
Pointing as an Instrumental Gesture: Gaze Representation Through Indication
Upload
independentCategory
view
3download
0
Nitric Oxide 20 (2009) 207–216
Contents lists available at ScienceDirect
Nitric Oxide
journal homepage: www.elsevier .com/ locate/yniox
Fluorescent indication that nitric oxide formation in NTS neurons is modulatedby glutamate and GABA
Gisela P. Pajolla a, Daniela Accorsi-Mendonça a, Gerson J. Rodrigues b, Lusiane M. Bendhack b,Benedito H. Machado a,*, Claure N. Lunardi c,*
a Department of Physiology, School of Medicine of Ribeirão Preto, University of São Paulo, Av. Bandeirantes, 3900, 14049-900 Ribeirão Preto, SP, Brazilb Department of Physics and Chemistry, School of Pharmaceutical Sciences of Ribeirão Preto, University of São Paulo, 14049-900, Ribeirão Preto, SP, Brazilc College of Ceilândia, University of Brasília, 70910-900, Brasília, DF, Brazil
a r t i c l e i n f o a b s t r a c t
Article history:Received 10 November 2008Revised 18 December 2008Available online 25 January 2009
Keywords:Nitric oxideNucleus tractus solitariiDAF-2 DAGlutamateGABANeurons
1089-8603/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.niox.2009.01.001
* Corresponding authors. Fax: +55 16 3633 0017 (BE-mail addresses: [email protected] (B.H. Ma
Lunardi).
Nitric oxide (NO) in NTS plays an important role in regulating autonomic function to the cardiovascularsystem. Using the fluorescent dye DAF-2 DA, we evaluated the NO concentration in NTS. Brainstem slicesof rats were loaded with DAF-2 DA, washed, fixed in paraformaldehyde and examined under fluorescentlight. In different experimental groups, NTS slices were pre-incubated with 1 mM L-NAME (a non-selec-tive NOS inhibitor), 1 mM D-NAME (an inactive enantiomere of L-NAME), 1 mM kynurenic acid (a non-selective ionotropic receptors antagonist) or 20 lM bicuculline (a selective GABAA receptors antagonist)before and during DAF-2 DA loading. Images were acquired using a confocal microscope and the intensityof fluorescence was quantified in three antero-posterior NTS regions. In addition, slices previously loadedwith DAF-2 DA were incubated with NeuN or GFAP antibody. A semi-quantitative analysis of the fluores-cence intensity showed that the basal NO concentration was similar in all antero-posterior aspects of theNTS (rostral intermediate, 15.5 ± 0.8 AU; caudal intermediate, 13.2 ± 1.4 AU; caudal commissural,13.8 ± 1.4 AU, n = 10). In addition, the inhibition of NOS and the antagonism of glutamatergic receptorsdecreased the NO fluorescence in the NTS. On the other hand, D-NAME did not affect the NO fluorescenceand the antagonism of GABAA receptors increased the NO fluorescence in the NTS. It is important to notethat the fluorescence for NO was detected mainly in neurons. These data show that the fluorescenceobserved after NTS loading with DAF-2 DA is a result of NO present in the NTS and support the conceptthat NTS neurons have basal NO production which is modulated by L-glutamate and GABA.
� 2009 Elsevier Inc. All rights reserved.
Nitric oxide (NO) synthesized from L-arginine by NO synthasehas a large variety of physiological actions in the periphery as wellin the central nervous system [1,2]. Studies performed in thenucleus tractus solitarii (NTS), an important site in brainstem forthe integration of afferent information, show the NO involvementin the cardiovascular and respiratory signal transduction [3–6].
In spite of a well documented presence of neuronal (nNOS) andendothelial (eNOS) isoforms of enzyme that synthesizes NO in theNTS [7] and also of several pieces of evidence pointing to theinvolvement of NO in NTS in regulation of the cardiovascular sys-tem, using different experimental approaches, none of these previ-ous studies measured NO basal levels in NTS [8–15]. The extremelylabile nature of the free radical NO (t1/2 = 3–6 s) makes it very dif-ficult to be determined directly. Therefore, effective measurementof NO presence in NTS remains as a challenge for a better under-standing of its physiological and pathophysiological significance
ll rights reserved.
.H. Machado).chado), [email protected] (C.N.
on this important integrative site in brainstem. The direct mea-surement of NO in different tissues has been achieved by severalinvestigators using chemiluminescence detection [16], electronspin resonance [17], or electrochemical detection with a porphyri-nic sensor [18]. Nevertheless, these methods have drawbacks inoperating procedure, detection sensitivity, or limitations in prepar-ing the NO-sensing electrode. Several diaminofluorescein analogs(DAFs) were developed by Kojima et al. [19] for determinationand bioimaging of NO in a variety of cells and this method seemsto be more reliable to identify the NO presence in different tissuesthan any other approach [19–21]. Most importantly is the fact thatDAF-2 DA was previously used with success in the central nervoussystem to evaluate the NO production in hippocampus and cortexof rats [20] as well in the supraoptic nucleus [22].
Processing of cardiovascular reflexes afferents in NTS involvesseveral neurotransmitters and modulators [23–26]. However,L-glutamate and GABA are the main excitatory and inhibitoryneurotransmitters in NTS, respectively, [27–30] and they are in-volved with the processing of the cardiovascular and respiratoryafferent informations [31–33]. There is evidence that responses

208 G.P. Pajolla et al. / Nitric Oxide 20 (2009) 207–216
to activation of glutamate and GABA receptors involve NO-depen-dent mechanisms [34–38] and also that activation of ionotropicglutamate receptors results in NO synthesis [34,35]. Thus, interac-tions of L-glutamate and GABA on NO production in NTS appear tobe complex and this interaction may modulate basal level of NOproduction with important implications for autonomic regulationof cardiovascular function. In the present study, we used a fluores-cence method with the dye DAF-2 DA for detection of NO presencein the NTS and also to evaluate the modulation of NO productionby L-glutamate and GABA in NTS neurons using confocal micro-scope. Immunoreactivity to NeuN or GFAP was also used to identifywhether NO production was present in neurons and/or in glialcells.
Fig. 1. Line drawing of transverse section of the brainstem at the rostral and caudalintermediate NTS and caudal commissural NTS (adapted from Paxinos and Watson,[39]). The digital images from the gray squares areas were used to quantify thefluorescence intensity in a confocal microscope.
Experimental procedures
Animals
The experiments were performed on male juvenile Wistar rats,weighing 50–60 g, from the Animal Care of the University of SãoPaulo at Ribeirão Preto, Brazil. All experimental protocols were ap-proved by the Ethical Committee on Animal Experimentation ofthe School of Medicine of Ribeirão Preto, University of São Paulo(protocol # 030/2004).
Three or four brainstem slices containing the NTS were obtainedfor each animal (see Section ‘NTS cell loading with the NO fluores-cent indicator DAF-2 DA’) and were loaded with DAF-2 DA (5 lM).The concentration of DAF-2 DA used to evaluate the nitric oxideproduction in the NTS was based in previous works by Zhangand Stern [22], Kasim et al. [39] and Kojima et al. [19] and on theseprevious studies the DAF-2 DA concentration used was 10 lM.Studies by Rodriguez et al. [40] documented that with this concen-tration (10 lM) commonly used, intracellular DAF-2 accumulatesto concentrations near the mM range and such elevated concentra-tions are associated with high levels of background autofluores-cence from the probe. In order to minimize this problem, in thepresent study we used 5 lM DAF-2 DA. Moreover, this concentra-tion seems to be appropriated to evaluate a decrease/increase ofNO production using DAF-2 DA in our system, since in our experi-ments NO production varied in accordance with the experimentalprotocol.
