
Dysregulation of HSG triggers vascular proliferative disorders
Upload
independentCategory
view
2download
0
Pergamon J. Steroid Biochem. Molec. Biol. Vol. 63, No. 4-6, pp. 211-218, 1997
© 1997 Published by Elsevier Science Ltd. All rights reserved Printed in Great Britain
P I I : S 0 9 6 0 - 0 7 6 0 ( 9 7 ) 0 0 1 2 2 - 2 0960-0760/97 S19.00 + o.00
Expres s ion of Novel Genes Linked to the A n d r o g e n - i n d u c e d , Prol i ferat ive Shutof f in
Prostate Cancer Ceils Peter Geck, Jozsef Szelei, Jesus Jimenez, Tien-Min Lin,
Carlos Sonnenschein and Ana M. Soto* Department of Anatomy and Cell Biology, Tufts University School of Medicine, Boston, MA 0211 I, U.S.A.
Androgens control cell numbers in the prostate through three separate pathways: (a) inhibit ion of cell death, (b) induct ion of cell prol iferat ion (Step-l) and (c) inhibit ion of cell proliferat ion (Step-2, proliferative shutoff). The mechan i sms under lying these phenomena are incompletely unders tood. The h u m a n prostate carc inoma LNCaP variants express these pathways as follows: LNCaP-FGC express both steps, LNCaP-LNO expresses Step-2, LNCaP-TAC expresses Step- l , and LNCaP-TJA cells express nei ther step. These cells facil i tated the search for media tors of the androgen- induced proliferative shutoff pathway. Androgen exposure for 24 h or longer induced an irreversible prolif- erative shutoff in LNCaP-FGC cells. The Wang and Brown approach for identifying differentially expressed mRNAs was used to search for media tors of Step-2. Ten unique inserts were identified and f rom those ten, three genes were fur ther studied. The basal expression of these genes in shut- off-negative var iants was not affected by androgen exposure. They were induced by androgens in shutoff-posit ive LNCaP var iants and the androgen receptor- t ransfected, shutoff-positive, MCF7- AR1 cells. These genes were induced only in the range of androgen concentrat ions tha t elicited the shutoff response. Time course analysis showed that their induct ion precedes the c o m m i t m e n t point by 12-I8 h. In addit ion, they were expressed in the normal prostate during proliferative shutoff. These features suggest that the candidate genes have a role in the regulat ion cascade for prolifera- tive shutoff. © 1997 Publ ished by Elsevier Science Ltd. All rights reserved
J. Steroid Biochem. Molec. Biol., Vol. 63, No. 4-6, pp. 211-218, 1997
INTRODUCTION
Cell numbers in the prostate are regulated by andro- gens through three distinct pathways: (a) inhibition of cell death (apoptosis) [1], (b) induction of cell pro- liferation (Step-l), and (c) inhibition of cell prolifer- ation (proliferative shutoff, Step-2) [2,3]. Two of these mechanisms [(a) and (b)] have been used for therapeutic manipulation in prostate cancer by andro- gen ablation [4, 5]; the potential therapeutic use of the proliferative shutoff mechanism (c) remains unex- plored. The complexity of interactions among various cell types in animals hinders the understanding of these androgen-mediated phenomena. Established
*Correspondence to A. M. Soto. Tel: 636 6954; Fax: 636 6536; e-mail: [email protected].
Received 12 May 1997; ~ccepted 21 Jul. 1997.
21
hormone target cell lines together with recombinant DNA technology now offer an opportunity for testing hypotheses regarding the control of cell proliferation, without being affected by variables present in intact animals. The human prostate LNCaP-FGC cell line has been extensively used for this purpose. These cells are inhibited from proliferating by sex-steroid stripped human serum (CDHuS); low levels of andro- gens reverse this effect (Step-l) and at higher concen- trations androgens induce an irreversible proliferative shutoff (Step-2) [3,6]. LNCaP variants expressing only one of these two androgen-controlled pathways were isolated. From LNCaP-FGC cells, the LNCaP- TAC variant that expresses Step-1 only was selected [7]. The LNCaP-LNO cells proliferate maxi- mally in the presence of CDHuS, while responding to androgens by undergoing G0/G~ arrest. Finally, the
1

212 Peter Geck et al.
LNCaP-TJA variant is impervious to the inhibitory effect of both CD serum and androgens [6].
By using a novel differential subtractive amplifica- tion procedure [8], we are now able to describe candi- date genes responsible for the androgen-mediated proliferative shutoff.
M A T E R I A L S A N D M E T H O D S
Cell lines and tissue culture protocols
L N C a P - F G C cells and L N C a P - L N O cells derived from a metastatic lymph node of a patient with pros- tate adenocarcinoma were generously supplied by Dr. Julius Horoszewicz from the State University of New York at Buffalo [9]. The L N C a P - L N O variant was continuously maintained in 5% CD stripped fetal bovine serum (FBS) (HYCLONE, Logan, UT) sup- plemented to Dulbecco's modified Eagle medium (DME) (GIBCO-BRL, Gaithersburg, MD) [10]. The L N C a P - F G C , TAC and TJA variants were routinely grown in 5% FBS [6,7]. The MCF7-AR1 cell line resulted from transfection of the androgen receptor into the MCF7 cell line; it was propagated routinely in 5% F B S [ l l ] .
Rat prostate experiments
Sprague-Dawley male rats (200g) were used (Taconic, Germantown, NY). Ventral lobes of intact, castrated and castrated rats treated with testosterone propionate for 3 and 7 days [2] were dissected and frozen on dry ice and used to prepare RNA (see below).
