
Gel and gel-free proteomics to identify Saccharomyces cerevisiae cell surface proteins
13
Gel and gel-free proteomics to identify Saccharomyces cerevisiae cell surface proteins María Rosa Insenser a , María Luisa Hernáez b , César Nombela a , María Molina a , Gloria Molero a, ⁎ , Concha Gil a,b a Departamento de Microbiología II, Facultad de Farmacia, Universidad Complutense de Madrid, Plaza de Ramón y Cajal s/n, 28040 Madrid, Spain b Unidad de Proteómica UCM-Parque Científico de Madrid, Facultad de Farmacia, Universidad Complutense de Madrid, Plaza de Ramón y Cajal s/n, 28040 Madrid, Spain ARTICLE INFO ABSTRACT The study of Saccharomyces cerevisiae cell surface proteins was performed because of their important role in cell wall biogenesis and in the physiology of the yeast. Two different proteomic approaches were carried out. First, proteins loosely associated or S–S linked to structural wall components were released by treatment of whole intact cells with dithiothreitol, separated by 2D-PAGE and identified by mass spectrometry. Second, cell surface-exposed proteins (surfome) were digested with trypsin and DTT from whole intact cells, and analyzed by LC–MS/MS. Ninety-nine different proteins were identified: 67 with DTT treatment and 52 with DTT and trypsin digestion. These proteins were classified in different cellular processes: control of cell wall organization, cell rescue, defence, and virulence, protein fate, protein synthesis and metabolism. Most of the proteins have already been reported as present on the cell surface showing that the yeast cell surface is composed not only by typical but also by atypical cell wall proteins. “Bona fide” cell wall proteins were identified by both protocols but a higher number with the non-gel strategy. However, only 20% of the proteins identified were common to both protocols, thus, for a complete knowledge of the cell surface proteome, several strategies have to be used. © 2010 Elsevier B.V. All rights reserved. Keywords: Saccharomyces cerevisiae Cell surface Surfome Gel proteomics Non-gel proteomics 1. Introduction The fungal cell wall is a dynamic structure that protects the cell from osmotic and physical stresses. It is also involved in interactions with other cells and with the environment. The cell wall structure and organization of Saccharomyces cerevisiae has been extensively studied and it has served as a model for other fungi [1–3]. The cell wall exhibits a dynamic nature, undergoing profound changes during various cellular process- es, namely budding, mating and dimorphic transition [1,4]. The cell wall has two layers: an internal layer, which consists of a network of branched β-1,3-glucan molecules extended with covalently attached β-1,6-glucan and chitin molecules, and a fibrillar outer layer, which is mainly composed of a varied set of mannoproteins that collectively form yeast mannan [5]. Cell wall proteins are crucial components of this structure and are important for the physiology of the yeast. They participate in mating, colony morphology changes, biofilm formation, fruiting body development, and interactions with mammalian and plant hosts [6]. The composition of the cell wall proteome is highly dynamic, and depends on growth ⁎ Corresponding author. Departamento de Microbiología II, Facultad de Farmacia, Universidad Complutense, Plaza de Ramón y Cajal s/n, 28040-Madrid, Spain. Tel.: +34913941744; fax: +34 913941745. E-mail address: [email protected] (G. Molero). JOURNAL OF PROTEOMICS 73 (2010) 1183 – 1195 1874-3919/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.jprot.2010.02.005 available at www.sciencedirect.com www.elsevier.com/locate/jprot
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Gel and gel-free proteomics to identify Saccharomyces cerevisiae cell surface proteins

J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te / j p ro t
Gel and gel-free proteomics to identify Saccharomyces cerevisiaecell surface proteins
María Rosa Insensera, María Luisa Hernáezb, César Nombelaa, María Molinaa,Gloria Moleroa,⁎, Concha Gila,b
aDepartamento deMicrobiología II, Facultad de Farmacia, Universidad Complutense de Madrid, Plaza de Ramón y Cajal s/n, 28040Madrid, SpainbUnidad de Proteómica UCM-Parque Científico de Madrid, Facultad de Farmacia, Universidad Complutense de Madrid,Plaza de Ramón y Cajal s/n, 28040 Madrid, Spain
A R T I C L E I N F O
⁎ Corresponding author. Departamento de Mi28040-Madrid, Spain. Tel.: +34913941744; fax
E-mail address: [email protected] (G. M
1874-3919/$ – see front matter © 2010 Elsevidoi:10.1016/j.jprot.2010.02.005
A B S T R A C T
Keywords:
The study of Saccharomyces cerevisiae cell surface proteins was performed because of theirimportant role in cell wall biogenesis and in the physiology of the yeast. Two differentproteomic approaches were carried out. First, proteins loosely associated or S–S linked tostructural wall components were released by treatment of whole intact cells withdithiothreitol, separated by 2D-PAGE and identified by mass spectrometry. Second, cellsurface-exposed proteins (surfome) were digested with trypsin and DTT from whole intactcells, and analyzed by LC–MS/MS. Ninety-nine different proteins were identified: 67 withDTT treatment and 52 with DTT and trypsin digestion. These proteins were classified indifferent cellular processes: control of cell wall organization, cell rescue, defence, andvirulence, protein fate, protein synthesis andmetabolism. Most of the proteins have alreadybeen reported as present on the cell surface showing that the yeast cell surface is composednot only by typical but also by atypical cell wall proteins. “Bona fide” cell wall proteins wereidentified by both protocols but a higher number with the non-gel strategy. However, only20% of the proteins identified were common to both protocols, thus, for a completeknowledge of the cell surface proteome, several strategies have to be used.© 2010 Elsevier B.V. All rights reserved.
Saccharomyces cerevisiaeCell surfaceSurfomeGel proteomicsNon-gel proteomics
1. Introduction
The fungal cell wall is a dynamic structure that protects thecell from osmotic and physical stresses. It is also involved ininteractions with other cells and with the environment. Thecell wall structure and organization of Saccharomyces cerevisiaehas been extensively studied and it has served as a model forother fungi [1–3]. The cell wall exhibits a dynamic nature,undergoing profound changes during various cellular process-es, namely budding, mating and dimorphic transition [1,4].The cell wall has two layers: an internal layer, which consists
crobiología II, Facultad de: +34 913941745.olero).
er B.V. All rights reserved
of a network of branched β-1,3-glucan molecules extendedwith covalently attached β-1,6-glucan and chitin molecules,and a fibrillar outer layer, which is mainly composed of avaried set of mannoproteins that collectively form yeastmannan [5].
Cell wall proteins are crucial components of this structureand are important for the physiology of the yeast. Theyparticipate in mating, colony morphology changes, biofilmformation, fruiting body development, and interactions withmammalian and plant hosts [6]. The composition of the cellwall proteome is highly dynamic, and depends on growth
Farmacia, Universidad Complutense, Plaza de Ramón y Cajal s/n,
.

1184 J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
conditions such as nutrient availability, temperature, externalpH, and oxygen level. Furthermore, the expression andincorporation of proteins into the cell wall are temporallyand spatially controlled [7].
There are three main classes of cell wall proteins (CWPs),two of them covalently coupled to cell wall polysaccharides:Glycosyl-phosphatidylinositol cell wall proteins (GPI-CWPs),which are generally indirectly linked to β-1,3-glucan through aconnecting β-1,6-glucan moiety, and Pir proteins (Pir-CWPs),which are presumed to be directly linked to β-1,3-glucanthrough the γ-carboxyl group of glutamic acids, arising fromspecific glutamines [8].
The third class of proteins lacks the covalent attachment tothe polysaccharide matrix; these cell wall proteins can bereleased from intact cells using a reducing agent. Thissuggests that they might be disulphide-linked to other cellwall proteins [9].
