinant DNA methods
Total DNA was prepared from needles by the method of Wagner et el. (1987) as described previously (Sutton et aI. 1991). Mito- chondrial DNA of white spruce was obtained from an embryo- genic culture of white spruce (Webb et al. 1989). The culture was ground in liquid nitrogen and subjected to differential centrifu- gation followed by purification of a mitochondrial fraction on 6% 22% self-generating Percoll gradients (Pay and Smith 1988). This method enabled us to obtain mitochondrial DNA with some contaminating plastid DNA and was adequate for the cloning of mitochondrial sequences.
Purified mitochondrial DNA from white spruce was digest- ed with BamHI and ligated to pEMBL8- that had been digested with BamHI and treated with calf intestinal phosphatase (Boehringer Mannheim) (Maniatis et al. 1982). Following trans- formation into E. coli strain DH5~ approximately 500 recombi- nants were screened with the ATPasec~ (pTA22; Braun and Lev- ings 1985) and cytochrome oxidase II (pZME- 1; Fox and Leaver 198 t) probes derived from maize mitochondria. Clones carrying homologous fragments were recovered, and the inserts from these plasmids were used in subsequent hybridizations to spruce DNA.
Electrophoresis was carried out in TAE buffer 0.8 % agarose gels for chloroplast polymorphisms or 0.4% agarose gels for mitochondrial polymorphisms (Maniatis et al. 1982). The gels were blotted to Hybond (Amersham) according to the manufac- turers' instructions. For the preparation of probes, plasmid in- serts were separated in 1% low-melting-point agarose, and re- striction fragments excised and labelled using the oligolabelling method (Feinberg and Vogelstein 1983, 1984). Prehybridization was carried out in 6XSSPE, 1% sodium lauryl sarcosine and 100 gg/ml sheared and heat-denatured calf thymus DNA. Hy- bridization was carried out in the same solution with the addi- tion of 3-10 x 10 6 cpm of 32p-dATP-labelled probe and 10% dextran sulfate.
Results
Isolation and characterization o f mitochondrial probes
