
Triggering by Paf-acether and adrenaline of cyclo-oxygenase-independent platelet aggregation

Enhanced induction of heme oxygenase-1 suppressesthrombus formation and affects the protein C systemin sepsis
DONGSHENG FEI, XIANGLIN MENG, MINGRAN ZHAO, KAI KANG, GANG TAN,SHANGHA PAN, YUNPENG LUO, WEN LIU, CHUANCHUAN NAN, HONGCHI JIANG,GEOFFREY W. KRISSANSEN, MINGYAN ZHAO, and XUEYING SUN
HARBIN, CHINA; AND AUKLAND, NEW ZEALAND
From the Department of ICU; Key H
Department of General Surgery, The
Medical University, Harbin, China;
Hospital of Harbin City, Harbin,
Medicine & Pathology, the Uni
New Zealand.
Supported by grants 30872987, 30
National Natural Scientific Foun
ZJY0704-02 from the Heilongjia
Foundation, China.
Heme oxygenase-1 (HO-1) displays anti-inflammatory and cytoprotective activitiesin sepsis. Here, we investigated the effects of HO-1 on thrombus formation and theprotein C system in a septic C57BL/6 mouse model induced by cecal ligation andperforation (CLP). Septic mice were either preinjected with the vehicle, pretreatedwith hemin (an HO-1 inducer) or zinc protoporphyrin IX (ZnPP, an HO-1 inhibitor), orgiven a combination of hemin 1 ZnPP. CLP increased significantly the hepatic ex-pression of HO-1; increased thrombosis in livers, kidneys, and lungs; shortened theprothrombin time (PT) and activated partial thromboplastin time (APTT); elevatedthe levels of tumor necrosis factor-1a (TNF-1a), interleukin-6 (IL-6), and thrombomo-dulin (TM); reduced the levels of protein C (PC) and activated protein C (aPC); anddownregulated hepatic expression of PC and TM. The preadministration of heminto septic mice increased the expression and activity of HO-1; inhibited thrombosisin the preceding 3 organs; prolonged PT and APTT; inhibited the production of TNF-aand IL-6; upregulated the expression of PC and TM in livers; elevated the plasmalevels of PC and aPC; and reduced the plasma levels of TM. In contrast, ZnPP showedopposite effects to hemin and reversed the effects of hemin by inhibiting the activityof HO-1. The administration of tricarbonyl dichloro ruthenium (II) dimer (CORM-2),which is a CO-releasing molecule, had a similar effect to hemin on thrombosis andthe protein C system. The data indicate that the enhanced induction of HO-1 inhibitsthrombus formation and affects the protein C system in sepsis. (Translational Re-search 2012;159:99–109)
Abbreviations: aPC¼ activated protein C; APTT¼ activated partial thromboplastin time; CLP¼cecal ligation and perforation; CORM ¼ CO-releasing molecule; DIC ¼ disseminated intravas-cular coagulation; DMSO ¼ dimethyl sulfoxide; ELISA ¼ enzyme-linked immunosorbent assay;HE ¼ hematoxylin and eosin; HO-1 ¼ heme oxygenase-1; iCORM ¼ inactive CO-releasingmolecule; IL ¼ interleukin; LPS ¼ lipopolysaccharide; MSB ¼ martius/scarlet/blue; PBS ¼phosphate-buffered saline; PC ¼ protein C; PT ¼ prothrombin time; SD ¼ standard deviation;TM ¼ thrombomodulin; TNF-a ¼ tumor necrosis factor-a; ZnPP ¼ zinc protoporphyrin IX
epatosplenic Surgery Laboratory,
First Affiliated Hospital of Harbin
Department of Pediatrics, Child’s
China; Department of Molecular
versity of Auckland, Auckland,
973474, and 81171785 from the
dation of China and by Grant
ng Provincial Natural Scientific
Submitted for publication June 20, 2011; revision submitted October
18, 2011; accepted for publication October 25, 2011.
Reprint requests: Xueying Sun, MD, PhD, Department of Molecular
Medicine & Pathology, the University of Auckland, Auckland 1005,
New Zealand; e-mail: [email protected].
1931-5244/$ - see front matter
� 2012 Mosby, Inc. All rights reserved.
doi:10.1016/j.trsl.2011.10.009
99

AT A GLANCE COMMENTATRY
Fei D, et al.
Background
Heme oxygenase-1 (HO-1) displays extensive
physiologic activities including anti-inflammation
against septic shock and antithrombosis. The pro-
tein C system plays an important role in the mech-
anisms underlying coagulation derangement during
systemic inflammation, such as sepsis. However,
the effects of HO-1 on thrombosis induced by
sepsis and whether HO-1 influences the protein C
system remain unknown.
Translational Significance
Enhanced induction of HO-1 by hemin inhibits
thrombus formation and affects the protein C sys-
tem in sepsis.
