
Social perception of male and female computer synthesized speech
Upload
integraluniversityCategory
view
0download
0
G
C
Go
MAa
b
a
ARR3AA
KGGHSL
1
stTtbeaasosr[hgno
(
0h
ARTICLE IN PRESS Model
OLSUB-6160; No. of Pages 7
Colloids and Surfaces B: Biointerfaces xxx (2013) xxx– xxx
Contents lists available at ScienceDirect
Colloids and Surfaces B: Biointerfaces
jou rn al hom epage: www.elsev ier .com/ locate /co lsur fb
lycation-assisted synthesized gold nanoparticles inhibit growthf bone cancer cells
oniba Rahima, Sana Irama, Mohd Sajid Khana,∗, M. Salman Khana, Ankur R. Shuklab,.K. Srivastavaa, Saheem Ahmada
Nanomedicine & Nanobiotechnology Lab, Department of Biosciences, Integral University, Lucknow, Uttar Pradesh 226026, IndiaDepartment of Environmental Sciences, EIILM University, 8th Mile Budang, Sikkim West, India
r t i c l e i n f o
rticle history:eceived 31 August 2013eceived in revised form0 November 2013ccepted 3 December 2013vailable online xxx
a b s t r a c t
This study presents a novel approach to synthesize glycogenic gold nanoparticles (glycogenic GNps)capped with glycated products (Schiff’s base, Heyns products, fructosylamine etc.). These glycogenic GNpshave been found to be active against human osteosarcoma cell line (Saos-2) with an IC50 of 0.187 mM,while the normal human embryonic lung cell line (L-132) remained unaffected up to 1 mM concentration.The size of glycogenic GNps can also be controlled by varying the time of incubation of gold solution.Glycation reactions involving a combination of fructose and HSA (Human Serum Albumin) were found
eywords:lycationold nanoparticleseyns productaos-2-132
to be effective in the reduction of gold to glycogenic GNps whereas glucose in combination with HSAdid not result in the reduction of gold. The progress of the reaction was followed using UV–visible spec-troscopy and NBT (Nitroblue tetrazolium) assay. The glycogenic GNps were found to be spherical in shapewith an average size of 24.3 nm, in a stable emulsion. These GNps were characterized using UV–visiblespectroscopy, zeta potential analysis, transmission electron microscopy (TEM) and scanning electronmicroscopy (SEM).
. Introduction
Non-enzymatic glycation of proteins by reducing saccharidesuch as glucose and fructose results in the formation of fruc-osamine [1] and AGEs (Advanced Glycated End products) [2,3].he implication of glycation in a number of diseases has increasedhe focus on glycation since the last decade. Glycation alters theiological activities of proteins, leading to protein dysfunction. Theffects of glycation, in particular, are more pronounced in severalge-related diseases such as diabetes [4], cancer [5], cataracts [6],therosclerosis [7], Alzheimer’s [8], renal failure [9], etc. Thus, thetudy of glycation has become one of the most important areasf biomedical research. Glycation by glucose, among the reducingugars, has been extensively studied. In addition to glucose, othereducing sugars such as galactose [10], sialic acid [11], mannose12], glucose-6-phosphate [13], glyceraldehydes, and fucose [12]ave been used as glycating agents in vitro. The phenomenon of
Please cite this article in press as: M. Rahim, et al., Glycation-assisted synthSurf. B: Biointerfaces (2013), http://dx.doi.org/10.1016/j.colsurfb.2013.12.0
lucose glycation is a slow process, although it is associated withumerous cellular metabolic processes [14]. There is a great dealf information available on glycation by glucose, on the contrary,
∗ Corresponding author. Tel.: +91 522 2890812/2890730; fax: +91 522 2890809.E-mail addresses: sajid [email protected], [email protected]
M.S. Khan).
927-7765/$ – see front matter © 2013 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.colsurfb.2013.12.008
© 2013 Elsevier B.V. All rights reserved.
only a handful of studies have investigated the role of fructose inglycation. In vitro studies suggest that fructose is a more potentinitiator of Maillard reaction than glucose [15]. Elevated levels offructose in blood can lead to glycation of biomolecules, affectingtheir structure and function, through the formation of AGEs [16].Although the level of fructose in human plasma is relatively less ascompared to that of glucose, due to the increasing use of fructoseas a dietary sweetener and its highly reactive nature, the study ofthe effect of fructose mediated glycation gains significance.
Human serum albumin (HSA), being prone to glycation, has arelatively long half life (about 21 days) in comparison to other pro-teins [17]. In addition, proteins such as collagen [18], lens crystallins[19], red cell membrane [20], peripheral nerve proteins [21], ferritin[12], Apolipoprotein, hemoglobin [22], insulin, immunoglobins,laminin and fibronectin [23] are also susceptible to glycation andhave been widely studied. Even though, human serum albumin isthe major serum transport protein, very little information is avail-able on the effect of glycation of this protein [24]. The glycationreaction, also known as Maillard reaction, is a slow, non-enzymaticreaction involving the attachment of carbonyl group of reducingsugars (Glucose, fructose, etc.) to free amino groups of Human
esized gold nanoparticles inhibit growth of bone cancer cells, Colloids08
serum albumin to form glycated HSA (Schiff’s base), followed byamadori rearrangement to form more stable aminomethyl ketone[1,25]. This is the early glycation reaction wherein, Schiff’s baseand fructosamine are the early glycation adducts [26]. Subsequent

ING Model
C
2 ces B:
mrrcrftawpRos
gtc
2
2
t[eog
2
ifps
sdis
2
bsoa2t
2
dAfamimn1
ARTICLEOLSUB-6160; No. of Pages 7
M. Rahim et al. / Colloids and Surfa
odifications in early glycated end products such asearrangement, oxidation, polymerization and cleavage, giveise to irreversible advanced glycated end products [27]. In thease of fructose, the rearrangement of Schiff base is known as Heynsearrangement. The study of AGEs is important, since the loss ofunction of proteins and tissue damage in vivo, is directly relatedo the formation of AGEs. Early glycation reactions themselves canlter the function of a number of proteins. There are certain reportshich establish the role of glycated products against cancer withrobable mechanism of causing apoptosis, through activation ofOS, MAP kinases, the FOXO1 transcription factor [28], inhibitionf formation of nitrosamine, direct scavenging of carcinogenicpecies and performing anti-promoter activity [29].
