
Independent Component Analysis in the Study of Focal Seizures
Upload
independentCategory
view
5download
0
Febrile Seizures Impair Memory and cAMPResponse-Element Binding
Protein ActivationYing-Chao Chang, MD,1 A-Min Huang, PhD,2 Yu-Min Kuo, PhD,3 Shan-Tair Wang, PhD,4
Yung-Yee Chang, MD,5 and Chao-Ching Huang, MD6
The long-term effects of brief but repetitive febrile seizures (FS) on memory have not been as thoroughly investigated asthe impact of single and prolonged seizure in the developing brain. Using a heated-air FS paradigm, we subjected malerat pups to one, three, or nine episodes of brief FS on days 10 to 12 postpartum. Neither hippocampal neuronal damagenor apoptosis was noted within 72 hours after FS, nor was there significant hippocampal neuronal loss, aberrant mossyfiber sprouting, or altered seizure threshold to pentylenetetrazol in any FS group at adulthood. The adult rats subjectedto nine episodes of early-life FS, however, showed long-term memory deficits as assessed by the Morris water maze. Theyalso exhibited impaired intermediate and long-term memory but spared short-term memory in the inhibitory avoidancetask. Three hours after inhibitory avoidance training, phosphorylation of cAMP response-element binding (CREB) pro-tein in the hippocampus was significantly lower in nine-FS-group rats than in controls. Furthermore, rolipram admin-istration, which activated the cAMP–CREB signaling pathway by inhibiting phosphodiesterase type IV, reversed thelong-term memory deficits in nine-FS-group rats by enhancing hippocampal CREB phosphorylation. These results raiseconcerns about the long-term cognitive consequences of even brief frequently repetitive FS during early brain develop-ment.
Ann Neurol 2003;54:706–718
Febrile seizure (FS) is the most common convulsivedisorder in young children, affecting 2 to 4% beforeage 5 years.1,2 One third of those with FS have recur-rent FS by age 6 years, and the incidence of subsequentepilepsy is 2 to 8%.1–3 The causal relationship betweenFS and subsequent epilepsy and cognitive deficits hasalways been a focus of intense investigation. Populationstudies have not shown the associations between early-life FS and temporal lobe epilepsy.4,5 Nonetheless, ahistory of prolonged FS was commonly elicited in in-dividuals with mesial temporal lobe epilepsy, which fre-quently was associated with memory deficits.6,7 Al-though population studies did not show cognitivedisadvantages in children with FS,8–11 mental retarda-tion was reported in up to 22% of children with FSwho were seen in hospitals.12 These irreconcilable ob-servations from human studies may be resolved by us-ing animal models to induce FS and recording the cel-lular consequences.
Recent data indicated that single and prolonged FS
in immature rats could induce long-term functionalchanges in hippocampal circuitry and increase seizuresusceptibility.13–16 These studies examined the impactsof prolonged convulsions, whereas most children withFS experience brief and recurrent seizures.1,2 Findingson the effects of recurrent FS on mental developmentare contradictory. Verity and colleagues8 and Ellenbergand Nelson9 and found no difference in IQ betweenchildren with single FS and those with recurrent FS,whereas Kolfen and colleagues17 found that childrenwith multiple FS performed less well in all neuropsy-chological tests than controls or those with single FS.Studies on the effects of chemically induced repetitiveseizures in immature animals have demonstrated thateven relatively short-lasting seizures, when repetitive,can result in long-term detrimental effects on behavior,seizure susceptibility, and brain development.18–20 Theconvulsants used in these experimental paradigms,however, are also potent hippocampal neurotoxins.Thus, evaluation of repetitive seizure effects on the de-
From the 1Graduate Institute of Clinical Medicine and Departmentof Pediatrics, Chang Gung Memorial Hospital, Kaohsiung, Taiwan;2Department of Physiology, National Cheng Kung University, Col-lege of Medicine, Tainan, Taiwan; Institutes of 3Anatomy and CellBiology and 4Epidemiology, National Cheng Kung University, Col-lege of Medicine, Tainan, Taiwan; 5Department of Neurology,Chang Gung Memorial Hospital, Kaohsiung, Taiwan; 6Institute ofMolecular Medicine and Department of Pediatrics, National ChengKung University, College of Medicine, Tainan, Taiwan.
Received Mar 28, 2003, and in revised form Jul 7 and Aug 15.Accepted for publication Aug 20, 2003.
Address correspondence to Dr Huang, Department of Pediatrics,National Cheng Kung University Hospital, Tainan City 704, Tai-wan. E-mail: [email protected]
ORIGINAL ARTICLE
706 © 2003 American Neurological AssociationPublished by Wiley-Liss, Inc., through Wiley Subscription Services

veloping brain may unavoidably be influenced by theneurotoxic effects.
Relatively few studies have focused on the conse-quences of brief repetitive FS on learning, memory,and neuronal plasticity in the developing hippocampus.Therefore, the aim of this study was to determinewhether brief and repetitive FS in the immature rat (1)produced acute neuronal damage, chronic neuronalloss, and aberrant sprouting of mossy fiber in the hip-pocampus; (2) altered seizure susceptibility duringadulthood; or (3) impaired hippocampus-dependentlearning and memory.
The cAMP response element-binding protein(CREB) is a key mediator of stimulus-induced nuclearresponses that underlie the plasticity of the nervous sys-tem.21–24 CREB is a central molecular switch for theconversion of short-term memory to long-term mem-ory (LTM).23 LTM requires the phosphorylation ofCREB on Ser133 by cAMP-dependent protein kinase(PKA) and consequently binds to the cAMP responseelement of target genes.21–23 In this study, we alsotested the hypothesis that LTM deficits observed afterrepetitive early-life FS were associated with alteredCREB phosphorylation in the hippocampus.
Materials and MethodsExperimental Febrile Seizure in Rat PupsOn postnatal (P) days 10 to 12, a group of three rat pupswas placed into a 3 to l glass container (hyperthermia cham-ber) in which hyperthermia was induced by a regulatedstream of heated air from an adjustable hair dryer located50cm above the container.13 The chamber was in a waterbath maintained at 37°C, and the behaviors of rat pups inthe chamber were continously monitored. The rectal temper-ature of the rat pups was measured at baseline, 2-minute in-tervals after hyperthermia, and the onset of behavioral sei-zures. When their rectal temperature reached approximately40 to 41°C, the pups became agitated and showed head bob-bing or shaking, clonic twitching of hindlimbs, behavioralarrest or biting, and chewing of limbs (stage I FS). At 41 to43°C rectal temperature, the pups exhibited generalizedmyoclonic jerk, wide-running, ataxic walking, circling, fall-ing, or tonic-clonic seizures of limbs (stage II FS). After 10minutes in the chamber at 40 to 43°C rectal temperature,the pups were removed immediately and placed on a coolsurface until they regained posture and their core tempera-ture returned to baseline. The FS latency was determinedfrom the time the rats had been placed in the hyperthemicchamber to the onset of stage I FS. The seizure duration wasrecorded during the 10 minutes of hyperthermia within thechamber.
