Extracellular matrix remodeling accompanies axial muscle development and morphogenesis in the mouse
15
PATTERNS & PHENOTYPES Extracellular Matrix Remodeling Accompanies Axial Muscle Development and Morphogenesis in the Mouse Marianne Deries 1 * Andr e B. Gonc ¸ alves, 1† Raquel Vaz, 1† Gabriel G. Martins, 1,2 Gabriela Rodrigues, 1,2 and So ´ lveig Thorsteinsdo ´ ttir 1,2 * Background: Skeletal myogenesis is extensively influenced by the surrounding environment. However, how the extracellular matrix (ECM) affects morphogenesis of muscles is not well understood. Results: We mapped the three-dimensional (3D) organization of fibronectin, tenascin, and laminin by immunofluores- cence during early epaxial myogenesis in mouse embryos. We define four stages of dermomyotome/myotome development and reveal the 3D organization of myogenic cells within their ECM during those stages. Fibro- nectin is abundant in all interstitial tissues, while tenascin is restricted to intersegmental borders. Bundles of fibronectin and tenascin also penetrate into the myotome, possibly promoting myocyte alignment. A lami- nin matrix delineates the dermomyotome and myotome and undergoes dynamic changes, correlating with key developmental events. Conclusion: Our observations cast new light on how myotomal cells interact with their environment and suggest that, as the segmented myotomes transform into the epaxial muscle masses, the laminin matrix disassembles and myocytes use the abundant fibronectin matrix to reach their final organization. Developmental Dynamics 241:350–364, 2012. V C 2011 Wiley Periodicals, Inc. Key words: mouse embryo; skeletal muscle development; dermomyotome; myotome; extracellular matrix; fibronectin; tenascin; laminin; integrins; 3D analysis Key findings: Four stages of dermomyotome/myotome development are defined. The interstitial matrix around thde dermomyotome and myotome is rich in fibronectin. Fibronectin and tenascin are enriched at intersegmental borders and penetrate into the myotome. Laminin matrices line the dermomyotome and myotome. These laminin matrices are disassembled as the dermomyotome dissociates and the myotome transforms into the epaxial muscle masses. Accepted 8 November 2011 INTRODUCTION Although all body muscles of amniote embryos derive from the epithelial somites, skeletal muscle development occurs in several different ways (Evans et al., 2006). Muscle progenitor cells either differentiate within the somite to form the segmented myotome which then progressively expands both dorso- medially (epaxially) and ventro-later- ally (hypaxially), or they migrate to reach the site where they differentiate, or they adopt a developmental pro- gram which is a combination of these Developmental Dynamics Additional Supporting Information may be found in the online version of this article. 1 Centro de Biologia Ambiental/Departamento de Biologia Animal, Faculdade de Cie ˆncias, Universidade de Lisboa, Lisbon, Portugal 2 Instituto Gulbenkian de Cie ˆncia, Oeiras, Portugal Grant sponsor: Fundac ¸a ˜ o para a Cie ˆncia e a Tecnologia (Portugal); Grant numbers: PTDC/BIA-BCM/67437/2006, SFRH/BPD/65370/2009. † A.B. Gonc ¸alves and R. Vaz contributed equally to this work. *Correspondence to: Marianne Deries and So ´ lveig Thorsteinsdo ´ ttir, Centro de Biologia Ambiental/Departamento de Biologia Animal, Faculdade de Cie ˆ ncias, Universidade de Lisboa, Lisbon, Portugal. E-mail: [email protected] and [email protected] DOI 10.1002/dvdy.23703 Published online 29 November 2011 in Wiley Online Library (wileyonlinelibrary.com). DEVELOPMENTAL DYNAMICS 241:350–364, 2012 V C 2011 Wiley Periodicals, Inc.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Extracellular matrix remodeling accompanies axial muscle development and morphogenesis in the mouse

a PATTERNS & PHENOTYPES
Extracellular Matrix Remodeling AccompaniesAxial Muscle Development and Morphogenesisin the MouseMarianne Deries1* Andr�e B. Goncalves,1† Raquel Vaz,1† Gabriel G. Martins,1,2
Gabriela Rodrigues,1,2 and Solveig Thorsteinsdottir1,2*
Background: Skeletal myogenesis is extensively influenced by the surrounding environment. However, howthe extracellular matrix (ECM) affects morphogenesis of muscles is not well understood. Results: Wemapped the three-dimensional (3D) organization of fibronectin, tenascin, and laminin by immunofluores-cence during early epaxial myogenesis in mouse embryos. We define four stages of dermomyotome/myotomedevelopment and reveal the 3D organization of myogenic cells within their ECM during those stages. Fibro-nectin is abundant in all interstitial tissues, while tenascin is restricted to intersegmental borders. Bundlesof fibronectin and tenascin also penetrate into the myotome, possibly promoting myocyte alignment. A lami-nin matrix delineates the dermomyotome and myotome and undergoes dynamic changes, correlating withkey developmental events. Conclusion: Our observations cast new light on how myotomal cells interactwith their environment and suggest that, as the segmented myotomes transform into the epaxial musclemasses, the laminin matrix disassembles and myocytes use the abundant fibronectin matrix to reach theirfinal organization. Developmental Dynamics 241:350–364, 2012. VC 2011 Wiley Periodicals, Inc.
Key words: mouse embryo; skeletal muscle development; dermomyotome; myotome; extracellular matrix; fibronectin;tenascin; laminin; integrins; 3D analysis
Key findings:� Four stages of dermomyotome/myotome development are defined.� The interstitial matrix around thde dermomyotome and myotome is rich in fibronectin.� Fibronectin and tenascin are enriched at intersegmental borders and penetrate into the myotome.� Laminin matrices line the dermomyotome and myotome.� These laminin matrices are disassembled as the dermomyotome dissociates and the myotome transforms intothe epaxial muscle masses.
Accepted 8 November 2011
INTRODUCTION
Although all body muscles of amnioteembryos derive from the epithelialsomites, skeletal muscle development
occurs in several different ways (Evanset al., 2006). Muscle progenitor cellseither differentiate within the somiteto form the segmented myotome whichthen progressively expands both dorso-
medially (epaxially) and ventro-later-ally (hypaxially), or they migrate toreach the site where they differentiate,or they adopt a developmental pro-gram which is a combination of these
Dev
elop
men
tal D
ynam
ics
Additional Supporting Information may be found in the online version of this article.1Centro de Biologia Ambiental/Departamento de Biologia Animal, Faculdade de Ciencias, Universidade de Lisboa, Lisbon, Portugal2Instituto Gulbenkian de Ciencia, Oeiras, PortugalGrant sponsor: Fundacao para a Ciencia e a Tecnologia (Portugal); Grant numbers: PTDC/BIA-BCM/67437/2006, SFRH/BPD/65370/2009.†A.B. Goncalves and R. Vaz contributed equally to this work.*Correspondence to: Marianne Deries and Solveig Thorsteinsdottir, Centro de Biologia Ambiental/Departamento deBiologia Animal, Faculdade de Ciencias, Universidade de Lisboa, Lisbon, Portugal. E-mail: [email protected] [email protected]
DOI 10.1002/dvdy.23703Published online 29 November 2011 in Wiley Online Library (wileyonlinelibrary.com).
DEVELOPMENTAL DYNAMICS 241:350–364, 2012
VC 2011 Wiley Periodicals, Inc.

Dev
elop
men
tal D
ynam
ics
two processes (Kalcheim et al., 1999;Hollway and Currie, 2005; Evanset al., 2006).
Soon after their formation, somitesundergo a compartmentalization giv-ing rise to the sclerotome (origin ofvertebrae, ribs, and tendons of backmuscles) and the epithelial dermomyo-tome (origin of skeletal muscle, der-mal, some endothelial, smooth muscle,and brown fat precursors; Kalcheimand Ben-Yair, 2005; Buckingham,2006; Christ et al., 2007; Seale et al.,2008). Migratory muscle progenitorsdelaminate from the hypaxial lip of thedermomyotome at specific axial levelsand migrate to their final destination(e.g., limbs, diaphragm). Only afterreaching their target sites, they enterthe myogenic program, activating theexpression of the myogenic regulatoryfactors (MRFs), Myf5, MyoD, MRF4,and myogenin, which orchestrate theircommitment and differentiation (Buck-ingham et al., 2003). Then, myoblastselongate and fuse to form myotubes,eventually maturing into muscle fibers.In contrast, muscle progenitors whichdifferentiate within the myotomal com-partment of the somites are releasedfrom the dermomyotomal lips directlyinto the myotomal space at all axiallevels (Ben-Yair and Kalcheim, 2005;Hollway and Currie, 2005). They enterthe myogenic program in situ and formthe segmented muscle masses, themyotomes. At a later stage, a new poolof proliferating muscle progenitorsexpressing Pax3 and Pax7 enter themyotome from the central dermomyo-tome (Ben-Yair and Kalcheim, 2005;Gros et al., 2005; Kassar-Duchossoyet al., 2005; Relaix et al., 2005), contrib-uting to the growth of the myotome.Remarkably, in amniote embryos, themyotome is only a transient structureand does not get innervated (Derieset al., 2008). Rather, the segmentedmyotomes are transformed: epaxiallyinto the complex deep back musclesand hypaxially into the intercostal andbody wall muscles (Christ et al., 1983;Tremblay et al., 1998; Kalcheim et al.,1999; Deries et al., 2010).
All these events occur within the con-text of a complex environment com-posed of neighboring cells, the diffusiblesignaling molecules they produce andthe surrounding extracellular matrix(ECM). The ECM is composed of macro-molecules, primarily glycoproteins and
proteoglycans (Frantz et al., 2010). It isinvolved in tissue differentiation anddevelopment not only as a mechanicalsupport but also by transmitting biome-chanical and biochemical signals medi-ated by transmembrane receptors suchas the integrins (van der Flier andSonnenberg, 2001; Barczyk et al., 2010;Rozario and DeSimone, 2010).
Although it is well established thatthe ECM plays an important role inseveral steps of myogenesis, the pic-ture of its interaction with cells ofdeveloping muscles is far from com-plete (Thorsteinsdottir et al., 2011). Tofully understand the role of the ECMduring embryogenesis, it is essentialto know the exact organization of thismatrix in relation to the developingtissues. No studies have addressed thethree-dimensional (3D) organization ofthe ECM during the different phasesof myotome morphogenesis. Moststudies so far have analyzed the dis-tribution pattern of ECM componentseither in whole-mount analysis of lowmagnification images or with higherdetail on tissue-sections with the draw-backs of distorted tissues and limited3D spatial organization details. Thealternative views offered by confocalimaging of whole-mount immunofluo-rescence-stained embryos followed by3D image reconstruction techniques(Martins et al., 2007) provide valuablemissing details into the complex 3Darchitecture of tissues and their ECMs.
