
The mitochondrial heme exporter FLVCR1b mediates erythroid differentiation
Upload
independentCategory
view
0download
0
Li
P
P
Idaa
FBDS
SN
RJpe
0
©
d
1
ong-term effect of Heme oxygenase (HO)-1nduction in glomerular immune injury
RASUN K. DATTA, PU DUANN, and ELIAS A. LIANOS
HILADELPHIA, PENNSYLVANIA AND NEW BRUNSWICK, NEW JERSEY
In a rat model of macrophage-dependent glomerular immune injury induced byadministration of antibody against the glomerular basement membrane (anti-GBM), the authors assessed the anti-proteinuric effect of Heme Oxygenase-1(HO-1) induction. Rats received anti-GBM antibody alone, anti-GBM antibodyand treatment with the HO-1 inducer, hemin, or non-immune serum (controls).Urine protein, creatinine, and nitrite/nitrate excretion were measured on days 5,7, and 14 after administration of the anti-GBM antibody. In hemin-treated ani-mals with anti-GBM antibody-induced immune injury, HO-1 immunolocalized inmacrophages infiltrating glomeruli and in tubular epithelial cells. In these ani-mals, proteinuria was decreased. There was also a decrease in blood ureanitrogen (BUN) levels without a change in serum creatinine or systemic bloodpressure. The observations establish the anti-proteinuric effect of hemin induc-tion. This effect could be mechanistically linked to blunting of the ability ofinfiltrating macrophages to cause injury or to changes in tubular handling offiltered protein. (J Lab Clin Med 2006;147:150–155)
Abbreviations: ABC � avidin biotin complex; BP � blood pressure; BSA � bovine serumalbumin; BUN � blood urea nitrogen; CO � carbon monoxide; DAB � diaminobenzidinetetrahydrochloride; DTH � delayed-type hypersensitivity; GBM � glomerular basementmembrane; GFR � glomerular filtration rate; HO-1 � Heme oxygenase-1; iNOS � induciblenitric oxide synthetase; NO � nitric oxide; SD � standard deviation; Th1 � T helper cell 1;
TNF� � tumor necrosis factor-alpha; Ualb � urine albumin; Uc � urine creatinineteppaaoottasattld
n a rat model of glomerular injury induced byantibody against the GBM, the authors previouslydemonstrated that treatment with the HO-1 in-
ucer, hemin, attenuated proteinuria while it reducedctivity of the iNOS.1 These studies were performedt very early stages (24 h) of injury when upregula-
rom the Temple University Center for Neurovirology and Canceriology, Philadelphia, Pennsylvania; and Department of Medicine/ivision of Nephrology, UMDNJ-Robert Wood Johnson Medicalchool, New Brunswick, New Jersey.
ubmitted for publication September 7, 2005; revision submittedovember 11, 2005; accepted for publication November 11, 2005.
eprint requests: Elias A. Lianos, MD, PhD, UMDNJ-Robert Woodohnson Medical School, Department of Medicine/Division of Ne-hrology, P.O. Box 19-MEB 412, New Brunswick, N.J. 08903-0019;-mail: [email protected].
022-2143/$ – see front matter
2006 Mosby, Inc. All rights reserved.
toi:10.1016/j.lab.2005.11.009
50
ion of glomerular iNOS (a heme containing protein)xpression and enzyme activity occurs.1,2 It was pro-osed that the beneficial effect of HO-1 induction onroteinuria could be linked to the reduction of iNOSctivity, thereby reducing NO production. iNOS-cat-lyzed NO production can be sustained and of highutput, and NO can cause cell injury either on itswn3 or by interacting with superoxide (O2
�) to formhe potent and relatively stable oxidant peroxyni-rite.4 However, the upregulation of iNOS expressionnd activity in glomeruli after immune injury ishort-lived; although clearly detectable at 24 – 48 hfter onset of injury, it becomes barely detectablehereafter.2,5 The effect of HO-1 induction on pro-einuria when glomerular iNOS expression is noonger increased is unknown. These studies wereesigned to address this question and show that long-
erm HO-induction using treatment with the HO-1
if
M
saprarsaklptrGcwcmdsa
iiia(aPiodashtiomptcrcge
fscmiA
kl
mmcmcas
uNcdiNNmt
cmeaBmtpp
ay�
ahef(pa1ArfdrshisiA
Sd
J Lab Clin MedVolume 147, Number 3 Datta et al 151
nducer, hemin, has a sustained anti-proteinuric ef-ect.
