
Plasmodium vivax Duffy binding protein peptides specifically bind to reticulocytes
10
Plasmodium vivax Duffy binding protein peptides specifically bind to reticulocytes Marisol Ocampo*, Ricardo Vera, Luis Eduardo Rodriguez, Hernando Curtidor, Mauricio Urquiza, Jorge Suarez, Javier Garcia, Alvaro Puentes, Ramse ´s Lopez, Mary Trujillo, Elizabeth Torres, Manuel Elkin Patarroyo Fundacion Instituto de Inmunologı ´a de Colombia Carrera SO No. 26 – 00, Bogota ´, Colombia Received 7 May 2001; accepted 1 August 2001 Abstract Plasmodium vivax Duffy Binding Protein (Pv-DBP) is essential during merozoite invasion of reticulocytes. Reticulocyte binding region identification is important for understanding Pv-DBP reticulocyte recognition. Fifty 20 mer non-overlapping peptides, spanning Pv-DBP sequences, were tested in erythrocyte and reticulocyte binding assays. Ten HARBPs, mainly located in region II (Kd 50 –130 nM), were High Activity Reticulocyte Binding Peptides (HARBPs); one bound to erythrocytes. Reticulocyte trypsin-, chymotrypsin- or neuraminidase- treatment affects HARBP binding differently, suggesting that these peptides have different reticulocyte-binding-sites. Some peptides bound to a Coomasie non-stainable 40 Kda band. Some HARBPs were able to block recombinant PvRII binding (Pv-DBP region II) to Duffy positive reticulocytes. © 2002 Elsevier Science Inc. All rights reserved. Keywords: Plasmodium vivax; Duffy binding protein; HARBP (High Activity Reticulocyte Binding Peptide); HAEBP (High Activity Erythrocyte Binding Peptide) 1. Introduction The invasion of erythrocytes by merozoites from the Plasmodium species (the parasite causing malaria) is a pro- cess involving a series of defined events, having receptor- ligand interactions in which numerous parasite components, as well as receptor molecules on the erythrocyte surface, are involved [3,5,8,12]. It is a well established fact that the Duffy blood group is one of the main receptors used by Plasmodium vivax and Plasmodium knowlesi to invade red blood cells, since they cannot invade Duffy negative human erythrocytes [15]. Nevertheless, P. knowlesi merozoites attachment and reori- entation occurs in both Duffy positive and Duffy negative erythrocytes [13,23]. Plasmodium vivax show the capability of selecting young red blood cells to invade them [11]. In P. vivax and P. knowlesi merozoites a protein has been iden- tified, cloned and sequenced which exhibits specificity to- ward the Duffy human blood group glycoprotein [9]. This protein has a molecular weight of 140 KDa and 135 KDa for P. vivax and P. knowlesi, respectively. The molecule only binds to Duffy positive human erythrocytes, which are sus- ceptible to P. vivax invasion. This Pv DBP is located within the micronemes of late schizonts and free merozoites [1,10] and is presumably released after contact with another re- ceptor on the erythrocyte that mediates initial attachment and apical reorientation. According to the primary sequence, the protein has been divided into six regions. Region I presents a great number of charged residues and it has high homology between P. vivax and P. knowlesi. Region II or the 5 cystein-rich region, contains 12 cysteine residues whose positions are conserved between these two Plasmodia species. The aromatic amino acids in this region are also highly conserved. Furthermore, this region has been defined as the binding functional do- main [2,18]. There is in addition another cystein-rich re- gion, region VI, which precedes the transmembranal region. In this region all the aromatic residues and cysteines are conserved between Plasmodium vivax and Plasmodium knowlesi. Separating the cystein-rich regions there are re- * Corresponding author. Tel.: 57-12-207700 ext 494; fax: 57-1- 280-3999. E-mail address: [email protected] (M. Ocampo). Peptides 23 (2002) 13–22 0196-9781/02/$ – see front matter © 2002 Elsevier Science Inc. All rights reserved. PII: S0196-9781(01)00574-5
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Plasmodium vivax Duffy binding protein peptides specifically bind to reticulocytes

Plasmodium vivax Duffy binding protein peptides specifically bind toreticulocytes
Marisol Ocampo*, Ricardo Vera, Luis Eduardo Rodriguez, Hernando Curtidor,Mauricio Urquiza, Jorge Suarez, Javier Garcia, Alvaro Puentes, Ramses Lopez, Mary Trujillo,
Elizabeth Torres, Manuel Elkin PatarroyoFundacion Instituto de Inmunologıa de Colombia Carrera SO No. 26–00, Bogota, Colombia
Received 7 May 2001; accepted 1 August 2001
Abstract
Plasmodium vivax Duffy Binding Protein (Pv-DBP) is essential during merozoite invasion of reticulocytes. Reticulocyte binding regionidentification is important for understanding Pv-DBP reticulocyte recognition.