Using this approach in mouseembryos, we mapped the organizationof the fibronectin, tenascin, and lamininmatrices around and within the dermo-myotome/myotome in 3D, and relatedthat organization with the location andmorphology of the dermomyotomal andmyotomal cells. We focused exclusivelyon the epaxial region between the fore-limb and hindlimb axial levels becausethe mechanisms of its development arerelatively well known and do not varywithin this region.
Fibronectin is the major glycopro-tein of the interstitial matrix in theearly embryo (Duband et al., 1987;Ostrovsky et al., 1988; Cachaco et al.,2005) and lack of fibronectin leads todefects in somitogenesis and neuraltube morphogenesis (George et al.,1993; Georges-Labouesse et al., 1996;Rifes et al., 2007; Martins et al., 2009;Rozario and DeSimone, 2010). More-over, fibronectin has been shown to
play a role during myotome develop-ment in zebrafish (Snow et al., 2008).Unlike fibronectin, tenascin appears
to be restricted to very specific areasduring embryogenesis (Riou et al., 1992;Jones and Jones, 2000). It is firstdetected after gastrulation and isobserved around the somites. Then, it ispresent during muscle development butdeclines by late gestation and becomesconfined to tendons (Crossin et al., 1986;Saga et al., 1992;Wang et al., 1995).Laminins are the major glycoproteins
of basement membranes. They areformed by the combination of threechains, a, b, and g (Durbeej, 2010). Sev-eral laminins are essential for embryo-genesis and their assembly is the firststep of basement membrane formationand also a prerequisite for the incorpo-ration of the other basement membranecomponents (Smyth et al., 1999; Minerand Yurchenco, 2004; Yurchenco andPatton, 2009; Durbeej, 2010). Laminin111 and 511 are part of the basementmembrane of the epithelial somites,dermomyotome, and myotome (Bajancaet al., 2006; Anderson et al., 2009; Thor-steinsdottir et al., 2011). Laminin 211and 221 are expressed later and are themajor laminins of the skeletal musclebasement membrane (Cachaco et al.,2005; Durbeej, 2010). Their presence inthe myotube basement membrane isimportant for myotube stability duringcontraction, and lack of laminin a2chain leads to muscle dystrophy (Guoet al., 2003; Huh et al., 2005).To precisely analyze cell-ECM rela-
tionships during dermomyotome/myo-tome development, we first defined fourdevelopmental stages (DMM stages1–4), which provide a useful tool fordevelopmental studies of the dermo-myotome/myotome. Our detailed 3Danalysis shows that fibronectin formsan abundant matrix in all the inter-stitial spaces and is also localized inthe intersomitic border where it may,in combination with tenascin, serve atendon-like function for the elongatedmyocytes. In support for this hypothe-sis, we found that av-integrins, whichcan serve as fibronectin and tenascinreceptors, are enriched at myocytetips. Bundles of fibronectin and tenas-cin also extend along the myocytes ofthe myotome, thus being in a goodposition to provide support for cellelongation, alignment, and even fusionand translocation later on. Laminin
ECM 3D ORGANIZATION AND MYOGENESIS 351

Dev
elop
men
tal D
ynam
ics
matrices show a surprisingly dynamicpattern; they are first assembled andthen disassembled during myotomedevelopment, and produced again bythe forming muscle masses. Of inter-est, the basement membrane matriceslining the dermomyotome and myo-tome are never completely continuous,most likely due to the transient natureof both of these compartments, buttheir discontinuous nature may inaddition permit a direct communica-tion route with neighboring cells. Thisdetailed 3D analysis provides newinsights of the potential roles of fibro-nectin, tenascin, and laminin matricesduring myotome development andepaxial muscle morphogenesis.
RESULTS AND DISCUSSION
Definition of Stages of
Dermomyotome/Myotome
Development in the Mouse
In the mouse, the formation of the epax-ial myotome starts at embryonic day (E)8.5 in the rostral somites (Venters et al.,1999; Bajanca et al., 2004) and byE12.5, the myotomes have completelydisappeared as segmented structures(Deries et al., 2010). During these fewdays, the myotome develops gradually,and different stages of development canbe identified in different embryo agesand in different rostro-caudal levels ofthe same embryo. Due to differences inmaturity of embryos between andwithin each litter, it has been difficult torely on a staging of dermomyotomes/myotomes based solely on age and axiallevel of the embryo. To address this diffi-culty and to allow accurate descriptionsof the changes in ECM organizationalong epaxial dermomyotome and myo-tome development, we defined four keystages from E9.5 to E11.5 which we des-ignate dermomyotome/myotome (DMM)stages 1 to 4 (Fig. 1A–D). After DMMstage 4, the dermomyotome fully dissoci-ates and the myotome undergoes adevelopmentally regulated translocationand splitting (Deries et al., 2010), whichculminates in a new organization, visi-ble at E12.5, and is termed the earlyepaxial muscle masses (Fig. 1E). Verylittle is known about the formation ofcervical and tail epaxial muscle, there-fore, this study focuses only on theepaxial muscles from forelimb to hind-limb levels.
DMM stage 1 (as in E9.5 interlimbto E9.5 forelimb level) corresponds toinitial stages of myotome formation.The lips of the dermomyotome curlaround the forming epaxial myotomeand release the myogenic precursorcells (MPCs) into the myotomal space(Venters et al., 1999; Gros et al., 2004;Hollway and Currie, 2005). MRF-expressing cells are found in the dor-sal (epaxial) part of the myotome anda few myoblasts have differentiatedinto myosin-positive myocytes which
are elongating (Fig. 1A). The dorsal-most part of the dermomyotome isrounded (Fig. 1A).At DMM stage 2 (as in E10.5 hind-
limb level to E10.5 interlimb level),both the dermomyotome and the myo-tome have expanded dorso-ventrally.Myoblasts and myocytes are numer-ous and myocytes span the wholewidth of the segment, except for theepaxial-most ones which are youngerand therefore shorter (Venters et al.,1999) (Fig. 1B). The dorsal-most
Fig. 1. Developmental stages of the epaxial dermomyotome and myotome. Four key stages ofepaxial dermomyotome/myotome development (DMM stages 1–4) are shown from the formationof the myotome to the dissociation of the central dermomyotome (A–D). Only the axis levelsfrom forelimb to hindlimb were studied. A: DMM stage 1: Appearance of the first fully differenti-ated myocytes. B: DMM stage 2: Growth of the dermomyotome in the dorso-medial to ventro-lateral direction compared with stage 1, but epaxial lip still rounded; accumulation of differenti-ated myocytes. C: DMM stage 3: Dermomyotome has grown further, and epaxial lip hasbecome oblong. Fully differentiated myocytes have increased in number and nonelongated myo-blasts, derived from the Pax3/Pax7 precursors entering the myotome from the central dermo-myotome, are seen between the myotome and dermomyotome. D: DMM stage 4: Centraldermomyotome sheet has de-epithelialized, but epaxial dermomyotome lip is still epithelial;epaxial-most myocytes have started to translocate; presence of a large number of nonelongatedmyoblasts within the muscle mass. E: The stage of early epaxial muscles is depicted to showthe end of the dermomyotome/myotome as a morphological unit; it shows the full disappear-ance of the dermomyotome and the transformation of the myotome. For simplicity, the caudaland rostral lips of the dermomyotome are not drawn. The illustrations are schematic and are notintended to be to scale with each other. DMM, dermomyotome/myotome; C, caudal; D, dorsal;L, lateral; M, medial; R, rostral; V, ventral; TS, transversospinalis unisegmental muscles; LG,longissimus muscle; IC, iliocostalis muscle; NT, neural tube; FL, forelimb level; HL, hindlimblevel; IL, interlimb level; MHC, myosin heavy chain; MRFs, myogenic regulatory factors.
352 DERIES ET AL.
Dev
elop
men
tal D
ynam
ics
part of the dermomyotome remainsrounded (Fig. 1B).
DMM stage 3 (as in E10.5 forelimblevel to E11.5 hindlimb level) is char-acterized by the expansion of the myo-tome and the dermomyotome rostro-caudally and myocytes have becomemore elongated (Fig. 1C). The myo-tome keeps growing dorso-ventrally,and is now thicker in the medio-lat-eral direction (Fig. 1C). Furthermore,the dorsal-most region of the dermo-myotome has changed shape and
become oblong (Fig. 1C). It is duringthis stage that the first MPCsexpressing Pax3/Pax7 are releasedfrom the central part of the dermo-myotome into the myotome (Kassar-Duchossoy et al., 2005; Relaix et al.,2005) some of which enter myogenesis.Thus, non-elongated myoblasts can beseen between the dermomyotome andmyotome (Fig. 1C).
At DMM stage 4 (as in E11.5 inter-limb to E11.5 forelimb), the cells ofthe central region of the dermomyo-
tome have undergone an asymmetriccell division thus segregating themuscle and the dermis precursor cells(Ben-Yair and Kalcheim, 2005) leav-ing only the epaxial lip (as well as thehypaxial lip; Tajbakhsh and Bucking-ham, 2000) as epithelial remnants. Themyotome has continued to grow andsome myocytes of the dorsal-most part
Fig. 2.
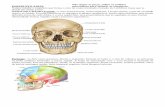
Fig. 2. A fibronectin matrix is predominant atall myotome stages. A–O: Three-dimensional(3D) reconstructions of individual segmentsobtained from confocal imaging of immunofluo-rescence in whole-mounts showing fibronectin(green) and myogenic cells (red, stained formyogenin and MHC at DMM stage 1 and 2, A–F; stained for MHC at DMM stage 3 and 4, G–O). Dorsal is always upward, rostral is to the lefton lateral views, and to the right on medialviews. Medial is to the right on transverse views.Transverse slabs of the 3D reconstructions areshown (C,F,J,N,O) and the level of the slabs isindicated by a dashed line (in B,E,H,L). For ananimated version of these 3D reconstructions,see Supplementary Movies S1–S4. Fibronectinis abundant around the dermomyotome and themyotome throughout all stages. A–K: At DMMstages 1–3, only the fibronectin matrix lining thesegment is shown to enable a clearer analysis ofthe matrix associated with the dermomyotomeand the myotome. A–C: At DMM stage 1, fibro-nectin-positive fibrils are present at the lateralside of the dermomyotome (A,C) and the medialside of the myotome (B,C). C: No fibronectinfibrils are seen within the myotome and betweenthe dermomyotome and myotome. D–K: AtDMM stages 2 and 3, the fibronectin matrix isdenser on the lateral side of the dermomyotome(D,F,G,I,J) and fibronectin-positive bundles arepresent in the area between the myotome anddermomyotome (F,J, arrow) and in the musclemass (J, arrowhead). A lateral view where the fi-bronectin labeling was made 70% transparentmedially and laterally evidences the strong fibro-nectin labeling between the dermomyotomeand the myotome (K). Due to this transparency,tips of myocytes are now visible within the ten-don-like structure at intersegmental boundaries(K, insert). Note the extension of fibronectin ma-trix lining a presumptive blood vessel growingfrom the basal side of the dermomyotomethrough the dermomyotome and reaching themyotome (J,K, asterisk). L–O: At DMM stage 4,all the fibronectin matrix is shown around thesegment (L) demonstrating that this matrix com-pletely ensheaths the myocytes (shown withoutfibronectin in M) and has invaded the space leftby the dermomyotome after its dissociation(N,O, arrowheads). While the epaxial lip of thedermomyotome remains epithelial, fibronectinsurrounds this epithelium (N, arrow). Two seg-ments rostral to N in the same embryo, theepaxial lip has de-epithelialized and fibronectinhas invaded the space (O, arrow). Note thespace occupied by a nerve growing toward themyotome (O, asterisk). DMM, dermomyotome/myotome; MHC, myosin heavy chain. Scalebars ¼ 100 mm.
