
Hepatocarcinogenesis: A polygenic model of inherited predisposition to cancer
Upload
independentCategory
view
0download
0
Genes involved in the WNT and vesicular trafficking pathwaysare associated with melanoma predisposition
Maider Ibarrola-Villava1, Rajiv Kumar2, Eduardo Nagore3, Meriem Benfodda4,5, Mickael Guedj6, Steven Gazal7,
Hui-Han Hu4,5, Jian Guan2, P. Sivaramakishna Rachkonda2, Vincent Descamps4,8, Nicole Basset-Seguin4,8,
Armand Bensussan4, Martine Bagot4,9, Philippe Saiag10, Dirk Schadendorf11, Manuel Martin-Gonzalez12, Matias Mayor13,
Bernard Grandchamp5, Gloria Ribas1 and Nadem Soufir4,5
1 Department of Haematology and Medical Oncology, Biomedical Research Institute INCLIVA, Valencia 46010, Spain2 Division of Molecular Genetic Epidemiology, German Cancer Research Center, Heidelberg 69120, Germany3 Department of Dermatology, Instituto Valenciano de Oncologia, Valencia 46009, Spain4 Inserm U976, Centre de Recherche Sur la Peau, Hopital Saint Louis, Universit�e Paris 7, Paris 75010, France5 D�epartement de G�en�etique, Hopital Bichat, APHP, Paris 75018, France6 Laboratoire Statistiques et Genomes, Evry 91000, France7 UMR S738, Facult�e de M�edecine Xavier Bichat, Paris 75018, France8 Department of Dermatology, Hopital Bichat, APHP, Paris 75018, France9 Department of Dermatology, Hopital Saint Louis, APHP, Paris 75010, France10 Department of Dermatology, Hopital Ambroise Par�e, APHP, Paris 92100, France11 Department of Dermatology, University Hospital Essen, Esse 45147, Germany12 Department of Dermatology, Hospital Ramon y Cajal, Madrid 28034, Spain13 Department of Dermatology, Hospital La Paz, Madrid 28046, Spain
Multifactorial predisposition to melanoma includes genes involved in pigmentation, immunity and DNA repair. Nonetheless,
missing heritability in melanoma is still important. We studied the role of 335 candidate SNPs in melanoma susceptibility by
using a dedicated chip and investigating 110 genes involved in different pathways. A discovery set was comprised of 1069
melanoma patients and 925 controls from France. Data were replicated using validation phases II (1085 cases and 801 con-
trols from Spain) and III (1808 cases and 1894 controls from Germany and a second set of Spanish samples). In addition, an
exome sequencing study was performed in three high-risk French melanoma families. Nineteen SNPs in 17 genes were initially
associated with melanoma in the French population. Six SNPs were replicated in phase II, including two new SNPs in the
WNT3 (rs199524) and VPS41 (rs11773094) genes. The role of VPS41 and WNT3 was confirmed in a meta-analysis (3940 mela-
noma cases and 3620 controls) with two-side p values of 0.002, (OR 5 0.86) and 4.07 3 10210 (OR 5 0.80), respectively.
Exome sequencing revealed a non-synonymous VPS41 variant in one family that was shown to be strongly associated with
familial melanoma (OR 5 4.46, p 5 0.001) in an independent sample of 178 melanoma families. WNT3 belongs to WNT path-
way known to play a crucial role in melanoma, whereas VPS41 regulates vesicular trafficking and is thought to play a role in
pigmentation. Our work identified two new pathways involved in melanoma predisposition. These results may be useful in the
future for identifying individuals highly predisposed to melanoma.
The knowledge of the genetic predisposition to melanoma hassignificantly increased during the last several years. Linkagestudies, candidate gene approaches, genome-wide associationstudies (GWAS) and whole-exome sequencing studies have
identified several loci that influence susceptibility to cutaneousmalignant melanoma (MM).1–3 Mutations in the highly pene-trant CDKN2A (MIM#60160), CDK4 (MIM#609048), TERT(MIM#187270) and POT1 (MIM#606478) genes are well
Key words: exome sequencing, case-control study, WNT3, VPS41, melanoma
Abbreviations: CI: confidence interval; GWAS: genome-wide association studies; HWE: Hardy Weinberg equilibrium; MAF: minor allele
frequency; MM: malignant melanoma; OR: odds ratio; SNP: single-nucleotide polymorphism
Additional Supporting Information may be found in the online version of this article.
Grant sponsor: Ministerio de Salud Carlos III; Grant number: PI10-0405, CP08-00069; Grant sponsor: La Societe Francaise de
Dermatologie (SFD), FEDER
DOI: 10.1002/ijc.29257
History: Received 7 Apr 2014; Accepted 9 Sep 2014; Online 10 Oct 2014
Correspondence to: Nadem Soufir, D�epartement de G�en�etique, Hopital Bichat Claude Bernard, 46 rue Henri Huchard, 75018 Paris, France.
Tel.: [33140258551], Fax: 1[33140258785], E-mail: [email protected]
Can
cerGenetics
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC
International Journal of Cancer
IJC

known to play a role in familial melanoma, which accountsfor about 10% of all MM cases.4–6 Moreover, low penetrancegenetic variants have been associated with sporadic form ofthe disease and, therefore, pigmentation, nevus count, immuneresponses, DNA repair, oxidative stress and the vitamin Dreceptor have been associated with melanomasusceptibility.7–12
Although different genetic pathways are involved in mela-noma onset, the human pigmentation pathway seems to playa critical role in the pathogenesis of the disease. MM predis-position is increased in individuals characterized by fair skincolour, high mole count, blond or red hair and high sun sen-sibility (Fitzpatrick’s phototypes I and II).4 Since the inci-dence of malignant melanoma in part reflects theconsequences of a complex interaction between skin pheno-type and UV exposure, variations in genes implicated in pig-mentation and tanning response to UV are associated withsusceptibility to MM. Thus, comprehensive knowledge of theentire network of genetic variants in pigmentation and mela-nogenesis genes may be a useful predictor of MM risk in thegeneral population.13,14
The pigmentation pathway is a multistep process thatinvolved differentiation and migration of melanocytes (pig-ment-producing cells), melanosome biogenesis, and migrationand melanin synthesis. More than 120 genes have been identi-fied through animal models. Nevertheless, only one gene, themelanocortin 1 receptor (MC1R, MIM#155555) is known toaccount for a substantial variation in the incidence of sporadicMM risk.15 Several other genes, such as OCA2 (MIM#611409),ASIP (MIM#600201), SLC45A2 (MIM#606202), TYR(MIM#606933) or TYRP1 (MIM#115501) have also emerged asbeing potentially important in melanoma susceptibility.1, 16–18
Considering the involvement of pigmentation in MM pre-disposition, the goal of the current study was to identifynovel variants associated with MM risk by using single-nucleotide polymorphism (SNP) array on three independentsporadic melanoma series of European samples: France, Ger-many and Spain. Moreover, we performed whole-exomesequencing on a series of three melanoma families.
Material and MethodsStudy subjects and data collection
Discovery phase I. A total number of 1069 melanoma caseswere recruited from 2002 to 2008 at the Departments of Der-matology of five different university-affiliated hospitals from
the Paris region (Saint Louis Hospital, Bichat Hospital, Tar-nier Hospital, Ambroise Par�e Hospital and Tenon Hospital).This cohort, called MelanCohort, comprised 888 sporadicmelanoma cases (83%), 61 multiple melanoma patients(5.7%) and 120 nonrelated familial melanoma cases (11.3%).Similarly, 925 skin cancer-free control subjects were recruitedfrom the EFS (Etablissement Francais du Sang) at Bichat andSaint Louis hospitals (Paris region) during the same period.