Fifty 20 mer non-overlapping peptides, spanning Pv-DBP sequences, were tested in erythrocyte and reticulocyte binding assays. TenHARBPs, mainly located in region II (Kd 50–130 nM), were High Activity Reticulocyte Binding Peptides (HARBPs); one bound toerythrocytes. Reticulocyte trypsin-, chymotrypsin- or neuraminidase- treatment affects HARBP binding differently, suggesting that thesepeptides have different reticulocyte-binding-sites. Some peptides bound to a Coomasie non-stainable 40 Kda band. Some HARBPs wereable to block recombinant PvRII binding (Pv-DBP region II) to Duffy positive reticulocytes. © 2002 Elsevier Science Inc. All rightsreserved.
Keywords: Plasmodium vivax; Duffy binding protein; HARBP (High Activity Reticulocyte Binding Peptide); HAEBP (High Activity Erythrocyte BindingPeptide)
1. Introduction
The invasion of erythrocytes by merozoites from thePlasmodium species (the parasite causing malaria) is a pro-cess involving a series of defined events, having receptor-ligand interactions in which numerous parasite components,as well as receptor molecules on the erythrocyte surface, areinvolved [3,5,8,12].
It is a well established fact that the Duffy blood group isone of the main receptors used by Plasmodium vivax andPlasmodium knowlesi to invade red blood cells, since theycannot invade Duffy negative human erythrocytes [15].Nevertheless, P. knowlesi merozoites attachment and reori-entation occurs in both Duffy positive and Duffy negativeerythrocytes [13,23]. Plasmodium vivax show the capabilityof selecting young red blood cells to invade them [11]. In P.vivax and P. knowlesi merozoites a protein has been iden-tified, cloned and sequenced which exhibits specificity to-
ward the Duffy human blood group glycoprotein [9]. Thisprotein has a molecular weight of 140 KDa and 135 KDa forP. vivax and P. knowlesi, respectively. The molecule onlybinds to Duffy positive human erythrocytes, which are sus-ceptible to P. vivax invasion. This Pv DBP is located withinthe micronemes of late schizonts and free merozoites [1,10]and is presumably released after contact with another re-ceptor on the erythrocyte that mediates initial attachmentand apical reorientation.
According to the primary sequence, the protein has beendivided into six regions. Region I presents a great number ofcharged residues and it has high homology between P. vivaxand P. knowlesi. Region II or the 5� cystein-rich region,contains 12 cysteine residues whose positions are conservedbetween these two Plasmodia species. The aromatic aminoacids in this region are also highly conserved. Furthermore,this region has been defined as the binding functional do-main [2,18]. There is in addition another cystein-rich re-gion, region VI, which precedes the transmembranal region.In this region all the aromatic residues and cysteines areconserved between Plasmodium vivax and Plasmodiumknowlesi. Separating the cystein-rich regions there are re-
* Corresponding author. Tel.: �57-12-207700 ext 494; fax: �57-1-280-3999.
E-mail address: [email protected] (M. Ocampo).
Peptides 23 (2002) 13–22
0196-9781/02/$ – see front matter © 2002 Elsevier Science Inc. All rights reserved.PII: S0196-9781(01)00574-5

gions III, IV and V, variable amongst Plasmodium vivaxand Plasmodium knowlesi sequences.
Sequence homology between the DBP protein from P.vivax and Erythrocyte Binding Antigen 175 (EBA 175)from Plasmodium falciparum has been established. In factEBA-175 has been included within the family of Duffybinding proteins due to gene structure similarity and homol-ogy in amino acid sequences in 3� and 5� cystein-richregions. The Plasmodium falciparum protein differs fromDuffy binding proteins in that it contains two copies (F1 andF2) of 5� cystein-rich regions [2].
Due to the importance already described for the Duffybinding protein in merozoite invasion, there is specific in-terest in looking for the reticulocyte binding sequences inthis protein that are possibly used by the parasite as ligandsto bind to the reticulocytes. In this paper, we describe thePlasmodium vivax Duffy binding protein peptides that havebeen chemically synthesized and, after being radiolabeledwith Na 125I, have been tested in binding assays to reticu-locytes and erythrocytes. Some of their functional activitieswere assessed, and the critical binding amino acids weredetermined, as well as the molecules to which they bind.
2. Materials and methods
2.1. Peptide synthesis
Sequential 20 mer amino acid long peptides from thePlasmodium vivax Salvador I strain Duffy binding protein[9], were chemically synthesized using the Multiple PeptideSynthesis technique in Solid Phase described in previousstudies [17,19,22]. The identity and purity of the peptideswere analyzed by mass spectrometry MALDI-TOF and an-alytical reversed phase-high-performance liquid chromatog-raphy (HPLC) (RP-HPLC). The sequence of synthesizedpeptides is in Fig. 1 represented with a one letter code.Some amino acids in bold type there were replaced byTyrosine to allow their radiolabeling.
