Creatinine-Based Equations Predicting Chronic Kidney Disease After Kidney Donation
Upload
khangminh22Category
view
2download
0
Volume 30 Issue 2 Article 3
2022
Exosomal proteomics in kidney disease: From technical Exosomal proteomics in kidney disease: From technical
approaches to clinical applications approaches to clinical applications
Follow this and additional works at: https://www.jfda-online.com/journal
Part of the Food Science Commons, Medicinal Chemistry and Pharmaceutics Commons,
Pharmacology Commons, and the Toxicology Commons
This work is licensed under a Creative Commons Attribution-Noncommercial-No Derivative
Works 4.0 License.
Recommended Citation Recommended Citation Brahmadhi, Ageng; Chuang, Yung-Kun; Wang, San-Yuan; Kao, Chih-Chin; and Tsai, I-Lin (2022) "Exosomal proteomics in kidney disease: From technical approaches to clinical applications," Journal of Food and Drug Analysis: Vol. 30 : Iss. 2 , Article 3. Available at: https://doi.org/10.38212/2224-6614.3409
This Review Article is brought to you for free and open access by Journal of Food and Drug Analysis. It has been accepted for inclusion in Journal of Food and Drug Analysis by an authorized editor of Journal of Food and Drug Analysis.

Exosomal proteomics in kidney disease: Fromtechnical approaches to clinical applications
Ageng Brahmadhi a,b,c, Yung-Kun Chuang d,e,f,1, San-Yuan Wang g,1,Chih-Chin Kao h,i,j,*, I-Lin Tsai a,b,g,k,l,**
a Department of Biochemistry and Molecular Cell Biology, School of Medicine, College of Medicine, Taipei Medical University, Taipei,Taiwanb International Ph.D. Program for Cell Therapy and Regeneration Medicine, College of Medicine, Taipei Medical University, Taipei,Taiwanc Faculty of Medicine, Universitas Muhammadiyah Purwokerto, Purwokerto, Central Java, Indonesiad School of Food Safety, College of Nutrition, Taipei Medical University, Taipei, Taiwane Nutrition Research Center, Taipei Medical University Hospital, Taipei, Taiwanf Master Program in Food Safety, College of Nutrition, Taipei Medical University, Taipei, Taiwang Master Program in Clinical Genomics and Proteomics, College of Pharmacy, Taipei Medical University, Taipei, Taiwanh Division of Nephrology, Department of Internal Medicine, School of Medicine, College of Medicine, Taipei Medical University, Taipei,Taiwani Division of Nephrology, Department of Internal Medicine, Taipei Medical University Hospital, Taipei, Taiwanj Taipei Medical University-Research Center of Urology and Kidney (TMU-RCUK), Taipei Medical University, Taipei, Taiwank Pulmonary Research Center, Wan Fang Hospital, Taipei Medical University, Taipei, Taiwanl Graduate Institute of Medical Sciences, College of Medicine, Taipei Medical University, Taipei, Taiwan
Abstract
Exosomes are small extracellular vesicles (sEVs) secreted from cells and have a general diameter ranging from30e150 nm. It was reported that exosomes have essential roles in intercellular communication and can be targeted asbiomarkers of disease or as therapeutic agents. Among the different techniques used for exosome investigation, the massspectrometry-based proteomics approach has accelerated the unraveling of the molecular composition of exosomes andhas contributed to improved knowledge of molecular processes in various diseases. In this review, we focused onproteomics-based studies of exosomes and clinical applications in kidney diseases. A general introduction of exosomes,isolation and characterization techniques, and proteomics-based study workflows are included in this article. We alsocategorized applications in acute kidney injury, chronic kidney disease, renal transplantation, congenital kidney disease,and malignant kidney disorder to show the important findings from proteomics-based exosomal investigations.
Keywords: Exosome, Extracellular vesicle, Kidney-related diseases, Mass spectrometry, Proteomics
1. Introduction
L iving cells can secrete particles called extracel-lular vesicles (EVs). EVs are categorized into
two major categories based on their origin: exo-somes and ectosomes. Exosomes are 30e150 nm indiameter and originate from the endosome, whileectosomes are larger vesicles (50 nme1 mm in
diameter) that originate from plasma membranebudding [1].Exosomes were discovered 30 years ago. The
term "exosome" was first introduced in 1981 todescribe plasma membrane-derived vesicles [2]. In1983, two study groups, Harding’s group andJohnstone’s group, isolated transferrin receptor-associated vesicles from reticulocytes [3,4].
Received 16 October 2021; revised 16 January 2022; accepted 21 March 2022.Available online 15 June 2022.
* Corresponding author at: Department of Internal Medicine, Taipei Medical University Hospital, 252 Wuxing Street, Taipei 11031. Taiwan.** Corresponding author at: Department of Biochemistry and Molecular Cell Biology, School of Medicine, College of Medicine, Taipei Medical University,250 Wuxing Street, Taipei 11031. Taiwan.E-mail addresses: [email protected] (C-C. Kao), [email protected] (I-L. Tsai).1 Both authors contributed equally to this manuscript.
https://doi.org/10.38212/2224-6614.34092224-6614/© 2022 Taiwan Food and Drug Administration. This is an open access article under the CC-BY-NC-ND license(http://creativecommons.org/licenses/by-nc-nd/4.0/).
REVIEW
ARTIC
LE

However, the term "exosome" was not used until1984, when Salem and colleagues described theexfoliation of membrane vesicles in cultured ratglioma cells. This group referred to exfoliatedmembrane vesicles as exosomes [5]. Later, in anarticle published by Johnstone and colleagues in1987 [6], the term exosome was highlighted andbecame widely used to describe EVs that originatefrom multivesicular bodies (MVBs).In recent years, scientists began studies of exo-
somes to investigate their biological functions,and techniques used for exosome isolation andcharacterization also were being developed.Although the heterogeneous properties of exosomesmake them difficult to isolate, important findingshave been continually reported that point out theirroles in cellular communication, immune modula-tion, and pathological/therapeutic effects [1].Several review articles have summarized and
discussed the application of omics analyses to kid-ney disease-related exosomes to understand path-ological mechanisms and the potential ofdiscovering exosomal biomarkers for variousdisorders. In this review article, we focused onproteomics-based exosomal studies and their ap-plications to clinical kidney diseases. Generalintroduction to exosomes and isolation methods areincluded, followed by characterization techniques,proteomics study workflows, and key findings fromkidney disease-related exosomal studies.
1.1. Exosome biogenesis
Exosome formation occurs within the endosomalcomplex. The endosomal complex can be distin-guished into three groups: early endosomes orsorting endosomes, late endosomes, and recyclingendosomes [7]. The early endosome undergoesseveral transformations in progressing to the lateendosome. During the process, early endosomesmight fuse with endocytic vesicles and sort theircontents for recycling, degradation, or exocytosis.Contents designated for exportation are furthersorted into 30e150-nm vesicles. These multiplevesicles are referred to as intraluminal vesicles.Together with the late endosome structure, thesestructures are referred to as multivesicular bodies(MVBs) [8]. Late endosomes fuse with lysosomes orplasma membranes. Fusion with lysosomes resultsin the degradation of contents in late endosomes.Fusion with the plasma membrane leads to thesecretion of intraluminal vesicles or exosomes(Fig. 1).
1.2. Literature survey for proteomics-based exosomestudies and kidney diseases
Generally, exosomes used for clinical studies aremostly obtained from blood or urine. To furtherunderstand the trends and applications of exosomeand proteomics research in kidney-related diseases,a literature survey was performed in the PubMeddatabase through July 2021. The keywords “exo-some”, “kidney disease”, and “proteomic” wereused as advanced search options, and 115 articleswere identified, among which 85 articles involvedhuman subjects (Fig. 2). To expand the search re-sults, additional keywords addressing kidney-related diseases were used, including “acute kidneyinjury”, “diabetic nephropathy”, “glomerular dis-ease”, “polycystic kidney disease”, “renal cell carci-noma”, and “renal fibrosis”. Some inclusion criteriawere adopted during manual screening: the articlehad to be an exosome-related original article, pub-lished in 2000e2021, with humans as the mainsubject of the research. In addition, articles wereexcluded if they were not a mass spectrometric(MS)-based proteomics study. After expansion, 105human-related articles were collected. The collectedarticles consisted of 33 original articles, 35 reviewarticles, and 37 nonrelated articles. Even though weused exosome as keyword for the literature survey,some screened-out articles used EV instead of exo-some to represent their purified vesicles. It needs tobe emphasized here that the consensus of nomen-clature in using the term “EV, exosome, or micro-vesicle” has just been discussed in recent years [9]. Itis suggested to use the name “EV” if there is no clearevidence of biogenetic origin or characterization forthe purified vesicles. Although we aimed to collectarticles focusing on small EVs or exosomes, thestudies we found used different terms to representtheir purified vesicles; therefore, we will use theterm “exosome/sEV” in the clinical application partto prevent confusion.In the collected literature, the general workflow
covered exosome/sEV isolation, characterization,and bottom-up proteomics. Urine samples werethe most common source of exosomes/sEVs inthese studies. Other biofluids, such as plasma,amniotic fluid, and peritoneal dialysis effluent(PDE), also were utilized in some cases. Sinceplasma and urine are still commonly used in manyclinical studies, general sample pretreatmentmethods, isolation and characterization techniquesfor exosomes/sEVs from both sample types areintroduced in the following section (Section 2). The
JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222 203
REVIEW
ARTIC
LE

Fig. 1. Schematic representation of exosome/sEVs biogenesis. Exosome/sEVs biogenesis involves the formation of endocytic vesicles, endosomematuration, and multivesicular body (MVB) formation. Subsequently, MVBs fuse with plasma membranes and release vesicular contents, which alsoare called exosomes.
Fig. 2. Bibliographic visualization of publication trends in proteomics-based exosomal studies related to kidney diseases. Publication numbers for eachyear are indicated at the top of the purple bars, and publication numbers for studies related to human species are indicated at the top of the blue bars.
204 JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222
REVIEW
ARTIC
LE