A standardised questionnaire was used to collect informa-tion on pigmentation characteristics, the presence of child-hood sunburns, Fitzpatrick’s skin type classification andpersonal and family history of cancer.
Validation phase II. A second phase of the study was com-posed of 1085 nonrelated MM cases obtained from theDepartments of Dermatology of three different hospitals inMadrid, Spain (Gregorio Mara~non Hospital, La Paz Hospitaland Ramon y Cajal Hospital) and from the Instituto Valen-ciano de Oncologia in Valencia, Spain. A total of 801 cancer-free controls from the college of lawyers (Madrid, Spain),Gregorio Mara~non Hospital and Instituto Valenciano deOncologia were recruited. All subjects came from the geo-graphical regions covered by the hospitals involved in thestudy.
Validation phase III. A third phase of the study was com-posed of two different sample sets. The first sample setincluded 872 nonrelated MM cases and 1158 cancer-free con-trols obtained from a different collection from the InstitutoValenciano de Oncologia. The second one included 936 MMpatients and 736 controls recruited from the German CancerResearch Centre (Heidelberg, Germany). A total of 1808 mel-anoma cases and 1894 control subjects were included in thisphase III validation. All subjects came from the geographicalregions covered by their respective study hospitals.
All enrolled patients and controls were from Caucasianorigin. Distribution of cases and controls in discovery andvalidation phases is summarised in Supporting InformationTable 1 and Supporting Information Table 2.
Genomic DNA from cases and controls was isolated fromperipheral blood lymphocytes using the MagNA Pure LCInstrument according to the manufacturer’s protocol (RocheApplied Science, Mannheim, Germany). DNA concentrationwas quantified using Quant-iT PicoGreen dsDNA Reagent(Invitrogen, Eugene, OR). All DNA samples were subjectedto a quality control before processing them on the Illumina
What’s new?
While our understanding of the molecular pathways that lead to malignant melanoma has increased over the past few years,
it is still incomplete. In this study, the authors identified two new pathways: one that involves WNT signaling (involved in mel-
anoblast development), and another that involves vesicular trafficking (involved in changes in pigmentation phenotype). These
results may be useful in developing screening tests for people who are genetically susceptible to melanoma. In addition, the
whole-exome sequencing techniques used in this study may aid researchers in identifying mutations in unknown disease-
candidate genes.
Can
cerGenetics
2 VPS41 and WNT3 and melanoma susceptibility
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

arrays (Illumina, San Diego, CA). All subjects gave writteninformed consent, and the study was approved by the EthicsCommittees of the Gregorio Mara~non Hospital, HospitalClinico Universitario of Valencia and the Ambroise Par�eHospital (Boulogne, France).
Gene and SNP selection
Previous literature and the information available on publicdatabases were used for gene selection. Eventually, 110 geneswere included in the study. The most overrepresented cate-gory with 96 genes was pigmentation. Pigmentation geneswere chosen from the albinism and pigmentation genes data-bases (http://www.espcr.org/micemut/). A further group of 14genes implicated in DNA repair (3 genes), cell cycle (5genes), immune responses and apoptosis (4), MAP kinasepathway (1 gene) and glutamate receptor (1 gene) wereincluded in the study.
Three databases were used to collect information aboutsingle nucleotide polymorphisms (SNPs): NCBI (www.ncbi.nlm.nih.gov/SNP), HapMap (www.hapmap.org) and Illuminadatabases (www.illumina.com). Finally, 384 SNPs were cho-sen that included (a) polymorphisms predicted to have afunctional impact (snpinfo.niehs.nih.gov/snpinfo/snpfunc-.htm) that comprise SNPs located in putative transcriptionfactor binding sites (66 SNPs) or in conserved regions (81SNPs), nonsynonymous polymorphisms (49 SNPs), SNPspredicted to affect splicing (65 SNPs) or miRNA binding sites(63 SNPs), and (b) some tag-SNPs (87 SNPs). Tag-SNPsrange from the hypothetical promoter area (�2000 Kbupstream) until �2000 Kb downstream of the gene. EightSNPs commonly associated with MM were included as posi-tive controls (rs4911414 in ASIP, rs11547464, rs1805007 andrs1805009 in MC1R, rs16891982 in SLC45A2, rs1126809 inTYR, rs2733832 in TYRP1 and rs910873 in the 20Q region).To ensure a high genotyping success rate, a minor allele fre-quency threshold of 0.05 in the European HapMap popula-tion and a recommended manufacturer “Illumina score” wereestablished in the SNP selection. SIFT and PolyPhen, (http://sift.jcvi.org/ and http://genetics.bwh.harvard.edu/pph2/,respectively), were used to evaluate predictive functional con-sequences of nonsynonymous changes.
SNP genotyping
SNPs were genotyped using the GoldenGate Genotyping Assaysystem according to the manufacturer’s protocol (Illumina,San Diego, CA). Genotyping was carried out by the genomicplatform of the Pasteur Institute (CNRS UMR 8090, InstitutBiologie de Lille, France) using 1.5 mcg of DNA per reaction.Genotyping specificity was assessed by including two negativecontrols in each 96-well plate genotyped, yielding 100% con-sistent results. Genotypes were called using the proprietarysoftware supplied by Illumina (BeadStudio, 2009 version).
Validation set genotyping was carried out using the KASPSNP genotyping System (LGC Genomics, Hoddesdon, UK).Five ng of DNA were used for each genotyping reaction. For
quality control measure, one sample duplicate and a nontem-plate sample per 96-well plate were included. For the first vali-dation set (Phase II), a total of 13 SNPs were genotyped while7 SNPs were studied in the second validation sample set (PhaseIII). Genotypes were scored by the LGC Genomics Company.
Exome sequencing
Three French families were included in an exome sequencingstudy with the following criteria: at least three individualsaffected in a family, and at least one patient affected withmultiple melanomas. Two affected relatives per family weresequenced. None of them carried CDKN2A, CDK4 or BAP1mutations. Sequencing was done with the V4 Agilent kitexon capture (Otogenetics Company), coverage 50X, accord-ing to the manufacturer’s protocol using HiSeq 2000 fromIllumina. Sequencing was carried out using 5 mcg of DNAper reaction. Variants were filtered using the DNA Nexussoftware (www.dnanexus.com). Only variants with allele fre-quency <1% were retained.
Statistical analysis
Univariate and multivariate analyses were performed usingthe R software (version 8). All statistical tests performed weretwo-sided. Deviation from null-hypothesis was consideredsignificant at the 5% level. All odds ratios (ORs) are reportedwith their 95% confidence interval (CI). Deviations fromHardy-Weinberg equilibrium (HWE) were verified for eachgenetic marker using Pearson chi-square test. Allelic and gen-otypic association was tested according to the additive modelsof inheritance using logistic regression (glm() R procedure)before and after adjustment for sex and known melanomarisk factors (nevi count, skin type, eye colour, and nevicount). Correction for multiple hypothesis testing was carriedout using the Bonferroni method.