ETHODS
Glomerular immune injury and experimental de-ign. This was induced in female Lewis rats (180–200 g) bydministration of immune serum raised in rabbits against ratarticulate GBM. To induce injury, rats were immunized withabbit IgG mixed with complete Freund’s adjuvant and givens a single dose intraperitoneally. Seven days later, animalseceived two doses (0.3 mL each) of rabbit anti-rat GBMerum given intravenously 24 h apart. This protocol results incomplement- and macrophage- dependent glomerular injurynown as accelerated anti-GBM nephritis, which histopatho-ogically resembles human rapidly progressive glomerulone-hritis. Infiltration of activated macrophages in glomeruli ishe result of a DTH reaction driven by T helper cell 1 (Th1)esponses to the disease-initiating globulin (the rabbit anti-ratBM globulin bound to the rat GBM).6 The DTH reaction
ontributes to crescent formation.6 Infiltrating macrophagesere shown to be major effectors of injury by releasing
ytokines, in particular TNF�,7 and reactive oxygen species,ainly superoxide.8 Injury is characterized by proteinuria and
ecreased GFR. The decrease in GFR occurs at the very earlytages of injury, ie, within hours of binding of the anti-GBMntibody, and recovers thereafter.9
The following groups were studied: (1) rats that weremmunized with rabbit IgG and given two doses of non-mmune rabbit serum 7 days later (controls, n � 6); (2) ratsmmunized similarly as those in group (1) but received rabbitnti-rat GBM immune serum to develop anti-GBM nephritisn � 8); and (3) rats that received rabbit anti-rat GBM serums in (2) and treated with the HO-1 inducer, hemin (Ferri-rotoporphyrin IX Chloride, Sigma) (n � 8). Hemin was
njected subcutaneously 30 �mol/kg bodyweight. A solutionf hemin (10 mM) was prepared immediately before use byissolving in 0.1N NaOH, adjusted to pH 7.4 with 0.1N HClnd further diluted with saline to the desired concentration forubcutaneous injections. Hemin injections were initiated 24ours before administration of the anti-GBM serum and con-inued on days 1, 3, and 6 thereafter to achieve sustainednduction of HO-1. On days 5, 7, and 14 after administrationf the anti-GBM serum, animals were placed in individualetabolic cages for a timed (18 h) urine collection in which,
rotein, creatinine, and nitrate (NO3�)/nitrite (NO2
�), collec-ively referred to as NOx, were measured. The metabolicages were placed on an automated refrigerated collectionack that freezes urine down to �20°C while it is beingollected (Nalgene, Braintree, Mass). This prevents bacterialrowth and provides stability of protein and NO metabolitesxcreted in the urine.
Systemic blood pressure (BP) measurements were per-ormed 24–48 hours before administration of the anti-GBMerum and on day 14 thereafter. Animals were sacrificed afterompletion of urine collection and blood pressure measure-ents on day 14. A blood sample was obtained from the
nferior vena cava for measurement of BUN and creatinine.
nimals were subsequently nephrectomized, and sections of cidney cortex were processed for immunohistochemistry toocalize HO-1 expression.
Urine protein and creatinine excretion. Total urine albu-in and creatinine were measured by established colorimetricethods using commercially available kits. (Bio-Rad, Her-
ules, Calif, and Sigma-Aldrich, St. Louis, Mo). Urine albu-in excretion was expressed as urine albumin (Ualb) to urine
reatinine (Uc) ratio. BUN was measured using an automatednalyzer or a commercially available kit as previously de-cribed.10
Urine NOx excretion. This was assessed by measuringrine nitrate (NO3
�) and nitrite (NO2�) collectively referred as
Ox. Urine samples were diluted in water, and NO3� was
onverted to NO2� using vanadium chloride. NO2
� wasetected in samples using the Griess reagent (Promega, Mad-son, Wis). Results were compared against a standard curve ofaNO3 treated in an identical manner as the urine samples.Ox were determined for each sample and expressed asicromoles per deciliter of urine. Values were factored by
hose of urine creatinine expressed in milligrams/deciliter.BP measurements. Tail systolic blood pressure was re-
orded based on the plethysmography method. The instru-ent used (Kent Scientific XBP/1000) allows BP monitoring
very 1 min. Individual animals were placed in a restrainernd allowed to become acclimated for 15 min before systolicP readings were recorded. Eight independent readings, 1in apart, were recorded for each BP recording session, and
he mean value was taken to represent the systolic BP for thatarticular session. BP measurements were performed at com-letion of the urine collection (see above).
