
T Helper 2 Cytokines Inhibit Autophagic Control of Intracellular Mycobacterium tuberculosis
Upload
independentCategory
view
2download
0
ISSN: 1524-4539 Copyright © 2009 American Heart Association. All rights reserved. Print ISSN: 0009-7322. Online
72514Circulation is published by the American Heart Association. 7272 Greenville Avenue, Dallas, TX
DOI: 10.1161/CIRCULATIONAHA.109.894642 published online Nov 16, 2009; Circulation
Tabrizian, Eric Thorin and Yahye Merhi Paul-Eduard Neagoe, Benoit Labarthe, Pierre Théroux, Martin G. Sirois, Maryam
Haissam Abou-Saleh, Daniel Yacoub, Jean-François Théorêt, Marc-Antoine Gillis, Formation
Endothelial Progenitor Cells Bind and Inhibit Platelet Function and Thrombus
http://circ.ahajournals.org/cgi/content/full/CIRCULATIONAHA.109.894642/DC1Data Supplement (unedited) at:
http://circ.ahajournals.org
located on the World Wide Web at: The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://circ.ahajournals.org/subscriptions/Subscriptions: Information about subscribing to Circulation is online at
by on May 18, 2011 circ.ahajournals.orgDownloaded from

Endothelial Progenitor Cells Bind and Inhibit PlateletFunction and Thrombus Formation
Haissam Abou-Saleh, PhD; Daniel Yacoub, MSc; Jean-François Théorêt, PhD;Marc-Antoine Gillis, MSc; Paul-Eduard Neagoe, MSc; Benoit Labarthe, PhD; Pierre Théroux, MD;
Martin G. Sirois, PhD; Maryam Tabrizian, PhD; Eric Thorin, PhD; Yahye Merhi, PhD
Background—Interactions of endothelial progenitor cells (EPCs) with vascular and blood cells contribute to vascularhomeostasis. Although platelets promote the homing of EPCs to sites of vascular injury and their differentiation intoendothelial cells, the functional consequences of such interactions on platelets remain unknown. Herein, we addressedthe interactions between EPCs and platelets and their impact on platelet function and thrombus formation.
Methods and Results—Cultured on fibronectin in conditioned media, human peripheral blood mononuclear cellsdifferentiated, within 10 days of culture, into EPCs, which uptake acetylated low-density lipoprotein, bind ulex-lectin,lack monocyte/leukocyte markers (CD14, P-selectin glycoprotein ligand-1, L-selectin), express progenitor/endothelialmarkers (CD34, vascular endothelial growth factor receptor-2, von Willebrand factor, and vascular endothelialcadherin), and proliferate in culture. These EPCs bound activated platelets via CD62P and inhibited its translocation,glycoprotein IIb/IIIa activation, aggregation, and adhesion to collagen, mainly via prostacyclin secretion. Indeed, thiswas associated with upregulation of cyclooxygenase-2 and inducible nitric oxide synthase. However, the effects onplatelets in vitro were reversed by cyclooxygenase and cyclooxygenase-2 inhibition but not by nitric oxide or induciblenitric oxide synthase inhibition. Moreover, in a ferric chloride–induced murine arterial thrombosis model, injection ofEPCs led to their incorporation into sites of injury and impaired thrombus formation, leading to an incomplete occlusionwith 50% residual flow.
Conclusions—Peripheral blood mononuclear cell–derived EPCs bind platelets via CD62P and inhibit platelet activation,aggregation, adhesion to collagen, and thrombus formation, predominantly via upregulation of cyclooxygenase-2 andsecretion of prostacyclin. These findings add new insights into the biology of EPCs and define their potential roles inregulating platelet function and thrombosis. (Circulation. 2009;120:2230-2239.)
Key Words: progenitor cells � nitric oxide � platelets � prostaglandins � thrombosis
Endothelial progenitor cells (EPCs) are believed to con-tribute to vascular biology and hemostasis. Although
EPCs have been localized in cord blood, bone marrow, andperipheral blood as well as in some regenerative tissues,controversy relative to their isolation, identification, andfunctionality still exists. Indeed, the term EPC has beenemployed to describe different populations of multipotentcells that have the capability to differentiate, depending onthe environmental cues, into mature endothelial cells (ECs).These cells are extremely rare in peripheral blood, but theirnumber can be markedly increased after treatment withmobilizing cytokines or vascular trauma.1 Nevertheless, cellswith progenitor and endothelial phenotypes have been gen-erated in vitro from peripheral blood mononuclear cells(PBMCs)2–10 and have shown potential therapeutic applicationsin vascular tissue engineering and cell-based therapy.5,11–14 The
use of ex vivo expanded EPCs in animal and clinical studies hasshown beneficial effects in reendothelialization and neovas-cularization, as well as in the prevention of hybrid graftthrombosis and rejection11,12,15 and late stent thrombo-sis.14,16 However, the mechanisms that regulate mobiliza-tion, migration, and differentiation of EPCs and theirhoming to sites of vascular injury are complex and involveseveral mediators and receptors, such as P-selectin glycopro-tein ligand-1 (PSGL-1), �4 integrin, CXC chemokinereceptor-2 and -4, and �1- and �2-integrins.17–20 Furthermore,it has been shown that the interaction of platelets with EPCsinfluences their chemotaxis, adhesion, activation, and differ-entiation into mature ECs during vascular repair.17,19,21–24
Collectively, these studies suggest the existence of a crosstalk between EPCs and platelets in the regulation of eachother’s function.
Received January 14, 2009; accepted September 23, 2009.From the Research Center, Montreal Heart Institute and Université de Montréal, Faculty of Medicine (H.A.-S., D.Y., J.T., M.G., P.N., B.L., P.T.,
M.G.S., E.T., Y.M.); and McGill University (M.T.), Montreal, Quebec, Canada.The online-only Data Supplement is available with this article at http://circ.ahajournals.org/cgi/content/full/CIRCULATIONAHA.109.894642/DC1.Correspondence to Yahye Merhi, PhD, Laboratory of Thrombosis and Hemostasis, Montreal Heart Institute, 5000 Belanger, Montreal, Quebec, Canada,
H1T 1C8. E-mail [email protected]© 2009 American Heart Association, Inc.
Circulation is available at http://circ.ahajournals.org DOI: 10.1161/CIRCULATIONAHA.109.894642
2230 by on May 18, 2011 circ.ahajournals.orgDownloaded from

