
Expression of human ANT2 gene in highly proliferative cells: GRBOX, a new transcriptional element,...
10
Expression of Human ANT2 Gene in Highly Proliferative Cells: GRBOX, a New Transcriptional Element, is Involved in the Regulation of Glycolytic ATP Import into Mitochondria Ste ´ phane Giraud, Christelle Bonod-Bidaud Micheline Wesolowski-Louvel and Georges Stepien* Centre de Ge ´ne ´tique Mole ´culaire et Cellulaire CNRS UMR-5534, Universite ´ Claude Bernard Lyon1 43 Bd du 11 Novembre 1918 69622 Villeurbanne Cedex France The adenine nucleotide translocator (ANT) is the most abundant mito- chondrial inner membrane protein which catalyses the exchange of ADP and ATP between cytosol and mitochondria. The human ANT protein has three isoforms encoded by three differentially regulated nuclear genes. The ANT gene expression was examined in several human cells. The gene encoding the ANT2 isoform was found specifically induced in Simian virus 40 (SV40)-transformed, tumoral and mtDNA lacking rho cell lines. Moreover, the ANT2 gene was preferentially expressed under a glycolytic metabolism. Functional complementation of a Saccharomyces cerevisiae mutant revealed that the human ANT2 protein specifically restores yeast cell growth under anaerobic conditions. Sequence analysis of the ANT2 proximal promoter in comparison to that of the third yeast adenine nucleotide translocator (AAC3) led us to identify a new motif termed GRBOX. Promoter-deletion transfection and mobility gel-shift assays revealed that this motif is recognized by a negative transcriptional regulator. This transcription factor might be involved in a molecular mechanism which selects the import of the glycolytic ATP in the mito- chondrial matrix. This ATP import is required in highly proliferative cells, such as tumour cells, which depend strongly on glycolysis for ATP synthesis. # 1998 Academic Press Keywords: cancer cell; mitochondria; ANT2; gene regulation; ATP *Corresponding author Introduction In eukaryotic cells, ATP production can vary substantially in different tissues. This results from the differential expression of nuclear genes involved in oxidative phosphorylations. Moreover, it was proposed that this variation of ATP pro- duction is mainly controlled by the kinetics of the ADP and ATP exchange mediated by the adenine nucleotide translocator (ANT). ANT is the most abundant protein in mitochondria, embedded in the inner membrane, where it links the mitochon- drial ATP production to cellular ATP consumption (Klingenberg, 1993). The human nuclear ANT protein has three iso- forms encoded by three distinct genes: ANT1 (Cozens et al., 1989; Li et al., 1989), ANT2 (Ku et al., 1990) and ANT3 (Cozens et al., 1989). ANT1 is primarily expressed in heart and skeletal muscle, whereas ANT3 is ubiquitously expressed. In contrast, ANT2 is either not or weakly expressed in all human tissues studied (Stepien et al., 1992). The ANT2 mRNA levels are high in neoplastically transformed cells with high glyco- lytic rates (Torroni et al., 1990), in tumours and tumoral cells (Faure-Vigny et al., 1996; Heddi et al., 1996), as well as in early log phase cell E-mail address of the corresponding author: [email protected] Abbreviations used: AAC, ATP/ADP carrier; ANT, adenine nucleotide translocator; mtDNA, mitochondrial DNA; CAT, chloramphenicol acetyltransferase; GRBOX, glycolysis regulated box; D-Glc, D-glucose; L-Gln, L-glutamine; OXPHOS, oxidative phoshorylation; RT-PCR, reverse trancription-polymerase chain reaction; MAPS, multiple antigen peptide system; PBS, phosphate-buffered saline; ELISA, enzyme-linked immunosorbent assay; PMSF, phenylmethylsulfonyl fluoride. Article No. mb981955 J. Mol. Biol. (1998) 281, 409–418 0022 – 2836/98/330409–10 $30.00/0 # 1998 Academic Press
-
Upload
centreleonberard -
Category
Documents
-
view
5 -
download
0
Transcript of Expression of human ANT2 gene in highly proliferative cells: GRBOX, a new transcriptional element,...

Article No. mb981955 J. Mol. Biol. (1998) 281, 409±418
Expression of Human ANT2 Gene in HighlyProliferative Cells: GRBOX, a New TranscriptionalElement, is Involved in the Regulation of GlycolyticATP Import into Mitochondria
SteÂphane Giraud, Christelle Bonod-BidaudMicheline Wesolowski-Louvel and Georges Stepien*
Centre de GeÂneÂtiqueMoleÂculaire et CellulaireCNRS UMR-5534, UniversiteÂClaude Bernard Lyon143 Bd du 11 Novembre 191869622 Villeurbanne CedexFrance
E-mail address of the [email protected]
Abbreviations used: AAC, ATP/adenine nucleotide translocator; mDNA; CAT, chloramphenicol acetyglycolysis regulated box; D-Glc, D-L-glutamine; OXPHOS, oxidative pRT-PCR, reverse trancription-polyMAPS, multiple antigen peptide sphosphate-buffered saline; ELISA,immunosorbent assay; PMSF, phe¯uoride.
0022±2836/98/330409±10 $30.00/0
The adenine nucleotide translocator (ANT) is the most abundant mito-chondrial inner membrane protein which catalyses the exchange of ADPand ATP between cytosol and mitochondria. The human ANT proteinhas three isoforms encoded by three differentially regulated nucleargenes. The ANT gene expression was examined in several human cells.The gene encoding the ANT2 isoform was found speci®cally induced inSimian virus 40 (SV40)-transformed, tumoral and mtDNA lacking rho�cell lines. Moreover, the ANT2 gene was preferentially expressed under aglycolytic metabolism. Functional complementation of a Saccharomycescerevisiae mutant revealed that the human ANT2 protein speci®callyrestores yeast cell growth under anaerobic conditions. Sequence analysisof the ANT2 proximal promoter in comparison to that of the third yeastadenine nucleotide translocator (AAC3) led us to identify a new motiftermed GRBOX. Promoter-deletion transfection and mobility gel-shiftassays revealed that this motif is recognized by a negative transcriptionalregulator. This transcription factor might be involved in a molecularmechanism which selects the import of the glycolytic ATP in the mito-chondrial matrix. This ATP import is required in highly proliferativecells, such as tumour cells, which depend strongly on glycolysis for ATPsynthesis.
# 1998 Academic Press
Keywords: cancer cell; mitochondria; ANT2; gene regulation; ATP
*Corresponding authorIntroduction
In eukaryotic cells, ATP production can varysubstantially in different tissues. This results fromthe differential expression of nuclear genesinvolved in oxidative phosphorylations. Moreover,it was proposed that this variation of ATP pro-
ding author:
ADP carrier; ANT,tDNA, mitochondrialltransferase; GRBOX,
glucose; L-Gln,hoshorylation;
merase chain reaction;ystem; PBS,enzyme-linked
nylmethylsulfonyl
duction is mainly controlled by the kinetics of theADP and ATP exchange mediated by the adeninenucleotide translocator (ANT). ANT is the mostabundant protein in mitochondria, embedded inthe inner membrane, where it links the mitochon-drial ATP production to cellular ATP consumption(Klingenberg, 1993).