Several chemical and/or enzymatic strategies for isolationof the CWPs have been described, both fromwhole intact cellsand from isolated cell walls after cell breakage (reviewed in[2]). The classic approach to their study is based on cell wallfractionation, protein separation by polyacrylamide gel elec-trophoresis (SDS-PAGE), and identification of proteins byN-terminal sequencing [10]. Sodium dodecylsulphate andreducing agents, such as mercaptoethanol or dithiothreitol,allow the extraction of mannoproteins that are proteinsloosely associated or S–S linked to structural wall com-ponents. Classical proteomics analyses involve the simul-taneous separation, visualization and quantification ofthousands of proteins by two-dimensional gel electrophore-sis (2D-PAGE) and their subsequent identification and char-acterization by mass spectrometry (MS). In previous studiesin C. albicans we have shown the utility of 2D-PAGE of yeastprotein preparations for the study of the cell wall proteome inorder to identify potential targets for new antifungal drugs[11] or immunogenic proteins [12]. Alternatively, anotherapproach for studying CWPs involves the analysis of proteinssecreted into the medium by regenerating cell wall proto-plasts [13].
Another strategy aimed at improving the detection of lowcopy-number proteins of functional interest and/or the resolu-tionefficacyofhydrophobicproteins, involves chromatographictechnology. Liquid chromatography (LC)-based technologieshave several advantages compared with 2D-gel-based techni-ques. LC–MS, which can be automated, combines high-speed,high-resolution and high-sensitivity separation of extremelycomplex peptide mixtures [11,14], allowing the relative quanti-fication of the proteins present on the cell surface [15]. Thus,using a combination of 2D-PAGE and LC to identify cell wallproteins can lead to a more complete understanding of the cellwall complexity at the protein level.
In the present study, two different protocols were used.First, proteins that were loosely associated or S–S linked tostructural wall components were released by treatment of S.cerevisiaewhole intact cells with a reducing agent, dithiothrei-tol (DTT), separated by 2D-PAGE and identified by massspectrometry. Second, cell surface-exposed proteins (sur-fome), covalently linked or not to polysaccharides or to otherproteins, were digested with trypsin and DTT from wholeintact cells, and analyzed by LC–MS/MS.
2. Material and methods
2.1. Strains and media
The S. cerevisiae strain used in this study was BY4741 (MATa;his3Δ1; leu2Δ0; met15Δ0; ura3Δ0). This strain was maintainedon YPD agar (2% D-glucose, 1% Difco yeast extract, 2% peptoneand 2% agar) and incubated at 24 °C for at least 2 days. Cellswere grown at 24 °C in YPD broth or in minimal medium brothat 28 °C (0.67% yeast nitrogen base, 2% glucose, with theappropriate supplements to allow growth) for yeast surfacedigestion.
2.2. Sample preparation
2.2.1. Cell surface protein extractionS. cerevisiae cell surface proteins released from whole intactcells by DTT were isolated as described previously [9] withsome modifications. Briefly, cells were grown in 2 l of YPDmedium at 24 °C and incubated for 12 h up to an optical densityof 1 at 600 nm (log-phase cells, 108 cells ml−1). Cells wereharvested by centrifugation and washed twice with pH 7.550 mM-Tris–HCl buffer. Subsequently, cells were resuspendedin 200ml of the same buffer containing 2 mM dithiothreitol(DTT). Cells were shaken vigorously for 2 h at 4 °C and thenpelleted. The supernatant was concentrated by ultrafiltrationusing a pore-size of 10,000 Da (Amicon®, Ultra, Millipore) at 4 °Candwashed five timeswith ice-coldwater. The concentratewasstored at−80 °C. Proteinquantificationwasperformedusing theBradford assay (Bio-Rad, Richmond, CA, USA).
2.2.2. Cell surface-exposed peptide extractionFor S. cerevisiae surface protein digestion we applied themethod described by Martinez-López and colleagues [16].Briefly, cells were grown in minimal medium at 28 °C up toan optical density of 1 at 600 nm (log-phase cells). Cells (109)were collected and washed several times with Milli-Ro water.Subsequently, cells were resuspended in 0.8 ml of 25 mMammonium bicarbonate buffer (pH 8.0) and incubated at 37 °Cfor 5 min with DTT and trypsin. After incubation, sampleswere centrifuged at 3500 ×g for 5 min and the supernatantswere collected. The samples were filtered through 0.22-µmpore-size filters before lyophilization.
2.2.3. Propidium iodide staining and flow cytometric analysisFor PI staining, cells were pelleted by centrifugation for 2 minat 3500 ×g, the supernatant was discarded, and the cells wereresuspended in PI solution (0.005% in phosphate-bufferedsaline).
The stained cells were analyzed with a FACScalibur flowcytometer (Becton Dickinson, Heidelberg, Germany). Cellswere acquired in each sample at different times. The lowerand upper fluorescence limits that included most of the cellsin the live control (more than 99.9%) were determined. Thestained cells within those limits were considered as intact ornon-depolarized cells, and stained cells with higher fluores-cence than the upper limit were regarded as permeabilized ordepolarized cells. Cells treated with 70% ethanol–PBS wereused as a control for dead cells.

1185J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
2.3. Protein and peptide isolation
2.3.1. Two-dimensional gel electrophoresis2D-PAGEwas performed as previously reported [11] with somemodifications. Samples containing 500–700 µg of protein weresolubilized in lysis buffer (7 M urea, 2 M thiourea, 2% CHAPS,65 mMdithioerythritol, 2% IPG buffer pH 3–10 non-linear or pH4.5–5.5 linear (GE Healthcare), bromphenol blue) and werethen applied onto Immobiline pH 3–10 non-linear DryStrips orpH 4.5–5.5 linear DryStrips (18 cm long, GE Healthcare). Thefirst dimension was carried out on an Ettan-IPGphor®Isoelectric focusing unit (GE Healthcare) at 15 °C using thefollowing program: 30 V (active rehydration) for 13 h, 500 V for1 h, 1000 V for 2 h, 2000 V for 2 h, 2000–5000 V for 4 h, and8000 V for 16 h. After this, immobilized pH gradient stripswere reduced (2% dithioerythritol) and then alkylated (2.5%iodoacetamide) in equilibration buffer (6 M urea, 50 mM Tris–HCl, pH 6.8, 30% glycerol, 2% SDS) [17]. The second dimensionwas carried out on homogeneous 10% T, 2.6% C polyacryl-amide gels using piperazine diacrylamide as a cross-linker.Electrophoresis was conducted at 40 mA/gel constant currentfor 6 h in a Protean II cell (Bio-Rad). Preparative gels weresilver-stained as described by Shevchenko [18]. Mr and pIvalues were estimated using external 2D-PAGE standards(Bio-Rad).
Silver-stained gelsweredigitizedusing a computingdensito-meter (GS-690 Imaging Densitometer, Bio-Rad). Different 2Dimages were processed, using Melanie 3.0 software (Bio-Rad).
2.3.2. Nano-liquid chromatographyPeptides from yeast surface digestion were separated by HPLCchromatography, spotted directly onto a MALDI target andanalyzed by MALDI-TOF/TOF (off-line coupled LC/MALDI-MS/MS). Samples were resuspended inmobile phase A and loadedat 20 µl min−1. Peptides were trapped and desalted in the trapcolumn for 5 min. Then, they were separated in the analyticalcolumn according to the following protocol: 90 min 5–50%buffer B, 10 min 50–70% buffer B, 10 min 90% buffer B. Eluateswere continuously spotted at 300 nL min−1 flow onto a 576MALDI target (Applied Biosystems) every 20 s using a Probotµfraction collector (LC Packings). The matrix was addedsimultaneously at the same flow using a µTee-piece.
2.4. Mass spectrometry analysis
2.4.1. Peptide mass fingerprintingThe two-dimensional electrophoresis gel spots weremanuallyexcised from preparative gels using biopsy punches. Proteinsselected for analysis were in-gel reduced, alkylated anddigestedwith trypsin according to Sechi and Chait [19]. Briefly,gel spots were incubated in 100 nM sodium thiosulphate and30 mM potassium fenicyamide, rinsed twice with water,shrunk for 15 min with 100% acetonitrile and dried in a SavantSpeedVac. Then the samples were reduced with 10 mMdithioerythritol in 25 mM ammonium bicarbonate for 30 minat 56 °C and subsequently alkylated with 55 mM iodoaceta-mide in 25 mM ammonium bicarbonate for 15 min in the dark.Gel pieces were alternately washed with 25 mM ambic andACN and dried under vacuum. Finally, samples were digestedwith 12.5 ng/µl sequencing grade trypsin (Roche Molecular
Biochemicals) in 25 mM ammonium bicarbonate (pH 8.5)overnight at 37 °C.