This study focuses on the synthesis of gold nanoparticles (glyco-enic GNps) using fructose and HSA, as the glycating agents andhe utility of these nanoparticles as anticancer agent against boneancer.
. Materials and methods
.1. Chemicals and reagents
d-glucose, d-fructose, human serum albumin, NBT (Nitroblueetrazolium) reagent were purchased from Merck. TetrachloroauricIII] acid (HAuCl4) and HEPES (4-(2-hydroxyethyl) piperazine-1-thanesulfonic acid) were purchased from Sigma–Aldrich. Unlesstherwise indicated, all solvents and chemicals were of analyticalrade and were used as received.
.2. Glycation assay
Glucose glycated or fructose glycated HSAs were prepared byncubating HSA (150 �g, 300 �g, 450 �g) with 80 mM of glucose orructose (80 mM, 150 mM, 250 mM) for 33 days at 37 ◦C in 10 mMhosphate buffered saline (pH 7.4) in 3 ml reaction volume, underterile conditions.
All the solutions were filter sterilized using PuradiscTM 0.2 �myringe filter (Whatman, GE Healthcare UK limited, UK) and wereispensed into culture tubes, under aseptic conditions. HSA under
dentical conditions except for the addition of reducing sugars,erved as the control.
.3. Absorbance spectroscopy
The absorption spectra of native and glycated samples incu-ated for 33 days were recorded on Shimadzu dual-beampectrophotometer (model UV-1601 PC) in the wavelength rangef 220–800 nm in quartz cuvette of 1 cm path length. Individualliquots of the reaction mixtures were analyzed for absorbance at79 nm. Absorption spectra in the range of 250–700 nm were foundo be optimal for detecting AGEs.
.4. Analysis of glycated albumin
The keto-amine moieties resulting from glycation of HSA wereetermined by a modified form of NBT reduction assay [30].bsorbance spectra were measured each day by removing aliquots
rom the reaction mixture to determine the presence of keto-
Please cite this article in press as: M. Rahim, et al., Glycation-assisted synthSurf. B: Biointerfaces (2013), http://dx.doi.org/10.1016/j.colsurfb.2013.12.0
mine moieties. 20 �l of native and glycated HSA samples wereixed with 200 �l of 100 mM carbonate buffer (pH 10.8) contain-
ng 0.25 mM NBT and incubated for 10 min at 37 ◦C and absorbanceeasured at 525 nm. The presence of keto-amine moieties in
mol/ml was determined based on an extinction coefficient of2,640 M−1 cm−1 for monoformazan [31].
PRESS Biointerfaces xxx (2013) xxx– xxx
2.5. Glycation-assisted synthesis of gold nanoparticles(glycogenic GNps)
For the in vitro synthesis of gold nanoparticles (glycogenicGNPs), 3 ml of reaction mixture containing glycated HSA (preparedby incubating 300 �g human serum albumin with 250 mM fruc-tose for one, two and three days, respectively, in three separatereactions) and 1.0 mM freshly prepared H[AuCl4] was incubatedat 37 ◦C for 24 h. Reactions performed with native HSA and goldsolution as well as with native fructose and gold solution, wereused as control. The synthesis of nanoparticles was followed bymeasuring the absorbance spectra of the reaction mixture at reg-ular intervals. On completion of the reaction, gold nanoparticleswere collected by centrifugation (30,000 × g, 30 min) and washedtwice with Milli-Q water. The unbound proteins were removedby extracting with 50% (v/v) of ethanol. Glycogenic GNPs wereseparated from unused HSA and fructose by passing the reactionmixture through Biogel P-30 gel filtration column pre-equilibratedwith 20 mM HEPES buffer (pH 6.0) containing 150 mM NaCl. Thefractions were scanned between 220 nm and 800 nm and the frac-tions that exhibited absorbance at 520 nm were pooled together.The pooled samples were dialyzed against distilled water and usedfor further characterization.
2.6. Characterization of gold nanoparticles
The glycogenic GNPs were characterized by absorbance spec-troscopy, zeta potential analysis, TEM and SEM. Absorbancemeasurements were performed on a Shimadzu dual beam spec-trophotometer (model UV-1601 PC) operated at a resolution of1 nm. Zeta potential was measured on Zetasizer Nano-ZS, ModelZEN3600 (Malvern Instrument Ltd, Malvern, UK). Scanning electronmicroscopy was performed by drying a drop of suspension of glyco-genic GNPs on glass slides and coated with gold. The morphologyof nanoparticles was examined under a scanning electron micro-scope (JEOL JSM 5200). Transmission electron microscopy (TEM)was performed on TecnaiTM G2 Spirit BioTWIN, FEI Company, bydrop coating gold nanoparticles suspension on carbon coated cop-per grids.