Clinical Perspective on p 2239Platelet activation and adhesion to damaged blood vessels
represent the first step in atherothrombosis.25 At sites ofvascular injury, platelets roll and interact with various com-ponents of the subendothelial matrix via a number of adhe-sive receptors expressed on the platelet surface. This, in turn,induces signaling that leads to platelet aggregation throughglycoprotein (GP) IIb/IIIa (�IIb�3 integrin) activation andbinding to fibrinogen.26 Platelet activation is accompanied bythe translocation of P-selectin (CD62P), which contributes tothe recruitment of other blood leukocytes.27 In this regard,CD62P has also been involved in the interaction of plateletswith EPCs.17,21,22
Although platelets promote the homing of EPCs at sites ofvascular injury and favor their differentiation into ECs, thefunctional consequences of such interactions on plateletsremain unknown. In fact, EPCs secrete many vasoactive andangiogenic factors that may modulate vascular thrombosisand hemostasis. Accordingly, we hypothesized that they
might influence platelet function. We therefore designed thepresent study to determine the impact and the mechanisms ofaction of EPCs on platelet function in vitro and in vivo. Wefound that human PBMC-derived EPCs bound platelets viaCD62P and inhibited platelet activation, aggregation, and adhe-sion to collagen in vitro and thrombus formation in vivo,predominantly via upregulation of cyclooxygenase-2 (COX-2)and secretion of prostacyclin (prostaglandin I2 [PGI2]).
MethodsMethods are available in the online-only Data Supplement. Thisstudy has been approved by the human and animal ethical commit-tees of the Montreal Heart Institute. All human subjects were healthyvolunteers of either sex aged from 20 to 55 years. They gaveinformed consent and were free from any drugs that interfere withplatelet function.
The authors had full access to and take full responsibility for theintegrity of the data. All authors have read and agree to themanuscript as written.
Figure 1. Characteristics of cultured cells. A, Immediately after plating, optical microscopy observation showed that freshly isolatedPBMCs with rounded morphology and irregular size appear either as single cells or as small clumps. At day 5, colonies are visible andscored as CFU-EPCs because they consist of a central core of round cells and elongated sprouting cells at the periphery. At day 10,cells form a flat monolayer of spindle-shaped cells characteristic of EPCs in culture (magnification �20). Staining of freshly isolatedPBMCs and 5- and 10- day cultured cells with ulex-lectin (green) and acetylated low-density lipoprotein (Ac-LDL) (red) under a laser-scanning confocal microscope is shown. Doubly stained cells appear after 5 days and remain positive up to 10 days of culture (bar�5�m). D.I.C. indicates differential interference contrast. B, Histogram showing the expression of different cell surface markers as deter-mined by flow cytometry. Single-color immunostaining of cultured cells was performed on day 0 (PBMCs) and days 5 and 10 with satu-rating concentrations of mouse anti-human PE-conjugated monoclonal antibodies directed against different mononuclear/leukocyte(CD14, PSGL-1, CD62L), progenitor (CD34, VEGFR2, CD62P), endothelial (VE-cadherin [VE-CAD], von Willebrand factor [vWF]), andplatelet (CD41) cell surface markers (n�3 to 6 independent experiments). C, Growth capacity of PBMC-derived EPCs after 10 days ofculture. On passaging, 10-day EPCs showed high proliferative potential (n�3).
Abou-Saleh et al Endothelial Progenitor Cells and Platelet Function 2231
by on May 18, 2011 circ.ahajournals.orgDownloaded from
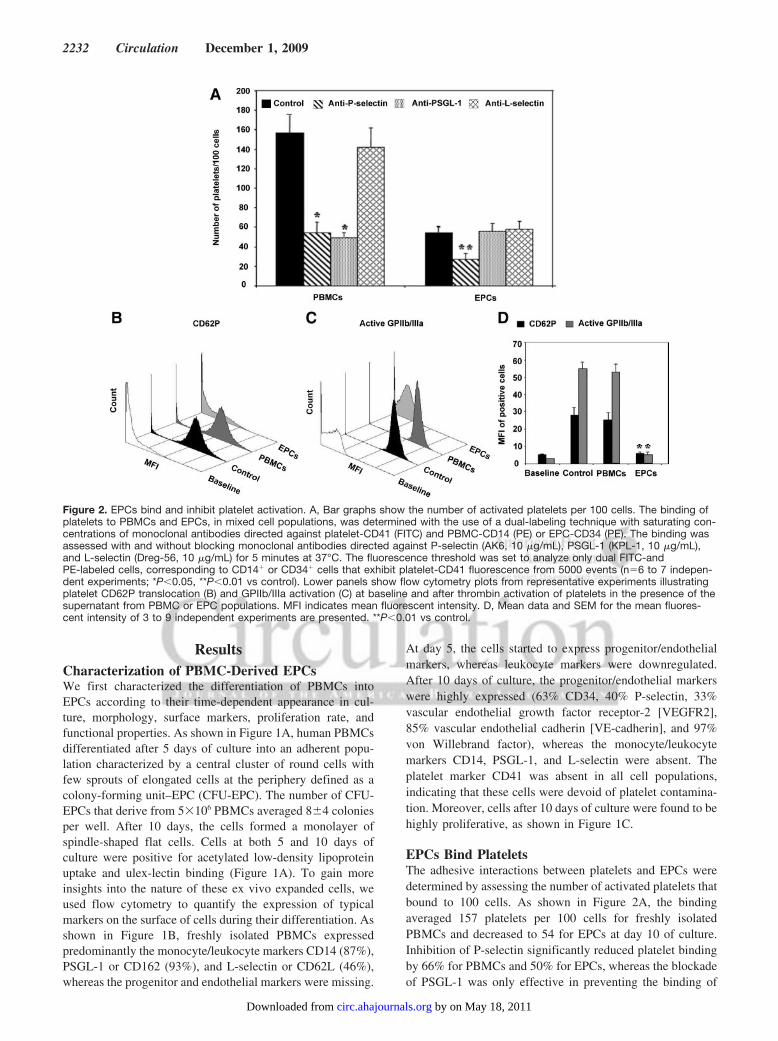
ResultsCharacterization of PBMC-Derived EPCsWe first characterized the differentiation of PBMCs intoEPCs according to their time-dependent appearance in cul-ture, morphology, surface markers, proliferation rate, andfunctional properties. As shown in Figure 1A, human PBMCsdifferentiated after 5 days of culture into an adherent popu-lation characterized by a central cluster of round cells withfew sprouts of elongated cells at the periphery defined as acolony-forming unit–EPC (CFU-EPC). The number of CFU-EPCs that derive from 5�106 PBMCs averaged 8�4 coloniesper well. After 10 days, the cells formed a monolayer ofspindle-shaped flat cells. Cells at both 5 and 10 days ofculture were positive for acetylated low-density lipoproteinuptake and ulex-lectin binding (Figure 1A). To gain moreinsights into the nature of these ex vivo expanded cells, weused flow cytometry to quantify the expression of typicalmarkers on the surface of cells during their differentiation. Asshown in Figure 1B, freshly isolated PBMCs expressedpredominantly the monocyte/leukocyte markers CD14 (87%),PSGL-1 or CD162 (93%), and L-selectin or CD62L (46%),whereas the progenitor and endothelial markers were missing.
At day 5, the cells started to express progenitor/endothelialmarkers, whereas leukocyte markers were downregulated.After 10 days of culture, the progenitor/endothelial markerswere highly expressed (63% CD34, 40% P-selectin, 33%vascular endothelial growth factor receptor-2 [VEGFR2],85% vascular endothelial cadherin [VE-cadherin], and 97%von Willebrand factor), whereas the monocyte/leukocytemarkers CD14, PSGL-1, and L-selectin were absent. Theplatelet marker CD41 was absent in all cell populations,indicating that these cells were devoid of platelet contamina-tion. Moreover, cells after 10 days of culture were found to behighly proliferative, as shown in Figure 1C.
EPCs Bind PlateletsThe adhesive interactions between platelets and EPCs weredetermined by assessing the number of activated platelets thatbound to 100 cells. As shown in Figure 2A, the bindingaveraged 157 platelets per 100 cells for freshly isolatedPBMCs and decreased to 54 for EPCs at day 10 of culture.Inhibition of P-selectin significantly reduced platelet bindingby 66% for PBMCs and 50% for EPCs, whereas the blockadeof PSGL-1 was only effective in preventing the binding of
Figure 2. EPCs bind and inhibit platelet activation. A, Bar graphs show the number of activated platelets per 100 cells. The binding ofplatelets to PBMCs and EPCs, in mixed cell populations, was determined with the use of a dual-labeling technique with saturating con-centrations of monoclonal antibodies directed against platelet-CD41 (FITC) and PBMC-CD14 (PE) or EPC-CD34 (PE). The binding wasassessed with and without blocking monoclonal antibodies directed against P-selectin (AK6, 10 �g/mL), PSGL-1 (KPL-1, 10 �g/mL),and L-selectin (Dreg-56, 10 �g/mL) for 5 minutes at 37°C. The fluorescence threshold was set to analyze only dual FITC-andPE-labeled cells, corresponding to CD14� or CD34� cells that exhibit platelet-CD41 fluorescence from 5000 events (n�6 to 7 indepen-dent experiments; *P�0.05, **P�0.01 vs control). Lower panels show flow cytometry plots from representative experiments illustratingplatelet CD62P translocation (B) and GPIIb/IIIa activation (C) at baseline and after thrombin activation of platelets in the presence of thesupernatant from PBMC or EPC populations. MFI indicates mean fluorescent intensity. D, Mean data and SEM for the mean fluores-cent intensity of 3 to 9 independent experiments are presented. **P�0.01 vs control.
2232 Circulation December 1, 2009
by on May 18, 2011 circ.ahajournals.orgDownloaded from
platelets to PBMCs but not to EPCs. As control, the bindingof platelets to PBMCs or EPCs was not affected by L-selectinblockade.
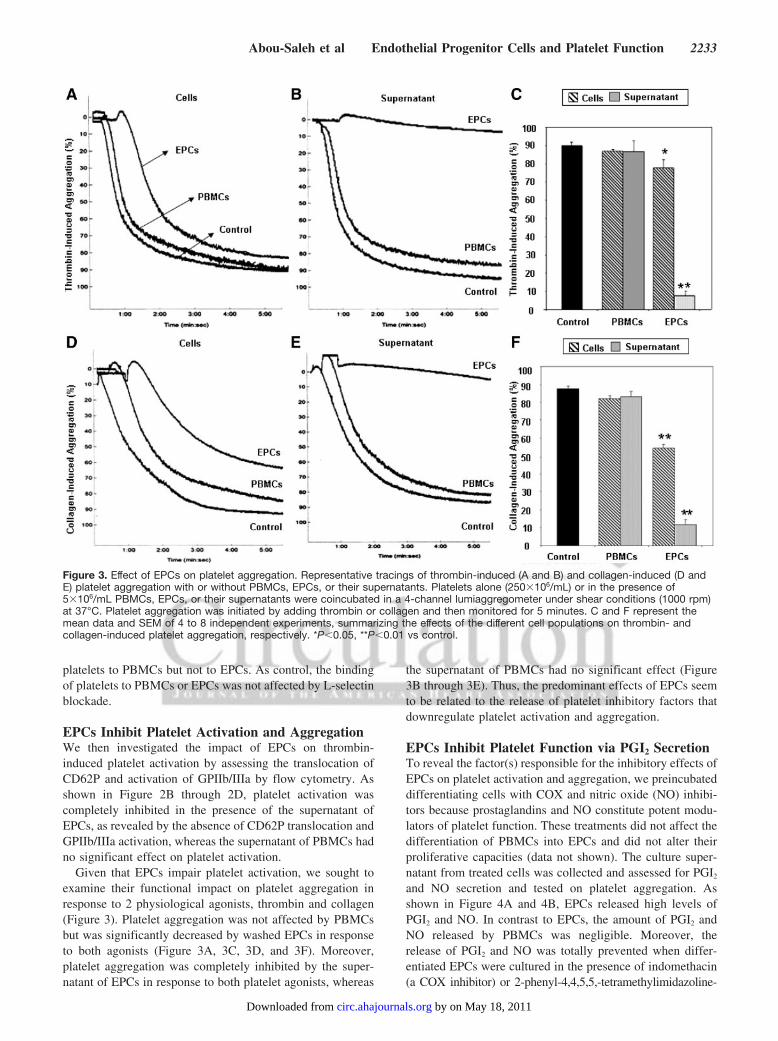
EPCs Inhibit Platelet Activation and AggregationWe then investigated the impact of EPCs on thrombin-induced platelet activation by assessing the translocation ofCD62P and activation of GPIIb/IIIa by flow cytometry. Asshown in Figure 2B through 2D, platelet activation wascompletely inhibited in the presence of the supernatant ofEPCs, as revealed by the absence of CD62P translocation andGPIIb/IIIa activation, whereas the supernatant of PBMCs hadno significant effect on platelet activation.
Given that EPCs impair platelet activation, we sought toexamine their functional impact on platelet aggregation inresponse to 2 physiological agonists, thrombin and collagen(Figure 3). Platelet aggregation was not affected by PBMCsbut was significantly decreased by washed EPCs in responseto both agonists (Figure 3A, 3C, 3D, and 3F). Moreover,platelet aggregation was completely inhibited by the super-natant of EPCs in response to both platelet agonists, whereas
the supernatant of PBMCs had no significant effect (Figure3B through 3E). Thus, the predominant effects of EPCs seemto be related to the release of platelet inhibitory factors thatdownregulate platelet activation and aggregation.
EPCs Inhibit Platelet Function via PGI2 SecretionTo reveal the factor(s) responsible for the inhibitory effects ofEPCs on platelet activation and aggregation, we preincubateddifferentiating cells with COX and nitric oxide (NO) inhibi-tors because prostaglandins and NO constitute potent modu-lators of platelet function. These treatments did not affect thedifferentiation of PBMCs into EPCs and did not alter theirproliferative capacities (data not shown). The culture super-natant from treated cells was collected and assessed for PGI2
and NO secretion and tested on platelet aggregation. Asshown in Figure 4A and 4B, EPCs released high levels ofPGI2 and NO. In contrast to EPCs, the amount of PGI2 andNO released by PBMCs was negligible. Moreover, therelease of PGI2 and NO was totally prevented when differ-entiated EPCs were cultured in the presence of indomethacin(a COX inhibitor) or 2-phenyl-4,4,5,5,-tetramethylimidazoline-
Figure 3. Effect of EPCs on platelet aggregation. Representative tracings of thrombin-induced (A and B) and collagen-induced (D andE) platelet aggregation with or without PBMCs, EPCs, or their supernatants. Platelets alone (250�106/mL) or in the presence of5�106/mL PBMCs, EPCs, or their supernatants were coincubated in a 4-channel lumiaggregometer under shear conditions (1000 rpm)at 37°C. Platelet aggregation was initiated by adding thrombin or collagen and then monitored for 5 minutes. C and F represent themean data and SEM of 4 to 8 independent experiments, summarizing the effects of the different cell populations on thrombin- andcollagen-induced platelet aggregation, respectively. *P�0.05, **P�0.01 vs control.
Abou-Saleh et al Endothelial Progenitor Cells and Platelet Function 2233
by on May 18, 2011 circ.ahajournals.orgDownloaded from