In different experimental groups, the slices of the NTS wereincubated with L-NAME, D-NAME, kynurenic acid or bicucullinebefore and during DAF-2 DA loading. Another group of NTS sliceswas loaded with DAF-2 DA and submitted to the immunofluores-cence for neuronal or glial cell marker.
The analyze the NO production in the NTS, the NO-induced fluo-rescence (NOIF) was acquired in three sub-regions of the NTS foreach brainstem slice, i.e., above the central canal and in the lateralaspects of the NTS near by the tractus solitarii (rostral intermediatecommissural NTS, caudal intermediate commissural NTS andcaudal commissural NTS), according to the atlas of Paxinos andWatson [41] (see Fig. 1)
Relation between 4,5-diaminofluorescein diacetate (DAF-2 DA) andNO availability
DAF-2 DA is a NO-sensitive indicator that permeates cellsmembrane and is hydrolyzed by cytosolic esterases formingDAF-2, a non-permeable and relatively non-fluorescent com-pound which is nitrosylated by products of oxidation of nitricoxide to form fluorescent heterocycles, a compound that becometrapped in the cytoplasm as DAF-2T. Thus, DAF-2T allows thevisualization and semi-quantitative analysis of basal NO availabil-ity at the cell level.
A cell-free system, as previously documented by Stern andZhang [22], was used to verify if the DAF-2 DA used was able tobe converted in DAF-2T in the presence of nitric oxide and emit afluorescent signal to our image acquisition system, and also to doc-ument that the physical calibration of the system (laser, pinhole,filters, etc.) were sensitive enough to detect changes in the NOIF.It is important to note that in this procedure DAF-2T intensity sig-nal from a solution was not compared to that confined within aneuron. DAF-2 DA (10 ll dissolved in DMSO) was hydrolyzed toform DAF-2 with 10 ll methanol (1:1) and treated with KOH(2 M–25 ll) for 1 h at room temperature followed by pH adjust-ment to 7.4 with HCl (2 M). A series of solutions containing a fixedconcentration of DAF-2 (5 lM) and increasing concentrations ofthe NO donor sodium nitroprusside (SNP; 0.1–10 mM) was usedto measure the emitted fluorescence of DAF-2T. The emitted fluo-
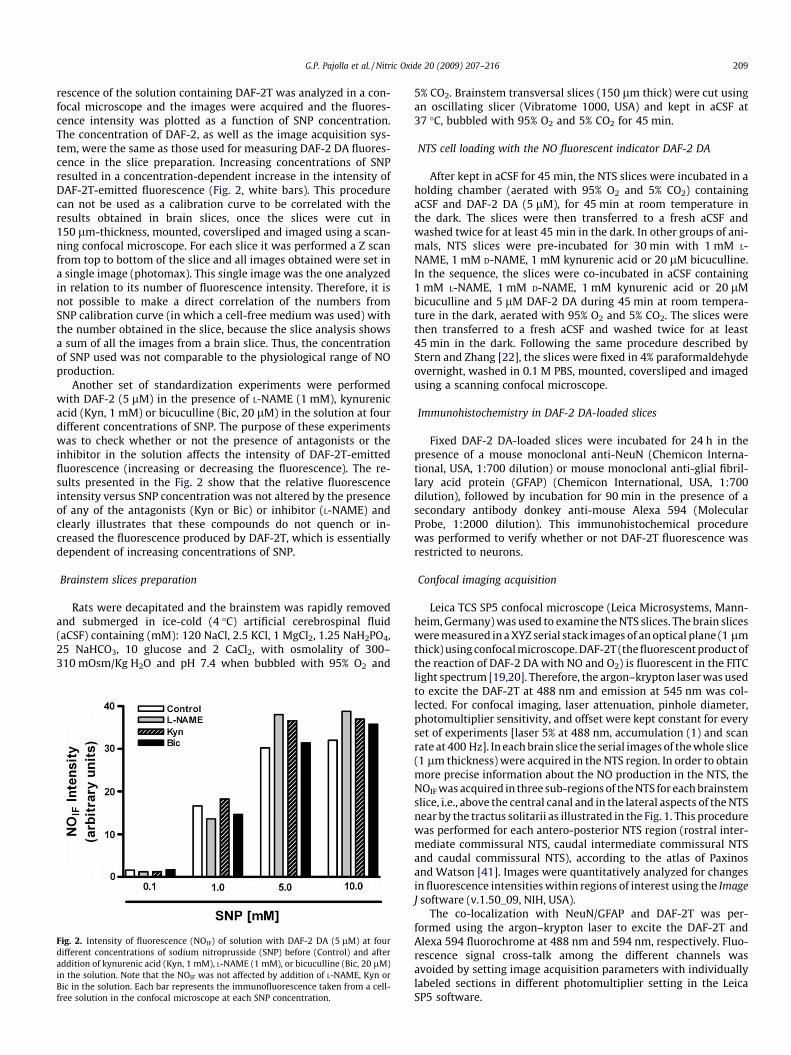
G.P. Pajolla et al. / Nitric Oxide 20 (2009) 207–216 209
rescence of the solution containing DAF-2T was analyzed in a con-focal microscope and the images were acquired and the fluores-cence intensity was plotted as a function of SNP concentration.The concentration of DAF-2, as well as the image acquisition sys-tem, were the same as those used for measuring DAF-2 DA fluores-cence in the slice preparation. Increasing concentrations of SNPresulted in a concentration-dependent increase in the intensity ofDAF-2T-emitted fluorescence (Fig. 2, white bars). This procedurecan not be used as a calibration curve to be correlated with theresults obtained in brain slices, once the slices were cut in150 lm-thickness, mounted, coversliped and imaged using a scan-ning confocal microscope. For each slice it was performed a Z scanfrom top to bottom of the slice and all images obtained were set ina single image (photomax). This single image was the one analyzedin relation to its number of fluorescence intensity. Therefore, it isnot possible to make a direct correlation of the numbers fromSNP calibration curve (in which a cell-free medium was used) withthe number obtained in the slice, because the slice analysis showsa sum of all the images from a brain slice. Thus, the concentrationof SNP used was not comparable to the physiological range of NOproduction.
Another set of standardization experiments were performedwith DAF-2 (5 lM) in the presence of L-NAME (1 mM), kynurenicacid (Kyn, 1 mM) or bicuculline (Bic, 20 lM) in the solution at fourdifferent concentrations of SNP. The purpose of these experimentswas to check whether or not the presence of antagonists or theinhibitor in the solution affects the intensity of DAF-2T-emittedfluorescence (increasing or decreasing the fluorescence). The re-sults presented in the Fig. 2 show that the relative fluorescenceintensity versus SNP concentration was not altered by the presenceof any of the antagonists (Kyn or Bic) or inhibitor (L-NAME) andclearly illustrates that these compounds do not quench or in-creased the fluorescence produced by DAF-2T, which is essentiallydependent of increasing concentrations of SNP.
Brainstem slices preparation
Rats were decapitated and the brainstem was rapidly removedand submerged in ice-cold (4 �C) artificial cerebrospinal fluid(aCSF) containing (mM): 120 NaCl, 2.5 KCl, 1 MgCl2, 1.25 NaH2PO4,25 NaHCO3, 10 glucose and 2 CaCl2, with osmolality of 300–310 mOsm/Kg�H2O and pH 7.4 when bubbled with 95% O2 and
Fig. 2. Intensity of fluorescence (NOIF) of solution with DAF-2 DA (5 lM) at fourdifferent concentrations of sodium nitroprusside (SNP) before (Control) and afteraddition of kynurenic acid (Kyn, 1 mM), L-NAME (1 mM), or bicuculline (Bic, 20 lM)in the solution. Note that the NOIF was not affected by addition of L-NAME, Kyn orBic in the solution. Each bar represents the immunofluorescence taken from a cell-free solution in the confocal microscope at each SNP concentration.