proteomics workflow and useful tools are intro-duced in Section 3. Selected studies that appliedthese methods to clinical investigations are sum-marized and discussed in Section 4.
2. Sample pretreatment and exosome isolation
Exosomes can be purified from biological samplesbased on their nanoscale size, density, and specificsurface markers. Ultracentrifugation (UC), precipi-tation reagents, size-exclusion chromatography(SEC), and immunoaffinity capture methods havebeen frequently reported. Other techniques, such asmicrofluidic binding affinity isolation [10], field flowfractionation, and fluorescence-activated cell sorting(FACS), also have been developed [11]. Beforeapplying isolation techniques to target samples, apretreatment step is important in achieving higher-quality exosome purification.
2.1. Sample pretreatment for urine and plasmasamples
The pretreatment stage is intended to preserveexosomes and reduce cell debris. The general pro-cess usually includes the addition of protease in-hibitors followed by sequential low-speedcentrifugation to obtain a clearer sample source [12].
2.1.1. Urine sample pretreatmentSecond-morning urine is preferred to be used for
isolating urinary exosomes. Protease inhibitors areadded immediately after urine collection to preservethem [13]. The collected urine can be stored at�80 �Cuntil extraction. Vigorous vortexing is suggestedafter samples are thawed, followed by centrifugationat 17,000�g for 10e15 min to remove large particlesand cell debris [14,15]. A lower speed of initialcentrifugation also was purposed at 4,000�g for15 min at 4 �C. This approach was proven to clear celldebris without decreasing the purification yield [13].The major obstacle of urine exosome isolation is
the presence of the TammeHorsfall protein (THP)or uromodulin. This protein has polymeric networksthat might bind the exosome. Vesicle entrapmentcan be visualized during TEM observations [16].This entrapment not only disturbs exosome visual-ization but also affects total protein yields andidentified proteins in proteomic studies. To mini-mize the disturbance due to THP, dithiothreitol(DTT) is added as a reducing agent to the samplemixture, which helps dissolve the THP polymericnetwork. This approach was successfully used forsamples before applying UC and precipitation
methods. Since DTT is a strong reducing agent, itcan affect the identified proteins in proteomicstudies and might lead to false identification ordetection. Therefore, the use of DTT requiresfurther consideration. We also found one study thatshowed that it is also possible to perform a proteo-mic study while allowing uromodulin to remain insamples. Dr. Lilley et al. developed an approach toovercome uromodulin in proteomic experiments byapplying a list of excluded uromodulin-relatedpeptide ions. By including a uromodulin-relatedexclusion list during MS analyses, the number ofidentified proteins was increased by 29.7%, for atotal of 288 identified proteins, compared to theconventional method with 222 proteins [17].
2.1.2. Plasma sample pretreatmentIt was suggested to utilize plasma, instead of
serum, as the main source when isolating exosomesfrom the blood of patients and healthy individuals.The majority of vesicles in serum might be releasedfrom platelets during clot formation due to theblood-collection process [18]. Plasma from bloodsamples can be collected by using tubes with anti-coagulants and a low speed of centrifugation(1,200e1,800�g for 10 min). Before introducingplasma to exosome isolation, additional low-speedcentrifugations, such as 1,500�g for 10 min followedby 10,000�g for 20 min, can be used to removeplatelets or large vesicles [19]. Centrifugation at2,500�g for 10 min also was conducted in somestudies [20].
2.2. Exosome isolation techniques
Exosome can be isolated from multiple types ofbiofluids, including plasma/serum, saliva, urine,pleural effusions, and amniotic fluids, and they alsocan be isolated from conditioned medium of cellcultures. Until recently, there was no single bestpractice for retrieving exosomes from originalsample sources, and all methods required fine-tuning of the experimental conditions.
2.2.1. Ultracentrifugation (UC)UC is the most commonly used technique for
isolating exosomes. Based on a survey reported in2016, UC applications accounted for 81% of allmethods worldwide [21]. Centrifugation utilizescentrifugal force to separate a heterogeneousmixture based on the particle density, size, andshape. The term UC refers to remarkably high-speed centrifugation up to 100,000e120,000�g. Thepellets of dead cells and debris were discarded with
JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222 205
REVIEW
ARTIC
LE

a series of low-speed centrifugations as described inthe sample pretreatment section, and the superna-tant was used for subsequent high-speed UC toobtain exosomes. It usually takes 1e2 h to get exo-some fraction. A wash step with phosphate-bufferedsaline (PBS) is recommended to eliminate contami-nating proteins, and the sample is again centrifugedat high speed (105 �g) to recover the exosome frac-tion [15]. This method results in relatively pureexosomes and is considered the gold standard ofisolation techniques. However, possible contami-nants were reported to be lipoproteins and aggre-gate particles with similar densities when using UCfor blood exosome isolation [22].Sucrose density gradient UC was developed to
achieve better purities. With this method, 30% su-crose cushioning is added to the bottom of the UCtube. The exosome sample purified by traditionalUC is added on top of the sucrose cushion, and thetube is subjected to UC. Exosomes are trapped inthe sucrose cushion and can be collected carefully.However, co-isolation of high-density lipoprotein(HDL) still occurs because exosomes and HDL havesimilar densities (1.06e1.21 g/mL). Therefore, exo-somes and HDL can be separated only based on sizedifferences, since a single vesicle has a largerdiameter (30e1,000 nm) than an HDL particle(7e13 nm) [23]. Although UC is considered the goldstandard of exosome isolation, it is relatively time-consuming and requires high volumes of startingmaterials, and vesicles can rupture due to the highvelocities in some cases [24].
2.2.2. Precipitation methodPolyethylene glycol (PEG) is one of the most
commonly used reagents in precipitation methods.The PEG polymer has the capability to interact withwater molecules to create a polymer network fortrapping exosomes, and the trapped exosome ag-gregates can be precipitated using low-speedcentrifugation at 1,500�g [22].Commercially available precipitation reagents
include ExoQuick (System Biosciences), Total Exo-some Isolation (Invitrogen, Thermo Fisher Scienti-fic), RIBO™ Exosome Isolation Reagent (REI),Trident High-Efficiency Exosome Precipitation Re-agent (GeneTex), and Minute™ High-EfficiencyExosome Precipitation Reagent (Invent Bio-technologies). The technique is quick, simple, andrequires relatively small amounts of starting mate-rials. However, the isolated exosome sample is lesspure due to the co-precipitation of other EVs, pro-teins, and aggregates. In addition, residual polymersmight be retained in the final isolated exosomesample [24].
2.2.3. Size-exclusion chromatography (SEC)Exosome-specific sizes of 30e150 nm are advan-
tageous for size-based particle separation. SEC is aliquid chromatographic (LC) technique that sepa-rates molecules based on their size, which dependson the molecular weight and shape. It utilizes aporous polymer as the stationary phase to separateparticles in sample mixtures. The advantages of SECare that it is simple and faster, no specializedequipment is required, and it is less destructive forisolated exosomes than UC [25]. However, sincelimited sample volumes can be loaded into an SECcolumn, SEC is more suitable for isolating exosomesfrom biological fluids, such as serum/plasma andsaliva, rather than conditioned medium. Biologicalfluids usually contain relatively abundant EVs;therefore, small volumes of 0.5e2 mL are sufficientfor subsequent analyses. In contrast, exosomes arerelatively diluted in conditioned medium, and theelution process results in more diluted isolatedexosomes [26]. For example, single-step exosomeisolation using SEC was demonstrated by the teamof Dr. Nieuwland. Sepharose CL-2B pores wereused as the porous polymer inside a 10-mL syringeas the SEC column of 6.2 cm in height and 1.6 cm indiameter. Sepharose CL-2B has a pore size diameterof approximately 75 nm, which limits particles witha diameter larger than 75 nm from entering thepores and traveling along with the elution buffer.With this approach, vesicles with diameters of>75 nm can be successfully isolated [27]. The qEVSEC column from Izon Science is one of the exam-ples of commercial SEC-based exosome isolationkits. Other products include ExoPure™ from Bio-Vision and SmartSEC™ from System Biosciences.
2.2.4. Immunoaffinity-based exosome isolationImmunoaffinity exosome isolation relies on spe-
cific markers expressed by the exosome membrane.Specific surface markers include cluster of differ-entiation 63 (CD63), CD09, and CD81, which aretargeted by immunoaffinity isolation using mag-netic beads [28]. The generic workflow for exosomeisolation using magnetic capture is divided intothree steps. The first step is the pre-enrichment ofthe initial sample, which is performed by severaloptions, including UC, precipitation, filtration, andSEC. The second step is the magnetic capture ofexosome-specific surface markers, which requiresthe overnight incubation of samples and magneticbeads (Dynabeads) coated with CD9 or CD81 anti-bodies. The following day, the isolation tube isplaced on a magnet, and the magnetic beads arewashed by removing the supernatant and adding0.1% bovine serum albumin (BSA) in PBS. The final
206 JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222
REVIEW
ARTIC
LE

steps are labeling the captured exosomes anddownstream analysis, such as the Western blottingof bead-bound exosomes [28].Many studies also have developed novel affinity
purification methods for higher exosome isolationpurity. For instance, Dr. Hanayama's team utilizedan affinity protein named T cell immunoglobulindomain and mucin domain-containing protein 4(Tim4), which strongly binds phosphatidylserine topurify EVs. The binding is Ca2þ dependent, and thecaptured EVs can be easily recovered by addingCa2þ chelators [29]. Dr. Kuroda et al. also found thatlysine peptides that bind phospholipids could beused to isolate exosomes. In their study, immobi-lized peptides containing 8 and 16 lysine residues onmagnetic beads were able to capture small EVs witha size of <0.2 mm [30].
2.2.5. Sequential isolation method and noveltechniquesIn addition to single isolation methods, combined
methods also should be considered. To separateexosomes into different size groups, additional frac-tionation is required. Asymmetric flow field-flowfractionation is able to separate exosomes into sub-populations: large exosome vesicles (90e120 nm),small exosome vesicles (60e80 nm), and non-mem-branous nanoparticles termed exomeres (<35 nm)[31]. Dr. Klein et al. compared nanomembrane ul-trafiltration, UC, and the combined methods of UC-SECs for exosome isolation from nephrotic urine.The combined UC-SEC method enhances theenrichment and purification of microparticles. Withthe proteomics analysis performed using tandemmatrix-assisted laser desorption ionization-tandemtime of flight mass spectrometry (MALDI-TOF/TOFMS), neprilysin, aquaporin-2, and podocalyxin werehighly enriched by the UC-SECmethod compared tothe other two isolation methods. The improvement invesicular isolation supported the identification ofnephrotic syndrome biomarkers [32].
2.3. Exosome characterization
Exosome characterization in terms of identifica-tion, biomolecules carried, total particle numbers,and size distributions requires multiple comple-mentary techniques. In the following paragraphs,we focus only on techniques used for exosomecharacterization. Other information such as purity,copurified components, and contaminants alsoshould be clarified. Although these are not dis-cussed here, more information can be found in theMinimal Information for Studies of Extracellular Vesicles(MISEV) guidelines (2018) [9].
Exosomes have simple spherical characteristicswith a lipid bilayer membrane as they originatefrom the endosome. Even though secreted from thesame cell type, the sizes of exosomes are signifi-cantly heterogeneous. The total particle number andsize distribution of exosomes can be determinedusing nanoparticle tracking analysis (NTA), stan-dard and high-resolution flow cytometry, andtunable resistive pulse sensing (TRPS). At mini-mum, it is important to report the obtained con-centration (particle numbers/mL) along with themaximum and minimum diameters of the exo-somes. Electron microscopy (EM) is considered astandard tool for examining exosome morphology[33]. EM techniques include scanning electron mi-croscopy (SEM), transmission electron microscopy(TEM), and cryo-EM. Under SEM, exosomes appearround-spheroid-shaped and relatively uniform.Meanwhile, TEM imaging displays a centraldepression [34]. Enhanced visualization by TEMmight be achieved using cryo-TEM. Under cryo-TEM, the majority of extracellular exosomes appearas round-shaped vesicles with double-layered elec-trodense plasma membranes [35]. In several cases,immunolabeling was added to improve the EM-specific output or function-dependent morphologyconfirmation [36].Exosomes contain diverse molecules, including
DNA, messenger (m)RNA, micro (mi)RNA, lipids,metabolites, and proteins [37]. Specific proteinsenriched in exosomes can be used for vesicle char-acterization. Examples of membrane proteins includetetraspanins, such as CD63, CD81, and CD9. Therealso are cell-specific membrane proteins, such asepithelial cell-associated epithelial cell adhesionmolecule (EpCAM) and mesenchymal stem/stromalcell (MSC)-associated CD90. Examples of soluble orcytosolic proteins include endosomal complexesrequired for transport (ESCRT-I/II/III), flotillins,apoptosis-linked gene 2 interacting protein X (ALIX),etc. Immunoblotting or Western blotting is widelyused to analyze exosome markers [9], and multipleexosome markers also can be detected usingenzyme-linked immunosorbent assays (ELISAs) [38].Total protein amounts of isolated exosomes are
important to be determined for purification evalu-ation and are useful in the following proteomicsanalysis. It can be measured by colorimetricmethods such as the Bradford assay and bicincho-ninic acid (BCA) protein assay [39]. By calculatingthe ratio of exosome particle numbers and proteinconcentrations, the purity of exosomes can bedetermined [39]. One must consider that proteinquantification can be interfered with by proteincontaminants from plasma/serum or the culture
JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222 207
REVIEW
ARTIC
LE