ECM 3D ORGANIZATION AND MYOGENESIS 353

Dev
elop
men
tal D
ynam
ics
of the segment have changed orienta-tion and are no longer parallel to therostro-caudal axis of the embryo (Fig.1D). Moreover, it is at this stage thatmyotomal myocytes are becoming mul-tinucleated (Deries et al., 2010). Fromthis point onward, the segmentedshape of the myotome progressivelytransforms into the complex organiza-tion of the axial muscle masses (Derieset al., 2010). An early epaxial musclemass stage (from E12.5 onward) is rep-resented in Figure 1E to illustrate thedisappearance of the dermomyotome/myotome in the mammalian embryo.The most dorsal muscle masses at thisstage are the anlagen of the transver-sospinalis muscles, some of which stayunisegmental. Ventro-lateral to them,the longissimus is formed, followed bythe iliocostalis (Fig. 1E), both of whichspan several segments (Vallois, 1922).
In this study, we focus on the rela-tionship between the ECM and thedermomyotome/myotome along thefour DMM stages. We also describethe relationship between the ECMand the translocating myocytes of thepresumptive unisegmental transver-sospinalis muscles, which are the firstto translocate (Deries et al., 2010).
The Interstitial Matrix
Around the Dermomyotome
and Myotome Is Rich in
Fibronectin
To better understand the relationshipof the dermomyotomal and myotomalcells with their surrounding intersti-tial ECM, the precise 3D organizationof fibronectin and tenascin aroundand within the dermomyotome/myo-tome was analyzed from DMM stages1 to 4 and after the dissociation of thedermomyotome and translocation ofthe myotome. Whole-mount embryoswere co-immunolabeled for myogenin/myosin heavy chain (MHC) and fibro-nectin or tenascin and confocalimages were rendered to obtain a 3Dreconstruction of these matrices withthe myogenic cells (see the Experi-mental Procedures section).
During all DMM stages, a fibronec-tin matrix (Fig. 2; Supp. Movies S1–S4, which are available online) fillsthe interstitial space on the lateralside of the dermomyotome (betweenthe dermomyotome and ectoderm;Fig. 2A,C,D,F,G,I,J,N,O; Supp. Mov-
ies). Fibronectin is highly enriched atintersegmental boundaries at allDMM stages (shown for DMM stage3; Fig. 2I,K) and is also found on themedial side of the myotome (Fig.2B,C,E,F,H,J,L,N,O).
Tenascin has a much more re-stricted distribution than fibronectin(Fig. 3; Supp. Movies S5–S7). Consist-ent to what has been reported for thechick embryo (Crossin et al., 1986), atDMM stage 1, tenascin is detected atintersegmental borders, although thelevels of the immunostaining werenot sufficiently above background toallow a reliable reconstruction. FromDMM stage 2 onward, tenascin progres-sively becomes highly enriched in theintersegmental space (Fig. 3A,B,D,E).Some tenascin matrix is present onthe lateral side of the dermomyotome,but the immunostaining was againweak and could not be reconstructed.
The epithelial and segmented orga-nization of the somite is dependent onthe surrounding fibronectin matrix asshown by blocking fibronectin matrixassembly in cultured chick embryoexplants (Rifes et al., 2007; Martinset al., 2009). In these explants, notonly is somitogenesis impaired, butalso already formed somites start to de-epithelialize. The fact that the dermo-myotome retains a fibronectin matrixafter somite compartmentalization sug-gests that the support of a fibronectinmatrix is still needed for its epithelialand segmented organization. Duringearly embryogenesis, tenascin hasbeen reported to be deposited after afibronectin matrix is in place (Crossinet al., 1986; Saga et al., 1992), which isconsistent with our observations.
Fibronectin and Tenascin Are
Enriched at Intersegmental
Borders, Possibly Serving a
Tendon-Like Function for
Myotomal Myocytes
When myotomal myocytes elongateacross the full length of the segment,their tips insert into the fibronectinand tenascin matrices at interseg-mental borders (Figs. 2K, insert; 3A–F, arrows). Thus, the fibronectin andtenascin matrices may serve as atendon-like structure providing ananchor for the elongated, differenti-ated myocytes. This intersegmental
structure has been shown to be essen-tial for myotome organization inzebrafish embryos because the disrup-tion of fibronectin leads to a disorga-nization of myotomal cells (Snowet al., 2008). It would be interesting toverify whether the disruption of thismatrix in the mouse leads to the samephenotype.The a4b1 integrin, a fibronectin
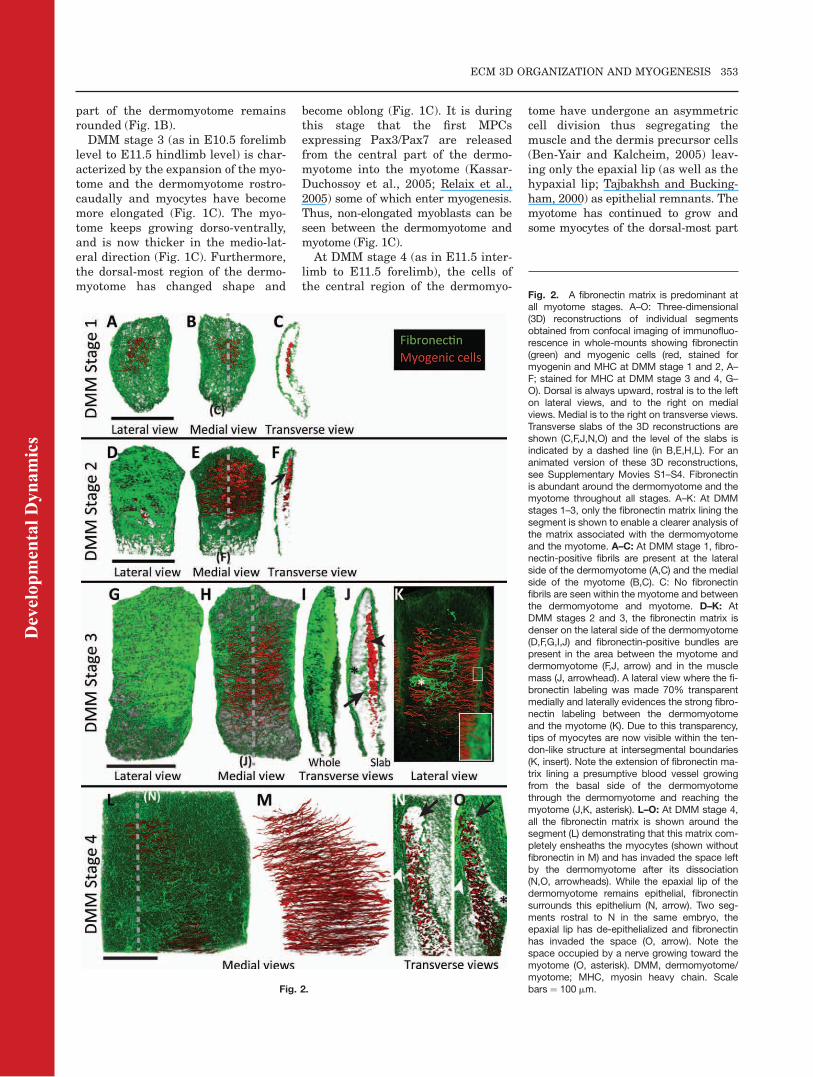
and VCAM1 receptor, is enriched atthe tips of elongated myocytes(Bajanca et al., 2004), thus being in aposition to mediate their attachmentto the intersegmental fibronectin ma-trix. Furthermore, mRNA for Itga5,the a chain of the a5b1 fibronectin re-ceptor, is present in the myotome(Bajanca et al., 2004; Cachaco et al.,2005). We have extended those obser-vations to other fibronectin receptorsas well as tenascin receptors on themyocytes. The integrins avb3 anda9b1 are two receptors for tenascinand both avb3 and avb1 bind to fibro-nectin (Thorsteinsdottir et al., 2011).Immunohistochemistry on frontal cry-osections for av and a9, combinedwith MHC staining, shows that boththese subunits are present in themature somite already at DMM stage2 onward (Fig. 4), earlier than previ-ously described using immunohisto-chemistry with an enzymatic reaction(Hirsch et al., 1994; Wang et al.,1995). The av subunit is present in allelongated myocytes from the forma-tion of the myotome to the morpho-genesis of epaxial muscle masses (Fig.4A–C and data not shown). This subu-nit is enriched at the myocyte tips(Fig. 4A–C, arrows), making it a goodcandidate as a receptor for both fibro-nectin and tenascin. In contrast, thea9 subunit is not present on myo-cytes, but is localized on the basal andapical sides of the dermomyotome aswell as in the sclerotome (Fig. 4D–F).Thus, although it is present at inter-segmental borders, where tenascin islocalized, a9b1 appears to be re-stricted to the dermomyotome and isnot present on myocyte tips. Thea9b1 also binds laminin (Durbeej,2010), which could be its major ligandat the basal side of the dermomyo-tome (see below).We conclude that myotomal myo-
cytes are in a position to attach to thefibronectin matrix through a4b1,a5b1 and av-containing integrin(s)
354 DERIES ET AL.
Dev
elop
men
tal D
ynam
ics
and may also attach to tenascinthrough avb3. These integrins couldbe playing an important role inanchoring myogenic cells to the ECMat intersegmental borders, providingmechanical support and signals whichwould promote their differentiationand/or maturation.
Fibronectin and Tenascin
Bundles Penetrate the Space
Between the Dermomyotome
and Myotome as Well as
Between Myocytes
By using immunolabeling in cryosec-tions, we have previously shown thata patchy immunoreactivity for fibro-nectin is present between the dermo-myotome and myotome as well aswithin the myotome (Cachaco et al.,
Fig. 3.
Fig. 4.