Association between genotyped variants in pigmentationgenes and various pigmentation characteristics and sun sensi-bility were also assessed via logistic regression. Finally, ameta-analysis involving all phases was performed applying aCochran-Mantel-Haenszel test,19 as described previously.20
ResultsThe role of 384 polymorphisms in 110 genes was initiallyinvestigated. Forty-two SNPs (11%) were discarded becauseof unsuccessful genotyping. Evidence for the departure fromHardy-Weinberg equilibrium (HWE) among controls wasobserved for 23 SNPs of which 7 remained statistically signif-icant after a conservative Bonferroni correction for multiplehypothesis testing. Overall, remaining 335 SNPs wereincluded in the analysis. Genotyping results do not differedsubstantially from other SNP array association publishedstudies of similar characteristics.21,22
Associations with melanoma risk
Representation of –log10 p values for the comparison of thegenotypes between the 1069 French cases and the 925
Can
cerGenetics
Ibarrola-Villava et al. 3
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC
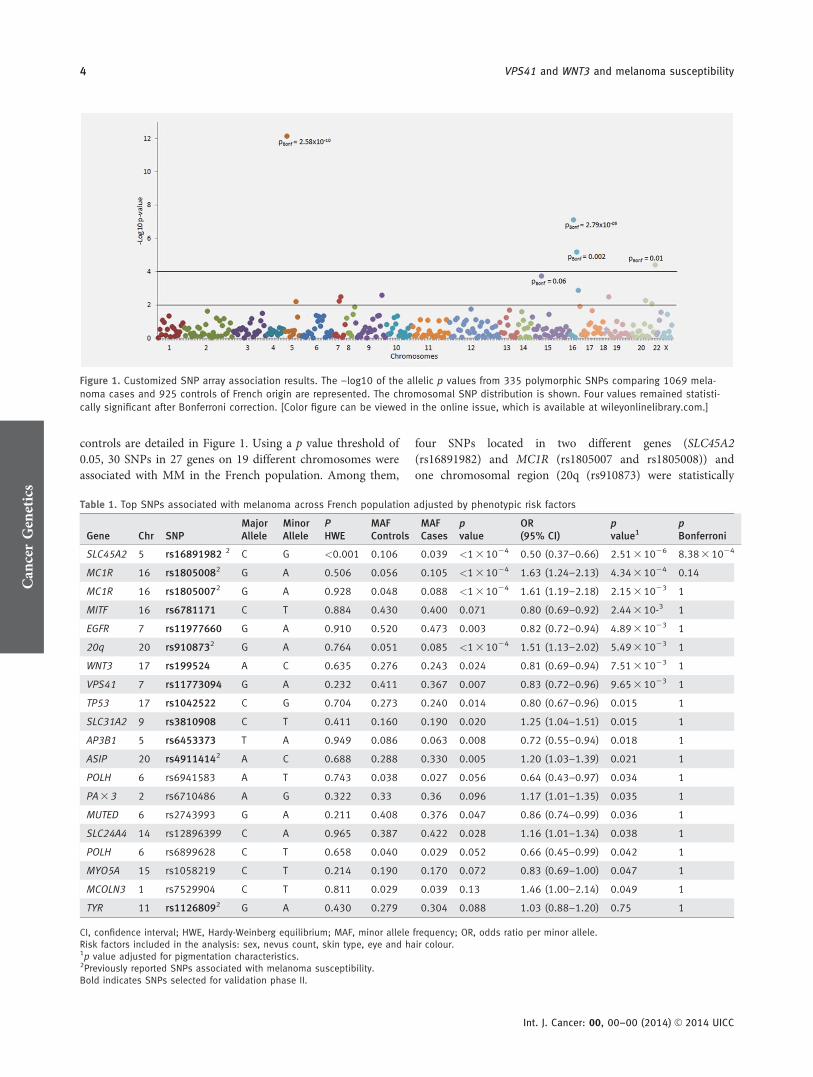
controls are detailed in Figure 1. Using a p value threshold of0.05, 30 SNPs in 27 genes on 19 different chromosomes wereassociated with MM in the French population. Among them,
four SNPs located in two different genes (SLC45A2(rs16891982) and MC1R (rs1805007 and rs1805008)) andone chromosomal region (20q (rs910873) were statistically
Figure 1. Customized SNP array association results. The –log10 of the allelic p values from 335 polymorphic SNPs comparing 1069 mela-
noma cases and 925 controls of French origin are represented. The chromosomal SNP distribution is shown. Four values remained statisti-
cally significant after Bonferroni correction. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Table 1. Top SNPs associated with melanoma across French population adjusted by phenotypic risk factors
Gene Chr SNPMajorAllele
MinorAllele
PHWE
MAFControls
MAFCases
pvalue
OR(95% CI)
pvalue1
pBonferroni
SLC45A2 5 rs16891982 2 C G <0.001 0.106 0.039 <1 3 1024 0.50 (0.37–0.66) 2.51 3 1026 8.38 3 1024
MC1R 16 rs18050082 G A 0.506 0.056 0.105 <1 3 1024 1.63 (1.24–2.13) 4.34 3 1024 0.14
MC1R 16 rs18050072 G A 0.928 0.048 0.088 <1 3 1024 1.61 (1.19–2.18) 2.15 3 1023 1
MITF 16 rs6781171 C T 0.884 0.430 0.400 0.071 0.80 (0.69–0.92) 2.44 3 10-3 1
EGFR 7 rs11977660 G A 0.910 0.520 0.473 0.003 0.82 (0.72–0.94) 4.89 3 1023 1
20q 20 rs9108732 G A 0.764 0.051 0.085 <1 3 1024 1.51 (1.13–2.02) 5.49 3 1023 1
WNT3 17 rs199524 A C 0.635 0.276 0.243 0.024 0.81 (0.69–0.94) 7.51 3 1023 1
VPS41 7 rs11773094 G A 0.232 0.411 0.367 0.007 0.83 (0.72–0.96) 9.65 3 1023 1
TP53 17 rs1042522 C G 0.704 0.273 0.240 0.014 0.80 (0.67–0.96) 0.015 1
SLC31A2 9 rs3810908 C T 0.411 0.160 0.190 0.020 1.25 (1.04–1.51) 0.015 1
AP3B1 5 rs6453373 T A 0.949 0.086 0.063 0.008 0.72 (0.55–0.94) 0.018 1
ASIP 20 rs49114142 A C 0.688 0.288 0.330 0.005 1.20 (1.03–1.39) 0.021 1
POLH 6 rs6941583 A T 0.743 0.038 0.027 0.056 0.64 (0.43–0.97) 0.034 1
PA 3 3 2 rs6710486 A G 0.322 0.33 0.36 0.096 1.17 (1.01–1.35) 0.035 1
MUTED 6 rs2743993 G A 0.211 0.408 0.376 0.047 0.86 (0.74–0.99) 0.036 1
SLC24A4 14 rs12896399 C A 0.965 0.387 0.422 0.028 1.16 (1.01–1.34) 0.038 1
POLH 6 rs6899628 C T 0.658 0.040 0.029 0.052 0.66 (0.45–0.99) 0.042 1
MYO5A 15 rs1058219 C T 0.214 0.190 0.170 0.072 0.83 (0.69–1.00) 0.047 1
MCOLN3 1 rs7529904 C T 0.811 0.029 0.039 0.13 1.46 (1.00–2.14) 0.049 1
TYR 11 rs11268092 G A 0.430 0.279 0.304 0.088 1.03 (0.88–1.20) 0.75 1
CI, confidence interval; HWE, Hardy-Weinberg equilibrium; MAF, minor allele frequency; OR, odds ratio per minor allele.Risk factors included in the analysis: sex, nevus count, skin type, eye and hair colour.1p value adjusted for pigmentation characteristics.2Previously reported SNPs associated with melanoma susceptibility.Bold indicates SNPs selected for validation phase II.
Can
cerGenetics
4 VPS41 and WNT3 and melanoma susceptibility
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

significant after applying the Bonferroni correction for multi-ple comparisons. In addition, when adjusted by phenotypicrisk factors 19 SNPs in 17 genes remained significantly asso-ciated to melanoma risk. A detailed information on SNP rs#number, genes, chromosome location, minor allele frequen-cies (MAF), Odds Ratio (OR), 95% confidence interval (CI)and p values for comparisons between cases and control sub-jects are represented in Table 1.