Cell proliferation analysis
The time-course of androgen exposure required for commitment to express the shutoff phenotype was studied in L N C a P - F G C cells. 40 000 cells/well were seeded in 12-well plates (Costar, Cambridge, MA) in 5% FBS. Medium was changed after 4 8 h to 5% CDHuS in phenol red-free DME. Cells were treated with 3 0 n M testosterone (T) for 0 to 120h. Subsequently, the medium was removed, cells were washed twice with D M E and fresh 5% C D H u S con- taining 30 nM estradiol (E2) was supplied to allow proliferation of cells not committed to the androgen- induced shutoff. Cells were lysed, and the nuclei were counted by a Coulter Counter Model ZM (Coulter, Hialeah, FL) [6].
The Wang-Brown differential amplification
This procedure first amplifies short fragments of cDNAs. Then, successive subtractions and amplifica- tions between the two cDNAs result in differential sequence pools in both directions [8] (Fig. 1). L N C a P - F G C cells expressing androgen-induced pro- liferative shutoff were harvested after 24 h of 30 nM R1881 treatment. R1881 is a synthetic, non-metab-
Shutoff* cDNA (R)
RsaI+AluI digestion adaptor ligation PCR
> +cDNA
Long hybridization
l +I cDNA
Short hybridization PCR
I *2 cDNA
I repeat procedure
cDNA-BD (shutoff- negative, biotinyl.) (CD)
cDNA-BD (CD)
Shutoff* > probe
subtracted library (R) (shutoff*)
Cloning into pCRII vector
Fig. 1. Subtrac t i ve ampl i f i ca t i on o f a n d r o g e n - i n d u c e d s e q u e n c e s . S h u t o f f ~ R N A w a s p r e p a r e d f r o m L N C a P - F G C ce l l s t rea ted wi th 3 0 n M R1881 for 2 4 h ; the R (for R1881- treated) s e q u e n c e s originate f rom this preparat ion. 'Dr iver ' c D N A w a s p r e p a r e d f r o m L N C a P - F G C ce l l s grown in CDHuS, a c o n d i t i o n in w h i c h the pro l i ferat ive shuto f f is no t e x p r e s s e d ; the shuto f f -nega t ive c D N A s e q u e n c e s are origi- na ted f rom this pool, called C D (for C D H u S - t r e a t e d ) . A s i m i l a r pro toco l w a s p e r f o r m e d to ident i fy s e q u e n c e s that w e r e s e l ec t i ve l y e x p r e s s e d in ce l l s m a d e q u i e s c e n t by ex- p o s u r e to C D H u S ; n a m e l y , shuto f f -nega t i ve c D N A s f r o m L N C a P - F G C ce l l s g r o w n in C D H u S w e r e s u b t r a c t e d with
c D N A s d e r i v e d f r o m shuto f f +, R1881- t rea ted cells.
olized androgen (methyltrienolone, RousselI-UCLAF, Romainville). A shutoff-negative control was obtained by using L N C a P - F G C cells harvested after 3 days of CD H u S treatment. Total RNA was prepared by the acidic guanidinium-thiocyanate method [12] and poly-A + RNA was purified by the FastTrack kit (Invitrogen, San Diego, CA). Double-stranded cDNA pools from both RNA preparations were synthesized using the Copy Kit (Invitrogen), with oligo-dT prim- ing. A 2 #g aliquot of cDNA was digested by AluI and RsaI enzymes; specific adapters with EcoRI and PCR primer sites were ligated to the ends, and the constructs were PCR amplified (GeneAmp Kit, Perkin Elmer, Foster City, CA). 50 /~g aliquots of amplified D N A were digested with EcoRI, photobioti- nylated (driver cDNA) and hybridized to a 2 #g ali- quot of non-biotinylated cDNA from the other pool. Long and short hybridizations were performed to increase subtraction efficiency for low and high abun- dance transcripts [8]. After 3 cycles, the final ampli- fied pools of the androgen-induced shutoff (R) and
Gene Expression in Prostate Cancer Arrest 213
serum arrested (CD) sequences were digested with EcoRI, and ligated into the BlueScript SK vector (Stratagene, La Jolla, CA) for transformation into E. coli (OneShot strain, Invitrogen). Colonies were trans- ferred to nylon filters and differentially screened using radiolabelled subtracted R and CD pools as probes. Multiple cross-hybridizations of dot-blot filters were used to investigate relatedness among cloned inserts.
DNA sequence analysis
PCR sequencing reactions were performed using the dsDNA Sequencing System (Life Technologies, Gaithersburg, MD). The DNA sequences were searched for homology to known sequences using the FASTA (Genetics Computer Group, Madison, WI) and BLAST (National Center for Biotechnology Information, Bethesda, MD) programs.
Northern blot analysis
Total RNA was prepared using the RNeasy kit (Qiagen, Chatsworth, CA). RNA samples (20 #g) were run in 1% agarose-formaldehyde gels and trans- ferred to nylon membranes using the Turbo-Blot System (Schleicher and Schuell, Keene, NH). The PCR amplified inserts were labelled by the Random Primed DNA Labeling Kit (Boehringer-Mannheim, Indianapolis, IN) and used as probes. Hybridizations were performed overnight at 68°C in ExpressHyb sol- ution (Clontech, Palo Alto, CA). Membranes were analyzed by PhosphorImager (Molecular Dynamics, Stanton, CA) using the ImageQuant program. Northern filters containing poly-A ÷ RNA preparations from an array of normal human tissues were pur- chased from Clontech. Hybridizations of rat Northern blots were at 65°C and the washing temperature was 48°C. RNA loading wets normalized to either glyceral- dehyde 3-phosphate dehydrogenase (GAPDH) or fl- actin (Clontech) signals in the same blot.
