
Denervation-induced proliferative changes of triads in rabbit skeletal muscle
14
Protein compositional and functional differences exist between longitudinal and junctional sarcoplasmic reticulum (SR) in relation to Ca transport and to Ca release. In light of this knowledge, we have reinvestigatedthe effects of denervation on SR of rabbit gastrocnemius, a predominantly fast muscle. Electron microscopy of 2-weeks denervated muscle showed proliferation of transverse tubules (lT), forming junctional contacts with SR terminal cister- nae (TC). At coincident periods, the yield of muscle microsomes was in- creased, and their fractionation by sucrose-density centrifugation demon- strated a relative increase of heavy vesicles. Thin-section electron microscopy of heavy SR from denervated muscle showed an increased number of vesicles containing calsequestrin (CS) as compared with control muscle. Electrophoretic analysis confirmed the relative decrease of Ca- ATPase protein and the striking increase of CS both in total microsomes and in heavy SR vesicles. Calcium loading and Ca-ATPase activity as well as the density of Ca-ATPase protein were decreased to a similar extent (20-30%) in denervated muscle microsomes. Stimulation of Ca-ATPase activity by Ca-ionophore A231 87 showed that the vesicles were tightly sealed. When probed by competitive ELSA with antibody to SR Ca- ATPase from pure fast muscle, the Ca-ATPase of denervated rnicrosomes was found to be highly cross reactive. Cleveland's peptide maps of the Ca- ATPase protein after partial digestion with S. aureus V8 protease also showed no significant change after denervation. Changes in cholesterol content and in the ratio of Mg-ATPase to Ca-ATPase activity of denervated muscle microsomes indicated a 4-fold increase of lT protein, i.e., from about 3% to not more than 12% of total protein, at 2 weeks after denerva- tion. All these changes were totally reversed upon reinnervation of muscle fibers, and the consequent muscle recovery, as obtained by nerve crushing instead of nerve sectioning. From these results, we conclude that dener- vated adult fast muscle, similarly to immature fast muscle, contains more junctional SR. However, the molecular and catalytic properties of the Ca- ATPase are unaffected by denervation. MUSCLE & NERVE 11A246-1259 1989 DENERVATION=INDUCEDPROLIFERATIVE CHANGES OF TRIADS IN RABBIT SKELETAL MUSCLE SERGIO SALVATORI, DScB, ERNEST0 DAMIANI, MD, FRANCESCO ZORZATO, MD, POMPEO VOLPE, MD, SANDRA PIEROBON, DScB, DANIELA QUAGLINO Jr., DScB, GlOVANNl SALVIATI, MD, and ALFRED0 MARGRETH, MD From the Centro di Studio della Biologia e Fisiopatologia Muscolare del CNR, lstituto di Patologia generale dell'Universit8 di Padova, via Loredan 16, 35131, Padova. Italy (Drs. Salvatori, Damiani, Zorzato. Volpe. Piero- bon, Salviati, and Margreth), and the lstituto di Patologia generale dell'Universit6 di Modena, via Campi 28, Modena, Italy (Dr. Quaglino). Acknowledgments. This work was supported by institutional funds from the Consiglio Nazionale delle Ricerche and from the Minister0 della Pub- blica lstruzione to A.M. The technical assistance of Mr. G.A. Tobaldin and of Miss S. Furlan is gratefully acknowledged. We also thank Dr. D. Biral for typing the fiber-type population of muscle specimens by his- tochemical and immunofluorescent staining and Mrs. Elena Golin for her kind editorial assistance. Address reprint requests to Dr. Margreth at the Institute of General Pa- thology, Via Loredan 16, 35131 Padova, Italia. Accepted for publication March 8, 1988. 01 48-639)(/1112/1246 $04.00/14 0 1988 John Wiley & Sons, Inc In skeletal muscle the contraction-relaxation cycle and duration of the active state are regulated by changes in the myoplasmic free calcium concen- tration, which are in turn mainly dependent on the rate and total amount of calcium released from the terminal cisternae (TC) of the sarcoplas- mic reticulum (SR) system and on the rate of cal- cium uptake into the SR lumen through the activ- ity of the Ca-ATPase protein. The existence of areas of membrane specialization within skeletal muscle SR, in relation to intracellular calcium fluxes, is now well e s t a b l i ~ h e d . ~ . ' ~ ~ ~ ~ Isolated TC from rabbit fast muscle have been characterized with regard to calcium release propertiesg and to 1246 Proliferation of Triads MUSCLE & NERVE December 1988
Transcript of Denervation-induced proliferative changes of triads in rabbit skeletal muscle

Protein compositional and functional differences exist between longitudinal and junctional sarcoplasmic reticulum (SR) in relation to Ca transport and to Ca release. In light of this knowledge, we have reinvestigated the effects of denervation on SR of rabbit gastrocnemius, a predominantly fast muscle. Electron microscopy of 2-weeks denervated muscle showed proliferation of transverse tubules (lT), forming junctional contacts with SR terminal cister- nae (TC). At coincident periods, the yield of muscle microsomes was in- creased, and their fractionation by sucrose-density centrifugation demon- strated a relative increase of heavy vesicles. Thin-section electron microscopy of heavy SR from denervated muscle showed an increased number of vesicles containing calsequestrin (CS) as compared with control muscle. Electrophoretic analysis confirmed the relative decrease of Ca- ATPase protein and the striking increase of CS both in total microsomes and in heavy SR vesicles. Calcium loading and Ca-ATPase activity as well as the density of Ca-ATPase protein were decreased to a similar extent (20-30%) in denervated muscle microsomes. Stimulation of Ca-ATPase activity by Ca-ionophore A231 87 showed that the vesicles were tightly sealed. When probed by competitive ELSA with antibody to SR Ca- ATPase from pure fast muscle, the Ca-ATPase of denervated rnicrosomes was found to be highly cross reactive. Cleveland's peptide maps of the Ca- ATPase protein after partial digestion with S. aureus V8 protease also showed no significant change after denervation. Changes in cholesterol content and in the ratio of Mg-ATPase to Ca-ATPase activity of denervated muscle microsomes indicated a 4-fold increase of lT protein, i.e., from about 3% to not more than 12% of total protein, at 2 weeks after denerva- tion. All these changes were totally reversed upon reinnervation of muscle fibers, and the consequent muscle recovery, as obtained by nerve crushing instead of nerve sectioning. From these results, we conclude that dener- vated adult fast muscle, similarly to immature fast muscle, contains more junctional SR. However, the molecular and catalytic properties of the Ca- ATPase are unaffected by denervation.
MUSCLE & NERVE 11A246-1259 1989
DENERVATION=INDUCED PROLIFERATIVE CHANGES OF TRIADS IN RABBIT SKELETAL MUSCLE
SERGIO SALVATORI, DScB, ERNEST0 DAMIANI, MD, FRANCESCO ZORZATO, MD, POMPEO VOLPE, MD, SANDRA PIEROBON, DScB, DANIELA QUAGLINO Jr., DScB, GlOVANNl SALVIATI, MD, and ALFRED0 MARGRETH, MD
From the Centro di Studio della Biologia e Fisiopatologia Muscolare del CNR, lstituto di Patologia generale dell'Universit8 di Padova, via Loredan 16, 35131, Padova. Italy (Drs. Salvatori, Damiani, Zorzato. Volpe. Piero- bon, Salviati, and Margreth), and the lstituto di Patologia generale dell'Universit6 di Modena, via Campi 28, Modena, Italy (Dr. Quaglino).
Acknowledgments. This work was supported by institutional funds from the Consiglio Nazionale delle Ricerche and from the Minister0 della Pub- blica lstruzione to A.M. The technical assistance of Mr. G.A. Tobaldin and of Miss S. Furlan is gratefully acknowledged. We also thank Dr. D. Biral for typing the fiber-type population of muscle specimens by his- tochemical and immunofluorescent staining and Mrs. Elena Golin for her kind editorial assistance.
Address reprint requests to Dr. Margreth at the Institute of General Pa- thology, Via Loredan 16, 35131 Padova, Italia.
Accepted for publication March 8, 1988.