The rat pups were divided into four groups: (1) single FSon P10 (single FS [SFS]); (2) one FS daily from P10 to P12(three FS total) (repetitive FS [RFS)]; (3) three FS daily, atan interval of 4 hours between seizures, from P10 to P12(nine FS total; frequently repetitive FS [FRFS]); and (4) con-trols. The controls were removed from the cage along withFS groups during the experiment but were not subjected to
the hyperthermic chamber. Another group of pups, servingas a hyperthermia control, was subjected to nine episodes ofhyperthermia along with FRFS group, but seizures were pre-vented each time by pretreatment with pentobarbital (20–30mg/kg, IP). In total, 28 dams and their litters (n � 310)were used in this study. Ten to 12 pups per dam were usedand housed with a 12-hour-light/12-hour-dark schedule. Thepups were weaned on P21 and housed in groups of our ratsper cage. This study was approved by the Animal Care Com-mittee at National Cheng Kung University College of Med-icine.
HistopathologyACUTE STAGE: CRESYL VIOLET STAIN AND IN SITU DNANICK END LABELING STAIN. After anesthetized with pen-tobarbital (60mg/kg, IP), brains from controls and experi-mental rats 24 and 72 hours after the last FS were removed,immediately frozen, and stored at �80°C. Frozen coronalsections (20�m) of entire hippocampus were fixed in form-aldehyde for cresyl violet or in situ DNA nick end labeling(TUNEL) stain as described previously.25 In brief, after pre-incubation in hydrogen peroxide and sodium azide, slideswere immersed in terminal deoxynucleotidyl transferase(TdT) buffer at room temperature for 15 minutes, followedby TdT and biotinylated 16-uridine-5�-triphosphate at 37°Cfor 60 minutes. The reaction was stopped with sodium ci-trate and sodium chloride, followed by incubation with 2%bovine serum albumin in phosphate-buffered saline. Thecleaved DNA was visualized using an avidin-biotin kit (Vec-tor Laboratories, Burlingame, CA). Sections from rats afterfocal cerebral ischemia were run in parallel as “positive con-trols.”25
CHRONIC STAGE: TIMM STAIN AND CRESYL VIOLET STAIN.On P60, rats were deeply anesthetized with pentobarbitaland then perfused transcardially with 0.58% sodium sulfidefollowed by 4% paraformaldehyde (PFA). Their brains werepostfixed in PFA for 24 hours and then placed in a 30%sucrose solution until they sank to the bottom. Coronal sec-tions (20�m) through the entire hippocampus were cut on afreezing microtome. Every sixth section was stained withTimm stain.19,20 In brief, the sections were developed in thedark for 10 to 45 minutes in a 200ml solution containing5.1gm citric acid, 4.7gm sodium citrate, 3.47gm hydroqui-none, 212.25mg AgNO3, and 120ml 50% arabic gum.Timm staining was assessed from the septal area to the tem-poral hippocampus (region between 2.8 and 3.8mm poste-rior to the bregma). The degree of mossy fiber sprouting wasevaluated with a semiquantitative scale in the pyramidal andinfrapyramidal area of the cornu ammonis region of the hip-pocampus (CA3) region, and granular cell and inner molec-ular layers of the dentate gyrus, avoiding the edge andcrest.20 The score criteria were 0 � no granules; 1 � occa-sional discrete granule bundles; 2 � occasional to moderategranules; 3 � prominent granules; 4 � prominent near-continuous granule band; and 5 � continuous or near-continuous dense granule band.
Cell numbers were counted in Nissl-stained sections(5�m). Unbiased stereological cell counts were performedusing the optical dissector method.19,20,26 The region ana-
Chang et al: Memory in Febrile Seizures 707

lyzed was similarly to the region for Timm stain scores.Counts were at �400 magnification using a computerizedimage analysis system (Media Cybernetics, Silver Spring,MD), and the hippocampal subfields were defined by animaginary line connecting the tips of the granule cell layerblades, which separated CA3c (medially) from CA3b (later-ally), and CA2 from CA1.27 A counting box 200�m �150�m was placed over the region of interest. For the stra-tum pyramidal and granule cell layer, the laminar profile ofneurons within a 200�m strip in each subfield was counted.The hilar neurons within 200�m strips from the crest, ex-cluding the CA3c pyramidal neurons, were counted.26 Val-ues from bilateral hippocampi were obtained blindly and av-eraged.
The same sections for cell counts were used for volumemeasurement. At �40 magnification, areas of CA3, CA1,the hilus, and dentate gyrus were obtained manually by trac-ing the histological area. Volume was calculated by multiply-ing the mean area of two adjacent specimens by the distancebetween the two and then adding the volumes for the entireanalyzed hippocampus length.19,20
Seizure SusceptibilityOn P60, rats were placed in individual cages and given pen-tylenetetrazol (PTZ; 10mg/kg IP every 5 minutes) until theonset of retropulsive myoclonus, defined as a myoclonic jerkresulting in backward movement of the head and shoulders.Dosage and time to the onset of myoclonic jerk and gener-alized clonic activity was calculated.19,20
Behavioral TestingMORRIS WATER MAZE. On P36, rats underwent the Mor-ris water maze to assess hippocampus-dependent spatialmemory.18–20 A circular pool (160cm diameter � 50cmhigh) was filled with water (26°C) to a depth of 25cm. Thepool, illuminated by room lights, was divided into fourquadrants. An 8 � 8cm platform, onto which rats couldescape from the water, was positioned in the center of one ofthe quadrants, 1cm below the water surface. Escape latency,
escape distance, and swimming patterns of rats were moni-tored by a computer program (EthoVision 2.3; Noldus In-formation Technology, Wageningen, the Netherlands).
On days 1 to 4, rats were given 24 training sessions (sixper day) to escape onto the submerged platform. The plat-form quadrant location remained constant, but the point ofentry varied in a quasi-random order. On day 5, the plat-form was removed, and the rats were placed in the quadrantopposite the previous platform position. The rats were al-lowed 60 seconds of free swimming, and the time in eachquadrant was recorded. After the task, the rats were submit-ted to a visually cued task to locate a green platform 2cmabove the water surface. The platform position and the entryin the pool were changed quasi-randomly across trials. Thevisual motor task was conducted over 2 days, with eight trialsper day split between the daily sessions, with 2 hours be-tween sessions.