Heterologous mi tochondr ia l probes from maize covering the ATPasec~ (pAT22) and cytochrome oxidase II ( pZME-I ) genes were tested in hybridizat ion to various individuals of each spruce species. Hybr id iza t ion was obtained with total DNA; however, the signal obtained was weak and, in the case of the ATPasec~ probe, exten- sive analysis showed that bands of chloroplast origin were detected in addi t ion to mi tochondr ia l bands (data not shown). Fo r these reasons we chose to clone the equivalent sequences from white spruce in order to in- crease the ease of routine screening experiments. Two BamHI fragments of 6.6 kb and 2.6 kb within spruce total D N A hybridized with the maize cytochrome oxi- dase I1 probe. Clones carrying both these fragments (three clones and one clone respectively) were recovered from the mi tochondr ia l library. The relative degree o f hybridizat ion is i l lustrated for the maize cytochrome ox- idase II probe and the homologous spruce probe in Fig. 2; the clone carrying the larger f ragment ( pWSml ) was used in subsequent analyses. Three BamHI frag- ments (7.8, 3.4 and 1.5 kb) hybridized with the maize
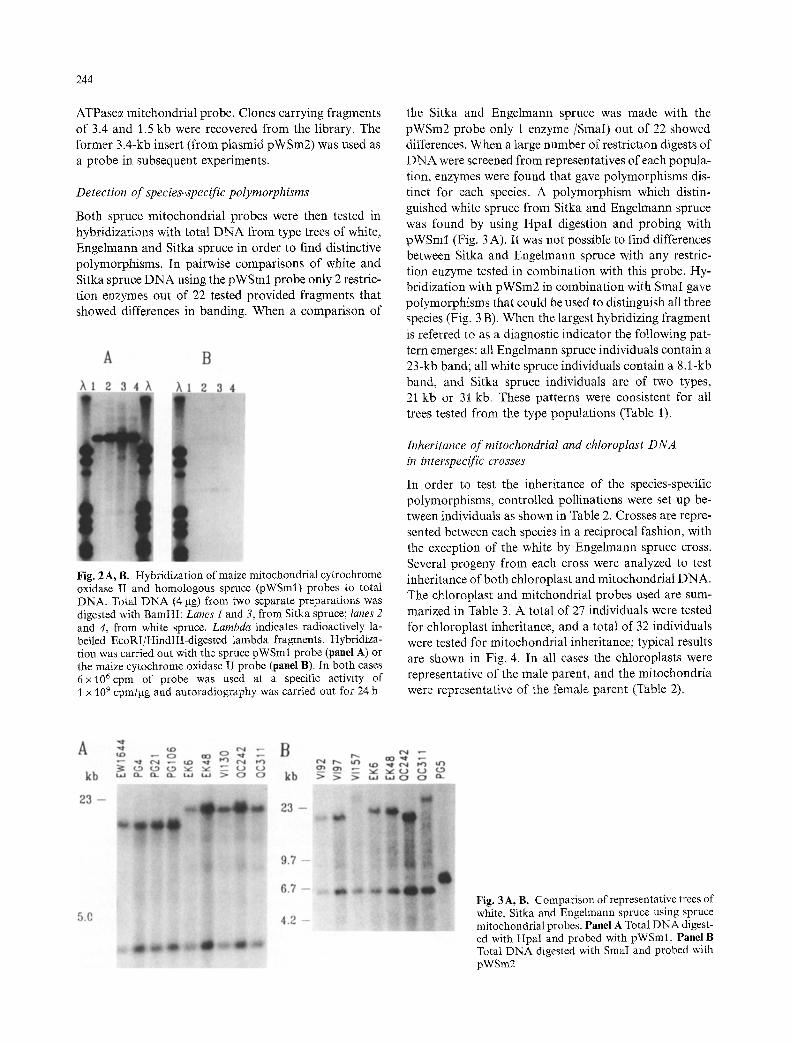
244
ATPasec~ mitchondrial probe. Clones carrying fragments of 3.4 and 1.5 kb were recovered from the library. The former 3.4-kb insert (from plasmid pWSm2) was used as a probe in subsequent experiments.
Detection of" species-specific polymorphisms
Both spruce mitochondrial probes were then tested in hybridizations with total D N A from type trees of white, Engelmann and Sitka spruce in order to find distinctive polymorphisms. In pairwise comparisons of white and Sitka spruce DNA using the pWSml probe only 2 restric- tion enzymes out of 22 tested provided fragments that showed differences in banding. When a comparison of
Fig. 2 A, B. Hybridization of maize mitochondrial cytrochrome oxidase II and homologous spruce (pWSml) probes to total DNA. Total DNA (4 gg) from two separate preparations was digested with BamHI: Lanes 1 and 3, from Sitka spruce; lanes 2 and 4, from white spruce. Lambda indicates radioactively la- belled EcoRI/HindlII-digested lambda fragments. Hybridiza- tion was carried out with the spruce pWSml probe (panel A) or the maize cytochrome oxidase II probe (panel B). In both cases 6x 106cpm of probe was used at a specific activity of 1 x 109 cpm/gg and autoradiography was carried oat for 24 h