01 48-639)(/1112/1246 $04.00/14 0 1988 John Wiley & Sons, Inc
In skeletal muscle the contraction-relaxation cycle and duration of the active state are regulated by changes in the myoplasmic free calcium concen- tration, which are in turn mainly dependent on the rate and total amount of calcium released from the terminal cisternae (TC) of the sarcoplas- mic reticulum (SR) system and on the rate of cal- cium uptake into the SR lumen through the activ- ity of the Ca-ATPase protein. The existence of areas of membrane specialization within skeletal muscle SR, in relation to intracellular calcium fluxes, is now well e s t a b l i ~ h e d . ~ . ' ~ ~ ~ ~ Isolated T C from rabbit fast muscle have been characterized with regard to calcium release propertiesg and to
1246 Proliferation of Triads MUSCLE & NERVE December 1988

the distinct ultrastructural and protein composi- tional features" of the free and junctional mem- brane regions. The calcium-binding protein calse- questrin (CS) not only is selectively localized in the lumen of TC'"'' but is seemingly anchored to the inner profile of the junctional membrane, oppo- site the site of attachment of the feet projections on the outer surface.'s241 On the other hand, the Ca-ATPase protein is diffusely distributed in SR membranes except for that in the junctional mem- brane of TC."
Denervation of skeletal muscles leads to a wide array of changes in the properties and protein composition of the surface membrane,47348 to pro- liferation of transverse tubules (TTs), and to a parallel increase of nitrendipine-sensitive calcium- channel^.^^,^^ Electron microscopic studies of amphibian'"34 and rat muscle^'^,^' have shown that an early response to denervation is the hyper- trophy of TC, followed by an increase in the num- ber of triads.46 Conflicting results have been re- ported concerning changes in membrane protein composition and in the properties of the calcium transport system of the SR of denervated skeletal muscle, depending also on the animal species, the fiber-type composition of the muscle, and the time-period after d e n e r v a t i ~ n . ~ ~ , ~ " ' ~ In addition, many of these previous results are open to criti- cism due to the heterogeneous membrane origin of muscle m i c r o ~ o m e s ~ ~ ~ ~ ~ and since it is only re- cently that the protein composition and functional characterization of junctional and extrajunctional SR, as well as of TT, became p ~ s s i b l e . " ~ ~ ' , ~ ~ ~ ~ ~
Here we report electron microscopic, immuno- logical, and biochemical observations on total mi- crosomes and on purified heavy and light SR vesi- cles from rabbit gastrocnemius muscle after mechanical denervation, either by nerve section- ing or by nerve crushing. Our results demonstrate that the immunological and biochemical proper- ties of the Ca-ATPase and the efficiency of the Ca- transport system are not affected at 2 weeks after denervation, even though the average membrane density of the Ca-ATPase units in SR membranes decreases by 20-30%, mostly as a result of a rela- tive increase of junctional SR. We also show that the transitions in membrane composition and properties of skeletal muscle SR after denervation are accompanied by a proliferation of T-tubules and that both these changes are reversible follow- ing reinnervation of the muscle.
MATERlALS AND METHODS
Chemicals. All chemicals were analytical grade reagents and were obtained from Merck A. G. (Darmstadt, FRG) and Carlo Erba (Milano, Italy). p-Nitrophenylphosphate, pyruvate kinase, lactate dehydrogenase, cholesterol oxidase, esterase, and peroxidase were obtained from Boehringer (Man- neheim, FRG). Ionophore A23 187 was purchased from Calbiochem (Behring Co., La Jolla, CA). Stains All 1 -ethyl-2-[3-( 1-ethylnaphto-[ 1, 2d]-thia- zolin-2-ylidene)-2-methylpropenyl-naphto][ 1, 2 4 thiazoliurn bromide, anti-(chicken) and anti- (rabbit) IgG were obtained from Sigma Chemical Co. (St. Louis, MO).
Preparative Procedures. New Zealand male adult rabbits were used. The gastrocnemius, a represen- tative fast-twitch muscle, was denervated by sec- tioning the common popliteal nerve at popliteal space. Alternatively, the nerve was crushed by pinching with a clamp precooled in liquid nitro- gen. The contralateral muscle from the unoper- ated leg was used as control. Animals were sacri- ficed by cervical dislocation at 1 week to 3 months after denervation, as reported in the figures and tables. Total microsomes were obtained as de- scribed previously (Ref. 44, see procedure B) and were fractionated by discontinuous sucrose- density gradient ~entrifugation.~' Protein concen- tration was determined according to Lowry et al.24 with bovine serum albumin as a standard.
Biochemical Assays. Cholesterol content was measured enzymatically as previously described.44 ATPase activity was determined at 37°C with an enzyme-linked spectrophotometric assay5' by mea- suring the oxidation of NADH at 340 nm.44 Basal, Mg-stimulated ATPase was measured in the pres- ence of 0.2 mM EGTA. Extra Ca-stimulated ATPase was measured after adding 0.2 mM CaC1, and 4 ph4 A23187. Ca uptake in the absence of Ca-precipitating anions was measured as de- scribed4' by a Ca-sensitive electrode (Mueller Glassblaserei, Geneve, Switzerland) at 25°C in a medium containing, in a final volume of 4 ml, 20 mM tris-maleate, pH 6.8, 100 mM KC1, 5 mM MgCl,, 2.5 mM ATP, 12 pg/ml creatine kinase, 5 mM creatine phosphate, and 200-300 pg of mi- crosomal protein. Phosphate-facilitated Ca loading was measured with 10-20 pg of protein in a me- dium having a composition identical to that used for Ca-uptake except that it was buffered at pH 7.0 with 100 mM Na-phosphate.
Proliferation of Triads MUSCLE & NERVE December 1988 1247

Gel Electrophoresis and Peptide Mapping. One-dimensional sodium dodecyl sulfate polyacry- lamide gel electrophoresis (SDS-PAGE) was car- ried out according to Laemmli.*’ Two-dimen- sional gel electrophoresis was carried out accord- ing to Michalak et al.32 Myosin heavy chains were electrophoretically separated as described by Biral et al.’ Peptide mapping of SR Ca-ATPase was car- ried out with S . aureus V8 protease, according to Cleveland et a1.l’ Slab gels were stained with Coomassie brilliant blue by a silver nitrate method31 or with Stains All.’ Densitometric scan- ning of slab gels was carried out as described by Volpe et a1.53
immunological Techniques. One-step noncompet- itive and two-step competitive enzyme-linked im- munosorbent assay (ELISA) were carried out as described previously.”
Electron Microscopy. Heavy SR fractions were prefixed at room temperature with 0.5% glutaral- dehyde in Tyrode’s buffer plus 0.5% tannic acid and centrifuged. Pellets were fixed with 2.5% glu-
4-r 0
.. 50
1 2 3
0
f. 0 I . .
0 0
0
0.
0
taraldehyde in Tyrode’s for 1 hr, treated with 1% OsO , , in Tyrode’s buffer for 30 min at room tem- perature, dehydrated in ethanol, and embedded in durcupan (Fluka, A.G. Switzerland). Sections were stained with 3% uranyl acetate in 50% etha- nol and lead citrate. Observations were made with a Philips 400 T electron microscope. Thin-section electron microscopy of muscle specimens was car- ried out as described by Pierobon-Bormioli et al.38
RESULTS
Muscle Weight and Fiber-Type Composition. The time course of changes in the fresh weight of rab- bit gastrocnemius after nerve sectioning (Fig. 1A) was similar to that described in previous ~tudies . ’”~ Experiments in which the muscle was denervated by nerve crushing showed a steady loss in weight, up to 30 days after the operation. Thereafter, the innervation of muscle fibers from the regenerating motor nerve resulted in about 80% recovery of the original muscle weight dur- ing the subsequent 2 months. At 2-3 weeks after denervation, the gastrocnemius had lost 40% in weight, as compared with the contralateral control
0 20 40 60 80 100
Days post- operatively FIGURE 1. Muscle atrophy and myosin heavy chain composition of denervated muscle. (A) Changes in fresh muscle weight after op- eration. The fresh weight of operated and contralateral gastrocnemius muscle of rabbits were determined at times indicated on the abscissa. Values, expressed as percentage of control values, are the means of at least three experiments in the case of nerve section- ing. Vertical bars represent +SEM when more than three determinations were carried out. Values obtained after nerve crushing were from individual rabbits. Key: (0) nerve sectioning; (0) nerve crushing. (6) Myosin HC were resolved by 5% SDS-PAGE.3 About 200 ng of protein were loaded on each lane. Slabs were stained with silver nitrate.31 Key to myosins: (1) control muscle, (2) denervated muscle, (3) rabbit soleus, a representative slow-twitch muscle. Abbreviations used; HCf, fast myosin HC; HCs, slow myosin HC.