1-oxyl 3-oxide (PTIO) (a NO chelator), respectively. Tofurther validate the release of PGI2 and NO from EPCs, weassessed the expression of the constitutive and inducibleforms of COX (COX-1 and COX-2) and NO synthase (NOS)(endothelial NOS [eNOS] and inducible NOS [iNOS] iso-forms) in cultured EPCs, in comparison to PBMCs, byWestern blot. As shown in Figure 4C, COX-1 was detectablein freshly isolated PBMCs (day 0) and increased in EPCs atday 10. However, COX-2 was specifically upregulated afterthe differentiation of PBMCs into EPCs. Similarly, iNOS waspoorly expressed in PBMCs and profoundly increased inEPCs, whereas eNOS was barely detectable in both cellpopulations. Interestingly, the antiaggregatory or inhibitoryeffect of EPCs on platelets was reversed by incubatingcultured EPCs with indomethacin (even in the presence ofsuperoxide dismutase) or NS-398 (a COX-2 inhibitor) but notwith PTIO, NG-nitro-L-arginine (a NOS inhibitor), or 1400W(an iNOS inhibitor) (Figure 4D and 4E).
Taken together, these data indicate that, although EPCsspecifically upregulate the expression of COX-2 and iNOSand release PGI2 and NO, their main effect on plateletactivation and aggregation appears to be PGI2 dependent.
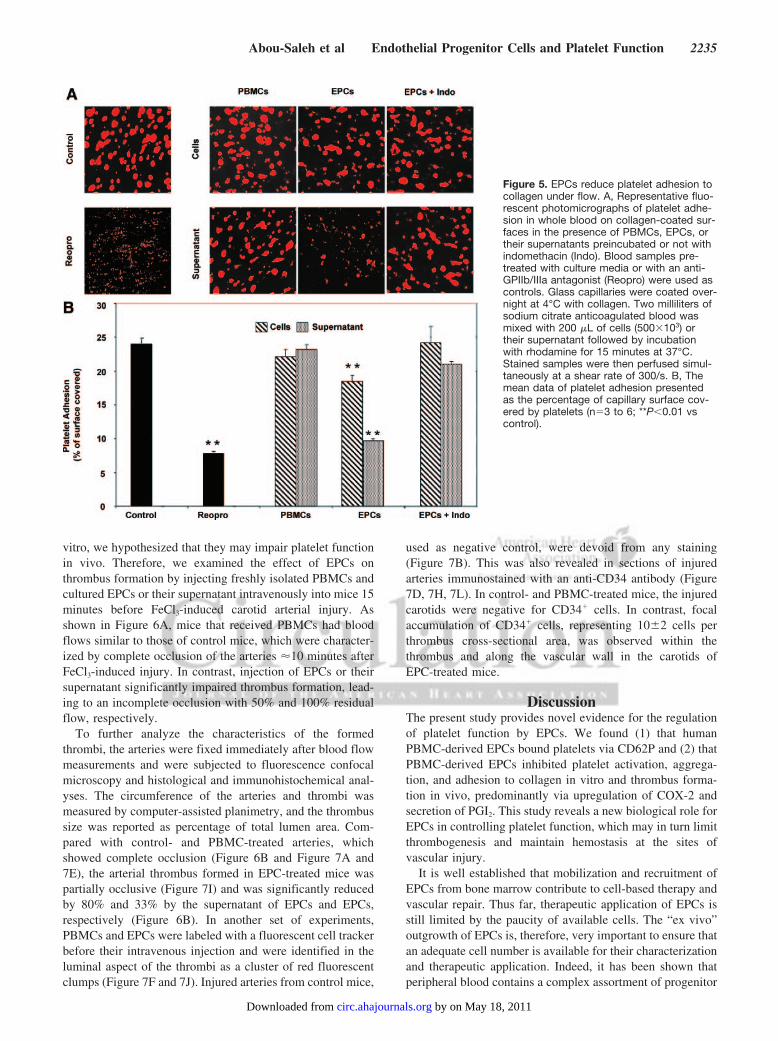
EPCs Reduce Platelet Adhesion Under FlowBecause EPCs inhibit activation and aggregation of washedplatelets, we then assessed their impact on platelet adhesionto collagen in a whole blood perfusion system. As shown inFigure 5, perfusion of whole blood resulted in almost 25%coverage of the collagen surface with platelets, which wasreduced to 7% by a GPIIb/IIIa antagonist, Reopro (positivecontrol). Coincubation of blood with PBMCs or their respec-tive supernatant had no significant effect on platelet adhesion.In contrast, the presence of EPCs or their supernatant reducedsignificantly platelet adhesion by 23% and 53%, respectively.The effects of EPCs and their supernatant were reversed, onceagain, by pretreating EPCs with the COX inhibitor indometh-acin. Hence, the effects of EPCs on activation and aggrega-tion of isolated platelets were also highlighted in a morephysiological approach of platelet adhesion under flow inwhole blood.
EPCs Incorporate at the Sites of Mouse CarotidInjury and Inhibit Thrombus FormationHaving shown the impact of EPCs on activation and aggre-gation of washed platelets and on adhesion in whole blood in
Figure 4. EPCs secrete PGI2 and NO and upregulate COX-2 and iNOS. Release of PGI2 (A) and NO (B) from PBMCs and EPCs pre-treated or not with indomethacin (Indo) (a COX inhibitor) or PTIO (a NO chelator) was assessed in the supernatants by radioimmunoas-say and nitrate/nitrite fluorometric assay, respectively (n�3; **P�0.01 vs EPCs). C, Expression of COX and NOS isoforms in PBMC andEPC populations by Western blot. �-Actin was used as an internal control. The blots shown are representative of 3 independent experi-ments. D, Representative tracings of thrombin-induced platelet aggregation with the supernatant of EPCs pretreated or not with indo-methacin or PTIO, showing the reversible effect of indomethacin, but not PTIO, on EPC-induced platelet aggregation inhibition. E, Themean data of thrombin-induced platelet aggregation are presented. The antiaggregatory or inhibitory effect of EPCs on platelets wasreversed by incubating cultured EPCs with indomethacin (even in the presence of superoxide dismutase [SOD]) or NS-398 (a COX-2inhibitor) but not with PTIO, NG-nitro-L-arginine (L-NNA, a NOS inhibitor), or 1400W (an iNOS inhibitor) (n�3 to 6; **P�0.01 vs control).
2234 Circulation December 1, 2009
by on May 18, 2011 circ.ahajournals.orgDownloaded from
vitro, we hypothesized that they may impair platelet functionin vivo. Therefore, we examined the effect of EPCs onthrombus formation by injecting freshly isolated PBMCs andcultured EPCs or their supernatant intravenously into mice 15minutes before FeCl3-induced carotid arterial injury. Asshown in Figure 6A, mice that received PBMCs had bloodflows similar to those of control mice, which were character-ized by complete occlusion of the arteries �10 minutes afterFeCl3-induced injury. In contrast, injection of EPCs or theirsupernatant significantly impaired thrombus formation, lead-ing to an incomplete occlusion with 50% and 100% residualflow, respectively.
To further analyze the characteristics of the formedthrombi, the arteries were fixed immediately after blood flowmeasurements and were subjected to fluorescence confocalmicroscopy and histological and immunohistochemical anal-yses. The circumference of the arteries and thrombi wasmeasured by computer-assisted planimetry, and the thrombussize was reported as percentage of total lumen area. Com-pared with control- and PBMC-treated arteries, whichshowed complete occlusion (Figure 6B and Figure 7A and7E), the arterial thrombus formed in EPC-treated mice waspartially occlusive (Figure 7I) and was significantly reducedby 80% and 33% by the supernatant of EPCs and EPCs,respectively (Figure 6B). In another set of experiments,PBMCs and EPCs were labeled with a fluorescent cell trackerbefore their intravenous injection and were identified in theluminal aspect of the thrombi as a cluster of red fluorescentclumps (Figure 7F and 7J). Injured arteries from control mice,
used as negative control, were devoid from any staining(Figure 7B). This was also revealed in sections of injuredarteries immunostained with an anti-CD34 antibody (Figure7D, 7H, 7L). In control- and PBMC-treated mice, the injuredcarotids were negative for CD34� cells. In contrast, focalaccumulation of CD34� cells, representing 10�2 cells perthrombus cross-sectional area, was observed within thethrombus and along the vascular wall in the carotids ofEPC-treated mice.
DiscussionThe present study provides novel evidence for the regulationof platelet function by EPCs. We found (1) that humanPBMC-derived EPCs bound platelets via CD62P and (2) thatPBMC-derived EPCs inhibited platelet activation, aggrega-tion, and adhesion to collagen in vitro and thrombus forma-tion in vivo, predominantly via upregulation of COX-2 andsecretion of PGI2. This study reveals a new biological role forEPCs in controlling platelet function, which may in turn limitthrombogenesis and maintain hemostasis at the sites ofvascular injury.
It is well established that mobilization and recruitment ofEPCs from bone marrow contribute to cell-based therapy andvascular repair. Thus far, therapeutic application of EPCs isstill limited by the paucity of available cells. The “ex vivo”outgrowth of EPCs is, therefore, very important to ensure thatan adequate cell number is available for their characterizationand therapeutic application. Indeed, it has been shown thatperipheral blood contains a complex assortment of progenitor
Figure 5. EPCs reduce platelet adhesion tocollagen under flow. A, Representative fluo-rescent photomicrographs of platelet adhe-sion in whole blood on collagen-coated sur-faces in the presence of PBMCs, EPCs, ortheir supernatants preincubated or not withindomethacin (Indo). Blood samples pre-treated with culture media or with an anti-GPIIb/IIIa antagonist (Reopro) were used ascontrols. Glass capillaries were coated over-night at 4°C with collagen. Two milliliters ofsodium citrate anticoagulated blood wasmixed with 200 �L of cells (500�103) ortheir supernatant followed by incubationwith rhodamine for 15 minutes at 37°C.Stained samples were then perfused simul-taneously at a shear rate of 300/s. B, Themean data of platelet adhesion presentedas the percentage of capillary surface cov-ered by platelets (n�3 to 6; **P�0.01 vscontrol).
Abou-Saleh et al Endothelial Progenitor Cells and Platelet Function 2235
by on May 18, 2011 circ.ahajournals.orgDownloaded from