Logarithmic phase RT-f~CR
This procedure was used to measure the expression of low copy number transcripts [13-15]. 2 #g aliquots of total RNA were used as templates for cDNA syn- thesis with gene specific primers (cDNA Synthesis Modul, Invitrogen). GAPDH levels were measured first to normalize the: assay for subsequent exper- iments. PCR reactions were performed with the GeneAmp kit. Primers were labelled with 32p T-ATP. From the 50 #1 reaction volume, 7 #1 aliquots were withdrawn between the 15th and 35th cycles at vari- able points, and resolved by polyacrylamide gel elec- trophoresis. Patterns were analyzed by Phosphorimager. Optical densities of the bands were integrated and plotted against the amplification factor. Only data within the logarithmic phase were used for further analyses.
R E S U L T S
Time-course of induction of the proliferative shutoff
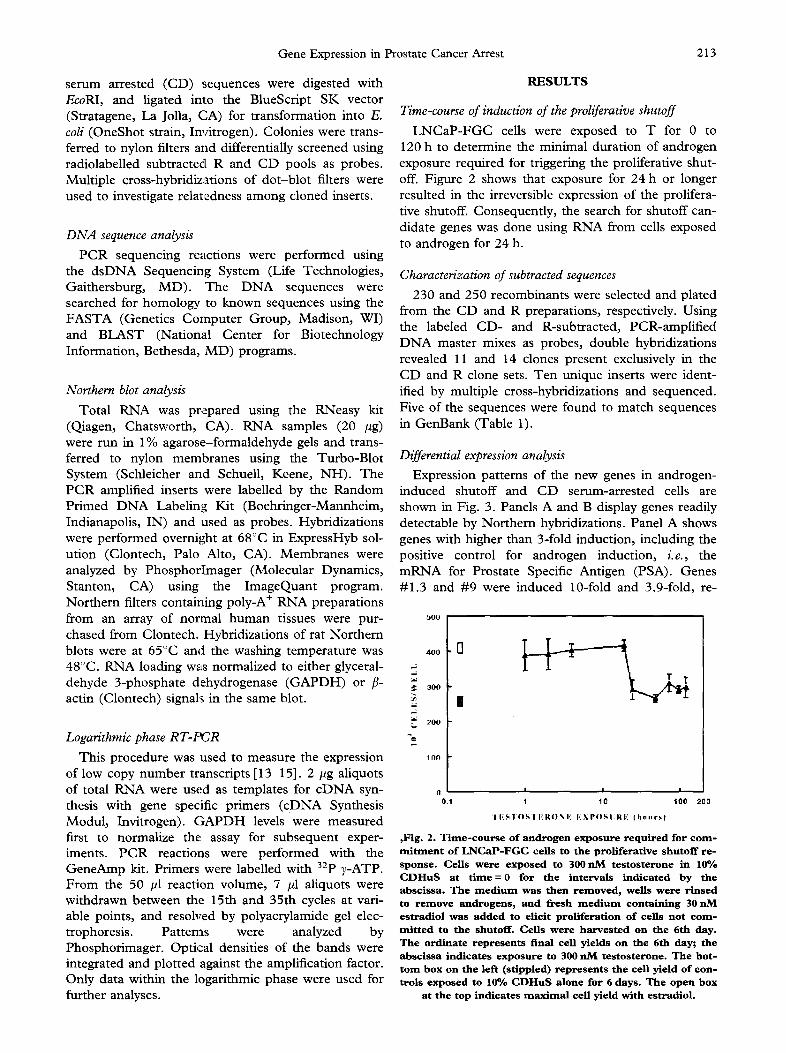
LNCaP-FGC cells were exposed to T for 0 to 120 h to determine the minimal duration of androgen exposure required for triggering the proliferative shut- off. Figure 2 shows that exposure for 24 h or longer resulted in the irreversible expression of the prolifera- tive shutoff. Consequently, the search for shutoff can- didate genes was done using RNA from cells exposed to androgen for 24 h.
Characterization of subtracted sequences
230 and 250 recombinants were selected and plated from the CD and R preparations, respectively. Using the labeled CD- and R-subtracted, PCR-amplified DNA master mixes as probes, double hybridizations revealed 11 and 14 clones present exclusively in the CD and R clone sets. Ten unique inserts were ident- ified by multiple cross-hybridizations and sequenced. Five of the sequences were found to match sequences in GenBank (Table 1).
Differential expression analysis
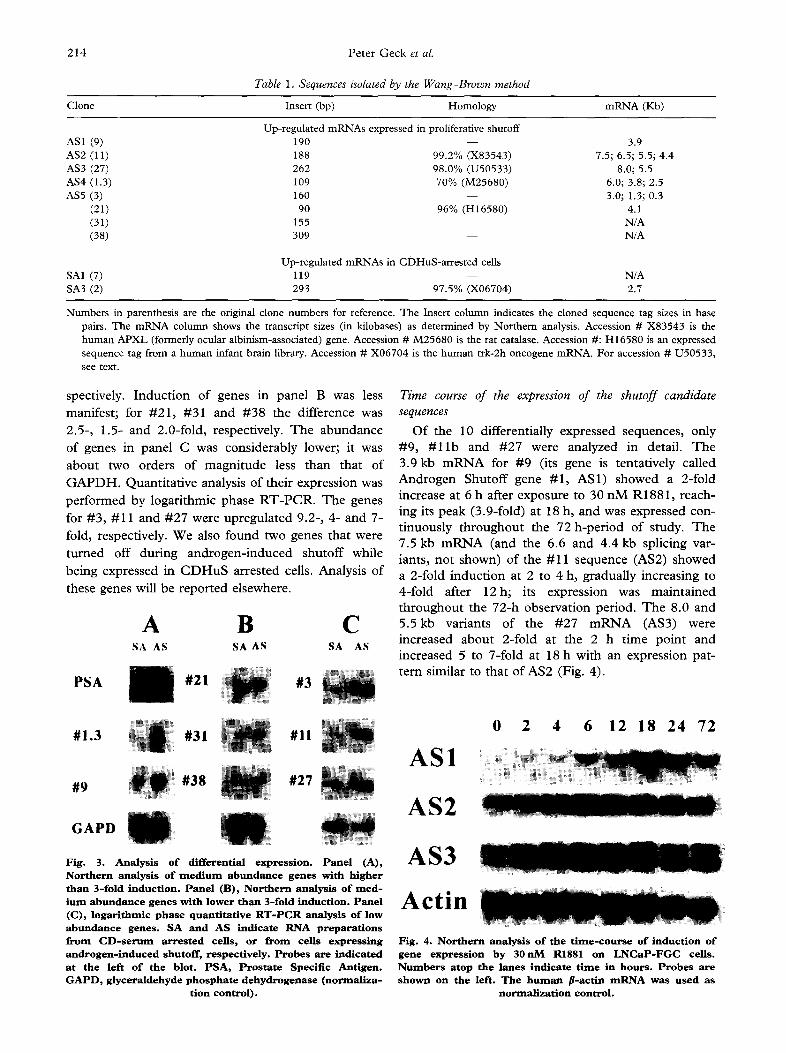
Expression patterns of the new genes in androgen- induced shutoff and CD serum-arrested cells are shown in Fig. 3. Panels A and B display genes readily detectable by Northern hybridizations. Panel A shows genes with higher than 3-fold induction, including the positive control for androgen induction, i.e., the mRNA for Prostate Specific Antigen (PSA). Genes #1.3 and #9 were induced 10-fold and 3.9-fold, re-
~00
400
30o
'-~ ~00
%
[]
100
N I I !