1248 Proliferation of Triads MUSCLE & NERVE December 1988

muscle by either denervation procedure. In agree- ment with earlier evidence, the rate of atroph
which in rabbit gastrocnemius account for about 90% of the total muscle fiber population, as typed by histochemical staining for myosin ATPase or by immunofluorescent staining, both using antibody to fast myosin and antibody to the fast-type of sar- coplasmic reticulum (SR) Ca-ATPase (results not shown). Despite that, the electrophoretic pattern of myosin heavy chains (Fig. lB), which in rabbit gastrocnemius is characterized by the predomi- nance of the fast heavy-chain isoform of lower mobility, appears to be maintained, as determined at 2 weeks after denervation.
was found to be faster for fast-twitch fibers, 37
Ultrastructural Changes In Denervated Murcle. When examined in the electron microscope 2- 3 weeks after nerve sectioning, gastrocnemius mus- cle fibers exhibited a marked atrophy of the myo- fibrils and extensive modifications in architecture and development both in TT and SR in mem- branes. The proliferation of TT, which is a char- acteristic response to d e n e r ~ a t i o n , ' * ~ ~ ' ~ ~ was best evidenced in lanthanum- stained ultrathin sec- tions, under which conditions the limiting mem- brane stains distinctly darker than SR rnembrane~.~' In contrast to normal fibers, in which TT exhibit a regular disposition with re- spect to sarcomeres, on each side of the I band (Fig. 2A), the TT of denervated fibers appear to be much more extensively developed and to branch and extend longitudinally (Fig. 2B). Despite the ir- regular disposition, the proliferating TT are iden- tified mainly as junctional TT from the presence of connecting feet processes with flattened SR cis- ternae containing dense, granular material and most often looking hypertrophic, as compared with normal cisternae (Fig. 2C). Consequently, in denervated muscle fibers, triads are increased in number and have an irregular disposition with re- spect to sarcomeres. Interestingly, at the same pe- riods after denervation, more coated vesicles are observed, most often lying in subsarcolemmal ar- eas, but also occasionally found located at interior sites in intermyofibrillar spaces, some adjacent to membrane profiles of SR cisternae and of TT (Fig. 2D).
Transitions in Membrane-Protein Composition of Muscle Microsomes. As shown in Figure 3 and in agreement with earlier observations on amphib- i a r ~ , ' ~ mammalian,26p36 and avian5' muscles, the yield of microsomal protein from rabbit gastroc-
nemius rises following denervation, the peak value being reached at a stage of atrophy corresponding to a loss in muscle weight of 20-30% (see Fig. 1A). When analyzed by SDS-PAGE, the mi- crosomes from denervated muscle exhibit a mem- brane protein composition qualitatively similar to that of purified SR membranes from normal gas- trocnemius, the main proteins being identified as the 100 kDa Ca-ATPase, and the 64 kDa calse- questrin (CS), in addition to several protein com- ponents in the 55-53 kDa range (Fig. 4A, lane 1).
The main significant changes seen at 2 weeks after denervation are a relative decrease of Ca- ATPase protein and a more prominent increase of CS (Fig. 4A, lane 2), as identified by its apparent M r 64 kDa, its characteristic blue staining with the cationic carbocyanine dye Stains All (Fig. 4A, lanes 7 and 8), and its off-diagonal position when ana- lyzed in the two-dimensional gel system of Micha- lak et al.32 (Fig. 4B). Densitometric measurements demonstrated that the ratio of CS to the Ca-ATPase protein increased from a control value of 0.11, on average, to 0.36 at 2 weeks as shown in Fig. 5.
The nature (Fig. 4A, lanes 5 and 6) as well as the time-course of changes (Fig. 6, lower panel) in membrane- protein composition were found to be quite similar in the experiments of nerve crushing for the duration of the denervated period (3-4 weeks). Afterwards, at times coincident with the onset of cytoplasmic growth in the reinnervated muscle fibers, the ratio of CS to the Ca-ATPase protein progressively decreased, approaching nor- mal values at 10 weeks (Fig. 6, lower panel).
Changes in Ca-ATPase activity, measured in the presence of the Ca-ionophore A23187, ap- peared to be time-correlated with percentage in the intramembrane content of the 100 kDa pro- tein, both in the falling phase associated with mus- cle atrophy (Fig. 3) and during muscle recovery, i.e., starting from about 2 months after nerve crushing (Fig. 6, upper panel).
In order to eliminate the possibility that the decrease in Ca-ATPase activity of the microsomes from denervated gastrocnemius could reflect a shift from the predominant fast isoform of SR Ca- ATPase to the slow form4s52" which characteristi- cally has a lower catalytic activity,44 the isolated membranes were probed with specific antibody to fast SR Ca-ATPase, which was previously demon- strated to be able to distinguish between the fast
competitive ELISA techniques. As shown in Fig. 7A, the extent of immunological reactivity of the ATPase protein bound to microsomes of dener-
and slow isoforms of SR b Y
Proliferation of Triads MUSCLE & NERVE December 1988 1249
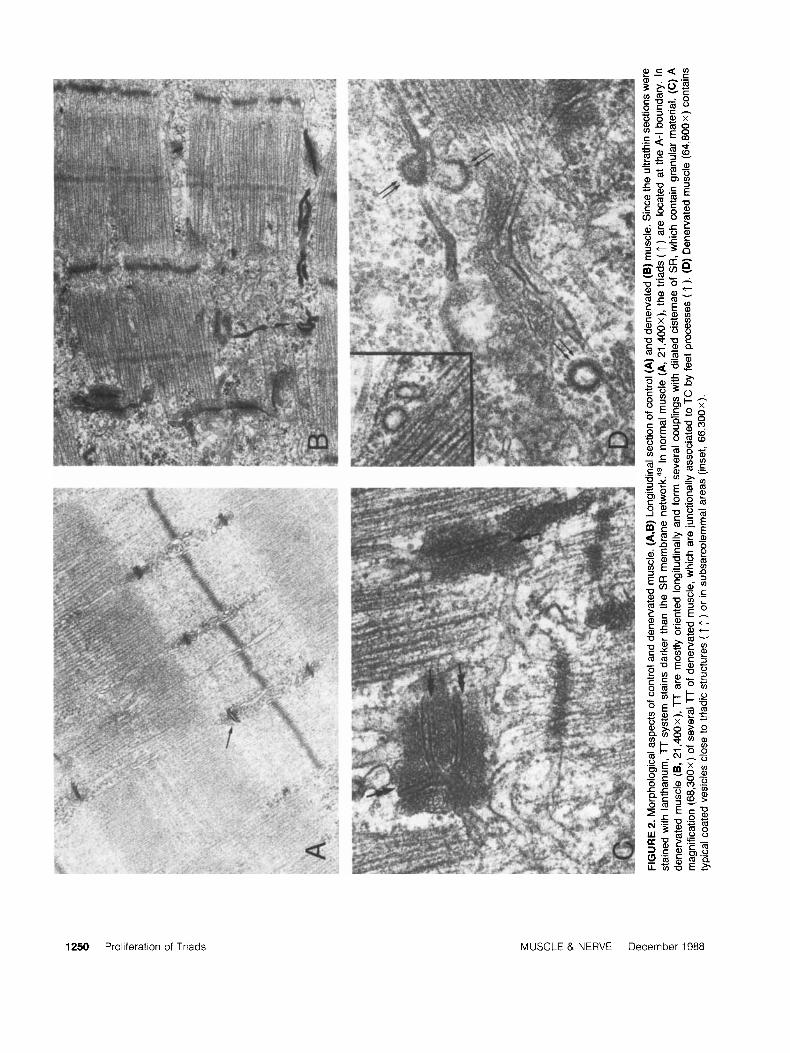
1250 Proliferation of Triads MUSCLE & NERVE December 1988

0 1 2 3 4 5 8 7
Weeks a f t e r denervation
0 0 l r l l l , , l , *
FIGURE 3. Effects of denervation on the yield of microsomal protein, on the Ca-ATPase activity, and on the Ca-ATPase membrane content of total microsomal fraction. Microsomes were isolated at times indicated on the abscissa from control and denervated muscle, as reported in Materials and Methods. Ca-ATPase activity was measured in the presence of calcium ionophore A23187. Percentage of Ca-ATPase protein was quantitated by densitometry of Coomassie blue-stained slab gels. Values are the means of at least three ex- periments with paired, denervated, and contralateral control muscles. Vertical bars represent *SEM, where more than three deterrnina- tions on different membrane preparations were carried out.
vated muscle appears to be virtually unmodified up to 4 weeks after the operation. Similarly, pep- tide mapping of the 100 kDa ATPase by Cleve- lands techniques using S . aureus V8 proteaseZ5 shows that the pattern specific to the fast form of SR Ca-ATPase is maintained until 7 weeks after nerve sectioning (Fig. 7B).