Of these 19 associated SNPs (p< 0.5), we selected 13SNPs that had a nominal p value less than 0.02 for a valida-tion phase II that consisted of an independent set of 1085Spanish MM cases and 801 controls. The genotype distri-bution for all the SNPs was in HWE except for three SNPsthat were excluded. Six SNPs had an unadjusted p< 0.05 inphase II. MC1R, SLC45A2 and 20q region SNPs were associ-ated with melanoma in this phase. Moreover, rs11773094in the VPS41 gene and rs199524 in WNT3 had an unadjustedp of 0.004 and 0.002, respectively, in phase II. All remainedsignificant after multiple testing corrections. None of theother 4 SNPs tested at this stage reached statisticalsignificance.
Nine SNPs were genotyped in 1808 melanoma patientsand 1894 control subjects in validation phase III. None of theSNPs presented evidence of a departure from HWE at thisstage. Five SNPs (SLC45A2 (rs16891982), MC1R (rs1805007and rs1805008), TYR (rs1126809) and WNT3 (rs199524)remained associated with MM susceptibility after correctionfor multiple testing.
Finally, six SNPs, located in SLC45A2, MC1R, TYR, VPS41and WNT3, showed an overall p< 0.05 when all the phaseswere considered together (3940 MM cases and 3620 controls).These six SNPs were the following: rs16891982 located in theSLC45A2 gene that causes the amino-acid change p.Phe374Leuwith an overall OR5 0.4 (95% CI: 0.35–0.46, p5 5.333
10245); rs1805007 and rs1805008 located in MC1R encodep.Arg151Cys and p.Arg160Trp amino-acid changes, respec-tively, and had an overall OR of 2.25 (95% CI: 1.89–2.69,p5<13 1024) and 1.90 (95% CI: 1.37–2.65, p5<13 1024)respectively, rs11773094, located in the 3’UTR of the VPS41gene, with an overall OR5 0.86 (95% CI: 0.79–0.95, p5 0.002);rs199524, located in an intronic region of the WNT3 gene,with a global OR5 0.80 (95% CI: 0.74–0.85, p5 4.073 10210);
Figure 2. Forest plot showing the already known SLC45A2 and TYR variants and two novel associated polymorphisms in VPS41 and WNT3
with melanoma susceptibility. Dots represent odds ratios in the treated sub-groups versus those in control groups. Diamond shapes repre-
sent pools results for phase III. Squares shapes represent the overall odds ratios. The size of the dots and squares are proportional to the
number of individuals, and error bars represent 95% confidence intervals.
Can
cerGenetics
Ibarrola-Villava et al. 5
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

Tab
le2
.A
sso
cia
tio
nw
ith
ph
en
oty
pic
cha
ract
eri
stic
sa
cro
ssth
eFr
en
chp
op
ula
tio
n
Ge
ne
SN
PS
kin
typ
eN
evu
sco
un
tS
kin
colo
rH
air
colo
rE
yeco
lor
Ep
he
llid
es
OR
(95
%C
I)p
valu
eO
R(9
5%
CI)
pva
lue
OR
(95
%C
I)p
valu
eO
R(9
5%
CI)
pva
lue
OR
(95
%C
I)p
valu
eO
R(9
5%
CI)
pva
lue
20
qrs
91
08
73
1.6
8(1
.28
–2
.20
)1
.70
31
02
4,2
1.1
6(0
.86
–1
.56
)0
.33
2.0
5(1
.04
–4
.04
)0
.04
1.2
4(0
.95
–1
.62
)0
.12
0.7
7(0
.59
–1
.02
)0
.07
1.5
9(1
.21
–2
.10
)9
.03
10
24
AD
AM
TS2
0rs
10
50
62
26
0.9
5(0
.80
–1
.12
)0
.54
1.1
0(0
.91
–1
.33
)0
.32
0.7
5(0
.56
–1
.01
)0
.05
0.8
7(0
.74
–1
.02
)0
.10
1.2
6(1
.07
–1
.48
)0
.00
71
.09
(0.9
1–
1.3
0)
0.3
5
DC
Trs
95
16
41
80
.95
(0.8
3–
1.1
0)
0.5
11
.03
(0.8
8–
1.2
0)
0.7
20
.79
(0.6
1–
1.0
2)
0.0
70
.78
(0.6
8–
0.8
9)
2.0
31
02
41
.22
(1.0
6–
1.4
0)
0.0
05
1.0
4(0
.89
–1
.20
)0
.63
ED
Ars
65
25
31
70
.67
(0.4
3–
1.0
3)
0.0
70
.60
(0.3
3–
1.0
9)
0.1
00
.32
(0.2
1–
0.4
9)
1.6
03
10
27
,10
.76
(0.5
1–
1.1
2)
0.1
61
.32
(0.8
9–
1.9
4)
0.1
60
.86
(0.5
5–
1.3
5)
0.5
2
GP
NM
Brs
19
93
47
1.0
4(0
.91
–1
.19
)0
.55
0.9
1(0
.77
–1
.06
)0
.22
0.8
2(0
.63
–1
.05
)0
.12
0.9
6(0
.84
–1
.10
)0
.54
1.2
1(1
.05
–1
.38
)0
.01
0.9
6(0
.83
–1
.12
)0
.62
HE
RC
2rs
12
91
38
32
0.6
7(0
.58
–0
.77
)1
.10
31
02
8,1
0.9
1(0
.78
–1
.07
)0
.25
0.3
7(0
.28
–0
.48
)2
.23
31
02
13
10
.31
(0.2
6–
0.3
6)
1.0
03
10
25
0,1
14
.14
(10
.92
–1
8.3
0)
6.4
93
10
29
0,1
0.7
4(0
.64
–0
.86
)7
.88
31
02
05
,1
LYS
Trs
11
46
41
.07
(0.4
8–
2.3
8)
0.8
80
.44
(0.1
3–
1.4
9)
0.1
90
.13
(0.0
5–
0.3
0)
2.1
93
10
26
,10
.31
(0.1
2–
0.7
8)
0.0
12
.58
(1.1
0–
6.0
7)
0.0
30
.25
(0.0
6–
1.0
9)
0.0
7
KIT
LGrs
12
82
12
56
1.1
4(0
.88
–1
.47
)0
.31
0.9
7(0
.72
–1
.30
)0
.84
1.3
9(7
.08
–3
.13
)0
.01
1.8
2(1
.39
–2
.37
)1
.36
31
02
5,1
0.7
4(0
.57
–0
.96
)0
.02
0.9
3(0
.71
–1
.23
)0
.63
MC
1R
rs1
80
50
07
2.5
5(1
.89
–3
.45
)1
.23
31
02
9,1
1.1
1(0
.81
–1
.53
)0
.51
1.0
0(0
.59
–1
.72
)0
.99
1.5
5(2
.83
–2
.09
)1
.56
31
02
60
.95
(0.7
2–
1.2
6)
0.7
13
.17
(2.3
6–
4.2
6)
1.5
13
10
21
41
MC
1R
rs1
80
50
09