Translational Research100 Fei et al February 2012
Sepsis remains a challenging problem for criticallyill patients, with a mortality rate of approximately40%.1 It is characterized by systemic inflammationand a hypercoagulable state, leading to a myriadof complications including microvascular collapse,multiple organ failure, and death.2 The inflammation-induced activation of coagulation is mediated by theoverproduction of proinflammatory cytokines, suchas interleukin (IL)-6, which plays a role in the initia-tion of coagulation activation, and tumor necrosisfactor-a (TNF-a), which inhibits physiologic anticoa-gulation.3,4 An extensive cross talk exists between thepathways mediating inflammation and coagulation,whereby inflammation triggers the coagulation cascadeand coagulation influences inflammatory activitymarkedly.The protein C (PC) system plays an important role in
the mechanisms underlying coagulation derangementduring systemic inflammation.5 PC, which is a key pro-tein member of the PC system, is a vitamin K-depen-dent glycoprotein synthesized by the liver, whichcirculates in plasma as an inactive zymogen and isactivated when binding to thrombin.6 Its activation isaccelerated greatly by thrombomodulin (TM), whichis an endothelial cell transmembrane glycoproteinthat binds to thrombin and functions as a cofactor inthe thrombin-induced activation of PC.7 Activated PC(aPC) is the central effector protease of the PC path-way. It interacts with the PC receptor on endothelialcells and with other cellular receptors to exert multipleeffects on hemostasis and immune cell function,5 thus
promoting fibrinolysis and inhibiting both coagulationand inflammation.The levels of aPC are deficient in septic patients, cor-
responding with increased morbidity and mortality,8
suggesting that increasing the levels of aPC might beof therapeutic benefit.9 Intravenous infusion of septicrats with aPC inhibited the production of inflammatorycytokines, reduced leukocyte trafficking, and preservedhepatic function, resulting in a 30% increase in sur-vival.10 Infusion of aPC improved cardiovascular dys-function in rats exposed to lipopolysaccharide (LPS)by decreasing LPS-induced inflammation and oxidativestress.11 In another study, the infusion of aPC reducedsystemic inflammation and preserved renal function inseptic rats.12
Heme oxygenase-1 (HO-1) is an inducible enzymethat converts heme into CO, biliverdin, and freeiron.13 It has attracted considerable interest becauseof its extensive physiologic functions and potentialtherapeutic application.14 HO-1 and the metabolites itproduces display anti-inflammatory,15 antioxidative,and antiapoptotic activities.16-18 HO-1 suppressed theinfiltration of neutrophils into rat livers during sepsis,19
contributed to the protection of rat kidneys againstseptic shock,20 and displayed beneficial effects in thepatients with severe sepsis and septic shock.21 HO-1–derived CO enhanced the host–defense responseagainst bacterial sepsis in mice.22 Furthermore, the in-duction of HO-1 also inhibited vascular arterial andvenous thrombosis in animal models,23-26 and in ac-cord, HO-1–deficiency accelerated formation of arterialthrombosis in mice.27 Here, we sought to determine theeffects of HO-1 on thrombosis induced by septicinflammation, and whether HO-1 affects the protein Csystem.
METHODS
Sepsis model. Cecal ligation and perforation (CLP)were performed on 8–10-week-old male C57BL/6mice supplied by the Animal Research Center at theFirst Affiliated Hospital of Harbin Medical University,Harbin, China. All surgical procedures and careadministered to the animals were approved by theinstitutional ethics committee. The mice were fastedfor 12 h but allowed free access to water beforeanesthesia, which was induced by an intraperitonealinjection of pentobarbital (70 mg/kg). A midlinelaparotomy was performed and the cecum identified.Stool contents were milked to the tip of the cecum,which was subsequently ligated 1 cm from the tip witha 2-0 silk tie. The cecum was then perforated with a 22G needle and returned into the abdomen. Theabdominal wall was closed with a continuous 3-0 silk

Translational ResearchVolume 159, Number 2 Fei et al 101
suture. No antibiotics were used, and the animals hadfree access to food and water postoperatively.
Chemicals. Hemin (an HO-1 inducer) or zincprotoporphyrin-IX (ZnPP, an HO-1 inhibitor) (Sigma-Aldrich, St. Louis, Mo) were dissolved in dimethylsulfoxide (DMSO) and then diluted with phosphate-buffered saline (PBS) to make a solution witha concentration of 10 mmol/mL. The CO-releasingmolecule, tricarbonyldichlororuthenium (II) dimer(CORM-2) (Sigma-Aldrich) was dissolved in DMSO(100 mmol/mL) and diluted with PBS to achievea concentration of 2 mmol/mL. Inactivated CORM-2(iCORM-2) was prepared by incubating CORM-2dissolved in DMSO (100 mmol/mL) for 24 h at 37�Cin a 5% CO2 atmosphere to liberate CO completelyand then diluted with PBS to achieve a concentrationof 2 mmol/mL. All solutions were freshly prepared.The drug doses and modes of application were chosenbased on previously published studies.28-30
Experimental groups. One hundred twenty mice wereassigned to 4 groups (each group had 30 mice) thateither were sham operated, or received a single intraper-itoneal injection of PBS (vehicle), hemin, or ZnPP ata dose of 50 mmol/kg, and then underwent CLP 12 hlater. The sham-operated mice underwent laparotomyand bowel manipulation, and they received anintraperitoneal injection of PBS without CLP. Tostandardize the experiments, all the mice receiveda similar volume of solution, which contained thesame level of DMSO (2% in volume). Six mice fromeach group were killed randomly 4, 8, 12, 18, and24 h post-CLP. The blood was collected via cardiacpuncture with heparin-treated syringe-needles,immediately mixed with 3.2% sodium citrate at a ratioof 9:1 (vol/vol), and then centrifuged at 10003 g for10 min to harvest the plasma. The inferior vena cavawas cut, and the tissues were perfused with 10 ml ofphysiologic saline first through the right ventricle andnext the left ventricle, and then the lungs, kidneys, andlivers were harvested. To investigate whether ZnPPcould reverse the effects of hemin in CLP mice, anexperiment was performed by including three groupsof animals (each group had 6 mice), which received anadministration of vehicle, hemin, or hemin 1 ZnPP asabove and killed 12 h post-CLP. To investigatewhether CO, which is the downstream product ofHO-1, has a similar beneficial effect to hemin, anotherexperiment was performed by including 4 groups ofanimals (each group had 6 mice), which receivedintravenous administration of PBS, CORM-2 (16mmole/kg), or iCORM2 (16 mmole/kg) via tail vein,underwent CLP as indicated, and then killed 12 hpost-CLP.