When examined for the content of total choles- terol, a distin uishing marker of TT versus SR membranes, 23’9*50 the total microsomal fraction shows a steady increase of cholesterol after dener- vation (Figs. 5 and 6, lower panel), these changes again being completely reversed following muscle reinnervation in the experiments with nerve crushing (Fig. 6, lower panel). The results in Fig. 5 show that the increase in membrane cholesterol after denervation appears to be time-correlated with the increase in the ratio between Mg-ATPase activity2. 13,35,40 and Ca-ATPase activity measured in the same fraction.
Characterization of Membrane Fractions Obtained by Sucrose-Density Centrifugation After Denervation. Microsomal vesicles comprise SR vesicles derived from junctional and free TC and from the longi- tudinal tubules, as well as a normally low propor-
tion of TT vesicles, that can be separated by su- crose-density gradient centrifugation. When ana- lyzed by the method of Rosemblatt et al.,39 the microsomes from 2 weeks denervated gastrocne- mius are separated into a very light, a light, and a heavy fraction (Fig. 8A) having buoyant density properties similar to those of the corresponding fractions from control muscle. However, dener- vated muscle microsomes appear to be relatively enriched in heavy as well as in very light vesicles, as confirmed by the respective protein yields in Table I.
The majority of SR vesicles composing the heavy fraction of normal gastrocnemius, when ex- amined in the electron microscope after staining with tannic acid (Fig. 9A), show the presence of intraluminal electron-dense material correspond- ing to CS, which is consistent with their predomi- nant origin from cisternal SR.29 The correspond- ing fraction from denervated muscle is differ- entiated by the presence of an increased number of filled vesicles (Fig. 9B), some of which, i.e., those richer in CS, exhibit junctional feet struc- tures on the outer membrane profile, opposite the internal site of attachment of CS (Fig. 9C). In agreement with these distinguishing morphologi-
Proliferation of Triads MUSCLE & NERVE December 1988 1251
A 0
9 * E h c .- .? 5
0 0 .- Y
c
1st-
0 1 2 3 4 5 8 7
W e e k s a f t e r d e n e r v a t i o n FIGURE 5. Cholesterol content, Mg- and Ca-dependent ATPase activities, and relative content of calsequestrin and Ca-ATPase protein of microsomes from control and denervated muscle. Mi- crosomes were isolated from control and denervated muscle at times indicated on the abscissa. Cholesterol (nonesterified cho- lesterol + cholesterol esters) was determined as previously described.44 Mg- and Ca-dependent ATPase activities were measured as described in Methods. Percentage of CS and of Ca-ATPase protein was quantitated as in Fig. 3. Values are the average of three determinations on paired muscles from differ- ent rabbits.
a c c\1
1
FIGURE 4. Protein profile of muscle rnicrosomes from control and denervated muscle. (A) One-dimensional 10% SDS-PAGE of microsomes from control and operated muscle, after either nerve sectioning (lanes 2, 3, 4, and 8) or nerve crushing (lanes 5 and 6), was carried out according to Laemmli.22 Gels were stained with Coornassie blue (lanes 1-6) or with Stains All (lanes 7 and 8); 30 pg of protein were loaded per gel. Key to lanes: 1 and 7) control muscle, (2 and 8) 2-weeks denervated muscle, (3) 3-weeks denervated muscle, (4) 6-weeks dener- vated muscle, (5) 2-weeks after nerve crushing, (6) 10-weeks after nerve crushing. Asterisks indicate blue-staining proteins. (6) Two-dimensional electrophoresis in 10% SDS-PAGE32 of microsomes from control (upper panel) and 2-weeks denervated (lower panel) muscle. Gels were stained with Coomassie blue; 30 pg of protein were loaded per gel.
cal features, the calculated values of the ratio of CS to Ca-ATPase protein, by gel densitometry of slab gels (see Fig. SB), are found to be more than three times higher for denervated than for control heavy SR (Table I). On the other hand, a very low content of cholesterol and of Mg-ATPase activity appear to be properties shared by the heavy SR of denervated and control gastrocnemius. This find- ing together with the evidence that both fractions are similarly weakly cross-reactive with specific an- tibody to the Mg-ATPase of chicken TT (Fig.
10D) does therefore indicate that the heavy SR fraction, as obtained by the method of Rosemblatt et a ~ , ~ ’ is relatively free of triads as well as of ‘IT fragments.
Measurements of Ca-ATPase rates, in the ab- sence and in the presence of Ca-ionophore A23187, are also reported in Table I. The results show that the extent of stimulation of Ca-ATPase activity by the ionophore does not differ signifi- cantly in microsomes from denervated muscle from that found with control preparations, thus suggesting that the SR vesicles are normally sealed. The Ca-ATPase activity values for control heavy SR (Table 1) are in agreement with re- ported values in the literature30344 as are rates of phosphate-facilitated Ca-uptake measured in the presence of 5 mM MgC1, (see Table V of Ref. 33). On the other hand, the Ca-ATPase activity and the Ca-loading rate of the heavy SR fraction from denervated muscle appear to be both reduced by about 40%, i.e., to about the same extent as the decrease in membrane content of Ca-ATPase pro- tein (see Table I). In agreement with these results, we found in some experiments that the Ca/A‘TP coupling ratio of heavy SR vesicles from dener-
1252 Proliferation of Triads MUSCLE & NERVE December 1988
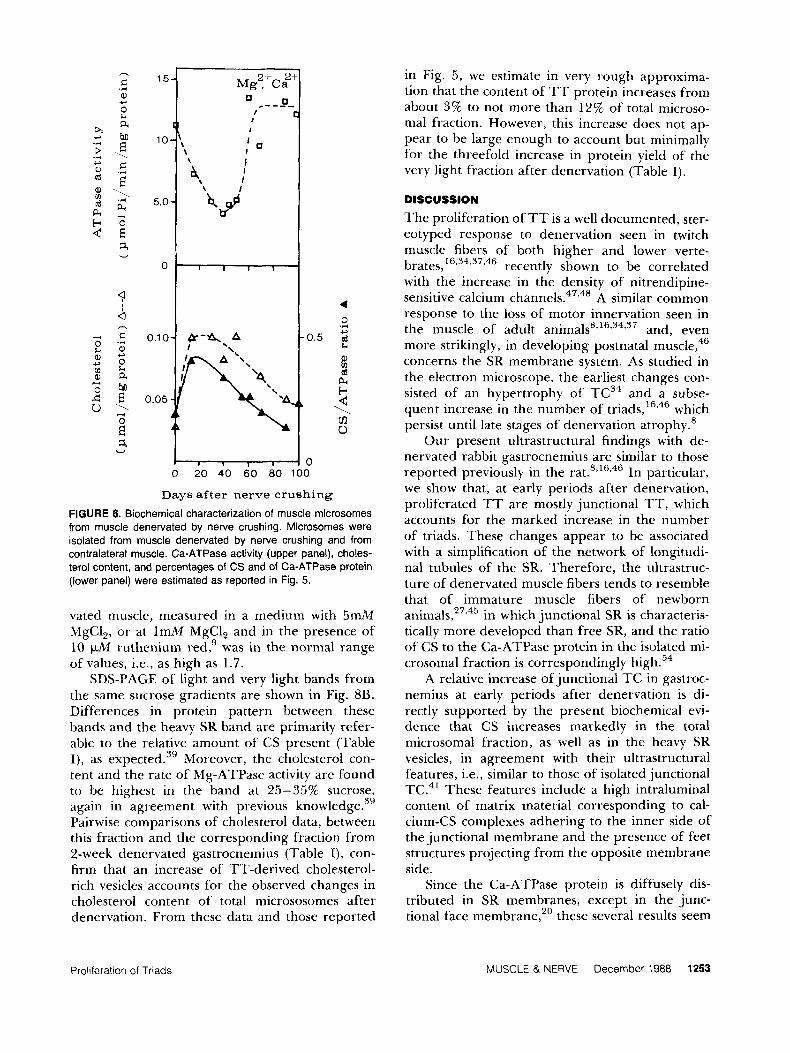
15
10
5.0
0
0.1 0
0.05
C r - 4 , A I
2+ 2 M g , Ca
0.5
\ I 1 4, I
‘4 w 4’
I 0
4 0
0 20 40 60 80 100
Days after nerve crushing FIGURE 6. Biochemical characterization of muscle microsomes from muscle denervated by nerve crushing. Microsomes were isolated from muscle denervated by nerve crushing and from contralateral muscle. Ca-ATPase activity (upper panel), choles- terol content, and percentages of CS and of Ca-ATPase protein (lower panel) were estimated as reported in Fig. 5.