2.7. In vitro anticancer studies of glycogenic GNps
2.7.1. Cell cultureHuman osteosarcoma cell line (Saos-2) and normal human
embryonic lung cell line (L-132) were obtained from National Cen-tre for Cell Science (NCCS), Pune, India. Saos-2 and L-132 cells weregrown as monolayer in Mac Coy’s and EMEM medium, respectively,supplemented with 10% fetal bovine serum and 1% antibiotic con-taining 10,000 units of penicillin, 10 mg streptomycin and 25 �gamphotericin B, in a humidified atmosphere in a 5% CO2 incubatorat 37 ◦C. Stocks were maintained in 75 cm2 tissue culture flasks.
2.7.2. Cell viability assaySaos-2 and L-132 cells were plated in 96-well plates at a
density of 1 × 104 cells per well and incubated for 24 h in a humid-ified 5% CO2 incubator at 37 ◦C. After 24 h, the cells were treatedwith glycogenic GNps at concentrations of 1, 0.5, 0.25, 0.125 and0.062 mM, in triplicates, and incubated for 48 h. After incubation,media was discarded and 50 �l MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide] (5 mg/ml in PBS), was added toeach well and incubated for 4 h in 5% CO2 incubator. The result-ing formazan crystals were solubilized in 150 �l DMSO (Dimethyl
esized gold nanoparticles inhibit growth of bone cancer cells, Colloids08
sulfoxide). The reduced MTT was quantified by measuring theoptical densities at a wavelength of 570 nm with reference filterof 655 nm using an ELISA reader (Microplate Reader (BIORAD-680)). Percentage inhibition of the cells was calculated using the

ARTICLE IN PRESSG Model
COLSUB-6160; No. of Pages 7
M. Rahim et al. / Colloids and Surfaces B: Biointerfaces xxx (2013) xxx– xxx 3
Table 1Showing rate of glycation determined by using UV absorbance at 279 nm, NBT assay at 525 nm and amount of ketoamine moieties at third day in different combinations ofconc. of HSA with fructose and glucose.
S. no. HSA Conc.(�g/3 ml)
Fructose Glucose
Conc. (mM) A525 Fructosamine(nmol/ml)
A279 Conc. (mM) A525 Glucosamine(nmol/ml)
A279
1150
80 1.856 293.67 0.36880 0.41 32.437 1.3372 150 1.461 231.171 0.328
3 250 1.653 261.551 0.432
4300
80 0.895 141.614 0.34780 0.411 32.516 1.4865 150 1.333 210.918 0.997
6 250 1.802 285.126 0.965
7450
80 1.044 165.189 1.00580 0.458 36.234 1.1178 150 1.356 214.557 0.989
9 250 1.608 254.43 1.212
fipoT6
2L
c5wE
2
dwwSbwrfc
FS
ormula X = 100 − (Atest − Ablank)/(Acontrol − Ablank) × 100; where, Xs percentage inhibition, Atest is the absorbance of the test sam-le, Ablank is the absorbance of blank and Acontrol is the absorbancef control sample. Doxorubicin served as the positive control.he IC50 value was calculated by fitting the data using ORIGIN.1.
.7.3. Measurement of cytomorphological changes in Saos-2 and-132
Saos-2 and L-132 cells were pre-treated with different con-entrations of glycogenic GNps and incubated for 48 h at 37 ◦C in% CO2 atmosphere. The gross morphological changes in the cellsere observed under an inverted phase contrast microscope (Nikon
CLIPSE Ti-S, Nikon Corporation, Tokyo Japan).
.7.4. Apoptosis assayThe apoptotic effect of glycogenic GNps on Saos-2 cells was
etermined using fluorescent nuclear dye, DAPI [32]. The cellsere seeded and treated as mentioned above. And the cells wereashed with PBS and fixed in 4% paraformaldehyde for 10 min.
ubsequently, the cells were permeabilized with permeabilizinguffer (3% paraformaldehyde and 0.5% Triton X-100) and stained
Please cite this article in press as: M. Rahim, et al., Glycation-assisted synthSurf. B: Biointerfaces (2013), http://dx.doi.org/10.1016/j.colsurfb.2013.12.0
ith DAPI dye. The stained cells were imaged using a fluo-escent microscope (Nikon ECLIPSE Ti-S, Japan). The cells withragmented and condensed nuclei were identified as apoptoticells.
ig. 1. Schematic representation of glycation assisted synthesis of gold nanoparticles (glyaos-2 bone cancer cells.
3. Results
3.1. Comparative study of the rate of glycation of glucose andfructose
When HSA was incubated independently with glucose and fruc-tose for 33 days at 37 ◦C and pH 7.4, in combinations of differentconcentrations of respective glycating agents (Table 1), the rates ofglycation were found to be different for similar reaction at differenttimes of reaction. Similarly, a difference in the rates of reactions wasalso observed for both the reactions. The variation in the rate of gly-cation was confirmed through UV–visible spectroscopy at 279 nm(Table 1 and Supplementary Fig. 1) and NBT assay by measuringthe absorbance at 525 nm (Table 1 and Supplementary Fig. 2). Thehighest rate of glycation was observed with fructose (300 �g/3 ml)and HSA (250 mM) on the third day (Fig. 1).