5% CO2. Brainstem transversal slices (150 lm thick) were cut usingan oscillating slicer (Vibratome 1000, USA) and kept in aCSF at37 �C, bubbled with 95% O2 and 5% CO2 for 45 min.
NTS cell loading with the NO fluorescent indicator DAF-2 DA
After kept in aCSF for 45 min, the NTS slices were incubated in aholding chamber (aerated with 95% O2 and 5% CO2) containingaCSF and DAF-2 DA (5 lM), for 45 min at room temperature inthe dark. The slices were then transferred to a fresh aCSF andwashed twice for at least 45 min in the dark. In other groups of ani-mals, NTS slices were pre-incubated for 30 min with 1 mM L-NAME, 1 mM D-NAME, 1 mM kynurenic acid or 20 lM bicuculline.In the sequence, the slices were co-incubated in aCSF containing1 mM L-NAME, 1 mM D-NAME, 1 mM kynurenic acid or 20 lMbicuculline and 5 lM DAF-2 DA during 45 min at room tempera-ture in the dark, aerated with 95% O2 and 5% CO2. The slices werethen transferred to a fresh aCSF and washed twice for at least45 min in the dark. Following the same procedure described byStern and Zhang [22], the slices were fixed in 4% paraformaldehydeovernight, washed in 0.1 M PBS, mounted, coversliped and imagedusing a scanning confocal microscope.
Immunohistochemistry in DAF-2 DA-loaded slices
Fixed DAF-2 DA-loaded slices were incubated for 24 h in thepresence of a mouse monoclonal anti-NeuN (Chemicon Interna-tional, USA, 1:700 dilution) or mouse monoclonal anti-glial fibril-lary acid protein (GFAP) (Chemicon International, USA, 1:700dilution), followed by incubation for 90 min in the presence of asecondary antibody donkey anti-mouse Alexa 594 (MolecularProbe, 1:2000 dilution). This immunohistochemical procedurewas performed to verify whether or not DAF-2T fluorescence wasrestricted to neurons.
Confocal imaging acquisition
Leica TCS SP5 confocal microscope (Leica Microsystems, Mann-heim, Germany) was used to examine the NTS slices. The brain sliceswere measured in a XYZ serial stack images of an optical plane (1 lmthick) using confocal microscope. DAF-2T (the fluorescent product ofthe reaction of DAF-2 DA with NO and O2) is fluorescent in the FITClight spectrum [19,20]. Therefore, the argon–krypton laser was usedto excite the DAF-2T at 488 nm and emission at 545 nm was col-lected. For confocal imaging, laser attenuation, pinhole diameter,photomultiplier sensitivity, and offset were kept constant for everyset of experiments [laser 5% at 488 nm, accumulation (1) and scanrate at 400 Hz]. In each brain slice the serial images of the whole slice(1 lm thickness) were acquired in the NTS region. In order to obtainmore precise information about the NO production in the NTS, theNOIF was acquired in three sub-regions of the NTS for each brainstemslice, i.e., above the central canal and in the lateral aspects of the NTSnear by the tractus solitarii as illustrated in the Fig. 1. This procedurewas performed for each antero-posterior NTS region (rostral inter-mediate commissural NTS, caudal intermediate commissural NTSand caudal commissural NTS), according to the atlas of Paxinosand Watson [41]. Images were quantitatively analyzed for changesin fluorescence intensities within regions of interest using the ImageJ software (v.1.50_09, NIH, USA).
The co-localization with NeuN/GFAP and DAF-2T was per-formed using the argon–krypton laser to excite the DAF-2T andAlexa 594 fluorochrome at 488 nm and 594 nm, respectively. Fluo-rescence signal cross-talk among the different channels wasavoided by setting image acquisition parameters with individuallylabeled sections in different photomultiplier setting in the LeicaSP5 software.

210 G.P. Pajolla et al. / Nitric Oxide 20 (2009) 207–216
Data analysis
NO measurement of brain slices with DAF-2 DASemi-quantitative analysis of NOIF intensity was performed
using imaging analysis software (Image J, v.1.50_09, NIH, USA).The background fluorescence for each set of experiment (i.e., brainslice) was subtracted and a photomax (z-project from the serialstack) was constructed. DAF-2 DA was used in the present studyto measure the NO reaction inside the cell and the background sub-traction was performed to avoid a false result in the quantification.The background subtracted from the NTS images was the low fluo-rescence signal not present in cells. This background seems to begenerated by three majors sources: from the fixative paraformal-dehyde that crosslink to bioamines such as dopamine increasingthe external fluorescence from the neurons [42,43], from autofluo-rescence of the tissue or from the probe itself [40].
Briefly, individual NTS cells were traced using the pluginparticle analysis (nucleus counter), and the mean gray value con-tained within these regions was measured as an index of NOIF,and expressed as fluorescence arbitrary units (AU), ranging from0 (absolute black) to 255 (absolute green). The mean value of thethree regions of the NTS for each set of experiment (i.e., each brain-stem slice) was quantified and analyzed in relation to the antero-posterior levels (rostral and caudal intermediate NTS and caudalcommissural NTS). The data are expressed as mean ± SEM of arbi-trary unit (AU) of NOIF intensity and statistical significance(P < 0.05) was determined by the ANOVA one way or Student t testusing the GraphPad Prism 4 software (GraphPad Prism, USA).
Co-localization of NeuN/GFAP in neurons with DAF-2 DATo determine co-localization of two fluorescent markers (DAF-2T
and/or NeuN, GFAP), confocal images obtained from the differentfluorophores were merged. A positive co-localization was consid-ered by the appearance of yellow (red + green) profile on the samespot in the merged image.
Drugs
The following drugs were used in these experiments: DAF-2 DA,L-NAME, D-NAME, kynurenic acid, bicuculline and DMSO from Sig-ma (St. Louis, MO, USA). The stock solution of bicuculline was dis-solved in DMSO and the final concentration of DMSO in the bathwas smaller than 0.1%. All other drugs were dissolved in water.
Results
Blockade of NO production by non-selective inhibitor of NOS L-NAMEwithin NTS
To verify whether the fluorescence is a result of NO presence, NTSslices were incubated with 1 mM of L-NAME or 1 mM of D-NAME (theinactive enantiomere of L-NAME) before and during DAF-2 DA load-ing. The NOIF was quantified in nine specific sites of NTS, i.e., threeantero-posterior sections in which one medial and two lateral sub-regions were analyzed. The data from these experiments are shownin Figs. 3 and 4. The use of L-NAME produces a significant reduction inNOIF in NTS neurons (88 ± 7 AU vs 32 ± 3 AU, n = 6, P < 0.05, Fig. 3).The use of D-NAME produced no change in the NOIF in NTS neurons(80 ± 5 AU vs 89 ± 6 AU, n = 4 Fig. 4). Therefore, these results stronglysupport the concept that the NOIF detect in all sets of experiments aredue to NO production within the NTS.
Basal nitric oxide (NO) production within NTS
To determine basal production of NO in NTS neurons and alsoto evaluate its production in different sub-regions of NTS (Fig. 5),
brainstem slices containing NTS were loaded with DAF-2 DA,fixed, and examined under fluorescent light (see Section ‘Experi-mental procedures’). Examples of fluorescence confocal photomi-crographs obtained from DAF-2 DA-loaded slices containingrostral and caudal intermediate commissural NTS and the caudalcommissural NTS are shown in Fig. 3. High levels of basal NOIF
were observed throughout the NTS and no significant differenceswere observed in the basal NO production in the three antero-posterior regions of NTS evaluated: (a) rostral intermediate com-missural NTS at the level of rostral aspect of area postrema(15.5 ± 0.8 AU); (b) caudal intermediate commissural NTS at levelof calamus scriptorius (13.2 ± 1.4 AU); and (c) caudal commissuralNTS [13.8 ± 0.8 AU (n = 8–10)]. Considering that no differenceswere observed in the intensity of fluorescence among these threeantero-posterior areas of the NTS, in the subsequent experimentswe used the overall average as a control of NOIF in the NTS(14.22 ± 0.57 AU).