medium. In addition to total biomolecule quantifi-cation, detailed profiling of exosome componentscan be performed through transcriptomics, lip-idomics, metabolomics, and proteomics analyses.
3. Proteomics analysis of exosomes
The protein contents of exosomes can be studiedusing MS-based analyses. With the advantages of thesensitivity and selectivity of MS, a wide range ofproteins can be identified and quantified [40]. Afterisolating exosomes from biofluids, a fraction of thesample is used for exosome characterization. A suf-ficient amount of exosomal proteins can be used forproteomics analysis. Although protein analysis byWestern blotting or ELISA is also categorized asconventional proteomics analysis, comprehensiveand quantitative bottom-up (shotgun) proteomicsstudies using MS have more frequently been appliedrecently. A successful proteomic experiment requiressuitable protein sample preparation and accurateinstrumental analysis, including peptide separationand MS detection, protein identification, pathwaymapping, and pathway interaction analysis [41]. InFig. 3, we summarize the workflow of proteomics-based exosomal studies, and detailed information onsample preparation, instrumentation, and dataanalysis are introduced in the following subsections.
3.1. Sample preparation for exosomal proteomicsanalysis
Since the sensitivity and discovery depth of MS inpeptide analysis are critically affected by the samplequality, fractionation of proteins and samplecleanup are usually necessary steps during prepa-ration. Sample pretreatment steps prior to the MSanalysis include (1) protein extraction, (2) proteinpreparation, (3) protein digestion, (4) peptidecleaning, detergent removal, and desalting, and (5)peptide separation [42].After exosome isolation, protein isolation is the
first step of sample preparation. Exosomal proteinsare usually extracted using lysis buffer such as so-dium dodecyl sulfate (SDS) or 8 M urea [43].Another option is to use physical lysis equipmentsuch as a homogenizer or sonicator. The combina-tion of chemical and physical lysis also can beapplied. During lysis, enzymes such as proteasesand phosphatase are released and may affect pro-tein profiles. Placing the sample on ice and adding acocktail of inhibitors are useful to halt enzyme ac-tivity and prevent protein loss [41].Crude extracted proteins are contaminated with
salts, detergents, and other contaminants, which
make samples incompatible with MS. Removal ofthose components is necessary, and this can beachieved using gel electrophoresis, chromatog-raphy, membrane filtration, and protein precipita-tion [41]. Gel electrophoresis can remove low-molecular-weight contaminants. In addition, thisprotein preseparation is beneficial by increasing thenumber of identified proteins, which eventually in-creases the depth of the analysis. A more advancedapproach can be achieved using filter-aided samplepreparation (FASP). FASP utilizes an ultrafiltrationdevice to separate proteins from contaminants. InFASP, the sample is solubilized in 4% SDS, con-taminants pass through an ultrafilter during centri-fugation, and proteins are retained on the filter unit[44]. The filter unit provides the function of deter-gent removal, buffer exchange, and solid supportduring protein digestion. Unlike in-gel digestion,FASP enables consecutive digestion using differentenzyme combinations [45]. Although FASP is ver-satile and efficient, sample loss occurs when thesample load contains less than 10 mg protein. This issuspected due to nonspecific protein binding to theinner membrane of the ultrafiltration device. Highprotein concentrations also lead to irreversibleprotein aggregation, which results in additionalsample loss [45]. The addition of poly-vinylpyrrolidone-40 (PVP-40) to the protein samplebefore FASP digestion improved peptide recoveryand identification. This approach also producescleaner samples than traditional FASP [46]. Dr.Lasch’s team developed a detergent-free samplepreparation method called sample preparation byeasy extraction and digestion (SPEED). It uses puretrifluoroacetic acid (TFA) for protein extraction.Compared to detergent-based sample preparations,the number of identified peptides and proteins wasincreased by >40%, and it was claimed to havesimplified sample preparation and enhanced thereproducibility of the results [47].In the protocols and methods mentioned above,
protein reduction using DTT and protein alkylationwith iodoacetamide are commonly conducted. Theprepared proteins are then digested with differentenzymes. Trypsin is the most widely used digestionenzyme due to its high specificity, availability, andpracticality. However, not all sequences are detect-able by protein tryptic digestion. Missing sequencecoverage might occur due to the uneven distributionand number of arginine and lysine residues [48],with the limitation restricting the total elucidation ofprotein segments, proteome information, and post-translational modification sites. Therefore, alterna-tive enzymes in addition to trypsin, such aschymotrypsin, Lys-C, Lys-N, Asp-N, Glu-C, and
208 JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222
REVIEW
ARTIC
LE

Arg-C, began to garner interest in proteomic studies[49]. Multiple protease utilization might be benefi-cial in large-scale mass spectrometry-based proteinsequencing, as there is an approximately 20% in-crease compared to the single protease approach,and a twofold increase in proteome sequencecoverage [50]. The peptides are further collected anddesalted using solid-phase extraction, followed byliquid chromatographic tandem MS (LC-MS/MS)analysis.
3.2. LC-MS/MS analysis
LC-MS is a high-throughput, sensitive analyticaltechnique. The technique utilizes the physical sep-aration by LC, coupled with the measurement of themass-to-charge ratio of ionized molecules by MS.The aims of MS-based proteomics studies are todetermine proteins and posttranslational modifica-tions of those proteins [51]. Reversed-phase LCcolumns, such as C18 packing material, are usuallyused as the stationary phase for peptide separation[52]. Compared to conventional high-performanceLC (HPLC) columns, nanoscale LC coupled withtandem MS (nano LC-MS/MS) has become themethod of choice because of its better sensitivity.
The inner diameter of the separation column canbe � 0.1 mm, a typical flow rate is < 1 mL/min, and itcan be utilized with limited sample volumes [53].Typical commercial nano-LC columns are 150 mmlong with a 50e75 mm inner diameter.Electrospray ionization (ESI) is the most
commonly used ion source for peptide analyses.Multiple positive charges of peptides are generatedwhen the nano-LC eluate passes through the ESIion source. Charged peptides are then analyzed by amass analyzer, such as a TOF mass analyzer, Orbi-trap, and Fourier transform-ion cyclotron resonance(FT-ICR) mass spectrometers. Some of the high-resolution MS techniques can provide resolvingpower of 105 resolution with a mass accuracy of2 ppm and 5 � 105 resolution with a mass accuracyof <2 ppm [54].There are two types of data acquisition modes for
bottom-up proteomics: data-dependent acquisition(DDA) and data-independent acquisition (DIA). TheDDA strategy selects precursor ions for fragmenta-tion based on the topmost abundant precursor ionsin MS1, the absolute signal intensity, and thecharges of the precursor ions. Peptides are identi-fied by matching the fragment spectra with a data-base of protein sequences [55]. DDA can identify
Fig. 3. General workflow of proteomics-based exosomal studies of kidney diseases. Exosomal proteomic studies were initiated by exosome isolationfrom body fluids. Exosomes/sEVs can be isolated from urine, and alternative sources are plasma and peritoneal fluids. Ultracentrifugation, size-exclusion chromatography, affinity purification, and polymer precipitation are commonly used purification methods. Exosomes/sEVs should becharacterized as a method of confirmation. After an LC-MS/MS analysis, proteins can be identified, thus enabling further data analyses and pathwayinterpretation.
JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222 209
REVIEW
ARTIC
LE

thousands of proteins; however, minor contentsmight be missing in some cases. This issue is causedby excessive coelution of peptide species and ap-pears in a single MS1 scan. Since DDA stochasticallyselects the most abundant peptides, peptides withlower abundance might be neglected [56]. Incontrast to DDA, isolation windows are pre-determined for the MS2 scan in the DIA approach.All precursor ions within the isolation windows arescanned regardless of their intensity [57]. Thisstrategy ensures the identification of all peptideswithin the selected m/z range, generally within300e1200 m/z isolation windows [52]. However, DIAMS2 scans contain peptide mixtures, which makesthem difficult to analyze. This condition also affectsthe measurement precision of very low abundancepeptides [56]. Among proteomics-based exosomalinvestigations related to kidney diseases, most sci-entists use DDA proteomics approaches.
3.3. Data interpretation and normalization
After obtaining MS/MS spectra, protein se-quences can be identified through de novosequencing or a database search. With de novosequencing, peptide sequences are obtained directlyfrom the spectra. Database searching is a commonstrategy for protein identification where a spectrumis matched against a protein database. A score isgiven for each match between theoretical and ac-quired spectra, and peptides with high matchingscores are used for protein identification [58].Many protein identification software programs
have been developed, such as Mascot, X!Tandem,SEQUEST, and OMSSA. From the open-sourcelineup, Skyline and MaxQuant are the most popularplatforms for MS-based proteomics analysis.Skyline was designed for targeted proteomicsstudies. Meanwhile, MaxQuant is a quantitativeproteomics software for mass spectrometry-basedshotgun proteomics. Database matching is robustfor already identified proteins but not for unknownproteins. This makes the number of identified pro-teins reliant on the availability of known proteins incorresponding databases, such as the UniProt andNCBI-nr protein sequence databases. The choice ofproteolytic enzymes used in digestion also alters theidentified peptide results in the database search.With proteins identified from experiments, it is
important to perform data interpretation andpathway analysis. General and kidney-specificprotein expression databases are both useful inrelated research [59]. General protein expression orsequence databases include PRIDE (PRoteomicsIDEntifications) [60], the Human Protein Atlas [61],
and UniProt [62]. For kidney-specific proteome da-tabases, HKUPP (Human Kidney and Urine Prote-ome Project) (http://www.hkupp.org/) and theUrinary Protein Biomarker Database are available[63]. ExoCarta is a compendium of exosomalcargoes and is devoted to covering exosomal pro-teins, RNAs, and lipids. The current ExoCartadatabase hosts 286 studies and records approxi-mately 41,860 protein entries. This database alsocomes with the FunRich (Functional EnrichmentAnalysis Tool), which is useful for analyses offunctional enrichment and interaction networks[64]. Other databases used for functional pathwayanalyses include the Database for Annotation,Visualization and Integrated Discovery (DAVID)[65] and Reactome [66].To discover biomarkers for clinical diseases, a
comparison of biomolecules among different groupsor samples is usually conducted. A key issue forbiomarkers from urine exosomes/sEVs is that thecomponents and concentrations of urine are alwaysvariable due to many factors, such as fluid intake,time of urine collection, biological status of the in-dividual, diet or medication, and even age. Theisolation approach also results in variation, whichrequires normalization. Normalization approachesinclude evaluating the relative excretion rate or ab-solute excretion rate. The relative excretion rate canbe calculated by dividing the abundance of bio-markers by the number of exosomes/sEVs or totalproteins from exosomes/sEVs. Meanwhile, the ab-solute excretion rate refers to biomarkers in exo-somes/sEVs excreted per unit of time, and theevaluation can be achieved through a timed collec-tion [67]. A recent report showed that urine creati-nine is highly correlated with urine EVconcentrations, suggesting the reliability of usingurine creatinine as a normalization factor for spotsamples [68]. Normalization should be taken intoaccount carefully during the study to achieve a moreaccurate investigation.
4. Proteomics-based studies of exosomes/sEVsfor kidney diseases
In this section, we summarized isolation tech-niques used in the collected clinical studies, fol-lowed by categorizing these studies into acutekidney injury, chronic kidney disease, renal trans-plantation, congenital kidney disease, and malig-nant kidney disorder. We also summarized therelated information of these studies in Table 1.Urine is one of the common sample types for
kidney disease studies, and among the screened-outarticles, ultracentrifugation (UC) is the most
210 JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222
REVIEW
ARTIC
LE
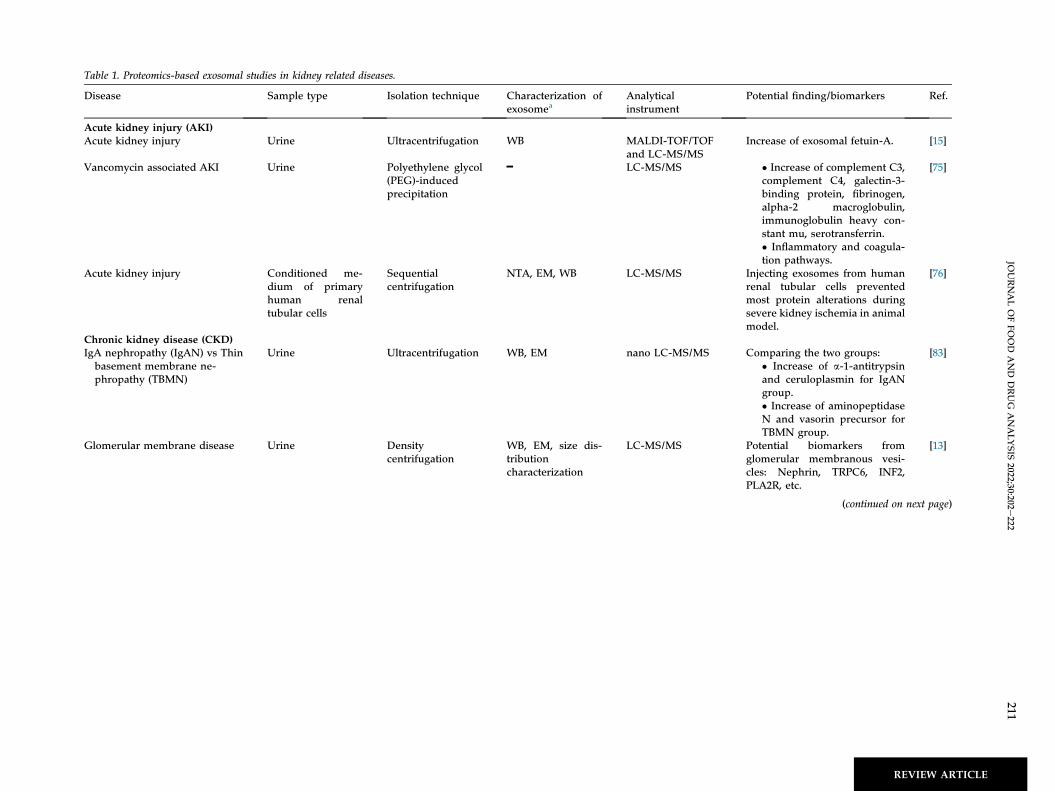
Table 1. Proteomics-based exosomal studies in kidney related diseases.
Disease Sample type Isolation technique Characterization ofexosomea
Analyticalinstrument
Potential finding/biomarkers Ref.
Acute kidney injury (AKI)Acute kidney injury Urine Ultracentrifugation WB MALDI-TOF/TOF
and LC-MS/MSIncrease of exosomal fetuin-A. [15]
Vancomycin associated AKI Urine Polyethylene glycol(PEG)-inducedprecipitation
━ LC-MS/MS � Increase of complement C3,complement C4, galectin-3-binding protein, fibrinogen,alpha-2 macroglobulin,immunoglobulin heavy con-stant mu, serotransferrin.� Inflammatory and coagula-tion pathways.
[75]
Acute kidney injury Conditioned me-dium of primaryhuman renaltubular cells
Sequentialcentrifugation
NTA, EM, WB LC-MS/MS Injecting exosomes from humanrenal tubular cells preventedmost protein alterations duringsevere kidney ischemia in animalmodel.
[76]
Chronic kidney disease (CKD)IgA nephropathy (IgAN) vs Thinbasement membrane ne-phropathy (TBMN)
Urine Ultracentrifugation WB, EM nano LC-MS/MS Comparing the two groups:� Increase of a-1-antitrypsinand ceruloplasmin for IgANgroup.� Increase of aminopeptidaseN and vasorin precursor forTBMN group.
[83]
Glomerular membrane disease Urine Densitycentrifugation
WB, EM, size dis-tributioncharacterization
LC-MS/MS Potential biomarkers fromglomerular membranous vesi-cles: Nephrin, TRPC6, INF2,PLA2R, etc.
[13]
(continued on next page)
JOURNALOFFO
OD
AND
DRUG
ANALYSIS
2022;30:202e222
211
REVIEW ARTICLE

