
Dialogue Journal Keeping to Scaffold Five- Paragraphing Writing

lable at ScienceDirect
Biomaterials 31 (2010) 1180–1190
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
Engineering a scaffold-free 3D tumor model for in vitro drug penetration studies
Siew-Min Ong a,b,c, Ziqing Zhao a, Talha Arooz a, Deqiang Zhao a,d, Shufang Zhang a,e, Tiehua Du f,Martin Wasser f, Danny van Noort a,j, Hanry Yu a,b,g,h, i, j,k,*
a Institute of Bioengineering and Nanotechnology, Agency for Science, Technology and Research (A* STAR), The Nanos,# 04-01, 31 Biopolis Way, Singapore 138669, Singaporeb NUS Graduate School for Integrative Sciences and Engineering, Centre for Life Sciences (CeLS), # 05-01, 28 Medical Drive, Singapore 117456, Singaporec Singapore Immunology Network (SIgN), Agency for Science, Technology and Research (A* STAR), 8A Biomedical Grove, Immunos # 4, Biopolis, Singapore 138648d Department of Chemical Engineering, Faculty of Engineering, National University of Singapore, Singapore 117576e LSCTE, School of Medicine, Zhejiang University, Hangzhou 310058, People’s Republic of Chinaf Bioinformatics Institute, Agency for Science, Technology and Research (A* STAR), Matrix # 07-01, 30 Biopolis Street, Singapore 138671, Singaporeg Department of Physiology, Yong Loo Lin School of Medicine, # 03-03, 2 Medical Drive, Singapore 117597, Singaporeh Singapore-MIT Alliance, E4-04-10, 4 Engineering Drive 3, Singapore 117576, Singaporei NUS Tissue-Engineering Programme, DSO Labs, National University of Singapore, Singapore 117597, Singaporej Centre for Mechanobiology, Temasek Laboratories, National University of Singapore, 5A Engineering Drive 1, Singapore 117411k Department of Mechanical Engineering, Massachusetts Institute of Technology, Cambridge, MA 02139, USA
a r t i c l e i n f o
Article history:Received 29 September 2009Accepted 20 October 2009Available online 3 November 2009
Keywords:SpheroidLiverSynthetic inter-cellular linkerSolid tumour modelDrug penetration
* Corresponding author. Institute of Bioengineeringfor Science, Technology and Research (A* STAR), ThWay, Singapore 138669, Singapore. Tel.: þ65 6516346
E-mail address: [email protected] (H. Yu).
0142-9612/$ – see front matter � 2009 Elsevier Ltd.doi:10.1016/j.biomaterials.2009.10.049
a b s t r a c t
Three-dimensional (3D) in vitro cultures are recognized for recapitulating the physiological microenvi-ronment and exhibiting high concordance with in vivo conditions. In cancer research, the multi-cellulartumor spheroid (MCTS) model is an established 3D cancer model that exhibits microenvironmentalheterogeneity close to that of tumors in vivo. However, the established process of MCTS formation istime-consuming and often uncontrolled. Here, we report a method for engineering MCTS using a tran-sient inter-cellular linker which facilitates cell-cell interaction. Using C3A cells (a hepatocellular carci-noma cell line) as a model, we formed linker-engineered spheroids which grew to a diameter of 250 mmin 7 days, as compared to 16 days using conventional non-adherent culture. Seven-day old linker-engineered spheroids exhibited characteristics of mature MCTS, including spheroid morphology, geneexpression profile, cell-cell interaction, extracellular matrix secretion, proliferation and oxygenconcentration gradients, and cellular functions. Linker-engineered spheroids also displayed a resistanceto drug penetration similar to mature MCTS, with dose-dependent extracellular accumulation of thedrug. The linker-engineered spheroids thus provide a reliable accelerated 3D in vitro tumor model fordrug penetration studies.
� 2009 Elsevier Ltd. All rights reserved.
1. Introduction
In vitro culture systems that mimic the in vivo conditions cangreatly advance biomedical research while minimizing controver-sial animal experimentation. Three-dimensional (3D) in vitrocultures are now recognized for recapitulating the physiologicalmicroenvironment and exhibiting high concordance with in vivoresponses [1,2]. In cancer research, the 3D multi-cellular tumorspheroid (MCTS) model is one of the most studied and establishedcancer models that exhibits microenvironmental heterogeneityclose to that of tumors in vivo [3,4]. MCTSs are compact aggregatesof cells with an extensive 3D network of cell–matrix and cell-cell
and Nanotechnology, Agencye Nanos,# 04-01, 31 Biopolis6; fax: þ65 68748261.
All rights reserved.
interactions [2,4–6], with cells on the periphery actively prolifer-ating while those innermost ones remaining quiescent or necroticwhen the spheroid enlarges beyond 400–600 mm, as a result ofdecreasing nutrient concentration from the exterior of the spheroidto the centre [3,6,7].
MCTSs are conventionally formed using non-adhesive culturemethods like the liquid overlay or rotational culture, relying on thecells’ ability to establish cell-cell contacts, a process which may takeup to weeks [8], or may not even occur at all for certain cell types[9]. To accelerate this process, a large number of cells can be madeto aggregate in a confined space, e.g. forming spheroids using thehanging drop method [10] or by centrifugation of cells in suspen-sion culture [11]. Biomaterials have also been used to confine cellsthree-dimensionally, like seeding cells in scaffolds [12,13] orencapsulating cells in hydrogels [14,15]. Soluble extracellularmatrix (ECM) components can also be added to the culture mediumto aid cell aggregation into 3D spheroids [11]. These methods have

S.-M. Ong et al. / Biomaterials 31 (2010) 1180–1190 1181
their respective advantages but spheroids formed using thesemethods have not been well-characterized to possess the essentialfeatures of MCTS [5]. In addition, employment of bulk biomaterialslike scaffolds and hydrogels would not produce cell-dense andmatrix-poor tumor spheroids reminiscent of in vivo tumors. Hence,engineering MCTS with minimal biomaterials or even as scaffold-free cellular constructs, while allowing the tumor cells to secreteand accumulate natural extracellular matrix (ECM) to maintaintheir 3D structural integrity, is of vital importance in mimickingtheir in vivo counterparts.
