
Mediators involved in the febrile response induced by Tityus serrulatus scorpion venom in rats
11
Toxicon 48 (2006) 556–566 Mediators involved in the febrile response induced by Tityus serrulatus scorpion venom in rats Andre´a C. Pessini a , Danielle R. Santos a , Eliane C. Arantes b , Glo´ria E.P. Souza a, a Laborato´rio de Farmacologia, Faculdade de Cincias Farmaceˆuticas de Ribeira˜o Preto, USP, Av. do Cafe´, s/n Campus USP, 14040-903 Ribeira˜o Preto, SP, Brasil b Fı´sico-Quı´mica, Depto. de Fı´sica e Quı´mica, Faculdade de Cieˆncias Farmaceˆuticas de Ribeira˜o Preto, USP, 14040-903 Ribeira˜o Preto, SP, Brasil Received 25 October 2005; received in revised form 24 June 2006; accepted 3 July 2006 Available online 7 July 2006 Abstract Tityus serrulatus venom (Tsv) was intraperitoneally (ip) injected at doses of 75, 150 and 300 mg/kg and IL-1b (2.0 mg/kg) was given intravenously (iv) to male Wistar rats. Rectal temperature was measured by radiotelemetry. Vagotomy was performed according to Bluthe et al. [1994. Lipopolysaccharide induces sickness behaviour in rats by a vagal mediated mechanism. C R Acad. Sci. 317(6), 499–503]. Cerebrospinal fluid (CSF) and peritoneal fluid (PF) levels of bradykinin (BK) were measured by ELISA. B 1 (des-Arg 9 -[Leu 8 ]-BK; DALBK) and B 2 kinin receptor (icatibant) antagonists (1.0 mg/kg each), the induced nitric oxide synthase inhibitor aminoguanidine (50.0 mg/kg), the neuronal nitric oxide synthase inhibitor 7-nitroindazole (30.0 mg/kg), the dual cyclooxygenase inhibitor ibuprofen (10.0 mg/kg), the selective interleukin-1 receptor antagonist IL-ra (2.0 mg/kg) and dipyrone (120 mg/kg) were given ip. Celecoxib (5 mg/kg) was given per os (po). Tsv at doses of 75 mg/kg evoked no change in rectal temperature while at doses of 150 and 300 mg/kg it promoted long-lasting fever (2 1C70.1). Tsv (150 mg/kg) increased by nearly 3 and 5 times, respectively BK concentration in the CSF and in the PF. Subdiaphragmatic vagotomy or 7-nitroindazole reduced, icatibant, DALBK, IL-1ra, aminoguanidine and dipyrone abolished, while ibuprofen and celecoxib failed to affect Tsv-induced fever. These results suggest that PGs do not play a relevant role, whereas, kinins via their B 1 and B 2 receptors, IL-1, nitric oxide and vagal neurotransmission are involved in Tsv-induced fever. r 2006 Elsevier Ltd. All rights reserved. Keywords: Fever; Tityus serrulatus venom; BK; NO; IL-1; Vagus nerve 1. Introduction Fever, an important clinical sign of the acute phase response to pathogens, is mediated by several endogenous pyrogens from host cells, among which interleukin (IL)-1a, IL-1b, IL-6, IL-8, tumour necrosis factor-a (TNF-a; for a review see Roth and Souza, 2001), bradykinin (BK; Coelho et al., 1997), endothelins (Fabricio et al., 1998, 2005a) and nitric oxide (NO; Roth et al., 1999). While the precise mechanism by which these pyrogens activate the hypothalamus to induce fever remains unclear (Zeisberger, 1999), the current model of fever points towards induction of prostaglandins ARTICLE IN PRESS www.elsevier.com/locate/toxicon 0041-0101/$ - see front matter r 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.toxicon.2006.07.006 Corresponding author. Tel.: +55 16 3602 4184; fax: +55 16 3602 4880. E-mail address: [email protected] (G.E.P. Souza).
Transcript of Mediators involved in the febrile response induced by Tityus serrulatus scorpion venom in rats

ARTICLE IN PRESS
0041-0101/$ - se
doi:10.1016/j.to
�Correspondifax: +5516 360
E-mail addre
Toxicon 48 (2006) 556–566
www.elsevier.com/locate/toxicon
Mediators involved in the febrile response inducedby Tityus serrulatus scorpion venom in rats
Andrea C. Pessinia, Danielle R. Santosa, Eliane C. Arantesb, Gloria E.P. Souzaa,�
aLaboratorio de Farmacologia, Faculdade de Cincias Farmaceuticas de Ribeirao Preto, USP, Av. do Cafe, s/n Campus USP,
14040-903 Ribeirao Preto, SP, BrasilbFısico-Quımica, Depto. de Fısica e Quımica, Faculdade de Ciencias Farmaceuticas de Ribeirao Preto, USP,
14040-903 Ribeirao Preto, SP, Brasil
Received 25 October 2005; received in revised form 24 June 2006; accepted 3 July 2006
Available online 7 July 2006
Abstract
Tityus serrulatus venom (Tsv) was intraperitoneally (ip) injected at doses of 75, 150 and 300 mg/kg and IL-1b (2.0 mg/kg)was given intravenously (iv) to male Wistar rats. Rectal temperature was measured by radiotelemetry. Vagotomy was
performed according to Bluthe et al. [1994. Lipopolysaccharide induces sickness behaviour in rats by a vagal mediated
mechanism. C R Acad. Sci. 317(6), 499–503]. Cerebrospinal fluid (CSF) and peritoneal fluid (PF) levels of bradykinin (BK)
were measured by ELISA. B1 (des-Arg9-[Leu8]-BK; DALBK) and B2 kinin receptor (icatibant) antagonists (1.0mg/kg
each), the induced nitric oxide synthase inhibitor aminoguanidine (50.0mg/kg), the neuronal nitric oxide synthase inhibitor
7-nitroindazole (30.0mg/kg), the dual cyclooxygenase inhibitor ibuprofen (10.0mg/kg), the selective interleukin-1 receptor
antagonist IL-ra (2.0mg/kg) and dipyrone (120mg/kg) were given ip. Celecoxib (5mg/kg) was given per os (po). Tsv at
doses of 75 mg/kg evoked no change in rectal temperature while at doses of 150 and 300 mg/kg it promoted long-lasting
fever (2 1C70.1). Tsv (150 mg/kg) increased by nearly 3 and 5 times, respectively BK concentration in the CSF and in the
PF. Subdiaphragmatic vagotomy or 7-nitroindazole reduced, icatibant, DALBK, IL-1ra, aminoguanidine and dipyrone
abolished, while ibuprofen and celecoxib failed to affect Tsv-induced fever. These results suggest that PGs do not play a
relevant role, whereas, kinins via their B1 and B2 receptors, IL-1, nitric oxide and vagal neurotransmission are involved in
Tsv-induced fever.
r 2006 Elsevier Ltd. All rights reserved.
Keywords: Fever; Tityus serrulatus venom; BK; NO; IL-1; Vagus nerve
1. Introduction
Fever, an important clinical sign of the acutephase response to pathogens, is mediated by severalendogenous pyrogens from host cells, among which
e front matter r 2006 Elsevier Ltd. All rights reserved
xicon.2006.07.006
ng author. Tel.: +55 16 3602 4184;
2 4880.
ss: [email protected] (G.E.P. Souza).
interleukin (IL)-1a, IL-1b, IL-6, IL-8, tumournecrosis factor-a (TNF-a; for a review see Rothand Souza, 2001), bradykinin (BK; Coelho et al.,1997), endothelins (Fabricio et al., 1998, 2005a) andnitric oxide (NO; Roth et al., 1999).