Ionotropic glutamate receptor antagonism decreased basalproduction of NO in NTS
To determine whether baseline production of NO in NTS neu-rons is modulated by activation of glutamate receptors, NTS sliceswere incubated with 1 mM of kynurenic acid, a non-selectiveantagonist of ionotropic glutamate receptors, before and duringDAF-2 DA loading. The NOIF was quantified in nine specific sitesof NTS, i.e., three antero-posterior sections in which one medialand two lateral sub-regions were analyzed. Antagonism of gluta-mate receptors with kynurenic acid significantly decrease NOIF inNTS neurons (13.5 ± 0.8 AU vs 8.9 ± 0.6 AU, n = 8, P < 0.05, Fig. 6).
GABAA receptor antagonism increased basal production of NO in NTS
To evaluate whether basel production of NO in NTS neurons ismodulated by GABAA receptors activation, NTS slices were incu-bated with 20 lM bicuculline, a GABAA receptors antagonist, be-fore and during DAF-2 DA loading. The NOIF was quantified innine specific sites of NTS, i.e., three antero-posterior sections inwhich one medial and two lateral sub-regions were analyzed.Blockade of GABAA receptors with bicuculline significantly in-creased NOIF in the NTS (control = 15.8 ± 0.5 AU; Bicucul-line = 18.0 ± 0.3 AU, n = 4, P < 0.05, Fig. 7).
NOIF colocalizes with immunostained for neurons but not for glial cellsin NTS
To determine whether NOIF was restricted to neurons DAF-2DA-loaded slices were subsequently immunostained for NeuN(neurons) or GFAP (glial cells). Typical photomicrographs showingNOIF, immunostain for NeuN and superimposed images of NeuNand NOIF staining are shown on the panels A–C of Fig. 8, respec-tively. Typical photomicrographs showing NOIF, immunostain forGFAP and superimposed images of GFAP and NOIF staining areshown on the panels D–F of Fig. 6, respectively. Panel C showsthe double staining for NOIF and NeuN and panel F shows that NOIF
staining is not co-localized with GFAP.
Discussion
In the present study we used DAF-2 DA fluorescence method todetermine the NO presence and its modulation by L-glutamate andGABA in the NTS neurons. This study provides the first evidence ofNO presence in the neurons of the rostral and caudal intermediatecommissural NTS and caudal commissural NTS and also that NOproduction in the NTS neurons is reduced by the antagonism of

Fig. 3. L-NAME decreases basal NOIF in NTS neurons. Confocal photomicrographs showing two representative images of the basal NOIF production in the NTS in control aCSF(control, A) and in aCSF containing L-NAME (1 mM, B). Scale bar = 50 lm. (C) Graphic showing that the L-NAME significantly decreased NOIF in NTS neurons. The values of eachbar correspond to the average of the data from three antero-posterior regions of the NTS for each brainstem analyzed (n = 6). (�) different in relation to the control group,P < 0.05.
G.P. Pajolla et al. / Nitric Oxide 20 (2009) 207–216 211
glutamate ionotropic receptors and increased by the antagonism ofGABAA receptors, indicating a modulatory role for L-glutamate andGABA on the basal production of NO.
NO is a gaseous free radical and it has been shown to be a phys-iological neurotransmitter or neuromodulator [1]. Due to its insta-bility and low concentration in biological systems, it is not simpleto perform NO detection with a reliable sensitive method. A familyof fluorescence dyes (diaminofluorescein analogs—DAFs), devel-oped by Kojima et al. [19] for the detection of NO in a variety ofcells is sensitive enough to detect nanomolar concentrations. Theseindicators contain a benzoic group attached to a fluorophore thatpossesses a charge transfer state located energetically betweenthe ground and excited states of the fluorophore moiety, whichserves as an effective deactivation pathway for the highly fluores-cent excited state [19,44]. When nitrosated by reactive nitrogenoxides such as N2O3 at one of its vicinal amino moieties, an internaldiazotation reaction takes place that leads to formation of the cor-responding triazole. The latter may be formed by autooxidation ofNO in the air according to the reaction 4NO2 + O2 ? 2N2O3, and inaqueous solution will produce nitrite (N2O3 + H2O ? NO2
� + 2H+)[45]. Therefore, the fluorescence intensity is dependent not onlyof NO production, but also on the rate of NO oxidation, which re-quires oxygen and it is dependent of NO concentration. This factimplies that DAF cannot be used under anoxia. This chemical mod-ification of the dye by a secondary product of NO is associated witha lowering of the energetic charge transfer state below the groundstate of the fluorophore, thus removing the quenching effect of thebenzoic structure and allowing the fluorophore efficiently to emitthe fluorescence. DAFs do not interact with NO2�, NO3� or otherreactive oxygen species, including OONOO�, O2� and H2O2
[19,44]. The high yield of fluorescence of the triazole forms of DAFs
makes these indicators suitable for the detection of NO productionin biological systems, a property that has been exploited in severalstudies using fluorometry, flow cytometry, high-pressure liquidchromatography (HPLC), and fluorescence microscopy as describedby Rodriguez et al. [40]. Among DAF analogs, DAF-2 or its ester,DAF-2 diacetate (DAF-2 DA), has been widely used for NO assayand NO imaging in vascular endothelial cells and brain neurons[19,45]. DAF-2 DA readily enters cells where it is converted to4,5-diaminofluorescein (DAF-2), which in turn reacts with NOautooxidation product to yield triazolofluorescein (DAF-2T), a fluo-rescent compound not permeable throughout the cell membrane[19].
The NTS contains several putative neurotransmitters that havebeen implicated in processing of cardiovascular reflex signals[23,25,26,33,46,47]. NO is a modulatory molecule which acts as aneurotransmitter in the central nervous system [48] and NO-pro-ducing neurons are found in brain regions known to influenceautonomic activity, including the NTS [49–53]. Furthermore, ithas been shown that NO in the NTS modulates the sympathetic re-nal nerve activity [54,55] as well as arterial blood pressure in rats[56].
The physiological role of NO within the NTS on the central neu-ral control of the circulation has been suggested since Lewis et al.[4] demonstrated that NO donors microinjected into the NTS pro-duced baroreflex-like responses and Tagawa et al. [57] showed thatL-arginine, a NO precursor, increased the firing of NTS neurons.Moreover, L-arginine microinjected into the NTS also decreased,in a dose-dependent manner, arterial blood pressure and heart ratein the anaesthetized rat [55]. Studies by Paton et al. [11] docu-mented that the depression of the gain of the baroreflex inducedby angiotensin II was mediated by endothelial NOS in the NTS.

Fig. 4. D-NAME does not change basal NOIF in NTS neurons. Confocal photomicrographs showing two representative images of the basal NOIF production in the NTS in controlaCSF (control, A) and in aCSF containing D-NAME (1 mM, B). Scale bar = 50 lm. (C) Graphic showing that the D-NAME caused no significant changes in NOIF in NTS neurons.The values of each bar correspond to the average of the data from three antero-posterior regions of the NTS for each brainstem analyzed (n = 4).