vated muscle, measured in a medium with 5mh4 MgCl,, or at 1mM MgC1, and in the presence of 10 pA4 ruthenium red,g was in the normal range of values, i.e., as high as 1.7.
SDS-PAGE of light and very light bands from the same sucrose gradients are shown in Fig. 8B. Differences in protein pattern between these bands and the heavy SR band are primarily refer- able to the relative amount of CS present (Table I), as e~pec ted .~’ Moreover, the cholesterol con- tent and the rate of Mg-ATPase activity are found to be highest in the band at 25-35% sucrose, again in agreement with previous kn~wledge.~’ Pairwise comparisons of cholesterol data, between this fraction and the corresponding fraction from 2-week denervated gastrocnemius (Table I), con- firm that an increase of TT-derived cholesterol- rich vesicles accounts for the observed changes in cholesterol content of total micrososomes after denervation. From these data and those reported
in Fig. 5 , we estimate in very rough approxima- tion that the content of TT protein increases from about 3% to not more than 12% of total microso- ma1 fraction. However, this increase does not ap- pear to be large enough to account but minimally for the threefold increase in protein yield of the very light fraction after denervation (Table I).
DISCUSSION
The proliferation of TT is a well documented, ster- eotyped response to denervation seen in twitch muscle fibers of both higher and lower verte-
recently shown to be correlated with the increase in the density of nitrendipine- sensitive calcium channels.47348 A similar common response to the loss of motor innervation seen in the muscle of adult animals8316334v37 and, even more strikingly, in developing postnatal muscle,46 concerns the SR membrane system. As studied in the electron microscope, the earliest changes con- sisted of an hypertrophy of TC34 and a subse- quent increase in the number of triads,’6246 which persist until late stages of denervation atrophy.*
Our present ultrastructural findings with de- nervated rabbit gastrocnemius are similar to those reported previously in the In particular, we show that, at early periods after denervation, proliferated TT are mostly junctional TT, which accounts for the marked increase in the number of triads. These changes appear to be associated with a simplification of the network of longitudi- nal tubules of the SR. Therefore, the ultrastruc- ture of denervated muscle fibers tends to resemble that of immature muscle fibers of newborn
in which junctional SR is characteris- tically more developed than free SR, and the ratio of CS to the Ca-ATPase protein in the isolated mi- crosomal fraction is correspondingly high.54
A relative increase of junctional TC in gastroc- nemius at early periods after denervation is di- rectly supported by the present biochemical evi- dence that CS increases markedly in the total microsomai fraction, as well as in the heavy SR vesicles, in agreement with their ultrastructural features, i.e., similar to those of isolated junctional TC.41 These features include a high intraluminal content of matrix material corresponding to cal- cium-CS complexes adhering to the inner side of the junctional membrane and the presence of feet structures projecting from the opposite membrane side.
Since the Ca-ATPase protein is diffusely dis- tributed in SR membranes, except in the junc- tional face membrane,,’ these several results seem
brates, 16,34,37,46
Proliferation of Triads MUSCLE & NERVE December 1988 1253
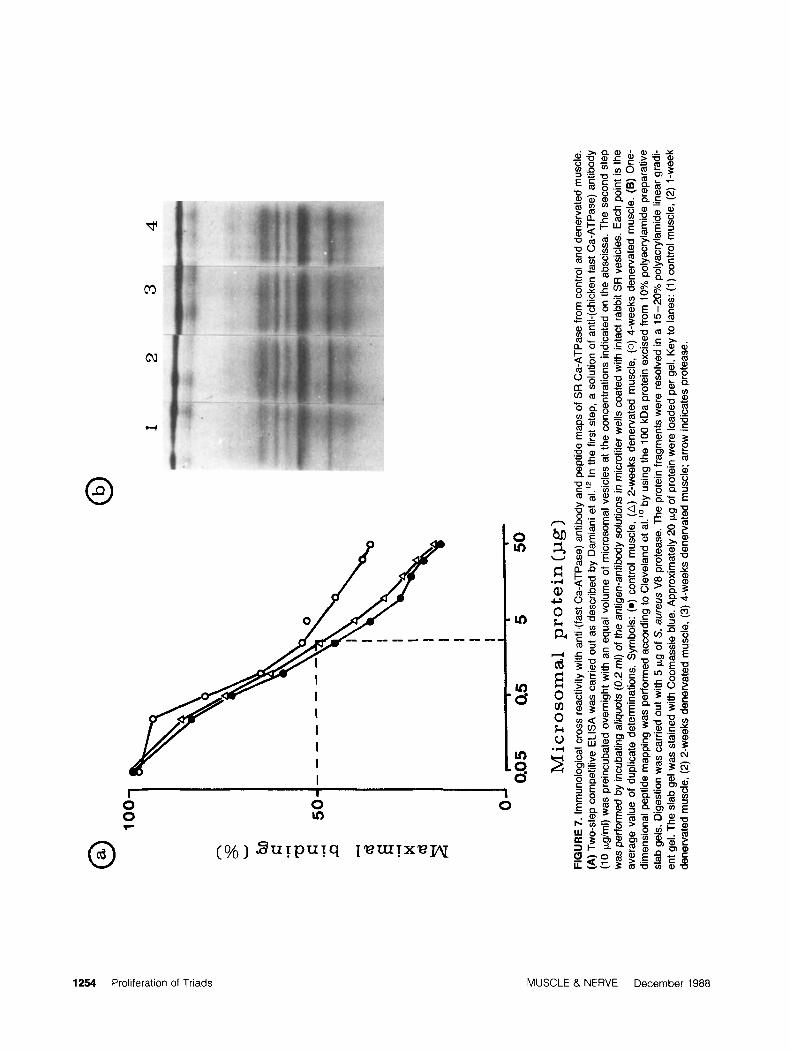
A
N e z
rn
ID
<
rn
A
(D
03
03
100
50
0
-------
0.05
Q
5 5
50
Mic
roso
mal
pro
tein
(pg)
1 2
3
4
FIG
UR
E 7.
Im
mun
olog
ical
cro
ss re
activ
ity w
ith a
nti-(
fast
Ca-
ATP
ase)
ant
ibod
y an
d pe
ptid
e m
aps
of S
R C
a-A
TPas
e fro
m c
ontro
l and
den
erva
ted
mus
cle.
(A
) Tw
o-st
ep co
mpe
titiv
e E
LIS
A w
as c
arrie
d ou
t as
desc
ribed
by
Dam
iani
et a
I.’*
In th
e fir
st s
tep,
a s
olut
ion
of a
nti-(
chic
ken f
ast
Ca-
ATP
ase)
ant
ibod
y (1
0 pg
lml)
was
pre
incu
bate
d ov
erni
ght w
ith a
n eq
ual v
olum
e of
mic
roso
mal
ves
icle
s at
the
conc
entra
tions
indi
cate
d on
the
absc
issa
. The
sec
ond
Step
w
as p
erfo
rmed
by
incu
batin
g atiq
uots
(0.2
ml)
of th
e an
tigen
-ant
ibod
y sol
utio
ns in
mic
rotit
er w
ells
coa
ted
with
inta
ct ra
bbit
SR v
esic
les.