While the precise mechanism by which thesepyrogens activate the hypothalamus to induce feverremains unclear (Zeisberger, 1999), the current modelof fever points towards induction of prostaglandins
.

ARTICLE IN PRESSA.C. Pessini et al. / Toxicon 48 (2006) 556–566 557
(PGs, namely PGE2 and PGF2a) as a commonpathway leading to the rise of the hypothalamicthermoregulatory set point (Morimoto et al., 1988).
Tityus serrulatus scorpion is considered one of themost dangerous species for humans in Brazil,responsible for many clinical cases of envenomationin the southern region of the country. It containsseveral basic proteins of low molecular weightwith neurotoxic activity (for a review see Becerrilet al., 1997). Injection of scorpion venom intoexperimental animals produces systemic signsand symptoms similar to those observed followinghuman envenomation such as hyperthermia, psy-chomotor agitation, salivation, lachrymation, in-creased gastrointestinal mobility, cardiac andrespiratory arrhythmias, arterial hypertension fol-lowed by hypotension, cardiac failure, pulmonaryoedema and shock (Freire-Maia and Campos,1989). Hyperthermia was reported to occur in about50% of the patients stung by the scorpion Centrur-
oides margaritatus (Marinkelle and Stahnke, 1965),and constitutes an important sign in victims of thescorpions Androctomus australis and Buthus occita-
nus (Goyffon et al., 1982). Fatal hyperthermiaoccurred in rabbits following intraventricular injec-tion of venom from the Leiurus quinquestriatus
scorpion (Osman et al., 1973). Four fatal casesfollowing L. quinquestriatus sting in children werereported by Ismail et al. (1990). Three victims hadrectal temperatures over 40 1C, while the fourth washypothermic. The hyperthermia was successfullytreated by acetaminophen, but all of them devel-oped hypothermia 48 h after the sting.
The involvement of the kinin system followingenvenomation has been reported in several species.Heterometrus bengalensis scorpion venom activatesthe plasma kinin system in rabbits (Lahiri and NagChaudhuri, 1983) and L. quinquestriatus venomincreases the plasma level of BK (Fatani et al., 1998)in the same species. Moreover, Ismail et al. (1992)observed that aprotinin, a kallikrein-kinin inhibitor,protected rabbits and rats from the cardiac effects ofthe L. quinquestriatus venom such as bradycardiaand rhythm and conduction defects.
Fukuhara et al. (2004) has suggested that thekinin system may participate in the pathogenesis ofhuman Tityus serrulatus envenomation. An increaseof kinins could culminate in neuropeptide releasefrom sensory fibers (Geppetti, 1993), synthesis ofcytokines (Tiffany and Burch, 1989; Paegelow et al.,1995a, b; Pan et al., 1996; Pang and Knox, 1998), ofPG, leukotrienes (O’Neill and Lewis, 1989; Saka-
moto et al., 1994) and NO (Bogle et al., 1991; Volpeet al., 1996). In fact, after Tityus serrulatus venom(Tsv) envenomation an increased plasma levelof IL-1, IL-6 and TNF-a was found in humansand rats (Magalhaes et al., 1999; Pessini et al.,2003) and an elevation in serum level of NOwas observed in mice (Barros et al., 1998; Petrice-vich et al., 2000; Petricevich and Pena, 2002;Tambourgi et al., 1998).
Recent findings suggest that peripheral afferentnerves, especially those contained in the vagusnerve, play an important signalling role betweeninflammatory sites and the brain. Among otherresponses to intraperitoneal (ip) injection of eitherLPS or IL-1b, sub-diaphragmatic vagotomy blocksfever (Watkins et al., 1995; Goldbach et al., 1997;Werner et al., 2003), central induction of IL-1bmRNA (Laye et al., 1995), hyperalgesia andpain (Watkins et al., 1995), and reduces residentcell numbers and influx of neutrophils (Werneret al., 2003).
Using conscious rats, the present study wasundertaken aiming to: (1) assess whether theincrease in body temperature caused by ip Tsvinjection constitutes solely a hyperthermic action ora true integrated fever; (2) investigate the contribu-tion of kinins, IL-1, NO, PGs and vagus nerve tothis response; and (3) establish a possible relation-ship between Tsv-induced fever and changes incerebrospinal and peritoneal fluid BK levels.
2. Material and methods
2.1. Animals
Experiments were conducted using male Wistarrats weighing 180–200 g, housed at 2471 1C under a12:12 h light-dark cycle (lights on at 06:00 AM) withfree access to food and tap water. Each animal,including the controls, was utilised for experimentalprocedures only once. The experiments were carriedout in accordance with the current guidelines for thecare of laboratory animals and ethical guidelines forinvestigations in conscious animals set by theEthical Committee of University of Sao Paulo,Campus of Ribeirao Preto, SP, protocol number03.1.1055.53.9.
2.2. Scorpion venom and drugs
Desiccated Tityus serrulatus scorpion venom waspurchased from Phoneutria Biotechnology and

ARTICLE IN PRESSA.C. Pessini et al. / Toxicon 48 (2006) 556–566558
Services (Belo Horizonte, Brazil) and kept at �20 1Cuntil use. The following drugs were employed:icatibant [D-Arg{Hyp3Thi5-DTic7-Oic8}BK] anddipyrone were kindly donated by Aventis, Ger-many. DALBK [des-Arg9-Leu8]-BK, iNOS selectiveinhibitor aminoguanidine (AG), nNOS selectiveinhibitor 7-nitroindazole (7-NI) and ibuprofen werefrom Sigma Chemical Co. (St Louis, MO, USA);recombinant human interleukin-1 receptor antago-nist (IL-1ra, Synergen, Bouder CO); murine IL-1b(R&D Systems, Inc., Minneapolis, USA) andcelecoxib (Celebras, Pharmacia-Pfizer, Sao Paulo,Brazil). All drugs were dissolved in pyrogen-freesterile saline, except 7-NI and celecoxib which weredissolved in a solution of sesame oil/dimethylsulfoxide (9:1) and water, respectively. Proteaseinhibitors used were aprotinin (Sigma); SBTI/Tripsin Inhibitor (Sigma); Captopril (Sigma);1,10—Phenanthroliniumchloride (Merck); EDTA(Merck) dissolved in PBS, pH 8.0.
2.3. Subdiaphragmatic vagotomy
Animals were anaesthetised with sodium pento-barbital (40mg/kg, ip). The peritoneal cavity wasexposed and the ventral and dorsal vagal trunkswere removed just below the diaphragm, togetherwith 2 cm of the tissue between the gastric arteryand the oesophagus. Sham-operated animals weresubmitted to the same procedure without handlingthe nerves. Animals received antibiotic treatment(oxytetracycline hydrochloride, 400mg/kg, sc) priorto surgery, and 2 days after the procedure. Tominimise weight loss, animals received a comple-mentary liquid caloric meal (13.3% protein, 70.5%carbohydrates, 2.6% fat, and 4.7% fibres). Experi-ments were conducted 15 days after surgeryand animals with abnormal patterns of body weightgain were excluded from the study (Bluthe et al.,1994).