INHIBITORY AVOIDANCE TASK. The single-trial inhibitoryavoidance (IA) task, another hippocampus-dependent behav-ior test, was used to measure different phases of memory inrats on P45.24,28 The apparatus consisted of one illuminatedcompartment and one dark compartment. A shock generatorwas connected to the floor of the dark compartment. Beforethe experiment, the rat was kept in a dim room for 1 hour toadjust to the environment. In the training phase, the rat wasplaced in the illuminated compartment facing away from thedoor. As the rat turned around, the door was opened. Whenthe rat entered the dark compartment, the door was closed,and the rat was given a 1.0mA/1-second shock. The reactionto the shock was graded as flinch, vocalization, or locomo-tion. The rat then was removed from the alley and returnedto its home cage. The retention test was given 1, 3, or 24hours after training for the measurement of short-term, in-termediate, and long-term memory, respectively. The rat wasagain placed in the illuminated compartment, and the la-tency to step into the dark compartment was recorded as themeasure of retention performance. Rats that did not enter
Table. Magnitude of Hyperthermia at Stage I and II FS, and Latency of FS in Rat Pups with Different RecurrentRates of FS (n � 40 per group)
Group
BodyWeight
(gm)
BasalTemperature
(°C)
MaximumTemperature
(°C)
Stage I FSTemperature
Threshold(°C)
Stage II FSTemperature
Threshold(°C)
FSDuration
(min)
FSLatency(min)
First FS on P10Control 20.1 � 0.6 33.6 � 0.2 — — — — —SFS 20.4 � 0.5 33.5 � 0.2 42.7 � 0.1 41.0 � 0.2 42.5 � 0.3 4.4 � 0.2 4.1 � 0.2RFS 19.9 � 0.6 33.5 � 0.2 42.6 � 0.2 41.1 � 0.3 42.5 � 0.2 4.3 � 0.2 4.1 � 0.2FRFS 20.0 � 0.5 33.4 � 0.2 42.7 � 0.2 41.1 � 0.3 42.6 � 0.3 4.5 � 0.2 4.2 � 0.2
Last FS on P12Control 24.9 � 0.7 33.6 � 0.2 — — — — —SFS 24.9 � 0.5 33.5 � 0.2 — — — — —RFS 22.8 � 0.5a 33.5 � 0.2 42.7 � 0.1 41.2 � 0.3 42.5 � 0.2 4.4 � 0.2 4.0 � 0.2FRFS 22.6 � 0.6a 33.4 � 0.2 42.7 � 0.2 41.1 � 0.3 42.6 � 0.3 4.3 � 0.2 3.9 � 0.2
FS � febrile seizure; SFS � single FS group; RFS � repetitive FS group; FRFS � frequently repetitive FS group.ap � 0.05.
708 Annals of Neurology Vol 54 No 6 December 2003

the dark compartment within 600 seconds were removedfrom the alley.
For Western blot assessment, the hippocampi were dis-sected 3 hours after IA training. Another two groups of con-trols, untrained (naive) controls and unpaired shock controlsthat had stayed in the dark compartment for 20 minutesbefore receiving the shock (unpaired learning control), wereused for comparison.
Effect of Rolipram on Inhibitory Avoidance Task inFrequently Repetitive Febrile Seizure GroupIn another experiment, rolipram (5mg/kg, IP) or its vehicle(dimethyl sulfoxide) was administered 20 minutes after IAtraining in the control, SFS, and FRFS groups on P45.Twenty-four hours after training, the latency before each ratstepped into the dark compartment was measured. For theassessment of rolipram’s effect on CREB phosphorylation,the hippocampi were removed 3 hours after IA training fromrats treated with rolipram or vehicle.
Western Blot AnalysisHippocampi were homogenized in ice-cold lysis buffer andresolved by sodium dodecyl sulfate–polyacylamide gel beforeelectroblotting. Membranes were incubated with primary an-tibodies and visualized with an enhanced chemiluminescence
detection system (Pierce Biotechnology, Rockford, IL) beforedensitometric analysis. The blots then were stripped with Re-store Western Blot Stripping Buffer (Pierce Biotechnology)for 15 minutes and incubated with the antibodies. TheWestern blots were repeated in at least four independent ex-periments. The increase in phosphorylation was normalizedto the naive control and expressed as a fold increase. Thefollowing primary antibodies were used: anti-phospho-Ser133-CREB (pCREB, diluted 1:1,000; Upstate Biotech-nology, Lake Placid, VA), anti-CREB (diluted 1:1,000; Up-state Biotechnology), and anti-synaptophysin (diluted1:1,000; Santa Cruz Biotechnology, Santa Cruz, CA).
Phospho-Ser133-CREB ImmunohistochemistrypCREB immunohistochemistry was performed according toGinty and colleagues.29 Three hours after IA training, ratswere perfused with 4% PFA, and the brains were postfixedin PFA for 24 hours and then cryoprotected in 30% su-crose solutions. Cryostat coronal sections (20�m) were in-cubated with anti–pCREB antibody (1:200), and specificimmune complexes were visualized with an avidin-biotinsystem (Vector Laboratories). In each pCREB immunohis-tochemistry staining, negative control staining also was per-formed with normal rabbit serum as a substitute for theprimary antibody.
Fig 1. Frequently repetitive febrile seizures in the immature brain did not cause acute neuronal damage or apoptosis in the hip-pocampus. Photomicrographs of cresyl violet–stained coronal sections of the hippocampus from control (A, E, I, M) and 72 hoursafter the last febrile seizure (FS) from single FS (B, F, J, N), repetitive FS (C, G, K, O) and frequently repetitive FS (D, H, L,P) groups. Higher magnification of the boxed areas showing no gross cell loss after FS in CA1 (E, F, G, H), CA3 (I, J, K, L), orhilus (M, N, O, P) regions (n � 8 for each group) in any FS group. TUNEL stain showed no apoptotic neurons in the CA1 re-gion 72 hours after the last FS from rat pups that had undergone frequently repetitive FS (Q; n � 6 for each group). In contrast,sections from rats subjected to focal cerebral ischemia contained abundant labeled neurons in the CA1 region (R). Scale bars �400�m (A–D) or 50�m (E–R).
Chang et al: Memory in Febrile Seizures 709

StatisticsStatistical significance (p � 0.05) was determined using one-way analysis of variance (SPSS, Chicago, IL). When datawere not normally distributed, analyses were performed usinga nonparametric analysis of variance by ranks (Kruskal–Wal-lis one-way test) followed by Dunn’s multiple comparisontests. Repeated measures analysis of variance was used wheredays or quadrants were treated as a within-subject factor andgroups as a between-subject factor. Continuous data are ex-pressed as mean � SEM unless indicated otherwise.
ResultsTemperature Threshold for Inducing Febrile SeizureDid Not Change with Repetitive SeizuresMortality was not observed in any FS group duringhyperthermia or follow-up. There was no significantdifference in the temperature threshold for inducing FSbetween any FS group across different stages of FS (Ta-ble). Likewise, there were no significant differences inseizure latency, FS duration, or maximum rectal tem-perature between them. From P10 to P12, pups un-dergoing RFS or FRFS showed significantly less bodyweight gain than did the other groups. In adulthood,however, body weights were comparable in all fourgroups.
Febrile Seizures Did Not Cause Acute HippocampalNeuronal Damage or ApoptosisThe hippocampus, including its subregions, did notdemonstrate gross neuronal loss at 24 or 72 hours afterthe last FS in any FS group (Fig 1, A–P). No apoptoticneurons could be labeled in the hippocampus in con-trols or any FS group 72 hours after the last FS (seeFig 1Q, R). Brain sections from rats subjected to focalcerebral ischemia contained abundant labeled neuronsin the hippocampus, confirming the validity ofTUNEL method.