Immunohistochemistry. Kidney cortical sections obtainedfter nephrectomies were frozen by immersing first in meth-lbutyl solvent and then in liquid nitrogen. Cryostat section (4m) were air dried at room temperature and washed with PBSnd 100% acetone. Sections were then incubated with a 3%ydrogen peroxide/methanol solution for 30 min to blockndogenous peroxide and subsequently with 1% horse serumor 15 min followed by application of Avidin/Biotin solutionVector Laboratories, Burlingame, Calif) to block nonspecificrotein binding sites. Biotin conjugated monoclonal HO-1ntibody (StressGen, Victoria, BC, Canada) was diluted:250 or 1:500 with 1% BSA/PBS and applied to the sections.fter overnight incubation, Avidin Biotin Complex (ABC)
eagent (Vector Laboratories, Burlingame, Calif) was appliedor l h at room temperature followed by 3, 3=Diaminobenzi-ine Tetrahydrochloride (DAB) solution (DAKO, Carpinte-ia, Calif) for 20 min in darkness to lend color to the reactionites. Sections were counterstained with Toluldine Blue, de-ydrated with xylene, mounted, and photographed. Specific-ty of the anti-HO-1 antibody employed was assessed inections either by omitting this antibody or by using anrrelevant monoclonal antibody followed by application of thevidin/Biotin system as described above.Statistical analyses. Results were expressed as mean �
D. Differences between groups were analyzed by the Stu-ent’s t-test for unpaired observations, P values �0.05 were
onsidered significant.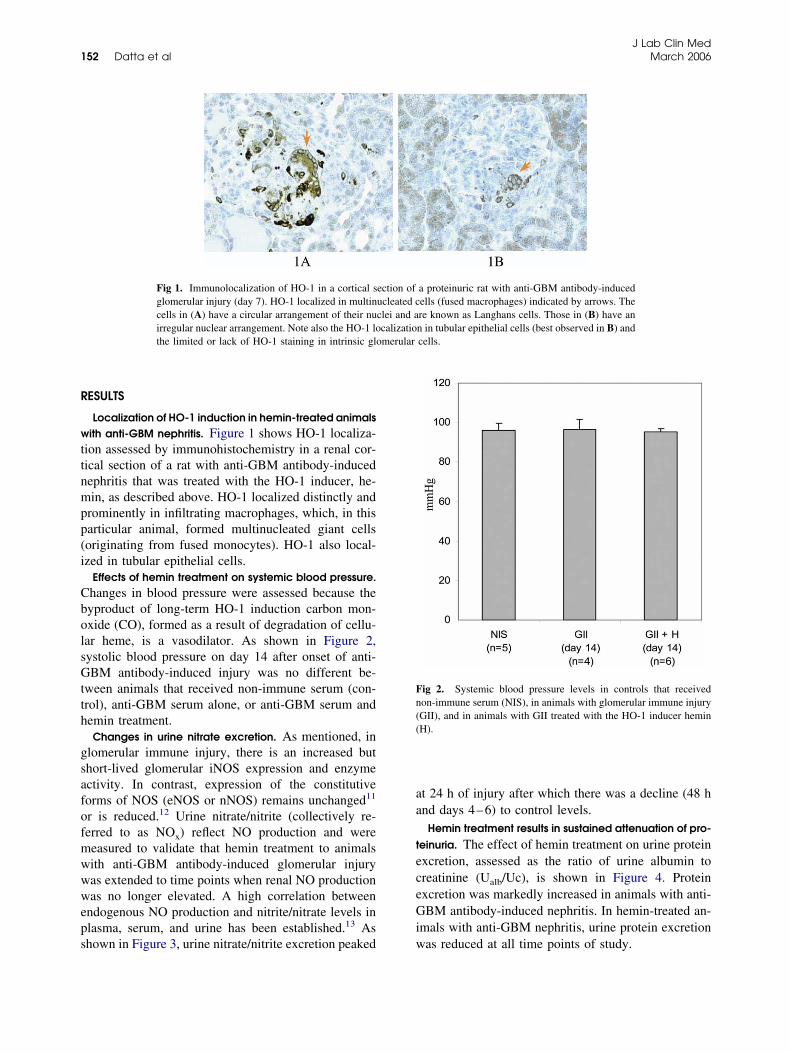
R
wttnmpp(i
CbolsGtth
gsafofmwwweps
aa
teceGi
merular
Fn((
J Lab Clin Med152 Datta et al March 2006
ESULTS
Localization of HO-1 induction in hemin-treated animalsith anti-GBM nephritis. Figure 1 shows HO-1 localiza-
ion assessed by immunohistochemistry in a renal cor-ical section of a rat with anti-GBM antibody-inducedephritis that was treated with the HO-1 inducer, he-in, as described above. HO-1 localized distinctly and
rominently in infiltrating macrophages, which, in thisarticular animal, formed multinucleated giant cellsoriginating from fused monocytes). HO-1 also local-zed in tubular epithelial cells.