the Sitka and Engelmann spruce was made with the pWSm2 probe only 1 enzyme/SmaI) out of 22 showed differences. When a large number of restriction digests of DNA were screened from representatives of each popula- tion, enzymes were found that gave polymorphisms dis- tinct for each species. A polymorphism which distin- guished white spruce from Sitka and Engelmann spruce was found by using HpaI digestion and probing with pWSm/(Fig . 3 A). It was not possible to find differences between Sitka and Engelmann spruce with any restric- tion enzyme tested in combination with this probe. Hy- bridization with pWSm2 in combination with SmaI gave polymorphisms that could be used to distinguish all three species (Fig. 3 B). When the largest hybridizing fragment is referred to as a diagnostic indicator the following pat- tern emerges: all Engelmann spruce individuals contain a 23-kb band; all white spruce individuals contain a 8.1-kb band, and Sitka spruce individuals are of two types, 21 kb or 31 kb. These patterns were consistent for all trees tested from the type populations (Table 1),
Inheritance o f mitochondrial and chloroplast D N A in interspecific crosses
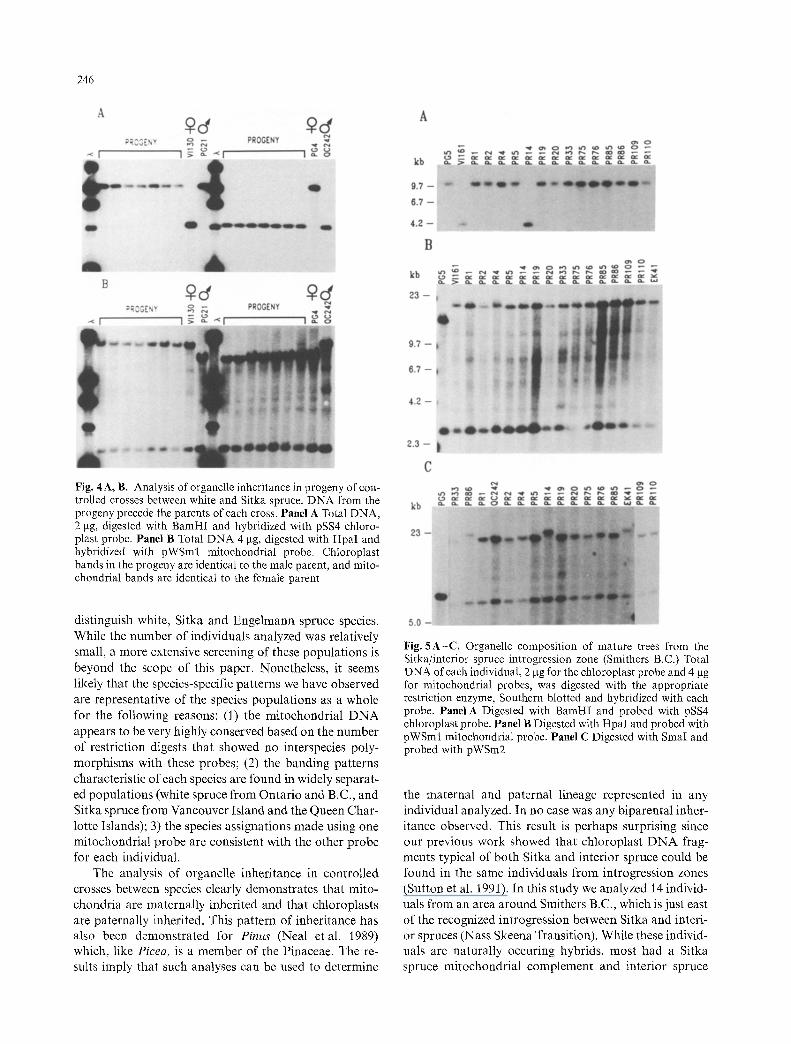
In order to test the inheritance of the species-specific polymorphisms, controlled pollinations were set up be- tween individuals as shown in Table 2. Crosses are repre- sented between each species in a reciprocal fashion, with the exception of the white by Engelmann spruce cross. Several progeny from each cross were analyzed to test inheritance of both chloroplast and mitochondrial DNA. The chloroplast and mitchondrial probes used are sum- marized in Table 3. A total of 27 individuals were tested for chloroplast inheritance, and a total of 32 individuals were tested for mitochondrial inheritance; typical results are shown in Fig. 4. In all cases the chloroplasts were representative of the male parent, and the mitochondria were representative of the female parent (Table 2).