After digestion, the supernatants were collected and 1 µl ofeach sample was spotted onto a MALDI target plate (96×2 spotTeflon®-coated plates) and allowed to air-dry for 10 min atroom temperature. Then, 0.4 µl of a 3 mg/ml α-cyano-4-hydroxy-trans-cinnamic acid matrix (Sigma) in 50% acetoni-trile/0.1% trifluoroacetic acid was added to the dried peptidedigest spots and allowed to air-dry again for another 10 min atroom temperature. MALDI-TOF MS analyses were performedin a Voyager-DE™ STR instrument (PerSeptives Biosystems), amodel fitted with a 337 nm nitrogen laser and operated inreflector mode (accumulating 100 spectra of single laser shotsunder threshold irradiance), with an accelerating voltage of20,000 V. All mass spectra were externally calibrated using astandard peptide mixture: angiotensin I (1296.7), adenocorti-cotropic hormone fragment 18–39 (2465.2) and 1–17 (2093.1)(Sigma). Peptides from the auto digestion of trypsin were usedfor the internal calibration. Some of the samples wereanalyzed in a MALDI-tandem time-of-flight mass spectrome-ter 4700 Proteomics Analyser (Applied Biosystems, Framing-ham, MA).
2.4.2. Tandem mass spectrometryFor surface-exposed peptides, mass spectrometry analysiswas performed automatically with a 4700 Analyser MALDI-TOF/TOF instrument (Applied Biosystems). First, the MSspectra of all the spotted fractions were acquired in positivereflector mode for peak selection (S/N>20, excluded precursorwith 200 resolution) and further MS/MS spectra acquisitionusing the CID of selected peaks.
2.4.3. Database searchesThe peptide mass fingerprinting obtained by MALDI-TOFanalyses from2D gels was used to search for protein candidatein Swiss-Prot sequence databases. The identification ofprotein spots was carried out submitting the monoisotopicpeptide masses assigned to Mascot software available on line(http://www.matrixscience.com). Database Swiss-Prot,TrEMBL, YPD and MIPS were consulted.
The MASCOT parameters were: (1) species: all; (2) allowednumber of missed cleavages (only for trypsin digestion): 1;(3) variable post-translational modifications: methionine oxi-dation and Cys as S-carboamidomethyl derivate; (4): peptidecharge: +1; (5) isoelectric point (3–10) and (6) protein massrange from 10 to 100 kDa. Positive identifications wereaccepted when at least five peptides matched, and at least20% of the peptide coverage of the theoretical sequencesmatched within a mass accuracy of 50 ppm or 25 ppm withinternal calibration. Peptides were excluded if their massescorresponded to those for trypsin, human keratins or otherirrelevant proteins.
The search of filtered peptides of LC–MS/MS from yeastsurface digestion was performed in batch mode using GPSExplorer v3.5 software with a licensed version of MASCOT, ina local database. The MASCOT search parameters were:(1) species: all; (2) allowed number of missed cleavages (onlyfor trypsin digestion): 1; (3) variable post-translational mod-ifications: methionine oxidation (only for DTT treatment);(4) peptide tolerance: ±150 ppm; (5) MS/MS tolerance: ±0.40 Da

1186 J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
and (6): peptide charge: +1. In order to estimate the falsepositive rate (FDR), a search of a composite S. cerevisiaereversed-sequence database was performed. None of thefalse positive proteins were identified in the reverse-sequencedata base with the search parameters used (Supplementaldata, Table S2).
3. Results
3.1. Isolation of cell surface proteins from S. cerevisiae
We set up two different approaches in order to analyze surfaceproteins from S. cerevisiae. The followed strategies aredescribed in Fig. 1: the first, treating whole yeast cells withDTT and separating them by 2D-PAGE (gel proteomic proto-col). And the second, digesting whole intact cells with trypsinand DTT to release the external peptides and separating themby LC (non-gel proteomic protocol).
3.1.1. Cell surface proteins released by DTT treatmentDTT extracts were released under milder conditions fromwhole intact cells as described in Material and methods. Thismethod allows the extraction of proteins that are non-covalently attached to the cell wall or attached by disulphidebridges.
To verify the integrity of yeast cells, selective permeabilitywas assayed by PI staining and quantified by flow cytometry.The unstained cells were considered as intact or non-depolarized cells, and stained cells with higher fluorescencethan the upper limit were regarded as permeabilized ordepolarized cells. Only samples with a high percentage(99.9%) of intact cells were subsequently analyzed (Fig. 2).
Fig. 1 – Strategies used to identify
Protein extracts released from whole intact cells by DTTwere separated by 2D-PAGE. Four different biological repli-cates were analyzed. Fig. 3A shows a pH 3–10 reference gel ofthese proteins in which between 400 and 500 protein spotswere detected. In addition, a narrow pH range of 4.5–5.5 wasused to amplify the region where most of the “bona fide” cellwall proteins were detected (Fig. 3B). Proteins were identifiedby peptide mass fingerprinting (PMF). A total of 68 differentproteins from 81 spots were identified. These proteins arelisted in Table 1, and indicated by name on the 2-DE referencemaps (Fig. 3).
3.1.2. Cell surface-exposed peptide extractionCell surface proteins (surfome) from whole intact cells wereextracted using the protocol described in Materials andmethods by Martínez-López [16]. Cell lysis was controlled byPI staining (Fig. 2). Yeast cell integrity was tested before andafter the incubation step. The results confirmed that most ofthe cells were not altered with respect to their selectivepermeability. The extracts were analyzed by nano-HPLC–MS/MS. Tryptic peptides present in the supernatant were concen-trated and separated by reverse-phase nano-HPLC and theproteins were identified using MALDI-TOF/TOF. Four differentbiological replicates were analyzed. Identified proteins arelisted in Table 2 and in Table S1 (Supplemental data).
3.2. Functional classification of cell surface proteins fromS. cerevisiae
Tables 1 and 2 show the proteins identified by both methodsand classified according to their biological function. Proteinsthat could be in more than one group were included in themost representative group. We employed the functional
S. cerevisiae cell wall proteins.

Fig. 2 – S. cerevisiae cell integrity measurement by flow cytometry. A. Negative fluorescence control: yeast cell culture time 0.B. Positive fluorescence control: cells permeabilized by 70% ethanol–PBS treatment. C. Yeast cells treated with DTT for 2 h.D. Yeast cells collected after 5 min of cell surface digestion with trypsin and DTT. These measurements were performed forevery experiment. Graphics show representative experiments.
1187J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
catalogue of the CYGD (comprehensive yeast genome data-base) from MIPS http://mips.gsf.de/proj/yeast/CYGD/db/ andthe data in the BIOBASE Knowledge Library.
Of the 67 identified proteins released fromwhole intact cellsby DTT (Table 1), 9 proteins are classified in control of cellularorganization (cell wall and cytoskeleton), 39 proteins belong tobiosynthetic routes of carbohydrate, amino acid, nucleotide andphosphate metabolism, and 16 of them are enzymes fromdifferent pathways of energy generation (glycolysis, gluconeo-genesis, pentose-phosphate and tricarboxylic pathway). Wealso identified 5 proteins involved in transcription and proteinsynthesis, 9 related to protein fate, 5 involved in cell rescue,defence and virulence and 2 in cell signalling. Of the 52 surface-exposed proteins identified (Table 2), 13 are involved inmolecular and cellular organization (cell wall and cytoskeleton),and 14 are enzymes from different pathways of energygeneration (glycolysis, gluconeogenesis, pentose-phosphateand tricarboxylic pathway). We also identified 16 proteinsinvolved in transcription and protein synthesis, 6 related toprotein fate and 2 involved in cell rescue, defence and virulence.