cells that possess the ability to differentiate into EPCs invitro.28–30 In this connection, we succeeded in adequatelydifferentiating EPCs from PBMCs. The morphology of thesecells changed from a population of irregular and smallrounded cells to a flattened, spindle-shaped population thatforms CFU-EPCs during the differentiation process, uptakesDil–acetylated low-density lipoprotein, and binds ulex-lectin.Additionally, the expression pattern of surface markers ofcultured cells changed during differentiation, as demonstratedby a decrease in pan-monocyte/leukocyte markers and anincrease in progenitor/endothelial markers, as well as by theircapacity to proliferate after 10 days of culture. Such a subtypeof differentiated EPCs displayed some of the morphological,phenotypical, and functional characteristics of late EPCs thatderived from the adherent population of PBMCs when platedon collagen or fibronectin in the presence of endothelial-specific growth media.9,10,30,31
Accumulating evidence suggests that platelets play animportant role not only in hemostasis and thrombosis but alsoin inflammation and tissue repair through paracrine mecha-nisms or direct interactions with other blood cells.25,32 Re-cently, it has been shown that platelets promote the homingand differentiation of EPCs at sites of vascular injury.17–24
Conversely, EPCs may influence platelet function and mod-ulate their thrombogenic properties during vascular repair. Toanalyze and characterize the action of EPCs on plateletfunction, we first showed that platelets can bind EPCs but toa lower extent than the binding to PBMCs. The decrease inbinding during the differentiation process may be explainedby the dynamic changes in the expression of cell surfacemarkers, as well as by the upregulation of platelet inhibitingfactors, throughout the process of PBMC differentiation intoEPCs. Indeed, we observed a decrease in the expression ofPSGL-1 and an increase in the secretion of PGI2 and NO, thusproviding a possible explanation for the significant decreasein the binding of activated platelets to EPCs. We also showedthat PBMC-derived EPCs bind activated platelets in aCD62P-dependent manner, thus confirming previous findingsdemonstrating that the interaction between platelets and EPCsoccurs, in part, via CD62P and its high-affinity receptorPSGL-1.17,19,21–24 In addition to PSGL-1, other mediatorssuch as stromal cell–derived factor-1�, CXC chemokinereceptor-2 and -4, and �1- and �2-integrins participate inhoming of EPCs at sites of vascular injury.17–20
Moreover, incubation of platelets with the releasate ofEPCs resulted in a profound inhibition of platelet activation,
Figure 6. EPCs reduce thrombus formation in vivo. A, PBMCs and culture-derived EPCs (500�103 cells) or their supernatant wereinjected 15 minutes before FeCl3-induced injury of mouse carotid arteries, and thrombus formation was monitored by residual bloodflow measurements. Control experiments were done with culture media alone. After a stabilization period and application of FeCl3 for 3minutes, blood flow was monitored continuously for 20 minutes after FeCl3 injury. Tracings represent the mean data of 8 mice in thecontrol and EPCs groups, 6 in the PBMCs, and 3 in the supernatant of the EPC group (**P�0.01 vs control and PBMCs). B, Bargraphs show the mean data of thrombus size measured on cross-sectional area of arterial thrombi and expressed as percentage oflumen area. ** P�0.01 vs control and PBMCs.
2236 Circulation December 1, 2009
by on May 18, 2011 circ.ahajournals.orgDownloaded from