0.1 ~ 10 100 200
'I I .~STO,~I E R O N E E X P O S U R E ( h o u r s )
,Fig. 2. Time-course of androgen exposure required for com- mi tment o f L N C a P - F G C ce l l s to t h e p r o l i f e r a t i v e s h u t o f f r e - s p o n s e . C e l l s w e r e e x p o s e d to 300 n M t e s t o s t e r o n e in 10% C D H u S at t i m e = 0 for t h e i n t e r v a l s i n d i c a t e d b y t h e a b s c i s s a . T h e m e d i u m w a s t h e n r e m o v e d , w e l l s w e r e r i n s e d to r e m o v e a n d r o g e n s , a n d f r e s h m e d i u m c o n t a i n i n g 30 n M e s t r a d i o l w a s a d d e d to e l i c i t p r o l i f e r a t i o n o f ce l l s n o t c o m - m l t t e d to t h e shutof f . C e l l s w e r e h a r v e s t e d o n t h e 6 th d a y . T h e o r d i n a t e r e p r e s e n t s f ina l ce l l y i e l d s o n t h e 6 th day; t h e a b s c i s s a i n d i c a t e s e x p o s u r e to 300 n M t e s t o s t e r o n e . T h e b o t - t o m b o x o n t h e l e f t ( s t ipp led) r e p r e s e n t s t h e ce l l y i e l d o f c o n - t ro i s e x p o s e d to 10% C D H u S a l o n e for 6 d a y s . T h e o p e n b o x
at t h e top i n d i c a t e s m a x i m a l ce l l y i e l d w i t h e s t r a d i o L
214 Peter Geck et al.
Table 1. Sequences isolated by the Wang-Brown method
Clone Insert (bp) Homology mRNA (Kb)
Up-regulated mRNAs expressed in proliferative shutoff AS1 (9) 190 - - 3.9 AS2 (11) 188 99.2% (X83543) 7.5; 6.5; 5.5; 4.4 AS3 (27) 262 98.0% (U50533) 8.0; 5.5 AS4 (1.3) 109 70% (M25680) 6.0; 3.8; 2.5 AS5 (3) 160 - - 3.0; 1.3; 0.3
(21) 90 96% (H16580) 4.1 (31) 155 N/A (38) 309 - - N/A
Up-regulated mRNAs in CDHuS-arrested cells SA1 (7) 119 - - N/A SA3 (2) 293 97.5% (X06704) 2.7
Numbers in parenthesis are the original clone numbers for reference. The Insert column indicates the cloned sequence tag sizes in base pairs. The mRNA column shows the transcript sizes (in kilobases) as determined by Northern analysis. Accession # X83543 is the human APXL (formerly ocular albinism-associated) gene. Accession # M25680 is the rat catalase. Accession #: H16580 is an expressed sequence tag from a human infant brain library. Accession # X06704 is the human trk-2h oncogene mRNA. For accession # U50533, see text.
spectively. Induction of genes in panel B was less manifest; for #21, #31 and #38 the difference was 2.5-, 1.5- and 2.0-fold, respectively. The abundance of genes in panel C was considerably lower; it was about two orders of magnitude less than that of GAPDH. Quantitative analysis of their expression was performed by logarithmic phase RT-PCR. The genes for #3, #11 and #27 were upregulated 9.2-, 4- and 7- fold, respectively. We also found two genes that were
turned off during androgen-induced shutoff while being expressed in CDHuS arrested cells. Analysis of these genes will be reported elsewhere.
PSA
#1.3
#9
A B C SA AS SA AS
/ #21
#31
#38
SA AS
#3
#11 ~
#27
GAPD
Fig. 3. Analysis o f differential express ion. Pane l (A), Northern analysis of m e d i u m abundance genes with higher than 3-fold induction. Panel (B), Northern analysis o f reed- Jura abundance genes with lower than 3-fold induction. Pane l (C), logarithmic phase quantitative R T - P C R analysis o f low abundance genes . SA and AS indicate RNA preparations from C D - s e r u m arrested cells, or from cells express ing androgen- induced shutoff, respectively. Probes are indicated at the left of the blot. PSA, Prostate Specific Antigen. G A P D , glyceraldehyde phosphate dehydrogenase (normal iza-
tion control) .