Early-life Febrile Seizure Did Not Cause ChronicHippocampal Neuronal Loss or Aberrant MossyFiber SproutingCresyl violet staining showed no detectable hippocam-pal neuronal loss in adult rats in any FS group. Therewere no differences in cell counts in the hippocampalCA3a, CA3b, CA1, hilus, or dentate granular cell layerbetween controls and any FS group (Fig 2A). Nor didthe volumes of these subregions differ between the fourgroups (see Fig 2B). Similarly, the degree of mossy fi-ber sprouting into the hippocampal CA3 or dentategyrus in FRFS group did not differ from that in anyother group (Fig 3, A–J).
Early-life Febrile Seizure Did Not Alter SeizureThreshold to Pentylenetetrazol at AdulthoodPTZ injection, a nonspecific GABA receptor antagonist,elicited seizures in all groups. There were no significant
differences between the four groups in the PTZ dosage(mg/kg) required to elicit seizures (controls: 63.8 � 4.0;SFS: 64.1 � 5.7; RFS: 63.0 � 5.3; FRFS: 60.2 � 4.5).Likewise, there were no significant differences in the la-tency (seconds) of myoclonic jerks (controls: 1,708 �96; SFS: 1,653 � 93; RFS: 1,742 � 103; FRFS:1,578 � 72) or generalized clonic activity (controls:1,733 � 90; SFS: 1,725 � 88; RFS: 1,742 � 95;FRFS: 1,698 � 72), although the FRFS group tendedto have a shorter latency for both seizure types.
Early-life Frequently Repetitive Febrile SeizureResulted in Hippocampus-dependent Long-termMemory Deficits at AdulthoodMORRIS WATER MAZE. During acquisition, all groupsshowed a decrease in escape time across days, indicat-ing learning of the platform position (p � 0.001).There was, however, a significant group differencefrom days 2 to 4 (p � 0.05). Post hoc multiple com-parisons showed that the FRFS group spent signifi-cantly more time finding the submerged platform thaneach of the remaining groups (all p � 0.05). On theprobe test, there was a significant group–quadrant in-
Fig 2. Frequently repetitive febrile seizures (FRFSs) in earlylife did not cause chronic neuronal loss in the hippocampus atadulthood (postnatal day 60). No significant differences in celldensity (A) or reference volume (B) in the various hippocam-pal subfields were found between control, single febrile seizure(SFS), repetitive febrile seizures (RFS), and FRFS groups(n � 8 for each group). DG � dentate gyrus.
710 Annals of Neurology Vol 54 No 6 December 2003

teraction on time distribution in the various quadrants(p � 0.05). The FRFS group spent significantly lesstime in the target quadrant than did controls, SFS, andRFS groups (all p � 0.05), but there was no significantdifference between control, SFS, and RFS groups. Toexclude the possibility of motor performance deficits,we gave rats a visually cued task that all four groupsacquired in a similar manner (Fig 4A–C).
INHIBITORY AVOIDANCE TASK. The initial latency inentering the dark compartment during training did notdiffer between the four groups. At 1 hour after train-ing, control and FS groups showed similar retentiontime, whereas, at 3 hours and especially at 24 hoursafter training, only the FRFS group showed signifi-cantly shorter retention time than controls (p � 0.01;see Fig 4D). There were no differences between the
groups in the pain responses of flinch, vocalization, orlocomotion to foot shock (see Fig 4E).
Rats with Frequently Repeated Hyperthermia AloneDid Not Have Long-term Memory Deficitsat AdulthoodTo examine whether the LTM deficits observed in theFRFS group were specifically caused by FRFS insteadof repeated hyperthermia, we pretreated rat pups withpentobarbital or normal saline each time before sub-jecting them to hyperthermia. There was no differencein maximum rectal temperature between the FRFS(42.7 � 0.1°C) and frequently repeated hyperthermiagroups (42.6 � 0.2°C) during the 10 minutes in thehyperthermic chamber. In contrast with the FRFSgroup, the rats with frequently repeated hyperthermiahad comparable retention time 24 hours after IA train-
Fig 3. Frequently repetitive febrile seizures (FRFSs) in early life did not cause aberrant mossy fiber sprouting in the hippocampus atadulthood as demonstrated by Timm stain. Representative photomicrographs in the CA3 region showed no Timm granules in thestratum pyramidale (sp) or stratum oriens (so; a Timm score of 0) in one rat (A), and prominent granules occurring in SP (aTimm score of 3) in another (B). In the dentate gyrus (C, D), occasional granules (arrow) in the suprapyramidal (E) and infrapy-ramidal blade (G) of the granular cell layer (GCL; a Timm score of 1) was noted in one rat, and prominent Timm-stained fibers(arrows) crossing the suprapyramidal (F) and infrapyramidal (H) blade of GCL with a light band of near-continuous granules (ar-rowheads) in the GCL-inner molecular layer (IML) boundary (a Timm score of 3) was seen in another. Quantitative analysis ofmossy fiber sprouting into the CA3 region (I) or dentate gyrus (J) in the FRFS group did not differ from that of controls, singlefebrile seizure (SFS), repetitive febrile seizure (RFS) groups (n � 7 for each group). Scale bars � 100�m (A, B), 200�m (C, D),or 20�m (E–H).
Chang et al: Memory in Febrile Seizures 711

ing with controls (see Fig 4F). Again, there were nodifferences in the pain response to foot shock betweenthe three groups.
Hippocampal cAMP Response-element BindingPhosphorylation after Inhibitory Avoidance LearningWas Impaired in the Frequently Repetitive FebrileSeizure Group at AdulthoodSeveral studies have reported that a circumscribed andpersistent CREB phosphorylation (lasting up to 20hours) occurs in the hippocampus after IA train-ing.24,30–32 We investigated whether CREB phosphor-ylation in the hippocampus after learning was relatedto the LTM deficits induced by early-life FRFS. West-ern blots of hippocampal extracts showed comparablebasal levels of total CREB and pCREB in the control
and FS groups (Fig 5A). Three hours after IA training,however, significantly increased pCREB levels, com-pared with those in naive controls, were found in allbut the FRFS group. The pCREB levels in trainedFRFS group were significantly lower than in trainedcontrols (p � 0.01) (see Fig 5B, C). There were nodifferences in total CREB expression between thegroups. To exclude the effects of exposure to the ap-paratus or shock stress, we measured hippocampalpCREB levels in the unpaired shock control. Nochanges in pCREB levels, compared with those in na-ive controls, were observed. Furthermore, there was asignificantly positive correlation between the fold in-crease in hippocampal pCREB levels and retentiontime 3 hours after IA training (see Fig 5D, p � 0.01).