3.7
8(2
.43
–5
.87
)3
.27
31
02
90
.79
(0.4
9–
1.2
7)
0.3
42
.00
(0.7
3–
5.4
6)
0.1
72
.17
(1.4
1–
3.3
3)
0.0
00
40
.88
(0.6
0–
1.3
1)
0.5
42
.98
(1.9
9–
4.4
6)
1.1
03
10
27
,1
MC
1R
rs1
15
47
46
42
.69
(2.0
6–
3.5
1)
3.4
43
10
21
3,1
0.9
3(0
.71
–1
.23
)0
.63
1.1
4(0
.69
–1
.88
)0
.62
2.0
0(1
.54
–2
.60
)2
.66
31
02
7,1
0.8
7(0
.68
–1
.11
)0
.27
3.1
5(2
.44
–4
.08
)2
.75
31
02
18
1
MIT
Frs
76
41
32
01
.08
(0.9
3–
1.2
4)
0.3
00
.90
(0.7
6–
1.0
6)
0.2
00
.95
(0.7
2–
1.2
4)
0.6
91
.01
(0.8
8–
1.1
7)
0.8
40
.83
(0.7
2–
0.9
6)
0.0
11
.04
(0.8
9–
1.2
2)
0.5
8
MIT
Frs
67
81
17
11
.19
(1.0
3–
1.3
6)
0.0
10
.92
(0.7
9–
1.0
8)
0.,
32
1.0
8(0
.83
–1
.40
)0
.58
1.0
3(0
.90
–1
.18
)0
.66
0.7
9(0
.69
–0
.91
)0
.00
11
.16
(1.0
0–
1.3
5)
0.0
5
MY
O5
Ars
17
54
20
17
1.0
2(0
.89
–1
.17
)0
.80
1.0
1(0
.86
–1
.18
)0
.89
1.0
9(0
.83
–1
.41
)0
.54
0.8
7(0
.76
–1
.00
)0
.05
1.2
0(1
.04
–1
.37
)0
.01
1.0
4(0
.90
–1
.21
)0
.57
OC
A2
rs1
80
04
07
1.1
5(0
.90
–1
.46
)0
.26
1.4
9(1
.15
–1
.93
)0
.00
30
.95
(2.8
8–
1.6
5)
0.0
80
.89
(0.7
0–
1.1
2)
0.3
21
.40
(1.1
0–
1.7
8)
0.0
10
.67
(0.5
0–
0.8
8)
0.0
05
SLC
24
A4
rs1
28
96
39
91
.04
(0.9
0–
1.1
9)
0.6
11
.01
(0.8
7–
1.1
8)
0.8
71
.23
(0.9
4–
1.6
0)
0.1
31
.33
(1.1
6–
1.5
2)
3.8
43
10
25
,10
.87
(0.7
6–
0.9
9)
0.0
41
.07
(0.9
2–
1.2
3)
0.4
0
SLC
24
A5
rs2
67
53
46
0.0
9(0
.01
–0
.73
)0
.02
0.6
0(0
.13
–2
.73
)0
.51
0.0
7(0
.03
–0
.21
)1
.69
31
02
6,1
0.2
2(0
.06
–0
.80
)0
.02
4.8
2(1
.33
–1
7.5
3)
0.0
20
.19
(0.0
2–
1.4
9)
0.1
1
SLC
45
A2
rs1
68
91
98
20
.29
(0.2
1–
0.4
1)
5.3
33
10
21
3,1
0.5
8(0
.41
–0
.82
)0
.00
20
.17
(0.1
3–
0.2
4)
2.8
23
10
22
6,1
0.3
8(0
.29
–0
.50
)1
.48
31
02
12
12
.01
(1.5
5–
2.6
0)
1.5
43
10
20
7,1
0.4
9(0
.35
–0
.69
)4
.05
31
02
5,1
SLC
45
A2
rs6
86
35
60
––
0.5
4(0
.07
–4
.47
)0
.57
0.0
5(0
.01
–0
.21
)4
.36
31
02
5,1
0.3
1(0
.06
–1
.53
)0
.15
1.7
5(0
.41
–7
.42
)0
.45
––
SLC
7A
11
rs4
56
04
61
1.1
0(0
.88
–1
.37
)0
.39
1.0
3(0
.81
–1
.32
)0
.81
0.9
1(0
.61
–1
.36
)0
.66
0.9
4(0
.76
–1
.17
)0
.59
0.7
4(0
.59
–0
.93
)0
.01
1.0
8(0
.85
–1
.36
)0
.54
TFA
P2
ArS
16
21
70
01
.02
(0.8
6–
1.2
0)
0.8
41
.34
(1.1
1–
1.6
1)
0.0
02
1.6
3(1
.13
–2
.37
)0
.01
1.0
1(0
.85
–1
.19
)0
.94
0.9
0(0
.76
–1
.07
)0
.22
1.0
1(0
.85
–1
.21
)0
.89
TPC
N2
rs1
07
50
83
60
.19
(0.0
9–
0.4
1)
3.1
03
10
25
,10
.56
(0.2
7–
1.1
5)
0.1
10
.19
(0.1
1–
0.3
3)
1.0
93
10
28
,10
.41
(0.2
3–
0.7
0)
0.0
01
0.3
8(0
.22
–0
.66
)0
.00
10
.45
(0.2
1–
0.9
4)
0.0
3
TYR
rs1
12
68
09
1.5
0(1
.29
–1
.75
)9
.32
31
02
8,1
1.2
3(1
.04
–1
.45
)0
.02
1.3
6(1
.00
–1
.84
)0
.05
1.0
7(0
.92
–1
.24
)0
.37
0.8
4(0
.72
–0
.98
)0
.02
1.0
8(0
.92
–1
.26
)0
.36
TYR
P1
rs2
73
38
32
0.8
8(0
.77
–1
.01
)0
.06
0.8
6(0
.74
–1
.00
)0
.06
0.7
1(0
.55
–0
.92
)0
.01
0.8
3(0
.72
–0
.94
)0
.00
51
.27
(1.1
1–
1.4
5)
6.3
03
10
–4
1.0
6(0
.92
–1
.23
)0
.40
TYR
P1
rs1
40
87
99
0.8
6(0
.75
–0
.99
)0
.04
0.9
7(0
.83
–1
.14
)0
.73
0.8
0(0
.62
–1
.05
)0
.10
0.8
8(0
.76
–1
.00
)0
.06
1.2
6(1
.09
–1
.45
)0
.00
20
.99
(0.8
5–
1.1
5)
0.8
8
WR
Nrs
18
00
39
10
.93
(0.7
3–
1.2
0)
0.6
01
.15
(0.8
7–
1.5
3)
0.3
31
.26
(0.7
4–
2.1
5)
0.3
91
.30
(1.0
1–
1.6
7)
0.0
40
.69
(0.5
3–
0.8
9)
0.0
10
.94
(0.7
1–
1.2
3)
0.6
4
Bo
ldin
dic
ate
sst
ati
stic
all
ysi
gn
ifica
nt
resu
lts.
CI,
con
fid
en
cein
terv
al;
OR
,o
dd
sra
tio
.1In
dic
ate
sst
ati
stic
all
ysi
gn
ifica
nt
resu
lts
aft
er
Bo
nfe
rro
ni
corr
ect
ion
.

and rs1126809, which encodes the amino-acid changeArg402Gln in the TYR gene, with and overall OR5 1.15 (95%CI: 1.07–1.24, p5 2.523 1024) (Supporting Information Table3, Supporting Information Table 4 and Figure 2). SNPs locatedin SLC45A2, VPS41, WNT3 and TYR remained significant aftermultiple testing corrections.
Evidence of new associations with pigmentation character-istics for some of the 335 SNPs analysed in the discoveryphase was also observed (see Table 2).
Exome sequencing: T52R VPS41 variant
Exome sequencing was performed in six patients from threedifferent French melanoma families. A rare nonsynonymousT52R variant (rs147210031), located in the VPS41 gene,appeared in the two individuals sequenced of the first family.Therefore, six additional individuals were genotyped for thismutation in the family; five of them were carriers of the vari-ant. Thus, nonsynonymous T52R variant appeared to partlysegregate with the disease in the first family (Figure 3). Inter-estingly, this variant has a low frequency in the Caucasianpopulation (MAF5 0.005), and was predicted to be damag-ing by PolyPhen and SIFT algorithms.