Table 1. (continued )
Disease Sample type Isolation technique Characterization ofexosomea
Analyticalinstrument
Potential finding/biomarkers Ref.
Hypertensive nephropathy(albuminuria)
Urine Ultracentrifugation EM, Proteomicexosomal markerdetection
LC-MS/MS Pathway and proteins inresponse to hypertension andalbuminuria:
� Glycosaminoglycan degra-dation: Beta-galactosidase(GLB1), beta-glucuronidase(GUSB), N-acetyl-glucos-amine-6-sulfatase (GNS), N-acetyl-galactosamine-6-sulfa-tase (GALNS), etc.� Coagulation/Complementsystem: Antithrombin III,protein C inhibitor, C4a, C4b,C3, etc.� Oxidative stress: Cerulo-plasmin, transferrin myelo-peroxidase (MPO), etc.
[85]
Diabetic nephropathy Urine and kidneybiopsy
Ultracentrifugation EM, WB MALDI-TOF MS Downregulation of regucalcin. [88]
Diabetic nephropathy Urine Ultracentrifugation EM, WB nano LC-MS/MS � Increase of a-micro-globulin/bikunin precursor(AMBP), histone-lysine N-methyltransferase (MLL3).� Downregulation of voltage-dependent anion-selectivechannel protein 1 (VDAC1).
[14]
Immunoglobulin light chainamyloidosis (AL)
Urine Density gradientcentrifugation
WB, Immuno-goldEM
nano LC-MS/MS High molecular weight lightchain oligomers were found forAL patients (active disease).
[90]
Immunoglobulin light chainamyloidosis (AL)
Urine Density gradientcentrifugation
WB nano LC-MS/MS &MALDI-TOF MS
Two monoclonal lambda lightchains were identified.
[91]
End-stage renal disease Peritoneal dialysiseffluent (PDE)
Size-exclusionchromatography(SEC)
Flow cytometry an-alyses (anti-CD9,anti-CD63)
nano LC-MS/MS Enriched proteins were found inthe PD patients with stable peri-toneal membrane (PM) func-tions: Endoglin (ENG), Thy-1membrane glycoprotein (THY-1or CD90), biglycan (BGN), kini-nogen-1 (KNG1).
[94]
Renal transplantationRenal transplantation Conditioned me-
dium of humanproximal tubularepithelial cells,serum (murine)
Ultracentrifugation EM, Immunoblot-ting, Flowcytometry
nano-LC-MS/MS Vascular injury enhanced theproteasome activity in apoptoticexosome-like vesicles whichincreased the production of renaldamage-related autoantibodies(anti-LG3).
[96]
212JO
URNALOFFO
OD
AND
DRUG
ANALYSIS
2022;30:202e222
REVIEWARTICLE

Acute T cell-mediated rejectionin kidney transplant (TCMR)
Urine Ultracentrifugation EM, WB nano-UPLC MS/MS
Apolipoprotein A1, Hemopexin,Polymeric immunoglobulin re-ceptor, Lectin galactoside-bind-ing soluble 3 binding protein,Tetraspanin-1 were potentialdiagnostic proteins.
[97]
Renal transplantation andmedication
Urine Size-exclusionchromatography(SEC)
Flow cytometry(CD9 and CD63)
LC-MS/MS Proteins of uroplakin (UPK) andplakin families were significantlyupregulated in the patients withcalcineurin inhibitors toxicity.
[99]
Kidney related congenital diseasesAutosomal dominant polycystickidney disease (ADPKD)
Urine Sucrose densitygradientultracentrifugation
EM, WB Mass spectrometry Polycystine1 (PC-1), PC2, fibro-cystin/polyductin (FCP), andtheir interacting proteins wereidentified. Cystin and ADP-ribo-sylation factor-like 6 were detec-ted which were involved in cysticdisease.
[102]
Autosomal dominant polycystickidney disease (ADPKD)
Urine Sucrose densitygradientultracentrifugation
EM, WB nano-LC-MS/MS Post-translational proteolyticprocesses were discovered forpolycystine1 (PC1), PC2, andfibrocystin in human urine exo-some like vesicles.
[103]
Autosomal dominant polycystickidney disease (ADPKD)
Urine Sucrose densitygradientultracentrifugation
EM, WB nano-LC-MS/MS Ratios of exosomal polycystin-1/transmembrane protein 2 (PC1/TMEM2) or polycystin-2/trans-membrane protein 2 (PC2/TMEM2) have potential to beindicators for polycystic kidneydisease.
[104]
Autosomal dominant polycystickidney disease (ADPKD)
Urine Ultracentrifugation WB LC-MS/MS Periplakin, envoplakin, villin-1,and complement C3 and C9 werehighly expressed in exosomesfrom ADPKD patients.
[105]
Autosomal dominant polycystickidney disease (ADPKD) &Medullary sponge kidney(MSK)
Urine Ultracentrifugation Dynamic light scat-tering, WB, Flowcytometry, ELISA
nano LC-MS/MS � Higher level in ADPKD:Prominin 1 (CD133), cellularrepressor of E1A stimulatedgenes 1 (CREG1), Inter-alpha-trypsin inhibitor heavychain 5 (ITIH5), Guanylatecyclase activator 2B(GUCA2B), Myelin andlymphocyte protein (MAL).� Higher level in MSK:Osteopontin (SPP1).
[38]
(continued on next page)
JOURNALOFFO
OD
AND
DRUG
ANALYSIS
2022;30:202e222
213
REVIEW ARTICLE

Table 1. (continued )
Disease Sample type Isolation technique Characterization ofexosomea
Analyticalinstrument
Potential finding/biomarkers Ref.
Medullary sponge kidney (MSK)disease
Urine Ultracentrifugation Dynamic light scat-tering, WB, ELISA
nano LC-MS/MS Ficolin 1 (FCN1) and comple-ment component 4-binding pro-tein beta (C4BPB) were moreabundant in the exosomes fromMSK patients. Lectin comple-ment pathway was pointed out tobe associated with MSK.
[106]
Ureteropelvic junction obstruc-tion (UPJO)
Amniotic fluid Exosome extractionkit
EM, WB LC-MS/MS Amniotic fluid exosomes frompatient with congenital UPJOhad less angiotensin-convertingenzyme (ACE) and aminopepti-dase N (AP-N).
[108]
Galactosemia related kidneyinsufficiency
Urine Differentialcentrifugation
WB LC fractionation,MALDI-MS/MS
Premature protein dysglycosyla-tion disrupts the renal basementmembrane.
[109]
Cystinuria Urine Ultracentrifugation EM, WB nano LC-MS/MS, Protein rBAT which was codedby the SLC3A1 gene, was detec-ted only in healthy controls. Theother 38 proteins which wereoverexpressed in patients withcystinuria were associated withkidney injury, circulating pro-teins, and neutrophil signature.
[111]
Kidney malignancyRenal cell carcinoma (RCC) Urine Differential
centrifugationEM, WB, exosomesizecharacterization
LC-MS/MS Exosomal proteins from RCCpatients were different fromhealthy controls: Matrix metal-loproteinase 9 (MMP-9), Cerulo-plasmin (CP), Podocalyxin(PODXL), Dickkopf related pro-tein 4 (DKK4), and CarbonicAnhydrase IX (CAIX).
[113]
Kidney cancer and Chronic kid-ney disease (CKD)
Plasma Chemical affinitypurification(amphiphilic beads)
EM, NTA, Tunableresistive pulsesensing, WB
nano LC-MS/MS, The cardiomyopathy-associatedprotein 5 (CMYA5), phosphory-lated Crk-like protein (CRKL),and LYRIC (MTDH) were higherin the kidney cancer group.The increase of apolipoproteinA-IV (APOA4) was found forCKD.
[20]
a Some studies provided concise methods or the characterizations were referred to their previously published work. WB: Western blot; EM: Electron microscopy; NTA: nanoparticletracking analysis.
214JO
URNALOFFO
OD
AND
DRUG
ANALYSIS
2022;30:202e222
REVIEWARTICLE