The human nuclear ANT protein has three iso-forms encoded by three distinct genes: ANT1(Cozens et al., 1989; Li et al., 1989), ANT2 (Kuet al., 1990) and ANT3 (Cozens et al., 1989). ANT1is primarily expressed in heart and skeletalmuscle, whereas ANT3 is ubiquitously expressed.In contrast, ANT2 is either not or weaklyexpressed in all human tissues studied (Stepienet al., 1992). The ANT2 mRNA levels are high inneoplastically transformed cells with high glyco-lytic rates (Torroni et al., 1990), in tumours andtumoral cells (Faure-Vigny et al., 1996; Heddiet al., 1996), as well as in early log phase cell
# 1998 Academic Press

Figure 1. Analysis of ANT2 gene expression in humancell lines. Total RNA samples (10 mg/lane) from cul-tured cells were blotted and successively hybridizedwith speci®c [a-32P]dCTP-labelled ANT2 and 18S rRNAprobes. WI-38, human lung cells; WI-38SV40, SV40-transformed derivatives of WI-38; CAKI 1, renal tumoralcell line; rho� 143B, human osteosarcoma lackingmtDNA.
410 Regulation of the ANT2 Gene Expression
cultures such as myoblasts (Stepien et al., 1992).Finally, ANT2 is induced in rapidly dividing cellssuch as serum-stimulated ®broblasts (Battini et al.,1987). The ANT genes share three highly homolo-gous coding sequences but are made up of dis-tinct promoter sequences (Ku et al., 1990),supporting the view that the expression of ANTgenes is under distinct regulatory controls. TheANT2 gene is localized on the long arm of the Xchromosome, has at least seven pseudogenes(Chen et al., 1990; Schiebel et al., 1994) and its 50-¯anking region contains sequence motifs (TATA,SP1 sites and enhancer-like segments: Ku et al.,1990) that are similar to those of other growth-related genes (Li et al., 1996).
For several years, rapidly growing cancer cellshave been shown to exhibit an abnormal energymetabolism, in comparison with normal mamma-lian cells which derive most of their energysupplies from oxidative phosphorylations. In con-trast, cancer cells can maintain an increased rate ofglucose utilization and sustain high rates of gly-colysis under aerobic conditions (Dickman &Mandel, 1989; Nakashima et al., 1984; Pedersen,1978). Therefore, tumoral cell growth may dependstrongly on glycolysis for ATP synthesis.
In yeast, the mitochondrial ADP/ATP transloca-tor exists, as in human, as three isoforms encodedby the AAC1, AAC2, and AAC3 genes (Adrian et al.,1986; Kolarov et al., 1990; Lawson & Douglas,1988). Aac1 and Aac2 proteins allow yeast cells togrow aerobically. On the other hand, Aac3p isessential for anaerobic growth of yeast cells (Drgonet al., 1991). The AAC3 gene is regulated by hemethrough the ROX1 factor (Sabova et al., 1993),which was shown to mediate the oxygen repres-sion of several other yeast genes (Lowry &Zitomer, 1988; Zitomer & Lowry, 1992; Zitomeret al., 1987).
Here, comparison of the ANT2 and AAC3 proxi-mal promoters reveals a ROX1-like binding motifin the ANT2 promoter region. This new motif, notfound in the two other human ANT or yeast AAC1and AAC2 gene promoters, was termed GRBOX(glycolysis-regulated box). The particular role ofthe human ANT2 isoform is demonstrated by itsability to restore anaerobic growth to a yeast triplenull mutant, in which the three AAC genes are dis-rupted. Furthermore, to characterize the involve-ment of GRBOX in the regulation of the ANT2gene, we performed promoter-reporter gene con-structs including or not the GRBOX sequence.Moreover, gel-shift assays were carried out with40 bp wild-type or mutant oligonucleotides inorder to characterize the sequence involved in theDNA-protein interactions.
As indicated in detail below, our experimentsshow that the ANT2 isoform should import theglycolytic ATP into mitochondria, and haverevealed a new motif implicated in ANT2 geneexpression.
Results
Expression of the ANT2 gene in transformed,tumoral and rho� cell lines
Total RNA samples from different exponentiallygrowing cell lines were subjected to electrophor-esis, blotted, and hybridized with ANT2 and 18SrRNA probes (Figure 1). The 18S rRNA probe wasused as an internal standard. The ANT2 probe wasa fragment encompassing the 30 930 to 1123 ntcDNA sequence. This probe exclusively hybridizesto the ANT2 isoform mRNA. The ANT2 gene wasnot expressed in the differentiated WI-38 ®bro-blasts as described (Stepien et al., 1992). On thecontrary, this gene was found highly expressed intransformed (WI-38SV40) and tumoral (CAKI 1;rho� 143B) cells. These results, in accordance withprevious reports, clearly showed the induction ofthe ANT2 gene transcription in highly proliferativecells (Battini et al., 1987; Torroni et al., 1990). More-over, our work demonstrates that this ANT2mRNA over expression was linked to a simul-taneous ANT2 protein accumulation (Figure 2).Total protein extracted from the same control,transformed and tumoral cell lines were blotted onnitrocellulose membrane and immuno-revealedwith a polyclonal antibody directed to the N-term-inal region of the ANT2 isoform. In the WI-38 con-trol cells, the faint signal might be due to cross-reactivity with the ubiquitously expressed ANT3isoform, the peptide sequences of the two ANT2and ANT3 isoforms sharing 70% of homology inthe N-terminal sequence.
Effect of metabolic conditions on ANT2 geneexpression in CAKI 1 cells
In order to investigate the effect of metabolicconditions on ANT2 gene expression, CAKI 1cells were cultured in media supplemented withD-Glc (glycolytic metabolism) or L-Gln (oxidativemetabolism). The oxidative or glycolytic path-
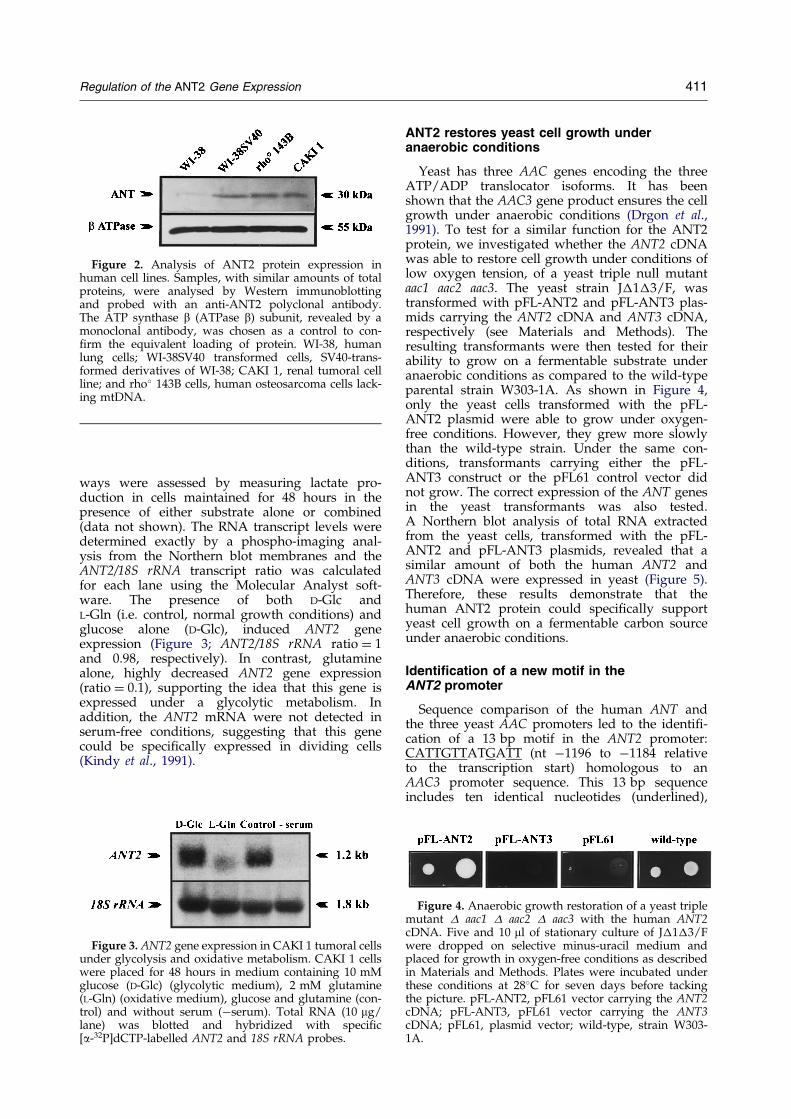
Figure 2. Analysis of ANT2 protein expression inhuman cell lines. Samples, with similar amounts of totalproteins, were analysed by Western immunoblottingand probed with an anti-ANT2 polyclonal antibody.The ATP synthase b (ATPase b) subunit, revealed by amonoclonal antibody, was chosen as a control to con-®rm the equivalent loading of protein. WI-38, humanlung cells; WI-38SV40 transformed cells, SV40-trans-formed derivatives of WI-38; CAKI 1, renal tumoral cellline; and rho� 143B cells, human osteosarcoma cells lack-ing mtDNA.