Here, we report the engineering of cell-dense MCTS by con-necting cells directly but transiently with an inter-cellular linker[16] to facilitate cell-cell interaction and aggregation. As the inter-cellular linker resides on cell surfaces with a half-life of w2 days[16], the spheroids formed (hereafter referred to as ‘‘linker-engineered spheroids’’) are scaffold-free and cell-dense. The linker-engineered spheroids thus provide a well-characterized andaccelerated 3D tumor model for in vitro drug penetration studies.
2. Materials and methods
2.1. Formation of natural and linker-engineered spheroids
Spheroids were formed naturally from C3A cells cultured to a density of2 million/well in a 6-well dish, on an orbital shaker (Spectra-teknik, USA) rotating at50 rpm at 37 �C in a humidified environment with 5% CO2. For linker-engineeredspheroids, single C3A cells were re-suspended in 0.5 mM sodium periodate andincubated at 4 �C in the dark for 15 min. Cold PEI-hy was then added and incubatedwith the cells for 30 min at 4 �C on an orbitron shaker (Model 260200, BoekelScientific, US). The cellular aggregates formed were washed with PBS, re-suspendedin culture medium and placed on an orbital shaker (Spectra-teknik, USA) at a densityof w1 million/well, rotating at 50 rpm at 37 �C in a humidified environment with 5%CO2. The growth of linker-engineered spheroids and naturally-formed spheroidswas quantified by measuring their diameters on images taken with a phase contrastmicroscope (Olympus, Japan). Error bars represent the s.e.m. of three independentexperiments, with at least 10 images taken per experiment.
2.2. Gene expression analysis
The sequences of the seven human genes analyzed were obtained fromEntrezGene web database. Primers were designed for each gene using Primer-BLAST web application as shown: CDH1: 5-AATCCAAGCAGAATTGCTCA-3 forwardand 5-GGAAATGGGCCTTTTTCATT-3 reverse; CDH2: 5-GACAATGCCCCTCAAGTGTT-3 forward and 5-GGTCCAGCATTTGGATCAAT-3 reverse; ITGA6: 5-CCCTTCCAACC-CAAGGAATC-3 forward and 5-AGGGAAAGCCCTCAGTTCTC-3 reverse; ITGB1:5-ATCCCAGAGGCTCCAAAGAT-3 forward and 5-TGCTGTGGTTGGATCTGAGT-3reverse; CYP1A1: 5-GCCAAGAGTGAAGGGAAGAG-3 forward and 5-GGGCA-GAGGAATGTGATGTT-3 reverse; CYP3A4: 5-TGTTTTCAGCCCATCTCCTT-3 forwardand 5-CATTGCATCGAGACAGTTGG-3 reverse; CASP3a: 5-TTGAGTCCTTTTCCTTT-GACG-3 forward and 5-AACCACCAACCAACCATT TC-3 reverse. RNA was isolatedfrom frozen cell samples with RNeasy Kit (QIAGEN), and reverse transcribed usingHigh Capacity RNA-to-cDNA Kit (Applied Biosystems). PCR was performed ona 7500 Fast Real-Time PCR System (Applied Biosystems) using Power SYBR GreenPCR Master Mix (Applied Biosystems). The relative quantity (RQ) of expression ofeach gene was obtained by normalizing the measured expression level against thatof housekeeping gene GAPDH and a monolayer C3A culture as the referencesample, according to the requirements of the Applied Biosystems 7500/7500 FastReal-Time PCR Software v2.0.
2.3. Sample fixation, cryosection preparation, and H&E staining
Samples of linker-engineered spheroids are taken on day 3, 5, 7, 8, 9 and 10, andsamples of naturally-formed spheroids are taken on day 16. Samples were fixed with3.7% PFA for 2 h at room temperature and stored in PBS at 4 �C. To prepare thecryosections, samples were immersed in 30% sucrose overnight at 4 �C and placingthem into a cryo-mould filled with cryo-embedding compound (OCT), snappedfrozen in a slush of dry ice and sectioned using a cryostat at 5 mm. Routine hema-toxylin and eosin staining was carried out on paraffin sections followed by mountingwith a glass coverslip and synthetic resin.
2.4. Immunostaining
The following steps were all carried out on cryo-sections in a humidifiedchamber. The PBS was replaced with a blocking solution of 10% goat serum (Dako-Cytomation) in PBS for 20 min, followed by 1-h incubation with the desired primary
antibody diluted in PBS. The sections were washed for 10 min in running tap waterthen incubated for 30 min with the secondary antibody – labelled polymer- HRP antimouse Envision Kit (Dako Cat K4007), followed by washing in running tap water for10 min. DAB substrate solution from labelled polymer-HRP anti mouse Envision Kit(Dako Cat K4007) was placed on the sections for 30 s to 2 min. The slides were thenplaced into water to stop the staining reaction before counterstaining with hae-matoxylin for 2 min haematoxylin and washed in running tap water for 5 min. Thesections were then dehydrated by a series of 5-min immersions in 70%, 80%, 90%,3�100% ethanol followed by 3� xylene. Coverslips were mounted on top of thesections using DPX (CellPath, Singapore) and left to dry overnight.
2.5. Hydroxyproline assay
Cellular spheroids or cells grown on 2D monolayer for 24 h were lysed in 6 M HClat 110 �C for 24 h. The HCl was evaporated by placing the samples in open glass vialson a heating stage at 70 �C in the fume hood. The residue was dissolved in 200 ml of 1�assay buffer, a 10� dilution of the stock buffer. (the stock buffer: 50 g citric acidmonohydrate,12 ml acetic acid,120 g sodium acetate, 34 g sodium hydroxide, toppedup to 1 l with deionized water). One hundred ml of choramin-T-reagent was added tothe mixture and incubated at room temperature for 20 min (Chloramin-T-reagent:0.3525 g Chloramine T freshly dissolved in 5.175 ml water, 6.5 ml m-propanol and13.325 ml stock buffer). One hundred ml of dimethyl-aminobenzaldehyde (DMAB)reagent was then added to the mixture and incubated at 60 �C for 15 min (DMABreagent: 3.75 g DMAB freshly dissolved in 15 ml n-propanol and 6.5 ml perchloricacid). All chemicals are used without dilution. The resulting solution was then cooledand the absorbance measured at 540 nm within 30 min. Collagen (PureCol, Inamed,USA) standard was used as positive control for calibration of a standard curve. Thecollagen content measured was normalized to the number of cells analysed byPicoGreen DNA quantification assay (below) and plotted as a percentage of the day0 value. Error bars represent the s.e.m. of three independent experiments.