commonly used method to purify uEVs. However,the recovery yield needs to be evaluated carefullyfor UC to prevent the loss of important proteins [69].Other isolation methods include sucrose densitygradient ultracentrifugation, differential centrifuga-tion, size-exclusion chromatography, polyethyleneglycol (PEG)-induced precipitation, and exosomeextraction kits were used in clinical studies as well(Table 1).
4.1. Acute kidney injury (AKI)
AKI is characterized by swift escalation of serumcreatinine, an accelerated reduction in urine output,or a combination of these two conditions. AKI wasmainly caused by the mismatch of oxygen/nutrientdelivery and nephron energy demand [70]. Com-mon causes include cardiorenal syndrome, shock,transplantation, sepsis, drug-related necrosis, andurinary tract obstruction, which cause impairedperfusion [71]. Novel AKI biomarkers, namely, tis-sue inhibitor of metalloproteinase-2, neutrophilgelatinase-associated lipocalin, kidney injurymarker-1, and cystatin C, may provide earlierdiagnostic and prognostic value. However, moreinvestigations are needed to define the clinical rolesof these biomarkers [72].Many drugs have been reported to cause AKI,
including antimicrobial agents, NSAIDs, herbalmedicines, chemotherapeutic agents, and check-point inhibitors [73]. Proteomics analyses of uri-nary exosomes/sEVs have been applied to discovermedication-induced AKI biomarkers such ascisplatin and vancomycin. Renal tubular cells arevulnerable to the toxic effects of medications. Dr.Star et al. found that fetuin-A was increased inurinary exosomes/sEVs in both an animal modeland patients with cisplatin-induced AKI. The exactfunctions of fetuin-A in AKI remain elusive; how-ever, it was hypothesized that exosomes/sEVscontaining fetuin-A may be secreted fromdamaged proximal tubule cells [15]. Increasedurinary ATF3 (activating transcription factor 3)protein was reported in acute kidney injury bycisplatin or ischemia-reperfusion injury [74]. Theincrease in ATF3 in urinary exosomes/sEVs pre-cedes serum creatinine elevation and may serve asan early biomarker. Therefore, urinary exosomalfetuin-A and ATF3 might serve as indicators ofkidney tubular injury. In the case of vancomycin-related (V)-AKI, complement C3 and C4 aresignificantly increased in urinary exosomes/sEVs.According to the results, inflammation and thecomplement system might play important roles invancomycin-related kidney injury [75].
In addition to research on biomarkers, the appli-cation of exosomes/sEVs in AKI therapy also wasevaluated. In a hypoxic AKI model of athymic nuderats, injections of exosomes/sEVs originating fromharvested human kidneys prevented apoptosis ofkidney cells and preserved the microvascular andrenal tubular structures. These results suggestedthat exosomes/sEVs might act as therapeutic agentsin kidney diseases [76].
4.2. Chronic kidney disease (CKD)
CKD is a pathological condition characterized bythe gradual loss of normal kidney functions andkidney structural changes. While this medical con-dition slowly progresses, the damage is irreversible.The disease prevalence is approximately 10%e13%of the population and is associated with increasedcardiovascular risks [77]. Currently, CKD is definedby a glomerular filtration rate (GFR) of <60 mL/min/1.73 m2, microalbuminuria, or an indication of kid-ney failure [78]. Glomerulonephritis, hypertension,and diabetes are common causes of CKD.
4.2.1. GlomerulonephritisGlomerulonephritis (GN) refers to inflammation
of glomeruli in response to infections, malignancies,drugs, toxins, or autoimmune diseases [79]. PrimaryGN is kidney oriented without accompanying con-ditions, such as idiopathic nephrotic syndrome, thinbasement membrane nephropathy (TBMN), IgAnephropathy (IgAN), Alport syndrome, and mem-branous nephropathy. Secondary GN is morerelated to systemic diseases such as lupus nephritis,diabetic nephropathy, and postinfectious GN [80].Glomerular injuries can disturb the glomerularfiltration barrier and result in a decreased glomer-ular filtration rate (GFR), hematuria, and proteinuria[81]. Currently, the gold standard for diagnosingglomerulonephritis is kidney biopsy, which isinvasive and inconvenient. Therefore, novel bio-markers would substantially improve the diagnosisand therapy of this disease [82]. Isolated microscopichematuria is mostly due to Alport syndrome,immunoglobulin A nephropathy (IgAN), and thinbasement membrane nephropathy (TBMN).Although IgAN and TBMN have identical clinicalpresentations, patients with IgAN develop worseclinical outcomes than those with TBMN. Currently,there are no diagnostic biomarkers, and a renal bi-opsy is the only diagnostic tool for differentiatingIgAN from TBMN. From the proteomics profiling ofurinary exosomes/sEVs, aminopeptidase N, vasorinprecursor, a-1-antitrypsin, and ceruloplasmin wereshown to be potential biomarkers to differentiate
JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222 215
REVIEW
ARTIC
LE

early IgAN from TBMN. IgAN was found to havehigher a-1-antitrypsin, which could induce fibrino-lysis in response to hematuria, and the increase inceruloplasmin is an indication of mesangialinflammation. The TBMN group with higheraminopeptidase N is capable of modulating MAPKpathways, galectin (Lgals) molecules, Fc receptors,and immune regulators. The higher vasorin pre-cursor was reported to be associated with the vesselrepair process [83].Dr. Hogan et al. isolated glomerular membrane
vesicles (GMVs) from human urine samples andapplied proteomics analysis for biomolecule inves-tigation. In the GMV-enriched samples from 3 pa-tients with glomerular membrane diseases, 5657unique proteins were identified. Some of them wereassociated with nephrotic syndrome, includingnephrin, short transient receptor potential channel 6(TRPC6), inverted formin 2 (INF2), and secretoryphospholipase A2 receptor (PLA2R). Although theseidentified proteins require further investigation asclinical biomarkers, the results of this study showedthat GMVs have the potential to be used forbiomarker discovery [13].
4.2.2. Hypertensive nephropathy and diabeticnephropathyThe relationship between hypertension and
chronic kidney disease (CKD) is a reciprocalcausation. Hypertension can result in renal impair-ment, and CKD can contribute to the developmentof secondary hypertension [84]. The renin-angio-tensin system (RAS) plays an important role inblood pressure regulation. RAS suppression isusually found in hypertensive patients. However,even with the presence of RAS suppression, albu-minuria occurs in some cases in which the mecha-nism is unclear. Dr. Gonzalez-Calero and Ruilopeet al. investigated urinary exosomes/sEVs from pa-tients with normoalbuminuria, de novo albuminuria,and maintained albuminuria. In their study, threefunctional clusters, including glycosaminoglycandegradation, the coagulation/complement system,and oxidative stress, were identified [85]. The hitproteins and mapped pathways were mostly relatedto glomerular basement membrane (GBM) disrup-tion and proinflammatory signs.Diabetic nephropathy (DN), also known as dia-
betic kidney disease, is characterized by perpetualalbuminuria and a gradual diminution of normalrenal functions. DN occurs in 20%e50% of patientswith diabetes mellitus (DM), and disease progres-sion arises in both type 1 and 2 DM. Since DNsilently progresses and shows no obvious clinicalsymptoms in the early stages, a potential biomarker
of this disease needs to be discovered. The currentnoninvasive marker is microalbuminuria; however,this marker is not very sensitive and cannot predictthe prognosis [86,87]. Dr. Alvarez-Llamas et al.found that the expression of regucalcin was reducedin human kidney tissues and in urinary exosomes/sEVs from DN patients. Regucalcin plays essentialroles in regulating intracellular calcium transport,cell signaling, DNA synthesis, gene expression,proliferation, and apoptosis in kidney cells. Sinceurine is an easily accessible and noninvasive bio-fluid, regucalcin of urinary exosomes/sEVs might bea novel target for the early diagnosis of DN [88]. Inaddition to regucalcin, a-microglobulin/bikuninprecursor (AMBP) and histone-lysine N-methyl-transferase (MLL3) were found at higher levels inDN patients. AMBP is a membrane glycoproteinwith serine protease inhibitor activity, and MLL3 isa histone methyltransferase. In contrast, voltage-dependent anion-selective channel protein 1(VDAC1) was decreased in DN patient urinaryexosomes/sEVs. It is a transmembrane channel thatplays a crucial role in regulating cell survival,growth, and death [14].
4.2.3. Amyloidosis nephropathyAmyloidosis is a rare disease characterized by
misfolded amyloid deposition in particular tissues,and it can lead to organ failure. Kidney tissues areone of the main sites of amyloid deposition [89]. Dr.Ramirez-Alvarado et al. investigated the proteomicprofiling of urinary exosomes/sEVs from AL pa-tients and revealed enormous numbers of free light-chain immunoglobulins. In addition, high-molecu-lar-weight oligomers (light chain decamers) werespecifically found in patients with light-chainamyloidosis [90]. They further analyzed proteins ofurinary exosomes/sEVs by MS and identified twomonoclonal lambda light chains that could not bedetected by current clinical methods. Immuno-globulin light-chain oligomers in urinary exosomes/sEVs were correlated with the formation of amyloidand might predict the prognosis of immunoglobulinlight-chain (AL) amyloidosis [91].
4.2.4. End-stage renal disease (ESRD)ESRD is referred to as the final stage of chronic
kidney disease. To define ESRD, uremia must bepresent, and the patient needs renal replacementtherapy. Alternatively, ESRD might be defined as acondition with an estimated glomerular filtrationrate (eGFR) < 15 mL/min/1.73 m2 [92]. Hemodial-ysis and peritoneal dialysis are two treatment op-tions for ESRD patients. Peritoneal dialysis effluent(PDE) contains a diverse variety of cells and
216 JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222
REVIEW
ARTIC
LE