4. Discussion
In the present work, two different proteomic approaches havebeen carried out to analyze cell surface proteins from S.cerevisiae: 2D-PAGE and nano-LC–MS/MS. Cell integrity wasassured during both extraction protocols to guarantee surfaceprotein enrichment. A total of 99 different proteins wereisolated and identified using the two methods: 67 with DTTtreatment and 52 with cell surface digestion (Tables 1 and 2).Only 20 were common to both procedures (Fig. 4). We foundproteins classified in different cellular processes: control ofcellular organization, cell rescue, defence, and virulence,protein fate, protein synthesis and metabolism. These proteingroups are discussed below.
4.1. Control of cellular organization
Most of the proteins classified in this group are related to cellwall organization. Four glucanases (Bgl2p, Exg1p, Scw4p, andScw3p) and one PIR protein (Cis3p/Pir4p)were identified byDTT
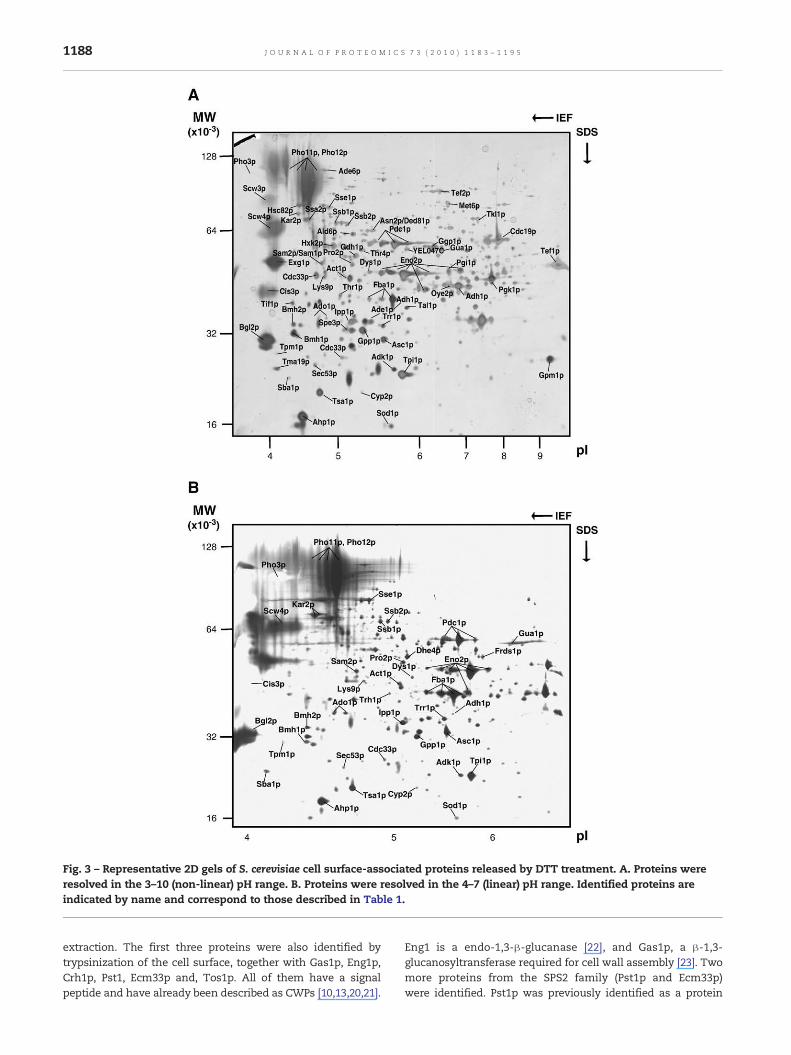
Fig. 3 – Representative 2D gels of S. cerevisiae cell surface-associated proteins released by DTT treatment. A. Proteins wereresolved in the 3–10 (non-linear) pH range. B. Proteins were resolved in the 4–7 (linear) pH range. Identified proteins areindicated by name and correspond to those described in Table 1.
1188 J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
extraction. The first three proteins were also identified bytrypsinization of the cell surface, together with Gas1p, Eng1p,Crh1p, Pst1, Ecm33p and, Tos1p. All of them have a signalpeptide and have already been described as CWPs [10,13,20,21].
Eng1 is a endo-1,3-β-glucanase [22], and Gas1p, a β-1,3-glucanosyltransferase required for cell wall assembly [23]. Twomore proteins from the SPS2 family (Pst1p and Ecm33p)were identified. Pst1p was previously identified as a protein

Table 1 – Clustering of S. cerevisiae DTT-extractable proteins from whole cells from separated by 2D-PAGE and identified byMALDI-TOF MS. Proteins are classified according to Gene Ontology Molecular Function.
Accessionnumber a
Protein name Description Mr (kDa) b pI b No. matchedpeptides c
% covered ofthe Protd
Control of cellular organizationCell wallP15703 Bgl2p Endo-beta-1,3-glucanase 33 4.2 8–13 25–45P47001 Cis3p Cell wall protein member of the PIR family 43 4.0 5 15P23776 Exg1p Exo-beta-1,3-glucanase 43 3.9 12 26P53616 Scw3p Cell wall protein related to glucanases 78 4.1 2 6P53334 Scw4p Cell wall protein related to glucanases 66 4.2 9 26
Actin cytoskeletonP60010 Act1p Actin 43 5.0 7–13 21–41P17536 Tpm1p Tropomiosin 31 4.2 4 19
Mitochondria localizationP35691 Tma19p Microtubule and mitochondria interacting
protein26 4.0 6 39
Cell rescue, defence and virulenceStress responseP38013 Ahp1p Alkyl hydroperoxide reductase 22 4.8 11 45P00445 Sod1p Copper–zinc superoxide dismutase 16 5.5 7 59P29509 Trr1p Thioredoxin reductase 35 5.6 4 25Q035528 Oye2p NADPH dehydrogenase 44 6.7 6P34760 Tsa1p Thioredoxin peroxidase 21 4.6 5–7 24–40
Cell signalingP29311 Bmh1p 14–3–3 protein involved in rapamycin-
sensitive signalling31 4.3 5 17
P34730 Bmh2p 14–3–3 protein involved in rapamycin-sensitive signalling
34 4.3 5 19
Protein fateProtein folding and stabilizationP23285 Cyp2p Peptidyl-prolyl cis-trans isomerase
(cyclophilin)21 5.1 10 46
P38879 Egd2p Alpha subunit of the nascent polypeptide-associated complex
18* 4.6* 7 44
P15108 Hsc82p Heat shock protein member of HSP90 family 83 4.4 23–30 38–47P16474 Kar2p Heat shock protein member of HSP70 family 73 4.3–4.4 8–13 15–24P28707 Sba1p HSP90 associated co-chaperone 25 4.2 4 14P10592 Ssa2p Heat shock protein member of HSP70 family 72 4.5 6–12 11–26P11484 Ssb1p Heat shock protein member of HSP70 family 69 4.8 7–11 13–19P40150 Ssb2p Heat shock protein member of HSP70 family 69 4.9 6–14 13–26P32589 Sse1p Heat shock protein member of HSP70 family 82 4.8 9 17
MetabolismCarbohydrate metabolismP07283 Sec53p Phosphomannomutase 26–39 4.5–4.3 7–11 28–46P32614 YEL047C Fumarate reductase 52 5.8 6 10
Glycolytic enzymesP00549 Cdc19p Pyruvate kinase 1 59 7.8 35 71P00925 Eno2p Enolase 2 47 5.3–5.8 8–21 28–49P14540 Fba1p Fructose-biphosphate aldolase 41 5.2–5.6 11–22 32–51P00950 Gpm1p Phosphoglycerate mutase 1 27 9.6 4 17P04807 Hxk2p Hexokinase B 55 4.9 11–31 30–62P12709 Pgi1p Glucose-6-phosphate isomerase 55 6.4 30 59P00560 Pgk1p Phosphoglycerate kinase 46 7.7 16–34 53–81P00942 Tpi1p Triosephosphate isomerase 24 5.7 8–12 35–41