3.2. Synthesis of glycogenic GNps
Gold solution (1 mM AuHCl4) was incubated overnight at 37 ◦Cat the 1st, 2nd, 3rd and 4th day of glycation reaction, when the rateof glycation was found to be higher in comparison to other days
esized gold nanoparticles inhibit growth of bone cancer cells, Colloids08
of reactions. The formation of glycogenic GNps were identified bytheir characteristic ruby red color which was further confirmed bytheir representative surface plasmon resonance (SPR) band cen-tered at 529 nm, 526 nm, 520 nm and 534 nm. When the diameter
cogenic GNps) encapsulated with glycated Human Serum Albumin (HSA) to combat
ARTICLE IN PRESSG Model
COLSUB-6160; No. of Pages 7
4 M. Rahim et al. / Colloids and Surfaces B: Biointerfaces xxx (2013) xxx– xxx
ta pot
obadapsaU3
3
aspmsr
3
3
ehlGtSicc
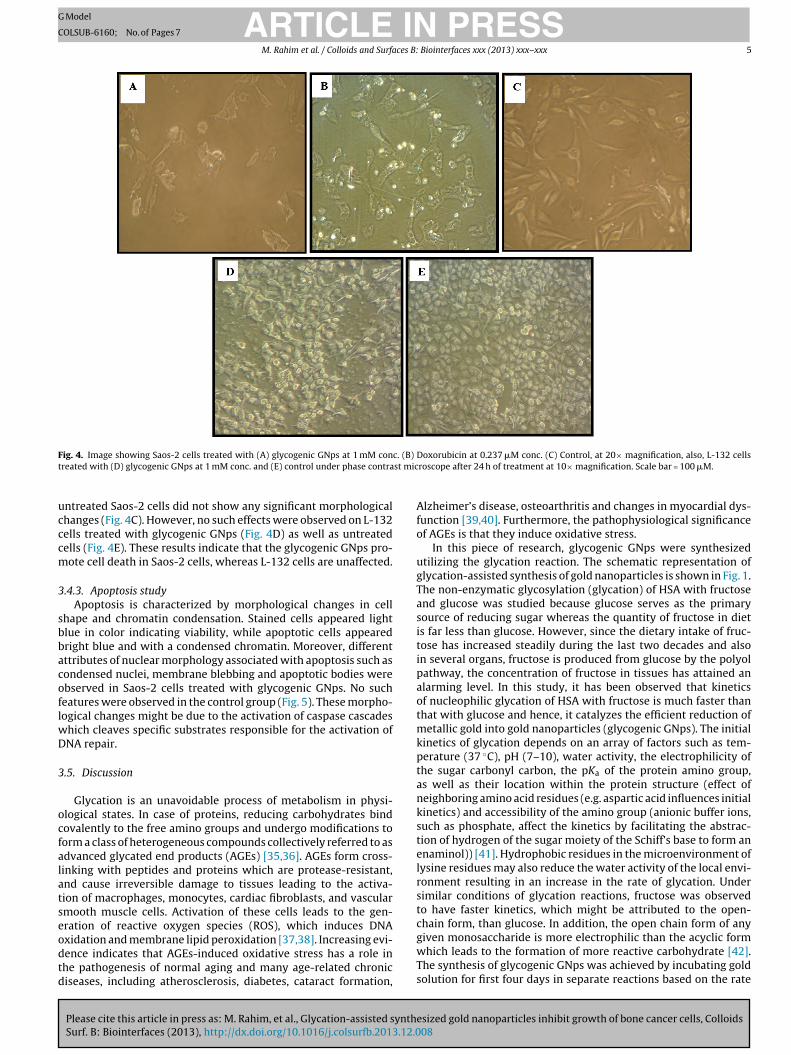
Remarkable morphological variations such as loss of membraneintegrity, inhibition of cell growth, necrosis, cytoplasmic conden-sation and cell clumping were observed in Saos-2 cells treatedwith glycogenic GNps (Fig. 4A) and Doxorubicin (Fig. 4B) whereas
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0 1.10
10
20
30
40
50
60
70
80
90
100
Perc
enta
ge In
hibi
tion
(%)
Conc entr ation ( mM)
L-13 2 cells trea ted with gl ycogenic GNpsSaos -2 cel ls trea ted with gl ycoge nic GNps (IC
50=0.18 7mM)
Fig. 3. The cytotoxicity (dose dependent) study of glycogenic GNps on Saos-2
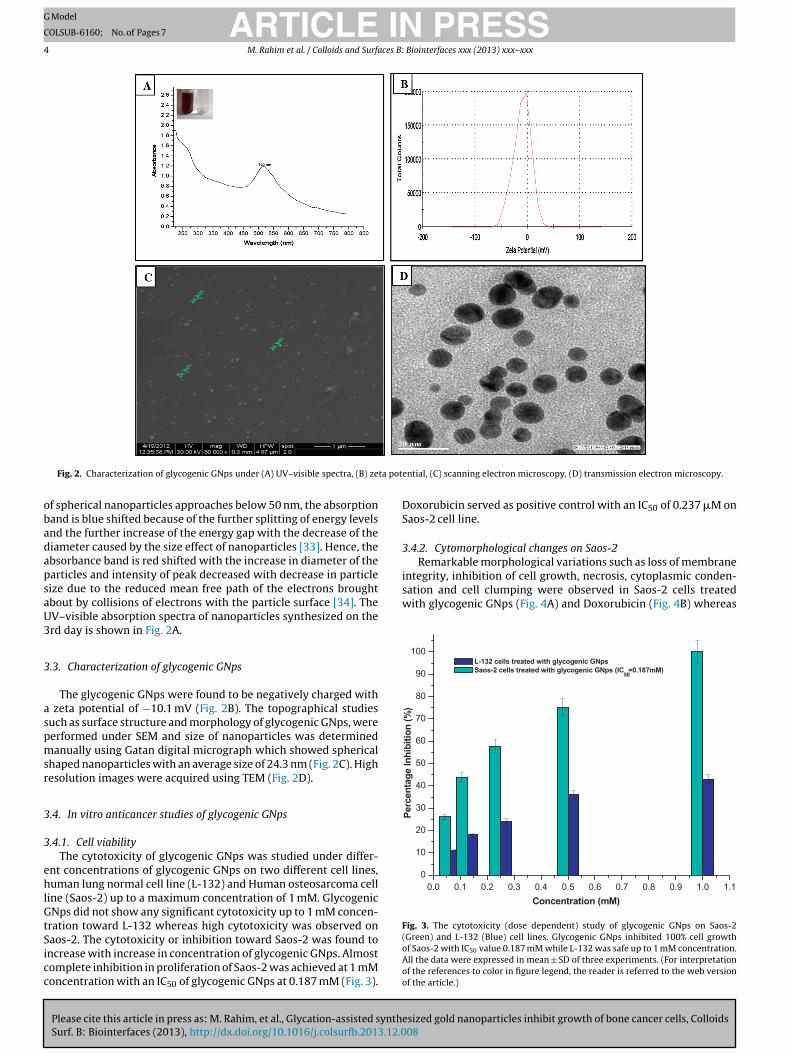
Fig. 2. Characterization of glycogenic GNps under (A) UV–visible spectra, (B) ze
f spherical nanoparticles approaches below 50 nm, the absorptionand is blue shifted because of the further splitting of energy levelsnd the further increase of the energy gap with the decrease of theiameter caused by the size effect of nanoparticles [33]. Hence, thebsorbance band is red shifted with the increase in diameter of thearticles and intensity of peak decreased with decrease in particleize due to the reduced mean free path of the electrons broughtbout by collisions of electrons with the particle surface [34]. TheV–visible absorption spectra of nanoparticles synthesized on therd day is shown in Fig. 2A.
.3. Characterization of glycogenic GNps
The glycogenic GNps were found to be negatively charged with zeta potential of −10.1 mV (Fig. 2B). The topographical studiesuch as surface structure and morphology of glycogenic GNps, wereerformed under SEM and size of nanoparticles was determinedanually using Gatan digital micrograph which showed spherical