Fig. 5. Basal NO presence in NTS cells loaded with DAF-2 DA. Confocal photomicrographs showing three representative images of the basal NOIF in the intermediate NTS [(A)rostral aspect at the area postrema (AP) level and (B) caudal aspect at the calamus scriptorium (CS) level] and (C) in the caudal commissural NTS (CAU). Scale bar = 50 lm. TheNOIF signal was verified in the three antero-posterior NTS sub-regions evaluated. (D) Graphic showing that the basal NOIF in all NTS sub-regions analyzed were notstatistically different. The values of each bar are the average of the data from the three antero-posterior regions of the NTS for each brainstem analyzed (n = 10) or the overallaverage in NTS.
212 G.P. Pajolla et al. / Nitric Oxide 20 (2009) 207–216
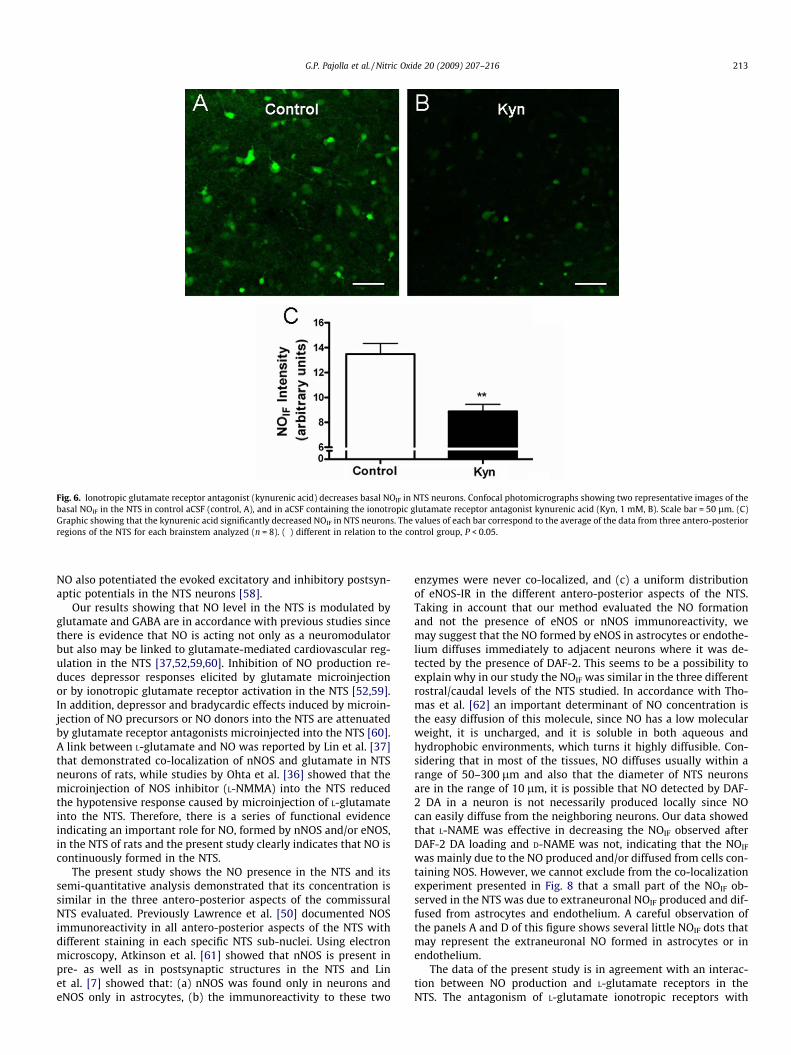
Fig. 6. Ionotropic glutamate receptor antagonist (kynurenic acid) decreases basal NOIF in NTS neurons. Confocal photomicrographs showing two representative images of thebasal NOIF in the NTS in control aCSF (control, A), and in aCSF containing the ionotropic glutamate receptor antagonist kynurenic acid (Kyn, 1 mM, B). Scale bar = 50 lm. (C)Graphic showing that the kynurenic acid significantly decreased NOIF in NTS neurons. The values of each bar correspond to the average of the data from three antero-posteriorregions of the NTS for each brainstem analyzed (n = 8). (�) different in relation to the control group, P < 0.05.
G.P. Pajolla et al. / Nitric Oxide 20 (2009) 207–216 213
NO also potentiated the evoked excitatory and inhibitory postsyn-aptic potentials in the NTS neurons [58].
Our results showing that NO level in the NTS is modulated byglutamate and GABA are in accordance with previous studies sincethere is evidence that NO is acting not only as a neuromodulatorbut also may be linked to glutamate-mediated cardiovascular reg-ulation in the NTS [37,52,59,60]. Inhibition of NO production re-duces depressor responses elicited by glutamate microinjectionor by ionotropic glutamate receptor activation in the NTS [52,59].In addition, depressor and bradycardic effects induced by microin-jection of NO precursors or NO donors into the NTS are attenuatedby glutamate receptor antagonists microinjected into the NTS [60].A link between L-glutamate and NO was reported by Lin et al. [37]that demonstrated co-localization of nNOS and glutamate in NTSneurons of rats, while studies by Ohta et al. [36] showed that themicroinjection of NOS inhibitor (L-NMMA) into the NTS reducedthe hypotensive response caused by microinjection of L-glutamateinto the NTS. Therefore, there is a series of functional evidenceindicating an important role for NO, formed by nNOS and/or eNOS,in the NTS of rats and the present study clearly indicates that NO iscontinuously formed in the NTS.
The present study shows the NO presence in the NTS and itssemi-quantitative analysis demonstrated that its concentration issimilar in the three antero-posterior aspects of the commissuralNTS evaluated. Previously Lawrence et al. [50] documented NOSimmunoreactivity in all antero-posterior aspects of the NTS withdifferent staining in each specific NTS sub-nuclei. Using electronmicroscopy, Atkinson et al. [61] showed that nNOS is present inpre- as well as in postsynaptic structures in the NTS and Linet al. [7] showed that: (a) nNOS was found only in neurons andeNOS only in astrocytes, (b) the immunoreactivity to these two
enzymes were never co-localized, and (c) a uniform distributionof eNOS-IR in the different antero-posterior aspects of the NTS.Taking in account that our method evaluated the NO formationand not the presence of eNOS or nNOS immunoreactivity, wemay suggest that the NO formed by eNOS in astrocytes or endothe-lium diffuses immediately to adjacent neurons where it was de-tected by the presence of DAF-2. This seems to be a possibility toexplain why in our study the NOIF was similar in the three differentrostral/caudal levels of the NTS studied. In accordance with Tho-mas et al. [62] an important determinant of NO concentration isthe easy diffusion of this molecule, since NO has a low molecularweight, it is uncharged, and it is soluble in both aqueous andhydrophobic environments, which turns it highly diffusible. Con-sidering that in most of the tissues, NO diffuses usually within arange of 50–300 lm and also that the diameter of NTS neuronsare in the range of 10 lm, it is possible that NO detected by DAF-2 DA in a neuron is not necessarily produced locally since NOcan easily diffuse from the neighboring neurons. Our data showedthat L-NAME was effective in decreasing the NOIF observed afterDAF-2 DA loading and D-NAME was not, indicating that the NOIF
was mainly due to the NO produced and/or diffused from cells con-taining NOS. However, we cannot exclude from the co-localizationexperiment presented in Fig. 8 that a small part of the NOIF ob-served in the NTS was due to extraneuronal NOIF produced and dif-fused from astrocytes and endothelium. A careful observation ofthe panels A and D of this figure shows several little NOIF dots thatmay represent the extraneuronal NO formed in astrocytes or inendothelium.