Fig. 3. Tenascin matrix is dense at interseg-mental borders. A–I: Three-dimensional (3D)reconstructions of individual segmentsobtained from confocal imaging of immunoflu-orescence in whole-mounts showing tenascin(green) and myogenic cells (red, stained formyogenin and MHC at DMM stage 2 A–C; andfor MHC at DMM stage 4 and early epaxialmuscles, D–I). Epaxial is always upward, rostralis to the left on lateral views, and to the right onmedial views. Medial is to the right on transverseviews. Transverse slabs of the 3D reconstruc-tions are shown (C,G) and the levels of the slabsare indicated with dashed lines (in B,E). For ananimated version of these 3D reconstructionssee Supplementary Movies S5–S7. A–G: Thetenascin matrix lining the dermomyotome is tooweak to be reconstructed. Tenascin is highlyabundant at intersegmental borders (A,B,D,E,arrows) and myocyte tips elongate through thematrix (F, arrow). Bundles of tenascin matrix areextended along the medial and lateral sides ofthe myotome (A,B,D,E, arrowheads) and withinthe muscle mass (G, arrowhead). H,I: When themost dorsal epaxial muscles are formed (trans-versospinalis unisegmented and multiseg-mented), tenascin matrix is concentrated at thetendon sites (arrows). MHC, myosin heavy chain;DMM, dermomyotome/myotome; US, uniseg-mented transversospinalis; MS, multisegmentedtransversospinalis. Scale bars¼ 100 mm.
Fig. 4. The av integrin subunit, but not a9, ispresent on myocytes. A–F: Co-immunofluores-cence on frontal cryosections stained for integ-rin subunit av (A–C) or a9 (D–F) and MHC (red)both at DMM stage 2. A–C: av is present on themyocytes of the forming myotome and isenriched at their tips (arrows, A,C). D–F: The a9is present around the dermomyotome and in thesclerotome but not in the myotome. MHC, myo-sin heavy chain; DM, dermomyotome; M, myo-tome; Scl, sclerotome. Scale bars ¼ 50 mm.
ECM 3D ORGANIZATION AND MYOGENESIS 355

Dev
elop
men
tal D
ynam
ics
2005). The 3D reconstruction of theorganization of the fibronectin matrixreveals that the patches seen in sec-tions represent fibronectin bundleswhich are spread on the surface ofand within the myotome, generallyrunning parallel to the myocytes (Fig.2J,K). At DMM stage 1, these bundlesare present on the medial side of themyotome (Fig. 2B,C). From DMMstage 2, they also appear in the spacebetween the myotome and dermomyo-tome (Fig. 2F, arrow), and becomeparticularly evident in this locationfrom DMM stage 3 onward (Fig. 2J,arrow). They are also seen within themuscle mass (Fig. 2J, arrowhead). Fi-bronectin bundles remain abundanton the medial side of the myotome atall DMM stages (Fig. 2B,E,H). Atearly stages, the bundles are thin(Fig. 2B,E) but as the myotome grows,they become thicker (Fig. 2H,K). 3Dreconstruction of the tenascin matrixat DMM stage 2 shows that fibrils oftenascin, originating in the denseintersegmental matrix, are also run-ning along the surface of the myotome,on both the medial and the lateralsides (Fig. 3A,B, arrowheads; Supp.Movie S5). By DMM stage 4, thicktenascin bundles spread over the myo-tome both laterally and medially (Fig.3D,E, arrowheads; Supp. Movie S6)and tenascin bundles are also detectedwithin the muscle mass (Fig. 3G,arrowhead; Supp. Movie S6). Thesefibrils are distinct from the ones of fi-bronectin and their shape is different(compare Fig. 2H,K and 3D,E). As inthe case of the fibronectin fibrils, thetenascin fibrils grow in size, extendfrom the rostral and caudal interseg-mental borders and join in the middleof the myotome (Fig. 3D,E).
The fact that the bundles of fibro-nectin run mainly parallel to the myo-cytes (Fig. 2H,K) suggests that themyocytes may have a role in theirassembly and/or that these bundles aidmyocyte alignment along the axis. Thetenascin matrix may give structuralsupport to the fibronectin fibrils asreported in other systems (Jones andJones, 2000). It is tempting to proposethat when muscle cells become multi-nucleated, cell alignment for myocytefusion is guided by the fibronectin ma-trix. Although a5b1, a4b1 and avintegrins are not essential for myogeniccell fusion itself in vitro (Yang et al.,
1996; Blaschuk et al., 1997; Tavernaet al., 1998), cell engagement with thefibronectin matrix present within themyotome in vivo may serve to promotethe approximation of myogenic cells,increasing the opportunities for fusion.The fact that b1D integrin knock-inand conditional b1 integrin knock-outmouse embryos show defects in myo-tube formation (Cachaco et al., 2003;Schwander et al., 2003) supports thishypothesis.
It is interesting to note that the fi-bronectin fibrils on the lateral side ofthe myotome, i.e., between the myo-tome and the dermomyotome (Fig. 2J,arrow, K), are present at the timewhen the central part of the epithelialdermomyotome gives rise to the mus-cle progenitor cells expressing Pax3and Pax7 (Kassar-Duchossoy et al.,2005; Relaix et al., 2005). It has beenshown by live imaging that epithelialsomite cells are very dynamic, beingable to extend protrusions, reachingfor the fibronectin matrix and shut-tling between the somitic epitheliumand the somitocoel (Martins et al.,2009). It is therefore tempting to sug-gest that the Pax3/Pax7 progenitorcells may use the fibronectin matrixto translocate from the dermomyo-tome into the myotome in a similarway as the cells of epithelial somitesenter and exit the somitocoel.
When the Dermomyotome
Dissociates, a Fibronectin
Matrix Ensheaths and
Penetrates the Myotome,
While the Tenascin Matrix
Remains at Myocyte Ends
After the dissociation of the centraldermomyotome at DMM stage 4,fibronectin has invaded all the tissuessurrounding the myotome, includingthe forming dermis (Fig. 2L–O,arrowheads in N,O; Supp. Movie S4).This fibronectin matrix is dense andis tightly associated with the myo-tome. While the epaxial dermomyo-tome lip remains epithelial, the areaoccupied by it is free of fibronectin(Fig. 2N, arrow; transverse slab). How-ever, as soon as the lip de-epithelial-izes, the fibronectin matrix invadesthe space and becomes tightly associ-ated with the muscle cells (Fig. 2O,arrow; transverse slab, showing a
more mature myotome, two segmentsrostral to the one depicted in Figure2N). The interstitial fibronectin matrixremains present and is very denseduring the whole transformation ofthe myotome. The fibronectin recep-tors a4b1, a5b1 and av-containingintegrins are expressed on the myo-cytes during the whole process(Cachaco et al., 2005; and data notshown). Fibronectin is thus in a posi-tion to guide the reorganization of themyogenic cells of the myotome by pro-moting directional fusion or by serv-ing as a substrate for their transloca-tion (Deries et al., 2010), or both.In contrast to the almost ubiquitous
presence of fibronectin around theDMM stage 4 myotome and early dor-sal-most muscle masses, the tenascinmatrix has a specific 3D organization,being enriched at the ends of and alongmyotomal myocytes (Fig. 3H,I; Supp.Movie S7). Tenascin is known to play acrucial role during tissue repair and tu-mor development (Jones and Jones,2000; Chiquet-Ehrismann and Tucker,2004; Calve et al., 2010), but tenascin-null mice do not show any dramaticphenotype during development (Sagaet al., 1992; Forsberg et al., 1996; Gar-cion et al., 2001). It is possible thatother matrices compensate for itsabsence or that the early defects arecompensated later in development. Itwould be interesting to investigateearly stages of development and verifythe phenotype of early myogenesis inthese mutants. Tenascin and fibronec-tin matrices are both localized at theintersegmental border and along themyocytes, so fibronectin may alsocompensate for the absence of tenascin.However, when the myotome trans-forms into the dorsal-most epaxialmuscle masses, their pattern becomesdifferent. A fibronectin matrix fills thespace around and within the myotome(as shown for DMM stage 4; Fig. 2L–O;Supp. Movie S4), whereas tenascin isprimarily found at the tips of elongatedmyocytes/early myotubes and betweencleaving muscle masses (Fig. 3H,I,arrows; Supp. Movie S7). This indicatesthat, at this stage, their roles diverge.We suggest that fibronectin may guidethe reorientation of the myotomal myo-cytes whereas tenascin is in a positionto play a tendon- and muscle connec-tive tissue-like role (Deries et al., 2010;Mathew et al., 2011).
356 DERIES ET AL.

Dev
elop
men
tal D
ynam
ics
The Laminin Matrices of the
Dermomyotome and Myotome
Increase in Complexity From
DMM Stages 1 to 3, Without
Ever Forming Fully
Continuous Basement
Membranes
Two basement membranes are essen-tial during the first stages of myogen-esis in the mouse: one lines the basalside of the dermomyotome and plays asignificant role in preventing muscleprogenitors from entering the myo-genic program too early (Bajancaet al., 2006). The other is the basementmembrane covering the medio-ventralsurface of the myotome, which sepa-rates it from the sclerotome and keepsmyogenic cells from getting dispersedinto the sclerotome (Tosney et al.,1994; Tajbakhsh et al., 1996; Bajancaet al., 2006; Anderson et al., 2009). Itshould also be noted that both are dis-tinct from the basement membranethat later surrounds the maturingmuscle fibers (Cachaco et al., 2005).