Time course of the expression of the shutoff candidate sequences
Of the 10 differentially expressed sequences, only #9 , # 1 1 b and #27 were analyzed in detail. T h e 3.9 kb m R N A for #9 (its gene is tentat ively called
A n d r o g e n Shutoff gene #1 , AS1) showed a 2-fold increase at 6 h after exposure to 30 n M R1881, reach- ing its peak (3.9-fold) at 18 h, and was expressed con-
t inuous ly t h roughou t the 72 h-per iod of study. T h e 7.5 kb m R N A (and the 6.6 and 4.4 kb splicing var-
iants, no t shown) of the #11 sequence (AS2) showed a 2-fold i nduc t i on at 2 to 4 h, gradual ly increasing to
4-fold after 1 2 h ; its expression was m a i n t a i n e d th roughou t the 72-h observat ion period. T h e 8.0 and 5 . 5 k b variants of the #27 m R N A (AS3) were increased abou t 2-fold at the 2 h t ime poin t and increased 5 to 7-fold at 18 h with an expression pat- te rn similar to that of AS2 (Fig. 4).
AS1
AS2
0 2 4 6 12 18 24 72
AS3
Act in
Fig. 4. Northern analysis o f the t ime-course of induction of gene express ion by 3 0 n M R1881 on L N C a P - F G C cells. Numbers atop the lanes indicate t ime in hours . Probes are shown on the left. The h u m a n p-actin m R N A was used as
normal izat ion control .
Gene Expression in Prostate Cancer Arrest 215
AS1
AS2
AS3
FGC LNO AR-I TAC TJA
1 2 1 2 1 2 1 2 1 2
iiii!i!!i!i!:!~ii~i!ii ¸ iii~iii!:i!!iiii~!~i~/i~ii~
i iiiiii!!~!~i i~//~i/!~! ~
Androgen concentration-dependent induction of candidate shutoff genes
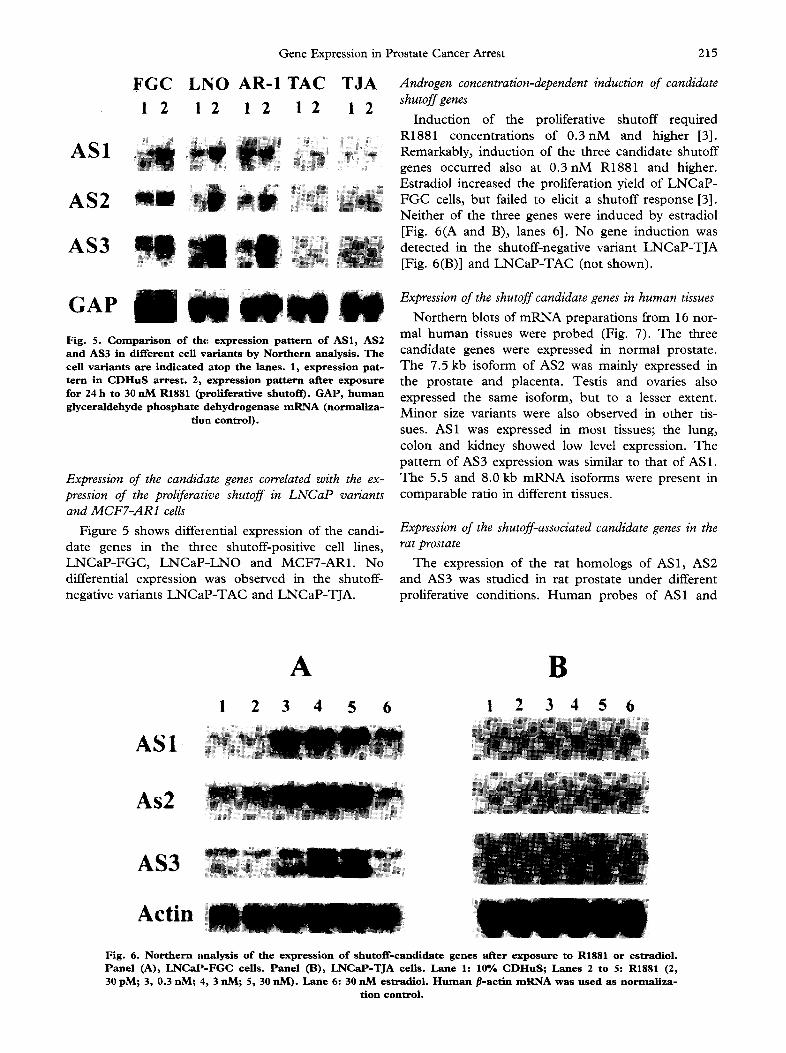
Induct ion of the proliferative shutoff required R1881 concentrations of 0 . 3 n M and higher [3]. Remarkably, induction of the three candidate shutoff genes occurred also at 0 . 3 n M R1881 and higher. Estradiol increased the proliferation yield of L N C a P - F G C cells, but failed to elicit a shutoff response [3]. Nei ther of the three genes were induced by estradiol [Fig. 6(A and B), lanes 6]. N o gene induction was detected in the shutoff-negative variant L N C a P - T J A [Fig. 6(B)] and L N C a P - T A C (not shown).
GAP I / Fig. 5. Comparison of the expression pattern of AS1, AS2 and AS3 in different cell variants by Northern analysis. The cell variants are indicated atop the lanes. 1, expression pat- tern in C D H u S arrest. 2, expression pattern after exposure for 24 h to 30 nM R1881 (proliferative shutoff). GAP, human glyceraldehyde phosphate dehydrogenase m R N A (normaliza-
tion control).
Expression of the candidate genes correlated with the ex- pression of the proliferative shutoff in LNCaP variants and MCF7-AR1 cells
Figure 5 shows differential expression of the candi- date genes in the three shutoff-positive cell lines, L N C a P - F G C , L N C a P - L N O and MCFT-AR1. No differential expression was observed in the shutoff- negative variants L N C a P - T A C and LNCaP-TJA.