The data of the present study is in agreement with an interac-tion between NO production and L-glutamate receptors in theNTS. The antagonism of L-glutamate ionotropic receptors with

Fig. 7. GABAA receptor antagonist increases basal NOIF in NTS neurons. Confocal photomicrographs showing two representative images of the basal NOIF production in theNTS in control aCSF (control, A) and in aCSF containing the GABAa receptor antagonist bicuculine (Bic, 1 mM, B). Scale bar = 50 lm. NOIF signal was increased in the presenceof bicuculline. (C) Graphic showing that the bicuculline significantly increased NOIF in NTS neurons. The values of each bar correspond to the average of the data from threeantero-posterior regions of the NTS for each brainstem analyzed (n = 4). (�) different in relation to the control group, P < 0.05.
Fig. 8. NeuN and GFAP immunoreactivity and basal NOIF in the NTS. Confocal photomicrographs showing representative images of basal NOIF (A), NeuN immunoreactivity (B)in a NTS slice incubated in DAF-2 DA. (C) Superimposed images showing the co-localization of both signals. Arrows point the examples of double-labeled neurones. Confocalphotomicrographs showing basal NOIF (D), GFAP immunoreactivity (E) in a NTS slice incubated in DAF-2 DA. (F) Superimposed images showing that NOIF and GFAP are notco-localized. Scale bar = 50 lm.
214 G.P. Pajolla et al. / Nitric Oxide 20 (2009) 207–216

G.P. Pajolla et al. / Nitric Oxide 20 (2009) 207–216 215
kynurenic acid decreased DAF-2 DA fluorescence in NTS slices andthis effect was similar in the different antero-posterior aspects ofthe intermediate and caudal commissural NTS studied, suggestingthat activation of L-glutamate receptors increases the NO produc-tion in NTS. This finding for NTS neurons is similar to a previousstudy by Stern and Zhang [22] showing that the production ofNO in neurons of the supraoptic nucleus is affected by antagonistsof glutamate ionotropic receptors. Studies by Garthwaite and Boul-ton [63] documented that activation of NMDA receptors increasesintracellular Ca2+ concentration, which may activate the NOS witha consequent increase in the NO production.
In the present study we also verified that the antagonism of GA-BAA receptors with bicuculline increased the NO production in theNTS, suggesting an important inhibitory role for GABA on the pro-duction of NO in the NTS. However, further experiments are neces-sary to elucidate the mechanism by which GABA decreases the NOproduction in NTS as well as the physiological implications of thismodulation on the autonomic function to the cardiovascular system.Although little is known about the NO interaction with inhibitoryGABAergic transmission, there is a report showing that the overex-pression of endothelial NOS (eNOS) in the rostral ventrolateral me-dulla led to an increase in the extracellular concentrations of GABA[64]. Studies by Wang et al. [65] showed that NO alters firing of theGABAergic neurons in the NTS and change the release of this inhibi-tory transmitter, indicating that NO can also modulate GABAergictransmission. There is also evidence that the antagonism of GABAA
receptors in the hippocampus induces a potentiation of adenosineA1 receptor-mediated inhibitory action, an effect that involves NOacting through guanylyl cyclase [38]. This study by Fragata et al.[38] demonstrated that GABA receptor antagonist potentiated theinhibitory effect of adenosine and this effect was abolished whenNOS was inhibited, suggesting that GABA decreases the NO produc-tion induced by adenosine in hippocampus. Our data show thatGABA decreases the NO production, which can be related to a director indirect effect of GABA on NO production. To answer this impor-tant question further experiments are required.
In the central nervous system the nNOS has been shown to be themost prevalent isoform in neurons under physiological conditions,although, the presence of the endothelial and inducible isoformshas also been demonstrated in physiological conditions [66] butnot in the NTS [7]. The enzyme responsible for neuronal synthesisof NO has been demonstrated in neurons and terminals in the NTS[7,61,67,68]. However, the expression of NOS and NO productionwere also demonstrated in astrocytes and microglia [7,22,69–71]and for these reasons we also studied the source of basal NO detectedin the NTS slices. We used NeuN and GFAP to identify neurons andglial cells, respectively, and the double fluorescence with DAF-2DA was analyzed. Our results demonstrated that NO source reportedin our study is mainly from neurons, since NOIF presence was co-localized with NeuN fluorescence but not with GFAP fluorescence.
In conclusion the present study show that (a) it is possible todetect NO presence in NTS using the DAF-2 DA; (b) NO concentra-tion is similar in three antero-posterior levels of NTS studied and(c) the NO is present in the NTS neurons. Moreover, the NO produc-tion is modulated by glutamate and GABA. Further studies are re-quired for a better understanding the mechanisms underlying theinteractions among NO, GABA and glutamate in the NTS and itsimplications on the cardiovascular function in physiological andpathophysiological conditions.
Acknowledgments
This work was supported by FAPESP (Grants 2004/08712-0,2004/03285-7 and 2004/08868-0) and CNPq (Grant 472704/2004-4).
References
[1] S. Moncada, R.M.J. Palmer, E.A. Higgs, Nitric oxide: physiology,pathophysiology, and pharmacology, Pharmacol. Rev. 43 (1991) 109–142.
[2] D.S. Bredt, S.H. Snyder, Nitric oxide, a novel neuronal messenger, Neuron 8(1992) 3–11.
[3] S.J. Lewis, B.H. Machado, H. Ohta, W.T. Talman, Processing of cardiopulmonaryafferent input within the nucleus tractus solitarii involves activation of solubleguanylate cyclase, Eur. J. Pharmacol. 203 (1991) 327–328.
[4] S.J. Lewis, H. Ohta, B.H. Machado, J.N. Bates, W.T. Talman, Microinjection of S-nitrosocysteine into the nucleus tractus solitarii decreases arterial pressureand heart rate via activation of soluble guanylate cyclase, Eur. J. Pharmacol.202 (1991) 135–136.
[5] B.H. Machado, L.G.H. Bonagamba, Microinjection of S-nitrosocysteine into thenucleus tractus solitarii of conscious rats decreases arterial pressure but L-glutamate does not, Eur. J. Pharmacol. 221 (1992) 179–182.
[6] H. Ogawa, A. Mizusawa, Y. Kikuchi, W. Hida, H. Miki, K. Shirato, Nitric oxide asa retrograde messenger in the nucleus tractus solitarii of rats during hypoxia, J.Physiol. 486 (1995) 495–504.
[7] L.H. Lin, O. Taktakishvili, W.T. Talman, Identification and localization of celltypes that express endothelial and neuronal nitric oxide synthase in the ratnucleus tractus solitarii, Brain Res. 1171 (2007) 42–51.
[8] S. Harada, S. Tokunaga, M. Momohara, H. Masaki, T. Tagawa, T. Imaizumi, A.Takeshita, Inhibition of nitric oxide formation in the nucleus tractus solitariusincreases renal sympathetic nerve activity in rabbits, Circ. Res. 72 (1993) 511–516.
[9] A.J. Lawrence, Nitric oxide as a modulator of medullary pathways, Clin. Exp.Pharmacol. Physiol. 24 (1997) 760–763.
[10] H.C. Lin, F.J. Wan, K.K. Cheng, C.J. Tseng, Nitric oxide signaling pathwaymediates the L-arginine-induced cardiovascular effects in the nucleus tractussolitarii of rats, Life Sci. 65 (1999) 2439–2451.
[11] J.F.R. Paton, J. Deuchars, Z. Ahmad, L-F. Wong, D. Murphy, S. Kasparov,Adenoviral vector demonstrates that angiotensin II-induced depression of thecardiac baroreflex is mediated by endothelial nitric oxide synthase in thenucleus tractus solitarii of the rat, J. Physiol. 531 (2) (2001) 445–458.
[12] Y. Hirooka, Adenovirus-mediated gene transfer into the brain stem to examinecardiovascular function: role of nitric oxide and Rho-kinase, Prog. Biophys.Mol. Biol. 84 (2004) 233–249.