2.6. Bromodeoxyuridine (BrdU) labelling and hypoxia staining
BrdU labelling was carried out with the BrdU Labelling and Detection Kit II(Roche). Samples were incubated with BrdU reagent (Roche) at 10 mmol L�1 for 1 h,fixed with 70% ethanol for 20 min at �20 �C, and kept in PBS at 4 �C until cryo-sectioning. Cryo-sections were then incubated with anti-BrdU antibodies (Roche),and visualized with indirect immunoperoxidase staining according to the manu-facturer’s protocol. Hypoxic regions in the spheroids were stained by incubating thesample with pimonidazole HCl (Natural Pharmacia International, Inc) at200 mmol L�1 for 2 h at 37 �C. They were then washed and fixed in 3.7% PFA for 2 h atroom temperature, and stored in PBS at 4 �C until cryo-sectioning. Sections are thendetected for pimonidazole with anti-pimonidazole antibody and visualized withindirect immunoperoxidase staining according to the manufacturer’s protocol.
2.7. Functional assessments
The amount of albumin secreted into the culture medium was determined bya sandwich enzyme-linked immunosorbent assay, using a human albumin ELISA kit(Bethyl Laboratories, Montgomery, TX). The standard curve was established usinghuman albumin. Absorbance was measured in a 96-well microtitre plate using Tecanmicroplate Reader, program: XFluor4SafireII. The specific secretion rate werecalculated from the albumin levels obtained in a 24 h culture supernatant normal-ized to the total number of cells obtained from the DNA quantification. CYP activitywas determined by incubating spheroids in 40 mM of 3-cyano-7-ethoxycoumarin(CEC) (Molecular Probes, OR, USA) in culture medium for 3 h. The medium wascollected and the metabolic product, 3-cyano-7-hydroxycoumarin (CHC), was ana-lysed using capillary electrophoresis with laser induced fluorescence (CE-LIF)detection (Prince Technologies B.V., Netherlands) at an excitation wavelength of325 nm [17]. UGT activity was determined by incubating spheroids in 100 mM of4-methylumbelliferone (4-MU) (Merck, Singapore) in culture medium for 4 h. Themedium was collected and the metabolic product, 4-methylumbelliferyl glucuro-nide (4-MUG), was analysed using capillary electrophoresis with laser inducedfluorescence (CE-LIF) detection (Prince Technologies B.V., Netherlands) at an exci-tation wavelength of 325 nm.[17] Total cell number was quantified using Quant-iT�PicoGreen dsDNA Assay Kit (Invitrogen). Spheroids were lysed in 0.1% SDS solution.50 ml of this mixture was diluted to 500 ml with PBS and reacted with the DNA dye(diluted 200� in PBS) for 5 min in the dark at room temperature. The mixture wasthen excited at 480 nm and the fluorescence read at 520 nm. A calibration curvewith known cell numbers was used for quantification.
2.8. Doxorubicin penetration studies
Linker-engineered spheroids (7-day old), naturally-formed spheroids andconfluent 2D cultures of C3A were tested with doxorubicin (Sigma). Concentrationsof 1, 5 and 10 PPC were chosen for the experiment, i.e., 0.4, 2 and 4 mg mL�1. Pene-tration of the drug was determined by incubating the samples with 25 mM Cell-Tracker� Blue (Molecular Probes, USA) and 250 nM of Sytox Green� (MolecularProbes, USA) in culture medium for 30 min at 37 �C and imaging immediately with

Fig. 1. Growth of linker-engineered spheroids. Growth kinetics of naturally-formed spheroids (a) is compared to that of linker-engineered spheroids (b). The dotted lines mark thetarget diameter of 250 mm. H&E staining of histological cross-sections of naturally-formed spheroids at day 16 (c) and linker-engineered spheroids at day 7 (d) shows similarmorphology of the two spheroid species. Scale bar: 50 mm.
S.-M. Ong et al. / Biomaterials 31 (2010) 1180–11901182
confocal microscope (Carl Zeiss). Doxorubicin was excited with laser wavelength of543 nm and its emission collected with a 590 nm long pass filter. Drug penetrationwas quantified with analysis of the confocal images using the software ImageProPlus, expressed as a percentage of cells with doxorubicin-stained nuclei.
2.9. Image quantification
All the images were analysed using our customized program (available uponrequest) written in Matlab (MathWorks, Massachusetts, USA). First, the images wereconverted to greyscale. An adaptive thresholding method [1] was applied to separatethe foreground from background. Morphological operations were applied to removebackground staining. Finally, the biggest region at the foreground was chosen foranalysis (as there might be more than one region (spheroid) in the image). For allimage quantification, the boundary of the region of interest (ROI) was first detec-ted..To quantify BrdU and hypoxia staining (transmission light images, Fig. 4), theinverse intensity of the images was used, and the distance of each positive stainingfrom the spheroid boundary was recorded. Multiple images were quantified andaveraged. To determine drug penetration (Fig. 7), the minimum distance of eachpixel to the boundary was determined and the pixel intensity recorded. Bothdistance and intensity values were normalized to a 0–1 scale for comparisonbetween images.
3. Results
Mature MCTS are compact structures of cells approximately200–250 mm in diameter that possess the following structuralfeatures: cell-cell interaction, development of extracellular matrix[18–20], as well as gradients of nutrient concentration and cellproliferation from the exterior to the spheroid center [3,7]. On theother hand, a cellular aggregate is a loose pack of cells withoutthese distinct features. It is therefore crucial to assess the linker-engineered spheroids for the development of mature MCTSfeatures in order for them to be used as a model for cancer research.
We have characterized these mature features during the aggregate-to-spheroid transformation for C3A cells, and compared them tomature spheroids of 250 mm in diameter formed naturally from C3Acells under rotational culture (hereafter referred to as ‘‘naturally-formed spheroids’’). The target size of 250 mm was chosen formature naturally-formed spheroids as MCTS of 200–250 mm havebeen reported to possess the essential features described above androutinely used for drug penetration tests [7].