Expression of the shutoff candidate genes in human tissues
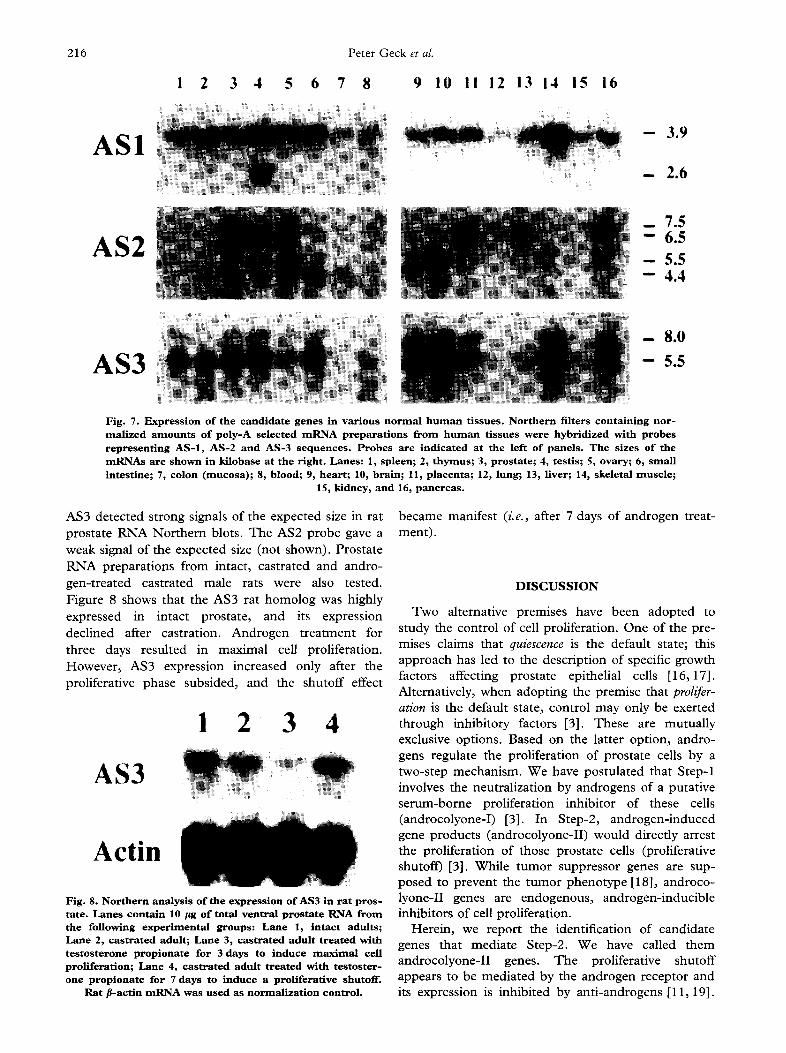
Northern blots of m R N A preparations f rom 16 nor- mal human tissues were probed (Fig. 7). The three candidate genes were expressed in normal prostate. The 7.5 kb isoform of AS2 was mainly expressed in the prostate and placenta. Testis and ovaries also expressed the same isoform, but to a lesser extent. Minor size variants were also observed in other tis- sues. AS1 was expressed in most tissues; the lung, colon and kidney showed low level expression. The pattern of AS3 expression was similar to that of AS1. The 5.5 and 8.0 kb m R N A isoforms were present in comparable ratio in different tissues.
Expression of the shutoff-associated candidate genes in the rat prostate
The expression of the rat homologs of AS1, AS2 and AS3 was studied in rat prostate under different proliferative conditions. H u m a n probes of AS1 and
AS1
As2
AS3
Actin
A I 2 3 4 5 6
B ! 2 3 4 5 6
Fig. 6. Northern analysis o f the expression of shutoff-candldate genes after exposure to R1881 or estradiol. Panel (A), LNCaP-FGC cells. Panel (B), LNCaP-TJA cells. Lane 1: 10% CDHuS; Lanes 2 to 5:R1881 (2, 30 pM; 3, 0.3 nM; 4, 3 nM; 5, 30 nM). Lane 6 :30 nM estradiol. H u m a n fl-actin m R N A was used as normaliza-
tion control.
216 Peter Geck et al.
! 2 3 4 5 6 7 8 9 10 I I 12 13 14 15 16
ASI - - 3 . 9
- - 2 . 6
AS2 i! ! :iii ¸ iiiiiiiiiii ! _ 7 .5 - - 6 . 5
- - 5 . 5 - - 4 . 4
AS3 - 8 . 0
- - 5 . 5
Fig. 7. Expression of t h e c a n d i d a t e genes in various normal human tissues. Nor thern filters containing nor- realized amounts of poly-A selected mRNA prepara t ions f rom h u m a n t i s s u e s w e r e hybridized with probes r e p r e s e n t i n g AS- l , AS-2 and AS-3 sequences. P r o b e s a r e indicated at the left of panels. The sizes of t h e mRNAs are shown in kilobase at the right. Lanes: 1, spleen; 2, thymus; 3, prostate; 4, testis; 5, ovary; 6, small i n t e s t i n e ; 7, colon (mucosa); 8, blood; 9, heart ; 10, brain; 11, placenta; 12, lung; 13, Hver; 14, skeletal m u s c l e ;
15, kidney, and 16, pancreas.
AS3 detected strong signals of the expected size in rat prostate RNA Northern blots. The AS2 probe gave a weak signal of the expected size (not shown). Prostate RNA preparations from intact, castrated and andro- gen-treated castrated male rats were also tested. Figure 8 shows that the AS3 rat homolog was highly expressed in intact prostate, and its expression declined after castration. Androgen treatment for three days resulted in maximal cell proliferation. However, AS3 expression increased only after the proliferative phase subsided, and the shutoff effect
1 2 3 4
AS3
Actin
Fig. 8. Nor thern analysis of the expression of AS3 in r a t p r o s - ta te . Lanes contain 10 /~g of total ventral prosta te RNA from t h e following exper imental groups: Lane 1, intact adults; Lane 2, cas t ra ted adult; Lane 3, c a s t r a t e d a d u l t t r e a t e d with testosterone propionate for 3 days to induce maximal cell proliferation; Lane 4, c a s t r a t e d a d u l t t r e a t e d w i t h t e s t o s t e r - o n e propionate for 7 days to induce a p r o l i f e r a t i v e shutof f .