We genotyped the T52R variant in DNA from 178 Frenchfamilial melanoma probands stored in the Genetic Departmentof Bichat Hospital, under the French MelanCohort and in the
Spanish Phase II population (in total, 1633 controls and 1882sporadic melanoma cases). Minor allele frequencies for controlsubjects, familial MM and sporadic MM cases were 0.006,0.028 and 0.007, respectively. A significant association withmelanoma predisposition was clearly observed for familialcases with an OR of 4.46 (CI 95%: 1.75–11.26; p5 13 1023,Table 2). Of these, two melanoma families (each one compris-ing two melanoma patients) were further been investigated forT52R variant segregation, and all melanoma patients carriedthe VPS41 variant. Although no association was reported forthe French and Spanish sporadic cases, when the T52R variantdata from 1000 genomes Caucasian control (N5 7237) wasevaluated against the same sporadic sample set, the initialtrend observed was significant. Minor allele frequency in 1000genomes database was 0.004, and a lesser, but still significantassociation with sporadic melanoma was observed (OR5 1.68;CI 95%: 1.03–2.72; p5 0.032) (Table 3).
DiscussionIn this work, by using a customized association approachcoupled to an exome sequencing study, we were able to iden-tify two new pathways involved in MM susceptibility.
An interesting finding was the relatively strong associationof the WNT3 gene with melanoma. The human WNT3 gene(MIM#165330) is located in chromosome 17q21 and has
Figure 3. Pedigree. Five generation family tree shows the segregation of the T52R variant of the VPS41 gene and the clinical manifestations
of melanoma affected individuals. Squares indicate males and circles indicate females. In black solid symbols are represented melanoma
patients. In grey solid symbols are represented other non-melanoma cancer patients. Symbols with a slash indicate deceased individuals.
An arrow points to the proband.
Can
cerGenetics
Ibarrola-Villava et al. 7
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

been shown to play a crucial role in the WNT pathway.WNT genes encode a family of 19 secreted glycoproteins thatact as ligands to activate receptor-mediated signalling path-ways that control cell fate and differentiation, cell prolifera-tion, and cell motility.23 Aberrations in these pathways havebeen implicated in a number of chronic diseases, such ascancer. Specifically, mutations leading to a constitutive activa-tion of WNT/b-catenin signalling are observed in colorectalcancer and in some kidney tumours, where activation of thepathway has been directly implicated in disease pathogene-sis.24–26 The WNT pathway is also known to play a crucialrole in melanoma oncogenesis and progression.27,28 Wnt/b-catenin signalling regulates mouse pigmentation duringdevelopment by determining the number of melanoblasts,
the precursor cells of melanocytes.29 In adults, high levels ofb-catenin are expressed in the cytoplasm and nucleus ofbenign nevi,30 which are spatially associated with approxi-mately 20% of cutaneous melanomas and are considered tobe direct melanoma precursors. Interestingly, WNT3 is highlyhomologous to the WNT3A gene that has been shown toplay an important role in differentiation of melanoma cells.31
The melanoma-associated SNP rs199524, located in a con-served region within intron 2, was selected as the tag for oneof the three most frequent haplotypes of this gene. In addi-tion, this SNP is in strong linkage disequilibrium with fourother SNPs located in WNT3 and nearby the NSF gene,which are predicted to affect transcription factor binding sites(rs199452, rs199453, rs199454 and rs199455). rs199524
Table 3. Genotypic frequencies across VPS41 T52R polymorphism and assessment of individual association with malignant melanoma
T52R variant genotyping across French population. Association with familial melanoma
France Controls (N 5 911) N Familial cases (N 5 178) N OR (95% CI) p value
G/G 899 G/G 169
G/C 12 G/C 8
C/C – C/C 1
MAF 0.006 MAF 0.028 4.46 (1.75–11.26) 0.001
T52R variant genotyping across French and Spanish population. Association with sporadic melanoma
France Controls (N 5 911) N Cases (N 5 821) N OR (95% CI) p value
G/G 899 G/G 808
G/C 12 G/C 13
C/C – C/C –
MAF 0.006 MAF 0.008 1.20 (0.51–2.83) 0.69
Spain Controls (N 5 722) N Cases (N 5 1061) N OR (95% CI) p value
G/G 715 G/G 1048
G/C 7 G/C 13
C/C – C/C –
MAF 0.005 MAF 0.006 1.26 (0.47–3.51) 0.66
Total Controls (N 5 1633) N Cases (N 5 1882) N OR (95% CI) p value
G/G 1614 G/G 1856
G/C 19 G/C 26
C/C – C/C –
MAF 0.006 MAF 0.007 1.19 (0.63–2.24) 0.65
T52R variant genotyping across Caucasian population and controls included in the 1000 genomes project. Association with sporadicmelanoma
Caucasian population Controls (N 5 7237)1 N Cases (N 5 1882) N OR (95% CI) p value
G/G 7177 G/G 1856
G/C 60 G/C 26
C/C – C/C –
MAF 0.004 MAF 0.007 1.68 (1.03–2.72) 0.032
Bold indicates significant results.p-value obtained using Fisher exact test.OR, Odds Ratio; CI, Confidence Interval1Controls used at this stage were from the 1000 genomes project
Can
cerGenetics
8 VPS41 and WNT3 and melanoma susceptibility
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC
represents an extended haplotype including the WNT3 gene,except exon 1, and the entire NSF gene located at the 5’ ofthe WNT3 gene. Remarkably, NSF is involved in vesiculartrafficking and glutamate pathway.
Population stratification was unlikely to explain the asso-ciation between the WNT3 rs199524 SNP and melanomarisk. In European populations, the MAF of this rs199524SNP is highly variable with a gradient increasing from theNorth to the South of Europe; 0.15 in Finnish controls, 0.25in UK controls and 0.32 in Iberian controls (data from the1000 genomes project, www.browser.1000genomes.org).The MAF values in our controls from Germany, France andSpain are completely coherent with these values (0.26, 0.28and 0.33, respectively). In addition, all enrolled cases andcontrols in the four cohorts were from Caucasian origin. Fur-thermore, the risk associated with this variant is in the sameorder of magnitude in all the phases of the study. Finally, ameta-analysis was performed, after adjustment for each pop-ulation, by a Cochran–Mantel–Haenszel test,19 as previouslydescribed.32 All these reasons stress the association betweenthe WNT3 rs199524 SNP and melanoma risk.
Therefore, the WNT pathway, and especially WNT3,appeared as an attractive candidate pathway that might be ofinterest to other pigmentation and melanoma studies.
Eukaryotic cells are divided by membrane-enclosed organ-elles where there is complicated traffic of molecules by vesic-ular transport. In this transport, intracellular molecules moveback and forth via membrane enclosed vesicles along theorganelles, including the endoplasmic reticulum, Golgi appa-ratus, endosome, secretory vesicle, and lysosome.33,34 Recentexome-wide studies have highlighted different rare variantsthat contribute to the inheritable risk for some monogenicdiseases.35,36 Using this approach, we could identify a nonsy-nonymous variant T52R, located in at a highly conserved res-idue in the VPS41 gene that strongly associates with familialmelanoma (p5 0.001), and segregates in three differentFrench melanoma families. In addition, this variant mightalso play a role in sporadic cases. Thus, the T52R variantmight be an intermediate penetrant risk allele, and the VPS41gene appears as a new susceptibility gene in melanoma. Con-sistent with this finding, similar results were detected for theE318K MITF variant, first detected in a whole-genomesequencing of probands from several melanoma families andlater significantly associated with sporadic melanoma cases.37
Functional studies are welcome to establish the exact conse-quences of this new melanoma predisposing variant.