Fermentative enzymesP00330 Adh1p Alcohol dehydrogenase 19 4.4 5 18P54115 Ald6p Aldehyde dehydrogenase 61 5.1 16 38P41277 Gpp1p DL-glycerol phosphate phosphatase 32 5.2 14 46P06169 Pdc1p Pyruvate decarboxylase isozyme 1 60 5.3–5.7 9–11 20–34P15019 Tal1p Transaldolase 39 5.8 6 21
(continued on next page)
1189J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
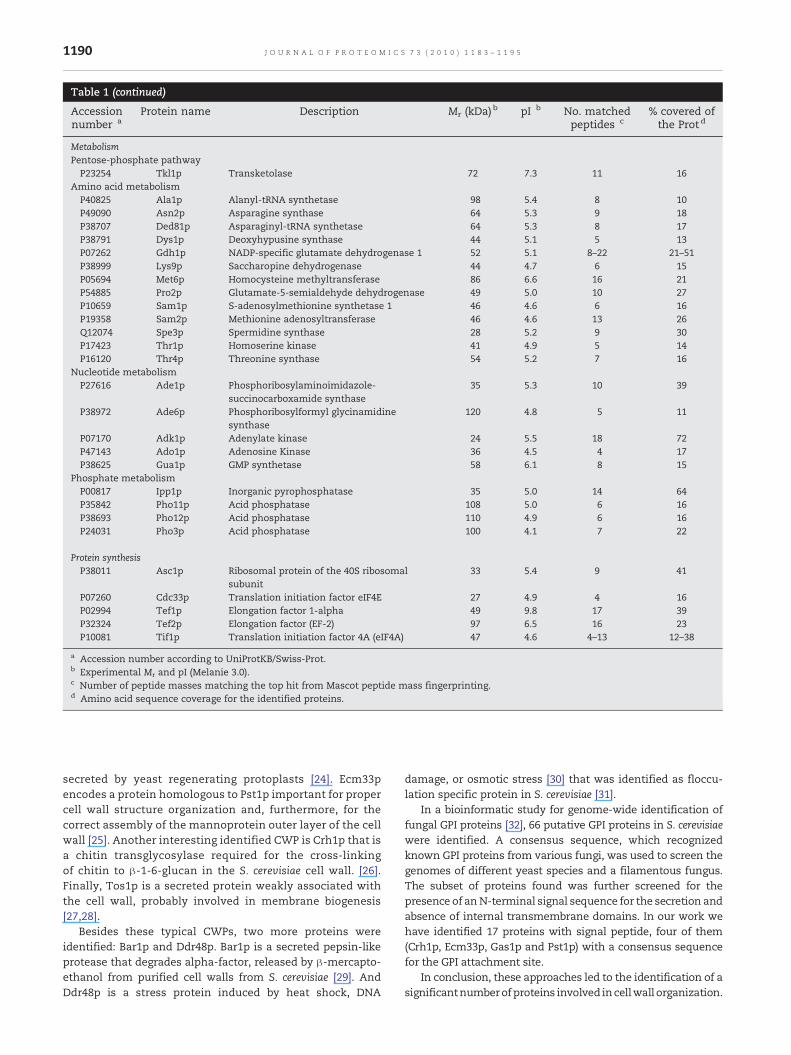
Table 1 (continued)
Accessionnumber a
Protein name Description Mr (kDa) b pI b No. matchedpeptides c
% covered ofthe Protd
MetabolismPentose-phosphate pathwayP23254 Tkl1p Transketolase 72 7.3 11 16
Amino acid metabolismP40825 Ala1p Alanyl-tRNA synthetase 98 5.4 8 10P49090 Asn2p Asparagine synthase 64 5.3 9 18P38707 Ded81p Asparaginyl-tRNA synthetase 64 5.3 8 17P38791 Dys1p Deoxyhypusine synthase 44 5.1 5 13P07262 Gdh1p NADP-specific glutamate dehydrogenase 1 52 5.1 8–22 21–51P38999 Lys9p Saccharopine dehydrogenase 44 4.7 6 15P05694 Met6p Homocysteine methyltransferase 86 6.6 16 21P54885 Pro2p Glutamate-5-semialdehyde dehydrogenase 49 5.0 10 27P10659 Sam1p S-adenosylmethionine synthetase 1 46 4.6 6 16P19358 Sam2p Methionine adenosyltransferase 46 4.6 13 26Q12074 Spe3p Spermidine synthase 28 5.2 9 30P17423 Thr1p Homoserine kinase 41 4.9 5 14P16120 Thr4p Threonine synthase 54 5.2 7 16
Nucleotide metabolismP27616 Ade1p Phosphoribosylaminoimidazole-
succinocarboxamide synthase35 5.3 10 39
P38972 Ade6p Phosphoribosylformyl glycinamidinesynthase
120 4.8 5 11
P07170 Adk1p Adenylate kinase 24 5.5 18 72P47143 Ado1p Adenosine Kinase 36 4.5 4 17P38625 Gua1p GMP synthetase 58 6.1 8 15
Phosphate metabolismP00817 Ipp1p Inorganic pyrophosphatase 35 5.0 14 64P35842 Pho11p Acid phosphatase 108 5.0 6 16P38693 Pho12p Acid phosphatase 110 4.9 6 16P24031 Pho3p Acid phosphatase 100 4.1 7 22
Protein synthesisP38011 Asc1p Ribosomal protein of the 40S ribosomal
subunit33 5.4 9 41
P07260 Cdc33p Translation initiation factor eIF4E 27 4.9 4 16P02994 Tef1p Elongation factor 1-alpha 49 9.8 17 39P32324 Tef2p Elongation factor (EF-2) 97 6.5 16 23P10081 Tif1p Translation initiation factor 4A (eIF4A) 47 4.6 4–13 12–38
a Accession number according to UniProtKB/Swiss-Prot.b Experimental Mr and pI (Melanie 3.0).c Number of peptide masses matching the top hit from Mascot peptide mass fingerprinting.d Amino acid sequence coverage for the identified proteins.
1190 J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
secreted by yeast regenerating protoplasts [24]. Ecm33pencodes a protein homologous to Pst1p important for propercell wall structure organization and, furthermore, for thecorrect assembly of the mannoprotein outer layer of the cellwall [25]. Another interesting identified CWP is Crh1p that isa chitin transglycosylase required for the cross-linkingof chitin to β-1-6-glucan in the S. cerevisiae cell wall. [26].Finally, Tos1p is a secreted protein weakly associated withthe cell wall, probably involved in membrane biogenesis[27,28].
Besides these typical CWPs, two more proteins wereidentified: Bar1p and Ddr48p. Bar1p is a secreted pepsin-likeprotease that degrades alpha-factor, released by β-mercapto-ethanol from purified cell walls from S. cerevisiae [29]. AndDdr48p is a stress protein induced by heat shock, DNA
damage, or osmotic stress [30] that was identified as floccu-lation specific protein in S. cerevisiae [31].
In a bioinformatic study for genome-wide identification offungal GPI proteins [32], 66 putative GPI proteins in S. cerevisiaewere identified. A consensus sequence, which recognizedknown GPI proteins from various fungi, was used to screen thegenomes of different yeast species and a filamentous fungus.The subset of proteins found was further screened for thepresence of anN-terminal signal sequence for the secretion andabsence of internal transmembrane domains. In our work wehave identified 17 proteins with signal peptide, four of them(Crh1p, Ecm33p, Gas1p and Pst1p) with a consensus sequencefor the GPI attachment site.
In conclusion, these approaches led to the identification of asignificantnumberofproteins involved incellwall organization.