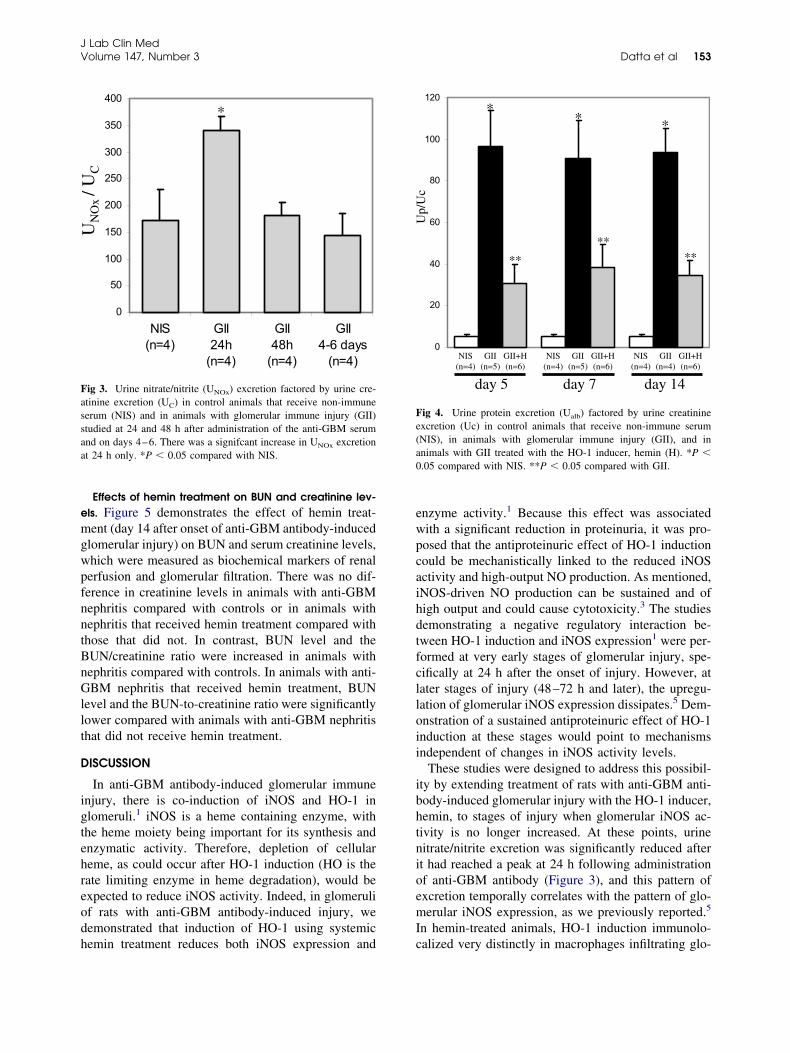
Effects of hemin treatment on systemic blood pressure.hanges in blood pressure were assessed because theyproduct of long-term HO-1 induction carbon mon-xide (CO), formed as a result of degradation of cellu-ar heme, is a vasodilator. As shown in Figure 2,ystolic blood pressure on day 14 after onset of anti-BM antibody-induced injury was no different be-
ween animals that received non-immune serum (con-rol), anti-GBM serum alone, or anti-GBM serum andemin treatment.Changes in urine nitrate excretion. As mentioned, in
lomerular immune injury, there is an increased buthort-lived glomerular iNOS expression and enzymectivity. In contrast, expression of the constitutiveorms of NOS (eNOS or nNOS) remains unchanged11
r is reduced.12 Urine nitrate/nitrite (collectively re-erred to as NOx) reflect NO production and wereeasured to validate that hemin treatment to animalsith anti-GBM antibody-induced glomerular injuryas extended to time points when renal NO productionas no longer elevated. A high correlation between
ndogenous NO production and nitrite/nitrate levels inlasma, serum, and urine has been established.13 As
Fig 1. Immunolocalization of HO-1 in a cortical sglomerular injury (day 7). HO-1 localized in multincells in (A) have a circular arrangement of their nuirregular nuclear arrangement. Note also the HO-1 lothe limited or lack of HO-1 staining in intrinsic glo
hown in Figure 3, urine nitrate/nitrite excretion peaked w
t 24 h of injury after which there was a decline (48 hnd days 4–6) to control levels.