Fig. 3A, B. Comparison of representative trees of white, Sitka and Engelmann spruce using spruce mitochondrial probes. Panel A Total DNA digest- ed with HpaI and probed with pWSml. Panel B Total DNA digested with SmaI and probed with pWSm2
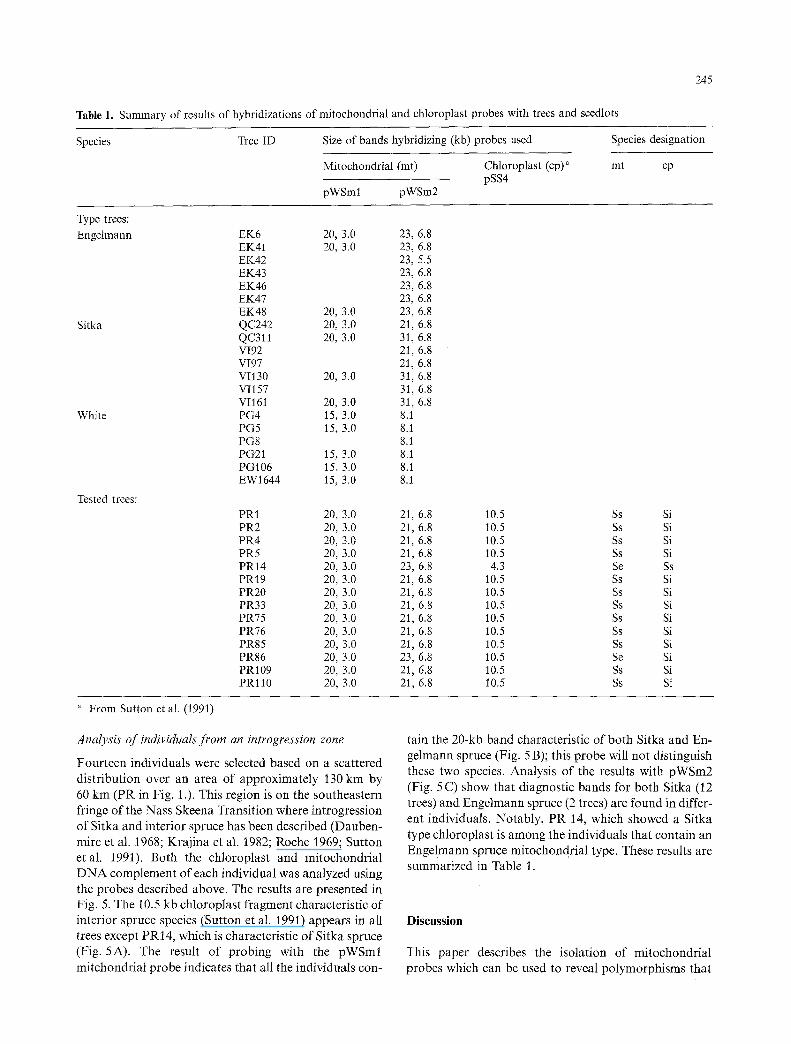
Table 1. Summary of results of hybridizations of mitochondrial and chloroplast probes with trees and seedlots
245
Species Tree ID Size of bands hybridizing (kb) probes used
Mitochondrial (mt) Chloroplast (cp)" pSS4
pWSml pWSm2
Species designation
mt cp
Type trees: Engelmann
Sitka
White
Tested trees:
EK6 20, 3.0 23, 6.8 EK41 20, 3.0 23, 6.8 EK42 23, 5.5 EK43 23, 6.8 EK46 23, 6.8 EK47 23, 6.8 EK48 20, 3.0 23, 6.8 QC242 20, 3.0 21, 6.8 QC311 20, 3.0 31, 6.8 vi92 21, 6.8 VI97 21, 6.8 VI130 20, 3.0 31, 6.8 VI157 31, 6.8 VI161 20, 3.0 31, 6.8 PG4 15, 3.0 8.1 PG5 15, 3.0 8.1 PG8 8.1 PG21 15, 3.0 8.1 PGI06 15, 3.0 8.1 EW1644 15, 3.0 8.1
PR1 20, 3.0 21, 6.8 10.5 Ss Si PR2 20, 3.0 21, 6.8 10.5 Ss Si PR4 20, 3.0 21, 6.8 10.5 Ss Si PR5 20, 3.0 21, 6.8 10.5 Ss Si PRt4 20, 3.0 23, 6.8 4.3 Se Ss PR19 20, 3.0 21, 6.8 10.5 Ss Si PR20 20, 3.0 21, 6.8 10.5 Ss Si PR33 20, 3.0 21, 6.8 10.5 Ss Si PR75 20, 3.0 21, 6.8 10.5 Ss Si PR76 20, 3.0 21, 6.8 10.5 Ss Si PR85 20, 3.0 21, 6.8 10.5 Ss Si PR86 20, 3.0 23, 6.8 10.5 Se Si PR109 20, 3.0 21, 6.8 10.5 Ss Si PRt10 20, 3.0 21, 6.8 10.5 Ss Si
a From Sutton et al. (1991)
Analysis of individuals fi'om an introgression zone
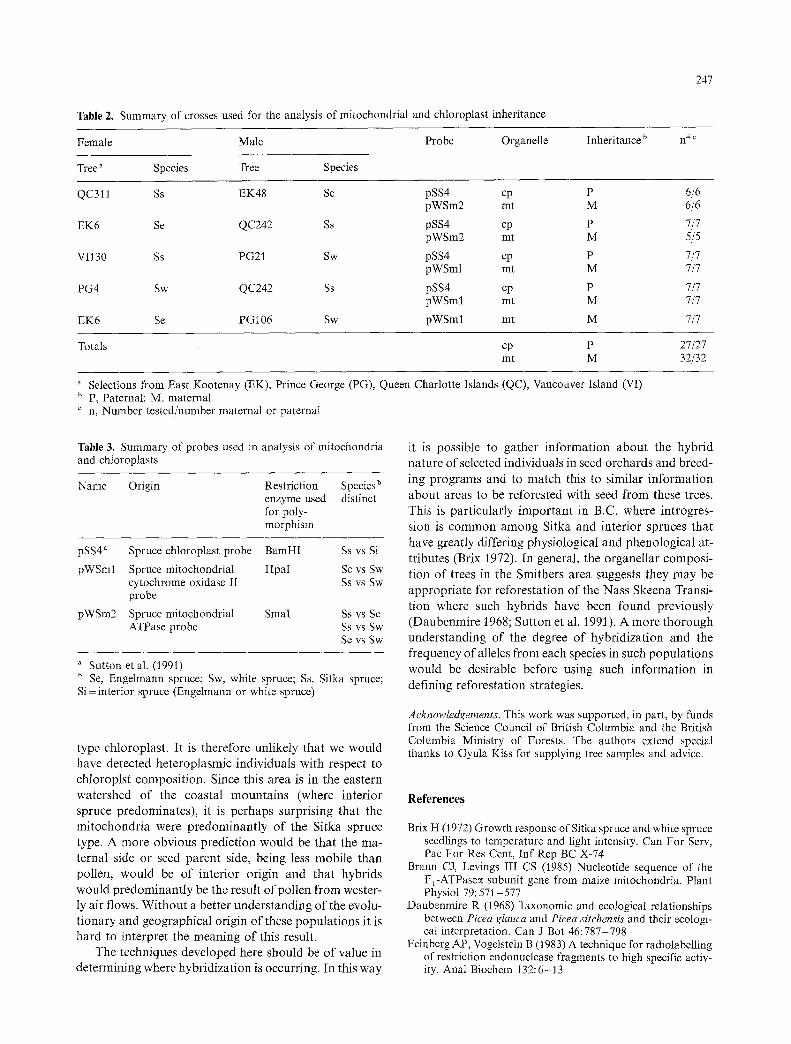
Fourteen individuals were selected based on a scattered distribution over an area of approximately 130 km by 60 km (PR in Fig. 1.). This region is on the southeastern fringe of the Nass Skeena Transition where introgression of Sitka and interior spruce has been described (Dauben- mire et al. 1968; Krajina et al. 1982; Roche 1969; Sutton et al. 1991). Both the chloroplast and mitochondrial D N A complement of each individual was analyzed using the probes described above. The results are presented in Fig. 5. The 10.5 kb chloroplast fragment characteristic of interior spruce species (Sutton et al. 1991) appears in all trees except PR14, which is characteristic of Sitka spruce (Fig. 5A). The result of probing with the pWSml mitchondrial probe indicates that all the individuals con-