We next examined the spatial profiles of pCREB im-
Fig 4. Frequently repetitive febrile seizures (FRFS) in early life caused hippocampal-dependent long-term memory deficits at adult-hood. In the Morris water maze, the control, single febrile seizure (SFS), repetitive febrile seizures (RFS), and FRFS groups alllearned to find the submerged platform during the training sessions (p � 0.001); however, the FRFS group was significantly slowerin finding the platform than each of the three remaining groups (* p � 0.05) (A). On the probe test, the platform was removedand the memory for the platform position was measured. There was a significant group–quadrant interaction on time distributionin the various quadrants (p � 0.05), and the FRFS group spent significantly less time in the target quadrant than did each of thethree remaining groups (* p � 0.05, n � 22 to 26 for each condition) (B). In contrast, all four groups acquired a similar perfor-mance on the visually cued version of the water maze (C). In the inhibitory avoidance task, retention time was assessed 1, 3, or 24hours after training. At 1 hour after training, control and FS groups showed similar retention time; whereas at 3 hours and espe-cially at 24 hours after training, only the FRFS group showed significantly shorter retention time than controls (*p � 0.01 byDunn’s multiple comparison test after Kruskal–Wallis analyses of variance, n � 18–22 for each condition) (D). There were nosignificant differences between the four groups in the pain response of flinch, vocalization, or locomotion to foot shock (E). In con-trast with the performance of the FRFS group, the rats with frequently repeated hyperthermia had comparable retention time 24hours after inhibitory avoidance training with controls (n � 15–18 for each group; F).
712 Annals of Neurology Vol 54 No 6 December 2003

munoreactivity in the hippocampus after IA learning.There was barely visible pCREB immunoreactivitythroughout the hippocampus of naive adult controlrats, comparable to the findings by others.33,34 Those
unpaired shock controls had low pCREB immunoreac-tivity in the hippocampus, suggesting that the shock orstress per se had few effects on CREB phosphorylationin the hippocampus, in line with our Western blot re-sults. Three hours after IA training, however, there wasprominent pCREB immunoreactivity in the CA1, den-tate gyrus especially the infrapyramidal blade, and tosome extent, CA3 of the controls, SFS and RFSgroups, which was in accordance with the findings byothers.24,31,32 In contrast, the hippocampal pCREBimmunoreactivity was significantly less in the trainedFRFS group than that in the other trained animals(Figs 6 and 7). Thus, decreased pCREB immunoreac-tivity in the hippocampus after IA learning was closelycorrelated with the memory consolidation deficits inrats with early-life FRFS.
To determine whether the decreased pCREB levelscould be attributed to decreased synaptogenesis afterFRFS, we examined the expression of a synapticmarker, synaptophysin, in the hippocampus. Therewere no differences in the synaptophysin level betweenthe groups (see Fig 5A).
Rolipram Administration Rescued Long-term MemoryDeficits in the Frequently Repetitive Febrile SeizureGroup by Enhancing cAMP Response-elementBinding PhosphorylationStudies have shown that the cAMP cascade leading toCREB phosphorylation can be pharmacologically up-regulated in vivo by administration of rolipram, an in-hibitor of phosphodiesterase type IV.35,36 We next ex-amined whether activation of the cAMP–CREBsignaling pathway with rolipram could rescue the LTMdeficits in the FRFS group. Twenty-four hours after IA
Š Fig 5. CAMP response-element binding (CREB) phosphoryla-tion in the hippocampus after inhibitory avoidance (IA) learn-ing was impaired in the frequently repetitive febrile seizures(FRFSs) group at adulthood. Representative Western blotsshowed no differences in the basal levels of pCREB and totalCREB, or synaptophysin among controls, single febrile seizure(SFS), repetitive febrile seizures (RFS), and FRFS groups(n � 6 in each group) (A). Western blots demonstrated thehippocampal pCREB and their respective total CREB levelsfrom naive (N) and unpaired shock control (S), and fromcontrol, SFS, RFS and FRFS groups 3 hours after IA training(B). The corresponding densitometry showed significantly in-creased pCREB levels, compared with those in naive controls,in all the trained animals but the FRFS group (n � 7 ineach group). The pCREB levels in the trained FRFS groupwere significantly lower than in trained controls (*p � 0.01)(C). The total CREB levels were similar in the control andFS groups. There was a significantly positive correlation be-tween the fold increase of hippocampal pCREB levels and re-tention time 3 hours after IA training (r2 � 0.33, p �0.01) (D).
Chang et al: Memory in Febrile Seizures 713

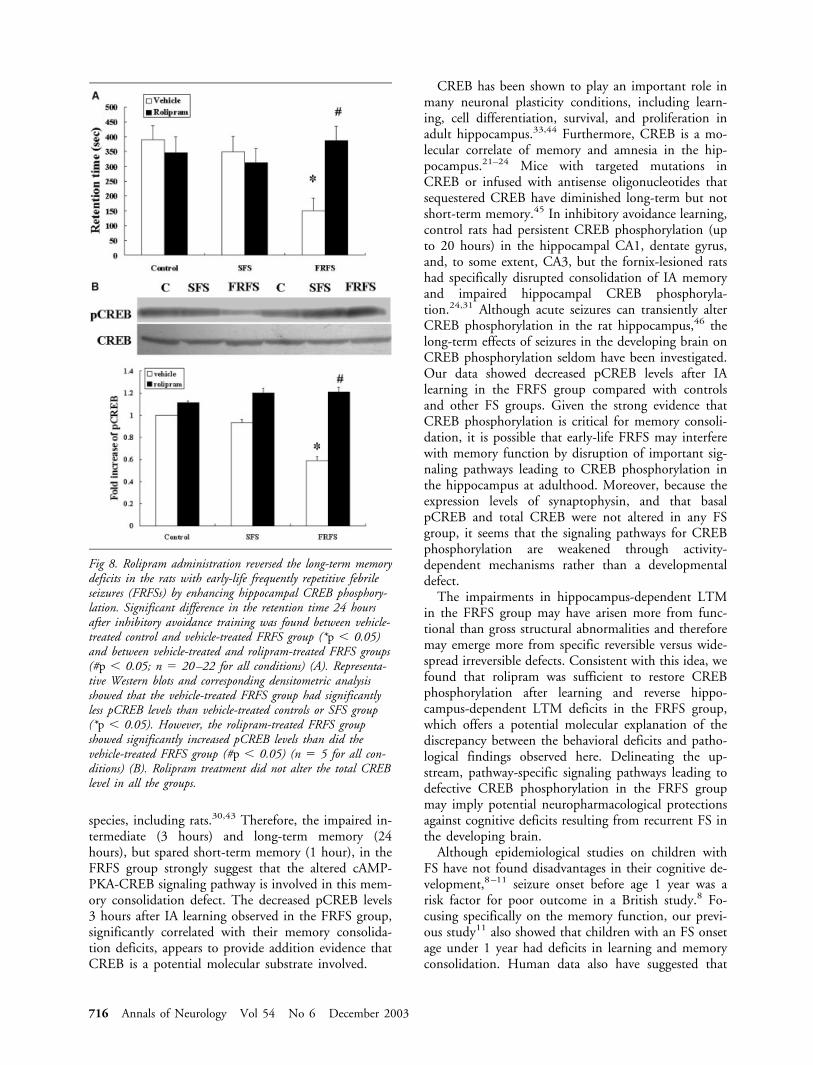
learning, the rolipram-treated FRFS group showed sig-nificantly longer retention time than the vehicle-treatedFRFS group (p � 0.05, Fig 8A). There was no signif-icant difference in the retention time between any ofthe rolipram-treated groups. There was also no signif-icant difference in the performance between rolipram-and vehicle-treated rats in either the control or SFSgroup. Thus, rolipram did not influence the memoryperformance in controls or SFS group, but rolipramreversed the LTM deficits of the FRFS group.