haped nanoparticles with an average size of 24.3 nm (Fig. 2C). Highesolution images were acquired using TEM (Fig. 2D).
.4. In vitro anticancer studies of glycogenic GNps
.4.1. Cell viabilityThe cytotoxicity of glycogenic GNps was studied under differ-
nt concentrations of glycogenic GNps on two different cell lines,uman lung normal cell line (L-132) and Human osteosarcoma cell
ine (Saos-2) up to a maximum concentration of 1 mM. GlycogenicNps did not show any significant cytotoxicity up to 1 mM concen-
ration toward L-132 whereas high cytotoxicity was observed on
Please cite this article in press as: M. Rahim, et al., Glycation-assisted synthSurf. B: Biointerfaces (2013), http://dx.doi.org/10.1016/j.colsurfb.2013.12.0
aos-2. The cytotoxicity or inhibition toward Saos-2 was found toncrease with increase in concentration of glycogenic GNps. Almostomplete inhibition in proliferation of Saos-2 was achieved at 1 mMoncentration with an IC50 of glycogenic GNps at 0.187 mM (Fig. 3).
ential, (C) scanning electron microscopy, (D) transmission electron microscopy.
Doxorubicin served as positive control with an IC50 of 0.237 �M onSaos-2 cell line.
3.4.2. Cytomorphological changes on Saos-2
esized gold nanoparticles inhibit growth of bone cancer cells, Colloids08
(Green) and L-132 (Blue) cell lines. Glycogenic GNps inhibited 100% cell growthof Saos-2 with IC50 value 0.187 mM while L-132 was safe up to 1 mM concentration.All the data were expressed in mean ± SD of three experiments. (For interpretationof the references to color in figure legend, the reader is referred to the web versionof the article.)
ARTICLE IN PRESSG Model
COLSUB-6160; No. of Pages 7
M. Rahim et al. / Colloids and Surfaces B: Biointerfaces xxx (2013) xxx– xxx 5
F c. (B)
t st mic
ucccm
3
sbbacoflwD
3
ocfalatseodtd
ig. 4. Image showing Saos-2 cells treated with (A) glycogenic GNps at 1 mM conreated with (D) glycogenic GNps at 1 mM conc. and (E) control under phase contra
ntreated Saos-2 cells did not show any significant morphologicalhanges (Fig. 4C). However, no such effects were observed on L-132ells treated with glycogenic GNps (Fig. 4D) as well as untreatedells (Fig. 4E). These results indicate that the glycogenic GNps pro-ote cell death in Saos-2 cells, whereas L-132 cells are unaffected.