2.4. Temperature measurements
By inserting a thermistor probe (no. 402, YellowSprings Instruments Co. Inc., OH, USA) 4 cm intothe animal’s rectum body temperature was mea-sured for 1min every 30min for 6 h. During eachtemperature measurement, the animal was pickedup gently and held manually, but was not removedfrom its home cage. This procedure was performedat least twice on the day before the experiment tominimise changes secondary to handling. On the
day of the experiment, the basal temperature of eachanimal was determined four times at 30min inter-vals prior to any injection. Only animals displayingmean basal rectal temperatures between 36.8 and37.2 1C were selected for the study. Tail skintemperature was measured by using a thermistorprobe attached with adhesive tape to the lateralsurface of the proximal segment of the tail (2 cmfrom its base), as originally proposed by Young andDawson (1982). The mean basal value of tail skintemperature ranged between 31 and 33 1C. Allexperiments were conducted between 08:00 and17:00 h in a temperature-controlled room (2871 1C;Gordon et al., 2002).
2.5. Harvesting of cerebrospinal and peritoneal fluid
For the determination of the BK content in thecerebrospinal fluid (CSF), 2 h after Tsv or salineinjections, the animals were anaesthetised withsodium pentobarbital (40mg/kg, ip) and a singlesample of CSF was collected from each animalaccording to the method described by Consiglio andLucion (2000). Briefly, each rat was fixed to thestereotaxic apparatus, with its body flexed down-ward, and the skin covering the base of the skull andback of the neck was trichotomised and moistenedwith a cotton swab soaked in ethanol to reveal asmall depression between the occipital protuberanceand the atlas. A scalp cannula connected to asyringe was then inserted through the depressioninto the cisterna magna, to collect 60–100 mlsamples. Samples contaminated with blood werediscarded. For evaluation of BK level in theperitoneal fluid, 10ml of PBS, pH 7.4 (composition,in mmol/l: NaCl 137.8, KCl 27, NaH2PO4 14,NaHPO4 81) containing 5U/ml of heparin wasinjected via a needle placed near the sternum 2 hafter the ip Tsv injection. The abdominal cavity wasgently massaged for 1min and 5ml of the peritonealfluid were collected by a needle inserted into theinguinal region (Souza et al., 1988). To preventformation or breakdown of kinins (Bond et al.,1997) the CSF and peritoneal fluid samples wereplaced into eppendorf tubing and plastic tubes,respectively, containing protease inhibitors [aproti-nin (0.5mg/ml); SBTI/Tripsin Inhibitor (4mg/ml);Captopril (10mM); 1,10—Phenanthroliniumchlor-ide (6mM); EDTA (60mM)], and maintained in adark box over ice until centrifugation (1300g, during15min), and immediately frozen to �70 1C untilanalysed.

ARTICLE IN PRESSA.C. Pessini et al. / Toxicon 48 (2006) 556–566 559
2.6. Determination of BK concentrations
The BK concentration in the CSF and peritonealfluid was determined by the quantitative sandwichenzyme immunoassay technique (ELISA) using theMARKIT-M BKskits (Dainippon PharmaceuticalCo., Ltd, Osaka, Japan) following the manufac-turer’s directions. Detection limits of the assay were490 pg/ml and intra- and inter-assay coefficients ofvariation were o10%. The analysis gave noevidence of cross-reactions with any protein. Peri-toneal fluid ðn ¼ 6Þ and CSF ðn ¼ 6Þ were harvested2.0 h following ip injection of 150 mg/kg of Tsv or ofsterile saline ðn ¼ 6Þ.
2.7. Experimental protocols
Tsv was dissolved in sterile saline, centrifuged at11269g for 5min, and filtered through sterilisingmembranes (cellulose ester filters: 0.22mm, Milli-pore). Protein concentration in the supernatant wasdetermined by the microbiuret method (Itzhaki andGill, 1964). To determine the adequate dose, Tsv wasadministered intraperitoneally (500ml) to rats at 75,150 and 300mg/kg and compared to control animalsthat received sterile saline. Other animals receivedip injections of either B1 (Des-Arg9-[Leu]8-BK,DALBK, 1mg/kg), or B2 receptor antagonist (icati-bant, 1mg/kg), iNOS inhibitor (AG, 50mg/kg),nNOS inhibitor (7-NI, 30mg/kg), IL-1 receptorantagonist (IL-1ra, 2mg/kg), ibuprofen (10mg/kg),dipyrone (120mg/kg) or po injection of celecoxib(5mg/kg) 30min prior to Tsv injection (150mg/kg,ip). IL-1b was given intravenously (iv) at a dose of(2.0mg/kg). Control animals were similarly treatedwith the appropriate vehicles (saline, sesame oil/dimethyl sulfoxide (9:1) or water). The dosesof all substances employed were established inprevious studies (Fabricio et al., 1998, 2005b;Cartmell et al., 1999; Souza et al., 2002; Rothet al., 1999). Pyrogenic stimuli were always injectedbetween 10:00 and 11:00 AM.
2.8. Statistical analysis
For data analysis, the baseline temperature prior toany injection was determined for each animal and allsubsequent temperatures expressed as changes fromthis averaged value (DT). The heat loss index (HLI)was used to evaluate thermoeffector responses of tailskin vasculature. The HLI was calculated accordingto the formula: HLI ¼ ðT sk � TaÞ � ðTc � TaÞ
�1
where Tsk refers to skin tail temperature, Ta toambient temperature and Tc to core body tempera-ture. The values of HLI varied between 0 and 1.0,representing respectively, conditions of maximalvasoconstriction and maximal vasodilatation (Roma-novsky et al., 2002).
Values are reported as means7standard error ofthe mean (SEM). The levels of BK were analysed byone-way ANOVA followed by Tukey’s test. DT andHLI responses were compared across treatmentsand time points by a two-way ANOVA for repeatedmeasures, using a SPSSs statistical software (SPSSInc., Chicago, IL, USA). Differences were consid-ered significant when Po0:05.
3. Results
3.1. Temperature changes induced by Tsv
Animals treated with only the vehicle or with75 mg/kg of Tsv showed no significant variations inrectal temperature over baseline values for up to 6 hafter administration. In sharp contrast, ip injectionsof 150 or 300 mg/kg of Tsv resulted in an increase inrectal temperature. At a dose of 150 mg/kg, Tsvincreased body temperature from 1 h, peaked after1.5 h by nearly 2 1C, and decreased thereafter,reaching basal values after approximately 4.5 h.The increase in body temperature caused by 300 mg/kg Tsv was similar to that evoked by 150 mg/kg, ofapproximately 2 1C; however, this increase startedalready 0.5 h following administration, peaked after1.5 h, and started to decrease after 2 h (Fig. 1A). Thedose of 150 mg/kg was selected for the remainingexperiments because no statistical differences be-tween the effects of this dose, and the dose of300 mg/kg were observed at any point. The increasein rectal temperature induced by 150 mg/kg of Tsvwas preceded by a significant decrease in skin tailtemperature of almost �2 1C, between 0.5 and 1.5 h,indicating vasoconstriction of the local vascular bed(Fig. 1B).
3.2. Effect of vagotomy on fever induced by Tsv
Fig. 2 shows that vagotomized or sham-operatedanimals did not show significant basal temperaturechanges after saline administration. In contrastto the sham operated, the vagotomized animalsshowed a significantly reduced fever responseto Tsv.

ARTICLE IN PRESS
Fig. 1. Time course of changes in rectal or skin tail temperatures
and heat loss index (HLI) of rats following ip injection of 500mlof Tsv. (A) represents the change in rectal temperature (DT, 1C)
of rats induced by different doses of Tsv (n ¼ 75, � ¼ 150 and
& ¼ 300mg/kg). (B) and (C) represent, respectively, the changes
in tail skin temperature (DTsk, 1C) and heat loss index (HLI) of
rats induced by 150mg/kg of Tsv. Control (6–7) animals received
an equivalent volume of saline (’ ¼ sal). (*) and (#), indicate
significant differences between fever responses of Tsv injected,
and control rats ðpo0:05Þ.