3.1. Growth of naturally-formed and linker-engineered spheroids
We followed the growth kinetics of the naturally-formedspheroids and linker-engineered spheroids by measuring theirdiameters (Fig. 1a and b). Naturally-formed spheroids took 15–17days to reach the target size of w250 mm in diameter, while linker-engineered spheroids took only 7 days. The 16 day old naturally-formed spheroids are thus used as the mature spheroid control forall subsequent comparisons with linker-engineered spheroids.Hematoxylin and Eosin (H&E) staining of cross-sections of thespheroids showed that the linker-engineered spheroids at day 7had a compact structure closely resembling the naturally-formedmature spheroids at day 16, with distinct nuclei and cytoplasmstaining (Fig. 1c and d).
3.2. Gene expression profiles of naturally-formed and linker-engineered spheroids
To further assess the time required for linker-engineeredspheroids to reach the maturity level of naturally-formed

Fig. 2. Relative quantity (RQ) of gene expression in naturally-formed and linker-engineered spheroids. Expression levels of 7 genes in naturally-formed spheroids (N) at days 14, 16and 19 are compared to those in linker-engineered spheroids (E) at days 5, 7 and 8. Each spot represents a gene. Average root-mean-square deviation (drms) of gene expression levelfrom the line y¼ x for each N/E pair is shown at the top of each graph.
S.-M. Ong et al. / Biomaterials 31 (2010) 1180–1190 1183
spheroids at 16 days, we compared the gene expression profiles ofthe linker-engineered spheroids at days 5, 7 and 8 (E5, E7, and E8respectively) with naturally-formed spheroids at days 14, 16 and 19(N14, N16, and N19 respectively). Genes relevant to the formationand maintenence, cellular functions and viability of an MCTS werecompared. These include cell-cell interaction molecules like cad-herins (CDH1, CDH2), cell–matrix intereaction molecules likeintegrins (Itga6, Itgb1), metabolic enzymes (Cyp1a1, Cyp3a4) andan apoptosis mediator (Casp3a). The expression of these genes wasanalyzed in both the naturally-formed (N) and linker-engineered(E) spheroids using qRT-PCR. The relative quantity (RQ) of eachgene is plotted for each N/E pair (Fig. 2). The closer each point is tothe line y¼ x (dash), the closer is the gene expression levelbetween the N and E spheroids. More quantitatively, the root-mean-square deviation (drms) of each point in Fig. 2 from the liney¼ x is calculated, and the average of the drms for all 7 genes isindicated at the top of each graph (Fig. 2). Among the nine N/E
pairs, the N16/E7 pair has a drms value (0.42) significantly smallerthan those of all other pairs (�1.00), indicating the highest simi-larity in gene expression profiles between these two species. Thisconfirms, on the gene expression level, that linker-engineeredspheroids at day 7 closely mimic mature naturally-formed spher-oids at day 16.
3.3. Structural characterization of linker-engineered spheroids
The formation of cell-cell interaction is an essential feature ofa 3D spheroid. E-cadherin is an integral membrane protein thatforms part of the adherens junction in epithelial cells [21]. To checkfor the development of natural cell-cell interaction in linker-engineered spheroids, expression of E-cadherin proteins in E7spheroids was assessed using immuno-staining and compared withN16 spheroids (Fig. 3) Abundant E-cadherin expression wasobserved in both the E7 spheroids (Fig. 3a) and N16 spheroids

Fig. 3. Cell-cell interaction and ECM secretion in mature spheroids. Histological cross-sections of linker-engineered spheroids at day 16 (a, c, e) and naturally-formed spheroids atday 7 (b, d, f) are immunostained for (a–b) E-cadherin, (c–d) fibronectin and (e–f) laminin. Scale bar: 50 mm. (g) Relative amounts of collagen secreted by linker-engineeredspheroids are expressed as percentages of collagen secreted by naturally-formed spheroids at day 16 (dotted line).

Fig. 4. Development of gradients in mature spheroids. BrdU labelling (a–b) and hypoxia staining (d–e) of histological cross-sections of mature naturally-formed spheroids at day 16and linker-engineered spheroids at day 7, respectively. Scale bar: 50 mm. The intensity of the respective labelling for both spheroid species is plotted across the spheroid radius in (c)and (f).
S.-M. Ong et al. / Biomaterials 31 (2010) 1180–1190 1185
(Fig. 3b). To better assess the level of expression of E-cadherin,quantification of the staining pattern was carried out using imageanalysis. The level of E-cadherin expression (number of brownpixels/number of nuclei, in arbitrary units) in E7 was normalized tothat of N16 (indicated at the bottom right corner of each image). E-cadherin level in E7 (103� 25) was similar to that of N16 (100�18),indicating sufficient formation of cell-cell interactions of linker-engineered spheroids by day 7.
Secretion and accumulation of extracellular matrix is anotheressential feature of a 3D spheroid. Major components of the hepaticextracellular matrix include fibronectin, laminin, and collagen typeIII and IV [22]. Both types of spheroids were assessed for fibronectinand laminin using immuno-staining, and for collagen using thehydroxyproline test. The expression of fibronectin (Fig. 3c) andlaminin (Fig. 3e) in E7 was uniform and highly similar to those inN16 (Fig. 3d and f). Quantification of their expression levels revealedthat both fibronectin (96� 3) and laminin (98� 19) expression inE7 were very close to those in N16 (100� 7 and 100�13 respec-tively). Total collagen content of E7, when normalized to cellnumber, also reached a similar level as that of N16 (Fig. 3g).Together, these results confirm, on the protein level, that the 7 dayold linker-engineered spheroids resemble the natural spheroidsstructurally in their cell-cell interaction and ECM secretion.