Hemin treatment results in sustained attenuation of pro-einuria. The effect of hemin treatment on urine proteinxcretion, assessed as the ratio of urine albumin toreatinine (Ualb/Uc), is shown in Figure 4. Proteinxcretion was markedly increased in animals with anti-BM antibody-induced nephritis. In hemin-treated an-
mals with anti-GBM nephritis, urine protein excretion
a proteinuric rat with anti-GBM antibody-inducedells (fused macrophages) indicated by arrows. There known as Langhans cells. Those in (B) have ann in tubular epithelial cells (best observed in B) andcells.
ig 2. Systemic blood pressure levels in controls that receivedon-immune serum (NIS), in animals with glomerular immune injuryGII), and in animals with GII treated with the HO-1 inducer heminH).
ection ofucleated cclei and acalizatio
as reduced at all time points of study.
emgwpfnntBnGllt
D
igtehreodh
ewpcaihdtfclloii
ibhtnioemI
Fassaa
Fe(a0
J Lab Clin MedVolume 147, Number 3 Datta et al 153
Effects of hemin treatment on BUN and creatinine lev-ls. Figure 5 demonstrates the effect of hemin treat-ent (day 14 after onset of anti-GBM antibody-induced
lomerular injury) on BUN and serum creatinine levels,hich were measured as biochemical markers of renalerfusion and glomerular filtration. There was no dif-erence in creatinine levels in animals with anti-GBMephritis compared with controls or in animals withephritis that received hemin treatment compared withhose that did not. In contrast, BUN level and theUN/creatinine ratio were increased in animals withephritis compared with controls. In animals with anti-BM nephritis that received hemin treatment, BUN
evel and the BUN-to-creatinine ratio were significantlyower compared with animals with anti-GBM nephritishat did not receive hemin treatment.
ISCUSSION
In anti-GBM antibody-induced glomerular immunenjury, there is co-induction of iNOS and HO-1 inlomeruli.1 iNOS is a heme containing enzyme, withhe heme moiety being important for its synthesis andnzymatic activity. Therefore, depletion of cellulareme, as could occur after HO-1 induction (HO is theate limiting enzyme in heme degradation), would bexpected to reduce iNOS activity. Indeed, in glomerulif rats with anti-GBM antibody-induced injury, weemonstrated that induction of HO-1 using systemic
ig 3. Urine nitrate/nitrite (UNOx) excretion factored by urine cre-tinine excretion (UC) in control animals that receive non-immuneerum (NIS) and in animals with glomerular immune injury (GII)tudied at 24 and 48 h after administration of the anti-GBM serumnd on days 4–6. There was a signifcant increase in UNOx excretiont 24 h only. *P � 0.05 compared with NIS.
emin treatment reduces both iNOS expression and c
nzyme activity.1 Because this effect was associatedith a significant reduction in proteinuria, it was pro-osed that the antiproteinuric effect of HO-1 inductionould be mechanistically linked to the reduced iNOSctivity and high-output NO production. As mentioned,NOS-driven NO production can be sustained and ofigh output and could cause cytotoxicity.3 The studiesemonstrating a negative regulatory interaction be-ween HO-1 induction and iNOS expression1 were per-ormed at very early stages of glomerular injury, spe-ifically at 24 h after the onset of injury. However, atater stages of injury (48–72 h and later), the upregu-ation of glomerular iNOS expression dissipates.5 Dem-nstration of a sustained antiproteinuric effect of HO-1nduction at these stages would point to mechanismsndependent of changes in iNOS activity levels.