tain the 20-kb band characteristic of both Sitka and En- gelmann spruce (Fig. 5 B); this probe will not distinguish these two species. Analysis of the results with pWSm2 (Fig. 5 C) show that diagnostic bands for both Sitka (12 trees) and Engelmann spruce (2 trees) are found in differ- ent individuals. Notably, PR 14, which showed a Sitka type chloroplast is among the individuals that contain an Engelmann spruce mitochondrial type. These results are summarized in Table 1.
Discussion
This paper describes the isolation of mitochondrial probes which can be used to reveal polymorphisms that
246
Fig. 4A, B. Analysis of organelle inheritance in progeny of con- trolled crosses between white and Sitka spruce. DNA from the progeny precede the parents of each cross. Panel A Total DNA, 2 p~g, digested with BamHI and hybridized with pSS4 chloro- plast probe. Panel B Total DNA 4 gg, digested with Hpal and hybridized with pWSml mitochondrial probe. Chloroplast bands in the progeny are identical to the male parent, and mito- chondrial bands are identical to the female parent
distinguish white, Sitka and Engelmann spruce species. While the number of individuals analyzed was relatively small, a more extensive screening of these populations is beyond the scope of this paper. Nonetheless, it seems likely that the species-specific patterns we have observed are representative of the species populations as a whole for the following reasons: (1) the mitochondrial D N A appears to be very highly conserved based on the number of restriction digests that showed no interspecies poly- morphisms with these probes; (2) the banding patterns characteristic of each species are found in widely separat- ed populations (white spruce from Ontario and B.C., and Sitka spruce from Vancouver Island and the Queen Char- lotte Islands); 3) the species assignations made using one mitochondrial probe are consistent with the other probe for each individual.