Two other key features of an MCTS are the development ofa proliferative gradient and an oxygen concentration gradientacross the spheroid radius. Proliferating cells in both linker-engi-neered spheroids and naturally-formed spheroids were labelledwith bromodeoxyuridine (BrdU), a synthetic nucleoside whichbinds to DNA of cells in S-phase. The preferential distribution ofproliferative cells on the periphery of E7 spheroids, as evidenced bytheir dark nucleus staining, compares favourably with the distri-bution of proliferative cells in N16 spheroids (Fig. 4a and b). More
quantitatively, the gradient of BrdU staining intensity across thespheroid radius from the boundary to the centre also shows goodagreement between the N16/E7 pair (Fig. 4c). To investigate thepresence of an oxygen concentration gradient, hypoxic regions(with oxygen concentration <10 mmHg) in the spheroids weredetected with pimonidazole and visualized with immunoperox-idase staining (Fig. 4d and e). Hypoxic regions were detected at theinterior of the E7 spheroids (more darkly stained regions) while theperipheral regions exhibited higher oxygen concentration, similarto the staining pattern in mature N16 spheroids. Quantitatively,N16 and E7 spheroid species exhibit high correspondence in thegradient of oxygen concentration across the spheroid radius fromthe centre to the periphery (Fig. 4f). These results verify that linker-engineered spheroids possess two concentration gradients essen-tial to MCTS by day 7.
3.4. Functional assessment of linker-engineered spheroids
To further evaluate the maturity of linker-engineered spheroids,three hepatocellular functions were monitored at different timepoints over 10 days and compared to the functional levels of N16spheroids. Albumin production as well as phase I and II metabolicactivities, as indicated by cytochrome P450 (CYP) 1A1/2 isoen-zymes and UDP-glucuronyltransferase (UGT) activities respectively,were assessed and normalized to total cell number (Fig. 5). Albuminproduction in linker-engineered spheroids rose to a level similar tothat of N16 spheroids around day 7–8 (Fig. 5a), whereas CYP1A1/2and UGT activities of linker-engineered spheroids resembled thoseof N16 from as early as on day 3 (Fig. 5b and c) and lasted till day 9.Taking all 3 functional assessments into account, the 7-day oldlinker-engineered spheroids proved to best resemble mature MCTSformed naturally.

Fig. 5. Functional assessments of linker-engineered spheroids. Normalized levels of (a)albumin production, (b) CYP1A1/2 activity, and (c) UGT activity of linker-engineeredspheroids over 10 days, compared to those of mature naturally-formed spheroids(dotted line).
S.-M. Ong et al. / Biomaterials 31 (2010) 1180–11901186
3.5. Resistance to drug penetration of mature linker-engineeredspheroids
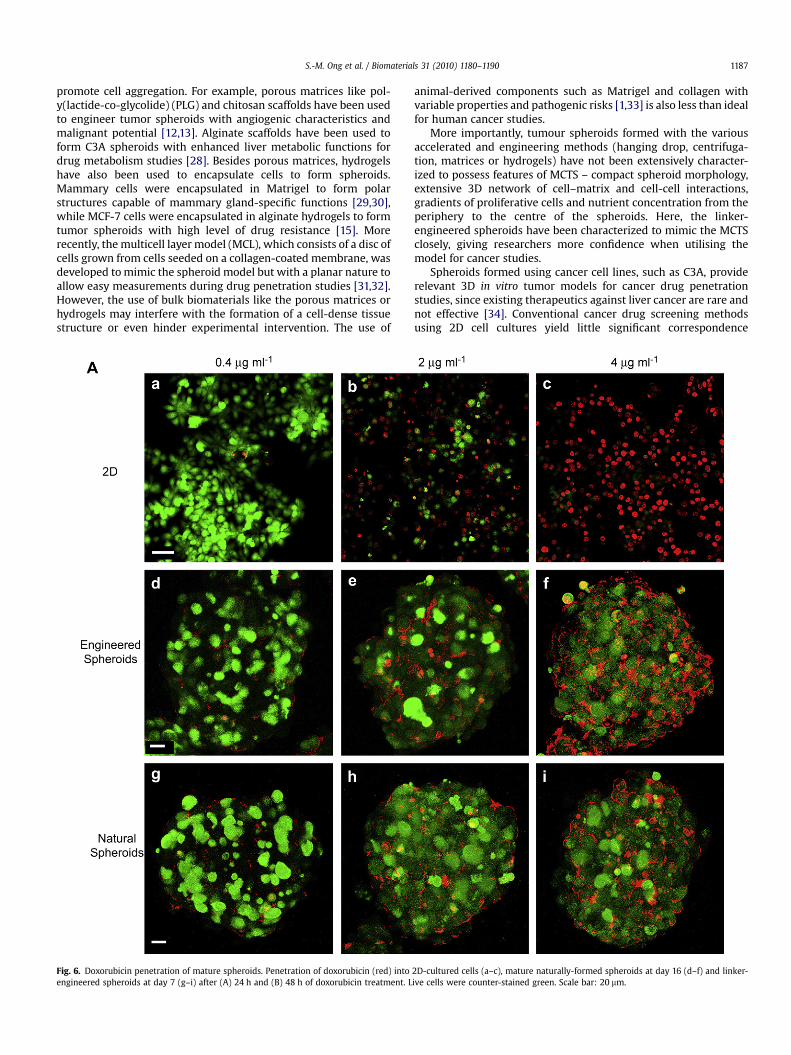
To investigate the application of inter-cellular linker to formmulti-cellular tumor spheroids (MCTS) for cancer studies, E7spheroids were tested for their resistance to drug penetration. Ananti-cancer drug, doxorubicin, with a reported peak plasmaconcentration (PPC) of 0.4 mg/ml, was used [23]. E7 spheroids weretreated with doxorubicin at concentrations of 1, 5 and 10 times thePPC; for comparison, cells cultured in 2D and N16 spheroids weretreated similarly. The penetration of doxorubicin, which fluorescesred [24–26], was assessed with confocal microscopy. For visuali-zation, live cells were counter-stained green. After 24 h of treat-ment, doxorubicin penetrated into the nuclei of 2D-cultured cells in
a dose-dependent manner (Fig. 6A a–c). On the other hand, doxo-rubicin hardly penetrated the cell nuclei of E7 and N16 spheroids at0.4 mg/ml (Fig. 6A d and g), and showed minimal penetration intocell nuclei at higher concentrations of 2 and 4 mg/ml (Fig. 6A e, f, hand i). Instead, doxorubicin displayed a dose-dependent accumu-lation at the extra-nuclei (either extracellular or cytoplasmic)region of the E7 and N16 spheroids, a phenomenon not observed in2D-cultured cells. It was only after 48 h of treatment that doxoru-bicin showed slight penetration into cell nuclei of E7 and N16spheroids at 2 mg/ml (Fig. 6B e and h), and extensive penetration at4 mg/ml (Fig. 6B f and i). We further quantified the fluorescenceimages to evaluate the kinetics of the drug penetration into thespheroids (Fig. 7). The distributions of doxorubicin across thespheroid radius in both N16 and E7 species after 24 h and 48 h oftreatment agree well at low drug concentration (0.4 and 2 mg/ml).As the drug concentration increases (4 mg/ml) the natural spheroidsshow a higher doxorubicin penetration than the engineered ones(Fig. 7a–c), which suggests that the naturally-formed spheroids areless compact functionally than the structurally similar linker-engineered spheroids and thus more readily allows the penetrationof drugs at very high concentrations. A similar trend is observed inthe total amount of doxorubicin penetrated into the spheroids(Fig. 7d). Nevertheless, the observation of doxorubicin penetrationpattern and amount at lower concentrations of 0.4 and 2 mg/mlagrees with reports of tumor cells rapidly acquiring and sustaininga drug-resistant phenotype when grown as 3D multi-cellular tumorspheroids [27], and further confirms the resemblance of our linker-engineered spheroids with naturally-formed MCTSs.