Fig. 2. Effect of subdiaphragmatic vagotomy (VGX) on fever
evoked by 150mg/kg of Tsv. Vagotomized and SHAM animals
(see methods) received ip saline or Tsv injections. Values
represent means7SEM of changes in rectal temperature (DT,
1C). (*) indicates significant difference between fever responses of
VGX and SHAM operated rats following Tsv injection,
ðpo0:05Þ.
A.C. Pessini et al. / Toxicon 48 (2006) 556–566560
3.3. Involvement of kinins on fever induced by Tsv
Compared to control animals, ip injection of Tsv(150 mg/kg) increased BK concentrations in the
cerebrospinal fluid nearly threefold (from56.777.5 to 163.2738.4 pg/ml) and in the perito-neal fluid nearly fivefold (from 53.479.6 to289.4755.0 pg/ml). Ip treatment with the B2 or B1
BK receptor antagonists, icatibant or DALBK,respectively (1.0mg/kg each), did not significantlymodify the basal temperature of saline-treated rats,but abolished the fever induced by Tsv (Fig. 3A).
3.4. Effects of IL-1ra on fever induced by Tsv
Ip injection of IL-1b caused increases in bodytemperature (0.9670.18 1C, 3 h) and pre-treatmentwith IL-1ra (2.0mg/kg) abolished this effect(0.0370.03 1C, 3 h), but did not significantly modifythe basal temperature of saline-treated rats (datanot shown).
Ip treatment with IL-1ra (2.0mg/kg) also abol-ished the fever evoked by Tsv (Fig. 3B).
3.5. Effects of NO synthase inhibitors on fever
induced by Tsv
In the next series of experiments, AG, a selectiveinhibitor of inducible NOS enzyme (50mg/kg, ip),or 7-NI, a selective inhibitor of neuronal NOSenzyme (30mg/kg, ip) were tested for their abilityto alter Tsv-induced fever. AG fully abolished(Fig. 4A), and 7-NI significantly reduced thisresponse (Fig. 4B). Neither ip treatment with either
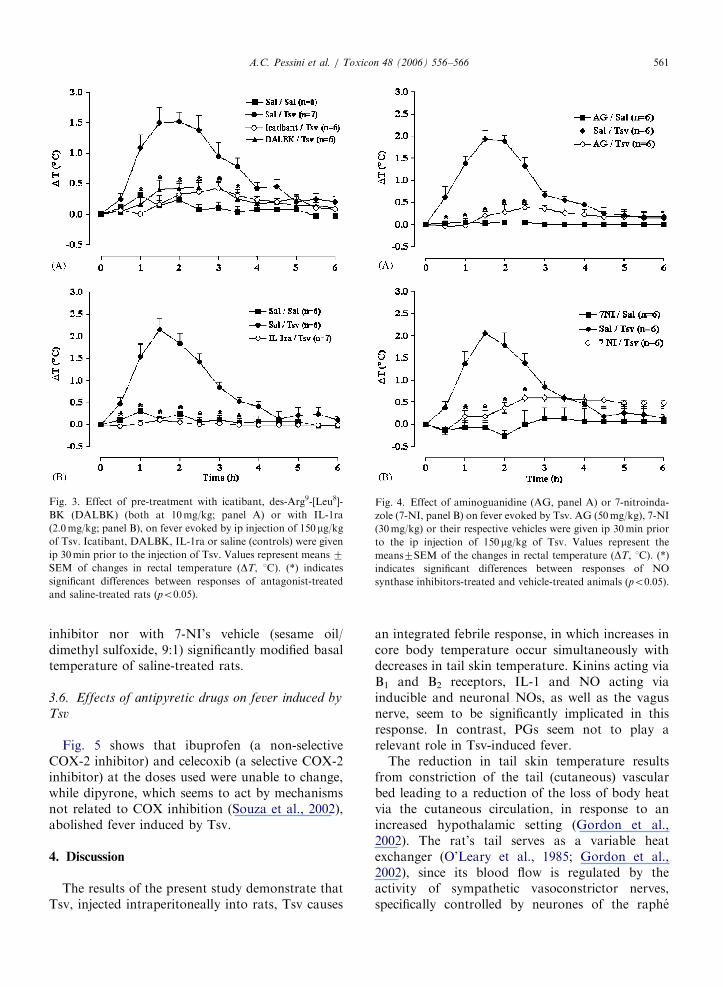
ARTICLE IN PRESS
Fig. 3. Effect of pre-treatment with icatibant, des-Arg9-[Leu8]-
BK (DALBK) (both at 10mg/kg; panel A) or with IL-1ra
(2.0mg/kg; panel B), on fever evoked by ip injection of 150mg/kgof Tsv. Icatibant, DALBK, IL-1ra or saline (controls) were given
ip 30min prior to the injection of Tsv. Values represent means 7SEM of changes in rectal temperature (DT, 1C). (*) indicates
significant differences between responses of antagonist-treated
and saline-treated rats (po0:05).
Fig. 4. Effect of aminoguanidine (AG, panel A) or 7-nitroinda-
zole (7-NI, panel B) on fever evoked by Tsv. AG (50mg/kg), 7-NI
(30mg/kg) or their respective vehicles were given ip 30min prior
to the ip injection of 150mg/kg of Tsv. Values represent the
means7SEM of the changes in rectal temperature (DT, 1C). (*)
indicates significant differences between responses of NO
synthase inhibitors-treated and vehicle-treated animals (po0:05).
A.C. Pessini et al. / Toxicon 48 (2006) 556–566 561
inhibitor nor with 7-NI’s vehicle (sesame oil/dimethyl sulfoxide, 9:1) significantly modified basaltemperature of saline-treated rats.
3.6. Effects of antipyretic drugs on fever induced by
Tsv
Fig. 5 shows that ibuprofen (a non-selectiveCOX-2 inhibitor) and celecoxib (a selective COX-2inhibitor) at the doses used were unable to change,while dipyrone, which seems to act by mechanismsnot related to COX inhibition (Souza et al., 2002),abolished fever induced by Tsv.
4. Discussion
The results of the present study demonstrate thatTsv, injected intraperitoneally into rats, Tsv causes
an integrated febrile response, in which increases incore body temperature occur simultaneously withdecreases in tail skin temperature. Kinins acting viaB1 and B2 receptors, IL-1 and NO acting viainducible and neuronal NOs, as well as the vagusnerve, seem to be significantly implicated in thisresponse. In contrast, PGs seem not to play arelevant role in Tsv-induced fever.
The reduction in tail skin temperature resultsfrom constriction of the tail (cutaneous) vascularbed leading to a reduction of the loss of body heatvia the cutaneous circulation, in response to anincreased hypothalamic setting (Gordon et al.,2002). The rat’s tail serves as a variable heatexchanger (O’Leary et al., 1985; Gordon et al.,2002), since its blood flow is regulated by theactivity of sympathetic vasoconstrictor nerves,specifically controlled by neurones of the raphe

ARTICLE IN PRESS
Fig. 5. Effect of ibuprofen (IBU, panel A), celecoxib (CELE,
panel B) and dipyrone, (DIP, panel C) on fever evoked by Tsv.
IBU (10mg/kg), CELE (5mg/kg) or DIP (120mg/kg) or their
vehicles were given ip 30min prior to the ip injection of 150mg/kgof Tsv. Values represent the means7SEM of the changes in rectal
temperatures (DT, 1C). (*) indicates significant differences
between responses of antipyretic-treated, and vehicle-treated
animals, ðpo0:05Þ.