Coagulation parameters. The prothrombin time (PT)was measured by incubating 50 mL plasma for 1 minat 37�C and then adding 100 mL antiheparin calciumthromboplastin (Steellex Scientific Instrument Com-pany, Beijing, China). The activated partial thrombo-plastin time (APTT) was measured by mixing 500 mLAPTT reagent (Steellex Scientific Instrument Com-pany) and 50 mL plasma followed by incubation for3 min at 37�C and recalcification with 50 mL40 mmol/L CaCl2.
Enzyme-linked immunosorbent assay (ELISA). Theplasma levels of IL-6, TNF-a, PC, aPC, and TM weremeasured by using ELISA kits (Shanghai BlueGeneBiotech Co., Ltd., Shanghai, China) according to themanufacturer’s instructions.
Histologic analysis and fibrin staining. The specimenswere fixed with 10% buffered formalin, embeddedin paraffin, sectioned, and examined under lightmicroscopy. Transverse sections made at 5 differentlevels to cover the entire organ, and 4 fields wereselected randomly from each section; thus, 20 fields(magnification 203) were examined in total for eachorgan for counting the number of thrombi. An addi-tional 2 sections from each organ were prepared forstaining fibrin by the martius/scarlet/blue (MSB)methods31 to confirm the formation of thrombi, andthe numbers of MSB-stained thrombi were countedalso.
Immunohistochemical analysis. The tissue sectionswere blocked with 3% bovine serum albumin andincubated with an Ab against mouse HO-1 (SantaCruz Biotechnology, Inc, Santa Cruz, Calif). Anisotype-matched Ab was used as a control. They wereincubated subsequently with the appropriate secondaryAbs using the Ultra Sensitive TMS-P kit (ZhongshanCo, Beijing, China), and immunoreactivity developedwith Sigma FAST DAB (3,3’-diaminobenzidinetetrahydrochloride) and CoCl2 enhancer tablets (Sigma-Aldrich, Shanghai, China). The sections were counter-stained with hematoxylin, mounted, and examined bymicroscopy.
Western blot analysis. The methodology has beendescribed previously.32 Briefly, the tissues werehomogenized, and the proteins were resolved on poly-acrylamide SDS gels and transferred electro-phoretically to polyvinylidene difluoride membranes.The membranes were blocked; incubated with Absagainst mouse HO-1, PC, or TM (Santa CruzBiotechnology, Inc), respectively; and incubatedsubsequently with an alkaline phosphatase-conjugatedsecondary Ab. They were developed with 5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium(Tiangen Biotech Co. Ltd., Beijing, China). The levels

Translational Research102 Fei et al February 2012
of proteins were normalized with respect to the banddensity of b-actin as an internal control.
Measurement of HO activity. Hepatic HO-1 activitywas determined by measuring the production ofbilirubin as described previously33 with modifications.Liver tissues were homogenized in homogenizationbuffer (20 mmol/L potassium phosphate buffer (pH7.4), 250 mmol/L sucrose, 2 mmol/L ethylenedi-aminetetraacetic acid, 2 mmol/L phenylmethyl sulfonylfluoride, and 10 lg/mL leupeptin). The homogenateswere centrifuged at 10,000 3 g for 30 min, and thesupernatant was further centrifuged at 100,000 3 g for1 h at 4�C. The pellet was suspended with potassiumphosphate buffer followed by sonication for 2 s at 4�C(microsome fraction). The microsomal protein (1 mg),cytosolic fraction of liver tissues (1 mg; for source ofbiliverdine reductase), 10 mmol/L hemin, and 0.8mmol/L nicotinamide adenine dinucleotide phosphateswere mixed in 1 mL potassium phosphate buffer.The reaction mixture was incubated for 60 min at 37�Cin dark. The reaction was terminated by addition of1 mL chloroform. The bilirubin concentration wasdetermined spectrophotometically by measuring thedifference in absorbance between 465 and 530 nm,with a molar extinction coefficient of 40/mol/L/cm.
Statistical analysis. The results were expressed asmeanvalues 6 standard deviation (SD). A 1-way analysis ofvariance followed by the post-hoc Dunnett’s test wasused for evaluating statistical significance (SPSS 17.0).A value of P , 0.05 was considered significant.
RESULTS
HO-1 expression and activity in livers. CLP induceda marked increase (P , 0.001) in the expression ofHO-1 protein 12 h post-CLP (Fig 1, B) compared withthe levels of HO-1 observed in sham-operated mice(Fig 1, A), as illustrated by immunohistochemicalanalysis of liver sections and confirmed by Westernblot analysis (Fig 1, E). The administration of hemincaused a significant (P , 0.05) increase of HO-1protein expression in livers of the CLP mice (Fig 1, Cand E), whereas ZnPP had a neglect effect on theexpression of HO-1 (Fig 1, D and E) compared withthe vehicle-injected CLP mice. We also measured theactivity of HO-1 in the livers. As shown in Fig 1, F,the HO-1 activity was (P , 0.001) increased by CLPsignificantly compared with that in sham-operatedmice, whereas hemin significantly (P , 0.001)increased HO-1 activity even more compared withthat in vehicle-injected CLP mice. However, ZnPP(P , 0.05) reduced HO-1 activity significantlycompared with vehicle-injected CLP mice.