4. Discussion
We have shown that linker-engineered spheroids are structur-ally and functionally mature by day 7, and they displayed similarresistance to doxorubicin penetration as mature MCTS formednaturally in 16 days. Hence, the engineering process did not alterthe ability of C3A cells to remodel upon culture to display pheno-types of mature MCTS, and the inter-cellular linker can be used toaccelerate the MCTS formation. The methods of linker introductionand spheroid culture are also not cell type-specific [16], and thuscould potentially be applied to all cell types.
The formation of a mature spheroid can be divided into twosteps. The first step is the ‘‘pooling’’ of many individual cells to formcell aggregates, which may be the result of natural aggregation ina rotational culture and/or the proliferation of single cells to formcellular aggregates. The second step is the remodelling of the 3Dcellular microenvironment, including the formation of cell-cellinteractions, secretion of ECM and development of proliferative andnutrient gradients. The inter-cellular linker accelerated theformation of spheroids by skipping the first step, i.e. by forcing theaggregation of a large number of individual cells. To date, variousengineering methods have also been developed to accelerate theaggregation of cells to form spheroids. For instance, cells can beencouraged to aggregate by confining them in ‘‘hanging drops’’, orcentrifuged to collect at the foot of round-bottom wells. However,these methods do not work for all cell types. Cell lines like MCF-10a,Caco-2 [10], MDA-MB and SK-BR-3 [11], though forced togetherphysically, dissociate easily and do not form compact spheroids. Inthis sense, the inter-cellular linker provides a good alternativemethod to aggregate cells. The inter-cellular linker consists ofpolyethyleneimine (PEI) molecules with multiple hydrazide armsthat react with aldehyde groups on modified cell surfaces to formcell aggregates [16]. This method is not cell type-specific and wouldbe especially useful for cells that do not aggregate spontaneously.
Other engineering methods to accelerate the formation of cellspheroids make use of biomaterials to seed cells close together to
S.-M. Ong et al. / Biomaterials 31 (2010) 1180–1190 1187
promote cell aggregation. For example, porous matrices like pol-y(lactide-co-glycolide) (PLG) and chitosan scaffolds have been usedto engineer tumor spheroids with angiogenic characteristics andmalignant potential [12,13]. Alginate scaffolds have been used toform C3A spheroids with enhanced liver metabolic functions fordrug metabolism studies [28]. Besides porous matrices, hydrogelshave also been used to encapsulate cells to form spheroids.Mammary cells were encapsulated in Matrigel to form polarstructures capable of mammary gland-specific functions [29,30],while MCF-7 cells were encapsulated in alginate hydrogels to formtumor spheroids with high level of drug resistance [15]. Morerecently, the multicell layer model (MCL), which consists of a disc ofcells grown from cells seeded on a collagen-coated membrane, wasdeveloped to mimic the spheroid model but with a planar nature toallow easy measurements during drug penetration studies [31,32].However, the use of bulk biomaterials like the porous matrices orhydrogels may interfere with the formation of a cell-dense tissuestructure or even hinder experimental intervention. The use of
Fig. 6. Doxorubicin penetration of mature spheroids. Penetration of doxorubicin (red) intoengineered spheroids at day 7 (g–i) after (A) 24 h and (B) 48 h of doxorubicin treatment. L
animal-derived components such as Matrigel and collagen withvariable properties and pathogenic risks [1,33] is also less than idealfor human cancer studies.
More importantly, tumour spheroids formed with the variousaccelerated and engineering methods (hanging drop, centrifuga-tion, matrices or hydrogels) have not been extensively character-ized to possess features of MCTS – compact spheroid morphology,extensive 3D network of cell–matrix and cell-cell interactions,gradients of proliferative cells and nutrient concentration from theperiphery to the centre of the spheroids. Here, the linker-engineered spheroids have been characterized to mimic the MCTSclosely, giving researchers more confidence when utilising themodel for cancer studies.
Spheroids formed using cancer cell lines, such as C3A, providerelevant 3D in vitro tumor models for cancer drug penetrationstudies, since existing therapeutics against liver cancer are rare andnot effective [34]. Conventional cancer drug screening methodsusing 2D cell cultures yield little significant correspondence
2D-cultured cells (a–c), mature naturally-formed spheroids at day 16 (d–f) and linker-ive cells were counter-stained green. Scale bar: 20 mm.

Fig. 6. (continued).
S.-M. Ong et al. / Biomaterials 31 (2010) 1180–11901188
between preclinical models and actual clinical results [8,35]. Thislack of predictability has been attributed to the lack of attention inculturing of the cells to phenotypically represent their in vivomicroenvironment [7,36]. To better recapitulate the in vivo situa-tion in vitro, 3D models like MCTS model are necessary, with keyfeatures of in vivo tumors such as cell-cell and cell–matrix inter-actions that are relevant to the penetration of drugs [3,4,33]. Thedifferent cell populations present in MCTS are also similar toavascular tumor nodules or micro-regions of solid tumors [3,9].Proliferating cells located in the periphery of spheroids with directaccess to culture medium are comparable to tumor cells situatedclose to capillaries which remain active in cell growth and prolif-eration. The quiescent or necrotic cells in the center of the spher-oids with poor access to culture medium are representative oftumor cells at a distance from capillaries which stop growing andeventually die [3,8]. These features in the MCTS model better reflectthe in vivo patho-physiological situation in human cancer tissuesthan conventional 2D cultures, and thus empower the model withmore predictive capability of the tumor behaviour in vivo.