[13] W.C. Lo, M. Hsiao, C.S. Tung, C.J. Tseng, The cardiovascular effects of nitricoxide and carbon monoxide in the nucleus tractus solitarii of rats, J. Hypertens.22 (2004) 1182–1190.
[14] K. Sakai, Y. Hirooka, H. Shigematsu, T. Kishi, K. Ito, H. Shimokawa, A. Takeshita,K. Sunagawa, Overexpression of eNOS in brain stem reduces enhancedsympathetic drive in mice with myocardial infarction, Am. J. Physiol. HeartCirc. Physiol. 289 (2005) H2159–H2166.
[15] A.C. Dias, M. Vitela, E. Colombari, S.W. Mifflin, Nitric oxide modulation ofglutamatergic, baroreflex, and cardiopulmonary transmission in the nucleus ofthe solitary tract, Am. J. Physiol. Heart Circ. Physiol. 288 (2005) H256–H262.
[16] K. Kikuchi, T. Nagano, H. Hayakawa, Y. Hirata, M. Hirobe, Detection of nitricoxide production from a perfused organ by a luminal-H2O2 system, Anal.Chem. 65 (1993) 1794–1799.
[17] C.M. Arroyo, C. Forray, E.E. el-Fakahany, G.M. Rosen, Receptor-mediatedgeneration of an EDRF-like intermediate in a neuronal cell line detected byspin trapping techniques, Biochem. Biophys. Res. Commun. 170 (1990) 1177–1183.
[18] T. Malinski, Z. Taha, Nitric oxide release from a single cell measured in situ by aporphyrinic-based microsensor, Nature 358 (1992) 676–678.
[19] H. Kojima, N. Nakatsubo, K. Kikuchi, S. Kawahara, Y. Kirino, H. Nagoshi, Y.Hirata, T. Nagano, Detection and imaging of nitric oxide with novel fluorescentindicators: diaminofluoresceins, Anal. Chem. 70 (1998) 2446–2453.
[20] H. Kojima, N. Nakatsubom, K. Kikuchim, Y. Uranom, T. Higuchi, J. Tanaka, Y.Kudo, T. Nagano, Direct evidence of NO production in rat hippocampus andcortex using a new fluorescent indicator: DAF-2 DA, Neuroreport 9 (1998)3345–3348.
[21] N. Nakatsubo, H. Kojima, K. Kikuchi, H. Nagoshi, Y. Hirata, D. Maeda, Y. Imai, T.Irimura, T. Nagano, Direct evidence of nitric oxide production from bovineaortic endothelial cells using new fluorescent indicators: diaminofluoresceins,FEBS Lett. Chem. 427 (1998) 263–266.
[22] J.E. Stern, W. Zhang, Cellular sources, targets and actions of constitutive nitricoxide in the magnocellular neurosecretory system of the rat, J. Physiol. 562(2005) 725–744.
[23] B.E. Maley, Immunohistochemical localization of neuropeptides andneurotransmitters in the nucleus solitarius, Chem. Senses 21 (1996) 367–376.
[24] D.A. Ruggiero, S.L. Cravo, E. Golanov, R. Gomez, M. Anwar, D.J. Reis, Adrenergicand non-adrenergic spinal projections of a cardiovascular-active pressor areaof medulla oblongata: quantitative topographic analysis, Brain Res. 663 (1994)107–120.
[25] B.H. Machado, Neurotransmission of the cardiovascular reflexes in the nucleustractus solitarii of awake rats, Ann. NY Acad. Sci. 940 (2001) 179–196.
[26] B.H. Machado, Chemoreflex and sympathoexcitation, in: N.J. Dun, B.H.Machado, P.M. Pilowsky (Eds.), Neural Mechanisms of CardiovascularRegulation, Kluwer Academic Press, Boston, 2004, pp. 31–58.
[27] F. Kato, E. Shigetomi, Distinct modulation of evoked and spontaneous EPSCs bypurinoceptors in the nucleus tractus solitarii of the rat, J. Physiol. 530 (2001)469–486.

216 G.P. Pajolla et al. / Nitric Oxide 20 (2009) 207–216
[28] V. Baptista, W.A. Varanda, Glycine binding site of the synaptic NMDA receptorin subpostremal NTS neurons, J. Neurophysiol. 94 (2005) 147–152.
[29] W.N. Ogawa, V. Baptista, J.F. Aguiar, W.A. Varanda, Neurotensin modulatessynaptic transmission in the nucleus of the solitary tract of the rat,Neuroscience 130 (2005) 309–315.
[30] D. Accorsi-Mendonca, R.M. Leao, J.F. Aguiar, W.A. Varanda, B.H. Machado,Urethane inhibits the GABAergic neurotransmission in the nucleus of thesolitary tract of rat brain stem slices, Am. J. Physiol. Regul. Integr. Comp.Physiol. 292 (2007) R396–R402.
[31] A. Mizusawa, H. Ogawa, Y. Kikuchi, W. Hida, H. Kurosawa, S. Okabe, T.Takishima, K. Shirato, In vivo release of glutamate in nucleus tractus solitarii ofthe rat during hypoxia, J. Physiol. 478 (1994) 55–66.
[32] R.M. Bradley, M.S. King, L. Wang, X. Shu, Neurotransmitter andneuromodulator activity in the gustatory zone of nucleus tractus solitarius,Chem. Senses 21 (1996) 377–385.
[33] A.J. Lawrence, B. Jarrot, Neurochemical modulation of cardiovascular control inthe nucleus tractus solitarius, Prog. Neurobiol. 48 (1996) 21–53.
[34] J. Garthwaite, Glutamate, Nitric oxide and cell–cell signaling in the nervoussystem, Trends Neurosci. 14 (1991) 60–67.
[35] P.R. Montague, C.D. Gancayco, M.J. Winn, R.B. Marchase, M.J. Friedlander, Roleof NO production in NMDA receptor-mediated neurotransmitter release incerebral cortex, Science 263 (1994) 973–977.
[36] H. Ohta, J.N. Bates, S.J. Lewis, W.T. Talman, Actions of S-nitrosocysteine in thenucleus tractus solitarii are unrelated to release of nitric oxide, Brain Res. 746(1997) 98–104.
[37] L.H. Lin, P.C. Emson, W.T. Talman, Apposition of neuronal elements containingnitric oxide synthase and glutamate in the nucleus tractus solitarii of rat: aconfocal microscopic analysis, Neuroscience 96 (2000) 341–350.
[38] I.R. Fragata, J.A. Ribeiro, A.M. Sebastião, Nitric oxide mediates interactionsbetween GABAA receptors and adenosine A1 receptors in the rat hippocampus,Eur. J. Pharmacol. 543 (2006) 32–39.
[39] N. Kasim, R.L. Branton, D.J. Clarke, Neuronal nitric oxide synthaseimmunohistochemistry and 4,5-diaminofluorescein diacetate: tools for nitricoxide research, J. Neurosci. Methods 112 (2001) 1–8.
[40] N. Rodriguez, F. Pincet, S. Cribier, Giant vesicles formed by gentle hydrationand electroformation: a comparison by fluorescence microscopy, Colloids Surf.B Biointerfaces 42 (2005) 125–130.
[41] G. Paxinos, C. Watson, The Rat Brain in Stereotaxic Coordinates, AcademicPress, New York, 1998.
[42] H. Corridi, G. Jonsson, The formaldehyde fluorescence method for thehistochemical demonstration of biogenic monoamines—A review on themethodology, J. Histochem. Cytochem. 15 (1967) 65–78.
[43] D.R. Dunn, R.F. Barth, Identification of melanoma cells by formal dehyde-induced fluorescence, Cancer 33 (1974) 701–706.