These studies were designed to address this possibil-ty by extending treatment of rats with anti-GBM anti-ody-induced glomerular injury with the HO-1 inducer,emin, to stages of injury when glomerular iNOS ac-ivity is no longer increased. At these points, urineitrate/nitrite excretion was significantly reduced aftert had reached a peak at 24 h following administrationf anti-GBM antibody (Figure 3), and this pattern ofxcretion temporally correlates with the pattern of glo-erular iNOS expression, as we previously reported.5
n hemin-treated animals, HO-1 induction immunolo-
ig 4. Urine protein excretion (Ualb) factored by urine creatininexcretion (Uc) in control animals that receive non-immune serumNIS), in animals with glomerular immune injury (GII), and innimals with GII treated with the HO-1 inducer, hemin (H). *P �.05 compared with NIS. **P � 0.05 compared with GII.
alized very distinctly in macrophages infiltrating glo-

mitcotHsHl
isrmstio
pit(fhipctGigk
HkistmrartigdtpbiicbtinRsttly
mflm
d with NI
J Lab Clin Med154 Datta et al March 2006
eruli and in tubules but was limited or absent inntrinsic glomerular cells (Figure 1). This reemphasizeshe observation that intrinsic glomerular cells are lessapable of upregulating HO-1 when compared withther nephron segments. In this regard, we have shownhat systemic treatment with potent and dissimilarO-1 inducers including hemin, cobalt chloride, or
tannous chloride results in a prominent upregulation ofO-1 in the nephron. In these studies, HO-1 immuno-
ocalized in tubular but not in glomerular cells.14,15
In hemin-treated animals with anti-GBM antibody-nduced injury, there was no significant changes inystemic blood pressure (Figure 2). One might expect aeduction in blood pressure on the basis of carbononoxide (CO) production (a vasodilator) after the
ustained HO-1 induction (repeated hemin dosing) inhese animals. To this end, it was reported that HO-1nduction reduces systemic blood pressure in spontane-usly hypertensive but not in normotensive rats.16
Long-term hemin treatment significantly reducedroteinuria in rats with anti-GBM antibody-inducednjury, and this effect was of similar magnitude at allime points (days 5, 7, and 14) after onset of injuryFigure 4). The mechanism of this anti-proteinuric ef-ect is unclear. In reviewing Figure 1, HO-induction inemin-treated animals was most prominent in infiltrat-ng macrophages and in tubular cells. This raises twoossibilities: (1) Induction of HO-1 in macrophagesould blunt their potential to cause injury after infiltra-ion in glomeruli. To this end, in the model of anti-BM antibody-induced injury employed in these stud-
es, infiltrating macrophages play a key role in causinglomerular injury by releasing proinflammatory cyto-
BUN
0
10
20
30
40
50
NIS(n=6)
GII(n=5)
GII+H(n=6)
0
0.2
0.4
0.6
0.8
1
N(n
mg
/dl **
*
mg
/dl
Fig 5. Changes in BUN, serum creatinine (Cr), andnon-immune serum (NIS), in animals with glomerulthe HO-1 inducer, hemin (H). *P � 0.05 compare
ines such as TNF�7 and reactive oxygen species.17 p
O-1 suppresses the release of proinflammatory cyto-ines18,19 and mediates the salutary effects of anti-nflammatory cytokines, such as suppression of TNF�ynthesis, as shown in activated macrophages exposedo IL-10.20 (2) The induction of HO-1 in tubular cells