Ra t #-actin mRNA was used as normal izat ion control.
became manifest (i.e., after 7 days of androgen treat- ment).
D I S C U S S I O N
Two alternative premises have been adopted to study the control of cell proliferation. One of the pre- mises claims that quiescence is the default state; this approach has led to the description of specific growth factors affecting prostate epithelial cells [16,17]. Alternatively, when adopting the premise that prolifer- ation is the default state, control may only be exerted through inhibitory factors [3]. These are mutually exclusive options. Based on the latter option, andro- gens regulate the proliferation of prostate cells by a two-step mechanism. We have postulated that Step-1 involves the neutralization by androgens of a putative serum-borne proliferation inhibitor of these cells (androcolyone-I) [3]. In Step-2, androgen-induced gene products (androcolyone-II) would directly arrest the proliferation of those prostate cells (proliferative shutoff) [3]. While tumor suppressor genes are sup- posed to prevent the tumor phenotype [18], androco- lyone-II genes are endogenous, androgen-inducible inhibitors of cell proliferation.
Herein, we report the identification of candidate genes that mediate Step-2. We have called them androcolyone-II genes. The proliferative shutoff appears to be mediated by the androgen receptor and its expression is inhibited by anti-androgens [11, 19].

Gene Expression in Prostate Cancer Arrest 217
To identify candidate genes that would code for inhi- bition of cell proliferation, we based our strategy on a subtractive system whereby two populations of the same human prostate I_2',ICaP cell line were prolifera- tively arrested by two different means; namely, CD serum (Step-l) and androgen (Step-2). These two agents act through pathways that are segregated in discrete L N C a P variants. This strategy was expected to eliminate the background noise and the down- stream common effector elements of the two regulat- ory pathways. In theory, the genes that ultimately remained in the subtracted pools were expected to be specific, high-ranking elements in the regulation of proliferation, and specifically of its inhibition.
Expression levels of' the genes identified by this approach are in the medium (AS1) and low (AS2 and AS3) copy number range. After androgen induction, AS1 is expressed in 200-300 copies/cell, AS2 is pre- sent in 40-50 copies/cell, and the AS3 copy number is about 60-80 copies/cell. These estimates stem from adopting G A P D H copy numbers as normalization control [20].
Expression patterns of the androcolyone-II candi- date genes were also investigated among different human tissues. Prostate and placenta were the organs where the full-size AS2 transcript was preferentially expressed. The expression pattern of the AS3 gene correlated positively with the occurrence of the prolif- erative shutoff in norraal rat prostate [2]. These fea- tures together with the data obtained using the L N C a P cell variants and the MCF7-AR1 cell line warrant pursuing further studies on these proliferation control candidate genes. Expression of AS1 and AS3 occurred in most of the tissues tested. It would be premature, however, to rule out their role in the androgen-induced sh~ttoff based on this expression pattern.
Several families of genes have been linked to the proliferative arrest of a few cell types in culture conditions [21-24]. It is unclear, however, whether or not the G A D D , gas and D D I family of genes are re- lated in any way to the shutoff genes described here, based on the lack of homology found so far. The AS2 sequence is homologous to a gene (APXL) originally reported in the human ocular albinism syndrome, an X-linked genetic disorder with retinal hypopigrnentation [25]. The gene is not involved in the syndrome, however, and its loss from the genome has been associated with sterility due to low sperm counts. The C-terminal of the putative polypeptide shares homology with the APX gene of Xenopus, a regulator of sodium channel proteins. Another domain of A P X L shows homology with syntrophin proteins that form a complex with dystrophins. The putative polypeptide has no membrane spanning domain, it has several phosphorylation sites and leu- cine zipper patterns. Its inducibility by androgens together with hypospermia-sterility upon deletion in-
dicate a role in endocrine-reproductive regulation. The AS3 sequence is homologous with a short partial transcript (CG008) in the BRCA2 region on chromo- some 13. This region is frequently lost in familial breast cancer, in leukemias and in some prostate can- cer cases [26]. The putative open reading frame of AS3 shows protein kinase motifs and nuclear localiz- ation signals, but it is not homologous with other known sequences.
The observations described here regarding the androgen-induced proliferative shutoff in culture con- ditions are relevant as well to diagnostic and thera- peutic approaches to prostate cancer patients; they point to a novel set of genes whose expression may be manipulated to control the proliferation of prostate cancer cells in situ.
In summary, our results represent the first efforts in a detailed analysis of androgen-induced, proliferative shutoff genes in the human prostate. Their expression appears to be closely associated with the shutoff phenomenon, as suggested by the expression pattern of these genes in shutoff-negative and positive L N C a P variants and MCF7-AR1 cells; they are induced only in the range of androgen concentrations that elicits the shutoff response. Time course analysis showed that their induction precedes the commitment point by 12-18 h, and this feature suggests a role for these genes in the regulation cascade for proliferative shutoff in the prostate.
Acknowledgements This work was supported in part by PHS NIH grants CA-55574, CA-13410, AG-13807 and NSF DCB-8711746. J.J. was a recipient of a postdoctoral fellowship from Fundacion Ramon Areces (Madrid, Spain). The able technical help of Marcelle Desronvil, Anita Oles, Mariana Fernandez and Maria Luizzi is acknowledged. We are grateful to Dr. Marion Gordon for her suggestions. The assistance of The Center for Reproductive Research at Tufts University (P30 HD 28897) is acknowledged.