Regulation of the ANT2 Gene Expression 411
ways were assessed by measuring lactate pro-duction in cells maintained for 48 hours in thepresence of either substrate alone or combined(data not shown). The RNA transcript levels weredetermined exactly by a phospho-imaging anal-ysis from the Northern blot membranes and theANT2/18S rRNA transcript ratio was calculatedfor each lane using the Molecular Analyst soft-ware. The presence of both D-Glc andL-Gln (i.e. control, normal growth conditions) andglucose alone (D-Glc), induced ANT2 geneexpression (Figure 3; ANT2/18S rRNA ratio � 1and 0.98, respectively). In contrast, glutaminealone, highly decreased ANT2 gene expression(ratio � 0.1), supporting the idea that this gene isexpressed under a glycolytic metabolism. Inaddition, the ANT2 mRNA were not detected inserum-free conditions, suggesting that this genecould be speci®cally expressed in dividing cells(Kindy et al., 1991).
Figure 3. ANT2 gene expression in CAKI 1 tumoral cellsunder glycolysis and oxidative metabolism. CAKI 1 cellswere placed for 48 hours in medium containing 10 mMglucose (D-Glc) (glycolytic medium), 2 mM glutamine(L-Gln) (oxidative medium), glucose and glutamine (con-trol) and without serum (ÿserum). Total RNA (10 mg/lane) was blotted and hybridized with speci®c[a-32P]dCTP-labelled ANT2 and 18S rRNA probes.
ANT2 restores yeast cell growth underanaerobic conditions
Yeast has three AAC genes encoding the threeATP/ADP translocator isoforms. It has beenshown that the AAC3 gene product ensures the cellgrowth under anaerobic conditions (Drgon et al.,1991). To test for a similar function for the ANT2protein, we investigated whether the ANT2 cDNAwas able to restore cell growth under conditions oflow oxygen tension, of a yeast triple null mutantaac1 aac2 aac3. The yeast strain J�1�3/F, wastransformed with pFL-ANT2 and pFL-ANT3 plas-mids carrying the ANT2 cDNA and ANT3 cDNA,respectively (see Materials and Methods). Theresulting transformants were then tested for theirability to grow on a fermentable substrate underanaerobic conditions as compared to the wild-typeparental strain W303-1A. As shown in Figure 4,only the yeast cells transformed with the pFL-ANT2 plasmid were able to grow under oxygen-free conditions. However, they grew more slowlythan the wild-type strain. Under the same con-ditions, transformants carrying either the pFL-ANT3 construct or the pFL61 control vector didnot grow. The correct expression of the ANT genesin the yeast transformants was also tested.A Northern blot analysis of total RNA extractedfrom the yeast cells, transformed with the pFL-ANT2 and pFL-ANT3 plasmids, revealed that asimilar amount of both the human ANT2 andANT3 cDNA were expressed in yeast (Figure 5).Therefore, these results demonstrate that thehuman ANT2 protein could speci®cally supportyeast cell growth on a fermentable carbon sourceunder anaerobic conditions.
Identification of a new motif in theANT2 promoter
Sequence comparison of the human ANT andthe three yeast AAC promoters led to the identi®-cation of a 13 bp motif in the ANT2 promoter:CATTGTTATGATT (nt ÿ1196 to ÿ1184 relativeto the transcription start) homologous to anAAC3 promoter sequence. This 13 bp sequenceincludes ten identical nucleotides (underlined),
Figure 4. Anaerobic growth restoration of a yeast triplemutant � aac1 � aac2 � aac3 with the human ANT2cDNA. Five and 10 ml of stationary culture of J�1�3/Fwere dropped on selective minus-uracil medium andplaced for growth in oxygen-free conditions as describedin Materials and Methods. Plates were incubated underthese conditions at 28�C for seven days before tackingthe picture. pFL-ANT2, pFL61 vector carrying the ANT2cDNA; pFL-ANT3, pFL61 vector carrying the ANT3cDNA; pFL61, plasmid vector; wild-type, strain W303-1A.

Figure 5. Analysis of the human ANT2 and ANT3cDNA expression in the yeast triple mutant � aac1� aac2 � aac3. Total RNA samples (7 mg/lane) fromS. cerevisiae J�1�3/F cells, transfected with either pFL-ANT2 or pFL-ANT3 constructs, and from human 143BTK cells, were blotted and successively hybridized withspeci®c [a-32P]dCTP-labelled ANT2 and ANT3 probes.
412 Regulation of the ANT2 Gene Expression
with seven contiguous nucleotides (ÿ1196 toÿ1190), to the ROX1 element. This motif was notfound in the two other human ANT and yeastAAC1 and AAC2 gene promoters. It was termedGRBOX (for glycolysis regulated box) in accord-ance with the previously characterized ANT1transcriptional elements OXBOX-REBOX (Li et al.,1990; Chung et al., 1992). As a ®rst approach tostudy the involvement of this motif in ANT2gene regulation, we performed transfection del-etion promoter experiments using the CAT repor-ter gene. A construct with an 80 bp deletion(ÿ1237 to ÿ1158) in the human ANT2 promoterregion including the GRBOX sequence was usedto transfect WI-38SV40 transformed cells, CAKI 1adenocarcinoma cells, and rho� 143B cells(Figure 6). Cells were transfected with pCAT-GRBOX (wild-type fragment of the ANT2 promo-ter) or pCAT-�GRBOX (mutant construct) plas-mid in the presence of the Exgen 500 reagent. Asdescribed in Materials and Methods, data werecorrected by the CAT/b-Gal ratio, and the corre-
Figure 6. GRBOX ablation mutant analysis. Fragmentsfrom the 50-¯anking region of the human ANT2 gene (ntÿ1237 to �46 and nt ÿ1158 to �46), cloned ahead ofthe CAT reporter gene in the pCATbasic (Promega),were transfected for 24 hours into human WI-38SV40,CAKI 1 and rho� 143B cell lines. Arrows, transcriptionstart (�1). The level of CAT enzyme was normalized tothe values obtained with the ÿ1237 to �46 fragment ineach cell line. All experimental measures were run induplicate. The mean standard error (bars) is given forthree separate experiments.