.4.3. Apoptosis studyApoptosis is characterized by morphological changes in cell
hape and chromatin condensation. Stained cells appeared lightlue in color indicating viability, while apoptotic cells appearedright blue and with a condensed chromatin. Moreover, differentttributes of nuclear morphology associated with apoptosis such asondensed nuclei, membrane blebbing and apoptotic bodies werebserved in Saos-2 cells treated with glycogenic GNps. No sucheatures were observed in the control group (Fig. 5). These morpho-ogical changes might be due to the activation of caspase cascades
hich cleaves specific substrates responsible for the activation ofNA repair.
.5. Discussion
Glycation is an unavoidable process of metabolism in physi-logical states. In case of proteins, reducing carbohydrates bindovalently to the free amino groups and undergo modifications toorm a class of heterogeneous compounds collectively referred to asdvanced glycated end products (AGEs) [35,36]. AGEs form cross-inking with peptides and proteins which are protease-resistant,nd cause irreversible damage to tissues leading to the activa-ion of macrophages, monocytes, cardiac fibroblasts, and vascularmooth muscle cells. Activation of these cells leads to the gen-ration of reactive oxygen species (ROS), which induces DNA
Please cite this article in press as: M. Rahim, et al., Glycation-assisted synthSurf. B: Biointerfaces (2013), http://dx.doi.org/10.1016/j.colsurfb.2013.12.0
xidation and membrane lipid peroxidation [37,38]. Increasing evi-ence indicates that AGEs-induced oxidative stress has a role inhe pathogenesis of normal aging and many age-related chroniciseases, including atherosclerosis, diabetes, cataract formation,
Doxorubicin at 0.237 �M conc. (C) Control, at 20× magnification, also, L-132 cellsroscope after 24 h of treatment at 10× magnification. Scale bar = 100 �M.
Alzheimer’s disease, osteoarthritis and changes in myocardial dys-function [39,40]. Furthermore, the pathophysiological significanceof AGEs is that they induce oxidative stress.
In this piece of research, glycogenic GNps were synthesizedutilizing the glycation reaction. The schematic representation ofglycation-assisted synthesis of gold nanoparticles is shown in Fig. 1.The non-enzymatic glycosylation (glycation) of HSA with fructoseand glucose was studied because glucose serves as the primarysource of reducing sugar whereas the quantity of fructose in dietis far less than glucose. However, since the dietary intake of fruc-tose has increased steadily during the last two decades and alsoin several organs, fructose is produced from glucose by the polyolpathway, the concentration of fructose in tissues has attained analarming level. In this study, it has been observed that kineticsof nucleophilic glycation of HSA with fructose is much faster thanthat with glucose and hence, it catalyzes the efficient reduction ofmetallic gold into gold nanoparticles (glycogenic GNps). The initialkinetics of glycation depends on an array of factors such as tem-perature (37 ◦C), pH (7–10), water activity, the electrophilicity ofthe sugar carbonyl carbon, the pKa of the protein amino group,as well as their location within the protein structure (effect ofneighboring amino acid residues (e.g. aspartic acid influences initialkinetics) and accessibility of the amino group (anionic buffer ions,such as phosphate, affect the kinetics by facilitating the abstrac-tion of hydrogen of the sugar moiety of the Schiff’s base to form anenaminol)) [41]. Hydrophobic residues in the microenvironment oflysine residues may also reduce the water activity of the local envi-ronment resulting in an increase in the rate of glycation. Undersimilar conditions of glycation reactions, fructose was observedto have faster kinetics, which might be attributed to the open-chain form, than glucose. In addition, the open chain form of any
esized gold nanoparticles inhibit growth of bone cancer cells, Colloids08
given monosaccharide is more electrophilic than the acyclic formwhich leads to the formation of more reactive carbohydrate [42].The synthesis of glycogenic GNps was achieved by incubating goldsolution for first four days in separate reactions based on the rate

ARTICLE IN PRESSG Model
COLSUB-6160; No. of Pages 7
6 M. Rahim et al. / Colloids and Surfaces B: Biointerfaces xxx (2013) xxx– xxx
reated
o(Gs(oa−sntsoagssat([twgwfhbmotceagaeirtcrnn
Fig. 5. DAPI staining for cellular apoptosis on Saos-2 cells (A) Control (B) t
f kinetics calculated using NBT assay and UV–visible spectroscopyTable 1 and Supplementary Figs. 2 and 3). Although synthesis ofNps took place for all four days within 8–10 h, variations in theizes of the GNps were observed for different periods of reactionData not shown). The glycogenic GNps obtained on the third dayf reaction were smallest in size and hence, chosen for further char-cterization. The zeta potential of glycogenic GNps was found to be10.1 mV which might be one of the reasons for their long term
tability (Fig. 2B); the electrostatic repulsive forces between theanoparticles might prevent them from getting closer to each otherhereby preventing their agglomeration or clumping in aqueoususpension. SEM and TEM analyses confirm the spherical shapef glycogenic GNps with an average diameter of 24.3 nm (Fig. 2Cnd D). This method was also adopted to monitor the growth oflycogenic GNps and regulate the particle size by incubating goldolution at different days of glycation. However, glycogenic GNpsynthesized on the 3rd day were found to be best in terms of sizend stability (Data not shown). The glycation reaction started fromhe very first day and attained the maximum rate on the third dayTable 1) with the amount of glycated protein reaching up to 40%43]. It is well documented that glycation of HSA is able to generatehermodynamically more stable high molecular weight aggregatesith a high �-sheet structure content as compared to its non-
lycated form [44]. On the contrary, short-term incubation (7 days)ith reducing sugars induces HSA to undergo rapid misfolding to
orm globular amyloid-like aggregations without any change in �-elix/�-sheet proportions [45]. Therefore, it is argued that Schiff’sase, fructosylamine and Heyns products with nonglycated HSAight have been present in the reaction mixture after three days
f glycation. The glycated HSA being more hydrophilic in naturehan nonglycated HSA, it might serve, as an ideal candidate, as aapping agent [46]. There are very few reports which suggest thatarly glycation products possess anticarcinogenic properties [30]nd promote AGE-induced apoptosis [29]. Based on these findings,lycogenic GNps capped with early glycation products were testedgainst human osteosarcoma cell line, Saos-2 and normal humanmbryonic lung cell line, L-132. These nanoparticles were found tonhibit the growth of cancerous cells (Fig. 4A) while normal cellsemained unaffected (Fig. 4D). The glycogenic GNPs were foundo induce nuclear condensation (Fig. 5) and eventually, caused
Please cite this article in press as: M. Rahim, et al., Glycation-assisted synthSurf. B: Biointerfaces (2013), http://dx.doi.org/10.1016/j.colsurfb.2013.12.0
ell death via apoptosis in Saos-2 cells while normal cells (L-132)emained unaffected (Fig. 5). It has been reported that colloidal goldanoparticles within the size range of 3–100 nm do not show sig-ificant toxicity, provided that the threshold dose does not exceed
with 0.25 mM glycogenic GNps at 20× magnification. Scale bar = 100 �M.