A.C. Pessini et al. / Toxicon 48 (2006) 556–566562
magnus/pallidum nucleus (Blessing and Nalivaiko,2001). Thus, it seems that by stimulating vasocon-striction of cutaneous vessels, intraperitoneallyadministered Tsv evokes a concerted thermoregula-
tory response in which the reduced efficacy of thetail skin to dissipate body heat results in fever andnot hyperthermia in which elevation of coretemperature is accompanied by tail skin vasodilata-tion (Briese, 1998). Changes in body blood pressuredo not seem to be responsible for the vasoconstric-tion and decrease in tail skin temperature caused byTsv in rats. Previous findings from our laboratoryshow that after iv administration of 100 mg/kg Tsv,the hypertensive effect started 2.5–3.5min afterinjection, and gradually decreased after 30min(Vasconcelos et al., 2005) a timing at which the tailskin temperature begins to decrease. The bloodpressure returned to normal after 90min a timeperiod at which the decrease in tail skin temperatureis still low.
Over the last few years, some studies have shownthat the information generated by peripheral stimulimight reach the central nervous system via periph-eral sensory nerves, including certain fibres of thevagus nerve (for a review see Roth and Souza,2001). Regarding the febrile response, it is wellestablished that subdiaphragmatic vagotomy blocksfever to ip injections of LPS or IL-1 (Watkins et al.,1995; Goldbach et al., 1997; Simons et al., 1998;Werner et al., 2003). In the present study we showedthat the development of fever due to ip injection ofTsv was attenuated by subdiaphragmatic vagotomy,indicating an important role of the vagus nerve inthe cascade of events leading to this fever.
The present study also showed that ip Tsvinjection increased BK concentration in CSF andperitoneal fluid suggesting an involvement of thekinin system in this response. Confirming thishypothesis we observed that the kininogen-kininreceptor antagonists, icatibant and DALBK, abol-ished Tsv-induced fever. Similar actions were alsoobserved in rat paw oedema induced by the venomof the Apis melifera bee (Calixto et al., 2003).
Cytokines also seem to be involved in thesystemic and local inflammatory response duringenvenomation caused by Tsv (Magalhaes et al.,1999; Pessini et al., 2003). We have previously found(Pessini et al., 2003), that Tsv was able to induce acharacteristic systemic and local inflammatoryreaction revealed by high serum levels of IL-1a,IL-6 and TNF-a and increased cell migration tothe peritoneal cavity. In the present study, theefficacy of ip treatment with IL-1ra on Tsv-inducedfever suggests a crucial involvement of IL-1 in thisresponse. Supporting this suggestion is the recog-nised role of this cytokine in the fever response to

ARTICLE IN PRESSA.C. Pessini et al. / Toxicon 48 (2006) 556–566 563
different exogenous pyrogenic stimuli (Netea et al.,2000), and the ability of BK, via B1 and B2
receptors, to stimulate a variety of cell types tosynthesise and release IL-1 and others cytokines(Paegelow et al., 1995b; Koyama et al., 1995; Satoet al., 2000; Santos et al., 2003), that can reach thethermoregulatory centre triggering critical mechan-isms for fever development (Strijibos et al., 1992;Blatteis and Sehic, 1998; Roth and Souza, 2001).Furthermore, the ability of IL-1 to induce B1
receptor expression has been well-established(Campos et al., 1999, 2002). Thus, it appearslikely that IL-1 is implicated in B1 expression inthe fever response to Tsv as suggested by Calixtoet al. (2003) for the paw oedema induced by venomof A. melifera bee. Blockade of B2 receptor inhibitsB1 expression (Phagoo et al., 1999); therefore, theantipyretic effect of icatibant on Tsv-induced fevermight also result in the inhibition of B1 expression.Since the Tsv venom caused a fever of quickeffervescence and slow deffervescence, it is possiblethat B2 activation, IL-1 release, and B1 expression/activation occur subsequently within a short timeperiod resulting in a similar fever blockade by therespective receptor antagonists. The constitutivepresence of functionally active BK B2 receptorson rat vagal afferent neurons (Krstew et al., 1998),and the constitutive presence of binding sitesfor IL-1 ligands on glomus cells located withinabdominal vagus nerve-associated paraganglia(Goeheler et al., 1997), may also contributeimportantly to the transmission of signals generatedin the periphery to the central nervous system(Watkins et al., 1995).
BK-potentiating peptides (peptide T) have beenfound in scorpion venom (Araujo and Gomes, 1976;Meki et al., 1995). It is therefore plausible thatpeptide T strengthens kinin effects during the feverto Tsv.
Cytokines and kinins are known to stimulate NOand PGs synthases (Bogle et al., 1991; O’Neill andLewis, 1989), that seem to participate in tempera-ture regulation and fever response to bacterialproducts for instance, LPS (Gerstberger, 1999,Roth et al., 1999; Morimoto et al., 1988). IncreasedNO plasma levels have also been described inenvenomed mice (Barros et al., 1998; Petricevichet al., 2000; Petricevich and Pena, 2002; Tambourgiet al., 1998) and high NO2—serum levels were alsoobserved after Tsv injection (Petricevich and Pena,2002). We showed here that ip treatment withinhibitors for iNOS (AG) or nNOS (7-NI), respec-
tively abolished and drastically reduced the febrileresponse induced by Tsv; this suggests that NO actsas a pyretic agent in this response. Considering thediversity of the distribution of various NOSs indifferent tissues, the role of these tissues in feverdevelopment and the autocrine nature of the actionof NO, it is conceivable that the sum of these effectscan culminate in favour of fever production. Forinstance, iNOS has been shown to be widespreadthroughout the body and is present in peripheralcells and tissues directly engaged in the generationof metabolic heat (De Luca et al., 1995), release ofpyrogenic mediators (Geller et al., 1994), and inbrain centres responsible for fever (Minano et al.,1997). Expression of constitutive nNOS, althoughalso demonstrated in the kidney, occurs mainly innervous tissues, including hypothalamic preopticnuclei (Azad et al., 2001).
PGs have been considered key mediators of feverin response to exogenous and endogenous pyrogens(Kluger, 1991); an increased level of PGE2 wasfound in the cerebrospinal fluid of rats after ipinjection of LPS (Fabricio et al., 2005b) and in thepre-optic area of the anterior hypothalamus (thecentre of temperature regulation) of cats challengedwith endotoxin or IL-1 (Sirko et al., 1989).
Unexpectedly, ibuprofen, a non-selective COX-1/-2(Tegeder et al., 2001) and celecoxib, a selective COX-2inhibitor (Botting, 2004) did not affect the feverresponse to Tsv, thus suggesting that PGs are notinvolved in it. In contrast, dipyrone a classicalantipyretic and analgesic drug (Souza et al., 2002;Levy and Carpenter, 1995) blocked fever due to Tsv.The reason to investigate the effect of dipyrone onTsv-induced fever in rats comes from the fact that thisdrug is clinically used in humans to treat the increasein body temperature on ongoing envenomation afterstinging by T. serrulatus (Freire-Maia and Campos,1989). Furthermore, in our hands dipyrone did notblock cyclooxygenase-2 (the enzyme responsible forthe production of PG during fever), but abolishedfever induced by prostaglandin F2a showing that thispotent antipyretic agent does not act by inhibitingprostaglandin synthesis (Souza et al., 2002). It has alsobeen shown that dipyrone produces peripheral, spinaland supraspinal antinociception when assessed inglutamate-induced hyperalgesia in mice (Beirith et al.,1998). Moreover, Nakamura et al., (2004) showedthat neurons expressing the vesicular glutamatetransporter 3 (VGLUT3) are distributed in ratmedullary raphe regions. VGLUT3-expressing me-dullary raphe neurons innervate rat interscapular

ARTICLE IN PRESSA.C. Pessini et al. / Toxicon 48 (2006) 556–566564
brown adipose tissue (BAT) and tail throughsympathetic preganglionic neurons of specific thoracicsegments. Furthermore, microinjection of glutamateinto the intermediolateral cell column of BATproduced thermogenesis in this tissue showing theinvolvement of glutamate in thermoregulatory func-tions, including fever (Nakamura et al., 2004). It isthus possible that dipyrone can reduce fever byblocking this glutamatergic neurotransmission as itdoes in hyperalgesia.