The effects of rolipram on the pCREB levels in thehippocampus 3 hours after IA training were examinedby Western immunoblotting. In vehicle-treated ani-mals, the FRFS group showed significantly less pCREBexpression than the control or SFS group (p � 0.05).However, the rolipram-treated FRFS group showedsignificantly increased pCREB expression than did thevehicle-treated FRFS group (p � 0.05; see Fig 8B).There was no difference in the pCREB levels betweenany of the rolipram-treated groups. Treatment with ro-lipram did not alter the total CREB level in any ofthese groups.
DiscussionThis study sought to answer whether brief and recur-rent FSs are detrimental to the developing brain and, if
they are, what the underlying mechanisms are. Ourmajor findings were that FRFS in immature rats didnot (1) cause acute neuronal damage or apoptosis, (2)result in chronic neuronal loss or aberrant mossy fiberssprouting in the hippocampus, or (3) alter seizure sus-ceptibility to the chemical convulsant (PTZ) at adult-hood, but that they did (4) impair hippocampus-dependent long-term, but not short-term, memory,and (5) decrease activity-dependent CREB phosphory-lation at Ser133 in the hippocampus after learningtasks. These findings were not observed in rats that hadexperienced early-life SFS or RFS. In addition, rolip-ram, an agent that enhances the cAMP–CREB signal-ing pathway, can rescue the LTM deficits observed inFRFS group by enhancing CREB phosphorylation inthe hippocampus. These data demonstrate that early-life FRFS caused functional impairment rather thansignificant morphological changes in the hippocampusof adult rats, and that alteration of cAMP signalingcascades in regulating CREB phosphorylation may un-derlie the cellular mechanism for the impaired neuro-nal plasticity and memory function in these rats.
Adult rats with early-life FRFS, in contrast with ratsin the control, SFS, and RFS groups, had LTM deficitsas assessed by Morris water maze and IA tasks. Thefrequency of FS in early life did not alter the sensori-
Fig 6. Frequently repetitive febrile seizures (FRFSs) in early life resulted in decreased pCREB immunoreactivity in the hippocampusafter inhibitory avoidance (IA) training at adulthood. The negative control brain slice without first antibody (A) showed nopCREB immunoreactivity throughout the hippocampus, whereas the naive control (B) had barely visible pCREB and the unpairedshock control animals (C) had low levels of pCREB immunoreactivity in the hippocampus. Three hours after IA training, there wasprominent immunoreactivity in the CA1 and dentate gyrus especially the infrapyramidal blade and, to some extent CA3 of the con-trol (D), single febrile seizure (E), and repetitive febrile seizure groups (F). In contrast, hippocampal pCREB immunoreactivity wassignificantly less in the FRFS group than that in the other trained animals (G) (n � 4 per group). Scale bar � 200�m.
714 Annals of Neurology Vol 54 No 6 December 2003

motor or motivational function in adult rats, becausethe visual-motor performance in the visually cued wa-ter maze and the pain responses in the IA training weresimilar between control and FS groups. These resultsclearly point to LTM deficits in the FRFS group. Theabsence of neuronal loss and mossy fiber sprouting inthe hippocampi of FRFS-group rats suggests that long-term alterations in neuronal plasticity and intracellularsignaling transduction were probably sufficient to ac-count for the significant changes in the memory defi-cits observed. Memory impairment of mature brainsexposed to seizures has been well documented andseems to relate to marked neuronal death and mossyfiber sprouting.37 Seizures in early life, however, do notproduce selective hippocampal damage but can havepermanent morphological and functional effects.38 Inrat pups, seizures after global hypoxia, hot water hyper-thermic seizures, flurothyl-induced seizures, or hip-pocampal tetanus toxin injections resulted in decreasedseizure thresholds and aberrant mossy fiber sprouting,despite no apparent neuronal injury.38–40 In contrastwith the previous rat pup seizure models, we found nomorphological changes in adult rats with early-life
FRFS, despite the magnitude of their memory deficit.The alteration in the activity-dependent neuronal plas-ticity observed in our study implies that the importantconsequences of early-life seizures are not limited tohistopathological alterations.
One prominent component of hippocampal memorydeficits in the FRFS group may be related to defects inthe ability to consolidate short-term memory to LTM.Studies have identified multiple phases of memory thatare distinguishable by both temporal dynamics and dis-tinct molecular requirements.41 Short-term memoryformation (which persists for minutes to hours) doesnot require transcription and translation. Intermediate-term memory (which persists for �24 hours) requiresprotein but not RNA synthesis, whereas LTM (whichlasts for days) requires both.41,42 Furthermore,intermediate-term memory requires the persistent acti-vation of PKA-CREB cascade, suggesting its potentialrole as a gate for LTM consolidation.41 In avoidancelearning paradigm, memory consolidation requires asecond time window of protein synthesis at 3 to 7hours after training, which parallels the time course ofthe increased PKA activity and pCREB levels in several
Fig 7. Frequently repetitive febrile seizures (FRFSs) in early life resulted in decreased pCREB immunoreactivity throughout the hip-pocampus after inhibitory avoidance (IA) training at adulthood. Higher magnification of Fig 6 in the CA1 (A–F), infrapyramidalblade (G–L) and suprapyramidal blade (M–R) of dentate gyrus, and CA3 (S–X) areas showed prominently enhanced pCREB im-munoreactivity in the trained control (C, I, O, U), single febrile seizure (D, J, P, V), and repetitive febrile seizure groups (E, K,Q, W), compared with that of the naive controls (A, G, M, S) or unpaired shock controls (B, H, N, T). In contrast, the trainedFRFS group had significantly less pCREB immunoreactivity (F, L, R, X) than the other trained animals throughout the hippocam-pal subregions (n�4 per group). Scale bars � 25�m (A–R) or 50�m (S–X).
Chang et al: Memory in Febrile Seizures 715
species, including rats.30,43 Therefore, the impaired in-termediate (3 hours) and long-term memory (24hours), but spared short-term memory (1 hour), in theFRFS group strongly suggest that the altered cAMP-PKA-CREB signaling pathway is involved in this mem-ory consolidation defect. The decreased pCREB levels3 hours after IA learning observed in the FRFS group,significantly correlated with their memory consolida-tion deficits, appears to provide addition evidence thatCREB is a potential molecular substrate involved.