as revealed by the inhibition of CD62P translocation andGPIIb/IIIa activation, both of which are involved in theformation and stabilization of platelet thrombus.33,34 Thefunctional impact of EPCs on platelets was also highlightedby their inhibitory effects on thrombin- and collagen-inducedaggregation of washed platelets and adhesion to collagenunder flow in whole blood. The functional impact of EPCs onplatelets was further depicted in a mouse arterial thrombosismodel, in which injection of EPCs impaired thrombus massby 33%, leading to an incomplete occlusion with 50%residual flow. Interestingly, the supernatant of the EPCs wasmore efficacious than EPCs in inhibiting thrombus formationin vivo, thus confirming the in vitro findings that the mainimmediate antithrombotic effect of EPCs is related to therelease of platelet inhibitory factors. These antiplatelet prop-erties may ultimately lead to the development of novelEPC-derived antithrombotic therapies.
These findings may be of important physiopathologicalrelevance because the acute vascular response to injury
involves the adhesion, activation, and aggregation of plate-lets, followed by the recruitment of other platelets, leuko-cytes, and possibly circulating EPCs. The recruitment ofcirculating EPCs at the site of injury may contribute (1) toattenuate the acute thrombotic response by inhibiting plateletfunction and (2) to promote reendothelialization and vascularrepair. This may explain previous observations showing thatex vivo expanded EPCs seeded onto the lumen of a small-diameter vascular graft and implanted in vivo have a muchlower chance of thrombosis and subsequent rejection com-pared with uncoated controls.11,12 Moreover, the use of EPCcapture stents during percutaneous coronary interventionreduces the incidence of late stent thrombosis.14,16 Thus,EPCs modulate platelet function and appear to play animportant role in the management of thrombotic reactions.
The fundamental role of EPCs on platelets may be facili-tated primarily by the secretion of thromboresistant factorssuch as PGI2 and NO, which are well-known antiplateletmediators. In fact, we have shown that PBMC-derived EPCs
Figure 7. EPCs are recruited into arterial thrombi in vivo and reduce thrombus formation. Representative histological transverse sec-tions of FeCl3-injured mouse carotid arteries, stained with hematoxylin-eosin (H&E) and observed by optical microscopy, are shown(left). Arterial thrombus was completely occlusive in control- and PBMC-treated mice (A and E), whereas it was partially occlusive inarteries from EPC-treated mice (I) (magnification �10). Middle panels show confocal imaging of mouse carotid arteries after vascularinjury. PBMCs and EPCs were labeled with intracellular fluorescence marker (DiL), injected before arterial injury, and assessed by con-focal fluorescence on cryostat cross sections (F and J). Injured arteries from untreated mice were used as negative control (B). The cor-responding differential interference contrast (D.I.C.) of the identical sections is shown on the right (C, G, K) (bar�50 �m). Right panelsshow CD34 immunostaining of the injured arteries. CD34-positive cells appeared only in the EPC-treated mice, confirming the recruitment ofEPCs to the luminal aspect of arterial thrombi in vivo (arrows in L). The cells in D and H are CD34 negative (magnification �100).
Abou-Saleh et al Endothelial Progenitor Cells and Platelet Function 2237
by on May 18, 2011 circ.ahajournals.orgDownloaded from

specifically upregulate COX-2 and iNOS during their differ-entiation and produce PGI2 and NO. In the present study,however, we identify PGI2 as the principal mediator involvedin the inhibitory action of EPCs on platelet function. This isbased on our findings that, in contrast to PBMCs, whichconstitutively expressed COX-1 but did not produce PGI2,EPCs specifically upregulated COX-2 and produced PGI2.Additionally, the effects of EPCs on platelet function werereversed by a pan-COX inhibitor (indomethacin) even in thepresence of superoxide dismutase, which ruled out an impli-cation of NO in this phenomenon. Indeed, the effects of EPCswere also reversed by a specific COX-2 inhibitor (NS-398),whereas NO scavenging with PTIO, NOS inhibition withNG-nitro-L-arginine, or iNOS inhibition with 1400W waswithout any significant effects. Thus, PGI2 most likely rep-resents the major EPC-derived platelet inhibitor. The highlevels of NO released from EPCs could be correlated with ahigh expression of iNOS, whereas eNOS expression wasnegligible in EPCs. Our findings are in accordance with arecent study showing that iNOS, but not eNOS, is detectablein peripheral blood–derived EPCs.35 The low expression ofeNOS in EPCs derived from peripheral blood may beexplained by the fact that they are not fully matured ECs andare preferentially prone to produce NO in a monocytic/mac-rophage fashion through iNOS upregulation. This may sug-gest that the level of NO production in EPCs via iNOS is notsufficient to affect platelet function. Recently, Marjanovic etal36 have shown that NO synthesis during platelet activationnot only involves eNOS but is also mediated by the consti-tutively expressed platelet iNOS. iNOS-derived NO is impor-tant in promoting platelet activation, both ex vivo and in vivo,via a cGMP-dependent mechanism. Although our data pointto the lack of effects of EPC-derived NO on platelet aggre-gation, the contribution of platelet-derived NO and its signal-ing pathways, which are believed to play biphasic roles inplatelet function, on the effects of EPCs remained to beexplored. Moreover, the role of iNOS in EPCs appears to beimportant in angiogenesis, perhaps by promoting the recruit-ment of EPCs near the damaged endothelium.37 Takentogether, this may explain, in part, our observation that theinhibitory effects of EPCs on platelet function in vitro werepredominantly PGI2 dependent but not NO dependent. Thisdoes not negate, however, the role of EPC-derived NO as aregulatory factor involved in the secondary phase of vascularrepair, including angiogenesis.37
Our data demonstrate that EPCs regulate platelet functionvia upregulation of COX-2 and a PGI2-dependent inhibitionof platelet activation, aggregation, adhesion, and thrombusformation. In addition to the well-documented roles of EPCsin angiogenesis and vascular repair, our findings highlight anew biological role for EPCs in regulating platelet function,which may, in turn, limit thrombogenesis and maintainhemostasis at sites of vascular injury.
AcknowledgmentsWe are grateful to M. Louis Villeneuve for his assistance withconfocal microscopy.
Sources of FundingThis study was supported by grants from the Canadian Institute forHealth Research (MOP-82767 to Y.M.; IAP-73374 to E.T. andY.M.; and RMF-79023 to M.T. and Y.M.).
DisclosuresNone.
References1. Rafii S, Lyden D. Therapeutic stem and progenitor cell transplantation for
organ vascularization and regeneration. Nat Med. 2003;9:702–712.2. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T,
Witzenbichler B, Schatteman G, Isner JM. Isolation of putative progenitorendothelial cells for angiogenesis. Science. 1997;275:964–967.
3. Rehman J, Li J, Orschell CM, March KL. Peripheral blood “endothelialprogenitor cells” are derived from monocyte/macrophages and secreteangiogenic growth factors. Circulation. 2003;107:1164–1169.
4. Hur J, Yoon CH, Kim HS, Choi JH, Kang HJ, Hwang KK, Oh BH, LeeMM, Park YB. Characterization of two types of endothelial progenitorcells and their different contributions to neovasculogenesis. ArteriosclerThromb Vasc Biol. 2004;24:288–293.
5. Kalka C, Masuda H, Takahashi T, Kalka-Moll WM, Silver M, KearneyM, Li T, Isner JM, Asahara T. Transplantation of ex vivo expandedendothelial progenitor cells for therapeutic neovascularization. Proc NatlAcad Sci U S A. 2000;97:3422–3427.
6. Urbich C, Heeschen C, Aicher A, Dernbach E, Zeiher AM, Dimmeler S.Relevance of monocytic features for neovascularization capacity of cir-culating endothelial progenitor cells. Circulation. 2003;108:2511–2516.
7. Romagnani P, Lasagni L, Romagnani S. Peripheral blood as a source ofstem cells for regenerative medicine. Expert Opin Biol Ther. 2006;6:193–202.
8. Elsheikh E, Uzunel M, He Z, Holgersson J, Nowak G, Sumitran-Holgersson S. Only a specific subset of human peripheral-bloodmonocytes has endothelial-like functional capacity. Blood. 2005;106:2347–2355.
9. Ingram DA, Mead LE, Tanaka H, Meade V, Fenoglio A, Mortell K,Pollok K, Ferkowicz MJ, Gilley D, Yoder MC. Identification of a novelhierarchy of endothelial progenitor cells using human peripheral andumbilical cord blood. Blood. 2004;104:2752–2760.
10. Yoder MC, Mead LE, Prater D, Krier TR, Mroueh KN, Li F, Krasich R,Temm CJ, Prchal JT, Ingram DA. Redefining endothelial progenitor cellsvia clonal analysis and hematopoietic stem/progenitor cell principals.Blood. 2007;109:1801–1809.
11. Kaushal S, Amiel GE, Guleserian KJ, Shapira OM, Perry T, SutherlandFW, Rabkin E, Moran AM, Schoen FJ, Atala A, Soker S, Bischoff J,Mayer JE Jr. Functional small-diameter neovessels created using endo-thelial progenitor cells expanded ex vivo. Nat Med. 2001;7:1035–1040.
12. Shirota T, He H, Yasui H, Matsuda T. Human endothelial progenitorcell-seeded hybrid graft: proliferative and antithrombogenic potentials invitro and fabrication processing. Tissue Eng. 2003;9:127–136.
13. Werner N, Nickenig G. Clinical and therapeutical implications of EPCbiology in atherosclerosis. J Cell Mol Med. 2006;10:318–332.
14. Co M, Tay E, Lee CH, Poh KK, Low A, Lim J, Lim IH, Lim YT, Tan HC.Use of endothelial progenitor cell capture stent (Genous Bio-EngineeredR Stent) during primary percutaneous coronary intervention in acutemyocardial infarction: intermediate- to long-term clinical follow-up. AmHeart J. 2008;155:128–132.
15. Shirota T, Yasui H, Shimokawa H, Matsuda T. Fabrication of endothelialprogenitor cell (EPC)-seeded intravascular stent devices and in vitroendothelialization on hybrid vascular tissue. Biomaterials. 2003;24:2295–2302.
16. Miglionico M, Patti G, D’Ambrosio A, Di Sciascio G. Percutaneouscoronary intervention utilizing a new endothelial progenitor cellsantibody-coated stent: a prospective single-center registry in high-riskpatients. Catheter Cardiovasc Interv. 2008;71:600–604.
17. Massberg S, Konrad I, Schurzinger K, Lorenz M, Schneider S,Zohlnhoefer D, Hoppe K, Schiemann M, Kennerknecht E, Sauer S,Schulz C, Kerstan S, Rudelius M, Seidl S, Sorge F, Langer H, Peluso M,Goyal P, Vestweber D, Emambokus NR, Busch DH, Frampton J, GawazM. Platelets secrete stromal cell-derived factor 1alpha and recruit bonemarrow-derived progenitor cells to arterial thrombi in vivo. J Exp Med.2006;203:1221–1233.
2238 Circulation December 1, 2009
by on May 18, 2011 circ.ahajournals.orgDownloaded from