VPS genes encode proteins involved in vesicular trans-port.38 Although some VPS proteins are involved in cancer-
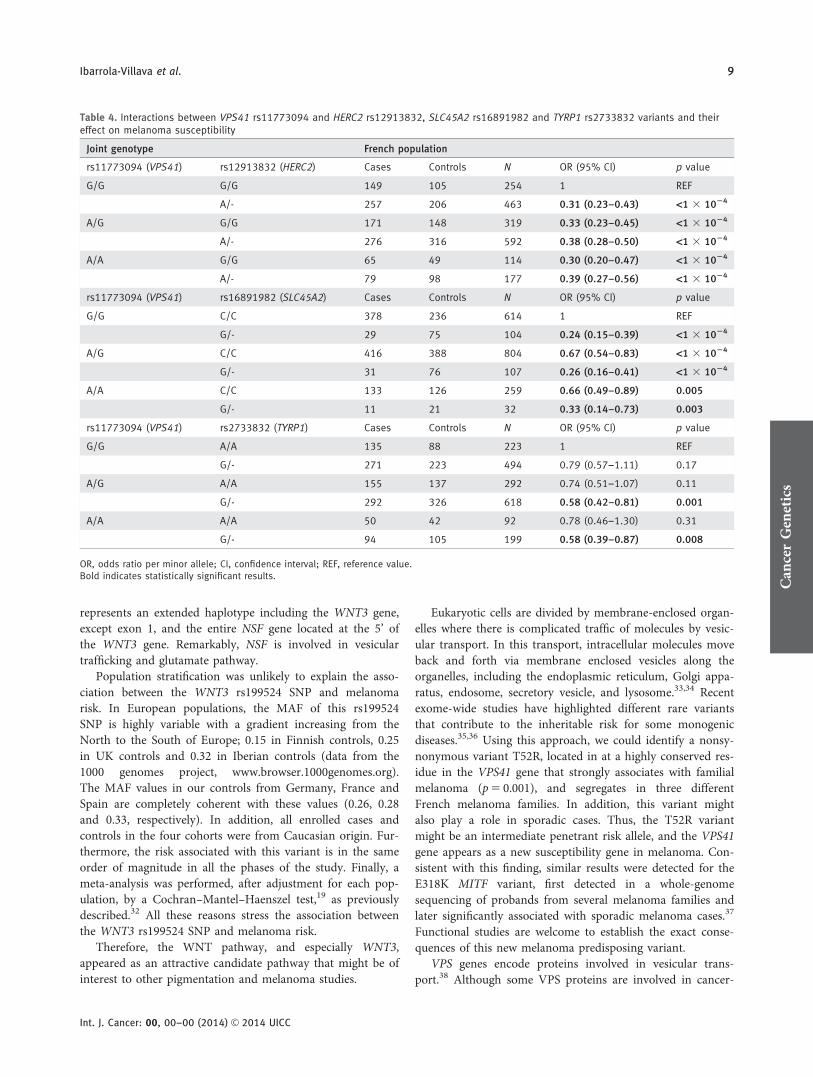
Table 4. Interactions between VPS41 rs11773094 and HERC2 rs12913832, SLC45A2 rs16891982 and TYRP1 rs2733832 variants and theireffect on melanoma susceptibility
Joint genotype French population
rs11773094 (VPS41) rs12913832 (HERC2) Cases Controls N OR (95% CI) p value
G/G G/G 149 105 254 1 REF
A/- 257 206 463 0.31 (0.23–0.43) <1 3 1024
A/G G/G 171 148 319 0.33 (0.23–0.45) <1 3 1024
A/- 276 316 592 0.38 (0.28–0.50) <1 3 1024
A/A G/G 65 49 114 0.30 (0.20–0.47) <1 3 1024
A/- 79 98 177 0.39 (0.27–0.56) <1 3 1024
rs11773094 (VPS41) rs16891982 (SLC45A2) Cases Controls N OR (95% CI) p value
G/G C/C 378 236 614 1 REF
G/- 29 75 104 0.24 (0.15–0.39) <1 3 1024
A/G C/C 416 388 804 0.67 (0.54–0.83) <1 3 1024
G/- 31 76 107 0.26 (0.16–0.41) <1 3 1024
A/A C/C 133 126 259 0.66 (0.49–0.89) 0.005
G/- 11 21 32 0.33 (0.14–0.73) 0.003
rs11773094 (VPS41) rs2733832 (TYRP1) Cases Controls N OR (95% CI) p value
G/G A/A 135 88 223 1 REF
G/- 271 223 494 0.79 (0.57–1.11) 0.17
A/G A/A 155 137 292 0.74 (0.51–1.07) 0.11
G/- 292 326 618 0.58 (0.42–0.81) 0.001
A/A A/A 50 42 92 0.78 (0.46–1.30) 0.31
G/- 94 105 199 0.58 (0.39–0.87) 0.008
OR, odds ratio per minor allele; CI, confidence interval; REF, reference value.Bold indicates statistically significant results.
Can
cerGenetics
Ibarrola-Villava et al. 9
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

related cellular mechanisms, alterations of human VPS genesin cancer are largely unknown.39 The human VPS41 gene(MIM#605485) is located in chromosome 7p14.1, and itsencoded protein is required for vacuolar assembly and vacuo-lar traffic from the Golgi. Interestingly, mutations of the flyhomologue vps41 are responsible for the light phenotype inDrosophila melanogaster, suggesting its role in pigmentation,which could also explain the melanoma predisposition.40 Fur-thermore, the melanoma-associated SNP rs11773094 is locatedin 3’UTR and is predicted to disrupt a binding site of twomicroRNAs (hsa-miR-196a and hsa-miR-9) that have beenpreviously implicated in cancer susceptibility.41 It looks verypromising that this new candidate gene to melanoma suscepti-bility is implicated in functions related to vesicle transportationand has been demonstrated to affect pigmentation.
Since the VPS41 and HERC2, SLC45A2 and TYRP1 genesfunctions have been related to vacuolar traffic and melanoma,it seemed biologically plausible that genetic interactionswould be detected among these gene variants. Putative inter-action effects could disrupt the downstream pathway. Addi-tive protective effects were observed when summing up rarealleles of VPS41 and HERC2, SLC45A2 and TYRP1 separately(Table 4). These results reinforce the role of the vesiculartrafficking pathway in melanoma.
We cannot exclude that there was potential for misclassifi-cation of phenotypic characteristics because of the subjectivenature of the attributes considered. The retrospective natureof the study raises the potential for recall bias, particularly
for exposures such as childhood sunburn, number of solarlentigines and number of nevi. Other published studies havebeen very similar in terms of data collection and althoughthere are some differences between the populations includedin the study our results are concordant with those obtainedin the other Mediterranean populations.
In summary, we conducted a three-stage melanoma suscep-tibility case-control study (candidate SNP selection and twophases of replication). First we screened 335 SNPs in a largeFrench population. Then we validated the observed associa-tions in an independent Spanish and German series. One SNP(WNT3 rs199524) was protectively associated with melanomain each phase indicating that a common variation in WNT3 isassociated with protection from the disease. In addition, oneSNP (VPS41 rs11773094) was protectively associated with mel-anoma when all phases were considered together. Moreover,the implementation of whole-exome sequencing technologiesin diseases with genetic basis allows for the identification ofmutations in unknown candidate genes, as in our study, wherethe role of VPS41 has been highlighted.
AcknowledgementsThis work was supported by grants from La Societe Francaise de Der-matologie (SFD) and Ministerio de Salud Carlos III [PI10–0405,CP08-00069] to G.R. and co-funded with FEDER. The authors thankthe Madrid College of Lawyers and the patients from the participatinghospitals. They also thank the participants of URC (D�epartement deRecherche Clinique) of Ambroise Par�e Hospital. The authors declareno conflicts of interest.
References
1. Brown KM, Macgregor S, Montgomery GW,et al. Common sequence variants on 20q11.22confer melanoma susceptibility. Nat Genet 2008;40:838–40.
2. Han J, Kraft P, Nan H, et al. A genome-wideassociation study identifies novel alleles associatedwith hair color and skin pigmentation. PLoSGenet 2008;4:e1000074.