The analysis of organelle inheritance in controlled crosses between species clearly demonstrates that mito- chondria are maternally inherited and that chloroplasts are paternally inherited. This pattern of inheritance has also been demonstrated for Pinus (Neal et al. 1989) which, like Picea, is a member of the Pinaceae. The re- sults imply that such analyses can be used to determine
Fig. 5A-C. Organelle composition of mature trees from the Sitka/interior spruce introgression zone (Smithers B.C.) Total DNA of each individual, 2 gg for the chloroplast probe and 4 gg for mitochondrial probes, was digested with the appropriate restriction enzyme, Southern blotted and hybridized with each probe. Panel A Digested with BamHI and probed with pSS4 chloroplast probe. Panel B Digested with HpaI and probed with pWSml mitochondrial probe. Panel C Digested with SmaI and probed with pWSm2
the maternal and paternal lineage represented in any individual analyzed. In no case was any biparental inher- itance observed. This result is perhaps surprising since our previous work showed that chloroplast D N A frag- ments typical of both Sitka and interior spruce could be found in the same individuals from introgression zones (Sutton et al. 1991). In this study we analyzed 14 individ- uals from an area around Smithers B.C., which is just east of the recognized introgression between Sitka and interi- or spruces (Nass Skeena Transition). While these individ- uals are naturally occuring hybrids, most had a Sitka spruce mitochondrial complement and interior spruce
247
Table 2. Summary of crosses used for the analysis of mitochondrial and chloroplast inheritance
Female Male Probe Organelle Inheritance b n ~ o
Tree a Species Tree Species
QC3 t 1 Ss EK48 Se pSS4 cp P 6/6 pWSm2 mt M 6/6
EK6 Se QC242 Ss pSS4 cp P 7/7 pWSm2 mt M 5/5
VIi30 Ss PG21 Sw pSS4 cp P 7/7 pWSml mt M 7/7
PG4 Sw QC242 Ss pSS4 cp P 7/7 pWSml mt M 7/7
EK6 Se PG106 Sw pWSmt mt M 7/7
Totals cp P 27/27 mt M 32/32
Selections from East Kootenay (EK), Prince George (PG), Queen Charlotte Islands (QC), Vancouver Island (VI) b p, Paternal; M, maternal
n, Number tested/number maternal or paternal
Table 3. Summary of probes used in analysis of mitochondria and chloroplasts
Name Origin Restriction Species b enzyme used distinct for poly- morphism
pSS4 ~ Spruce chloroplast probe BamHI Ss vs Si
pWSml Spruce mitochondrial HpaI Se vs Sw cytochrome oxidase II Ss vs Sw probe
pWSm2 Spruce mitochondrial SmaI Ss vs Se ATPase probe Ss vs Sw
Se vs Sw
a Sutton et al. (1991) b Se, Engelmann spruce; Sw, white spruce; Ss, Sitka spruce; Si = interior spruce (Engelmann or white spruce)
type chloroplast . It is therefore unlikely that we would have detected heteroplasmic individuals with respect to chloroplst composi t ion. Since this area is in the eastern watershed of the coastal mountains (where interior spruce predominates) , it is perhaps surprising that the mi tochondr ia were predominant ly of the Sitka spruce type. A more obvious predict ion would be that the ma- ternal side or seed parent side, being less mobile than pollen, would be of interior origin and that hybrids would predominant ly be the result of pollen from wester- ly air flows. Wi thout a better unders tanding of the evolu- t ionary and geographical origin of these populat ions it is hard to interpret the meaning of this result.
The techniques developed here should be of value in determining where hybridizat ion is occurring. In this way
it is possible to gather informat ion about the hybrid nature of selected individuals in seed orchards and breed- ing programs and to match this to similar informat ion about areas to be reforested with seed from these trees. This is part icular ly impor tan t in B.C. where introgres- sion is common among Sitka and interior spruces that have greatly differing physiological and phenological at- tributes (Brix 1972). in general, the organellar composi- tion of trees in the Smithers area suggests they may be appropr ia te for reforestat ion of the Nass Skeena Transi- tion where such hybrids have been found previously (Daubenmire 1968; Sutton et al. 1991). A more thorough understanding of the degree of hybridizat ion and the frequency of alleles from each species in such populat ions would be desirable before using such informat ion in defining reforestat ion strategies.