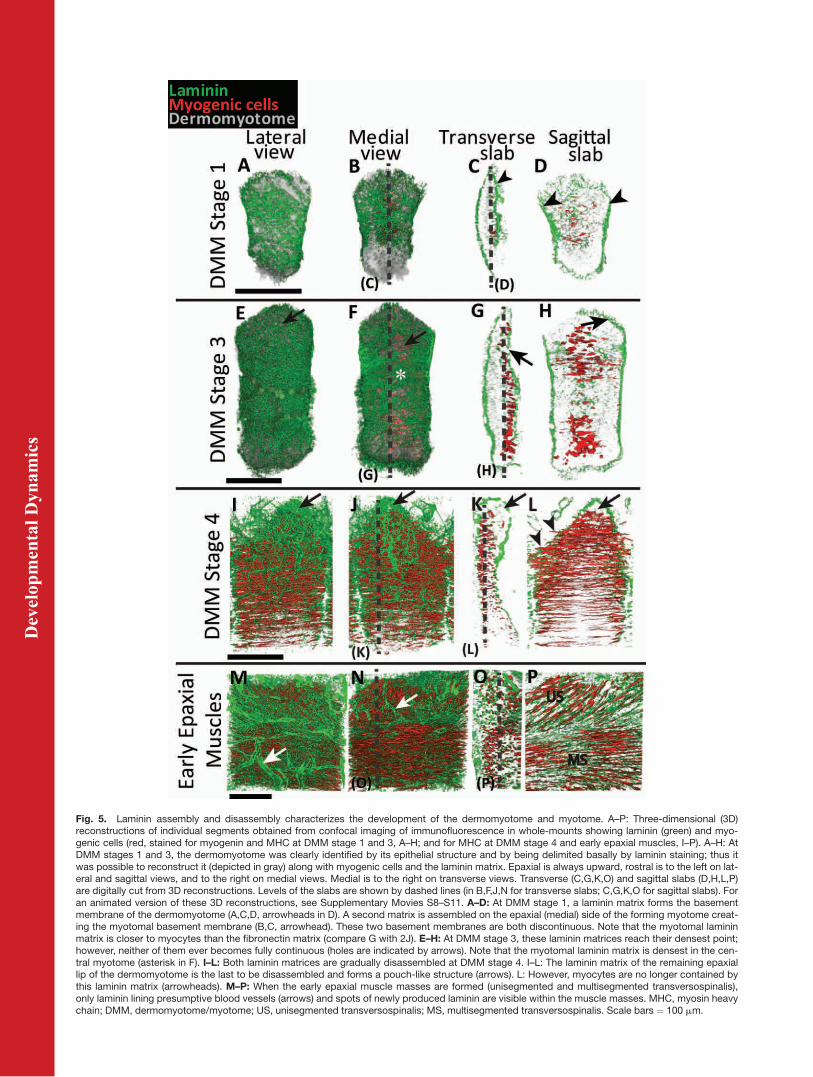
Earlier analyses of laminin distribu-tion in mouse embryo sections, usingan antibody that recognizes all lami-nins present (except the epidermal lam-inin 332), reveals a continuous line ofimmunoreactivity on the dermomyo-tome and a line of matrix separatingthe myotome from the sclerotome(Bajanca et al., 2004; Cachaco et al.,2005; Anderson et al., 2009). The otherbasement membrane molecules (colla-gen type IV, nidogen, and perlecan)have an identical pattern (Bajancaet al., 2006; Anderson et al., 2009). Our3D analysis of laminin distributionwith the same antibody (see the Experi-mental Procedures section) confirmsthat laminin is organized into a base-ment membrane sheet on the basal sideof the dermomyotome and between themyotome and sclerotome. However, italso provides new information, reveal-ing that this basement membrane sheetchanges significantly along the DMMstages defined here. At DMM stage 1(Fig. 5A–D; Supp. Movie S8), the basalside of the dermomyotome is lined bya loose basement membrane matrix,containing several holes (Fig. 5A,C),but this matrix becomes slightlymore dense at the rostral and caudalsegmental boundaries of the dermo-myotome (Fig. 5D, arrowheads). Themyotomal laminin matrix, which is
progressively assembled by the Myf5-positive myoblasts (Bajanca et al.,2006) is visible as a discontinuous sheeton the medial side of the myotome atDMM stage 1 (Fig. 5B,C, arrowhead;Supp. Movie S8). It is present only inassociation with myoblasts.
As the dermomyotome/myotomedevelops, the laminin basement mem-brane of the dermomyotome becomesdenser and the laminin matrix of themedial side of the myotome growsconcomitantly with the addition ofnew cells to the myotome. The centralpart of this matrix is denser than thedorsal and the ventral part (shown atDMM stage 3, Fig. 5F, asterisk, Supp.Movie S9), probably because it is themost mature part of the myotome(Venters et al., 1999). At DMM stage 3(Fig. 5E–H; Supp. Movie S9), both ofthese laminin matrices have reachedtheir highest density. Strikingly, theselaminin matrices are never completelycontinuous basement membrane sheets(see holes, Fig. 5E–H, arrows; Supp.Movie S9) such as those of the ectodermand the neural tube (data not shown).These discontinuities in the matrix arenot visible on sections (Bajanca et al.,2006; Anderson et al., 2009).
The observation that these two lami-nin matrices never form a completebasement membrane is surprising.Nonetheless, they maintain the epi-thelial dermomyotome and contain themyogenic cells within the myotomalcompartment. Furthermore, lamininengagement by dermomyotomal cellshas been shown to promote the undif-ferentiated state of the dermomyotome(Bajanca et al., 2006) while the myoto-mal laminin matrix aids the alignmentof myotomal myocytes (Wilson-Rawlset al., 1999; Bajanca et al., 2006; Seoet al., 2006). It is possible that, due tothe transient nature and rapid growthof both the dermomyotome and themyotome, the production of a completebasement membrane is an unneces-sary investment and an incompletebasement membrane is sufficient inthis context. Furthermore, the holes inthe basement membrane give the cellsof both the dermomyotome and myo-tome access to the surrounding inter-stitial matrix which could providestructural support for both of these tis-sues. Another hypothesis to explainthe incomplete dermomyotomal base-ment membrane could be that this
epithelium is very dynamic with pro-trusions of cells sent out and retractedagain (Gros et al., 2009; Martins et al.,2009), not permitting the formation of acontinuous laminin sheet. The holes inthe laminin matrix may also facilitatejuxtacrine cell–cell interactions. In thiscontext, it is interesting to note thatpassing migrating neural crest cellshave been shown to touch epaxial der-momyotome cells and thus induce notchactivation in these dermomyotomalcells which triggers their entry in themyogenic program (Rios et al., 2011).
DMM Stage 4 Is
Characterized by a
Progressive Disassembly of
the Laminin Matrices
At DMM stage 4, the dermomyotomallaminin matrix has largely disappeared(Fig. 5I–L; Supp. Movie S10), possiblythrough the action of matrix metallo-proteases (MMPs; Duong and Erickson,2004), and most of the dermomyotomehas dispersed to release the mesenchy-mal dermis and muscle precursors(Ben-Yair and Kalcheim, 2005). Thelaminin matrix has also been disas-sembled on the medial side of the myo-tome. Only the dorsal-most part of thelaminin matrix remains (Fig. 5I–L).Intriguingly, the 3D reconstructionsreveal that this matrix forms a pouch-like structure on the medial side of themyotome (Fig. 5I–L, arrows; Supp.Movie S10). However, the dorsal-mostmyocytes, which at this stage areacquiring a new orientation, cross thelaminin matrix at the rostral and cau-dal segment borders, and extendthrough the matrix (Fig. 5L, arrow-head; Supp. Movie S10). This indicatesthat the basement membrane liningthis pouch-like refuge of epithelial cellsis no longer capable of containingmyocytes.After the reorganization of the epax-
ial muscles, the dermomyotomal andmyotomal lamininmatrices have disap-peared and laminin is observed in a‘‘spotty’’ form around and within theearlymusclemasses, indicating de novoproduction (Fig. 5M–P; Supp. MovieS11). It is tempting to suggest that thislamininmay, at least in part, contributeto the basement membrane seen sur-rounding each muscle fiber from E13.5onward (Cachaco et al., 2005).
ECM 3D ORGANIZATION AND MYOGENESIS 357
Dev
elop
men
tal D
ynam
ics
Fig. 5. Laminin assembly and disassembly characterizes the development of the dermomyotome and myotome. A–P: Three-dimensional (3D)reconstructions of individual segments obtained from confocal imaging of immunofluorescence in whole-mounts showing laminin (green) and myo-genic cells (red, stained for myogenin and MHC at DMM stage 1 and 3, A–H; and for MHC at DMM stage 4 and early epaxial muscles, I–P). A–H: AtDMM stages 1 and 3, the dermomyotome was clearly identified by its epithelial structure and by being delimited basally by laminin staining; thus itwas possible to reconstruct it (depicted in gray) along with myogenic cells and the laminin matrix. Epaxial is always upward, rostral is to the left on lat-eral and sagittal views, and to the right on medial views. Medial is to the right on transverse views. Transverse (C,G,K,O) and sagittal slabs (D,H,L,P)are digitally cut from 3D reconstructions. Levels of the slabs are shown by dashed lines (in B,F,J,N for transverse slabs; C,G,K,O for sagittal slabs). Foran animated version of these 3D reconstructions, see Supplementary Movies S8–S11. A–D: At DMM stage 1, a laminin matrix forms the basementmembrane of the dermomyotome (A,C,D, arrowheads in D). A second matrix is assembled on the epaxial (medial) side of the forming myotome creat-ing the myotomal basement membrane (B,C, arrowhead). These two basement membranes are both discontinuous. Note that the myotomal lamininmatrix is closer to myocytes than the fibronectin matrix (compare G with 2J). E–H: At DMM stage 3, these laminin matrices reach their densest point;however, neither of them ever becomes fully continuous (holes are indicated by arrows). Note that the myotomal laminin matrix is densest in the cen-tral myotome (asterisk in F). I–L: Both laminin matrices are gradually disassembled at DMM stage 4. I–L: The laminin matrix of the remaining epaxiallip of the dermomyotome is the last to be disassembled and forms a pouch-like structure (arrows). L: However, myocytes are no longer contained bythis laminin matrix (arrowheads). M–P: When the early epaxial muscle masses are formed (unisegmented and multisegmented transversospinalis),only laminin lining presumptive blood vessels (arrows) and spots of newly produced laminin are visible within the muscle masses. MHC, myosin heavychain; DMM, dermomyotome/myotome; US, unisegmented transversospinalis; MS, multisegmented transversospinalis. Scale bars ¼ 100 mm.
Dev
elop
men
tal D
ynam
ics
The Last Assembled Laminin
Matrix Correlates With the
Epithelial State of the
Dermomyotomal Epaxial Lip
Because the epaxial pouch-like lami-nin matrix is no longer present as abarrier for myocytes, it may representthe basement membrane of the
remaining epithelial lip of the epaxialdermomyotome, thus containing alast refuge of dermomyotomal epithe-lial cells, in an otherwise fully mesen-chymal milieu (Fig. 5I–L, arrows).