Thrombus formation. A quantitative analysis of thenumbers of thrombi in the hematoxylin and eosin(HE)-stained sections of livers, lungs, and kidneysshowed that CLP had increased the numbers of thrombisignificantly (all P , 0.001) in the 3 organs (Fig 2, A).Preadministration of hemin to the CLP mice reducedthe numbers of thrombi significantly (P , 0.05) in the3 organs (Fig 2, A) compared with the vehicle.Preadministration of ZnPP resulted in a significant (bothP , 0.05) increase in the number of thrombi in kidneysand lungs, but only a slight increase (P . 0.05) in thenumber of thrombi in livers compared with the vehicle.The tissue sections were stained even more by MSB toview fibrin. As shown in Fig 2, B, CLP induceda marked thrombus formation in the livers, kidneys, andlungs, whereas the 3 organs from the sham-operatedmice had almost no thrombi. Preadministration ofhemin reduced, whereas ZnPP increased, the formationof thrombi in the 3 organs compared with the vehicle(Fig 2, B). A quantitative analysis of the numbers ofthrombi in MSB-stained sections (Fig 2, C) showeda similar pattern to that shown in HE-stained sections(Fig 2, A). However, the numbers of thrombi inMSB-stained sections were lower than that obtained inHE-stained sections, implying that some thrombi shownin the HE-stained sections may be formed loosely andpossibly caused by delayed perfusion of the organs.
Coagulation parameters. As shown in Fig 2, D and E,vehicle-injected CLP mice had significantly (both P ,0.01) shorter PT and APTT than the sham-operatedmice. The administration of hemin significantly (bothP , 0.05) prolonged the PT and APTT in the CLPmice compared with the vehicle. However, ZnPP-treated CLP mice had even shorter PT and APTT thanvehicle-injected CLP mice, but the difference did notreach significance.
Plasma levels of inflammatory cytokines. As shown inFig 3, the plasma levels of TNF-a and IL-6 wereincreased dramatically in the CLP mice 6 h post-CLP.The increase of TNF-a was (P , 0.05) diminishedsignificantly by the preadministration of hemin butincreased slightly (P . 0.05) by ZnPP in the CLPmice (Fig 3, A). Similarly, the increased level of IL-6was attenuated significantly (P , 0.05) by hemin, butincreased significantly (P , 0.05) by ZnPP (Fig 3, B).
HO-1 regulates the expression of PC and TM. As shownin Fig 4, A, PC and TM were both downregulated in thelivers of vehicle-injected CLP mice. The preadmini-stration of hemin attenuated the reduced expression ofPC and TM, whereas ZnPP reduced their expressioneven more compared with the vehicle. The plasmalevels of PC (Fig 4, B) and aPC (Fig 4, C) werereduced significantly (all P , 0.001) at all the

Fig 1. HO-1 expression and activity in livers. (A–D) Liver sections from sham-operated mice (A) or CLP mice
pretreated with vehicle (B), hemin (C), or ZnPP (D) were immunostained with an anti–HO-1 Ab and examined by
microscopy. (E) The liver tissues from sham-operated mice (lane 1) or CLP mice pretreated with vehicle (lane 2),
hemin (lane 3), or ZnPP (lane 4) were homogenized and Western blotted with an anti-HO-1 Ab. b-actin served as
an internal control. Band density was measured and normalized to the internal control to calculate relative band
density. (F) HO-1 activity in livers from the preceding mice was measured. The percent activity was compared
with that in untreated healthy mice, which are set to 100. Results were expressed as means 6 SD (n 5 6).
*P , 0.05 or **P , 0.001 vs sham-operated mice. †P , 0.05, significant increase from vehicle-treated CLP
mice. ‡P, 0.05, significant reduction from vehicle-treated CLPmice. (Color version of figure is available online.)
Translational ResearchVolume 159, Number 2 Fei et al 103

Fig 2. Thrombus formation and coagulation parameters. (A) Sections of the livers, kidneys, and lungs from sham-
operated mice or CLP mice pretreated with vehicle, hemin, or ZnPP were stained with HE and examined by
Translational Research104 Fei et al February 2012

Fig 3. Plasma levels of TNF-a and IL-6. The blood samples were
collected from sham-operated mice, or CLP mice pre-treated with ve-
hicle, hemin, or ZnPP 6 h post-CLP. The plasma levels of TNF-a (A)
and IL-6 (B) were measured. The results were expressed as means 6SD (n 5 6). **P , 0.001 vs sham-operated mice. †P , 0.05, signif-
icant increase from vehicle-treated CLP mice. ‡P , 0.05, significant
reduction from vehicle-treated CLP mice.
=
Translational ResearchVolume 159, Number 2 Fei et al 105
indicated time points, whereas the plasma levels of TM(Fig 4,D) were increased significantly (allP, 0.001) invehicle-injected CLP mice compared with the sham-operated mice. The preadministration of hemin led tosignificant (all P , 0.05) increases in the levels of PCand aPC at all the indicated time points, whereasZnPP lowered the plasma levels of PC and aPCsignificantly compared with vehicle-injectedCLP mice (Fig 4, B and C). The preadministration ofhemin reduced the plasma levels of TM significantlyat all the indicated time points, whereas ZnPPincreased the plasma levels of TM significantlycompared with the vehicle-injected CLP mice(Fig 4, D).