ay alter tubular handling of filtered protein, therebyeducing protein excretion. It is now recognized thatlthough changes in urine protein excretion generallyeflect changes in glomerular permeability to protein,he tubules may significantly contribute to the process-ng (uptake and degradation) of protein filtered by thelomerular capillaries.21 The possibility that HO-1 in-uction in tubules is caused by exposure to or reabsorp-ion of filtered protein is unlikely according to studieserformed in a model of glomerular proteinuria inducedy daily injections of BSA, which is associated with anncreased tubular uptake of filtered protein. These stud-es showed no induction by HO-1 in proximal tubularells. They further showed that direct exposure of tu-ular cells to BSA did not induce HO-1.22 Based onhese studies, it is unlikely that the observed HO-1nduction in proximal tubules (Figure 1) was mecha-istically linked to the uptake of filtered protein.ather, it was most likely due to hemin treatment as
hown.14,15 To examine whether tubular HO-1 induc-ion alters tubular handling of protein, studies designedo achieve targeted induction of HO-1 in specific tubu-ar segments of the nephron are required and are be-ond the scope of these studies.In a recent publication,23 we demonstrated that in theodel of anti-GBM antibody-induced glomerular in-ammation employed in these studies, superoxide is aediator of oxidant stress and albuminuria. This, cou-
r
I5)
GII+H(n=6)
BUN/Cr
0
20
40
60
80
NIS(n=6)
GII(n=5)
GII+H(n=6)
*
**
-to-creatinine ratio in control animals that receivede injury (GII), and in animals with GII treated withS. **P � 0.05 compared with GII.
C
IS=6)
GI(n=
in BUNar immun
led with the well-established inductive effect of su-

pitmgfF
u(iwdmc(bpctatBaah
uiiipph
R
1
1
1
1
1
1
1
1
1
1
2
2
2
2
J Lab Clin MedVolume 147, Number 3 Datta et al 155
eroxide on HO-1, allows us to propose that superoxides a likely inducer of HO-1 in this model. Furthermore,he demonstration that infiltrating macrophages are aajor source of superoxide in the anti-GBM model of
lomerular in inflammation17 provides an explanationor the localization of HO-1 in these cells as shown inigure 1.In animals with anti-GBM antibody-induced glomer-
lar injury, BUN but not creatinine levels increasedFigure 5). As serum creatinine levels reflect GFR, thisndicates that, at the time point of glomerular injuryhen these studies were performed, GFR was not re-uced. This is in agreement with previous observationsade in this model of glomerular immune injury; spe-
ifically, GFR is reduced at very early stages of injurywithin hours) but recovers thereafter while proteinuriaecomes sustained.9 The increase in BUN without aarallel increase in serum creatinine indicates intravas-ular volume depletion. This can be expected in view ofhe heavy proteinuria found in animals with anti-GBMntibody-induced injury (Figure 4). Hemin treatment ofhese animals significantly reduced BUN levels and theUN-to-creatinine ratio (Figure 5). This could indicatecorrection in intravascular volume depletion brought
bout by the significant reduction in proteinuria in theemin-treated animals.In summary, these studies establish the antiprotein-
ric effect of long-term HO-1 induction using the HO-1nducer, hemin, in an infiltrative form of glomerularmmune injury. This effect was accompanied by prom-nent HO-1 expression in infiltrating activated macro-hages in which it may blunt their injury-inducingotential, and in renal tubular cells where it may alterandling of filtered protein.
EFERENCES
1. Datta PK, Koukouritaki SB, Hopp K, Lianos EA. Heme oxygen-ase-1 induction attenuates inducible nitric oxide synthase expres-sion and proteinuria in glomerulonephritis. J Am Soc Nephrol1999;10(12):2540–50.
2. Cook HT, Smith J, Salmon JA, Cattell V. Functional character-istics of macrophages in glomerulonephritis in the rat. O2- gen-eration, MHC class II expression, and eicosanoid synthesis. Am JPathol 1989;134(2):431–7.
3. Nathan C, Xie QW. Regulation of biosynthesis of nitric oxide.J Biol Chem 1994;269(19):13725–8.
4. Beckman JS, Ischiropoulos H, Zhu L, van der Woerd M, SmithC, Chen J, et al. Kinetics of superoxide dismutase- and iron-catalyzed nitration of phenolics by peroxynitrite. Arch BiochemBiophys 1992;298(2):438–45.