sponding ef®ciency was quanti®ed: 80% for trans-formed cells and 40% for the other cell lines(data not shown). If the two constructs are com-pared in each cell line, the results showed thatthe removal of the ÿ1237 to ÿ1158 nt (relative totranscription start) led to a two- to threefoldincrease of the CAT enzyme level. Moreover,when the same experiment was conducted withcells lacking the transcription factors allowing theexpression of the ANT2 episomic form (i.e. WI-38), the CAT enzyme level remained constant forthe two constructs (data not shown). This resultindicates that the 80 bp sequence of the ANT2promoter might include a novel regulatoryelement.
GRBOX binding factors
To investigate the presence of speci®c nuclearfactors which might interact with the GRBOXelement, a 40 bp DNA sequence from the ANT2promoter, including the GRBOX motif (Figure 7)was labelled, incubated with nuclear extracts pre-pared from control WI-38 ®broblasts and analysedby electrophoretic mobility shift assay. Two DNA-protein complexes, a and b, were detected in WI-38 ®broblasts where the ANT2 gene is notexpressed (Figure 8, lane 1). These complexes wereshown to be speci®c for the 40 bp sequence includ-ing the GRBOX motif according to the fact that a40 bp scrambled GRBOX sequence (lane 2) did notlead to the a and b complex formation. Further, todetermine the DNA sequence involved in recog-nition of the a and b complexes, mutant oligonu-cleotides were selected by random substitution ofseveral nucleotides of the ANT2 promoter sequence(Figure 7). Electrophoretic mobility shift assaysusing Mut1 and Mut2 showed that the 30 half partof the GRBOX motif and the 3' end of the probewere involved in the a complex formation(Figure 8): if corrected with the bound/free proberatio, Mut1 and Mut2 had a similar effect on the aband. In contrast, Mut3, Mut4 and Mut5 oligonu-cleotides did not lead to a and b complex for-mation (Figure 9, left). The absence of any bindingwith Mut5 (lane 6), covering the 13 nucleotides ofthe GRBOX motif, indicates that the core sequence(CATTGTT), identical to the yeast ROX1 sequence,was necessary for the formation of both complexes.
Figure 7. Oligonucleotides used in electrophoretic mobi-lity shift assays. The ®rst line shows the ANT2 promotersequence between nucleotides ÿ1214 to ÿ1171 relativeto the transcription start. Open box, GRBOX sequence;gray boxes, oligonucleotides carrying mutated nucleo-tides, Mut1, Mut2, Mut3, Mut4, Mut5, scrambled.

Figure 8. Binding of nuclear proteins to the GRBOXsequence. The gel mobility shift assay was run usingfour different labelled probes (sequences shown inFigure 6) and nuclear extracts from WI-38 ®broblasts.The binding reaction mixture contained 0.1 ng (105 cpm)of labelled DNA, 5 mg of nuclear proteins, 100 ng ofpoly(dI-dC) �poly(dI-dC) and 0.5% NP40. Lane 1, nativesequence without nuclear extract; lane 2, scrambledsequence; lane 3, native sequence; lane 4, Mut1; lane 5,Mut2.
Figure 10. Electrophoretic mobility shift assay patternsin mitochondrial DNA depleted (rho� 143B) cells andtumoral cells (CAKI 1). The binding reaction mixture isshown in Figure 7 and oligonucleotide sequences inFigure 6. Lane 1, native sequence without nuclearextract; lane 2, native sequence; lane 3, Mut3; lane 4,Mut4; lane 5, Mut5.
Regulation of the ANT2 Gene Expression 413
The binding of nuclear factors from transformedcells (WI-38SV40) to the GRBOX sequence wascompared to that from control WI-38 ®broblasts. Inthe transformed cells, complexes with the GRBOXnative sequence were weakly detectable (Figure 9).The a and b complexes were similarly revealedwhen the gel-shifts with tumoral and rho� cellswere overexposed (Figure 10). If parallelled to theinduction of the ANT2 gene expression in trans-formed (WI-38SV40), tumoral (CAKI 1) and rho�
Figure 9. Electrophoretic mobility shift assay patterns inWI-38 and WI-38SV40 transformed cells. The bindingreaction mixture is shown in Figure 7 and oligonucleo-tide sequences in Figure 6. Lane 1, native sequence with-out nuclear extract; lane 2, scrambled sequence; lane 3,native sequence; lane 4, Mut3; lane 5, Mut4; lane 6,Mut5.
143B cells, this experiment demonstrated that, inWI-38 control cells, the GRBOX motif was recog-nized by nuclear proteins which might act as nega-tive regulators.
Discussion
In eukaryotic cells, among all the nuclear-encoded protein subunits involved in the mito-chondrial oxidative phosphorylation (OXPHOS)process, several are tissue speci®cally or develop-mentally regulated. These mechanisms are believedto be necessary for tissue-speci®c energy require-ments. The ANT protein plays a central role inOXPHOS by exchanging ATP and ADP throughthe mitochondrial internal membrane. This ATP/ADP ¯ux is the last step of the mitochondrial oxi-dative phosphorylations and might be highly regu-lated at the level of gene expression.
The two isoforms expressed in differentiatedhuman tissues, ANT1 and ANT3, export the ATPsynthesized in the mitochondria to the cytosol. Onthe other hand, the ANT2 mRNA were initiallyfound highly expressed in transformed and tumor-al cells (Torroni et al., 1990; Faure-Vigny et al.,1996; Heddi et al., 1996) as well as in early logphase cell cultures such as myoblasts (Stepien et al.,1992). Also, ANT2 induction in highly proliferativecells might be directly linked to the particularmetabolism called ``aerobic glycolysis'' of thetumoral cells (Pedersen, 1978). Therefore, theANT2 protein may have kinetic properties whichcould realize an inverse exchange, translocating theglycolytic ATP, synthesized in the cytosol, to themitochondrial matrix. This ATP is essential to pro-vide energy for the intra-mitochondrial metabolicpathways and for the mtDNA replication andexpression.

414 Regulation of the ANT2 Gene Expression
Such an inverse ATP/ADP exchange is ®rst cor-related with the ANT2 transcript accumulation inrho� 143B cells, which is not detectable in controlhuman ®broblasts. This is the ®rst report of ANT2expression using a speci®c probe revealing exclu-sively the ANT2 mRNA. The rho� cells exhibitacidosis, uridine auxotrophy and have lost theirmitochondrial DNA (King & Attardi, 1989). Conse-quently, the cytosolic glycolysis is their exclusivesource of ATP. Nevertheless, these cells are able tomaintain an electrochemical gradient across themitochondrial membranes (Li et al., 1995). Thissuggests the presence, in rho� cells, of an adaptableresponse that preserves the structural integrity ofthe mitochondrial inner membrane as well as thetransmembrane electrochemical gradient, despitethe absence of all the protein components encodedby mtDNA. The electrochemical gradient shouldbe restored by a partial reverse running of the res-piratory chain through the glycolytic ATP hydroly-sis by the ATP synthase F1 complex. Therefore,ANT2 induction under glycolytic metabolism maybe a nuclear response of the cell to preserve mito-chondrial functions, even if oxidative phosphoryl-ations are completely impaired.