Ismail et al. (1990, 1995) showed the effectivenessof acetaminophen in treating L. quinquestriatus-induced hyperthermia in children and suggested theinvolvement of PGs in this response, a suggestionsupported by Kis et al. (2004) who showed thatacetaminophen blocks the synthesis of PGs inisolated brain microvessels by inhibiting COX-2.However, in our hands (data not shown) acetami-nophen given ip (75mg/kg) did not significantlychange fever caused by Tsv in rats (2 h: Tsv+vehicle ¼ 1.970.3 1C;Tsv+acetaminophen ¼ 1.470.3 1C). The lack ofeffectiveness of this agent may be related to animalspecies (rats versus human), route of administrationof the venom (ip versus rectal) and differences in thecomposition of the venoms from T. serrulatus andL. quinquestriatus. Moreover, as discussed above,fever induced by Tsv was not altered by ibuprofenand celecoxib indicating non-involvement of PGsin this response. Therefore, it may be expectedthat acetaminophen that acts by blocking COX-2(Kis et al., 2004) does not modify the fever responseto Tsv.
In conclusion, the present study shows for thefirst time, that Tsv promotes an integrated febrileresponse, that seems to depend on kinins (via B1 andB2 receptors), IL-1, NO and vagal neurotransmis-sion. Confirming its clinical efficacy, dipyroneblocked the fever to Tsv by mechanisms not relatedto inhibition of PGs synthesis, since ibuprofen andcelecoxib failed to alter this fever. The efficacy of thefever blockers used here could be helpful in treatingother signals, for instance pain from Tsv envenoma-tion, whose cast of mediators is essentially the sameas that for fever production.
Acknowledgement
We are grateful to Fundac- ao de Amparo asPesquisas do Estado de Sao Paulo (FAPESP) andCNPq for financial support.
References
Araujo, R.L., Gomes, M.V., 1976. Potentiation of bradykinin
action on smooth muscle by a scorpion venom extract. Gen.
Pharmacol. 7 (2–3), 123–126.
Azad, S.C., Marsicano, G., Eberlein, I., Putzke, J., Zieglgans-
berger, W., Spanagel, R., Lutz, B., 2001. Differential role of
the nitric oxide pathway on delta(9)-THC-induced central
nervous system effects in the mouse. Eur. J. Neurosci. 13 (3),
561–568.
Barros, S.F., Friedlanskaia, I., Petricevich, V.L., Kipnis, T.L.,
1998. Local inflammation, lethality and cytokine release in
mice injected with Bothrops atrox venom. Mediators Inflamm.
7 (5), 339–346.
Becerril, B., Marangoni, S., Possani, L.D., 1997. Toxins and
genes isolated from scorpions of the genus Tityus. Toxicon 35
(6), 821–835.
Beirith, A., Santos, A.R., Rodriguez, A.L., Creczynski-Pasa,
T.B., Calixto, J.B., 1998. Spinal and supraspinal antinocicep-
tive action of dipyrone in formalin, capsaicin and glutamate
tests. Study of the mechanism of action. Eur. J. Pharmacol.
345 (3), 233–245.
Blatteis, C.M., Sehic, E., 1998. Cytokines and fever. Ann. N. Y.
Acad. Sci. 840 (1), 608–618.
Blessing, W.W., Nalivaiko, E., 2001. Raphe magnus/pallidus
neurons regulate tail but not mesenteric arterial blood flow in
rats. Neuroscience 105 (4), 923–929.
Bluthe, R.M., Walter, V., Parnet, P., Laye, S., Lestage, J.,
Verrier, D., Poole, S., Stenning, B.E., Kelley, K.W., Dantzer,
R., 1994. Lipopolysaccharide induces sickness behaviour in
rats by a vagal mediated mechanism. C. R. Acad. Sci. 317 (6),
499–503.
Bogle, R.G., Coade, S.B., Moncada, S., Pearson, J.D., Mann,
G.E., 1991. Bradykinin and ATP stimulate L-arginine up-take
and nitric oxide release in vascular endothelial cells. Biochem.
Biophys. Res. Commun. 180 (2), 926–932.
Bond, A.P., Lemon, M., Dieppe, P.A., Bhoola, K.D., 1997.
Generation of kinins in synovial fluid from patients with
arthropathy. Immunopharmacology 36 (2–3), 209–216.
Botting, R., 2004. Antipyretic therapy. Front. Biosci. 9 (1),
956–966.
Briese, E., 1998. Normal body temperature of rats: the setpoint
controversy. Neurosci. Biobehav. Rev. 22 (3), 427–436.
Calixto, M.C., Triches, K.M., Calixto, J.B., 2003. Analysis of the
inflammatory response in the rat paw caused by the venom of
Apis melifera bee. Inflamm. Res. 52 (3), 132–139.
Campos, M.M., Souza, G.E., Calixto, J.B., 1999. In vivo B1
kinin-receptor upregulation. Evidence for involvement of
protein kinases and nuclear factor kappaB pathways. Br. J.
Pharmacol. 127 (8), 1851–1859.
Campos, M.M., Souza, G.E., Ricci, N.D., Pesquero, J.L.,
Teixeira, M.M., Calixto, J.B., 2002. The role of migrating
leukocytes in IL beta-induced up-regulation of kinin B1
receptor in rats. Br. J. Pharmacol. 135 (5), 1107–1114.
Cartmell, T., Luheshi, G.N., Rothwell, N.J., 1999. Brain sites of
action of endogenous interleukin-1 in the febrile response to
localized inflammation in the rat. J. Physiol. 518 (2), 585–594.
Coelho, M.M., Oliveira, C.R., Pajolla, G.P., Calixto, J.B., Pela,
I.R., 1997. Central involvement of kinin B1 and B2 receptors
in the febrile response induced by endotoxin in rats. Br. J.
Pharmacol. 121 (2), 296–302.

ARTICLE IN PRESSA.C. Pessini et al. / Toxicon 48 (2006) 556–566 565
Consiglio, A.R., Lucion, A.B., 2000. Technique for collecting
cerebrospinal fluid in the cisterna magna of non-anesthetized
rats. Brain Res. Brain Res. Protoc. 5 (1), 109–114.
De Luca, B., Monda, M., Sullo, A., 1995. Changes in eating
behavior and thermogenic activity following inhibition of
nitric oxide formation. Am. J. Physiol. 268 (6 Pt 2),
R1533–R1538.
Fabricio, A.S., Silva, C.A., Rae, G.A., D’Orleans-Juste, P.,
Souza, G.E., 1998. Essential role for endothelin ET(B)
receptors in fever induced by LPS (E. coli) in rats. Br. J.
Pharmacol. 125 (3), 542–548.
Fabricio, A.S., Rae, G.A., D’Orleans-Juste, P., Souza, G.E.,
2005a. Endothelin-1 as a central mediator of LPS-induced
fever in rats. Brain Res. 20 (1–2), 92–100.