Eac
h po
int i
s th
e av
erag
e va
lue
of d
uplic
ate
dete
rmin
atio
ns, S
ymbo
ls:
(0) co
ntro
l mus
cle,
(A) 2-
wee
ks d
ener
vate
d m
uscl
e, (
0) 4-
wee
ks d
ener
vate
d m
uscl
e. (
8) O
ne-
dim
ensi
onal
pep
tide
map
ping
was
per
form
ed a
ccor
ding
to C
leve
land
et a
1.l’
by u
sing
the
100
kDa
prot
ein
exci
sed
from
10%
pol
yacr
ylam
ide p
repa
rativ
e sl
ab g
els.
Dig
estio
n w
as c
arrie
d ou
t with
5 p
g of
S.
aure
us V
8 pr
otea
se. T
he p
rote
in fr
agm
ents
wer
e re
solv
ed in
a 1
5-20
% p
olya
cryl
amid
e lin
ear g
radi
- en
t gel
. The
sla
b ge
l was
sta
ined
with
Coo
mas
sie
blue
. App
roxi
mat
ely
20 p
g of
pro
tein
wer
e lo
aded
per
gel
. Key
to la
nes:
(1) c
ontro
l mus
cle,
(2) 1
-wee
k de
nerv
ated
mus
cle,
(2) 2
-wee
ks d
ener
vate
d m
uscl
e, (3) 4-
wee
ks d
ener
vate
d m
uscl
e; a
rrow
indi
cate
s pr
otea
se.

~~
Table 1. Comparison of biochemical properties of unfractionated SR and of very Iiqht, Iiqht, and heavy fractions from 15-davs control (C) and denervated (D) muscle:
-
Very light
C 0 30 t 0 02 (4) D 0 91 k 0 03 (4)
C 18 t 6 (6) D 87 t 20 (6)
C 0 5 4 * 11 (4) D 1 19 t 0 0 4 (4) C 0 04 t 0 01 (4) D 0 03 * 0 01 (4) C 1 93 2 0 41 (4) D 2 23 t 0 69 (4) C 6 81 rt 2 03 (4) D 6 97 t 1 56 (4)
D C D
Protein yield (mg/g m)
Cholesterol (nmolimg)
Mg-ATPase activity (pmol/min/mg)
CSiATPase pr ratio
Ca-ATPase activity (pnoliminimg, - A23187)
Ca-ATPase activity (pmolirninimg, + A23187)
Ca-loading rate (pmolirninimg, 25°C) C
100 kDa protein (percent of total protein)
Light Heavy Unfract. SR
0.52 2 0.05 (4) 0.42 t 0.04 (4)
11 t 1 (6) 21 2 8 (6)
0.42 ? 0.03 (4) 0.46 t 0.11 (4) 0.04 & 0.01 (4) 0.19 t 0.03 (4) 4.42 * 1.17 (4) 3.67 2 1 90 (4)
13.40 t 0.88 (4) 8.29 f 1.21 (4)
0.18 t 0.04 (4) 0.40 t 0 08 (4)
17 t 6 (5) 0.43 t 0.10 (4) 0.48 2 0.10 (4) 0.44 * 0.05 (4) 1.32 t 0.16 (4) 2.60 t 1.19 (4) 1.47 2 0.41 (4) 5.85 2 1.12 (4) 3.14 t 0.48 (4) 1.26 t 0.20 (5) 0.73 t 0.14 (5)
58 2 3 (6)
22 t 4 (4)
33 2 4 (7)
3.71 (3) 4.23 (3)
13 t 3 (5)
0.51 t 0.05 (4) 0.66 (3)
0.06 * 0.02 (5) 0.16 t 0.04 (5) 1.84 k 0.33 (4) 1.45 2 0.41 (4) 8.30 * 0.50 (4) 5.40 2 0.63 (4) 2.50 t 0.38 (4) 1.85 2 0.16 (4)
72 5 6 (5) 58 t 8 (5)
37 t 5 (5)
~ ~~
Note Microsomes were prepared as described in Materiais and Methods (see Ref 43) except that KCI treatment was omitted Protein yieid is given as milligrams of protein per gram of fresh weight muscle Mg-ATPase and Ca-ATPase activities were determined at 37"C, as described in Materials and Methods Values are means t S E M for the number of determinations on the different membrane preparations shown in parentheses
to account for the 20-30% decrease in average membrane density of Ca-ATPase and for the cor- responding decrease in Ca-ATPase activity and in Ca-loading rate of SR vesicles from 2-3 weeks denervated muscle. At these early stages after denervation, there is also no evidence from immu- nologic studies of the Ca-ATPase, as well as from Cleveland's peptide maps, which might suggest an
even partial transition from the predominant fast isoform of Ca-ATPase to the slow isoform having lower catalytic activity.44 Furthermore, our pre- sent results, and preliminary results obtained with purified junctional TC (Zorzato, Volpe, Quaglino, and Margreth, unpublished observations), demon- strate that muscle denervation does not primarily alter the efficiency of calcium transport rnecha-
VL
L
H $racking dye
FIGURE 8. Sedimentation profile and protein electrophoretic composition of isolated membranes from control and denervated muscle. (A) Microsomes from control (lane 1) and 2-weeks denervated (lane 2) muscles were fractionated by the method of Rosemblatt et al.39 Protein bands were collected at the 25-35% (very light fraction), 35-40% (light fraction), and 40-50% (heavy fraction) sucrose inter- faces of the gradient. (B) One-dimensional 10% SDS-PAGE of unfractionated SR membranes and of fractions obtained by sucrose density centrifugation as in panel A was carried out according to Laemmli.'' Slab was stained with Coomassie blue; 30 kg of protein were loaded per gel. Key: lanes 1 (unfractionated SR), 3 (very light fraction), 5 (light fraction) and 7 (heavy fraction) are from control muscle; lanes 2, 4, 6, and 8 are the corresponding fractions from 2-weeks denervated muscle.
Proliferation of Triads MUSCLE & NERVE December 1988 1255

1256 Proliferation of Triads MUSCLE & NERVE December 1988

? a b
0 0.01 0.1 1 lo m 0.1 I lo 100
Abs (pg/rnl)
- - - - - -
FIGURE 10. Immunological cross-reactivity with anti-(chicken TT Mg-ATPase) antibody of microsornes from control and de- nervated muscle. Membranes were obtained from control and 2- weeks denervated muscles as described in Fig. 8A. One-step ELISA was carried out in microtiter wells precoated with mem- branes at a protein concentration of 5 kg/ml. Anti-(chicken l 7 Mg-ATPase) antibody was used at the concentrations indicated on the abscissa. Symbols: (0) control muscle; (m) denervated muscle. Key to panels: (A) total SR; (B) very light fractions; (C) light fractions; (D) heavy fractions.
nism in the SR, since Ca/ATP coupling ratios are found to be in the upper range of nornmal values, under optimal assay conditions. The present re- sults are difficult to compare with earlier biochem- ical findings on the isolated SR from denervated niuscle of the and of the chicken51 due to differences in methods and since it is only recently that differences in membrane protein composition and functional properties between free and junc- tional SR have been elucidated. As a result of these studies, for instance, CS, long classified as a compartmental, extrinsic protein of the SR,6 is now regarded as a relatively specific marker of junctional SK.11.:’i’2‘1 Species-lin ked differences have likewise becoine known, such as differences in molecular size and in metachromatic staining
with Stains All, between skeletal CS of niaminalian and avian species,14 that need to be considered in reinterpreting earlier results. Thus, interestingly, Tate et al.” had reported a striking increase of a 55 kDa component in the isolated SR from dener- vated chicken pectoralis, which, unlike rabbit skel- etal CS (Mr 63 kDa), stained purple rather than blue after staining with Stains All, and was there- fore thought to be a different protein. These properties are, however, identical to those later re- ported for chicken CS.I4
The increase of junctional SR in rabbit gastroc- nemius after denervation appears to be explained by the increase in number of TC, as well as by an increase of the junctional contact area of TC with TT. T T proliferation might act as a morphoge- netic inducer for the formation of extrasynaptic connections with SR membranes, or the seemingly tight coordination between these processes could reflect a common mechanism.