a value of the order of 1012 particles/ml [47]. The most efficientcellular uptake of gold nanoparticles was observed with particlesranging from 20 to 50 nm. Charge, size and surface functionalitycontribute to determine the cellular uptake of functionalized goldnanoparticles. Smaller gold nanoparticles rapidly diffuse into thetumor matrix, whereas larger particles are retained near the vas-culature. It has been reported that cytotoxic gold nanoparticleswith structured ligand shells could directly pass through the plasmamembrane of cells without the creation of pores [48]. Hence, theglycogenic GNps under study might be inhibiting bone tumor cellsfrom proliferation while sparing normal cells.
4. Conclusion
This study establishes a novel method to synthesize glycogenicGNps with encapsulated glycated products having an anticancerproperty, specifically, against bone cancer cells while leaving thenormal cells unaffected. It still remains difficult to modulate thepreviously glycated products against cancer in spite of the avail-ability of reports on their efficacy. Using this method, previouslyglycated products were immobilized over gold nanoparticles toform a stable suspension with remarkable characteristics. Thisstudy also corroborates the fact that glycation of HSA with fructoseoccurs at a much higher rate than with glucose. The work requiresan-in-depth study to establish these nanoparticles as a potent drugmolecule.
Acknowledgements
The authors MR and SI thank University Grant commission(UGC), New Delhi, for Maulana Azad National Fellowship. Theauthors also acknowledge Integral University and Department ofBiosciences for providing instrumentation facilities and support forthe research.
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.colsurfb.2013.12.008.
esized gold nanoparticles inhibit growth of bone cancer cells, Colloids08
References
[1] J.F. Day, S.R. Thorpe, J.W. Baynes, J. Biol. Chem. 254 (3) (1979) 595–597.

ING Model
C
ces B:
[[[[[
[[
[[[
[[
[
[
[[
[
[
[
[
[[
[
[[
[[
[
[[
[[
[[
[
[
ARTICLEOLSUB-6160; No. of Pages 7
M. Rahim et al. / Colloids and Surfa
[2] J.T.P., Clin. Lab. 45 (1999) 263–273.[3] M.I. Ahmad, S. Ahmad, Moinuddin, Ind. J. Biochem. Biophys. 48 (2011) 290–296.[4] D.R. McCance, D.G. Dyer, J.A. Dunn, K.E. Bailie, S.R. Thorpe, J.W. Baynes, T.J.
Lyons, J. Clin. Invest. 91 (6) (1993) 2470–2478.[5] H. Stopper, R. Schinzel, K. Sebekova, A. Heidland, Cancer Lett. 190 (2003)
151–156.[6] T.J. Lyons, G. Silvestri, J.A. Dunn, D.G. Dyer, J.W. Baynes, Diabetes 40 (8) (1991)
1010–1015.[7] L.G. Bucciarelli, T. Wendt, W. Qu, et al., Circulation 106 (2002) 2827–2835.[8] T. Miyata, C. van Ypersele de Strihou, K. Kurokawa, J.W. Baynes, Kidney Int. 55
(2) (1999) 389–399.[9] M.E. Westwood, P.J. Thornalley, J. Protein Chem. 14 (5) (1995) 359–372.10] J.C. Urbanowski, M.A. Cohenford, J.A. Dain, J. Biol. Chem. 257 (1982) 111–115.11] R.A. McKinney, J.C. Urbanowski, J.A. Dain, Biochem. Znt. 4 (1982) 127–133.12] Z. Zaman, R.L. Verwilghen, Biochem. Biophys. Acta 669 (1981) 120–124.13] D.N. Haney, H.F. Bunn, Proc. Natl. Acad. Sci. U. S. A. 73 (1976) 3534–3538.14] A. Stolzing, R. Widmer, T. Jung, P. Voss, T. Grune, Free Radic. Biol. Med. 40 (6)
(2006) 1017–1027.15] H.F. Bunn, P.L. Higgins, Science (Washington, DC) 213 (1981) 222–224.16] I. Mustafa, S. Ahmad, K. Dixit, Moinuddin, J. Ahmad, A. Ali, Diabetes Res. Clin.