The spheroids generated using the inter-cellular linker couldbe used as an MCTS model for cancer studies. In addition tohaving the ability to remodel during culture to display maturefeatures of MCTS, the linker-engineered spheroids also displayedsimilar drug penetration resistance as naturally-formed 3Dspheroids. In contrast, cells cultured in 2D monolayers experienceextensive doxorubicin penetration into the cell nuclei, suggestingthat the extracellular matrix present in 3D cultures may posea barrier to drug penetration, a phenomenon likely to occur invivo but which cannot be recapitulated using conventional 2Dcultures in vitro. Since good correlation in resistance to drugpenetration has been observed between MCTS and tumors in vivo[37,38], the inter-cellular linker can thus be used to form MCTS asa reliable 3D in vitro tumor model for cancer studies. Otherpotential applications of the inter-cellular linker include formingcellular aggregates for organ printing [39,40] and constructingcomplex tissue structures with the help of stenciling methodssuch as dielectrophoretic cell patterning [41] or microfabricationtechniques [42].

Fig. 7. Quantitative analysis of doxorubicin penetration into spheroids. The distribution of doxorubicin across the spheroid radius from the center to the periphery is plotted for bothspheroid species treated with (a) 0.4 mg/ml, (b) 2 mg/ml, and (c) 4 mg/ml for a period of 24 and 48 h. The total amount of doxorubicin penetrated computed by integrating the areaunder the respective distributions is shown in (d).
S.-M. Ong et al. / Biomaterials 31 (2010) 1180–1190 1189
5. Conclusions
We have employed a transient inter-cellular polymeric linker toengineer MCTS in 7 days, versus 16 days with conventional non-adherent culture method. Seven-day old linker-engineered MCTSexhibited similar structural characteristics as the mature naturally-formed MCTS, such as cell-cell interaction, ECM secretion,peripheral distribution of proliferating cells and presence ofcentral hypoxic region. Linker-engineered MCTS and maturenaturally-formed MCTS also displayed similar resistance to doxo-rubicin penetration under all conditions tested with dose-depen-dent extracellular accumulation of the drug, in agreement withreports that tumor cells in MCTS rapidly acquire and sustaina drug-resistant phenotype in response to drug treatment. Themethods of linker introduction and spheroid culture are not celltype-specific, and thus could potentially be applied to all cell types.The scaffold-free linker-engineered spheroids thus provide a reli-able, accelerated 3D in vitro tumor model for drug penetrationstudies.
Acknowledgements
We thank Dr. Zhilian Yue, Ms Xuejun Mo, and all other membersof the Cell and Tissue Engineering Laboratory for stimulatingscientific discussions. This work is supported in part by theInstitute of Bioengineering and Nanotechnology, BMRC, A*STAR ofSingapore and grants from BMRC (R185-001-045-305), Ministry ofEducation (R-185-000-135-112), Jassen Cilag (R-185-000-182-592),Singapore-MIT Alliance Computational and Systems Biology Flag-ship Project funding (C-382-603-004-001), SMART BioSyM andCentre for Mechanobiology to HYU. S.-M.O. is an A*STAR graduatescholar. Z.Z. is an A*STAR National Science Scholar. D.Z. and S.Z areNational University of Singapore graduate research scholars.
Appendix
Figures with essential colour discrimination. Many of the figuresin this article have parts that are difficult to interpret in black andwhite. The full colour images can be found in the on-line version, atdoi:10.1016/j.biomaterials.2009.10.049.
References
[1] Griffith LG, Swartz MA. Capturing complex 3D tissue physiology in vitro. NatRev Mol Cell Biol 2006;7(3):211–24.
[2] Pampaloni F, Reynaud EG, Stelzer EH. The third dimension bridges the gapbetween cell culture and live tissue. Nat Rev Mol Cell Biol 2007;8(10):839–45.
[3] Kunz-Schughart LA. Multicellular tumor spheroids: intermediates betweenmonolayer culture and in vivo tumor. Cell Biol Int;23(3):157–161.
[4] Sutherland RM. Cell and environment interactions in tumor microregions: themulticell spheroid model. Science 1988;240(4849):177–84.
[5] Friedrich J, Ebner R, Kunz-Schughart LA. Experimental anti-tumor therapy in3-D: spheroids–old hat or new challenge? Int J Radiat Biol 2007;83(11–12):849–71.
[6] Mueller-Klieser W. Three-dimensional cell cultures: from molecular mecha-nisms to clinical applications. Am J Physiol 1997;273(4 Pt 1):C1109–1123.
[7] Minchinton AI, Tannock IF. Drug penetration in solid tumours. Nat Rev Cancer2006;6(8):583–92.
[8] Kunz-Schughart LA, Freyer JP, Hofstaedter F, Ebner R. The use of 3-D culturesfor high-throughput screening: the multicellular spheroid model. J BiomolScreen 2004;9(4):273–85.
[9] Mueller-Klieser W. Tumor biology and experimental therapeutics. Crit RevOncol Hematol 2000;36(2–3):123–39.
[10] Kelm JM, Timmins NE, Brown CJ, Fussenegger M, Nielsen LK. Method forgeneration of homogeneous multicellular tumor spheroids applicable toa wide variety of cell types. Biotechnol Bioeng 2003;83(2):173–80.
[11] Ivascu A, Kubbies M. Rapid generation of single-tumor spheroids for high-throughput cell function and toxicity analysis. J Biomol Screen2006;11(8):922–32.
[12] Fischbach C, Chen R, Matsumoto T, Schmelzle T, Brugge JS, Polverini PJ, et al.Engineering tumors with 3D scaffolds. Nat Methods 2007;4(10):855–60.
[13] Dhiman HK, Ray AR, Panda AK. Three-dimensional chitosan scaffold-basedMCF-7 cell culture for the determination of the cytotoxicity of tamoxifen.Biomaterials 2005;26(9):979–86.