3. Sulem P, Gudbjartsson DF, Stacey SN, et al.Genetic determinants of hair, eye and skin pig-mentation in Europeans. Nat Genet 2007;39:1443–52.
4. Bataille V. Genetic epidemiology of melanoma.Eur J Cancer 2003;39:1341–7.
5. Horn S, Figl A, Rachakonda PS, et al. TERT pro-moter mutations in familial and sporadic mela-noma. Science 2013;339:959–61.
6. Shi J, Yang XR, Ballew B, et al. Rare missensevariants in POT1 predispose to familial cutaneousmalignant melanoma. Nat Genet 2014;46:482–6.
7. Fernandez L, Milne R, Bravo J, et al. MC1R:three novel variants identified in a malignantmelanoma association study in the Spanish popu-lation. Carcinogenesis 2007;28:1659–64.
8. Mohrenweiser HW, Wilson DM, 3rd, Jones IM.Challenges and complexities in estimating boththe functional impact and the disease risk associ-ated with the extensive genetic variation inhuman DNA repair genes. Mutat Res 2003;526:93–125.
9. Ibarrola-Villava M, Pena-Chilet M, FernandezLP, et al. Genetic polymorphisms in DNA repair
and oxidative stress pathways associated withmalignant melanoma susceptibility. Eur J Cancer2011;47:2618–25.
10. Ibarrola-Villava M, Martin-Gonzalez M, LazaroP, et al. Role of glutathione S-transferases in mel-anoma susceptibility: association with GSTP1rs1695 polymorphism. Br J Dermatol 2012;166:1176–83.
11. Ibarrola-Villava M, Fernandez LP, Alonso S, et al.A customized pigmentation SNP array identifiesa novel SNP associated with melanoma predispo-sition in the SLC45A2 gene. PLoS One 2011;6:e19271.
12. Pena-Chilet M, Ibarrola-Villava M, Martin-Gonzalez M, et al. rs12512631 on the group spe-cific complement (vitamin D-binding proteinGC) implicated in melanoma susceptibility. PLoSOne 2013;8:e59607.
13. Lin JY, Fisher DE. Melanocyte biology and skinpigmentation. Nature 2007;445:843–50.
14. Miller AJ, Mihm MC, Jr. Melanoma. N Engl JMed 2006;355:51–65.
15. Gerstenblith MR, Goldstein AM, Fargnoli MC,et al. Comprehensive evaluation of allele fre-quency differences of MC1R variants acrosspopulations. Hum Mutat 2007;28:495–505.
16. Fernandez LP, Milne RL, Pita G, et al. SLC45A2:a novel malignant melanoma-associated gene.Hum Mutat 2008;29:1161–7.
17. Gudbjartsson DF, Sulem P, Stacey SN, et al. ASIPand TYR pigmentation variants associate with
cutaneous melanoma and basal cell carcinoma.Nat Genet 2008;40:886–91.
18. Guedj M, Bourillon A, Combadieres C, et al. Var-iants of the MATP/SLC45A2 gene are protectivefor melanoma in the French population. HumMutat 2008;29:1154–60.
19. Mantel N, Haenszel W. Statistical aspects of theanalysis of data from retrospective studies of dis-ease. J Natl Cancer Inst 1959;22:719–48.
20. Di Lucca J, Guedj M, Lacapere JJ, et al. Variantsof the xeroderma pigmentosum variant gene(POLH) are associated with melanoma risk. Eur JCancer 2009;45:3228–36.
21. Esteras C, Formisano G, Roig C, et al. SNP geno-typing in melons: genetic variation, populationstructure, and linkage disequilibrium. Theor ApplGenet 2013;126:1285–303.
22. Wong MM, Cannon CH, Wickneswari R. Devel-opment of high-throughput SNP-based genotyp-ing in Acacia auriculiformis x A. mangiumhybrids using short-read transcriptome data.BMC Genomics 2012;13:726.
23. Chien AJ, Moon RT. WNTS and WNT receptorsas therapeutic tools and targets in human diseaseprocesses. Front Biosci 2007;12:448–57.
24. Koesters R, Ridder R, Kopp-Schneider A, et al.Mutational activation of the beta-catenin proto-oncogene is a common event in the developmentof Wilms’ tumors. Cancer Res 1999;59:3880–2.
25. Giles RH, van Es JH, Clevers H. Caught up in aWnt storm: Wnt signaling in cancer. BiochimBiophys Acta 2003;1653:1–24.
Can
cerGenetics
10 VPS41 and WNT3 and melanoma susceptibility
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC

26. Major MB, Camp ND, Berndt JD, et al. Wilmstumor suppressor WTX negatively regulates WNT/beta-catenin signaling. Science 2007;316:1043–6.
27. O’Connell MP, Weeraratna AT. Hear the WntRor: how melanoma cells adjust to changes inWnt. Pigment Cell Melanoma Res 2009;22:724–39.
28. Weeraratna AT. A Wnt-er wonderland–the com-plexity of Wnt signaling in melanoma. CancerMetastasis Rev 2005;24:237–50.
29. Larue L, Luciani F, Kumasaka M, et al. Bypassingmelanocyte senescence by beta-catenin: a novelway to promote melanoma. Pathol Biol (Paris)2009;57:543–7.
30. Bachmann IM, Straume O, Puntervoll HE, et al.Importance of P-cadherin, beta-catenin, andWnt5a/frizzled for progression of melanocytictumors and prognosis in cutaneous melanoma.Clin Cancer Res 2005;11:8606–14.
31. Chien AJ, Moore EC, Lonsdorf AS, et al. Acti-vated Wnt/beta-catenin signaling in melanoma isassociated with decreased proliferation in patient
tumors and a murine melanoma model. ProcNatl Acad Sci USA 2009;106:1193–8.
32. Ibarrola-Villava M, Hu HH, Guedj M, et al.MC1R, SLC45A2 and TYR genetic variantsinvolved in melanoma susceptibility in southernEuropean populations: results from a meta-analy-sis. Eur J Cancer 2012;48:2183–91.
33. Klionsky DJ. Protein transport from the cyto-plasm into the vacuole. J Membr Biol 1997;157:105–15.
34. Mellman I, Warren G. The road taken: past andfuture foundations of membrane traffic. Cell2000;100:99–112.
35. Bilguvar K, Ozturk AK, Louvi A, et al. Whole-exome sequencing identifies recessive WDR62mutations in severe brain malformations. Nature2010;467:207–10.
36. Hoischen A, Gilissen C, Arts P, et al.Massively parallel sequencing of ataxia genes afterarray-based enrichment. Hum Mutat 2010;31:494–9.
37. Yokoyama S, Woods SL, Boyle GM, et al. A novelrecurrent mutation in MITF predisposes to fami-lial and sporadic melanoma. Nature 2011;480:99–103.
38. Wickner W, Haas A. Yeast homotypic vacuolefusion: a window on organelle traffickingmechanisms. Annu Rev Biochem 2000;69:247–75.
39. Rusten TE, Filimonenko M, Rodahl LM, et al.ESCRTing autophagic clearance of aggregatingproteins. Autophagy 2008;4.
40. Warner TS, Sinclair DA, Fitzpatrick KA, et al.The light gene of Drosophila melanogaster enco-des a homologue of VPS41, a yeast gene involvedin cellular-protein trafficking. Genome 1998;41:236–43.
41. Ben-Hamo R, Efroni S. Gene expression andnetwork-based analysis reveals a novel role forhsa-miR-9 and drug control over the p38 net-work in glioblastoma multiforme progression.Genome Med 2011;3:77.
Can
cerGenetics
Ibarrola-Villava et al. 11
Int. J. Cancer: 00, 00–00 (2014) VC 2014 UICC
Copyright © 2022 FDOKUMEN




