The hypothesis that ANT2 may be involved in acytoplasmic ATP/mitochondrial ADP inverseexchange was also con®rmed by experiments per-formed with the CAKI 1 cell line under differentmetabolic conditions: ANT2 gene expression wasinduced following the switch from oxidative toglycolytic pathways. On the contrary, the ANT2gene is neither expressed in an exclusive oxidativemetabolism nor in quiescent cells with low ener-getic requirements.
Considering the high amino acid sequence simi-larity between the three ANT isoforms, classicalpolyclonal antibodies do not allow the study of aspeci®c ANT isoform. Using the MAPS method-ology (multiple antigen peptide system), we havegenerated an ANT2 polyclonal antibody directedagainst a ten amino acid peptide from the N-term-inal sequence of the ANT2 protein. This shortsequence presents speci®c antigenic properties thatmight distinguish ANT2 from the two other iso-forms. Therefore, these immunological studies vali-dated for the ®rst time the previous geneexpression experiments which showed that theANT2 isoform is speci®cally expressed in trans-formed, tumoral and rho� cells.
Previous works in yeast con®rm the hypothesisof these particular kinetic properties of the ANT2isoform. In Saccharomyces cerevisiae, three ANT iso-forms, which are encoded by three different genes,have been also identi®ed: AAC1 (ATP/ADP Car-rier), AAC2 and AAC3 (Adrian et al., 1986; Kolarovet al., 1990; Lawson & Douglas, 1988). When thethree human and the three yeast proteins are com-pared, the yeast isoform encoded by AAC3 and thehuman isoform encoded by ANT2 are found themost related, with a percentage of identity of 53%.It is now clearly established that the yeast Aac3pisoform is required to sustain cell growth under
anaerobic conditions. It was also hypothesized thatthis protein provides glycolytic ATP to mitochon-dria (Visser et al., 1994). Here we show that a yeasttriple null mutant, where the three AAC geneswere disrupted, was able to grow under anaerobicconditions, when transformed with the ANT2cDNA construct. Alternatively, the ANT3 cDNAcould not restore yeast cell growth. Further, North-ern blot analysis showed that the anaerobicallygrowing yeast expresses the human ANT2 geneand that both ANT2 and ANT3 constructs wereexpressed in transformants growing under aerobicconditions. Thus, this functional complementationsuggests for the ®rst time that the ANT2 isoformcatalyses the inverse cytosolic ATP/mitochondrialADP exchange. This is corroborated by in vitroexperiments assessing that the ANT protein canrealize a speci®c ATP/ADP exchange through themitochondrial internal membrane in both direc-tions (KraÈmer & Klingenberg, 1980).
Under aerobic conditions, the speci®c ROX1 fac-tor binds to an AAC3 promoter motif and preventsAAC3 gene expression (Sabova et al., 1993). Wefound a similar motif, nt ÿ1196 to ÿ1184 upstreamof the transcription start (10 out of 13 with sevencontiguous nucleotides of the yeast motif recog-nized by ROX1), within the human ANT2 promo-ter sequence (Ku et al., 1990). This motif wastermed GRBOX for glycolysis regulated box.Through transient transfection studies in tumoraland transformed cells, a 80 nucleotide sequence(ÿ1237 to ÿ1158) including the GRBOX motif wasfound to encompass a regulatory element. Thetwo- to threefold increase in CAT enzyme level forthe construct, where this sequence is deleted, indi-cates the presence of a negative regulator of geneexpression.
Using gel mobility shift assays, the sequenceinvolved in the ANT2 regulation was con®rmedand de®ned more precisely. The experimentsrevealed a complex regulation in this area whereseveral regulatory proteins might be involved.A GRBOX ablation mutant completely preventsthe binding of speci®c proteins. These proteinshave a high presence in control ®broblasts andtheir binding is highly decreased in transformed,tumoral and rho� cells. According to the differentmutant oligonucleotides used in mobility shiftassay analysis, the GRBOX binding factor seems tointeract with other regulatory proteins which couldbind to GRBOX ¯anking sequences. Moreover,according to other promoter-deletion transfectionexperiments (Li et al., 1996), a complex proteininteraction with other putative downstream regu-latory elements could be involved in the ANT2regulation.
The expression from the ANT2 proximal promo-ter was previously found modulated by Sp1elements which are supposed to regulate thegrowth activation of the ANT2 gene (Li et al.,1996). The GRBOX cis-acting DNA element may bemore speci®c of OXPHOS regulation and may beconsidered a responsive element linked to the gly-

Regulation of the ANT2 Gene Expression 415
colytic metabolism of the highly proliferative cells.In S. cerevisiae, the responsive protein of the AAC3oxygen regulation is the ROX1 factor. This proteinis an HMG (high mobility group)-like DNA bind-ing protein (Balasubramanian et al., 1993) which isheme regulated (Zitomer et al., 1997; Zitomer &Lowry, 1992). Therefore, the ANT2 gene might be anew heme-regulated gene such as the well-knownerythropoietin human gene (Huang et al., 1997).
In conclusion, expression of the ANT2 gene intransformed, tumoral and rho� 143B cell cultures iscorrelated with the release of a factor binding tothe new speci®c GRBOX motif. This factor canmodulate the ANT2 transcript level during theswitch from oxidatif to glycolytic metabolism.Therefore, in this model, the GRBOX element maybe the ®nal molecular sensor determining the pathof the ATP/ADP exchange through the mitochon-drial internal membrane depending on the meta-bolic way of the cell.
Materials and Methods
Cell cultures
The human cell lines used were: WI-38 (human lung®broblasts, ATCC Rockville, MD CCL 75), WI-38SV40(WI-38SV40 T antigen transformed, ATCC, CCL 75.1),CAKI 1 (renal tumoral cell line ATCC HTB46), 143B TK(human osteosarcoma, ATCC CRL-8303) and rho� 143B(143BTK cell derivatives, a gift from Dr Morais, Univer-sity of Montreal, Quebec). All cells were cultured inmedium containing 10% (v/v) fetal calf serum (FCS),100 units/ml penicillin, 100 units/ml streptomycin. WI-38 and WI-38SV40 were cultured in Minimum EssentialMedium (MEM) supplemented with 2 mM glutamineand non-essential amino acids. CAKI 1 cells were cul-tured in RPMI supplemented with 2 mM glutamine. Forshort-term experiments CAKI 1 cells were placed inRPMI medium without L-Gln and D-Glc, supplementedwith either 2 mM L-Gln, 100 mM Ino, 40 mM dThd,100 mM Urd, or D-Glc 10 mM and 10% dialysed FCS(Goossens et al., 1996). 143B TK and rho� 143B cells werecultured in DMEM supplemented with 0.2 mM uridineand 1 mM sodium pyruvate. The cells were plated in100 mm dishes and maintained at 37�C in 5% (v/v) CO2
atmosphere. All products were from GIBCO, except ino-sitol, thymidine and uridine (Sigma).