18. Chavakis E, Aicher A, Heeschen C, Sasaki K, Kaiser R, El Makhfi N,Urbich C, Peters T, Scharffetter-Kochanek K, Zeiher AM, Chavakis T,Dimmeler S. Role of beta2-integrins for homing and neovascularizationcapacity of endothelial progenitor cells. J Exp Med. 2005;201:63–72.
19. Langer H, May AE, Daub K, Heinzmann U, Lang P, Schumm M,Vestweber D, Massberg S, Schonberger T, Pfisterer I, Hatzopoulos AK,Gawaz M. Adherent platelets recruit and induce differentiation of murineembryonic endothelial progenitor cells to mature endothelial cells in vitro.Circ Res. 2006;98:e2–e10.
20. Hristov M, Zernecke A, Bidzhekov K, Liehn EA, Shagdarsuren E,Ludwig A, Weber C. Importance of CXC chemokine receptor 2 in thehoming of human peripheral blood endothelial progenitor cells to sites ofarterial injury. Circ Res. 2007;100:590–597.
21. Lev EI, Estrov Z, Aboulfatova K, Harris D, Granada JF, Alviar C,Kleiman NS, Dong JF. Potential role of activated platelets in homing ofhuman endothelial progenitor cells to subendothelial matrix. ThrombHaemost. 2006;96:498–504.
22. Daub K, Langer H, Seizer P, Stellos K, May AE, Goyal P, Bigalke B,Schonberger T, Geisler T, Siegel-Axel D, Oostendorp RA, Lindemann S,Gawaz M. Platelets induce differentiation of human CD34� progenitorcells into foam cells and endothelial cells. FASEB J. 2006;20:2559–2561.
23. Stellos K, Gawaz M. Platelet interaction with progenitor cells: potentialimplications for regenerative medicine. Thromb Haemost. 2007;98:922–929.
24. de Boer HC, Verseyden C, Ulfman LH, Zwaginga JJ, Bot I, Biessen EA,Rabelink TJ, van Zonneveld AJ. Fibrin and activated platelets coopera-tively guide stem cells to a vascular injury and promote differentiationtowards an endothelial cell phenotype. Arterioscler Thromb Vasc Biol.2006;26:1653–1659.
25. Davi G, Patrono C. Platelet activation and atherothrombosis. N EnglJ Med. 2007;357:2482–2494.
26. Ruggeri ZM, Mendolicchio GL. Adhesion mechanisms in plateletfunction. Circ Res. 2007;100:1673–1685.
27. Theoret JF, Bienvenu JG, Kumar A, Merhi Y. P-selectin antagonism withrecombinant P-selectin glycoprotein ligand-1 (rPSGL-Ig) inhibits circu-
lating activated platelet binding to neutrophils induced by damaged arte-rial surfaces. J Pharmacol Exp Ther. 2001;298:658–664.
28. Schatteman GC, Awad O. Hemangioblasts, angioblasts, and adult endo-thelial cell progenitors. Anat Rec A Discov Mol Cell Evol Biol. 2004;276:13–21.
29. Balbarini A, Barsotti MC, Di Stefano R, Leone A, Santoni T. Circulatingendothelial progenitor cells characterization, function and relationshipwith cardiovascular risk factors. Curr Pharm Des. 2007;13:1699–1713.
30. Prater DN, Case J, Ingram DA, Yoder MC. Working hypothesis toredefine endothelial progenitor cells. Leukemia. 2007;21:1141–1149.
31. Lin Y, Weisdorf DJ, Solovey A, Hebbel RP. Origins of circulatingendothelial cells and endothelial outgrowth from blood. J Clin Invest.2000;105:71–77.
32. Gawaz M, Langer H, May AE. Platelets in inflammation and athero-genesis. J Clin Invest. 2005;115:3378–3384.
33. Caron A, Theoret JF, Mousa SA, Merhi Y. Anti-platelet effects ofGPIIb/IIIa and P-selectin antagonism, platelet activation, and binding toneutrophils. J Cardiovasc Pharmacol. 2002;40:296–306.
34. Theoret JF, Chahrour W, Yacoub D, Merhi Y. Recombinant P-selectinglycoprotein-ligand-1 delays thrombin-induced platelet aggregation: anew role for P-selectin in early aggregation. Br J Pharmacol. 2006;148:299–305.
35. Muscari C, Gamberini C, Carboni M, Basile I, Farruggia G, Bonafe F,Giordano E, Caldarera CM, Guarnieri C. Different expression of NOSisoforms in early endothelial progenitor cells derived from peripheral andcord blood. J Cell Biochem. 2007;102:992–1001.
36. Marjanovic JA, Stojanovic A, Brovkovych VM, Skidgel RA, Du X.Signaling-mediated functional activation of inducible nitric-oxidesynthase and its role in stimulating platelet activation. J Biol Chem.2008;283:28827–28834.
37. Mayr U, Zou Y, Zhang Z, Dietrich H, Hu Y, Xu Q. Accelerated arterio-sclerosis of vein grafts in inducible NO synthase(�/�) mice is related todecreased endothelial progenitor cell repair. Circ Res. 2006;98:412–420.
CLINICAL PERSPECTIVEEndothelial progenitor cells (EPCs) represent a promising therapeutic approach for the treatment of cardiovasculardiseases. However, comprehensive delineation of the biology of these cells and the manner in which they interact withother blood and vascular cells is critical to fully understand their potential therapeutic properties. Herein, we addressed theinteractions between EPCs and platelets and their impact on platelet function and thrombus formation, which may berelevant to the management of atherothrombosis during acute coronary syndromes and after percutaneous coronaryinterventions. We found that human peripheral blood monocyte–derived EPCs in culture bound platelets via CD62P andinhibited platelet activation, aggregation, and adhesion to collagen in vitro, mainly via upregulation of cyclooxygenase-2and secretion of prostacyclin. Moreover, in a murine arterial thrombosis model, injection of EPCs led to their incorporationinto sites of injury and impaired thrombus formation, leading to an incomplete occlusion. In addition to thewell-documented roles of EPCs in angiogenesis and vascular repair, our findings highlight a new biological role for EPCsin regulating platelet function, which may, in turn, limit thrombogenesis while maintaining hemostasis at sites of vascularinjury. These antiplatelet properties may ultimately lead to the development of novel EPC-derived antithrombotic therapies.
Abou-Saleh et al Endothelial Progenitor Cells and Platelet Function 2239
by on May 18, 2011 circ.ahajournals.orgDownloaded from