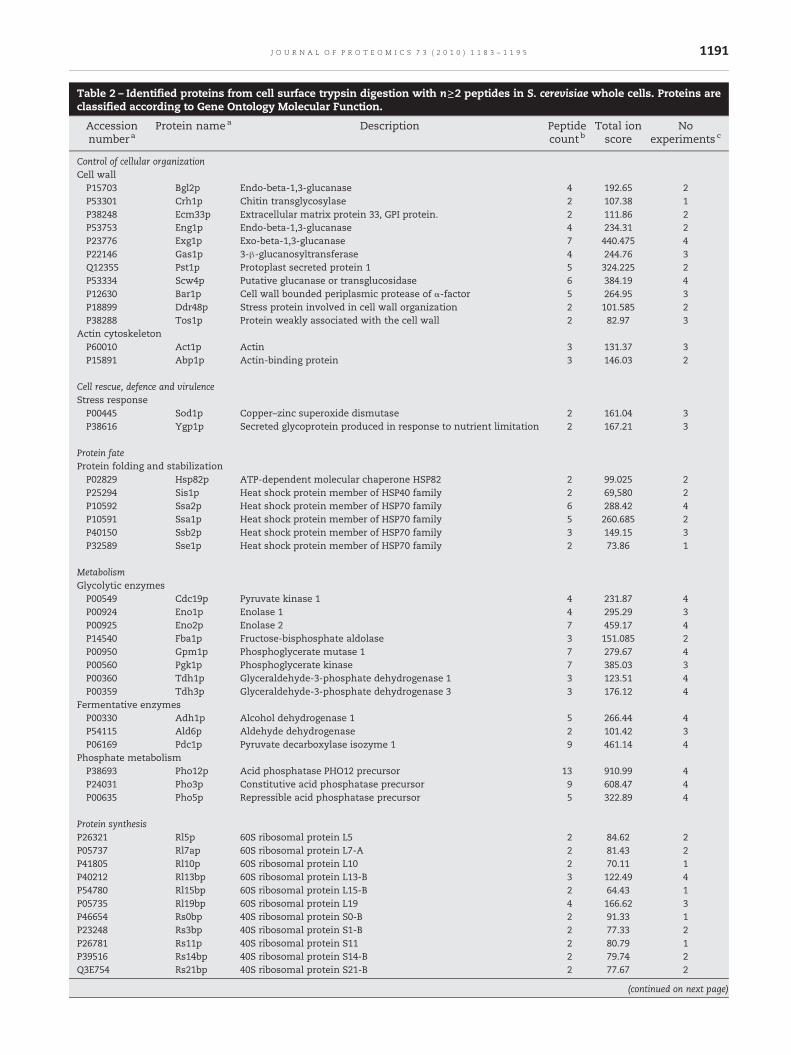
Table 2 – Identified proteins from cell surface trypsin digestion with n≥2 peptides in S. cerevisiae whole cells. Proteins areclassified according to Gene Ontology Molecular Function.
Accessionnumbera
Protein namea Description Peptidecountb
Total ionscore
Noexperiments c
Control of cellular organizationCell wallP15703 Bgl2p Endo-beta-1,3-glucanase 4 192.65 2P53301 Crh1p Chitin transglycosylase 2 107.38 1P38248 Ecm33p Extracellular matrix protein 33, GPI protein. 2 111.86 2P53753 Eng1p Endo-beta-1,3-glucanase 4 234.31 2P23776 Exg1p Exo-beta-1,3-glucanase 7 440.475 4P22146 Gas1p 3-β-glucanosyltransferase 4 244.76 3Q12355 Pst1p Protoplast secreted protein 1 5 324.225 2P53334 Scw4p Putative glucanase or transglucosidase 6 384.19 4P12630 Bar1p Cell wall bounded periplasmic protease of α-factor 5 264.95 3P18899 Ddr48p Stress protein involved in cell wall organization 2 101.585 2P38288 Tos1p Protein weakly associated with the cell wall 2 82.97 3
Actin cytoskeletonP60010 Act1p Actin 3 131.37 3P15891 Abp1p Actin-binding protein 3 146.03 2
Cell rescue, defence and virulenceStress responseP00445 Sod1p Copper–zinc superoxide dismutase 2 161.04 3P38616 Ygp1p Secreted glycoprotein produced in response to nutrient limitation 2 167.21 3
Protein fateProtein folding and stabilizationP02829 Hsp82p ATP-dependent molecular chaperone HSP82 2 99.025 2P25294 Sis1p Heat shock protein member of HSP40 family 2 69,580 2P10592 Ssa2p Heat shock protein member of HSP70 family 6 288.42 4P10591 Ssa1p Heat shock protein member of HSP70 family 5 260.685 2P40150 Ssb2p Heat shock protein member of HSP70 family 3 149.15 3P32589 Sse1p Heat shock protein member of HSP70 family 2 73.86 1
MetabolismGlycolytic enzymesP00549 Cdc19p Pyruvate kinase 1 4 231.87 4P00924 Eno1p Enolase 1 4 295.29 3P00925 Eno2p Enolase 2 7 459.17 4P14540 Fba1p Fructose-bisphosphate aldolase 3 151.085 2P00950 Gpm1p Phosphoglycerate mutase 1 7 279.67 4P00560 Pgk1p Phosphoglycerate kinase 7 385.03 3P00360 Tdh1p Glyceraldehyde-3-phosphate dehydrogenase 1 3 123.51 4P00359 Tdh3p Glyceraldehyde-3-phosphate dehydrogenase 3 3 176.12 4
Fermentative enzymesP00330 Adh1p Alcohol dehydrogenase 1 5 266.44 4P54115 Ald6p Aldehyde dehydrogenase 2 101.42 3P06169 Pdc1p Pyruvate decarboxylase isozyme 1 9 461.14 4
Phosphate metabolismP38693 Pho12p Acid phosphatase PHO12 precursor 13 910.99 4P24031 Pho3p Constitutive acid phosphatase precursor 9 608.47 4P00635 Pho5p Repressible acid phosphatase precursor 5 322.89 4
Protein synthesisP26321 Rl5p 60S ribosomal protein L5 2 84.62 2P05737 Rl7ap 60S ribosomal protein L7-A 2 81.43 2P41805 Rl10p 60S ribosomal protein L10 2 70.11 1P40212 Rl13bp 60S ribosomal protein L13-B 3 122.49 4P54780 Rl15bp 60S ribosomal protein L15-B 2 64.43 1P05735 Rl19bp 60S ribosomal protein L19 4 166.62 3P46654 Rs0bp 40S ribosomal protein S0-B 2 91.33 1P23248 Rs3bp 40S ribosomal protein S1-B 2 77.33 2P26781 Rs11p 40S ribosomal protein S11 2 80.79 1P39516 Rs14bp 40S ribosomal protein S14-B 2 79.74 2Q3E754 Rs21bp 40S ribosomal protein S21-B 2 77.67 2
(continued on next page)
1191J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5

Table 2 (continued)
Accessionnumbera
Protein namea Description Peptidecountb
Total ionscore
Noexperimentsc
Protein synthesisP32827 Rs23p 40S ribosomal protein S23 2 83.76 2P02994 Tef1p Elongation factor 1-alpha 3 194.848 3P32324 Tef2p Elongation factor 2 (EF-2) 3 156.12 2P16521 Tef3p Elongation factor 3A 2 96.545 3P34167 Tif3p Eukaryotic translation initiation factor 4B (eIF-4B) 2 90.27 1
Unknown functionP40159 Ynu8p Uncharacterized protein YNL208W 2 196.47 4
a Accession number based on UniProtKB/Swiss-Prot.b Number of identified peptides. Only individual ion CI ≥99% (related with p<0.01 Mascot score), was used.c Number of experiments where the protein has been identified.
1192 J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
4.2. Cell rescue, defence, and virulence
Six proteins related to stress responses have been identified.Ahp1p and Tsa1p have thioredoxin peroxidase activity.Tsa1p is specific for hydrogen peroxide and Ahp1p is specificfor organic peroxides. Trr1, thioredoxin reductase 1 thatfunctions in the response to oxidative stress. It's one of theproteins of the thioredoxin system (Trx1p, Trx2p, and Trr1p)which is required for the antioxidant function of Ahp1p [33].Oye2p is an oxidoreductase with modulating roles inoxidative stress and programmed cell death in yeast [34].And Sod1p is the superoxide dismutase 1 (copper–zincsuperoxide dismutase), an enzyme that functions in thestress response to temperature, oxidation, and radiation [35].Finally Ygp1p is a protein secreted in response to nutrientlimitation [36].