secretomes that reflect the pathophysiological stateof patients, and exosomes/sEVs can be isolatedfrom PDE [93]. A longitudinal study of peritonealdialysis patients was conducted by Dr. Troya-Sab-orido and Dr. Borr�as et al. to compare peritonealdialysis effluent containing extracellular vesicle(PDE-EV) proteomes related to the stability ofperitoneal membrane (PM) functions. The resultsrevealed that PDE-EVs of patients with stable PMfunctions were enriched in several proteins,including endoglin (ENG), Thy-1 membraneglycoprotein (THY-1 or CD90), biglycan (BGN), andkininogen-1 (KNG1). These proteins were found tobe associated with the ultrafiltration capacity andthe functional status of PM [94].
4.3. Renal transplantation
Large-scale analysis of urinary exosomes/sEVsfrom renal allografts has been used to discoverbiomarkers of graft rejection or tubular injury [95].Vascular injury enhanced proteasome activity inapoptotic exosome-like vesicles, which furtherincreased the production of renal damage-relatedautoantibodies, such as anti-LG3 IgG. The in-crease in anti-LG3 is known to be correlated withan increased risk of rejection post-transplantation[96]. In another report, tetraspanin-1 (TSPAN1),hemopexin (HPX), and the other 3 proteins weresignificantly increased in urine exosomes/sEVsfrom patients with acute T cell-mediated rejection(TCMR) compared to those with stablegraft function. The results revealed that exosomeproteins could be potential markers for TCMR[97].A proteomics study of urine exosomes/sEVs also
was conducted to discover biomarkers of immu-nosuppressant toxicity for renal transplant re-cipients. It was found that the use of calcineurininhibitors (CNIs) might lead to nephrotoxicity (CNItoxicity, CNIT) [98]. There were differences in pro-teomic profiles among renal transplant recipientswith normal kidney function, the CNIT group, andthe interstitial fibrosis/tubular atrophy (IFTA)group. Proteins of the uroplakin (UPK) and plakinfamilies, including UPK1A, UPK1B, UPK2, UPK3A,envoplakin (EVPL), and periplakin (PPL), weresignificantly upregulated in the CNIT group, whichmight have an important role in CNIT processes[99].
4.4. Congenital kidney diseases
Autosomal dominant polycystic kidney disease(ADPKD) is an inherited genetic disorder
characterized by numerous cysts and enormousbilateral kidney enlargement. Exosomes/sEVs fromADPKD urine samples can potentially serve as anoninvasive disease biomarker [100,101]. Dr. Wardet al. successfully identified polycystin-1 (PC1), PC2,fibrocystin/polyductin (FCP), and their interactingproteins from urinary exosome-like vesicles ofADPKD patients [102]. Posttranslational proteolyticprocesses also were found for PC1, PC2, and FCPproteins by studying urinary exosome-like vesiclesthat affect the progression of cystic disease [103]. Inaddition, PC1 and PC2 were found to be lower inurinary vesicles from patients with PKD1 mutations,and transmembrane protein 2 (TMEM2) was foundto have higher levels. The PC1/TMEM2 or PC2/TEME2 ratios were indicated as potential indicatorsfor ADPKD [104]. Comparing the exosomal proteinsbetween patients with ADPKD and medullarysponge kidney (MSK), Dr. Zaza et al. found higherexpression of prominin-1 (CD133), cellularrepressor of E1A-stimulated genes 1 (CREG1),interalpha-trypsin inhibitor heavy chain 5 (ITIH5),and guanylate cyclase activator 2B (GUCA2B) inADPKD patients [38]. In addition, complement C3,C9, plakins, and villin-1 were highly expressed inurinary exosomes/sEVs from ADPKD patientscompared to healthy controls [105].Medullary sponge kidney (MSK) disease is a rare
kidney disease characterized by cystic anomalies inthe precalyceal ducts and nephrocalcinosis. Byanalyzing the proteomics profiles of urine exo-somes/sEVs, Dr. Zaza and the team found that thelectin complement pathway was associated with thepathogenesis of the disease. Ficolin 1 (FCN1),mannan-binding lectin-associated serine protease-2(MASP2), and complement component 4-bindingprotein beta (C4BPB) are three proteins that can beused to discriminate the MSK group and the groupof patients with idiopathic calcium nephrolithiasis(ICN) [106].Ureteropelvic junction obstruction (UPJO) is an
obstruction of the urinary tract that results inreduced urine flow from the renal pelvis into theureter. Antenatal ultrasound can be used to detectcongenital UPJO during the second trimester [107].Dr. Wei et al. found that exosomes/sEVs from theamniotic fluid of patients with congenital UPJOhave significant alterations in the protein profiles interms of decreases in angiotensin-convertingenzyme (ACE) and aminopeptidase N (AP-N). Thebiological effects of ACE and AP-N reduction werestudied in vitro and were discovered to be associatedwith the curb of renal tubular epithelial cell prolif-eration, overproduction of reactive oxygen species,and enhanced inflammation [108].
JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222 217
REVIEW
ARTIC
LE

Galactosemia is caused by a deficiency of galac-tose-1-phosphate uridyltransferase due to a muta-tion of the GALT gene, which causes severe defectsin galactose metabolism. The proteomic profile fromurinary exosomes/sEVs depicts alterations related torenal failure, indicating drastic increases in abun-dant serum glycoproteins, including leucine-rich a-2-glycoprotein, albumin, fetuin, prostaglandin H2D-isomerase, a-1-microglobulin protein, and im-munoglobulins. It was hypothesized that renal fail-ure might occur due to premature proteindysglycosylation, which interferes with the forma-tion of the renal basement membrane. However,markers from urinary exosomes/sEVs of galacto-semia patients overlap with urinary protein markersfor IgA nephropathy, thin basement membranenephropathy, and acute kidney injury, making itdifficult to differentiate proteomic profiles acrossthese diseases [109].Cystinuria is an autosomal recessive disorder with
defects in the SLC3A1 and SLC7A9 genes [110]. Theresults of the exosomal proteomics study showedthat the rBAT protein, encoded by the SLC3A1 gene,was detected only in healthy individuals. Mean-while, 38 proteins were overexpressed in thecystinuria groups. The results suggested that one-quarter of overexpressed proteins in cystinuria pa-tients were neutrophil-derived, which might indi-cate intrarenal inflammatory reactions due tomicroscopic crystal formation [111].
4.5. Kidney malignancies
The roles of exosomes/sEVs in malignancies havebeen extensively studied; however, the majority ofexosome studies of renal cell carcinoma (RCC) havefocused on miRNAs [112]. Proteomic profiling ofurinary exosomes/sEVs from RCC patients showeddistinct protein profiles compared to urinary exo-somes/sEVs from healthy controls. The urinaryexosomal proteomic analysis of RCC patientsshowed enhanced expression of podocalyxin, ceru-loplasmin, matrix metalloproteinase 9, carbonicanhydrase IX, and dickkopf-related protein 4. Incontrast, the extracellular matrix metalloproteinaseinducers aquaporin-1, dipeptidase-1, syntenin-1,and neprilysin (CD10) exhibited significantlyreduced expression. From protein profiling andvalidation, urinary exosomes/sEVs manifested someportions of cancer protein profiles and might beinvolved in RCC progression [113]. Plasma EVspurified by using chemical affinity beads were alsoinvestigated for kidney cancer. In the study,
cardiomyopathy-associated protein 5 (CMYA5),phosphorylated Crk-like protein (CRKL), andphosphorylated LYRIC (MTDH) were found to behigher in the kidney cancer group [20].
4.6. Other biological findings from uEVs for kidneyfunction and disease
Recently, a large-scale animal study showed thatthe abundances of proteins in urine EVs (uEVs)are highly correlated with proteins in respectivekidney tissues. Positive correlations were foundfor approximately 1000 proteins from uEVs andkidney tissues, which reveals that investigatinguEV proteins is able to monitor particular (patho)physiological changes even without using renalbiopsy [114]. More clinical applications can befound when using uEV or exosome as the keywordin combination with specific kidney diseases. Forexample, Gonzales et al. investigated proteomicsand phosphoproteomics for human urine exo-somes. They successfully identified 1132 unam-biguous proteins, including solute and watertransporters, vacuolar Hþ-ATPase subunits, and 34proteins related to renal diseases. Through phos-phoproteomics investigation, serine-256-phos-phorylated AQP2 in uEVs was identified as apotential marker for monitoring the state ofvasopressin activation. The absence of sodium-potassium-chloride cotransporter 2 (NKCC2) inurine exosomes from patients with Bartter syn-drome was also reported in this study [115]. Inaddition, Raimondo et al. investigated the prote-ome of uEVs and set up a panel of 5 proteins,including carbonic anhydrase (CA2), vacuolar-typeATPase B subunit 1 (VATB1), Annexin A2(ANXA2), sodium chloride cotransporter (NCC),and NKCC2, which can differentiate Gitelman andBartter syndromes (salt-losing tubulopathies)[116]. Sung et al. also found that NCC and phos-phorylated-NCC were decreased in uEVs frompatients with Gitelman syndrome, while Naþ-hydrogen exchanger 3 (NHE3), epithelial Naþ
channel b (ENaCb), and pendrin were increased[117]. By investigating urinary microvesicles, Roodet al. found the increase of lysosome membraneprotein 2 (LIMP-2) from patients with idiopathicmembranous nephropathy, and the upregulationof LIMP-2 was identified in renal biopsies that theprotein was co-localized with IgG along theglomerular basement membrane [118]. All thestudies showed the potential of investigating uEVsfor kidney disease research.
218 JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222
REVIEW
ARTIC
LE

5. Summary and perspective
For kidney-related diseases, urinary exosomes/sEVs serve as a noninvasive biopsy and providebulk information on kidney-related diseases. Somestudies also investigated plasma exosomes/sEVs orthose from primary culture conditioned medium forpathological and biomarker discovery. Despite thediverse methods of urinary exosome/sEV isolation,ultracentrifugation was currently found to be themost popular tool. Characterization based on therepresentative protein markers, size distribution,and morphology observation is required to ensurethe precision of downstream analyses. With thenecessity and the emergence of novel techniques,exosomal proteomic analyses of clinical samples willenable the elucidation of disease-related molecularmechanisms and enhance our understanding ofkidney-related diseases.
Conflicts of interest
The authors declare no conflict of interest.
Acknowledgments
The authors acknowledge the academic and sci-ence graphic illustration service provided by theTMU Office of Research and Development. Thiswork was financially supported by the Higher Ed-ucation Sprout Project of the Ministry of Education(MOE) in Taiwan (DP2-110-21121-01-T-03-03).
References
[1] Kalluri R, LeBleu VS. The biology, function, and biomedicalapplications of exosomes. Science 2020;367:1e15.
[2] Trams EG, Lauter CJ, Norman Jr S, Heine U. Exfoliation ofmembrane ecto-enzymes in the form of micro-vesicles.Biochim Biophys Acta Biomembr 1981;645:63e70.
[3] Pan BT, Johnstone RM. Fate of the transferrinreceptor during maturation of sheep reticulocytes in vitro:Selective externalization of the receptor. Cell 1983;33:967e78.
[4] Harding C, Heuser J, Stahl P. Receptor-mediated endocy-tosis of transferrin and recycling of the transferrin receptorin rat reticulocytes. J Cell Biol 1983;97:329e39.
[5] Kassis S, Lauter CJ, Stojanov M, Salem Jr N. Exfoliation ofthe beta-adrenergic receptor and the regulatory compo-nents of adenylate cyclase by cultured rat glioma C6 cells.Biochim Biophys Acta 1986;886:474e82.
[6] Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C.Vesicle formation during reticulocyte maturation. Associa-tion of plasma membrane activities with released vesicles(exosomes). J Biol Chem 1987;262:9412e20.
[7] Naslavsky N, Caplan S. The enigmatic endosome e sortingthe ins and outs of endocytic trafficking. J Cell Sci 2018;131:1e14.
[8] Klumperman J, Raposo G. The complex ultrastructure ofthe endolysosomal system. Cold Spring Harbor PerspectBiol 2014;6:1e22.
[9] Th�ery C, Witwer KW, Aikawa E, Alcaraz MJ, Anderson JD,Andriantsitohaina R, et al. Minimal information for studies
of extracellular vesicles 2018 (MISEV2018): A positionstatement of the International Society for ExtracellularVesicles and update of the MISEV2014 guidelines.J Extracell Vesicles 2018;7:1e43.
[10] Kang YT, Purcell E, Palacios Rolston C, Lo TW, Ramnath N,Jolly S, et al. Isolation and profiling of circulating tumor-associated exosomes using extracellular vesicularlipideprotein binding affinity based microfluidic device.Small 2019;15:1e14.
[11] Stranska R, Gysbrechts L, Wouters J, Vermeersch P,Bloch K, Dierickx D, et al. Comparison of membrane af-finity-based method with size-exclusion chromatographyfor isolation of exosome-like vesicles from human plasma.J Transl Med 2018;16:1e9.
[12] Zubiri I, Vivanco F, Alvarez-Llamas G. Proteomic analysisof urinary exosomes in cardiovascular and associated kid-ney diseases by two-dimensional electrophoresis and LC-MS/MS. Methods Mol Biol 2013;1000:209e20.
[13] Hogan MC, Johnson KL, Zenka RM, Charlesworth MC,Madden BJ, Mahoney DW, et al. Subfractionation, charac-terization, and in-depth proteomic analysis of glomerularmembrane vesicles in human urine. Kidney Int 2014;85:1225e37.
[14] Zubiri I, Posada-Ayala M, Sanz-Maroto A, Calvo E, Martin-Lorenzo M, Gonzalez-Calero L, et al. Diabetic nephropathyinduces changes in the proteome of human urinary exo-somes as revealed by label-free comparative analysis.J Proteonomics 2014;96:92e102.
[15] Zhou H, Pisitkun T, Aponte A, Yuen PST, Hoffert JD,Yasuda H, et al. Exosomal Fetuin-A identified by prote-omics: A novel urinary biomarker for detecting acute kid-ney injury. Kidney Int 2006;70:1847e57.
[16] Hiemstra TF, Charles PD, Gracia T, Hester SS, Gatto L, Al-Lamki R, et al. Human urinary exosomes as innate immuneeffectors. J Am Soc Nephrol 2014;25:2017e27.
[17] Hiemstra TF, Charles PD, Hester SS, Karet FE, Lilley KS.Uromodulin exclusion list improves urinary exosomalprotein identification. J Biomol Tech 2011;22:136e45.
[18] Witwer KW, Buz�as EI, Bemis LT, Bora A, L€asser C, L€otvall J,et al. Standardization of sample collection, isolation andanalysis methods in extracellular vesicle research.J Extracell Vesicles 2013;2:1e25.
[19] Lobb RJ, Becker M, Wen Wen S, Wong CSF, Wiegmans AP,Leimgruber A, et al. Optimized exosome isolation protocolfor cell culture supernatant and human plasma. J ExtracellVesicles 2015;4:1e11.
[20] Iliuk A, Wu X, Li L, Sun J, Hadisurya M, Boris RS, et al.Plasma-derived extracellular vesicle phosphoproteomicsthrough chemical affinity purification. J Proteome Res 2020;19:2563e74.
[21] Gardiner C, Vizio DD, Sahoo S, Th�ery C, Witwer KW,Wauben M, et al. Techniques used for the isolation andcharacterization of extracellular vesicles: Results of aworldwide survey. J Extracell Vesicles 2016;5:1e6.
[22] Sidhom K, Obi PO, Saleem A. A review of exosomal isola-tion methods: Is size-exclusion chromatography the bestoption? Int J Mol Sci 2020;21:1e19.
[23] Brennan K, Martin K, FitzGerald SP, O'Sullivan J, Wu Y,Blanco A, et al. A comparison of methods for the isolationand separation of extracellular vesicles from protein andlipid particles in human serum. Sci Rep 2020;10:1e13.
[24] Chen BY, Sung CWH, Chen C, Cheng CM, Lin DPC,Huang CT, et al. Advances in exosomes technology. ClinChim Acta 2019;493:14e9.
[25] Guan S, Yu H, Yan G, Gao M, Sun W, Zhang X. Charac-terization of urinary exosomes purified with size-exclusionchromatography and ultracentrifugation. J Proteome Res2020;19:2217e25.
[26] Guerreiro EM, Vestad B, Steffensen LA, Aass HCD,Saeed M, Øvstebø R, et al. Efficient extracellular vesicleisolation by combining cell media modifications, ultrafil-tration, and size-exclusion chromatography. PLoS One2018;13:1e17.
JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222 219
REVIEW
ARTIC
LE