CIRCULATIONAHA/2009/894642/R1
SUPPLEMENTAL MATERIAL
Supplemental Methods
Culture and characterization of EPCs
Human PBMCs were isolated from peripheral blood by centrifugation on a Ficoll density
gradient according to the manufacturer’s instructions (StemCell Technologies, Vancouver, BC).
The procedures of PBMC-derived EPCs in culture were adapted with modifications from
previously published methods.1-4 Briefly, unfractioned mononuclear cells were seeded onto
fibronectin coated 35-mm diameter 6-well tissue culture plates (BD Biosciences, Mississauga, ON) at
a density of 5 x 106 cells/well, and supplemented with EndoCultTM Liquid Medium (StemCell
Technologies). After 48 hours, non adherent cells were discarded by gentle washing with
phosphate buffer saline (PBS), and fresh media was applied. The adherent cells were continually
cultured in a complete media that was changed every 48 hours to allow formation of a well-
circumscribed monolayer of spindle-shaped cells. Freshly isolated PBMCs and the differentiated
cells after 5 and 10 days of culture were examined by optical and confocal microscopy. At day 5
of culture, the number of colonies, per well for each sample, was counted. Colonies were defined
as a central core or round cells with elongated sprouting cells at the periphery. Direct fluorescent
staining was used to detect the binding of FITC-Conjugated Ulex-lectin (Sigma-Aldrich,
Oakville, ON) and the uptake of Dil-Acetylated Low Density Lipoprotein (Ac-LDL, Molecular
probes, Eugene, OR) by laser confocal microscopy. The cells were plated with fresh media onto
Microwell Dishes (MatTek corporation, Ashland, MA) coated with fibronectin (5 μg/cm2, Roche,
Laval, Qc) for 1 hour at 37ºC. Cells were then incubated with Ac-LDL (2.5 μg/mL) and Ulex-
lectin (10 μg/mL) for 1 hour at 37°C, and fixed with 2% paraformaldehyde for 10 minutes. Cells
1
by on May 18, 2011 circ.ahajournals.orgDownloaded from

CIRCULATIONAHA/2009/894642/R1
positive for both Ulex-lectin and Ac-LDL were classified as EPCs. Fluorescent confocal images
(Z stacks) were acquired with a LSM 510 confocal microscope (Zeiss, Oberkochen, Germany). A
100x/1.3 Plan-Neofluar objective (Zeiss) was used for magnification. Z stacks-images were
deconvolved with the Huygens Pro 3.0 software (Scientific Volume Imaging, Hilversum, The
Netherlands), and the final images were reconstructed with the LSM510 software.
The growth capacity of the ex vivo expanded EPCs was examined by passing the cells
after 10 days of culture onto fibronectine coated 35-mm diameter 6-well tissue culture plates, and
grown in complete endothelial media. Two, 5, and 11 days after plating, the grown cells were
passed with Trypsin 0.25% -EDTA 1 mM mixture (US. Biological) and counted with a
hematological counter under phase-contrast microscopy (Leicha), with non viable cells discarded
through utilization of 4% trypan blue staining (Invitrogen).
Surface marker expression
The expression of specific pan-monocyte/leukocyte and progenitor/endothelial surface markers
on freshly isolated PBMCs and EPCs were assessed by flow cytometry using saturating
concentrations of mouse anti-human PE-conjugated monoclonal antibodies against CD14 (AbD
Serotec, Raleigh, NC), CD34 (BD Biosciences), CD162 or PSGL-1 (Santa Cruz, Santa Cruz,
CA), CD62P or P-selectin (BD Biosciences), CD62L or L-selectin (AbD Serotec), VEGF
Receptor 2 (VEGFR2, Abcam, Cambridge, MA), vWF (AbD Serotec), VE-Cadherin (BD
Biosciences) or CD41 (Dako Mississauga, ON). Briefly, freshly isolated PBMCs or cultured cells
at days 5 and 10 were detached from the culture plates by gentle scraping in 1 mM EDTA-PBS
(pH 7.2) under inverted microscopy. Cells were then re-suspended in PBS containing 0.5%
bovine serum albumin, plated in 96-microwell plates at 200 x 103 cells/well, blocked with normal
2
by on May 18, 2011 circ.ahajournals.orgDownloaded from

CIRCULATIONAHA/2009/894642/R1
mouse serum for 15 minutes at 4ºC, washed, and labeled with saturating concentrations of the
above monoclonal antibodies for 20 minutes. The samples were then washed, fixed with
paraformaldehyde, and analyzed with an Altra flow cytometer (Beckman Coulter, Mississauga,
ON) as described previously.5, 6 Results were expressed as the percentage of positive cells
obtained from 5000 events. Fluorescent isotype-matched antibodies were used as negative
controls.
Platelet activation and binding
Platelets were prepared as previously described,5-7 and adjusted to a final concentration of 250 x
106/mL. The binding of thrombin-activated platelets to PBMCs and EPCs, in mixed cell
populations of 50:1 ratio, was determined using a dual-labeling technique with monoclonal
antibodies directed against platelet-CD41 (FITC) and PBMC-CD14 (PE) or EPC-CD34 (PE).
The binding was assessed with and without blocking monoclonal antibodies directed against P-
selectin (AK6, 10 μg/mL, AbD Serotec) and PSGL-1 (KPL-1, 10 μg/mL, Santa Cruz) for
5 minutes at 37ºC, as detailed previously.5-7 Control experiments for specificity were done with a
blocking anti-CD62L (Dreg-56, 10 μg/mL, BD Biosciences). The fluorescence threshold was set
to analyze only dual FITC-and PE-labeled cells, corresponding to CD14+ or CD34+ that exhibited
platelet-CD41 fluorescence from 5000 events. The number of platelets bound to 100 cells was
calculated, as described previously.8
Platelet (250 x 106/mL) activation was determined by assessing P-selectin translocation
(CD62P, BD Biosciences) and GPIIb/IIIa activation5-7 (PAC-1, BD Biosciences) at baseline and
after stimulation with thrombin in the presence of cell-free supernatant, which was obtained by
3
by on May 18, 2011 circ.ahajournals.orgDownloaded from

CIRCULATIONAHA/2009/894642/R1
centrifugation (600 g, 10 minutes) of 5 x 106/mL of freshly isolated PBMCs or EPCs culture
media.
Platelet aggregation
Platelets, alone (250 x 106/mL) or in the presence of 5 x 106/mL PBMCs, EPCs, or their cell-free
supernatants, were co-incubated in a 4-channel lumi-aggregometer (Chronolog, Havertown, PA)
under shear conditions (1,000 rpm) at 37°C. Platelet aggregation was initiated by adding
thrombin (0.1 U/mL) or collagen (2 μg/mL) and then monitored by Aggro-Link software
(Chronolog) for 5 minutes, as described previously.6, 7
Platelet adhesion
The effects of PBMCs and EPCs on platelet adhesion to collagen under flow were assessed in an
ex-vivo perfusion system.9 Cells (500 x 103/200 µL) or their supernatant were incubated with 2
mL of sodium citrate anticoagulated blood that was labeled with 10 µg/mL Rhodamine 6G
(Sigma-Aldrich) for 15 minutes at 37°C. Stained samples were then perfused simultaneously in
glass capillaries (0.2 x 2.0 mm I.D., Fiber Optic Center Inc., Bedford, MA), pre-coated overnight
at 4°C with 250 µg/mL of fibrillar equine type 1 collagen (Chronolog), at a shear rate of
300/seconds using a KD scientific syringe pump (Fisher Scientific, Ottawa, ON). After a washing
step, images of the surfaces were captured using a digital Camera (Scion Corp., Frederick, MD)
connected to a Nikon Diaphot 200 epifluorescence microscope (Plan 10x 0.25, Mississauga, ON).
The percentage of platelet-covered surface, which represents platelet adhesion, was then
quantified by morphometric analyses using the ImageJ software (NIH, Betheseda, MD)
4
by on May 18, 2011 circ.ahajournals.orgDownloaded from