[44] A. Gomes, E. Fernandes, J.L. Lima, Use of fluorescence probes for detection ofreactive nitrogen species: a review, J. Fluoresc. 16 (2006) 119–139.
[45] L.A. Brown, B.J. Key, T.A. Lovick, Bio-imaging of nitric oxide-producingneurones in slices of rat brain using 4,5-diaminofluorescein, J. Neurosci.Methods 92 (1999) 101–110.
[46] T. Kubo, J-L. Yue, Y. Goshima, S. Nakamura, Y. Misu, Evidence for L-DOPAsystems responsible for cardiovascular control in the nucleus tractus solitariiof the rat, Neurosci. Lett. 140 (1992) 153–156.
[47] J. Riley, L.H. Lin, D.A.J. Chianca, W.T. Talman, Ablation of NK1 receptors in ratnucleus tractus solitarii blocks baroreflexes, Hipertension 40 (2002) 823–826.
[48] E.M. Schuman, D.V. Madison, Nitric oxide and synaptic functions, Annu. Rev.Neurosci. 17 (1994) 153–183.
[49] J. Rodrigo, D.R. Springall, O. Uttenthal, M.L. Bentura, F. Abadia-Molina, V.Riveros-Moreno, R. Martinez-Murillo, J.M. Polak, S. Moncada, Localization ofnitric oxide synthase in the adult rat brain, Philos. Trans. R. Soc. Lond. B Biol.Sci. 345 (1994) 175–221.
[50] A.J. Lawrence, M. Castillo-Melendez, K.J. McLean, B. Jarrot, The distribution ofnitric oxide synthase-, adenosine deaminase- and neuropeptide Y-immunoreactivity through the entire rat nucleus tractus solitarius: effect ofunilateral nodose ganglionectomy, J. Chem. Neuroanat. 15 (1998) 27–40.
[51] F.O. Esteves, P.N. McWilliam, T.F. Batten, Nitric oxide producing neurones inthe rat medulla oblongata that project to nucleus tractus solitarii, J. Chem.Neuroanat. 20 (2000) 185–197.
[52] W.T. Talman, D.N. Dragon, H. Ohta, L.H. Lin, Nitroxidergic influences oncardiovascular control by NTS: a link with glutamate, Ann. NY Acad. Sci. 940(2001) 169–178.
[53] W.T. Talman, D.N. Dragon, Transmission of arterial baroreflex signals dependson neuronal nitric oxide synthase, Hypertension 43 (2004) 820–824.
[54] I. Sakuma, H. Togashi, M. Yoshioka, H. Saito, M. Yanagida, M. Tamura, T.Kobayashi, H. Yasuda, S.S. Gross, R. Levi, NG-methyl-L-arginine, an inhibitor ofL-arginine-derived nitric oxide synthesis, stimulates renal sympathetic activityin vivo, Circ. Res. 70 (1992) 607–611.
[55] C.J. Tseng, H.Y. Liu, H.C. Lin, L.P. Ger, C.S. Tung, M.H. Yen, Cardiovascular effectsof nitric oxide in the brainstem nuclei of rats, Hypertension 27 (1996) 36–42.
[56] H. Togashi, I. Sakuma, M. Yoshioka, T. Kobayashi, H. Yasuda, A. Kitabatake, H.Saito, S.S. Gross, R. Levi, A central nervous system action of nitric oxide inblood pressure regulation, J. Pharmacal. Exp. Ther. 262 (1992) 343–347.
[57] T. Tagawa, T. Imaizumi, S. Harada, T. Endo, M. Shiramoto, Y. Hirooka, A.Takeshita, Nitric oxide influences neuronal activity in the nucleus tractussolitarius of rat brainstem slices, Circ. Res. 75 (1994) 70–76.
[58] S. Wang, J.F. Paton, S. Kasparov, Differential sensitivity of excitatory andinhibitory synaptic transmission to modulation by nitric oxide in rat nucleustractus solitarii, Exp. Physiol. 92 (2007) 371–382.
[59] E.D. Di Paola, M.J. Vidal, G. Nistico, L-Glutamate evokes the release of anendothelium-derived relaxing factor-like substance from the rat nucleustractus solitarius, J. Cardiovasc. Pharmacol. 17 (1991) S269–S272.
[60] W.C. Lo, H.C. Lin, L.P. Ger, C.S. Tung, C.J. Tseng, Cardiovascular effects of nitricoxide and N-methyl-D-aspartate receptors in the nucleus tractus solitarii ofrats, Hypertension 30 (1997) 1499–1503.
[61] L. Atkinson, T.F. Batten, E.K.A. Corbett, J.K. Sinfield, J. Deuchars, Subcellularlocalization of neuronal nitric oxide synthase in the rat nucleus of the solitarytract in relation to vagal afferent inputs, Neuroscience 118 (2003) 115–122.
[62] D.D. Thomas, L.A. Ridnour, J.S. Isenberg, W. Flores-Santana, C.H. Switzer, S.Donzelli, P. Hussain, C. Vecoli, N. Paolocci, S. Ambs, C. Colton, C.C. Harris, D.D.Roberts, D.A. Wink, The chemical biology of nitric oxide: implications incellular signaling, Free Radic. Biol. Med. 45 (2008) 18–31.
[63] J. Garthwaite, C.L. Boulton, Nitric oxide signaling in the central nervoussystem, Annu. Rev. Physiol. 57 (1995) 683–706.
[64] T. Kishi, Y. Hirooka, K. Sakai, H. Shigematsu, H. Shimokawa, A. Takeshita,Overexpression of eNOS in the RVLM causes hypotension and bradycardia viaGABA release, Hypertension 38 (2001) 896–901.
[65] S. Wang, A.G. Teschemacher, J.F. Paton, S. Kasparov, Mechanism of nitric oxideaction on inhibitory GABAergic signaling within the nucleus tractus solitarii,FASEB J. 20 (2006) 1537–1539.
[66] G. Bhat, V.B. Mahesh, K. Aguan, D.W. Brann, Evidence that brain nitric oxidesynthase is the major nitric oxide synthase isoform in the hypothalamus of theadult female rat and that nitric oxide potently regulates hypothalamic cGMPlevels, Neuroendocrinology 64 (1996) 93–102.
[67] Z.K. Krowicki, K.A. Sharkey, S.C. Serron, N.A. Nathan, P.J. Hornby, Distributionof nitric oxide synthase in rat dorsal vagal complex and effects ofmicroinjection of nitric oxide compounds upon gastric motor function, J.Comp. Neurol. 377 (1997) 49–69.
[68] L.H. Lin, A. Sandra, S. Boutelle, W.T. Talman, Up-regulation of nitric oxidesynthase and its mRNA in vagal motor nuclei following axotomy in rat,Neurosci. Lett. 221 (1997) 97–100.
[69] R.C.C. Chang, P. Hudson, B. Wilson, B. Liu, H. Abel, J. Hemperly, J.S. Hong,Immune modulatory effects of neural cell adhesion molecules onlipopolysaccharide-induced nitric oxide production by cultured glia, Mol.Brain Res. 81 (2000) 197–201.
[70] Q. Si, Y. Nakamura, K. Kataoka, Factor enhances production of nitric oxide andtumor necrosis factor-a from cultured microglia, Exp. Neurol. 162 (2000) 89–97.
[71] A. Bal-Price, G.C. Brown, Inflammatory neurodegeneration mediated by nitricoxide from activated glia-inhibiting neuronal respiration, causing glutamaterelease and excitotoxicity, J. Neurosci. 21 (2001) 6480–6491.
Copyright © 2022 FDOKUMEN




