ZnPP reverses the effects of hemin. To investigatewhether ZnPP could reverse the beneficial effects ofhemin on sepsis-induced thrombosis, the vehicle,
microscopy. The numbers of thrombi were counted in 20 rand
(B) The preceding sections were stained by the MSB to view
indicated (C). (D, E) The blood samples were taken from the
measured. The results were expressed as means6 SD (n5 6†P , 0.05, significant increase from vehicle-injected CLP m
injected CLP mice. (Color version of figure is available onli
hemin, or ZnPP 1 hemin was preadministered intothe CLP mice, which were killed 12 h post-CLP.First, we could show that the administration ofZnPP 1 hemin reduced significantly (P , 0.001) theHO-1 activity in livers enhanced by hemin, resultingin no significant difference in HO-1 activity betweenZnPP 1 hemin-treated and vehicle-injected CLPmice. The mice treated with hemin 1 ZnPP hadsignificantly (all P , 0.05) higher numbers ofthrombi in livers, kidneys, and lungs than hemin-treated mice but had no significant difference fromvehicle-injected mice (Fig 5, B). Furthermore, themice treated with hemin 1 ZnPP had significantlyshorter PT and APTT than hemin-treated mice buthad no significant difference from vehicle-treatedmice (Fig 5, C). The data indicate that ZnPP couldreverse the effect of hemin on sepsis-inducedthrombosis completely.
CO suppresses thrombosis and affects the PCsystem. Because of the nonspecific feature of hemin,we investigated whether the administration ofCORM-2, which is a CO-releasing molecule, couldhave a similar beneficial effect to hemin on sepsis-induced thrombosis. The administration of CORM-2significantly prolonged PT (Fig 6, A) and APTT(Fig 6, B), reduced the numbers of thrombi in livers(Fig 6, C), kidneys (Fig 6, D), and lungs (Fig 6, E),increased plasma levels of PC (Fig 6, F) and aPC(Fig 6, G), and reduced the plasma levels of TM(Fig 6, H) compared with the vehicle. The changes ofthe preceding parameters in CORM-2-treated micewere in the similar patterns compared with that inhemin-treated mice (Figs 2 and 4). However, iCORM-2, which is an inactivated CORM-2 form, had negativeeffects compared with the vehicle. The results indicatethat supplementing CO, which is a downstreamproduct of HO-1, has a similar beneficial effect tohemin, which is an HO-1 inducer, on sepsis-inducedthrombosis by inhibiting thrombus formation andaffecting the protein C system.
DISCUSSION
Sepsis is the leading cause of death in critically illpatients, and its incidence continues to increase.22
Sepsis can evoke disseminated intravascular coagula-tion (DIC), resulting in multiple organ failure and
omly selected fields (magnification 203) per organ.
fibrin, and the MSB-stained thrombi were counted as
preceding mice, and the PT (D) and APTT (E) were
). *P, 0.05 or **P,0.001 vs sham-operated mice.
ice. ‡P , 0.05, significant reduction from vehicle-
ne.)

Fig 4. Levels of PC, TM, and aPC. (A) Liver tissues were harvested from sham-operated mice (lane 1) or CLP
mice pretreated with vehicle (lane 2), hemin (lane 3), or ZnPP (lane 4) 12 h post-CLP, homogenized, and Western
blotted with Abs against PC and TM. b-actin served as an internal control. The band density was measured and
normalized to the internal control to calculate the relative band density. (B–D) Blood samples were collected
from sham-operated mice or CLP mice pretreated with vehicle, hemin, or ZnPP at the indicated time points.
The plasma levels of PC (B), aPC (C), and TM (D) were measured. The results were expressed as means 6SD (n 5 6). *P , 0.05 or **P , 0.001 vs sham-operated mice. †P , 0.05, significant increase from vehicle-
treated CLP mice. ‡P , 0.05, significant reduction from vehicle-treated CLP mice.
Translational Research106 Fei et al February 2012
even death. Therefore, it is critical to explore patho-physiologic alterations in coagulant responses duringsepsis in an effort to seek new therapeutic options.The current study has demonstrated that hemin in-duced the overexpression of HO-1 and enhanced theactivity of HO-1 in the livers, and it inhibited sepsis-induced thrombosis significantly. It suppressed theproduction of TNF-a and IL-6, upregulated the he-patic expression of PC and TM, increased the plasmalevels of PC and aPC, and reduced the plasma levels ofTM. In marked contrast, ZnPP (which inhibited HO-1activity) had the opposite effects to hemin and re-versed the beneficial effects of hemin on sepsis-