CREB has been shown to play an important role inmany neuronal plasticity conditions, including learn-ing, cell differentiation, survival, and proliferation inadult hippocampus.33,44 Furthermore, CREB is a mo-lecular correlate of memory and amnesia in the hip-pocampus.21–24 Mice with targeted mutations inCREB or infused with antisense oligonucleotides thatsequestered CREB have diminished long-term but notshort-term memory.45 In inhibitory avoidance learning,control rats had persistent CREB phosphorylation (upto 20 hours) in the hippocampal CA1, dentate gyrus,and, to some extent, CA3, but the fornix-lesioned ratshad specifically disrupted consolidation of IA memoryand impaired hippocampal CREB phosphoryla-tion.24,31 Although acute seizures can transiently alterCREB phosphorylation in the rat hippocampus,46 thelong-term effects of seizures in the developing brain onCREB phosphorylation seldom have been investigated.Our data showed decreased pCREB levels after IAlearning in the FRFS group compared with controlsand other FS groups. Given the strong evidence thatCREB phosphorylation is critical for memory consoli-dation, it is possible that early-life FRFS may interferewith memory function by disruption of important sig-naling pathways leading to CREB phosphorylation inthe hippocampus at adulthood. Moreover, because theexpression levels of synaptophysin, and that basalpCREB and total CREB were not altered in any FSgroup, it seems that the signaling pathways for CREBphosphorylation are weakened through activity-dependent mechanisms rather than a developmentaldefect.
The impairments in hippocampus-dependent LTMin the FRFS group may have arisen more from func-tional than gross structural abnormalities and thereforemay emerge more from specific reversible versus wide-spread irreversible defects. Consistent with this idea, wefound that rolipram was sufficient to restore CREBphosphorylation after learning and reverse hippo-campus-dependent LTM deficits in the FRFS group,which offers a potential molecular explanation of thediscrepancy between the behavioral deficits and patho-logical findings observed here. Delineating the up-stream, pathway-specific signaling pathways leading todefective CREB phosphorylation in the FRFS groupmay imply potential neuropharmacological protectionsagainst cognitive deficits resulting from recurrent FS inthe developing brain.
Although epidemiological studies on children withFS have not found disadvantages in their cognitive de-velopment,8–11 seizure onset before age 1 year was arisk factor for poor outcome in a British study.8 Fo-cusing specifically on the memory function, our previ-ous study11 also showed that children with an FS onsetage under 1 year had deficits in learning and memoryconsolidation. Human data also have suggested that
Fig 8. Rolipram administration reversed the long-term memorydeficits in the rats with early-life frequently repetitive febrileseizures (FRFSs) by enhancing hippocampal CREB phosphory-lation. Significant difference in the retention time 24 hoursafter inhibitory avoidance training was found between vehicle-treated control and vehicle-treated FRFS group (*p � 0.05)and between vehicle-treated and rolipram-treated FRFS groups(#p � 0.05; n � 20–22 for all conditions) (A). Representa-tive Western blots and corresponding densitometric analysisshowed that the vehicle-treated FRFS group had significantlyless pCREB levels than vehicle-treated controls or SFS group(*p � 0.05). However, the rolipram-treated FRFS groupshowed significantly increased pCREB levels than did thevehicle-treated FRFS group (#p � 0.05) (n � 5 for all con-ditions) (B). Rolipram treatment did not alter the total CREBlevel in all the groups.
716 Annals of Neurology Vol 54 No 6 December 2003

hippocampal injury can occur in infants with repetitiveor prolonged FS.47,48 The current experimental FSmodel, which used rat pups during a brain-development age equivalent to that of the young in-fants,14,49 provides further evidence that FRFS ratherthan SFS at this vulnerable age can cause long-termcognitive deficits due to impaired activity-dependentneuronal plasticity in the hippocampus.
We found that brief repetitive FS did not cause al-teration in the seizure threshold at the acute stage or inPTZ-induced generalized seizure susceptibility at adult-hood. In contrast, Dube and colleagues16 found thatprolonged FS in immature rats caused increased limbicseizure susceptibility to kainate. The discrepancy be-tween these studies may be caused by the differences inthe seizure models, seizure duration, hyperthermicmethods, or different convulsant agents used.
Our study demonstrates the benign behavioral andmorphological outcome after brief SFS in rat pups, andthat long-term alteration in neuronal plasticity andmemory function without significant neuronal loss inthe hippocampus can occur after early-life FRFS. Ex-trapolation of a rodent experiment to humans is diffi-cult, and how many seizures would be required to elicitthese changes in humans is not known. Furthermore,childhood FS is induced by endogenous factors result-ing in an elevated temperature, whereas the experimen-tal hyperthermic seizures are induced by exogenoussources of heat. This study, however, raises concernsabout the long-term cognitive instead of epileptic con-sequences of even brief FRFS during early brain devel-opment.
This work was supported by grants from the National Health Re-search Institute (NHRI-EX91, 92-9131NN, C.C.H.), the ChangGung Memorial Hospital Research Program (CMRP 1104, Y.C.C.),and National Science Council (NSC 90-2314-B-182A-022,Y.C.C.).
We thank Dr L. Yu for his critical comments on the manuscript,and Dr L. T. Huang for technical support in the Timm stainingand Morris water maze.
References1. Shinnar S. Febrile seizures. In: Swaiman KF, Ashwal S, eds.
Pediatric neurology–principles and practice. 3rd ed. St. Louis:Mosby, 1999:676–682.
2. Huang CC, Wang ST, Chang YC, et al. Risk factors for a firstfebrile convulsion in children: a population study in southernTaiwan. Epilepsia 1999;40:719–725.
3. MacDonald BK, Johnson AL, Sander JW, Shorvon SD. Febrileconvulsions in 220 children—neurological sequelae at 12 yearsfollow-up. Eur Neurol 1999;41:179–186.
4. Annegers JF, Hauser WA, Shirts SB, Kurland LT. Factors prog-nostic of unprovoked seizures after febrile convulsions. N EnglJ Med 1987;316:493–498.
5. Berg AT, Shinnar S. Do seizures beget seizures? An assessmentof the clinical evidence in humans. J Clin Neurophysiol 1997;14:102–110.
6. Falconer MA, Taylor DC. Surgical treatment of drug-resistantepilepsy due to mesial temporal sclerosis. Etiology and signifi-cance. Arch Neurol 1968;19:353–361.
7. Shinnar S. Prolonged febrile seizures and mesial temporal scle-rosis. Ann Neurol 1998;43:411–412.
8. Verity CM, Greenwood R, Golding J. Long-term intellectualand behavioral outcomes of children with febrile convulsions.N Engl J Med 1998;338:1723–1728.
9. Ellenberg JH, Nelson KB. Febrile seizures and later intellectualperformance. Arch Neurol 1978;35:17–21.
10. Chang YC, Guo NW, Huang CC, et al. Neurocognitive atten-tion and behavior outcome of school-age children with a historyof febrile convulsions: a population study. Epilepsia 2000;41:412–420.
11. Chang YC, Guo NW, Wang ST, et al. Working memory ofschool-aged children with a history of febrile convulsions: apopulation study. Neurology 2001;57:37–42.
12. Wallace SJ, Cull AM. Long-term psychological outlook forchildren whose first fit occurs with fever. Dev Med Child Neu-rol 1979;21:28–40.
13. Baram TZ, Gerth A, Schultz L. Febrile seizures: an appropriate-aged model suitable for long-term studies. Dev Brain Res 1997;98:265–270.
14. Toth Z, Yan XX, Haftoglou S, et al. Seizure-induced neuronalinjury: vulnerability to febrile seizures in an immature ratmodel. J Neurosci 1998;18:4285–4294.
15. Chen K, Baram TZ, Soltesz I. Febrile seizures in the developingbrain result in persistent modification of neuronal excitability inlimbic circuits. Nat Med 1999;5:888–894.