Whole-mount co-immunofluores-cence staining for Pax3/Pax7 (mark-ing dermomyotomal cells and MPCs)and laminin showed that the persis-
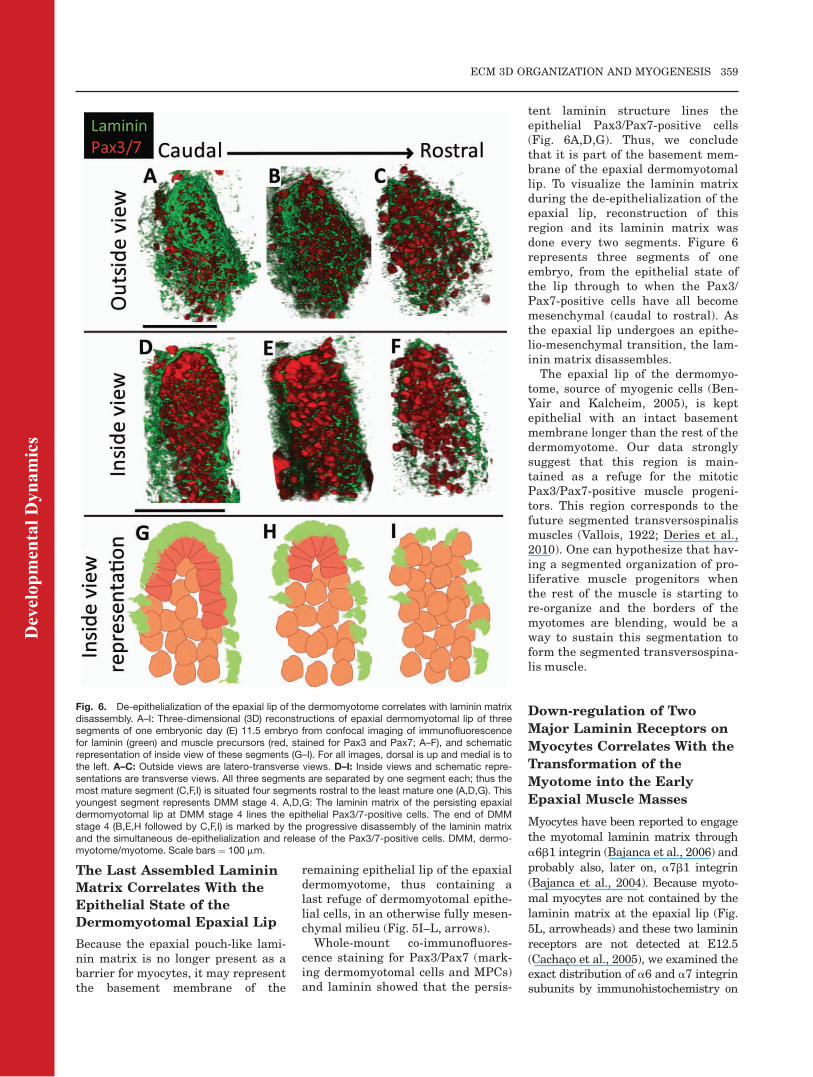
tent laminin structure lines theepithelial Pax3/Pax7-positive cells(Fig. 6A,D,G). Thus, we concludethat it is part of the basement mem-brane of the epaxial dermomyotomallip. To visualize the laminin matrixduring the de-epithelialization of theepaxial lip, reconstruction of thisregion and its laminin matrix wasdone every two segments. Figure 6represents three segments of oneembryo, from the epithelial state ofthe lip through to when the Pax3/Pax7-positive cells have all becomemesenchymal (caudal to rostral). Asthe epaxial lip undergoes an epithe-lio-mesenchymal transition, the lam-inin matrix disassembles.The epaxial lip of the dermomyo-
tome, source of myogenic cells (Ben-Yair and Kalcheim, 2005), is keptepithelial with an intact basementmembrane longer than the rest of thedermomyotome. Our data stronglysuggest that this region is main-tained as a refuge for the mitoticPax3/Pax7-positive muscle progeni-tors. This region corresponds to thefuture segmented transversospinalismuscles (Vallois, 1922; Deries et al.,2010). One can hypothesize that hav-ing a segmented organization of pro-liferative muscle progenitors whenthe rest of the muscle is starting tore-organize and the borders of themyotomes are blending, would be away to sustain this segmentation toform the segmented transversospina-lis muscle.
Down-regulation of Two
Major Laminin Receptors on
Myocytes Correlates With the
Transformation of the
Myotome into the Early
Epaxial Muscle Masses
Myocytes have been reported to engage
the myotomal laminin matrix through
a6b1 integrin (Bajanca et al., 2006) and
probably also, later on, a7b1 integrin
(Bajanca et al., 2004). Because myoto-
mal myocytes are not contained by the
laminin matrix at the epaxial lip (Fig.
5L, arrowheads) and these two laminin
receptors are not detected at E12.5
(Cachaco et al., 2005), we examined theexact distribution of a6 and a7 integrinsubunits by immunohistochemistry on
Fig. 6. De-epithelialization of the epaxial lip of the dermomyotome correlates with laminin matrixdisassembly. A–I: Three-dimensional (3D) reconstructions of epaxial dermomyotomal lip of threesegments of one embryonic day (E) 11.5 embryo from confocal imaging of immunofluorescencefor laminin (green) and muscle precursors (red, stained for Pax3 and Pax7; A–F), and schematicrepresentation of inside view of these segments (G–I). For all images, dorsal is up and medial is tothe left. A–C: Outside views are latero-transverse views. D–I: Inside views and schematic repre-sentations are transverse views. All three segments are separated by one segment each; thus themost mature segment (C,F,I) is situated four segments rostral to the least mature one (A,D,G). Thisyoungest segment represents DMM stage 4. A,D,G: The laminin matrix of the persisting epaxialdermomyotomal lip at DMM stage 4 lines the epithelial Pax3/7-positive cells. The end of DMMstage 4 (B,E,H followed by C,F,I) is marked by the progressive disassembly of the laminin matrixand the simultaneous de-epithelialization and release of the Pax3/7-positive cells. DMM, dermo-myotome/myotome. Scale bars ¼ 100 mm.
ECM 3D ORGANIZATION AND MYOGENESIS 359
Dev
elop
men
tal D
ynam
ics
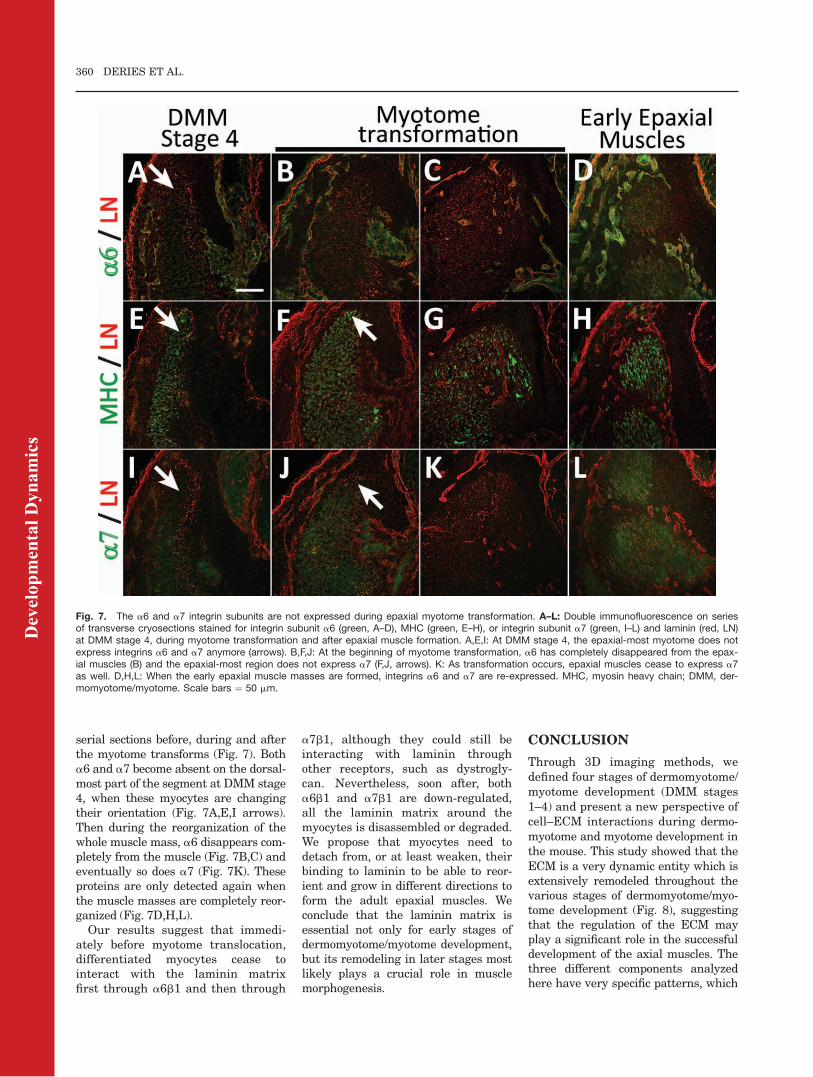
serial sections before, during and afterthe myotome transforms (Fig. 7). Botha6 and a7 become absent on the dorsal-most part of the segment at DMM stage4, when these myocytes are changingtheir orientation (Fig. 7A,E,I arrows).Then during the reorganization of thewhole muscle mass, a6 disappears com-pletely from the muscle (Fig. 7B,C) andeventually so does a7 (Fig. 7K). Theseproteins are only detected again whenthe muscle masses are completely reor-ganized (Fig. 7D,H,L).
Our results suggest that immedi-ately before myotome translocation,differentiated myocytes cease tointeract with the laminin matrixfirst through a6b1 and then through
a7b1, although they could still beinteracting with laminin throughother receptors, such as dystrogly-can. Nevertheless, soon after, botha6b1 and a7b1 are down-regulated,all the laminin matrix around themyocytes is disassembled or degraded.We propose that myocytes need todetach from, or at least weaken, theirbinding to laminin to be able to reor-ient and grow in different directions toform the adult epaxial muscles. Weconclude that the laminin matrix isessential not only for early stages ofdermomyotome/myotome development,but its remodeling in later stages mostlikely plays a crucial role in musclemorphogenesis.
CONCLUSION
Through 3D imaging methods, wedefined four stages of dermomyotome/myotome development (DMM stages1–4) and present a new perspective ofcell–ECM interactions during dermo-myotome and myotome development inthe mouse. This study showed that theECM is a very dynamic entity which isextensively remodeled throughout thevarious stages of dermomyotome/myo-tome development (Fig. 8), suggestingthat the regulation of the ECM mayplay a significant role in the successfuldevelopment of the axial muscles. Thethree different components analyzedhere have very specific patterns, which
Fig. 7. The a6 and a7 integrin subunits are not expressed during epaxial myotome transformation. A–L: Double immunofluorescence on seriesof transverse cryosections stained for integrin subunit a6 (green, A–D), MHC (green, E–H), or integrin subunit a7 (green, I–L) and laminin (red, LN)at DMM stage 4, during myotome transformation and after epaxial muscle formation. A,E,I: At DMM stage 4, the epaxial-most myotome does notexpress integrins a6 and a7 anymore (arrows). B,F,J: At the beginning of myotome transformation, a6 has completely disappeared from the epax-ial muscles (B) and the epaxial-most region does not express a7 (F,J, arrows). K: As transformation occurs, epaxial muscles cease to express a7as well. D,H,L: When the early epaxial muscle masses are formed, integrins a6 and a7 are re-expressed. MHC, myosin heavy chain; DMM, der-momyotome/myotome. Scale bars ¼ 50 mm.