induced thrombosis. Furthermore, the administrationof CORM-2, which is a CO-releasing molecule,showed a similar effect to hemin on thrombosis andthe PC system during sepsis. In accord, HO-1-derived CO has been reported to protect the hostagainst bacterial sepsis in mice.22 The results pre-sented herein suggest that the protective effect ofHO-1 may be attributed to its ability to inhibitTNF-a and IL-6 production, and to increase the levelsof PC, aPC, and TM in tissues. HO-1 has been re-ported previously to exert cytoprotective activity inhepatic injury.34 The latter activity may contribute tothe antithrombogenic activity of HO-1, as local
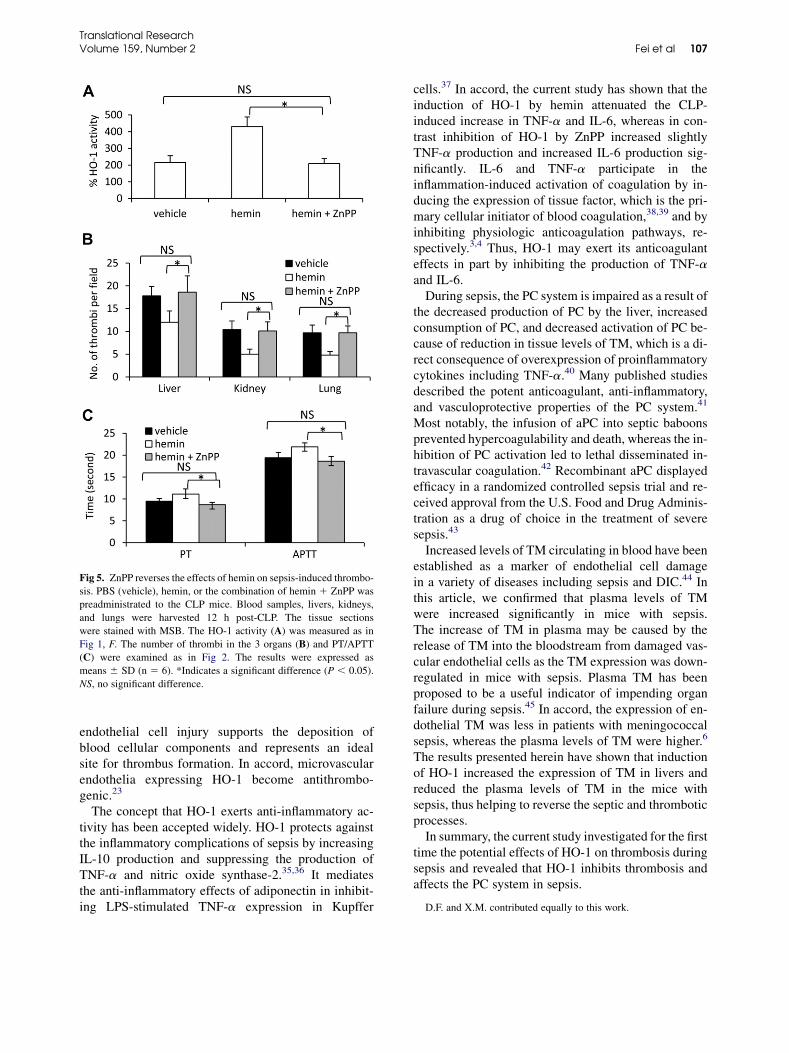
Fig 5. ZnPP reverses the effects of hemin on sepsis-induced thrombo-
sis. PBS (vehicle), hemin, or the combination of hemin 1 ZnPP was
preadministrated to the CLP mice. Blood samples, livers, kidneys,
and lungs were harvested 12 h post-CLP. The tissue sections
were stained with MSB. The HO-1 activity (A) was measured as in
Fig 1, F. The number of thrombi in the 3 organs (B) and PT/APTT
(C) were examined as in Fig 2. The results were expressed as
means 6 SD (n 5 6). *Indicates a significant difference (P , 0.05).
NS, no significant difference.
Translational ResearchVolume 159, Number 2 Fei et al 107
endothelial cell injury supports the deposition ofblood cellular components and represents an idealsite for thrombus formation. In accord, microvascularendothelia expressing HO-1 become antithrombo-genic.23
The concept that HO-1 exerts anti-inflammatory ac-tivity has been accepted widely. HO-1 protects againstthe inflammatory complications of sepsis by increasingIL-10 production and suppressing the production ofTNF-a and nitric oxide synthase-2.35,36 It mediatesthe anti-inflammatory effects of adiponectin in inhibit-ing LPS-stimulated TNF-a expression in Kupffer
cells.37 In accord, the current study has shown that theinduction of HO-1 by hemin attenuated the CLP-induced increase in TNF-a and IL-6, whereas in con-trast inhibition of HO-1 by ZnPP increased slightlyTNF-a production and increased IL-6 production sig-nificantly. IL-6 and TNF-a participate in theinflammation-induced activation of coagulation by in-ducing the expression of tissue factor, which is the pri-mary cellular initiator of blood coagulation,38,39 and byinhibiting physiologic anticoagulation pathways, re-spectively.3,4 Thus, HO-1 may exert its anticoagulanteffects in part by inhibiting the production of TNF-aand IL-6.During sepsis, the PC system is impaired as a result of
the decreased production of PC by the liver, increasedconsumption of PC, and decreased activation of PC be-cause of reduction in tissue levels of TM, which is a di-rect consequence of overexpression of proinflammatorycytokines including TNF-a.40 Many published studiesdescribed the potent anticoagulant, anti-inflammatory,and vasculoprotective properties of the PC system.41
Most notably, the infusion of aPC into septic baboonsprevented hypercoagulability and death, whereas the in-hibition of PC activation led to lethal disseminated in-travascular coagulation.42 Recombinant aPC displayedefficacy in a randomized controlled sepsis trial and re-ceived approval from the U.S. Food and Drug Adminis-tration as a drug of choice in the treatment of severesepsis.43
Increased levels of TM circulating in blood have beenestablished as a marker of endothelial cell damagein a variety of diseases including sepsis and DIC.44 Inthis article, we confirmed that plasma levels of TMwere increased significantly in mice with sepsis.The increase of TM in plasma may be caused by therelease of TM into the bloodstream from damaged vas-cular endothelial cells as the TM expression was down-regulated in mice with sepsis. Plasma TM has beenproposed to be a useful indicator of impending organfailure during sepsis.45 In accord, the expression of en-dothelial TM was less in patients with meningococcalsepsis, whereas the plasma levels of TM were higher.6
The results presented herein have shown that inductionof HO-1 increased the expression of TM in livers andreduced the plasma levels of TM in the mice withsepsis, thus helping to reverse the septic and thromboticprocesses.In summary, the current study investigated for the first
time the potential effects of HO-1 on thrombosis duringsepsis and revealed that HO-1 inhibits thrombosis andaffects the PC system in sepsis.