4.3. Metabolism
Sixteen and 14 metabolic proteins were identified by DTTextraction and cell surface digestion, respectively. The occur-
Fig. 4 – Number of proteins identified by the two methods.Proteins involved in cell wall organization are indicated byname.
rence of glycolytic enzymes as cell wall components and asimmunogenic proteins has been extensively reported in theliterature (reviewed in [37]), and thus we expected to identifysomeof them. For S. cerevisiae Eno1, for instance, there is geneticand proteomic evidence of its presence on the cell surface [38].This protein has recently been described as an anchorless S.aureus surface protein [39] and antibodies against it haveshowed opsonizing and killing capabilities against S. aureus.
In addition four acid phosphatases with signal peptide thatare exported to the cell surface by secretory vesicles [40] werefound, two of them (Pho3p and Pho12p) were identified in bothstrategies.
4.4. Protein synthesis and protein fate
With DTT extraction 8 proteins involved in protein synthesiswere identified. Tef1p has already been described as beingassociated with the cell surface in several organisms likeC. albicans [41], Trichoderma reesei [42], Mycobacterium leprae [43]and Bacillus subtilis [44]. With DTT and trypsin digestion,translation, elongation and initiation factors were also iden-tified, many of them already described on the cell surface byother authors [37]. In addition, 12 ribosomal proteins wereidentified by LC–MS after cell surface DTT and trypsindigestion. However none of these proteins were detected by2D-PAGE probably because their lowmolecular weight and thebasic pH make their detection impossible in a standard 2D gel(Fig. 5). Some ribosomal proteins have already been identifiedon the cell surface of Neisseria meningitidis [45], Plasmodiumfalciparum [46] and other microorganisms.
Besides 12 heat shock proteins were identified in oursamples. Six proteins from the HSP70 family were identifiedby the two strategies: Ssa1p, Ssa2p, Sse1p, Ssb1p, Ssb2pand Kar2p. Some members of the Hsp70 family proteins ofS. cerevisiae are located in the cell wall [47]. Five morechaperones were detected by DTT extraction: Sis1p (memberof Hsp 40 family) and Hsc82p, Cyd2p, Egd2p and Sba1p (Hsp 90family).
In conclusion, the twoprotein extractionprocedures allowedthe identification of a substantial number of S. cerevisiaecell surface proteins (Fig. 4). However, the DTT and trypsinproteindigestion followed by LC (non-gel-based strategy) seems

Fig. 5 – Virtual gel of the identified DTT-released proteins (in blue) and 117S. cerevisiae predicted ribosomal proteins (in red)using the software JVirGel 2.0 (http://www.jvirgel.de/). Protein spots in green represent the proteins identified also by LC. Therectangle shows the proteins that would be resolved the 3–10 (non-linear) pH range 2D-PAGE.
1193J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
to be more specific for “bona fide” cell wall proteins. Cell wallproteins are usually glycosylated, have a highmolecular weightand are hydrophobic. All these characteristics make theirisolation and identification by classical proteomics difficult.The DTT protein extraction followed by 2D-PAGE separationallowed the detection of a minor number of typically cellwall proteins and a higher number of proteins related withstress response, protein fate and metabolism, than withDTT and trypsin digestion of cell surface-exposed peptidesextraction. Only 20% of the proteins were common to bothprocedures, and thus, for a better knowledge of the cell wallproteome, several extractionmethods and proteomic strategiesshould be used.
Taking together these results with previous ones aboutC. albicans cell surface, from our group [11,16,38,48] and fromother groups [49], we can state that the yeast cell surface iscomposed not only by typical but also by atypical cell wallproteins. For a better understanding of the yeast cell surfaceproteome further studies about the mechanisms of secretionof these proteins and their role in the cell wall dynamics arerequired.
Acknowledgements
Protein identification was carried out in the Proteomics UnitUCM-Parque Científico, a member of the National Institute forProteomics, ProteoRed, funded by Genoma España. This workwas supported by BIO 01989-2006 from the Comisión Inter-ministerial de Ciencia y Tecnología (CYCIT, Spain), DEREMI-CROBIANA-CM from the Comunidad Autónoma de Madrid,
and REIPI, Spanish Network for the Research in InfectiousDiseases, RD06/0008/1027, from the Instituto de Salud CarlosIII and European Community STREP (Specific Targeted Re-search or Innovation Prrogram) LSHB-CT-2004-511952. C.Nombela is the director of the Merck, Sharp & Dohme (MSD)Special Chair in Genomics and Proteomics.
Appendix A. Supplementary data
Supplementary data associated with this article can be found,in the online version, at doi:10.1016/j.jprot.2010.02.005.
R E F E R E N C E S
[1] Orlean P. Biogenesis of yeast cell wall and surfacecomponents. Cell cycle and cell biology. New York: Cold SpringHarbor Laboratory Press; 1997. p. 229–362.
[2] Molina M, Gil C, Pla J, Arroyo J, Nombela C. Protein localisationapproaches for understanding yeast cell wall biogenesis.Microsc Res Tech 2000;51(6):601–12.
[3] Klis FM, Boorsma A, de Groot PW. Cell wall construction inSaccharomyces cerevisiae. Yeast 2006;23(3):185–202.
[4] Cid VJ, Duran A, del Rey F, Snyder MP, Nombela C, Sanchez M.Molecular basis of cell integrity and morphogenesis inSaccharomyces cerevisiae. Microbiol Rev 1995;59(3):345–86.
[5] Osumi M. The ultrastructure of yeast: cell wall structure andformation. Micron 1998;29(2–3):207–33.
[6] Dranginis AM, Rauceo JM, Coronado JE, Lipke PN. Abiochemical guide to yeast adhesins: glycoproteins for socialand antisocial occasions. Microbiol Mol Biol Rev 2007;71(2):282–94.

1194 J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
[7] Sakai Y, Azuma M, Takada Y, Umeyama T, Kaneko A, Fujita T,et al. Saccharomyces cerevisiae mutant displaying beta-glucanson cell surface. J Biosci Bioeng 2007;103(2):161–6.
[8] Ecker M, Deutzmann R, Lehle L, Mrsa V, Tanner W. Pirproteins of Saccharomyces cerevisiae are attached to beta-1,3-glucan by a new protein-carbohydrate linkage. J Biol Chem2006;281(17):11523–9.
[9] Cappellaro C, Mrsa V, Tanner W. New potential cell wallglucanases of Saccharomyces cerevisiae and their involvementin mating. J Bacteriol 1998;180(19):5030–7.
[10] Mrsa V, Seidl T, GentzshM, TannerW. Specific labelling of cellwall proteins by biotinylation. Identification of fourcovalently linked O-mannosylated proteins of Saccharomycescerevisiae. Yeast 1997;13(12):1145–54.
[11] Pitarch A, Sánchez M, Nombela C, Gil C. Sequentialfractionation and two-dimensional gel analysis unravels thecomplexity of the dimorphic fungus Candida albicans cell wallproteome. Mol Cell Proteomics 2002;1(12):967–82.
[12] Pitarch A, Abián J, Carrascal M, Sánchez M, Nombela C, Gil C.Proteomics-based identification of novel Candida albicansantigens for diagnosis of systemic candidiasis in patientswith underlying hematological malignancies. Proteomics2004;4(10):3084–106.
[13] Pardo M, Monteoliva L, Pla J, Sánchez M, Gil C, Nombela C.Two-dimensional analysis of proteins secreted bySaccharomyces cerevisiae regenerating protoplasts: a novelapproach to study the cell wall. Yeast 1999;15(6):459–72.
[14] Tjalsma H, Lambooy L, Hermans PW, Swinkels DW. Shedding& shaving: disclosure of proteomic expressions on a bacterialface. Proteomics 2008;8(7):1415–28.
[15] Yin QY, de Groot PW, de Jong L, Klis FM, De Koster CG. Massspectrometric quantitation of covalently bound cellwall proteinsin Saccharomyces cerevisiae. FEMS Yeast Res 2007;7(6):887–96.