The loss of motor innervation, rather than muscle inactivity, as such, appears to be responsi- ble for the hyperplasia of triads in muscle fibers, since the change appears to be readily reversible upon reinnervation of muscle fiber, after nerve crushing. Denervation-induced proliferation of TT is accompanied by an increased formation of nitrendipine-sensitive C a - c h a n n e l ~ , ~ ’ ~ ~ ~ which might be responsible for the increased influx of calcium, and for the increased concentration of in- tracellular calcium in denervated muscle fibers.” The augmented synthesis and accumulation of CS in the SR of denervated muscle might be caus- ally related to these changes, since calcium was shown to act as an inducer of CS synthesis in dif- ferentiating muscle.29 Interestingly, coated vesi- cles, recently shown to be involved in intrafiber transport of CS from the site of synthesis to the site of accumulation in the lumen of TC,52 are found in increased number in denervated gastroc- nemius fibers, occasionally adjacent to TC, as shown here.
A possible functional consequence of triad hy- perplasia and of the increase of intraluminal CS in the SR of inactivated muscle is that of increasing
FIGURE 9. (Opposite page) Electron microscopy of representative sections of HSR from control and denervated muscle. Thin section electron microscopy of heavy SR fraction from normal (A) and 15-days denervated (B) rat gastrocnemius muscle. In the fraction from normal muscle (25,OOOx), the majority of the vesicles appear empty, very few show ribosomes on their external surface, and the re- mainder contain electron-dense material corresponding to calsequestrin. After denervation (25,OoOx) most of the vesicles (about 90%) are filled with calsequestrin. (C) A higher magnification (150,000~) of heavy SR fraction from 15-days denervated rat gastrocnemius muscle. The lumen of the vesicles contains CS attached to the inner surface of the membranes in areas which present feet projections on the outer surface (arrows).
Proliferation of Triads MUSCLE & NERVE December 1988 1257

the steady-state concentration of calcium within the SR, which may be viewed as an adaptive change, by analogy with what we suggested for explaining the pathogenesis of tubular aggregates in hunian muscle.43 Furthermore, the calcium permeability of TC froin rabbit gastrocnenlius is
decreased after denel-vation, and that suggests the occurrence of concomitant changes in the proper- ties of calciurn release channels of junctional TC, which are n o w under investigation (Zorzato, Volpe, Quaglino, and Mai-greth, unpublished ob- servations).
REFERENCES 1. Barany hi. Barany K, Reckard T, Volpe A: Myosin of fast
and slow muscles of the rahhit. . 4 ~ r h Biut-hem Biop/+ 109:185- 1 9 1 , 1965.
3. Beeler TJ, Gable KS. Keffer JM: Characterization of the membrane bound Mg-ATPase of rat skeletal muscle. Bto- C / L I I I I Biophgs Acto 734:221-2Y1, 1983.
3. Biral D, Daniiani E, h i -g i - e th A. Scarpitii E: hlyosin sub- unit composition in human developing niiiscle. Biocheui J 224:%3-93 I , 1984.
4. Braiidl C.1, deLeon S. Martin S, MacLerrnan D H : Adult foi-ins of the (:a-ATPase of sarcoplasmic reticuluni. E x - pression i n developing skeletal muscle. J B i d Chrm 262:3768 - 3774, 19x7.
5 . Brandl CJ, Green NM, Korcrak B , MacLennari DII: Two Ca-ATPases genes: homologies arid niechmistic iniplica- tions of deduced amino acid sequences. Cell 44:597-607, 1986.
6. Campbell KP: Protein components and their roles in sarco- plasmic reticulum function, in Entmanti hlL, Van Winkle WB (eds). .Surco/hs?tiir Rctiriilrrm in hlusde P l i y o l o f l . Boca Raton, Florida. CR(: Pi-ess. 19x6. Vol 1. pp. 65-69.
7. Campbell KP, MacLeriiiari DH, Jorgenseii AO: Staining of thr Cktiinding proteins. calsequestrin, calmotluliri. rropo- nin c and S-100 with the cationic dye "Stains ,411." J Biol Chcrn 258: 11267- 11273, 1083.
8. Carraro U, Morale D, Miissini I , Lucke S, Cantini hl, Betto R, Cataiii C, Dalla Libera L, Danieli Betto D. Noventa L): Chronic denervation of rat hemidiaphragm: maintenance of fiber heterogeneity with associated incteasing unifor- mity of myosin is0forms.J Cell B i d 100:lfil- 174, 1985.
9. Chu A, Volpe P, Costello B, Fleischer S: Functional char- acterization of junctional terminal cisternae from Iiianinia- han fast skeletal muscle sarcoplasniic 1-eticulum. BtoChfwiJ- h? 25:8315-8324, 1986.
10. Cleveland. DW, Fischer SG. Kirschner MM', Laeminli UK: Peptide niapping h) limited proteolysis i i i sodium dodec-yl sulphare by gel electrophoresis. 1 Bzol Chrm 25?: 1 102- 1106. 1977.
11. Costello B, Chadwick C, Saito A. Chu A. hlaurer .4, Flei- scher S: Characterization of the juiictioiial facc nicnihraiie from terminal cisternae of sarcoplasinic rericuluni. J ( :d l B i d 103:741-753. 1986.
12. Damiani E. Betto R, Salvatori S, Volpe P, Salviati G. Mar- greth A: Polymorphism of sarcoplasinic reticulum adeno- sine triphosphatase of rahhit skeletal muscle. Biorhrrn J 197:245-248. 1981.
13. Damiani E, Margreth A, Furlan A, Dahms AS, Arnn J , Sabbadini R4 : Common sfructural donlains i n the sarco- plasniic reticulum Ca-ATPase and the ttansverse tubule Mg-ATP3se.J Cell B i d 104:461-47?. 1987.
14. Damiani E, Salvatori S, Zorzato F. Margreth A: Character- istics of skeletal muscle calsequestrin: comparison of mam- nialian, amphibian and avian muscles. 1 Muscle Res Ccll ,Mo- t i! 7:435-445. 1986.
15. Endo M : Calcium release from sarcoplasniic reticulum. Physiol Re11 57:71- 108, 1977.
16. Engel AG. Stonnington HH: Morphological effects of de-
IIervation of n1llsclr. A quantitative ultrastructural study. in DI-dchlllan DB (ed). 7',-op/ric Funcfio~7 01 the 1 ~ c ' 7 i J u J / . An- nals of the New York Acadeiriv of Scieirces, 1074. Vol. ?2X.
17. Fischer E: C;hanges in protein and enzymes in muscular degeneration suhsequrnr to denrrvation. .4vr J PIiuiml ,Me// 35:212-222, 1955.
18. Fraiizini-Ai-mstrong C:, Kenriey LI. \'ari-iano-Riartson E: I'he srriicture of cakeqiiestriii in triads of vei t eha te skele- tal muscle: a deepetch stu(ly. J C'd/ A i d lO5:49-.56, 19x7.
1 9 Jorgensen AO, Sheii AC'-Y, Campbell KP. MacLennan DH: L~lttastrtictu~~al localization of calsequestrin in rat skel- etal muscle by immunoferiitin lahelirrg of ultrathin frozen sectioith.J C r / / Liiol 97:1573- 1581. 1 9 S 3
20. Joi-gensen AO, Shen AC-Y. hlaclennan DH, Tokuyasu K T : l~ltrasti.iictiti~a1 localization of tile Ca-Mg-dependent ATPase o f sarcoplasniic reticulum in Iat skeletal muscle by ferriun labeling of ultrathin fr-ozen sections. J Ctd/ B I ~
21. Kirby A(:. Lindley BD: Calciuni content of' rat last and slow alter denervation. COIII /J Bcor/wnr P/i .y~zu/ iOA:583- 5Sti. 1981.
22. Laemmli LIK: Cleavage of sti urtiii-dl pic>trins during the assembly of the I i ~ d o f bacteriophage T4. NatvIc' (Loud) 227:680- 684 ~ 1970.
23. Lau Y. C:;iswell AH, Brunschwig JP. Baerwald R Y , Garcia R h l : Lipid analysis and freere-ti-ac-turr srudies on isolated transverse tubules and rarcoplasmic I-eticulurri subfmctioms of skeletal muscle. J Biol C ~ P I J ! 254:540-546, 1979.