[27] B€oing AN, van der Pol E, Grootemaat AE, Coumans FAW,Sturk A, Nieuwland R. Single-step isolation of extracellularvesicles by size-exclusion chromatography. J ExtracellVesicles 2014;3:1e11.
[28] Pedersen KW, Kierulf B, Neurauter A. Specific and genericisolation of extracellular vesicles with magnetic beads. In:Kuo WP, Jia S, editors. Extracellular vesicles: Methods andprotocols. New York: Springer; 2017. p. 65e87.
[29] Nakai W, Yoshida T, Diez D, Miyatake Y, Nishibu T,Imawaka N, et al. A novel affinity-based method for theisolation of highly purified extracellular vesicles. Sci Rep2016;6:1e11.
[30] Ishida T, Hashimoto T, Masaki K, Funabashi H, Hirota R,Ikeda T, et al. Application of peptides with an affinityfor phospholipid membranes during the automated -purification of extracellular vesicles. Sci Rep 2020;10:1e12.
[31] Zhang H, Freitas D, Kim HS, Fabijanic K, Li Z, Chen H,et al. Identification of distinct nanoparticles and subsets ofextracellular vesicles by asymmetric flow field-flow frac-tionation. Nat Cell Biol 2018;20:332e43.
[32] Rood IM, Deegens JK, Merchant ML, Tamboer WP,Wilkey DW, Wetzels JF, et al. Comparison of three methodsfor isolation of urinary microvesicles to identify biomarkersof nephrotic syndrome. Kidney Int 2010;78:810e6.
[33] Chuo STY, Chien JCY, Lai CPK. Imaging extracellularvesicles: Current and emerging methods. J Biomed Sci 2018;25:1e10.
[34] Wu Y, DengW, Klinke DJ, II. Exosomes: Improved methodsto characterize their morphology, RNA content, and surfaceprotein biomarkers. Analyst 2015;140:6631e42.
[35] Musante L, Bontha SV, La Salvia S, Fernandez-Pi~neros A,Lannigan J, Le TH, et al. Rigorous characterization of uri-nary extracellular vesicles (uEVs) in the low centrifugationpellet - a neglected source for uEVs. Sci Rep 2020;10:1e14.
[36] Jung MK, Mun JY. Sample preparation and imaging ofexosomes by transmission electron microscopy. JoVE 2018:1e5.
[37] Zhang M, Jin K, Gao L, Zhang Z, Li F, Zhou F, et al.Methods and technologies for exosome isolation and char-acterization. Small Methods 2018;2:1e10.
[38] Bruschi M, Granata S, Santucci L, Candiano G, Fabris A,Antonucci N, et al. Proteomic analysis of urinary micro-vesicles and exosomes in medullary sponge kidney diseaseand autosomal dominant polycystic kidney disease. Clin JAm Soc Nephrol : CJASN 2019;14:834e43.
[39] Soares Martins T, Catita J, Martins Rosa I, da Cruz eSilva AB, Henriques AG. Exosome isolation from distinctbiofluids using precipitation and column-based ap-proaches. PLoS One 2018;13:1e16.
[40] Rosa-Fernandes L, Rocha VB, Carregari VC, Urbani A,Palmisano G. A perspective on extracellular vesicles pro-teomics. Front Chem 2017;5:1e19.
[41] Rogers JC, Bomgarden RD. Sample preparation for massspectrometry-based proteomics; from proteomes to peptides.In: Mirzaei H, Carrasco M, editors. Modernproteomicsesample preparation, analysis and practical appli-cations. Cham: Springer International Publishing; 2016.p. 43e62.
[42] Suder P, Nov�ak P, Havlí�cek V, Bodzo�n-Kułakowska A.General strategies for proteomic sample preparation. In:Ciborowski P, Silberring J, editors. Proteomic profiling andanalytical chemistry. 2nd ed. Boston: Elsevier; 2016.p. 25e49.
[43] Li J, He X, Deng Y, Yang C. An update on isolation methodsfor proteomic studies of extracellular vesicles in biofluids.Molecules 2019;24:1e24.
[44] Wi�sniewski JR, Zougman A, Nagaraj N, Mann M. Universalsample preparation method for proteome analysis. NatMethods 2009;6:359e62.
[45] Wi�sniewski JR. Chapter two- Filter-aided sample prepara-tion: The versatile and efficient method for proteomicanalysis. In: Shukla AK, editor. Methods enzymol. Vols. 585.New york. Academic Press; 2017. p. 15e27.
[46] Tremblay T-L, Hill JJ. Adding polyvinylpyrrolidone to lowlevel protein samples significantly improves peptide re-covery in FASP digests: An inexpensive and simple modi-fication to the FASP protocol. J Proteonomics 2021;230:104000.
[47] Doellinger J, Schneider A, Hoeller M, Lasch P. Samplepreparation by easy extraction and digestion (SPEED)-Auniversal, rapid, and detergent-free protocol for proteomicsbased on acid extraction. Mol Cell Proteomics 2020;19:209e22.
[48] Guo X, Trudgian DC, Lemoff A, Yadavalli S, Mirzaei H.Confetti: A multiprotease map of the HeLa proteome forcomprehensive proteomics. Mol Cell Proteomics : MCP2014;13:1573e84.
[49] Giansanti P, Tsiatsiani L, Low TY, Heck AJR. Six alternativeproteases for mass spectrometryebased proteomics beyondtrypsin. Nat Protoc 2016;11:993e1006.
[50] Swaney DL, Wenger CD, Coon JJ. Value of using multipleproteases for large-scale mass spectrometry-based prote-omics. J Proteome Res 2010;9:1323e9.
[51] Matthiesen R, Bunkenborg J. Introduction to mass spec-trometry-based proteomics. In: Matthiesen R, editor. Massspectrometry data analysis in proteomics. Totowa: HumanaPress; 2013. p. 1e45.
[52] Gillet LC, Leitner A, Aebersold R. Mass spectrometryapplied to bottom-up proteomics: Entering the high-throughput era for hypothesis testing. Annu Rev AnalChem 2016;9:449e72.
[53] Wilson SR, Vehus T, Berg HS, Lundanes E. Nano-LC inproteomics: Recent advances and approaches. Bioanalysis2015;7:1799e815.
[54] Yates JR, Ruse CI, Nakorchevsky A. Proteomics by massspectrometry: Approaches, advances, and applications.Annu Rev Biomed Eng 2009;11:49e79.
[55] Guan S, Taylor PP, Han Z, Moran MF, Ma B. Datadependenteindependent acquisition (DDIA) proteomics.J Proteome Res 2020;19:3230e7.
[56] Hu A, Noble WS, Wolf-Yadlin A. Technical advances inproteomics: New developments in data-independentacquisition. F1000Res 2016;5:1e12.
[57] Fern�andez-Costa C, Martínez-Bartolom�e S, McClatchy DB,Saviola AJ, Yu NK, Yates JR. Impact of the identificationstrategy on the reproducibility of the DDA and DIA results.J Proteome Res 2020;19:3153e61.
[58] Suna G, Mayr M. Proteomics. In: Vasan RS, Sawyer DB,editors. Encyclopedia of cardiovascular research and med-icine. Oxford: Elsevier; 2018. p. 166e80.
[59] Papadopoulos T, Krochmal M, Cisek K, Fernandes M,Husi H, Stevens R, et al. Omics databases on kidney dis-ease: Where they can be found and how to benefit fromthem. Clin Kidney J 2016;9:343e52.
[60] Perez-Riverol Y, Csordas A, Bai J, Bernal-Llinares M,Hewapathirana S, Kundu DJ, et al. The PRIDE databaseand related tools and resources in 2019: improvingsupport for quantification data. Nucleic Acids Res 2019;47:442e50.
[61] Thul PJ, Åkesson L, Wiking M, Mahdessian D, Geladaki A,Ait Blal H, et al. A subcellular map of the human proteome.Science 2017;356:1e22.
[62] The UniProt C. UniProt: The universal protein knowl-edgebase in 2021. Nucleic Acids Res 2021;49:D480e9.
[63] Shao C. Urinary protein biomarker database 2.0: A litera-ture-curated database for protein biomarkers in urine. In:Gao Y, editor. Urine: promising biomarker source for earlydisease detection. Singapore: Springer Singapore; 2019.p. 65e72.
[64] Keerthikumar S, Chisanga D, Ariyaratne D, Al Saffar H,Anand S, Zhao K, et al. ExoCarta: A web-based compen-dium of exosomal cargo. J Mol Biol 2016;428:688e92.
[65] Huang DW, Sherman BT, Lempicki RA. Bioinformaticsenrichment tools: Paths toward the comprehensive func-tional analysis of large gene lists. Nucleic Acids Res 2009;37:1e13.
220 JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222
REVIEW
ARTIC
LE