D.F. and X.M. contributed equally to this work.

Fig 6. CO suppresses thrombus formation and affects the PC system. Sham-operated mice, or CLP mice pread-
ministered with PBS (vehicle), CORM-2, or iCORM-2 were killed 12 h post-CLP, and the blood samples, livers,
kidneys, and lungs were harvested. PT (A) and APTT (B) were measured. The sections of liver (C), kidney (D),
and lung (E) were stained withMSB, and the numbers ofMSB-stained thrombi were counted. The plasma levels of
PC (F), aPC (G), and TM (H) were measured. The results were expressed as means6 SD (n5 6). *P , 0.05 or
**P, 0.001 vs sham-operated mice. †P, 0.05, significant increase from vehicle-injected CLP mice. ‡P, 0.05,
significant reduction from vehicle-injected CLP mice.
Translational Research108 Fei et al February 2012
REFERENCES
1. Vincent JL, Sakr Y, Sprung CL, et al. Sepsis in European intensive
care units: results of the SOAP study. Crit Care Med 2006;34:
344–53.
2. Ince C. The microcirculation is the motor of sepsis. Crit Care
2005;9:S13–9.
3. Van der Poll T, Levi M, Hack CE, et al. Elimination of interleukin
6 attenuates coagulation activation in experimental endotoxemia
in chimpanzees. J Exp Med 1994;179:1253–9.
4. Van Deventer SJ, Buller HR, ten Cate JW, et al. Experimental en-
dotoxemia in humans: analysis of cytokine release and coagula-
tion, fibrinolytic, and complement pathways. Blood 1990;76:
2520–6.
5. Weiler H. Regulation of inflammation by the protein C system.
Crit Care Med 2010;38:S18–25.
6. Faust SN, LevinM, Harrison OB, et al. Dysfunction of endothelial
protein C activation in severe meningococcal sepsis. N Engl J
Med 2001;345:408–16.

Translational ResearchVolume 159, Number 2 Fei et al 109
7. DittmanWA, Majerus PW. Structure and function of thrombomo-
dulin: a natural anticoagulant. Blood 1990;75:329–36.
8. Fisher CJ Jr, Yan SB. Protein C levels as a prognostic indicator of
outcome in sepsis and related diseases. Crit Care Med 2000;28:
S49–56.
9. Joyce DE, Gelbert L, Ciaccia A, et al. Gene expression profile of
antithrombotic protein c defines new mechanisms modulating in-
flammation and apoptosis. J Biol Chem 2001;276:11199–203.
10. Huynh T, Nguyen N, Keller S, et al. Reducing leukocyte traffick-
ing preserves hepatic function after sepsis. J Trauma 2010;69:
360–7.
11. Sennoun N, Meziani F, Dessebe O, et al. Activated protein C im-
proves lipopolysaccharide-induced cardiovascular dysfunction by
decreasing tissular inflammation and oxidative stress. Crit Care
Med 2009;37:246–55.
12. Keller SA, Moore CC, Evans SL, McKillop IH, Huynh T. Acti-
vated protein C alters inflammation and protects renal function
in sepsis. J Surg Res 2011;168:e103–9.
13. Maines MD, Trakshel GM, Kutty RK. Characterization of two
constitutive forms of rat liver microsomal heme oxygenase.
Only one molecular species of the enzyme is inducible. J Biol
Chem 1986;261:411–9.
14. Yao P, Hao L, Nussler N, et al. The protective role of HO-1 and its
generated products (CO, bilirubin and Fe) in ethanol-induced hu-
man hepatocyte damage. Am J Physiol Gastrointest Liver Physiol
2009;296:G1318–23.
15. Otterbein LE, Bach FH, Alam J, et al. Carbon monoxide has anti-
inflammatory effects involving the mitogen-activated protein ki-
nase pathway. Nat Med 2000;6:422–8.
16. Yachie A, Niida Y,Wada T, et al. Oxidative stress causes enhanced
endothelial cell injury in human heme oxygenase-1 deficiency.
J Clin Invest 1999;103:129–35.
17. Tsui TY, Siu YT, Schlitt HJ, et al. Heme oxygenase-1-derived car-
bon monoxide stimulates adenosine triphosphate generation in
human hepatocyte. Biochem Biophys Res Commun 2005;336:
898–902.
18. Ke B, Buelow R, Shen XD, et al. Heme oxygenase 1 gene transfer
prevents CD95/Fas ligand-mediated apoptosis and improves liver
allograft survival via carbon monoxide signaling pathway. Hum
Gene Ther 2002;13:1189–99.
19. Lin YT, Chen YH, Yang YH, et al. Heme oxygenase-1 suppresses
the infiltration of neutrophils in rat liver during sepsis through in-
activation of p38 MAPK. Shock 2010;34:615–21.
20. Yu JB, Zhou F, Yao SL, et al. Effect of heme oxygenase-1 on the
kidney during septic shock in rats. Transl Res 2009;153:283–7.
21. Takaki S, Takeyama N, Kajita Y, et al. Beneficial effects of the
heme oxygenase-1/carbon monoxide system in patients with se-
vere sepsis/septic shock. Intensive Care Med 2010;36:42–8.