Northern blot analysis
Total cellular RNA were prepared with the ``RNeasyTotal RNA'' kit from Qiagen (Qiagen S.A., France) fol-lowing the manufacturer's instructions. RNA samples(10 mg) were denatured by incubation at 55�C for20 minutes in 22% (v/v) glyoxal, 65% (v/v) dimethylsulfoxide, 13% (w/v) 100 mM NaH2PO4 (pH 7) and sub-jected to electrophoresis in 1.3% (w/v) agarose gel (ICN)containing 10 mM NaH2PO4 (pH 7) (Sambrook et al.,1992). RNA was then transferred to Hybond-N� nylonmembrane (Amersham Corp, UK). Hybridization wasperformed as described (Faure-Vigny et al., 1996). Thespeci®c ANT3 probe was a full-lenght cDNA cloned intopBluescript II KS� (Stratagene) in EcoRI/BamHI cloningsites (Stepien et al., 1992). The speci®c ANT2 and 18Sprobes were generated by PCR ampli®cation from 25 ng
of the corresponding cDNA. Primer pairs were as fol-lows: for targeting the 18S rRNA, 50 primer nt 1196 to1212 and 30 primer nt 1790 to 1806; for targeting theANT2 mRNA, 50 primer nt 930 to 949 and 30 primer nt1104 to 1123. Ampli®cation of the cDNA was performedas follows: 94�C, 30 seconds, 55�C, 30 seconds, 72�C,30 seconds in an Eppendorf Mastercycler 5330 for 30cycles. The ANT2 fragment (193 nt) was cloned into thepGEM-T plasmid vector (Promega). The probes (193 ntfor ANT2 and 610 nt for 18S rRNA) were gel-puri®edand [a-32P]dCTP-labelled by random-primer DNA label-ling. The Northern blots were prehybridized for sixhours at 50�C in a Hybaid oven in 5 � standard salinephosphate (PBS) with EDTA, 5 � Denhardt's solution,50% (v/v) formamide, 0.2% (w/v) SDS, and 0.05% (w/v) denatured ®sh-sperm DNA (Serva Biochemicals, Para-mus, NJ). Hybridizations were performed for 16 hours at42�C in the same buffer supplemented with labelledprobe. For the ANT2 probe, the blots were washed threetimes for ®ve minutes in 2 � standard saline citrate and0.1% SDS at 50�C. For the 18S rRNA probe, the blotswere washed three times for 30 minutes in 1 � standardsaline citrate and 0.1% SDS at 60�C. Autoradiographswere obtained by exposing Kodak ®lms (BIOMAX MR).
Gel electrophoresis and immunoblotting
Cellular protein extraction
A total of 1.5 � 106 cells for each cell line were grownin 100 mm cell culture dishes. Cells were harvested bytrypsinization and lysed by sonication. Total proteins(200 mg) of each sample were boiled in Laemmli bufferfor ®ve minutes (Laemmli, 1970). Samples were then runon a 15% (w/v) acrylamide/bisacrylamide gel accordingto the SDS-PAGE method. Proteins were then transferredonto a nitrocellulose membrane (Schleicher and Schull)and immunoblotted. ATPsynthase b (gift from DrGodinot, UCBL Lyon I, France) and ANT2 antisera wereused as primary antibodies which were detected witheither goat anti-rabbit or goat anti-mouse immunoglobu-lins conjugated to horseradish peroxydase. Immunoblotswere revealed with the ECL kit (Amersham Corp, UK).
Production of an ANT2 polyclonal antibody
The N-terminal sequence of the human ANT2 carriercorresponding to amino acids 1 to 10 (MTDAAVSFAK)was synthesized by Neosystem (Strasbourg, France).Eight copies of this peptide were bound to a ligand pep-tide (K4-K2-bA) to increase its antigenicity. The conju-gates corresponding to 250 mg of coupled peptides in0.5 ml of PBS were supplemented with 0.5 ml of com-plete Freund's adjuvant, and female New Zealand rab-bits were immunized. The ability of antisera to reactwith the synthetic peptides was tested by ELISA. TheWestern blot technique was also used to test the reactiv-ity of the anti-peptide antisera against the ADP/ATPprotein.
Functional complementation in yeast
Yeast strains/media/transformation
S. cerevisiae strains used were W303-1A (Mata, ade2-1,leu2-3, leu2-112, his3-11, his3-15, trp1-1, ura3-1, can1-100)and J�1�3 (Mata, ade2-1, leu2-3, leu2-112, his3-11, his3-15, trp1-1, ura3-1, can1-100, aac2::HIS3, aac1::LEU2,aac3::URA3), a gift from Dr Kolarov (University of Brati-

416 Regulation of the ANT2 Gene Expression
slava, Slovakia). The ura3 recipient strain (J�1�3/F) wasobtained by selection on 5-¯uoro-orotic acid medium(Rose et al., 1990). Yeast media were as follows: completeglucose medium (YPD) 1% (w/v) yeast extract, 1% (w/v) BactoPeptone, 2% (w/v) glucose, Minimal glucosemedium, 0.67% (w/v) yeast nitrogen base, 2% glucoseand the appropriate nutritional requirements. Anaerobiccultures were grown in 5% glucose minimal mediumsupplemented with 12 mg/ml ergosterol, 0.2% (v/v)Tween 80. Cells were grown anaerobically in sealed jarscontaining Anaerocult A (Merck).
Yeast transformation
The ANT2 and ANT3 cDNA were cloned in the KpnI/XbaI (pFL61-ANT2), and KpnI/BamHI (pFL61-ANT3)sites of the pFL61 vector (gift from Professor Lacroute,Gif sur Yvette, France), respectively (Minet et al., 1992).pFL61 contains the origin of replication of the yeast 2 mmplasmid, the URA3 gene, and the promoter and termin-ator of the yeast phosphoglycerate kinase gene for theexpression of foreign cDNA in S. cerevisiae. Cells from anexponential phase culture of the J�1�3/F strain weretransformed by electroporation (Meilhoc et al., 1990)using a Gene Pulser system (BioRad).
Control of plasmid gene expression
Total RNA from yeasts were extracted with the``RNeasy total RNA'' kit from Qiagen following manu-facturer's instructions. 5 mg of RNA were then submittedto DNase I digestion (GIBCO). For reverse-transcriptionanalysis, 1 mg of DNase-treated RNA was reverse tran-scribed using Superscript II RT (GIBCO) in the presenceof 0.1 mM hexaprimer, 10 mM DTT, 0.5 mM dNTP forone hour at 42�C. PCR ampli®cation was then performedas described above using oligonucleotides 50 nt 1017 to1038 and 30 nt 1218 to 1239 as primers for the ANT3cDNA ampli®cation.
Transient transfection assays, CAT and bbb-Gal assays
The genomic clone of the human ANT2 gene was agift from Dr Wurzel (Temple University, Philadelphia,PA). A PstI/PstI fragment (nt ÿ1237 to �46) of the 50region was cloned upstream of the CAT reporter gene ina pCAT basic vector (Promega) (pCAT-GRBOX). Anadditional clone (pCAT-�GRBOX: nt ÿ1158 to �46) wasprepared by removing speci®c restriction fragmentsfrom pCAT-GRBOX and religating the plasmid withDNA ligase (Eurogentec S.A., France). Oligonucleotidesused as electrophoretic mobility shift assay probes(Figure 6) were synthesized and puri®ed by HPLC(Eurogentec).