[66] Jassal B, Matthews L, Viteri G, Gong C, Lorente P,Fabregat A, et al. The reactome pathway knowledgebase.Nucleic Acids Res 2020;48:D498e503.
[67] Erdbrügger U, Blijdorp CJ, Bijnsdorp IV, Borr�as FE,Burger D, Bussolati B, et al. Urinary extracellular vesicles: Aposition paper by the urine task force of the internationalsociety for extracellular vesicles. J Extracell Vesicles 2021;10:e12093.
[68] Blijdorp CJ, Tutakhel OAZ, Hartjes TA, van den Bosch TPP,van Heugten MH, Rigalli JP, et al. Comparing approachesto normalize, quantify, and characterize urinary extracel-lular vesicles. J Am Soc Nephrol 2021;32:1210e26.
[69] Musante L, Saraswat M, Ravid�a A, Byrne B, Holthofer H.Recovery of urinary nanovesicles from ultracentrifugationsupernatants. Nephrol Dial Transplant 2013;28:1425e33.
[70] Makris K, Spanou L. Acute kidney injury: Definition,pathophysiology and clinical phenotypes. Clin BiochemRev 2016;37:85e98.
[71] Kellum JA, Romagnani P, Ashuntantang G, Ronco C,Zarbock A, Anders HJ. Acute kidney injury. Nat Rev DisPrim 2021;7:52.
[72] Ronco C, Bellomo R, Kellum JA. Acute kidney injury.Lancet 2019;394:1949e64.
[73] Sales GTM, Foresto RD. Drug-induced nephrotoxicity. 2020Rev Assoc Med Bras 1992;66(Suppl 1):s82e90.
[74] Zhou H, Cheruvanky A, Hu X, Matsumoto T, Hiramatsu N,Cho ME, et al. Urinary exosomal transcription factors, anew class of biomarkers for renal disease. Kidney Int 2008;74:613e21.
[75] Awdishu L, Le A, Amato J, Jani V, Bal S, Mills RH, et al.Urinary exosomes identify inflammatory pathways in van-comycin associated acute kidney injury. Int J Mol Sci 2021:22.
[76] Dominguez 2nd JM, Dominguez JH, Xie D, Kelly KJ.Human extracellular microvesicles from renal tubulesreverse kidney ischemia-reperfusion injury in rats. PLoSOne 2018;13:1e16.
[77] Ammirati AL. Chronic kidney disease. Rev Assoc Med Bras(1992) 2020;66:s3e9.
[78] Glassock RJ, Warnock DG, Delanaye P. The global burdenof chronic kidney disease: Estimates, variability and pitfalls.Nat Rev Nephrol 2017;13:104e14.
[79] Madaio MP, Harrington JT. The diagnosis of glomerulardiseases: Acute glomerulonephritis and the nephrotic syn-drome. Arch Intern Med 2001;161:25e34.
[80] Khanna R. Clinical presentation & management ofglomerular diseases: Hematuria, nephritic & nephroticsyndrome. Mo Med 2011;108:33e6.
[81] Hebert LA, Parikh S, Prosek J, Nadasdy T, Rovin BH. Dif-ferential diagnosis of glomerular disease: A systematic andinclusive approach. Am J Nephrol 2013;38:253e66.
[82] Cavanaugh C, Okusa MD. The evolving role of novel bio-markers in glomerular disease: A review. Am J Kidney Dis2021;77:122e31.
[83] Moon PG, Lee JE, You S, Kim TK, Cho JH, Kim IS, et al.Proteomic analysis of urinary exosomes from patients ofearly IgA nephropathy and thin basement membrane ne-phropathy. Proteomics 2011;11:2459e75.
[84] Monhart V. Hypertension and chronic kidney diseases. CorVasa 2013;55:e397e402.
[85] Gonzalez-Calero L, Martínez PJ, Martin-Lorenzo M, Bal-dan-Martin M, Ruiz-Hurtado G, de la Cuesta F, et al.Urinary exosomes reveal protein signatures in hyperten-sive patients with albuminuria. Oncotarget 2017;8:44217e31.
[86] Selby NM, Taal MW. An updated overview of diabetic ne-phropathy: Diagnosis, prognosis, treatment goals and latestguidelines. Diabetes Obes Metabol 2020;22:3e15.
[87] Sulaiman MK. Diabetic nephropathy: Recent advances inpathophysiology and challenges in dietary management.Diabetol Metab Syndrome 2019;11:7.
[88] Zubiri I, Posada-Ayala M, Benito-Martin A, Maroto AS,Martin-Lorenzo M, Cannata-Ortiz P, et al. Kidney tissue
proteomics reveals regucalcin downregulation in responseto diabetic nephropathy with reflection in urinary exo-somes. Transl Res 2015;166:474e84.
[89] Picken MM. The pathology of amyloidosis in classification:A review. Acta Haematol 2020;143:322e34.
[90] Ramirez-Alvarado M, Ward CJ, Huang BQ, Gong X,Hogan MC, Madden BJ, et al. Differences in immunoglob-ulin light chain species found in urinary exosomes in lightchain amyloidosis (Al). PLoS One 2012;7:1e11.
[91] Ramirez-Alvarado M, Barnidge DR, Murray DL,Dispenzieri A,Marin-ArganyM, Dick CJ, et al. Assessment ofrenal response with urinary exosomes in patients with ALamyloidosis: A proof of concept. Am J Hematol 2017;92:536e41.
[92] Agarwal R. Defining end-stage renal disease in clinical tri-als: A framework for adjudication. Nephrol Dial Transplant2016;31:864e7.
[93] Pearson LJ, Klaharn IY, Thongsawang B, Manuprasert W,Saejew T, Somparn P, et al. Multiple extracellular vesicletypes in peritoneal dialysis effluent are prominent andcontain known biomarkers. PLoS One 2017;12:1e18.
[94] Carreras-Planella L, Soler-Majoral J, Rubio-Esteve C,Mor�on-Font M, Franquesa M, Bonal J, et al. Proteomicprofiling of peritoneal dialysis effluent-derived extracellularvesicles: A longitudinal study. J Nephrol 2019;32:1021e31.
[95] Pisitkun T, Gandolfo MT, Das S, Knepper MA, Bagnasco SM.Application of systems biology principles to proteinbiomarker discovery: Urinary exosomal proteome in renaltransplantation. Proteonomics Clin Appl 2012;6:268e78.
[96] Dieud�e M, Bell C, Turgeon J, Beillevaire D, Pomerleau L,Yang B, et al. The 20S proteasome core, active withinapoptotic exosome-like vesicles, induces autoantibodyproduction and accelerates rejection. Sci Transl Med 2015;7:318ra200.
[97] Lim JH, Lee CH, Kim KY, Jung HY, Choi JY, Cho JH, et al.Novel urinary exosomal biomarkers of acute T cell-medi-ated rejection in kidney transplant recipients: A cross-sectional study. PLoS One 2018;13:1e17.
[98] Flechner SM, Kobashigawa J, Klintmalm G. Calcineurininhibitor-sparing regimens in solid organ transplantation:Focus on improving renal function and nephrotoxicity. ClinTransplant 2008;22:1e15.
[99] Carreras-Planella L, Juega J, Taco O, Ca~nas L, Franquesa M,Lauzurica R, et al. Proteomic characterization of urinaryextracellular vesicles from kidney-transplanted patientstreatedwith calcineurin inhibitors. Int JMol Sci 2020;21:1e14.
[100] Choi DS. Urinary extracellular vesicles for biomarkersource to monitor polycystic kidney disease. ProteonomicsClin Appl 2015;9:447e8.
[101] Staubach S, Wenzel A, Beck BB, Rinschen MM, Müller S,Hanisch FG. Autosomal tubulointerstitial kidney disease-MUC1 type: Differential proteomics suggests that mutatedMUC1 (insC) affects vesicular transport in renal epithelialcells. Proteomics 2018;18:e1700456.
[102] Hogan MC, Manganelli L, Woollard JR, Masyuk AI,Masyuk TV, Tammachote R, et al. Characterization of PKDprotein-positive exosome-like vesicles. J Am Soc Nephrol2009;20:278e88.
[103] Lea WA, McGreal K, Sharma M, Parnell SC, Zelenchuk L,Charlesworth MC, et al. Analysis of the polycystin complex(PCC) in human urinary exosome-like vesicles (ELVs). SciRep 2020;10:1500.
[104] Hogan MC, Bakeberg JL, Gainullin VG, Irazabal MV,Harmon AJ, Lieske JC, et al. Identification of biomarkers forPKD1 using urinary exosomes. J Am Soc Nephrol 2015;26:1661e70.
[105] Salih M, Demmers JA, Bezstarosti K, Leonhard WN,Losekoot M, van Kooten C, et al. Proteomics of urinaryvesicles links plakins and complement to polycystic kidneydisease. J Am Soc Nephrol 2016;27:3079e92.
[106] Bruschi M, Granata S, Candiano G, Fabris A, Petretto A,Ghiggeri GM, et al. Proteomic analysis of urinary extra-cellular vesicles reveals a role for the complement system in
JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222 221
REVIEW
ARTIC
LE

medullary sponge kidney disease. Int J Mol Sci 2019;20:1e15.
[107] Jackson L, Woodward M, Coward RJ. The molecularbiology of pelvi-ureteric junction obstruction. PediatrNephrol 2018;33:553e71.
[108] Liu R, Zhang W, Luo M, Qin X, Yang F, Wei Q. iTRAQ-based proteomics and in vitro experiments reveals essentialroles of ACE and AP-N in the renin-angiotensin system-mediated congenital ureteropelvic junction obstruction. ExpCell Res 2020;393:112086.
[109] Staubach S, Pekmez M, Hanisch FG. Differential prote-omics of urinary exovesicles from classical galactosemicpatients reveals subclinical kidney insufficiency. J ProteomeRes 2016;15:1754e61.
[110] Claes DJ, Jackson E. Cystinuria: Mechanisms and man-agement. Pediatr Nephrol 2012;27:2031e8.
[111] Bourderioux M, Nguyen-Khoa T, Chhuon C, Jeanson L,Tondelier D, Walczak M, et al. A new workflow for prote-omic analysis of urinary exosomes and assessment incystinuria patients. J Proteome Res 2015;14:567e77.
[112] Thongboonkerd V. Roles for exosome in various kidneydiseases and disorders. Front Pharmacol 2020;10:1e14.
[113] Raimondo F, Morosi L, Corbetta S, Chinello C, Brambilla P,Della Mina P, et al. Differential protein profiling of renal
cell carcinoma urinary exosomes. Mol Biosyst 2013;9:1220e33.
[114] Wu Q, Poulsen SB, Murali SK, Grimm PR, Su X-T,Delpire E, et al. Large-scale proteomic assessment of uri-nary extracellular vesicles highlights their reliability inreflecting protein changes in the kidney. J Am Soc Nephrol2021;32:2195e209.
[115] Gonzales PA, Pisitkun T, Hoffert JD, Tchapyjnikov D,Star RA, Kleta R, et al. Large-scale proteomics and phos-phoproteomics of urinary exosomes. J Am Soc Nephrol :JASN (J Am Soc Nephrol) 2009;20:363e79.
[116] Raimondo F, Chinello C, Porcaro L, Magni F, Pitto M.Urinary extracellular vesicles and salt-losing tubulopathies:A proteomic approach. Proteomes 2020;8.
[117] Sung CC, Chen MH, Lin YC, Lin YC, Lin YJ, Yang SS, et al.Urinary extracellular vesicles for renal tubular transportersexpression in patients with gitelman syndrome. Front Med2021;8:679171.
[118] Rood IM, Merchant ML, Wilkey DW, Zhang T,Zabrouskov V, van der Vlag J, et al. Increased expression oflysosome membrane protein 2 in glomeruli of patients withidiopathic membranous nephropathy. Proteomics 2015;15:3722e30.
222 JOURNAL OF FOOD AND DRUG ANALYSIS 2022;30:202e222
REVIEW
ARTIC
LE
Copyright © 2022 FDOKUMEN