22. Chung SW, Liu X, Macias AA, Baron RM, Perrella MA. Heme
oxygenase-1-derived carbon monoxide enhances the host defense
response to microbial sepsis in mice. J Clin Invest 2008;118:
239–47.
23. Lindenblatt N, Bordel R, Schareck W, Menger MD, Vollmar B.
Vascular heme oxygenase-1 induction suppresses microvascular
thrombus formation in vivo. Arterioscler Thromb Vasc Biol
2004;24:601–6.
24. Johns DG, Zelent D, Ao Z, et al. Heme-oxygenase induction in-
hibits arteriolar thrombosis in vivo: effect of the non-substrate
inducer cobalt protoporphyrin. Eur J Pharmacol 2009;606:
109–14.
25. Desbuards N, Rochefort GY, Schlecht D, et al. Heme oxygenase-1
inducer hemin prevents vascular thrombosis. Thromb Haemost
2007;98:614–20.
26. Tracz MJ, Juncos JP, Grande JP, et al. Induction of heme
oxygenase-1 is a beneficial response in a murine model of venous
thrombosis. Am J Pathol 2008;173:1882–90.
27. True AL, OliveM, BoehmM, et al. Heme oxygenase-1 deficiency
accelerates formation of arterial thrombosis through oxidative
damage to the endothelium, which is rescued by inhaled carbon
monoxide. Circ Res 2007;101:893–901.
28. Lindenblatt N, Bordel R, Schareck W, et al. Vascular heme
oxygenase-1 induction suppresses microvascular thrombus for-
mation in vivo. Arterioscler Thromb Vasc Biol 2004;24:601–6.
29. AmonM,MengerMD, Vollmar B. Heme oxygenase and nitric ox-
ide synthase mediate cooling-associated protection against TNF-
a-induced microcirculatory dysfunction and apoptotic cell death.
FASEB J 2003;17:175–85.
30. Sun B, Sun Z, Jin Q, ChenX. CO-releasingmolecules (CORM-2)-
liberated CO attenuates leukocytes infiltration in the renal tissue
of thermally injured mice. Int J Biol Sci 2008;4:176–83.
31. Yang HL, Jiang XC, Zhang XY, et al. Thrombocytopenia in the
experimental leptospirosis of guinea pig is not related to dissem-
inated intravascular coagulation. BMC Infect Dis 2006;6:19.
32. Kang K, Zhao M, Jiang H, et al. The role of hydrogen sulfide in
hepatic ischemia reperfusion-induced injury in rats. Liver Transpl
2009;15:1306–14.
33. Hirai K, Sasahira T, Ohmori H, Fujii K, Kuniyasu H. Inhibition of
heme oxygenase-1 by zinc protoporphyrin IX reduces tumor
growth of LL/2 lung cancer in C57BL mice. Int J Cancer 2007;
120:500–5.
34. Carchman EH, Rao J, Loughran PA, Rosengart MR,
Zuckerbraun BS. Heme oxygenase-1-mediated autophagy pro-
tects against hepatocyte cell death and hepatic injury from infec-
tion/sepsis in mice. Hepatology 2011;53:2053–62.
35. Lee TS, Chau LY. Heme oxygenase-1 mediates the anti-
inflammatory effect of IL-10 in mice. Nat Med 2002;8:240–6.
36. Drechsler Y, Dolganiuc A, Norkina O, et al. Heme oxygenase-1
mediates the anti-inflammatory effects of acute alcohol on
IL-10 induction involving p38 mapk activation in monocytes.
J Immunol 2006;177:2592–600.
37. Mandal P, Park PH, McMullen MR, Pratt BT, Nagy LE. The anti-
inflammatory effects of adiponectin are mediated via a heme
oxygenase-1-dependent pathway in rat Kupffer cells. Hepatology
2010;51:1420–9.
38. Kambas K, Markiewski MM, Pneumatikos IA, et al. C5a and
TNF-alpha up-regulate the expression of tissue factor in intraal-
veolar neutrophils of patients with the acute respiratory distress
syndrome. J. Immunol 2008;180:7368–75.
39. Szotowski B, Antoniak S, PollerW, Schultheiss HP, RauchU. Pro-
coagulant soluble tissue factor is released from endothelial cells in
response to inflammatory cytokines. Circ Res 2005;96:1233–9.
40. Van der Poll T, Boer JD, Levi M. The effect of inflammation on
coagulation and vice versa. Curr Opin Infect Dis 2011;24:273–8.
41. Danese S, Vetrano S, Zhang L, et al. The protein C pathway in tis-
sue inflammation and injury: pathogenic role and therapeutic im-
plications. Blood 2010;115:1121–30.
42. Taylor FB Jr, Chang A, Esmon CT, et al. Protein C prevents the
coagulopathic and lethal effects of Escherichia coli infusion in
the baboon. J Clin Invest 1987;79:918–25.
43. Pet€aj€a J. Inflammation and coagulation. An overview. Thromb
Res 2011;127:S34–7.
44. Takano S, Kimura S, Ohdama S, Aoki N. Plasma thrombomodulin
in health and diseases. Blood 1990;76:2024–9.
45. Iba T, Yagi Y, Kidokoro A, Fukunaga M, Fukunaga T. Increased
plasma levels of soluble thrombomodulin in patients with sepsis
and organ failure. Surg Today 1995;25:585–90.
Copyright © 2022 FDOKUMEN




