Pract. 95 (2012) 98–104.17] R. Dolhofer, O.H. Wieland, FEBS Lett. 103 (1979) 282–286.18] A. Le Pape, J.P. Muh, A.J. Bailey, Biochem. J. 197 (1981) 405–412.19] V.J. Stevens, C.A. Rouzer, V.M. Monnier, A. Cerami, Proc. Natl. Acad. Sci. U. S. A.
75 (1978) 2918–2922.20] J.A. Miller, E. Gravallese, H.F. Bunn, J. Clin. Invest. 65 (1980) 896–901.21] H. Vlassara, M. Brownlee, A. Cerami, Proc. Natl. Acad. Sci. U. S. A. 78 (1981)
5190–5192.22] A. Biroccio, A. Urbani, R. Massoud, C. di Ilio, P. Sacchetta, S. Bernardini, C. Cortese,
G. Federici, Anal. Biochem. 336 (2005) 279–288.23] A.S. Charonis, L.A. Reger, J.E. Dege, K. Kouzi-Koliakos, L.T. Furcht, R.M.
Please cite this article in press as: M. Rahim, et al., Glycation-assisted synthSurf. B: Biointerfaces (2013), http://dx.doi.org/10.1016/j.colsurfb.2013.12.0
Wohlhueter, E.C. Tsilibary, Diabetes 39 (1990) 807–814.24] B.S. Lewis, J.J. Harding, Exp. Eye Res. 50 (1990) 463–467.25] J.W. Hartog, A.A. Voors, S.J. Bakker, A.J. Smit, D.J. van Veldhuisen, Eur. J. Heart
Fail. 9 (2007) 1146–1155.26] P.J. Thornalley, Clin. Lab. 5–6 (1999) 263–273.
[
[[
PRESS Biointerfaces xxx (2013) xxx– xxx 7
27] M.P. Cohen, E. Shea, S. Chen, C.W. Shearman, J. Lab. Clin. Med. 141 (2003)242–249.
28] M. Alikhani, C.M. Maclellan, M. Raptis, S. Vora, P.C. Trackman, D.T. Graves, Am.J. Physiol. Cell Physiol. 292 (2007) C850–C856.
29] H.U. Aeschbacher, P.A. Finot, H. Aeschbacher, R.F. Hurrel, R. Liardon (Eds.),Birkhauser Verlag, Basel (1990) 335–348.
30] R.N. Johnson, P.A. Metcalf, J.R. Baker, Clin. Chim. Acta 127 (1982) 87–95.31] R. Mironova, T. Niwa, Y. Handzhiyski, et al., Mol. Microbiol. 55 (2005)
1801–1811.32] M. Dhar-Mascareno, J.M. Carcamo, D.W. Golde, Free Radic. Biol. Med. 38 (2005)
1311–1322.33] Y.Q. He, S.P. Liu, L. Kong, Z.F. Liu, Spectrochim. Acta A 61 (2005) 2861–2866.34] W. Haiss, N.T.K. Thanh, J. Aveyard, D.G. Fernig, Anal. Chem. 79 (2007)
4215–4221.35] S.R. Thorpe, J.W. Baynes, Amino Acids 25 (2003) 275–281.36] F. Akhter, M.S. Khan, U. Shahab, Moinuddin, S. Ahmad, Int. J. Biol. Macromol. 58
(2013) 206–210.37] J. Uribarri, W. Cai, M. Peppa, et al., J. Gerontol. A: Biol. Sci. Med. Sci. 62 (4) (2007)
427–433.38] K. Zen, C.X. Chen, Y.T. Chen, et al., J. Immunol. 178 (4) (2007) 2483–2490.39] A. Simm, J. Wagner, T. Gursinsky, et al., Exp. Gerontol. 42 (7) (2007)
668–675.40] N. Ahmed, P.J. Thornalley, Diabetes Obes. Metab. 9 (3) (2007) 233–245.41] N.G. Watkins, C.I. Neglia-Fisher, D.G. Dyer, S.R. Thrope, J.W. Baynes, J. Biol. Chem.
262 (1987) 7207–7212.42] H.F. Bunn, P.J. Higgins, Science 213 (1981) 222–224.43] K. Faustinus, Yeboah, A. Inteaz, A. Varoujan Yaylayan, J. Agric. Food Chem. 47
(1999) 3164–3172.44] M.W. Khan, Z. Rasheed, W.A. Khan, R. Ali, Biochemistry (Mosc.) 72 (2007)
146–152.45] Y. Wei, L. Chen, J. Chen, L. Ge, R.Q. He, BMC Cell Biol. 10 (2009) 10.
esized gold nanoparticles inhibit growth of bone cancer cells, Colloids08
46] G. Brancolini, B. Daria, Kokh, L. Calzolai, C. Rebecca, Wade, S. Corni, ACS Nano6 (11) (2012) 9863–9878.
47] L.A. Dykman, N.G. Khlebtsov, Acta Nat. 3 (2) (2011) 34–55.48] M. Eghtedari, A.V. Liopo, J.A. Copland, A.A. Oraevsky, M. Motamedi, Nano Lett.
9 (2009) 287–291.
Copyright © 2022 FDOKUMEN




