One day before transfection, exponentially growingcells (WI-38SV40, CAKI 1, rho� 143B) were plated at adensity of 5 � 105 cells/60 mm dishe. They were trans-fected using the Exgen 500 reagent (Euromedex, Souffel-weyersheim, France: Boussif et al., 1995). Cells weregently washed with PBS, followed by incubation in2.6 ml of fresh culture medium. Eight mg of pCAT-GRBOX, or pCAT-�GRBOX, were added to 200 ml of150 mM NaCl and homogenized by vortexing (solutionA). Twelve ml (for WI-38SV40) or 16 ml (for CAKI 1 andrho� 143B) of Exgen 500 were similarly homogenized in200 ml of 150 mM NaCl (solution B). Solutions A and Bwere mixed for ten minutes and the mixture was addedto the cells with 2.6 ml of culture medium and incubated
for a further 24 hours. Cells were then harvested andstored at ÿ20�C. The ef®ciency of transfection was deter-mined in parallel experiments using pCMV plasmid car-rying the b-galactosidase gene. Transfected cells werewashed with cold PBS, scraped from the dishes, pelletedfor ®ve minutes at 1000 g and stored at ÿ20�C. The dayof experiment, pelleted cells were lysed and 50 mg oftotal cellular protein were analysed by the CAT ELISAtest (Boehringer). The percentage of cells expressing theb-galactosidase was monitored by 5-bromo-chloro-3indolyl b-D-galactosidase staining (Lim & Chae, 1989).
Electrophoretic mobility shift assays
WI-38, WI-38SV40, CAKI 1 and rho� 143B nuclearextracts were prepared according to Lane et al. (1992)with slight modi®cations: exponentially growing cellswere harvested, rinsed twice with PBS and stored atÿ80�C. The pellet was lysed by homogenization with aDounce potter at 4�C in 400 ml of buffer containing10 mM Hepes-NaOH, 1.5 mM MgCl2, 10 mM KCl, 0.1%(v/v) NP40 (Sigma), 0.5 mM DTT, 0.5 mM PMSF(pH 7.9). The procedure was repeated until nuclei werefree of cytoplasmic contaminations, as judged by micro-scopical analysis with a TMS inverted photo microscopeequipped with phase contrast (Nikon Inc., Garden City,NY). The lysate was then centrifuged for two minutes at2000 g at 4�C. The pellet was dissolved in 100 ml of buf-fer containing 20 mM Hepes-NaOH, 420 mM NaCl,1.5 mM MgCl2, 0.2 mM EDTA, 25% (v/v) glycerol,0.5 mM DTT, 0.5 mM PMSF (pH 7.9) and incubated for30 minutes on ice. After two minutes at 8000 g at 4�Ccentrifugation, the supernatant was stored at ÿ80�C.
Forty base-pair oligonucleotides (Figure 6) were end-labelled with [g-32P]dATP and phage T4 polynucleotidekinase (Eurogentec). All double-stranded oligonucleo-tides were puri®ed by electrophoresis through a 10%non-denaturing polyacrylamide gel (19:1, acrylamide/bisacrylamide ratio, in 445 mM Tris-HCl, 10 mM EDTA(pH 8.3)). Probes (0.1 ng) and nuclear extracts (5 mg)were incubated at 37�C for ten minutes in a 20 ml reac-tion volume (8 mM Tris-HCl, 8 mM MgCl2, 80 mMNaCl, 10% glycerol, 0.5% NP40, 100 ng poly(dI-dC) �poly(dI-dC) (Pharmacia) (pH 8)). The reaction productswere separated on a pre-run 6% polyacrylamide gel at4�C in electrophoresis buffer (90 mM Tris-HCl, 90 mMH3BO3, 2.5 mM EDTA (pH 8): Balasubramanian et al.,1993). The gel was dried onto 3MM Chr Whatman paperand exposed to Kodak autoradiograph ®lm (BIOMAXMR).
Acknowledgements
We gratefully acknowledge Dr K. Dabbagh and Dr B.Mousson for critical assessment and suggestions aboutthe manuscript. This work was supported by grantsfrom the Association pour la Recherche sur le Cancer(ARC), and from the Centre National de la RechercheScienti®que (CNRS) awarded to G.S.; from the Fondationpour la Recherche MeÂdicale (FRM) awarded to W.-L.M.;S.G. was supported by a doctoral fellowship from theFondation M. MeÂrieux and B.-B.C. by a post-doctoral fel-lowship from the Association FrancËaise contre les Myo-pathies (AFM).

Regulation of the ANT2 Gene Expression 417
References
Adrian, G. S., McCammon, M. T., Montgomery, D. L. &Douglas, M. G. (1986). Sequences required for deliv-ery and localization of the ADP/ATP translocatorto the mitochondrial membrane. Mol. Cell. Biol. 6,626±634.
Balasubramanian, B., Lowry, C. V. & Zitomer, R. S.(1993). the ROX1 repressor of the Saccharomyces cere-visiae hypoxic genes is a speci®c DNA-binding pro-tein with a high-mobility-group motif. Mol. Cell.Biol. 13, 6071±6078.
Battini, R., Ferrari, S., Kaczmarek, L., Calabretta, B.,Chen, S. & Baserga, R. (1987). Molecular cloning ofcDNA for a human ADP/ATP carrier which isgrowth regulated. J. Biol. Chem. 262, 4355±4359.
Boussif, O., Lezoualc'h, F., Zanta, M. A., Mergny, M. D.,Scherman, D., Demeneix, B. & Behr, J. P. (1995).A versatile vector for gene and oligonucleotidetransfer into cells in culture and in vivo: polyethyle-nimine. Proc. Natl Acad. Sci. USA, 92, 7297±7301.
Chen, S. T., Chang, C. D., Huebner, K., Ku, D. H.,McFarland, M., deRiel, J. K., Baserga, R. & Wurzel,J. (1990). A human ADP/ATP translocase gene hasseven pseudogenes and localizes to chomosome X.Somat. Cell. Mol. Genet. 16, 143±149.
Chung, A. B., Stepien, G., Haraguchi, Y., Li, K. &Wallace, D. C. (1992). Transcriptional control ofnuclear genes for the mitochondrial muscle ADP/ATP translocator and the ATP synthase û subunit.J. Biol. Chem. 267, 21154±21161.
Cozens, A. L., Runswick, M. J. & Walker, J. E. (1989).DNA sequences of two expressed nuclear genes forhuman mitochondrial ADP/ATP translocase. J. Mol.Biol. 206, 261±280.
Dickman, K. G. & Mandel, L. G. (1989). Glycolytic andoxidative metabolism in primary renal proximaltubule cultures. Am. J. Physiol. 257, C333±C340.
Drgon, T., Sabova, L., Nelson, N. & Kolarov, J. (1991).ADP/ATP translocator is essential only for anaero-bic growth of yeast Saccharomyces cerevisiae. FEBSLetters, 289, 159±162.
Faure-Vigny, H., Heddi, A., Giraud, S., Chautard, D. &Stepien, G. (1996). Expression of oxidative phos-phorylation genes in renal tumors and tumoral celllines. Mol. Carcinogen. 16, 165±172.
Goossens, V., Grooten, J. & Fiers, W. (1996). The oxi-dative metabolism of glutamine. J. Biol. Chem. 271,192±196.