360 DERIES ET AL.

Dev
elop
men
tal D
ynam
ics
change during the stages studied.While fibronectin is abundant in all in-terstitial tissues, including interseg-mental borders which are transientmyotome tendon-like structures, tenas-cin is restricted to these sites. Fibronec-tin and tenascin bundles are also in agood position to promote alignment ofmyocytes and guide the entry of muscle
precursors in the myotome. Lamininbasement membranes undergo organ-ized dynamic changes during dermo-myotome and myotome developmentwithout ever becoming continuous, pos-sibly because of the transient nature ofthese embryonic tissues. Our resultssuggest that during early epaxial mor-phogenesis, myocytes weaken the bind-
ing to laminin and might use fibronec-tin matrix as a substrate to acquiretheir final orientation.Unraveling the exact architecture
of the ECM environment duringmyotome development, raises newhypotheses regarding the developmen-tal roles for both the interstitial ECMcomponents fibronectin and tenascin,
Fig. 8. Model of the laminin, fibronectin, and tenascin matrices distribution pattern during DMM stages 1–4. This scheme summarizes the distributionpatterns of laminin (purple), fibronectin (green), and tenascin (light blue) at the four DMM stages analyzed in this study. For clarity, the intersegmental bor-der is only shown on the caudal border of each segment and the fibronectin present on the lateral side of the dermomyotome (between dermomyotomeand ectoderm) is not drawn. A: At DMM stage 1, a discontinuous laminin basement membrane lines the dermomyotome and the myoblasts entering themyotome are assembling laminin on the medial side of the myotome. Fibronectin and tenascin matrices are present in the intersegmental border(although tenascin is not abundant at this stage) and on the medial side of the myotome. B: At DMM stage 2, the laminin basement membranes haveexpanded with the growth of the dermomyotome and the myotome, but are still discontinuous. The tenascin matrix is now thicker at the intersegmentalborders and along the myocytes. The fibronectin matrix is abundant at the intersegmental borders and on the medial side of the myotome and a few fi-bronectin bundles are visible between the dermomyotome and the myotome. C: At DMM stage 3, the laminin basement membranes have reached theirdensest state but are still discontinuous. The tenascin matrix has grown in thickness and in number of bundles. The fibronectin matrix is even denserthan at the previous stage and is now clearly present between the dermomyotome and the myotome and generally runs along the elongated myocytes.D: At DMM stage 4, both laminin basement membranes have undergone a dramatic disassembly. The only basement membrane visible is lining theepaxial lip of the dermomyotome, the last remnant of the epithelial dermomyotome. A few spots of laminin are seen within the muscle mass. The tenascinmatrix has the same pattern as that of the earlier stages, but is denser. The fibronectin matrix is now even more abundant, filling all the spaces aroundand between the myocytes. The illustrations are schematic and not to scale with each other. DMM, dermomyotome/myotome; C, caudal; D, dorsal; L,lateral; M, medial; R, rostral; V, ventral; NT, neural tube; MHC, myosin heavy chain; MRFs, myogenic regulatory factors.
ECM 3D ORGANIZATION AND MYOGENESIS 361

Dev
elop
men
tal D
ynam
ics
as well as the laminins of basementmembranes. With the knowledgegained in this study, we are now in aposition to design experiments to testthese hypotheses.
EXPERIMENTAL
PROCEDURES
Embryo Collection
Dated pregnancies were obtained fromCharles River (CD1) mice (Harlan Iber-ica) and the day of the plug was E0.5.Embryos were collected from E9.5 toE13.5 after cervical dislocation of thefemale (project approved by DireccaoGeral de Veterinaria, Portugal).
Whole-Mount
Immunohistochemistry
Whole-mount immunohistochemistrywas performed as described in Derieset al., 2008. Briefly, embryos were fixedin 2% buffered paraformaldehyde, thenwashed and dehydrated in methanol.They were permeabilized with a solu-tion of dimethyl sulfoxide (DMSO) andmethanol (1:4) for 3 weeks. Then theywere stored in methanol 100% at�20�C until use. After rehydration,embryos were dissected as appropriateand processed for immunohistochemi-cal staining. Samples were incubatedwith primary antibodies (Table 1) inphosphate buffered saline (PBS) con-
taining 1% bovine serum albumin(BSA; Sigma) and 20% DMSO over-night at 4�C and were extensivelywashed in PBS during 2 days. Co-immunohistochemistry experimentswere performed to stain fibronectin,tenascin or laminins together withmyogenic cells. At early stages myo-genic cells were detected by a ‘‘cocktail’’of two mouse monoclonal antibodies:anti-myogenin (marking nuclei of dif-ferentiating cells) and anti-embryonicmyosin heavy chain (MHC, markingcytoplasm of fully differentiated myo-cytes). At later stages, only the anti-MHC antibody was used to detect myo-genic cells. Application of the secondaryantibodies (Table 1) was done as for theprimary antibodies. After immunolab-eling, samples were dehydrated inmethanol, cleared in methyl salicylate(Martins et al., 2009) and mountedbetween two coverslips sealed with par-affin. This mounting allowed us to viewthe thick specimens from two sides.Samples were imaged in a Leica SPEconfocal microscope (see below).
Immunohistochemistry on
Cryosections
Fluorescent immunohistochemistryon 12- or 30-mm cryosections was per-formed as in Bajanca et al. (2004).Embryos were fixed in 0.2% bufferedparaformaldehyde, gradually embed-ded in sucrose, followed by embedding
in gelatine and stored at �80�C. Aftersectioning in a Bright Clinicut cryo-stat, the samples were stained in pri-mary antibody overnight at 4�C andsecondary antibody for 2 hr at roomtemperature. Antibodies (see Table 1)were diluted in PBS containing 1%BSA. Sections were mounted in pro-pylgallate and viewed in a Leica SPEconfocal microscope.
Imaging and 3D
Reconstruction
To visualize the 3D matrix organiza-tion, confocal stacks were acquired ona Leica SPE confocal microscope witha �40 ACS APO 1.15 oil-immersionlens. From DMM stage 3 onward, thesegment spanned beyond the field ofthe �40 lens, therefore, the hypaxialregion of the myotome is not visible inthe images. The confocal stacks were3D reconstructed and analyzed usingthe Amira v5.3 software (Visage Inc).When appropriate, one individualsegment and its associated matrixwere isolated by manual contouring ofconfocal sections (i.e., other cells andthe matrix from surrounding tissueswere ‘‘digitally erased’’). The seg-ments are depicted in lateral, medialand transverse ‘‘3D views.’’
ACKNOWLEDGMENTSWe thank members of our group, inparticular Pedro Rifes, for helpful
TABLE 1. Antibodies Used
Antibodies Sourcea Dilution
Primary antibodies Monoclonal mouse anti-embryonic MHC, F59 DSHB 1:25Monoclonal mouse anti-myogenin, F5D Santa Cruz 1:100Monoclonal mouse anti-Pax3 DSHB 1:100Monoclonal mouse anti-Pax7 DSHB 1:100Monoclonal rat anti-tenascin-C antibody, LAT-2 van der Flier et al., 1997;
gift from A Sonnenberg1:50
Polyclonal rabbit anti-laminin 111, recognising alllaminins containing an a1, b1 or c1 chain, L-9393
Sigma 1:400
Polyclonal rabbit anti-fibronectin, F-3648 Sigma 1:400Polyclonal rabbit anti-av, CD51 Enzo 1:500Polyclonal rabbit anti-a9, H-198 Santa Cruz 1:50Monoclonal rat anti-integrin subunit a6, GoH3 Sonnenberg et al., 1988;
gift from A Sonnenberg1:25
Monoclonal rat anti-integrin subunit a7, CA5 Yao et al., 1996; gift fromA Sutherland
1:5
Secondary antibodies AlexafluorVR 488 /568 goat F(ab0)2 fragmentanti-mouse /rabbit IgG (HþL), as appropriate
Molecular Probes 1:1000
AlexafluorVR 488 goat whole antibody anti-rat IgG (HþL) Molecular Probes 1:1000
aDSHB, Developmental Studies Hybridoma Bank; MHC, myosin heavy chain.
362 DERIES ET AL.

Dev
elop
men
tal D
ynam
ics
discussions and Arnoud Sonnenbergand Ann Sutherland for their kind giftof antibodies (LAT-2/GoH3 and CA5).The F59, Pax3, and Pax7 antibodieswere developed by F.E. Stockdale, C.P.Ordahl, and A. Kawakami, respec-tively, and were obtained from theDevelopmental Studies HybridomaBank, developed under the auspices ofthe NICHD and maintained by TheUniversity of Iowa, Department ofBiology, Iowa City, IA52242, USA.This work, R.V., and M.D. were sup-ported by Fundacao para a Ciencia e aTecnologia (Portugal). A.B.G. is a stu-dent of the Masters in EvolutionaryandDevelopmental Biology (FCUL).
REFERENCES
Anderson C, Thorsteinsdottir S, BoryckiAG. 2009. Sonic hedgehog-dependentsynthesis of laminin a1 controls base-ment membrane assembly in the myo-tome. Development 136:3495–3504.
Bajanca F, Luz M, Duxson MJ, Thorsteins-dottir S. 2004. Integrins in the mousemyotome: developmental changes anddifferences between the epaxial andhypaxial lineage. Dev Dyn 231:402–415.
Bajanca F, LuzM, Raymond K, Martins GG,Sonnenberg A, Tajbakhsh S, BuckinghamM, Thorsteinsdottir S. 2006. Integrin a6b1-laminin interactions regulate earlymyotome formation in the mouse embryo.Development 133:1635–1644.
Barczyk M, Carracedo S, Gullberg D. 2010.Integrins. Cell Tissue Res 339:269–280.
Ben-Yair R, Kalcheim C. 2005. Lineageanalysis of the avian dermomyotomesheet reveals the existence of single cellswith both dermal and muscle progenitorfates. Development 132:689–701.
Blaschuk KL, Gu�erin C, Holland PC. 1997.Myoblast a v b3 integrin levels are con-trolled by transcriptional regulation ofexpression of the b 3 subunit and down-regulation of b 3 subunit expression isrequired for skeletal muscle cell differen-tiation. Dev Biol 184:266–277.
Buckingham M. 2006. Myogenic progenitorcells and skeletal myogenesis in verte-brates. Curr Opin Genet Dev 16:525–532.
Buckingham M, Bajard L, Chang T, Dau-bas P, Hadchouel J, Meilhac S, Montar-ras D, Rocancourt D, Relaix F. 2003.The formation of skeletal muscle: fromsomite to limb. J Anat 202:59–68.
Cachaco AS, Chuva de Sousa Lopes SM,Kuikman I, Bajanca F, Abe K, Baudoin C,Sonnenberg A, Mummery CL, Thorsteins-dottir S. 2003. Knock-in of integrin b 1Daffects primary but not secondary myogen-esis in mice. Development 130:1659–1671.
Cachaco AS, Pereira CS, Pardal RG,Bajanca F, Thorsteinsdottir S. 2005.Integrin repertoire on myogenic cellschanges during the course of primarymyogenesis in the mouse. Dev Dyn 232:1069–1078.
Calve S, Odelberg SJ, Simon HG. 2010. Atransitional extracellular matrixinstructs cell behavior during muscleregeneration. Dev Biol 344:259–271.
Chiquet-EhrismannR, Tucker RP. 2004. Con-nective tissues: signalling by tenascins. IntJ BiochemCell Biol 36:1085–1089.
Christ B, JacobM, Jacob HJ. 1983. On the or-igin and development of the ventrolateralabdominal muscles in the avian embryo.An experimental and ultrastructural study.Anat Embryol (Berl) 166:87–101.
Christ B, Huang R, Scaal M. 2007.Amniote somite derivatives. Dev Dyn236:2382–2396.
Crossin KL, Hoffman S, Grumet M,Thiery JP, Edelman GM. 1986. Site-restricted expression of cytotactin dur-ing development of the chicken embryo.J Cell Biol 102:1917–1930.
Deries M, Collins JJ, Duxson MJ. 2008.The mammalian myotome: a muscle withno innervation. Evol Dev 10:746–755.