S.-M. Ong et al. / Biomaterials 31 (2010) 1180–11901190
[14] Weaver VM, Lelievre S, Lakins JN, Chrenek MA, Jones JC, Giancotti F, et al. beta4integrin-dependent formation of polarized three-dimensional architectureconfers resistance to apoptosis in normal and malignant mammary epithe-lium. Cancer Cell 2002;2(3):205–16.
[15] Zhang X, Wang W, Yu W, Xie Y, Zhang X, Zhang Y, et al. Development of an invitro multicellular tumor spheroid model using microencapsulation and itsapplication in anticancer drug screening and testing. Biotechnol Prog2005;21(4):1289–96.
[16] Ong SM, He L, Thuy Linh NT, Tee YH, Arooz T, Tang G, et al. Transient inter-cellular polymeric linker. Biomaterials 2007;28(25):3656–67.
[17] Toh YC, Zhang J, Khong YM, Du YN, Sun WX, Yu H. Integrating sensitivequantification of hepatic metabolic functions by capillary electrophoresis withlaser-induced fluorescence detection. Analyst 2008;132(3):326–30.
[18] Bates RC, Edwards NS, Yates JD. Spheroids and cell survival. Crit Rev OncolHematol 2000;36(2-3):61–74.
[19] Santini MT, Rainaldi G. Indovina PL.Apoptosis, cell adhesion and the extra-cellular matrix in the three-dimensional growth of multicellular tumorspheroids. Crit Rev Oncol Hematol 2000;36(2–3):75–87.
[20] Santini MT, Rainaldi G. Three-dimensional spheroid model in tumor biology.Pathobiology 1999;67(3):148–57.
[21] Bruce Alberts AJ, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter. MolBiol Cell 2002.
[22] Burgess JW, Gould DR, Marcel YL. The HepG2 extracellular matrix containsseparate heparinase- and lipid-releasable pools of ApoE. Implications forhepatic lipoprotein metabolism. J Biol Chem 1998;273(10):5645–54.
[23] Kusumoto T, Sakaguchi Y, Maehara Y, Nakashima T, Furusawa M, Sugimachi K.Comparison of in vitro anticancer chemosensitivity between human squa-mous cell carcinoma and adenocarcinoma. Oncology 1992;49(5):343–6.
[24] Fornari FA, Randolph JK, Yalowich JC, Ritke MK, Gewirtz DA. Interference bydoxorubicin with DNA unwinding in MCF-7 breast tumor cells. Mol Pharmacol1994;45(4):649–56.
[25] Momparler RL, Karon M, Siegel SE, Avila F. Effect of adriamycin on DNA, RNA,and protein synthesis in cell-free systems and intact cells. Cancer Res1976;36(8):2891–5.
[26] Lee TK, Lau TC, Ng IO. Doxorubicin-induced apoptosis and chemosensitivity inhepatoma cell lines. Cancer Chemother Pharmacol 2002;49(1):78–86.
[27] Durand RE, Olive PL. Resistance of tumor cells to chemo- and radiotherapymodulated by the three-dimensional architecture of solid tumors and spher-oids. Methods Cell Biol 2001;64:211–33.
[28] Elkayam T, Amitay-Shaprut S, Dvir-Ginzberg M, Harel T, Cohen S.Enhancing the drug metabolism activities of C3A–a human hepatocyte cellline–by tissue engineering within alginate scaffolds. Tissue Eng 2006;12(5):1357–68.
[29] Schmeichel KL, Bissell MJ. Modeling tissue-specific signaling and organfunction in three dimensions. J Cell Sci 2003;116(Pt 12):2377–88.
[30] Nelson CM, Bissell MJ. Of extracellular matrix, scaffolds, and signaling: tissuearchitecture regulates development, homeostasis, and cancer. Annu Rev CellDev Biol 2006;22:287–309.
[31] Kyle AH, Huxham LA, Chiam AS, Sim DH, Minchinton AI. Direct assessment ofdrug penetration into tissue using a novel application of three-dimensionalcell culture. Cancer Res 2004;64(17):6304–9.
[32] Tannock IF, Lee CM, Tunggal JK, Cowan DS, Egorin MJ. Limited penetration ofanticancer drugs through tumor tissue: a potential cause of resistance of solidtumors to chemotherapy. Clin Cancer Res 2002;8(3):878–84.
[33] Abbott A. Cell culture: biology’s new dimension. Nature 2003;424(6951):870–2.
[34] Lo A, Lin CT, Wu HC. Hepatocellular carcinoma cell-specific peptide ligand fortargeted drug delivery. Mol Cancer Ther 2008;7(3):579–89.
[35] Johnson JI, Decker S, Zaharevitz D, Rubinstein LV, Venditti JM, Schepartz S,et al. Relationships between drug activity in NCI preclinical in vitro and in vivomodels and early clinical trials. Br J Cancer 2001;84(10):1424–31.
[36] Bhadriraju K, Chen CS. Engineering cellular microenvironments to improvecell-based drug testing. Drug Discov Today 2002;7(11):612–20.
[37] Desoize B, Jardillier J. Multicellular resistance: a paradigm for clinical resis-tance? Crit Rev Oncol Hematol 2000;36(2–3):193–207.
[38] Debnath J, Brugge JS. Modelling glandular epithelial cancers in three-dimen-sional cultures. Nat Rev Cancer 2005;5(9):675–88.
[39] Boland T, Mironov V, Gutowska A, Roth EA, Markwald RR. Cell and organprinting 2: fusion of cell aggregates in three-dimensional gels. Anat Rec ADiscov Mol Cell Evol Biol 2003;272(2):497–502.
[40] Jakab K, Neagu A, Mironov V, Markwald RR, Forgacs G. Engineering biologicalstructures of prescribed shape using self-assembling multicellular systems.Proc Natl Acad Sci U S A 2004;101(9):2864–9.
[41] Albrecht DR, Underhill GH, Wassermann TB, Sah RL, Bhatia SN. Probing therole of multicellular organization in three-dimensional microenvironments.Nat Methods 2006;3(5):369–75.
[42] Folch A, Toner M. Cellular micropatterns on biocompatible materials. Bio-technol Prog 1998;14(3):388–92.
Copyright © 2022 FDOKUMEN




