2-1. Lowry OH, Rosebrough NJ. F a n AL, Randall R,J: Proteln measurenients with the Foliu pheirol reagent. J B i d (;heJJl
193:365-275, 1951. ?5. Margreth A, Salwtori S, Daniiani E. Salviati G: Isomorfimi
of skeletal sai-roplasmic reticuluni (:a-ATPa.se in yelation t o fiber types. i i i Carpenedo F, Debctto P. Floi-eani M . Lu- ciaiii S, (eds), Prorrc.diti,q 01 ~ l J / C ? l l / l ~ l V l ~ f d Syiposiuttr UII . (:d- rii i in lo J J : .Alrrrrhiuiw 1 rtirrrpoi-1 nnd Ccdli i lo~ Rcgtrluliuti. I YR6.
?G. Margreth A , Salviati C;, DI hlauro S. T'ii~ati C, : Early h o - chemical corisqtiences ot' denci~\aiiort in fast and slow skeletal niurcle and their rehtionxhip to neural control over niiiscle differentiation. Biorhem ./ 1?6:109% 1 1 10, 1952.
27. Margreth .4. Salviati G, hlussini I . Cat-raro L': L"ltrastl-tlc- ture a n d niacromolecular coniposition o f the sarcoplasmlc reticuliini in relation to the t\\itch characteristics of skeletal muscles, in A. T. Milhorac (etl), E x p l o r a t o ~ (;onccpt.r 111 M I L S - r d n t DutIop/i? I I . Amsterdani. Escerpta hledica, 1974, pp 406- 4 18.
28. hlartonosi A: Transport of calcium by sarcoplasniic reticu- luni. in Cheung M'Y (ed). Calriitni mid C d f Functiori. Sali Di- ego, Acatleniic- Press. 1982. Vol. I l l . pp 57- 102.
29. hfartoriosi A : A possihle I O ~ For cytoplasmic (Ca) in the regulation of the synthesis of sat-coplasmic reticulum. in Martonosi A (ed), Memiit-utie c i d T J - Q ~ A ~ O ~ . ~ . New Y o r k , Ple- num. 1982, Vol. I , pp 593-600.
90. Meissner G : Isolation and characterization of two rypes ot
pp 68-88.
92:409- 4 14, 1 Y x2.
pp 75-86.
1258 Proliferation of Triads MUSCLE 8 NERVE December 1988

31.
32.
33.
34.
35.
36.
37.
38.
39.
40
41
sarcoplasmic reticulum vesicles. Biochim Biaplfj.! Artu 389 :.?I 1 - 68, 1 97 3 . Merril CR, Goldman D, Sedmari SA, Ebert MH: Ultrasen- sitive stain for proteins in polyacrylaniide gels shows re- gional variation in cerebrospinal fluids proteins. Science i?11:1437- 1438, 1981. Michalak M, Campbell KP, MacLennan DH: Localization of the high affinity calcium binding protein and of an in- trinsic glycoprotein in sarcoplasmic reticulum membranes. J Bzol Chem 255:1317-1326, 1980. Mitchell RD, Volpe P, Palade P, Fleischer S: Biochemical characterization, integrity, and sidedness of purified skele- tal muscle triads. J Biol Chem 258:9867-9877, 1983. Muscatello U, Margreth A, Aloisi M: On the differential response of sarcoplasni and myoplasm to denervation in frog muscle. J Cell Biol 27:l-24, 1965. Okamoto VR, Moulton MP, Runte EM, Kent CK, Lebherz HH, Dahms AS, Sahbadini RA: Characterization of trans- verse tubule membrane proteins: tentative identification of the Mg-ATPase. Arcli Biochfm Biophy.5 237:43-54, 1985. Palexas GN, Savage N, Isaacs H: Characteristics of sarco- plasmic reticulum from normal and denervated rat skeletal muscle. Biochem J 200: 11 - 15, 1982. Pellegrino C, Franzini C: An electron microscope study of denervation atrophy in red and white skeletal muscle fi- bers.] Cell B i d 17:327-349, 1963. Pierobon-Borniioli S, Armani hl, Ringel SP, Angelini C, Vergani L, Betto R, Salviati G: Faniilial neuromuscular dis- eases with tubular aggregates. M u s c l e Nerue 8:29 1-298, 1985. Rosemblatt hl, Hidalgo C, Vergara C, Ikemoto N: Immu- nological and biochemical properties of transverse tubule membranes isolated from rabbit skeletal muscle. J B i d Chem 256:8140-8148, 198 1. Sabhadini RA, Okamoto V: The distribution of AI'Pase ac- tivities in purified transverse tubular membranes. Arch Bzo- chrm Biophy 223:107-119, 1983. Saito A, Seiler S, Chu A, Flrischer S: Preparation and mor- phology of sarcoplasniic reticulum terminal cisternae from rabbit skeletal muscle. I (,'el/ Rzol 99:875-885. 1984.
Sarcoplasmic reticulum origin, calcium storage ability and functional implications. Muscle N t w e 8:299- 306, 1985.
44. Salviati G, Volpe P, Salvatori S, Betto R, Daniiani E, Mar- greth A, Pasquali-Konchetti l: Biochemical heterogeneity of skeletal-muscle microsomal membranes. hlenihrane ori- Fin, membrane specificity and fiber types. B7<Jrhem J LO2:289-30 1, 1982.
45. Schiaffino S, Margreth A: Coordinate development of the sarcoplasmic reticulum and T system during postnatal dif- ferentiation of rat skeletal muscle. J Cell Bzol 41 :855-875,
ffino S, Settembrini P: Studies on the effects of de- nervation in developing muscle. I . Differentiation of sar- cotubular system. Virchows Arch. B 4:345-356, 1970.
47. Schniid A, Kazazoglou T, Renaud JF, Lazdunski M: Com- parative changes of levels of nitrendipine Ca-channels, of tetrodoxin-sensitive Na-channels and of ouahain-sensitive (Na-K)-ATPase following denervation of rat and chick skeletal niuscle. FEBS Lett 172: 114- 118, 1984.
48. Schniid A, Renaud JF, Fosset M, Meaux JP, Lazdunski M: The nitreridipine-sensitive Ca-channel in chick muscle cells arid its appearance during niyogenesis in vitro and in vivo. J B i d Chem 2.59: 11366- 11372, 1984.
49. Soninier JR: The anatomy of sarcoplasmic reticulum in vertebrate skeletal muscle: its implication for excitation contraction coupling. 2 Naturfoi-sch 37:665-678, 1982.
50. Suninicht GE, Sabbadini RA: Lipid composition of trans- verse tubular membranes from normal and dystrophic skeletal muscle. Arch Bioch~in ifiophys 215:628-637, 1982.
51. Tate CA, Bick RJ, Myers TD, I'itts BJ, Van Winkle WB, Entmann XIL: Alteration of sarcoplasrnic reticulum after denervation of chicken pectoralis muscle. Riochem J 210:339-344, 1982.
52. 'l'honias KM, Benson RJJ, Fine RE: 'The role of coated ves- icles in the development of the sarcoplasmic reticulum. J Cell B i d 105: 122a, 1986.
53. V ~ l p e P, Biral D, Damiani E, hlargreth A: Characteriza- tion o f human muscle myosin with respect to the light chains. Bzocliem J 195:25 1 - 2.58, I 98 1.
i 4 . Volpe P, Daniiani E, Salviati C , Margreth A: Transition in mernbrane coniDosition during postnatal development of
~~~~ ~~ ~~
42. Salvatori S, Damiani E, hlargreth A, Fassina G, Caparrotta L: Effects of prostacyclins 011 sarcoplasmic reticulum Ca- pump, in Braquet P. Garay RP, Frolich .JC, Nicosia S, eds), Prostngkirkdzns aud MenibIar7es Ion ?-runsport. New York,
43. Salviati G, Pierobon S, Betto R, Daniiani E, Angelini C, Ringel SP, Salvatori S, Margreth A: Tubular aggregates:
rabbit fast n1tlscie.J Alusclr Kn"C;ll Motil 3:213-i30, 1982. 5.5. Warren GB, Toon PA, Birdsall NJhI, Lee AG, Metcalfe
JC: Reconstitution of a calcium pump using defined mem- brane components. pro^ Natl Acnd Sci LISA 71 :622-626, 1974. Raven Press, 1984, pp. 209-2 16.
Proliferation of Triads MUSCLE & NERVE December 1988 1259




