[16] Martínez-López R, Nombela C, Diez-Orejas R, Monteoliva L, GilC. Immunoproteomic analysis of the protective responseobtained from vaccination with Candida albicans ecm33 cellwall mutant in mice. Proteomics 2008;8(13):2651–64.
[17] Bjellqvist B, Pasquali C, Ravier F, Sánchez JC, Hochstrasser D.A nonlinear wide-range immobilized pH gradient for two-dimensional electrophoresis and its definition in a relevantpH scale. Electrophoresis 1993;14(12):1357–65.
[18] Shevchenko A,WilmM, VormO,MannM.Mass spectrometricsequencing of proteins silver-stained polyacrylamide gels.Anal Chem 1996;68(5):850–8.
[19] Sechi S, Chait BT. Modification of cysteine residues byalkylation. A tool in peptide mapping and proteinidentification. Anal Chem 1998;70(24):5150–8.
[20] de Groot PW, de Boer AD, Cunningham J, Dekker HL, de Jong L,Hellingwerf KJ, et al. Proteomic analysis of Candida albicanscell walls reveals covalently bound carbohydrate-activeenzymes and adhesins. Eukaryot Cell 2004;3(4):955–65.
[21] Yin QY, de Groot PW, Dekker HL, de Jong L, Klis FM, De KosterCG. Comprehensive proteomic analysis of Saccharomycescerevisiae cell walls: identification of proteins covalentlyattached via glycosylphosphatidylinositol remnants or mildalkali-sensitive linkages. J Biol Chem 2005;280(21):20894–901.
[22] Baladrón V, Ufano S, Dueñas E, Martín-Cuadrado AB, del ReyF, Vázquez de Aldana CR. Eng1p, an endo-1, 3-beta-glucanaselocalized at the daughter side of the septum, is involved in cellseparation in Saccharomyces cerevisiae. Eukaryot Cell 2002;1(5):774–86.
[23] Ragni E, Fontaine T, Gissi C, Latge JP, Popolo L. The Gas familyof proteins of Saccharomyces cerevisiae: characterization andevolutionary analysis. Yeast 2007;24(4):297–308.
[24] Pardo M,WardM, Bains S, Molina M, BlackstockW, Gil C, et al.A proteomic approach for the study of Saccharomyces cerevisiaecell wall biogenesis. Electrophoresis 2000;21(16):3396–410.
[25] Pardo M, Monteoliva L, Vázquez P, Martínez R, Molero G,Nombela C, et al. PST1 and ECM33 encode two yeast cell
surface GPI proteins important for cell wall integrity.Microbiology 2004;150(Pt 12):4157–70.
[26] Cabib E, Blanco N, Grau C, Rodríguez-Peña JM, Arroyo J. Crh1pand Crh2p are required for the cross-linking of chitin to beta(1-6)glucan in the Saccharomyces cerevisiae cell wall. MolMicrobiol 2007;63(3):921–35.
[27] Terashima H, Fukuchi S, Nakai K, Arisawa M, Hamada K,Yabuki N, et al. Sequence-based approach for identification ofcell wall proteins in Saccharomyces cerevisiae. Curr Genet2002;40(5):311–6.
[28] Gavin AC, Bosche M, Krause R, Grandi P, Marzioch M, Bauer A,et al. Functional organization of the yeast proteome bysystematic analysis of protein complexes. Nature 2002;415(6868):141–7.
[29] Moukadiri I, Jaafar L, Zueco J. Identification of twomannoproteins released from cell walls of a Saccharomycescerevisiae mnn1 mnn9 double mutant by reducing agents.J Bacteriol 1999;181(16):4741–5.
[30] Miralles VJ, Serrano R. A genomic locus in Saccharomycescerevisiae with four genes up-regulated by osmotic stress.Mol Microbiol 1995;17(4):653–62.
[31] Tonouchi A, Fujita A, Kuhara S. Molecular cloning of the geneencoding a highly expressed protein in SFL1 gene-disruptedflocculating yeast. J Biochem 1994;115(4):683–8.
[32] de Groot PW, Hellingwerf KJ, Klis FM. Genome-wideidentification of fungal GPI proteins. Yeast 2003;20(9):781–96.
[33] Lee J, Spector D, Godon C, Labarre J, Toledano MB. A newantioxidant with alkyl hydroperoxide defense properties inyeast. J Biol Chem 1999;274(8):4537–44 %19.
[34] Odat O, Matta S, Khalil H, Kampranis SC, Pfau R, Tsichlis PN,et al. Old yellow enzymes, highly homologous FMNoxidoreductases with modulating roles in oxidative stressand programmed cell death in yeast. J Biol Chem 2007;282(49):36010–23.
[35] Pereira MD, Eleutherio EC, Panek AD. Acquisition of toleranceagainst oxidative damage in Saccharomyces cerevisiae. BMCMicrobiol 2001;1:11.
[36] Destruelle M, Holzer H, Klionsky DJ. Identification andcharacterization of a novel yeast gene: the YGP1 gene productis a highly glycosylated secreted protein that is synthesized inresponse to nutrient limitation. Mol Cell Biol 1994;14(4):2740–54.
[37] Chaffin WL. Candida albicans cell wall proteins. Microbiol MolBiol Rev 2008;72(3):495–544.
[38] López-Villar E, Monteoliva L, Larsen MR, Sachon E, Shabaz M,Pardo M, et al. Genetic and proteomic evidences support thelocalization of yeast enolase in the cell surface. Proteomics2006;6(Suppl 1):S107–18.
[39] Glowalla E, Tosetti B, Kronke M, Krut O. Proteomics-basedidentification of anchorless cell wall proteins as vaccinecandidates against Staphylococcus aureus. Infect Immun2009;77(7):2719–29.
[40] Blinnikova EI, Mirjuschenko FL, Shabalin YA, Egorov SN.Vesicular transport of extracellular acid phosphatases inyeast Saccharomyces cerevisiae. Biochemistry (Mosc) 2002;67(4):485–90.
[41] Urban C, Sohn K, Lottspeich F, Brunner H, Rupp S.Identification of cell surface determinants in Candida albicansreveals Tsa1p, a protein differentially localized in the cell.FEBS Lett 2003;544(1–3):228–35.
[42] Lim D, Hains P, Walsh B, Bergquist P, Nevalainen H. Proteinsassociated with the cell envelope of Trichoderma reesei: aproteomic approach. Proteomics 2001;1(7):899–909.
[43] MarquesMA, Chitale S, Brennan PJ, PessolaniMC.Mapping andidentification of the major cell wall-associated components ofMycobacterium leprae. Infect Immun 1998;66(6):2625–31.
[44] Antelmann H, Tjalsma H, Voigt B, Ohlmeier S, Bron S, van DijlJM, et al. A proteomic view on genome-based signal peptidepredictions. Genome Res 2001;11(9):1484–502.

1195J O U R N A L O F P R O T E O M I C S 7 3 ( 2 0 1 0 ) 1 1 8 3 – 1 1 9 5
[45] Mendum TA, Newcombe J, McNeilly CL, McFadden J. Towardsthe immunoproteome of Neisseria meningitidis. PLoS One2009;4(6):e5940.
[46] Chatterjee S, Singh S, Sohoni R, Singh NJ, Vaidya A, Long C,et al. Antibodies against ribosomal phosphoprotein P0 ofPlasmodium falciparum protect mice against challengewith Plasmodium yoelii. Infect Immun 2000;68(7):4312–4318.
[47] López-Ribot JL, Chaffin WL. Members of the Hsp70 family ofproteins in the cell wall of Saccharomyces cerevisiae. J Bacteriol1996;178(15):4724–6.
[48] Nombela C, Gil C, Chaffin WL. Non-conventional proteinsecretion in yeast. Trends Microbiol 2006;14(1):15–21.
[49] Castillo L, Calvo E, Martinez AI, Ruiz-Herrera J, Valentin E,Lopez JA, et al. A study of the Candida albicans cell wallproteome. Proteomics 2008;8(18):3871–81.




