R E F E R E N C E S
1. Isaacs J. T., Antagonistic effect of androgen on prostatic cell death. Prostate 5 (1985) 545-557.
2. Bmchovsky N., Lesser B., Van Doom E. and Craven S., Hormonal effects on cell proliferation in rat prostate. Vit. Horm. 33 (1975) 61-102.
3. Sonnenschein C., Olea N., Pasanen M. E. and Soto A. M., Negative controls of cell proliferation: human prostate cancer cells and androgens. Cancer Res. 49 (1989) 3474-3481.
4. Isaacs J. T., Schulze H. and Coffey D. S., Development of androgen resistance in prostatic cancer. Prog. Clin. Biol. Res. 243A (1987) 21-31.
5. Akakura K., Bruchovsky N., Goldenberg S. L., Rennie P. S. Buckley A. R., Effects of intermittent androgen suppression on androgen-dependent tumors. Apoptosis and serum prostate- specific antigen. Cancer 71 (1993) 2782-2790.
6. Soto A. M., Lin T. M., Sakabe K., Olea N., Damassa D. A. Sonnenschein C., Variants of the human prostate LNCaP cell line as a tool to study discrete components of the androgen- mediated proliferative response. Oncol. Res. 7 (1995) 545-558.
7. T. -M. Lin, Biological effect of androgen on human prostate epithelial cell proliferation: LNCaP cell lines in culture as a model. Doctoral Thesis, Tufts Universiw, Boston, MA, 1993.
8. Wang Z. and Brown D. D., A gene expression screen. Proc. Natl. Acad. Sci. U.S.A. 88 (1991) 11505-11509.

218 Peter Geck et al.
9. Horoszewicz J. S., Leong S. S., Kawinski E., Karr J. P., Rosenthal H., Chu T. M., Mirand E. A. and Murphy G. P., LNCaP model of human prostatic carcinoma. Cancer Res. 43 (1983) 1809-1818.
10. Soto A. M. and Sonnenschein C., The role of estrogens on the proliferation of human breast tumor cells (MCF-7). J. Steroid Bioehem. 23 (1985) 87-94.
11. Szelei J., Jimenez J., Soto A. M., Luizzi M. F. and Sonnenschein C., Androgen-induced inhibition of proliferation in human breast cancer MCF7 cells transfected with androgen receptor. Endocrinology 138 (1997) 1406-1412.
12. Chomczynski P. and Sacchi N., Single step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 162 (1987) 156-159.
13. Chelly J., Monterras D., Pinset C., Berwald-Netter Y., Kaplan J. -C. and Kahn A., Quantitative estimation of minor mRNAs by cDNA polymerase chain reaction, Application to dystrophin mRNA in cultures of myogenic and brain cells. Eur. J. Biochem. 187 (1990) 691-698.
14. Noonan K. E., Beck C., Holzmayer T. A., Chin J. E., Wunder J. S., Andrulis I. L., Gazdar A. F., Willman C. L., Griffith B., Von Hoff D. D. and Roninson I. B., Quantitative analysis of MDR1 (multidrug resistance) gene expression in human tumors by polymerase chain reaction. Proc. Natl. Acad. ScL U.S.A. 87 (1990) 7160-7164.
15. Dallman M. J., Montgomery R. A., Larsen C. P., Wanders A. Wells A. F., Cytokine gene expression: Analysis using Northern blotting, polymerase chain reaction and in situ hy- bridization. Immunol. Rev. 119 (1991) 163-179.
16. Bettger W. J. and McKeehan W. L., Mechanisms of cellular nutrition. Physiol. Rev. 66 (1986) 1-35.
17. Ware J. L., Growth factors and their receptors as determinants in the proliferation and metastasis of human prostate cancer. CancerMet. Rev. 12 (1993) 287-301.
18. Harris A. L., p53 Expression in human breast cancer. Adv. Cancer Res. 59 (1992) 69-88.
19. Olea N., Sakabe K., Soto A. M. and Sonnenschein C., The proliferative effect of 'anti-androgens' on the androgen-sensitive human prostate tumor cell line LNCaP. Endocrinology 126 (1990) 1457-1463.
20. Gordon M. K., Foley J. W., Birk D. E., Fitch J. M. and Linsenmayer T. F., Type V collagen and Bowman's mem- brane. J. Biol. Chem. 269 (1994) 24959-24966.
21. Fornace A. J., Nebert D. W., Hollander M. C., Luethy J. D., Papathanasiou M., Fagnoli J. and Holbrook N. J., Mammalian genes coordinately regulated by growth arrest signals and DNA-damaging agents. Mol. Cell. Biol. 9 (1989) 4196-4203.
22. Del Sal G., Ruaro M. E., Philipson L. and Schneider C., The growth arrest-specific gene, gas1, is involved in growth suppres- sion. Cell 70 (1992) 595-607.
23. Schneider C., King R. M. and Philipson L., Genes specifically expressed at growth arrest of mammalian cells. Cell 54 (1988) 787-793.
24. Fornace A. J., Alamo I. and Hollander M. C., DNA damage- induced transcripts in mammalian cells. Br. J. Cancer 64 (1991) 47-53.
25. Schiaffino M. V., Bassi M. T., Rugarli E. I., Renieri A., Galli L. and Ballabio A., Cloning of a human homologue of the Xenopus laevis APX gene from the ocular albinism type 1 criti- cal region. Hum. MoL Genet. 4 (1995) 373-382.
26. Gudmundsson J., Johannesdottir G., Bergthorsson J. T., Arason A., Ingvarsson S., Egilsson V. and Barkardottir R. B., Different tumor types from BRCA2 carriers show wild type chromosome deletions on 13q12-q13. Cancer Res. 55 (1995) 4830-4832.
Copyright © 2022 FDOKUMEN




