Fabricio, A.S., Veiga, F.H., Cristofoletti, R., Navarra, P., Souza,
G.E., 2005b. The effects of selective and nonselective
cyclooxygenase inhibitors on endothelin-1-induced fever in
rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 288 (3),
R671–R677.
Fatani, A.J., Furman, B.L., Zeitlin, I.J., 1998. The involvement
of plasma kinins in the cardiovascular effects of Leiurus
quinquestriatus scorpion venom in anaesthetised rabbits.
Toxicon 36 (3), 523–536.
Freire-Maia, L., Campos, J.A., 1989. Pathophysiology and
treatment of scorpion poisoning. In: Ownby, C.L., Odell,
G.V. (Eds.), Natural Toxins: Characterization Pharmacology
and Therapeutics. Pergamon Press, Oxford, pp. 139–159.
Fukuhara, Y.D., Dellalibera-Joviliano, R., Cunha, F.Q., Reis,
M.L., Donadi, E.A., 2004. The kinin system in the
envenomation caused by the Tityus serrulatus scorpion sting.
Toxicol. Appl. Pharmacol. 196 (3), 390–395.
Geller, D.A., Freeswick, P.D., Nguyen, D., Nussler, A.K., Di
Silvio, M., Shapiro, R.A., Wang, S.C., Simmons, R.L.,
Billiar, T.R., 1994. Differential induction of nitric oxide
synthase in hepatocytes during endotoxemia and the acute-
phase response. Arch. Surg. 129, 165–171.
Gerstberger, R., 1999. Nitric oxide and body temperature
control. News Physiol. Sci. 14, 30–36.
Geppetti, P., 1993. Sensory neuropeptide release by bradykinin:
mechanisms and pathophysiological implications. Regul.
Peptides 47 (1), 1–23.
Goehler, L.E., Relton, J.K., Dripps, D., Kiechle, R., Tartaglia,
N., Maier, S.F., Watkins, L.R., 1997. Vagal paraganglia bind
biotinylated interleukin-1 receptor antagonist: a possible
mechanism for immune-to-brain communication. Brain Res.
Bull. 43 (3), 357–364.
Goldbach, J.M., Roth, J., Storr, B., Zeisberger, E., 1997.
Changes of abdominal temperature and circulating levels of
cortisol and interleukin-6 in response to intra-arterial infu-
sions of tumor necrosis factor-alpha or tumor necrosis factor-
beta in guinea pigs. Eur. J. Pharmacol. 334 (2–3), 249–254.
Gordon, C.J., Puckett, E., Padnos, B., 2002. Rat tail skin
temperature monitored noninvasively by radiotelemetry:
characterization by examination of vasomotor responses to
thermomodulatory agents. J. Pharmacol. Toxicol. Methods
47 (2), 107–114.
Goyffon, M., Vachon, M., Broglio, N., 1982. Epidemiological
and clinical characteristics of the scorpion envenomation in
Tunisia. Toxicon 20 (1), 337–344.
Ismail, M., Abd-Elsalam, M.A., Morad, A.M., 1990. Do changes
in body temperature following envenomation by the scorpion
Leiurus quinquestriatus influence the course of toxicity?
Toxicon 28 (11), 1265–1284.
Ismail, M., Fatani, A.J., Dabees, T.T., 1992. Experimental
treatment protocols for scorpion envenomation: a review of
common therapies and an effect of kallikrein-kinin inhibitors.
Toxicon 30 (10), 1257–1279.
Ismail, M., 1995. The scorpion envenoming syndrome. Toxicon
33 (7), 825–858.
Itzhaki, R.F., Gill, D.M., 1964. A microbiuret method for
estimating proteins. Anal. Biochem. 9, 401–410.
Kis, B., Snipes, A., Bari, F., Busija, D.W., 2004. Regional
distribution of cyclooxygenase-3 mRNA in the rat central
nervous system. Mol. Brain Res. 126, 78–80.
Kluger, M.J., 1991. Fever: role of pyrogens and cryogens.
Physiol. Rev. 71 (1), 93–127.
Koyama, S., Rennard, S.I., Robbins, R.A., 1995. Bradykinin
stimulates bronchial epithelial cells to release neutrophil and
monocyte chemotactic activity. Am. J. Physiol. 269 (1),
L38–L44.
Krstew, E., Jarrott, B., Lawrence, A.J., 1998. Bradykinin B2
receptors in nodose ganglia of rat and human. Eur.
J. Pharmacol. 348 (2–3), 175–180.
Lahiri, S.C., Nag Chaudhuri, A.K., 1983. Release of kinin by the
scorpion (Heterometrus bengalensis C.L. Koch) venom.
Indian J. Exp. Biol. 21, 198–202.
Laye, S., Bluthe, R.M., Kent, S., Combe, C., Medica, C., Parnet,
P., Kelley, K., Dantzer, R., 1995. Subdiaphragmatic vagot-
omy blocks induction of IL-1beta mRNA in mice brain in
response to peripheral LPS. Am. J. Physiol. 268 (5),
R1327–R1331.
Levy, B.S., Carpenter, R., 1995. Perioperative pain management.
J. Am. Assoc. Gynecol. Laparosc. 2 (4), 381–387.
Magalhaes, M.M., Pereira, M.E., Amaral, C.F., Rezende, N.A.,
Campolina, D., Bucaretchi, F., Gazzinelli, R.T., Cunha-
Melo, J.R., 1999. Serum levels of cytokines in patients
envenomed by Tityus serrulatus scorpion sting. Toxicon 37
(8), 1155–1164.
Marinkelle, C.J., Stahnke, H.L., 1965. Toxicological and clinical
studies on Centruroides margaritatus (Gervais), a common
scorpion in western Colombia. J. Med. Entomol. 2 (2),
197–199.
Meki, A.R., Nassar, A.Y., Rochat, H., 1995. A bradykinin-
potentiating peptide (peptide K12) isolated from the venom of
Egyptian scorpion Buthus occitanus. Peptides 16 (8),
1359–1365.
Minano, F.J., Armengol, J.A., Sancibrian, M., Pomares, F.,
Benamar, K., Myers, R.D., 1997. Macrophage inflammatory
protein-1 beta and inducible nitric oxide synthase immunor-
eactivity in rat brain during prostaglandin E2- or lipopoly-
saccharide-induced fever. Ann. N. Y. Acad. Sci. 15, 272–280.
Morimoto, A., Nakamori, T., Watanabe, T., Ono, T., Muraka-
mi, N., 1988. Pattern differences in experimental fevers
induced by endotoxin, endogenous pyrogen, and prostaglan-
dins. Am. J. Physiol. 254, R633–R640.
Nakamura, K., Matsumura, K., Hubschle, T., Nakamura, Y.,
Hioki, H., Fujiyama, F., Boldogkoi, Z., Konig, M., Thiel,
H.J., Gerstberger, R., Kobayashi, S., Kaneko, T., 2004.
Identification of sympathetic promotor neurons in medullary
raphe regions mediating fever and other thermoregulatory
functions. J. Neurosci. 24 (23), 5370–5380.

ARTICLE IN PRESSA.C. Pessini et al. / Toxicon 48 (2006) 556–566566
Netea, M.G., Kullberg, B.J., Van der Meer, J.W., 2000.
Circulating cytokines as mediators of fever. Clin. Infect.
Dis. 31 (5), S178–S184.
O’Leary, D.S., Johnson, J.M., Taylor, W.F., 1985. Mode of
neural control mediating rat tail vasodilation during heating.
J. Appl. Physiol. 59 (5), 1533–1538.
O’Neill, L.A., Lewis, G.P., 1989. Interleukinin-1 potentiates
bradykinin- and TNF alpha- induced PGE2 release. Eur.