5. Lianos EA, Liu J. Changes in inducible nitric oxide synthaseexpression in experimental glomerulonephritis. Proc Soc Exper
Biol Med 1997;215(4):405–11.6. Huang XR, Holdsworth SR, Tipping PG. Evidence for delayed-type hypersensitivity mechanisms in glomerular crescent forma-tion. Kidney Int 1994;46(1):69–78.
7. Khan SB, Cook HT, Bhangal G, Smith J, Tam FW, Pusey CD.Antibody blockade of TNF-alpha reduces inflammation and scar-ring in experimental crescentic glomerulonephritis. Kidney Int2005;67(5):1812–20.
8. Birtwistle RJ, Michael J, Howie AJ, Adu D. Reactive oxygenproducts in heterologous anti-glomerular basement membranenephritis in rats. Br J Exp Pathol 1989;70(2):207–13.
9. Lianos EA, Andres GA, Dunn MJ. Glomerular prostaglandin andthromboxane synthesis in rat nephrotoxic serum nephritis. Ef-fects on renal hemodynamics. J Clin Invest 1983;72(4):1439–48.
0. Cohen EP, Fish BL, Moulder JE. The renin-angiotensin systemin experimental radiation nephropathy. J Lab Clin Med 2002;139(4):251–7.
1. Datta PK, Dhupar S, Lianos E. Regulatory effects of induciblenitric oxide synthase on cycloxygenase-2 and heme oxygenase -1expression in glomerulonephritis. Nephrol Dial Transplant 2006;21(1):51–7.
2. Wagner L, Riggleman A, Erdely A, Couser W, Baylis C. Re-duced nitric oxide synthase activity in rats with chronic renaldisease due to glomerulonephritis [erratum appears in Kidney Int2002;62(5):1919.]. Kidney Int 2002;62(2):532–6.
3. Granger DL, Taintor RR, Boockvar KS, Hibbs JB Jr. Measure-ment of nitrate and nitrite in biological samples using nitratereductase and Griess reaction. Meth Enzymol 1996;268:142–51.
4. da Silva JL, Zand BA, Yang LM, Sabaawy HE, Lianos E,Abraham NG. Heme oxygenase isoform-specific expression anddistribution in the rat kidney. Kidney Int 2001;59(4):1448–57.
5. Shepard M, Dhulipala P, Kabaria S, Abraham NG, Lianos EA.Heme oxygenase-1 localization in the rat nephron. Nephron2002;92(3):660–4.
6. Levere RD, Martasek P, Escalante B, Schwartzman ML, Abra-ham NG. Effect of heme arginate administration on blood pres-sure in spontaneously hypertensive rats. J Clin Invest 1990;86(1):213–9.
7. Boyce NW, Tipping PG, Holdsworth SR. Glomerular macro-phages produce reactive oxygen species in experimental glomer-ulonephritis. Kidney Int 1989;35(3):778–82.
8. Kapturczak MH, Wasserfall C, Brusko T, Campbell-ThompsonM, Ellis TM, Atkinson MA, et al. Heme oxygenase-1 modulatesearly inflammatory responses: evidence from the heme oxygen-ase-1-deficient mouse. Am J Pathol 2004;165(3):1045–53.
9. Willis D, Moore AR, Frederick R, Willoughby DA. Heme oxy-genase: a novel target for the modulation of the inflammatoryresponse. Nat Med 1996;2(1):87–90.
0. Lee TS, Chau LY. Heme oxygenase-1 mediates the anti-inflam-matory effect of interleukin-10 in mice. Nature Med 2002;8(3):240–6.
1. Russo LM, Bakris GL, Comper WD. Renal handling of albumin:a critical review of basic concepts and perspective. Am J KidneyDis 2002;39(5):899–919.
2. Pedraza-Chaverri J, Murali NS, Croatt AJ, Alam J, Grande JP,Nath KA. Proteinuria as a determinant of renal expression ofHeme Oxygenase-1 studies in models of glomerular and tubularproteinuria in the rat. Am J Physiol Renal Physiol 2005;Aug23:[Epub ahead of print].
3. Duann P, Datta PK, Pan C, Blumberg J, Sharma M, Lianos EA.Superoxide dismutase mimetic preserves the glomerular capil-lary permeability barrier to protein. J Pharm Exp Therapeut. In
press.Copyright © 2022 FDOKUMEN




