Heddi, A., Faure-Vigny, H., Wallace, D. C. & Stepien, G.(1996). Coordinate expression of nuclear and mito-chondrial genes involved in energy production incarcinoma and oncocytoma. Biochim. Biophys. Acta,1316, 203±209.
Huang, L. E., Ho, V., Arany, Z., Krainc, D., Galson, D.,Tendler, D., Livingston, D. M. & Bunn, H. F. (1997).Erythropoietine gene regulation depends on heme-dependent oxygen sensing and assembly of interact-ing transcription factors. Kidney Int. 51, 548±552.
Kindy, M. S., Brown, K. E. & Sonenshein, G. E. (1991).Regulation of expression of the growth-state-relatedgenes 2F1 and 2A9 during entry of quiescentsmooth muscle cells into the cell cycle. J. Cell. Bio-chem. 46, 345±350.
King, M. P. & Attardi, G. (1989). Human cells lackingmtDNA:repopulation with exogenous mitochondriaby complementation. Science, 246, 500±503.
Klingenberg, M. (1993). Dialectics in carrier research: theADP/ATP carrier and the uncoupling protein.J. Bioenerg. Biomembr. 25, 447±457.
Kolarov, J., Kolarova, N. & Nelson, N. (1990). A thirdADP/ATP translocator gene in yeast. J. Biol. Chem.265, 12711±12716.
KraÈmer, R. & Klingenberg, M. (1980). Modulation of thereconstituted adenine nucleotide exchange by mem-brane potentiel. Biochemistry, 19, 556±560.
Ku, D., Kagan, J., Chen, S., Chang, C., Baserga, R. &Wurzel, J. (1990). The human ®broblast adeninenucleotide translocator gene. Molecular cloning andsequence. J. Biol. Chem. 265, 16060±16063.
Laemmli, U. K. (1970). Cleavage of structural proteinsduring the assembly of the head of bacteriophageT4. Nature, 227, 680±685.
Lane, D., Prentki, P. & Chandler, M. (1992). Use of gelretardation to analyze protein-nucleic acid inter-actions. Microbiol. Rev. 56, 509±528.
Lawson, J. E. & Douglas, M. G. (1988). Separate genesencodes functional equivalent ADP/ATP carrierproteins in Saccharomyces cerevisiae. J. Biol. Chem.263, 14812±14818.
Li, K., Warner, C. K., Hodge, J. A., Minoshima, S.,Kudoh, J., Fukuyama, R., Maekawa, M., Shimizu,Y., Shimizu, N. & Wallace, D. C. (1989). A humanmuscle adenine nucleotide translocator gene hasfour exons, is located on chromosome 4, and is dif-ferentially expressed. J. Biol. Chem. 264, 13998±14004.
Li, K., Hodge, J. A. & Wallace, D. C. (1990). OXBOX, apositive transcriptional element of the heart-skeletalmuscle ADP/ATP translocator gene. J. Biol. Chem.265, 20585±20588.
Li, K., Neufer, P. D. & Williams, R. S. (1995). Nuclearresponses to depletion of mitochondrial DNA inhuman cells. Am. J. Physiol. 269, C1265±C1270.
Li, R., Hodny, Z., Luciakova, K., Barath, P. & Nelson,B. D. (1996). Sp1 activates and inhibits transcriptionfrom separate elements in the proximal promoter ofthe human adenine nucleotide translocase 2 (ANT2)gene. J. Biol. Chem. 271, 18925±18930.
Lim, K. & Chae, C. B. (1989). A simple assay for DNAtransfection by incubation of the cells in culturedishes with substrates for beta-galactosidase. Bio-Techniques, 7, 576±580.
Lowry, C. V. & Zitomer, R. S. (1988). Rox1 encodes aheme-induced repression factor regulating ANB1and CYC7 of Saccharomyces cerevisiae. Mol. Cell. Biol.8, 4651±4658.
Meilhoc, E., Masson, J. M. & TeissieÂ, J. (1990). High ef®-ciency transformation of intact yeast cells by electric®eld pulses. Biotechnology, 8, 223±227.
Minet, M., Dufour, M. E. & Lacroute, F. (1992). Cloningand sequencing of a human cDNA coding for dihy-droorotate dehydrogenase by complementation ofthe corresponding yeast mutant. Gene, 121, 393±396.
Nakashima, R. A., Paggi, M. G. & Pedersen, P. L. (1984).Contributions of glycolysis and oxidative phos-phorylation to adenosine 50-triphosphate productionin AS-30D hepatoma cells. Cancer Res. 44, 5702±5706.
Pedersen, P. L. (1978). Tumor mitochondria and thebioenergetics of cancer cells. Prog. Exp. Tumor Res.22, 190±274.
Rose, M. D., Winston, F. & Hieter, P. (1990). Methods inYeast Genetics: A Laboratory Course Manual, Cold

418 Regulation of the ANT2 Gene Expression
Spring Harbor Laboratory Press, Cold Spring Har-bor, NY.
Sabova, L., Zeman, I., Supek, F. & Kolarov, J. (1993).Transcriptional control of AAC3 gene encodingmitochondrial ADP/ATP translocator in Saccharo-myces cerevisiae by oxygen, heme and ROX1 factor.Eur. J. Biochem. 213, 547±553.
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1992). Molecu-lar Cloning: A Laboratory Course Manual, Cold SpringHarbor Laboratory Press, Cold Spring Harbor, NY.
Schiebel, K., Mertz, A., Winkelmann, M., Nagaraja, R. &Rappold, G. (1994). Localization of the adeninenucleotide translocase gene ANT2 to chromosomeXq24±q25 with tight linkage to DXS425. Genomics,24, 605±606.
Stepien, G., Torroni, A., Chung, A., Hodge, J. A. &Wallace, D. C. (1992). Differential expression of ade-nine nucleotide translocator isoforms in mammaliantissues and during cell differentiation. J. Biol. Chem.267, 14592±14597.
Torroni, A., Stepien, G., Hodge, J. A. & Wallace, D. C.(1990). Neoplastic transformation is associated withcoordinate induction of nuclear and cytoplasmicoxidative phosphorylation genes. J. Biol. Chem. 265,20589±20593.
Visser, W., van der Baan, A. A., Batenburg-van derVegte, W., Scheffers, W. A., Kramer, R. & vanDijken, J. P. (1994). Involvement of mitochondria inthe assimilatory metabolism of anaerobic Saccharo-myces cerevisiae cultures. Microbiology, 140, 3039±3046.
Zitomer, R. S. & Lowry, C. V. (1992). Regulation of geneexpression by oxygen in Saccharomyces cerevisiae.Microbiol. Rev. 56, 1±11.
Zitomer, R. S., Sellers, J. W., McCarter, D. W., Hastings,P. A., Wick, P. & Lowry, C. V. (1987). Elementsinvolved in oxygen regulation of Saccharomyces cere-visiae CYC7 gene. Mol. Cell. Biol. 7, 2212±2220.
Zitomer, R. S., Carrico, P. & Deckert, J. (1997). Regu-lation of hypoxic gene expression in yeast. KidneyInt. 51, 507±513.
Edited by M. Yaniv
(Received 26 February 1998; received in revised form 20 May 1998; accepted 27 May 1998)




