CIRCULATIONAHA/2009/894642/R1
PGI2 and NO measurements
To assess the release of PGI2 and NO, cells were incubated with culture media containing either a
COX inhibitor (indomethacin, 10 μmol/L, Calbiochem, Darmstadt, Germany), or a selective NO
scavenger (phenyl-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide or PTIO, 100 μmol/L, Sigma-
Aldrich). PGI2 release was measured with a radioimmunoassay (RIA, Sigma-Aldrich) for the
stable metabolite of prostacyclin, 6-keto-PGF1α. NO release was measured with a nitrate/nitrite
fluorometric assay kit (Cayman chemical, Ann Arbor, MI) according to the manufacturer’s
instructions. The concentrations of PGI2 and NO were normalized per unit number for each cell
type. Cell culture supernatants from control, indomethacin (10 µmol/L) with and without
superoxide dismutase (SOD, 250 U/mL, Sigma ), COX-2 inhibitor (NS-398, 10 µmol/L,
Calbiochem), PTIO (100 µmol/L), NOS inhibitor (Nω-Nitro-L-arginine or L-NNA, 100 μmol/L,
Sigma-Aldrich), or iNOS inhibitor (1400W, 10 µmol/L, Calbiochem) treated cells were collected
and used for platelet function assays.
Western Blot of COX and NOS
Twenty-five μg of protein from PBMCs, and EPCs (5 × 106 cells/mL) lysates were separated on
8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and blotted onto
nitrocellulose membranes. After blocking of non-specific binding, the membranes were incubated
overnight at 4°C with antibodies (1:200) against COX-1, COX-2, eNOS or iNOS (Santa Cruz),
and labeled with HRP-conjugated secondary antibodies (Santa-Cruz) at room temperature for
1 hour. Proteins were detected by enhanced chemiluminescence (PerkinElmer Life Sciences,
Walthman, MA). β-actin was used as an internal control.
5
by on May 18, 2011 circ.ahajournals.orgDownloaded from

CIRCULATIONAHA/2009/894642/R1
Model of mouse carotid thrombosis
The effects of freshly isolated PBMCs and culture-derived EPCs (500 x 103 cells)3, 10-12 or their
supernatant on thrombus formation were determined in a FeCl3 mouse carotid injury model,
according to a standardized protocol.13 Control experiments were done with culture media alone.
Briefly, C57BL6 mice (14-16 weeks old, Jackson laboratory, Bar Harbor, ME) were anesthetized
with a mixture of ketamine (Vetalar: 1.5 mg/kg I.P., Bioniche, Belleville, ON) and medetomidine
(Domitor: 1 mg/kg I.P., Pfizer, Kirkland, QC) and the right carotid artery was carefully exposed.
A miniature ultrasound flow probe (0.5 VB 552; Transonic Systems, Ithaca, NY), interfaced with
a flow meter (T206; Transonic Systems) and a computer-based data acquisition program (Iox
2.2.17.19, Emka, Falls Church, VA), was positioned around the artery. After measuring baseline
blood flow, a 0.5 × 1.0-mm strip of filter paper (Whatman no. 1) soaked in 6.5% FeCl3 was
applied on the adventitial surface proximal to the flow probe for 3 minutes, after which blood
flow and time to thrombotic occlusion (blood flow of 0 mL/minutes) were monitored.
Histology, immunostaining and confocal fluorescence
After completion of in vivo flow measurements in mice, the injured and the contralateral non-
injured carotids were excised and fixed in 10% buffered formalin. These arterial segments were
then embedded in paraffin, sectioned at 6 microns, and stained with hematoxylin and eosin, or
with an anti-CD34 antibody (Santa Cruz). Samples were visualized using an Olympus BX60
microscope (Olympus imaging America Inc., Center Valley, PA) and images were captured with
a Retiga 2000R camera (QImaging Corporation, Surrey, BC). The final images were acquired
with Image Pro Plus 6.2 software (Media Cybernetics, Bethesda, MD).
6
by on May 18, 2011 circ.ahajournals.orgDownloaded from

CIRCULATIONAHA/2009/894642/R1
For confocal fluorescence, 500 x103 of PBMCs or EPCs were labeled with an intracellular
photostable fluorescence marker (CellTrackerTM CM-DiI, Molecular Probes) according to the
manufacturer’s instructions. Cells were then washed with PBS, resuspended in fresh media and
injected intravenously, and allowed to circulate for 15 minutes before inducing the mouse carotid
injury. The carotids were excised immediately after completion of the blood flow measurements
and immersed in liquid nitrogen. Labeled PBMCs and EPCs incorporated into the luminal aspect
of arterial thrombi were observed on cryostat sections of 14-μm thickness using confocal
microscopy. Fluorescent confocal images (Z stacks) were acquired with a LSM 510 confocal
microscope (Zeiss). A 10x/1.3 Plan-Neofluar objective (Zeiss) was used for magnification. Z
stacks-images were deconvolved with the Huygens Pro 3.0 software (Scientific Volume
Imaging), and the final images were reconstructed with the LSM510 software.
Statistical analysis
Results are presented as mean ± SEM. The number of experiments for each test is indicated in the
figure legends. Statistical comparisons were done using the InStat3 Software (GraphPad software
inc. La Jolla, CA) for one-way ANOVA, followed by a Dunnett-t-test for comparison against a
single group. Data with p <0.05 were considered statistically significant.
Supplemental References
1. Krenning G, Dankers PY, Jovanovic D, van Luyn MJ, Harmsen MC. Efficient
differentiation of CD14+ monocytic cells into endothelial cells on degradable
biomaterials. Biomaterials. 2007;28:1470-1479.
7
by on May 18, 2011 circ.ahajournals.orgDownloaded from

CIRCULATIONAHA/2009/894642/R1
2. Rehman J, Li J, Orschell CM, March KL. Peripheral blood "endothelial progenitor cells"
are derived from monocyte/macrophages and secrete angiogenic growth factors.
Circulation. 2003;107:1164-1169.
3. Hur J, Yoon CH, Kim HS, Choi JH, Kang HJ, Hwang KK, Oh BH, Lee MM, Park YB.
Characterization of two types of endothelial progenitor cells and their different
contributions to neovasculogenesis. Arterioscler Thromb Vasc Biol. 2004;24:288-293.
4. Yoder MC, Mead LE, Prater D, Krier TR, Mroueh KN, Li F, Krasich R, Temm CJ, Prchal
JT, Ingram DA. Redefining endothelial progenitor cells via clonal analysis and
hematopoietic stem/progenitor cell principals. Blood. 2007;109:1801-1809.
5. Theoret JF, Bienvenu JG, Kumar A, Merhi Y. P-selectin antagonism with recombinant p-
selectin glycoprotein ligand-1 (rPSGL-Ig) inhibits circulating activated platelet binding to
neutrophils induced by damaged arterial surfaces. J Pharmacol Exp Ther. 2001;298:658-
664.
6. Yacoub D, Theoret JF, Villeneuve L, Abou-Saleh H, Mourad W, Allen BG, Merhi Y.
Essential role of protein kinase C delta in platelet signaling, alpha IIb beta 3 activation,
and thromboxane A2 release. J Biol Chem. 2006;281:30024-30035.
7. Abou-Saleh H, Theoret JF, Yacoub D, Merhi Y. Neutrophil P-selectin-glycoprotein-
ligand-1 binding to platelet P-selectin enhances metalloproteinase 2 secretion and platelet-
neutrophil aggregation. Thromb Haemost. 2005;94:1230-1235.
8. Evangelista V, Manarini S, Rotondo S, Martelli N, Polischuk R, McGregor JL, de
Gaetano G, Cerletti C. Platelet/polymorphonuclear leukocyte interaction in dynamic
conditions: evidence of adhesion cascade and cross talk between P-selectin and the beta 2
integrin CD11b/CD18. Blood. 1996;88:4183-4194.
8
by on May 18, 2011 circ.ahajournals.orgDownloaded from

CIRCULATIONAHA/2009/894642/R1
9
9. Andre P, LaRocca T, Delaney SM, Lin PH, Vincent D, Sinha U, Conley PB, Phillips DR.
Anticoagulants (thrombin inhibitors) and aspirin synergize with P2Y12 receptor
antagonism in thrombosis. Circulation. 2003;108:2697-2703.
10. Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, Witzenbichler B,
Schatteman G, Isner JM. Isolation of putative progenitor endothelial cells for
angiogenesis. Science. 1997;275:964-967.
11. Kalka C, Masuda H, Takahashi T, Kalka-Moll WM, Silver M, Kearney M, Li T, Isner
JM, Asahara T. Transplantation of ex vivo expanded endothelial progenitor cells for
therapeutic neovascularization. Proc Natl Acad Sci U S A. 2000;97:3422-3427.
12. Hristov M, Zernecke A, Bidzhekov K, Liehn EA, Shagdarsuren E, Ludwig A, Weber C.
Importance of CXC chemokine receptor 2 in the homing of human peripheral blood
endothelial progenitor cells to sites of arterial injury. Circ Res. 2007;100:590-597.
13. Konstantinides S, Schafer K, Thinnes T, Loskutoff DJ. Plasminogen activator inhibitor-1
and its cofactor vitronectin stabilize arterial thrombi after vascular injury in mice.
Circulation. 2001;103:576-583.
by on May 18, 2011 circ.ahajournals.orgDownloaded from
Copyright © 2022 FDOKUMEN




