J. Pharmacol. 166 (2), 131–137.
Osman, O.H., Ismail, M., Wenger, T., 1973. Hyperthermic
response to intraventricular injection of scorpion venom: role
of brain monoamines. Toxicon 11, 361–368.
Paegelow, I., Werner, H., Reissmann, S., 1995a. Effects of
bradykinin and bradykinin analogues on spleen of mice. Eur.
J. Pharmacol. 279, 211–216.
Paegelow, I., Werner, H., Vietinghoff, G., Wartner, U., 1995b.
Release of cytokines from isolated lung strips by bradykinin.
Inflamm. Res. 44, 306–311.
Pan, Z.K., Zuraw, B.L., Lung, C.C., Prossnitz, E.R., Browning,
D.D., Ye, R.D., 1996. Bradykinin stimulates NF-kappaB
activation and interleukin 1beta gene expression in cultured
human fibroblasts. J. Clin. Invest. 98 (9), 2042–2049.
Pang, L., Knox, A.J., 1998. Bradykinin stimulates IL-8 produc-
tion in cultured human airway smooth muscle cells: role of
cyclooxygenase products. J. Immunol. 161 (5), 2509–2515.
Pessini, A.C., de Souza, A.M., Faccioli, L.H., Gregorio, Z.M.,
Arantes, E.C., 2003. Time course of acute-phase response
induced by Tityus serrulatus venom and TsTX-I in mice. Int.
Immunopharmacol. 3 (5), 765–774.
Petricevich, V.L., Teixeira, C.F., Tambourgi, D.V., Gutierrez,
J.M., 2000. Increments in serum cytokine and nitric oxide
levels in mice injected with Bothrops asper and Bothrops
jararaca snake venoms. Toxicon 38 (9), 1253–1266.
Petricevich, V.L., Pena, C.F., 2002. The dynamics of cytokine d
nitric oxide secretion in mice injected with Tityus serrulatus
scorpion venom. Mediators Inflamm. 11 (3), 173–180.
Phagoo, S.B., Poole, S., Leeb-Lundberg, L.M., 1999. Autoregu-
lation of bradykinin receptors: agonists in the presence of
interleukin-1b shift the repertoire of receptor subtypes from
B2 to B1 in human lung fibroblasts. Mol. Pharmacol. 56,
325–333.
Romanovsky, A.A., Ivanov, A.I., Shimansky, Y.P., 2002.
Selected contribution: ambient temperature for experiments
in rats: a new method for determining the zone of thermal
neutrality. J. Appl. Physiol. 92 (6), 2667–2679.
Roth, J., Storr, B., Goldbach, J., Voigt, K., Zeisberger, E., 1999.
Dose-dependent attenuation of lipopolysaccharide-fever by
inhibitors of inducible nitric oxide-synthase in guinea pigs.
Eur. J. Pharmacol. 383 (2), 177–187.
Roth, J., Souza, G.E., 2001. Fever induction pathways: evidence
from responses to systemic or local cytokine formation. Braz.
J. Med. Biol. Res. 34 (3), 301–314.
Sakamoto, W., Kaga, M., Handa, H., Gotoh, K., Suzuki, S.,
Fujie, K., Inoue, N., Nishihira, J., 1994. Bradykinin and Met-
T-kinin-Leu stimulated PGE2 production by rat macrophage
and fibroblast. Braz. J. Med. Biol. Res. 27 (8), 1923–1927.
Santos, D.R., Calixto, J.B., Souza, G.E., 2003. Effect of a kinin
B2 receptor antagonist on LPS- and cytokine-induced
neutrophil migration in rats. Br. J. Pharmacol. 139 (2),
271–278.
Sato, E., Nelson, D.K., Koyama, S., Hoyt, J.C., Robbins, R.A.,
2000. Bradykinin stimulates eotaxin production by a human
lung fibroblast cell line. J. Allergy Clin. Immunol. 106 (1),
117–123.
Simons, C.T., Kulchitsky, V.A., Sugimoto, N., Homer, L.D.,
Szekely, M., Romanovsky, A.A., 1998. Signaling the brain in
systemic inflammation: which vagal branch is involved in
fever genesis? Am. J. Physiol. 275 (1), R63–R68.
Sirko, S., Bishai, I., Coceani, F., 1989. Prostaglandin formation
in the hypothalamus in vivo: effect of pyrogens. Am.
J. Pshysiol. 256 (3), R616–R624.
Souza, G.E., Cunha, F.Q., Mello, R., Ferreira, S.H., 1988.
Neutrophil migration induced by inflammatory stimuli is
reduced by macrophage depletion. Agents Actions 24 (3–4),
377–380.
Souza, G.E., Cardoso, R.A., Melo, M.C., Fabricio, A.S., Silva,
V.M., Lora, M., De Brum-Fernandes, A.J., Rae, G.A.,
Ferreira, S.H., Zampronio, A.R., 2002. A comparative study
of antipyretic effects of indomethacin and dipyrone in rats.
Inflamm. Res. 51 (1), 24–32.
Strijbos, P.J., Hardwick, A.J., Relton, J.K., Carey, F., Rothwell,
N.J., 1992. Inhibition of central actions of cytokines on fever
and thermogenesis by lipocortin-1 involves CRF. Am. J.
Physiol. 263 (4), E632–E636.
Tambourgi, D.V., Petricevich, V.L., Magnoli, F.C., Assaf, S.L.,
Jancar, S., Dias da Silva, W., 1998. Endotoxemic-like shock
induced by Loxosceles spider venoms: pathological changes
and putative cytokine mediators. Toxicon 36 (2), 391–403.
Tegeder, I., Pfeilschifter, J., Geisslinger, G., 2001. Cycloxygenase-
independent actions of cyclooxygenase inhibitors. FASEB J.
15 (12), 2057–2072.
Tiffany, C.W., Burch, R.M., 1989. Bradykinin stimulates tumor
necrosis factor and interleukin-1 release from macrophages.
FEBS Lett. 247 (2), 189–192.
Vasconcelos, F., Lanchote, V.L., Bendhack, L.M., Giglio, J.R.,
Sampaio, S.V., Arantes, E.C., 2005. Effects of voltage-gated
Na+ channel toxins from Tityus serrulatus venom on rat
arterial blood pressure and plasma catecholamines. Comp.
Biochem. Physiol. C. Toxicol. Pharmacol. 141 (1), 85–92.
Volpe, A.R., Giardina, B., Preziosi, P., Carmignani, M., 1996.
Biosynthesis of endothelium-derived nitric oxide by bradyki-
nin as endogenous precursor. Immunopharmacology 33
(1–3), 287–290.
Watkins, L.R., Goehler, L.E., Relton, J.K., Tartaglia, N., Silbert,
L., Martin, D., Maier, S.F., 1995. Blockade of interleukin-1
induced hyperthermia by subdiaphragmatic vagotomy: evi-
dence for vagal mediation of immune-brain communication.
Neurosci. Lett. 183 (1-2), 27–31.
Werner, M.F., Fraga, D., Melo, M.C., Souza, G.E., Zampronio,
A.R., 2003. Importance of the vagus nerve for fever and
neutrophil migration induced by intraperitoneal LPS injec-
tion. Inflamm. Res. 52 (7), 291–296.
Young, A.A., Dawson, N.J., 1982. Evidence for on-off control of
heat dissipation from the tail of the rat. Can. J. Physiol.
Pharmacol. 60 (3), 392–398.
Zeisberger, E., 1999. From humoral fever to neuroimmunological
control of fever. J. Therm. Biol. 24, 287–326.




