16. Dube C, Chen K, Eghbal-Ahmadi M, et al. Prolonged febrileseizures in the immature rat model enhance hippocampal excit-ability long term. Ann Neurol 2000;47:336–344.
17. Kolfen W, Pehle K, Konig S. Is the long-term outcome of chil-dren following febrile convulsions favorable? Dev Med ChildNeurol 1998;40:667–671.
18. Holmes GL, Ben-Ari Y. Seizures in the developing brain: per-haps not so benign after all. Neuron 1998;21:1231–1234.
19. Holmes GL, Gairsa JL, Chevassus-Au-Louis N, Ben-Ari Y.Consequences of neonatal seizures in the rat: morphologicaland behavioral effects. Ann Neurol 1998;44:845–857.
20. Liu Z, Yang Y, Silveira DC, et al. Consequences of recurrentseizures during early brain development. Neuroscience 1999;92:1443–1454.
21. Kandel ER. The molecular biology of memory storage: a dia-logue between genes and synapses. Science 2001;294:1030–1038.
22. Weeber EJ, Sweatt JD. Molecular neurobiology of human cog-nition. Neuron 2002;33:845–848.
23. Lonze BE, Ginty DD. Function and regulation of CREB familytranscription factors in the nervous system. Neuron 2002;35:605–623.
24. Taubenfeld SM, Wiig KA, Bear MF, Alberini CM. A molecularcorrelate of memory and amnesia in the hippocampus. NatNeurosci 1999;2:309–310.
25. Huang CY, Fujimura M, Noshita N, et al. SOD1 down-regulates NF-kappaB amd c-Myc expression in mice after tran-sient focal cerebral ischemia. J Cereb Blood Flow Metab 2001;21:163–173.
26. West MJ, Gundersen HJ. Unbiased stereological estimation ofthe number of neurons in the human hippocampus. J CompNeurol 1990;296:1–22.
27. Brunson KL, Eghbal-Ahmadi M, Bender R, et al. Long-termprogressive hippocampal cell loss and dysfunction induced byearly-life administration of corticotropin-releasing hormone re-produce the effects of early-life stress. Proc Natl Acad Sci USA2001;98:8856–8861.
Chang et al: Memory in Febrile Seizures 717

28. Huang AM, Wang HL, Tang YP, Lee EH. Expression ofintegrin-associated protein gene associated with memory forma-tion in rats. J Neurosci 1998;18:4305–4313.
29. Ginty DD, Kornhauser JM, Thompson MA, et al. Regulationof CREB phosphorylation in the suprachiasmatic nucleus bylight and a circadian clock. Science 1993;260:238–241.
30. Bernabeu R, Bevilaqua L, Ardenghi P, et al. Involvement ofhippocampal cAMP/cAMP-dependent protein kinase signalingpathway in a late memory consolidation phase of aversively mo-tivated learning in rats. Proc Natl Acad Sci USA 1997;94:7041–7046.
31. Taubenfeld SM, Wiig KA, Monti B, et al. Fornix-dependentinduction of hippocampal CCAAT enhancer-binding proteinbeta and delta co-localize with phosphorylated cAMP responseelement-binding protein and accompanies long-term memoryconsolidation. J Neurosci 2001;21:84–91.
32. Impey S, Smith DM, Obrietan K, et al. Stimulation of cAMPresponse element (CRE)-mediated transcription during contex-ture learning. Nat Neurosci 1998;1:595–601.
33. Bender RA, Lauterborn JC, Gall CM, et al. Enhanced CREBphosphorylation in immature dentate gyrus granule cells pre-cedes neurotrophin expression and indicates a specific role ofCREB in granule cell differentiation. Eur J Neurosci 2001;13:679–686.
34. Thome J, Sakai N, Shin KH, et al. cAMP response element-mediated gene transcription is upregulated by chronic antide-pressant treatment. J Neurosci 2000;20:4030–4036.
35. Barad M, Bourtchouladze R, Winder DG, et al. Rolipram, atype IV-specific phosphodiesterase inhibitor, facilitates the es-tablishment of long-lasting long-term potentiation and im-proves memory. Proc Natl Acad Sci USA 1998;95:15020–15025.
36. Conti M, Jin SL. The molecular biology of cyclic nucleotidephosphodiesterases. Prog Nucleic Acid Res Mol Biol 1999;63:1–38.
37. Crusio WE, Schwegler H, Lipp HP. Radial-maze performanceand structural variation of the hippocampus in mice: a correla-tion with mossy fibre distribution. Brain Res 1987;425:182–185.
38. Jensen FE, Holmes GL, Lombroso CT. Age-dependent changesin long-term seizure susceptibility and behavior after hypoxia inrats. Epilepsia 1992;33:971–980.
39. Anderson AE, Hrachovy RA, Swann JW. Increased susceptibil-ity to tetanus toxin-induced seizures in immature rats. EpilepsyRes 1997;26:433–442.
40. Jiang W, Duong TM, de Lanerolle NC. The neuropathology ofhyperthermic seizures in the rat. Epilepsia 1999;40:5–19.
41. Sutton MA, Masters SE, Bagnall MW, Carew TJ. Molecularmechanisms underlying a unique intermediate phase of memoryin Aplysia. Neuron 2001;31:143–154.
42. Tully T, Preat T, Boynton SC, Del Vecchio M. Genetic dis-section of consolidated memory in Drosophila. Cell 1994;79:35–47.
43. Bourtchouladze R, Abel T. Berman N, et al. Different trainingprocedures recruit either one or two critical periods for contex-tual memory consolidation, each of which requires protein syn-thesis and PKA. Learn Mem 1998;5:365–374.
44. Nakagawa S, Kim JE, Lee R, et al. Localization of phosopho-rylated cAMP response element-binding protein in immatureneurons of adult hippocampus. J Neurosci 2002;22:9868–9876.
45. Guzowski JF, McGaugh JL. Antisense oligodeoxynucleotide-mediated disruption of hippocampal cAMP response element-binding protein levels impairs consolidation of memory for wa-ter maze training. Proc Natl Acad Sci USA 1997;94:2693–2698.
46. Moore AN, Waxham MN, Dash PK. Neuronal activity in-creases the phosphorylation of the transcription factor cAMPresponse element-binding protein (CREB) in rat hippocampusand cortex. J Biol Chem 1996;271:14214–14220.
47. Perez ER, Maeder P, Villemure KM, et al. Acquired hippocam-pal damage after temporal lobe seizures in 2 infants. Ann Neu-rol 2000;48:384–387.
48. VanLandingham KE, Heinz ER, Cavazos JE, Lewis DV. Mag-netic resonance imaging evidence of hippocampal injury afterprolonged focal febrile convulsions. Ann Neurol 1998;43:413–426.
49. Avishai-Eliner S, Brunson KL, Sandman CA, Baram TZ.Stressed-out, or in (utero)? Trends Neurosci 2002;25:518–524.
718 Annals of Neurology Vol 54 No 6 December 2003
Copyright © 2022 FDOKUMEN




