Deries M, Schweitzer R, Duxson MJ. 2010.Developmental fate of the mammalianmyotome. Dev Dyn 239:2898–2910.
Duband JL, Dufour S, Hatta K, Takeichi M,Edelman GM, Thiery JP. 1987. Adhesionmolecules during somitogenesis in theavian embryo. J Cell Biol 104:1361–1374.
Duong TD, Erickson CA. 2004. MMP-2plays an essential role in producing epi-thelial-mesenchymal transformations inthe avian embryo. Dev Dyn 229:42–53.
Durbeej M. 2010. Laminins. Cell TissueRes 339:259–268.
Evans DJ, Valasek P, Schmidt C, Patel K.2006. Skeletal muscle translocation in ver-tebrates. Anat Embryol (Berl) 211(suppl1):43–50.
Forsberg E, Hirsch E, Frohlich L, MeyerM, Ekblom P, Aszodi A, Werner S, Fass-ler R. 1996. Skin wounds and severednerves heal normally in mice lackingtenascin-C. Proc Natl Acad Sci U S A 93:6594–6599.
Frantz C, Stewart KM, Weaver VM. 2010.The extracellular matrix at a glance.J Cell Sci 123:4195–4200.
Garcion E, Faissner A, ffrench-ConstantC. 2001. Knockout mice reveal a con-tribution of the extracellular matrixmolecule tenascin-C to neural precursorproliferation and migration. Develop-ment 128:2485–2496.
George EL, Georges-Labouesse EN, Patel-King RS, Rayburn H, Hynes RO. 1993.Defects in mesoderm, neural tube andvascular development in mouseembryos lacking fibronectin. Develop-ment 119:1079–1091.
Georges-Labouesse EN, George EL, Ray-burn H, Hynes RO. 1996. Mesodermaldevelopment in mouse embryos mutantfor fibronectin. Dev Dyn 207:145–156.
Gros J, Scaal M, Marcelle C. 2004. A two-step mechanism for myotome formationin chick. Dev Cell 6:875–882.
Gros J, Manceau M, Thome V, MarcelleC. 2005. A common somitic origin forembryonic muscle progenitors and sat-ellite cells. Nature 435:954–958.
Gros J, Serralbo O, Marcelle C. 2009.WNT11 acts as a directional cue to
organize the elongation of early musclefibres. Nature 457:589–593.
Guo LT, Zhang XU, Kuang W, Xu H, LiuLA, Vilquin JT, Miyagoe-Suzuki Y,Takeda S, Ruegg MA, Wewer UM, Eng-vall E. 2003. Laminin a2 deficiency andmuscular dystrophy; genotype-pheno-type correlation in mutant mice. Neuro-muscul Disord 13:207–215.
Hirsch E, Gullberg D, Balzac F, AltrudaF, Silengo L, Tarone G. 1994. a v integ-rin subunit is predominantly located innervous tissue and skeletal muscle dur-ing mouse development. Dev Dyn 201:108–120.
Hollway G, Currie P. 2005. Vertebratemyotome development. Birth DefectsRes C Embryo Today 75:172–179.
Huh MS, Smid JK, Rudnicki MA. 2005.Muscle function and dysfunction inhealth and disease. Birth Defects Res CEmbryo Today 75:180–192.
Jones PL, Jones FS. 2000. Tenascin-C indevelopment and disease: gene regula-tion and cell function. Matrix Biol 19:581–596.
Kalcheim C, Ben-Yair R. 2005. Cell rear-rangements during development of thesomite and its derivatives. Curr OpinGenet Dev 15:371–380.
Kalcheim C, Cinnamon Y, Kahane N.1999. Myotome formation: a multistageprocess. Cell Tissue Res 296:161–173.
Kassar-Duchossoy L, Giacone E, Gayraud-Morel B, Jory A, Gomes D, Tajbakhsh S.2005. Pax3/Pax7 mark a novel popula-tion of primitive myogenic cells duringdevelopment. Genes Dev 19:1426–1431.
Martins GG, Rifes P, Amandio R, Cam-pinho P, Palmeirim I, ThorsteinsdottirS. 2007. 3D visualization and analysisof cell-matrix transformations in whole-mount and live embryos using confocaland multi-photon microscopy. In: Men-dez-Vilas A, Diaz J, editors. Modernresearch and educational topics inmicroscopy #3. Vol. I. Spain: Formatex.p 426–433.
Martins GG, Rifes P, Amandio R, RodriguesG, Palmeirim I, Thorsteinsdottir S. 2009.Dynamic 3D cell rearrangements guidedby a fibronectin matrix underlie somito-genesis. PLoS ONE 4:e7429.
Mathew SJ, Hansen JM, Merrell AJ,Murphy MM, Lawson JA, HutchesonDA, Hansen MS, Angus-Hill M, KardonG. 2011. Connective tissue fibroblastsand Tcf4 regulate myogenesis. Develop-ment 138:371–384.
Miner JH, Yurchenco PD. 2004. Lamininfunctions in tissue morphogenesis.Annu Rev Cell Dev Biol 20:255–284.
Ostrovsky D, Sanger JW, Lash JW. 1988.Somitogenesis in the mouse embryo.Cell Differ 23:17–25.
Relaix F, Rocancourt D, Mansouri A,Buckingham M. 2005. A Pax3/Pax7-de-pendent population of skeletal muscleprogenitor cells. Nature 435:948–953.
Rifes P, Carvalho L, Lopes C, AndradeRP, Rodrigues G, Palmeirim I, Thor-steinsdottir S. 2007. Redefining the roleof ectoderm in somitogenesis: a playerin the formation of the fibronectin
ECM 3D ORGANIZATION AND MYOGENESIS 363

Dev
elop
men
tal D
ynam
ics
matrix of presomitic mesoderm. Devel-opment 134:3155–3165.
Rios AC, Serralbo O, Salgado D, MarcelleC. 2011. Neural crest regulates myogen-esis through the transient activation ofNOTCH. Nature 473:532–535.
Riou JF, Umbhauer M, Shi DL, BoucautJC. 1992. Tenascin: a potential modula-tor of cell-extracellular matrix interac-tions during vertebrate embryogenesis.Biol Cell 75:1–9.
Rozario T, DeSimone DW. 2010. Theextracellular matrix in developmentand morphogenesis: a dynamic view.Dev Biol 341:126–140.
Saga Y, Yagi T, Ikawa Y, Sakakura T, AizawaS. 1992. Mice develop normally withouttenascin. Genes Dev 6:1821–1831.
Schwander M, Leu M, Stumm M, Dor-chies OM, Ruegg UT, Schittny J, MullerU. 2003. b1 integrins regulate myoblastfusion and sarcomere assembly. DevCell 4:673–685.
Seale P, Bjork B, Yang W, Kajimura S,Chin S, Kuang S, Scime A, Devara-konda S, Conroe HM, Erdjument-Brom-age H, Tempst P, Rudnicki MA, BeierDR, Spiegelman BM. 2008. PRDM16controls a brown fat/skeletal muscleswitch. Nature 454:961–967.
Seo KW, Wang Y, Kokubo H, KettlewellJR, Zarkower DA, Johnson RL. 2006.Targeted disruption of the DM domaincontaining transcription factor Dmrt2reveals an essential role in somite pat-terning. Dev Biol 290:200–210.
Smyth N, Vatansever HS, Murray P,Meyer M, Frie C, Paulsson M, Edgar D.1999. Absence of basement membranesafter targeting the LAMC1 gene results
in embryonic lethality due to failure ofendoderm differentiation. J Cell Biol144:151–160.
Snow CJ, Peterson MT, Khalil A, HenryCA. 2008. Muscle development is dis-rupted in zebrafish embryos deficient forfibronectin. Dev Dyn 237:2542–2553.
Sonnenberg A, Modderman PW, HogervorstF. 1988. Laminin receptor on platelets isthe integrin VLA-6. Nature 336:487–489.
Tajbakhsh S, Buckingham M. 2000. Thebirth of muscle progenitor cells in themouse: spatiotemporal considerations.Curr Top Dev Biol 48:225–268.
Tajbakhsh S, Bober E, Babinet C, Pour-nin S, Arnold H, Buckingham M. 1996.Gene targeting the myf-5 locus withnlacZ reveals expression of this myo-genic factor in mature skeletal musclefibres as well as early embryonic mus-cle. Dev Dyn 206:291–300.
Taverna D, Disatnik MH, Rayburn H,Bronson RT, Yang J, Rando TA, HynesRO. 1998. Dystrophic muscle in micechimeric for expression of a5 integrin.J Cell Biol 143:849–859.
Thorsteinsdottir S, Deries M, CachacoAS, Bajanca F. 2011. The extracellularmatrix dimension of skeletal muscle de-velopment. Dev Biol 354:191–207.
Tosney KW, Dehnbostel DB, EricksonCA. 1994.Neural crest cells prefer themyo-tome’s basal lamina over the sclerotome asa substratum.DevBiol 163:389–406.
Tremblay P, Dietrich S, Mericskay M,Schubert FR, Li Z, Paulin D. 1998. Acrucial role for Pax3 in the developmentof the hypaxial musculature and thelong-range migration of muscle precur-sors. Dev Biol 203:49–61.
Vallois HV. 1922. Les transformations dela musculature de l’�episome chez lesvert�ebr�es. Paris: Facult�e des sciencesde l’universit�e de Paris. 521 p.
van der Flier A, Gaspar AC, Thorsteins-dottir S, Baudoin C, Groeneveld E,Mummery CL, Sonnenberg A. 1997.Spatial and temporal expression of thea1D integrin during mouse develop-ment. Dev Dyn 210:472–486.
van der Flier A, Sonnenberg A. 2001.Function and interactions of integrins.Cell Tissue Res 305:285–298.
Venters SJ, Thorsteinsdottir S, Duxson MJ.1999. Early development of the myotomein the mouse. Dev Dyn 216:219–232.
Wang A, Patrone L, McDonald JA, Shep-pard D. 1995. Expression of the integ-rin subunit a 9 in the murine embryo.Dev Dyn 204:421–431.
Wilson-Rawls J, Hurt CR, Parsons SM,Rawls A. 1999. Differential regulationof epaxial and hypaxial muscle devel-opment by paraxis. Development 126:5217–5229.
Yang JT, Rando TA, Mohler WA, RayburnH, Blau HM, Hynes RO. 1996. Geneticanalysis of a 4 integrin functions in thedevelopment of mouse skeletal muscle. JCell Biol 135:829–835.
Yao CC, Ziober BL, Sutherland AE,Mendrick DL, Kramer RH. 1996. Lami-nins promote the locomotion of skeletalmyoblasts via the alpha 7 integrinreceptor. J Cell Sci 109(pt 13):3139–3150.
Yurchenco PD, Patton BL. 2009. Develop-mental and pathogenic mechanisms ofbasement membrane assembly. CurrPharmDes 15:1277–1294.
364 DERIES ET AL.