Acknowledgements. This work was supported, in part, by funds from the Science Council of British Columbia and the British Columbia Ministry of Forests. The authors extend special thanks to Gyula Kiss for supplying tree samples and advice.
References
Brix H (1972) Growth response of Sitka spruce and white spruce seedlings to temperature and light intensity. Can For Serv, Pac For Res Cent, Inf Rep BC-X-74
Braun CJ, Levings III CS (/985) Nucleotide sequence of the F1-ATPase~ subunit gene from maize mitochondria. Plant Physiol 79:571-577
Daubenmire R (1968) Taxonomic and ecological relationships between Picea glauca and Picea sitchensis and their ecologi- cal interpretation. Can J Bot 46:787-798
Feinberg AP, Vogelstein B (1983) A technique for radiolabelling of restriction endonuclease fragments to high specific activ- ity. Anal Biochem 132:6-13

248
Feinberg AP, Vogelstein B (1984) Addendum to "A technique for radiolabelling of restriction endonuclease fragments to high specific activity". Anal Biochem 137:266-267
Fox TD, Leaver CJ (1981) The Zea mays mitochondrial gene coding cytochrome oxidase subunit II has an intervening sequence and does not contain TGA codons. Cell 26: 315- 323
Krajina VT, Klinka K, Worrall J (1982) Distribution and eco- logical characteristics of trees and shrubs of British Colum- bia. UBC Fac For Pub1
Maniatis T, Fritsch EF, Sambrook J (1982) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
Neale DB, Sederoff RR (1989) Paternal inheritance of chloro- plast DNA and maternal inheritance of mitochondrial DNA in loblolly pine. Theor Appl Genet 77:212-216
Neale DB, Wheeler NC, Allard RW (1986) Paternal inheritance of chloroplast DNA in Douglas-fir. Can J For Res 6:1152- 1154
Neale DB, Marshall KA, Sederoff RR (1989) Chloroplast and mitochondrial DNA are paternally inherited in Sequoia sem- pervirens. DDon. Endl. Proc Natl Acad Sci USA 86: 9347- 9349
Palmer JD (1987) Chloroplast DNA evolution and biosystemat- ic uses of chloroplast DNA variation. Am Nat 130:$6 $29
Pay A, Smith MA (1988) A rapid method for the purification of organelles for DNA isolation: self-generated percoll gradi- ents. Plant Cell Rep 7:96-99
Roche L (1969) A genecological study of the genus Picea in British Columbia. New Phytol 68:505-554
Sederoff RR (1987) Molecular mechanisms of mitochondrial genome evolution. Am Nat 130:$30-$45
Sutton BCS, Flanagan DJ, E1-Kassaby YA (1991) A simple and rapid method for estimating representation of species in spruce seedlots using chloroplast DNA restriction fragment length polymorphism. Silvae Genet (in press)
Szmidt AE, Alden T, Hallgren JE (1987) Paternal inheritance of chloroplast DNA in Larix. Plant Mol Biol 9:59-64
Szmidt AE, E1-Kassaby YA, Sigurgeirsson A, Alden T, Lindgren D, Hallgren JE (1988) Classifyig seedlots of Picea sitchensis and P. glauca in zones of introgression, using restriction analysis of chloroplast DNA. Theor Appl Genet 76:841-845
Wagner DB, Furnier GR, Saghai-Maroof MA, Williams SM, Dancik BP, Allard RW (1987) Chloroplast DNA polymor- phism in lodgepole and jack pines and their hybrids. Proc Natl Acad Sci USA 84:2097-2100
Webb DT, Webster 17, Flinn BS, Roberts DR, Ellis DE (1989) Factors influencing the induction of embryogenic and caulo- genic callus from embryos of Picea glauea and P. engelman- nii. Can J For Res 19:1303-1308
White EE (1990) Chloroplast DNA in Pinus montieola: 2. Sur- vey of within species variability and detection of heteroplas- mic individuals. Theor Appl Genet 79:251-255







![genetic variation of norway spruce (picea abies [l.] karst ...](https://static.fdokumen.com/doc/165x107/633d6582b988f9753c06267d/genetic-variation-of-norway-spruce-picea-abies-l-karst-.jpg)