2.2. Radiolabeling
Peptides in 1 mg/ml concentration were radiolabeledwith Na125I using chloramine T [25], according to previousreports [17,19]. Radiolabeled peptide was purified in Seph-adex G-10 column (Pharmacia, Uppsala, Sweden).
2.3. Cell binding assay
Reticulocytes from a patient with �-thalasemia and withvery high proportions of reticulocytes in her blood (�85%)were used. Blood, obtained by venipuncture and collected inheparinized tubes, was washed with isotonic PBS andpassed through a cellulose column to remove white bloodcells. It was washed again several times with PBS andcentrifuged at 500� g for 5 min (at room temperature). The
cells were taken to a final hematocrit of 70% and 38 �l ofcells were placed in polystyrene tubes, in presence or ab-sence of excess unlabelled peptide (17 �M). For each tubeincreasing amounts of radiolabeled peptide (4, 8, 14 and 20pmol) were added according to the methodology previouslydescribed for this purpose [19,22].
These assays were carried out in triplicate for each pointon the curve. The added cpm was quantified by means of aninitial reading of the radioactivity. After one hour of reac-tion, the cells were successively washed three times with 25volumes of isotonic PBS. A new gamma counter readingallowed the determination of the amount of radiolabeledpeptide that has been bound to reticulocytes and that re-mains bound after the washings.
2.4. Reticulocyte enzyme treatment
The washed reticulocytes were treated with trypsin (1mg/mL) or chymotrypsin (1 mg/mL) at 37°C for 1 h. Theneuraminidase treatment (300 mU/mL) was carried out insodium acetate Buffer at pH 5.1, for 1 h at 37°C [17,22].Later on, the binding test with the treated cells was per-formed. Untreated reticulocytes were used as control.
2.5. Competition binding assay with analog peptides
In order to recognize the amino acids directly involved inreticulocyte binding, high binding activity peptide ana-logues were synthesized, replacing each one of the aminoacids from the peptide by glycine. A competition bindingassay was then carried out with the radiolabeled originalpeptide at constant 100 nM concentration, incubated inpresence or absence of non-radiolabeled original peptide oreach one of their analogues in increasing concentrations (10,100, 800 and 1,600 nM). Conditions for the test were thesame as for the binding assay [17].
2.6. Cross-linking assays
For the identification of the reticulocyte binding sites ofsome of the high binding activity peptides, some of themwere cross-linked to cell membranes. The binding test wasperformed using a final cell concentration of 4% and, afterincubation with the radiolabeled peptide for 1 h at roomtemperature and through washing with PBS, the boundpeptide was cross-linked with 10 �M BS3 (Bis[sulfosuccin-imidyl] suberate). The reaction was stopped with 20 nMTris-HCl (pH 7.4) and washed again with PBS. Then cellswere treated with lysis buffer (Tris-HCl 5 mM, NaCl 7 mM,EDTA 1 mM, PMSF 0.1 mM). The obtained membraneproteins were solubilized in Laemmli buffer and separatedin SDS-PAGE. The proteins cross-linked with radiolabeledpeptides were exposed on Kodak film (X-OMAT), during24 h to –70°C and the apparent molecular weight wasdetermined using molecular weight markers between 100KDa and 14 KDa (BIO-RAD).
14 M. Ocampo et al. / Peptides 23 (2002) 13–22

Fig. 1. Amino acid sequence and specific binding activity for chemically synthesized Sal-I strain peptides. The peptides are given on the left, with numberindicating their position within the native protein. To the right, the dotted line separates peptides with a binding capacity �2% to reticulocytes anderythrocytes.
15M. Ocampo et al. / Peptides 23 (2002) 13–22

2.7. Competition binding assay with PvRII
The region II (aa 194 to 521) of P. vivax Duffy bindingprotein, expressed in E. coli, purified and refolded [20],kindly provided by Dr Chitnis Chetan (PvRII)) was tested incompetition binding assay. Briefly, 15 �l reticulocytes(50%) were preincubated with inhibitor peptide at differentconcentrations (40, 100, 300 �M). Cells were incubatedwith Pv-RII radiolabeled (Iodogen technique) at 38 nM.Unbound protein was removed with PBS washes, followingincubation. The bound protein was quantified in a gammacounter.
3. Results
3.1. Pv-DBP screening
Fifty Pv-DBP peptides were synthesized and tested inerythrocyte and reticulocyte binding assays as indicatedabove. For each peptide, total and non-specific bindingassays at four different radiolabeled peptide concentrationswere determined. Total binding represents the quantity oflabeled peptide bound to reticulocytes in determined fromthe amount of absence of unlabelled peptide. Non-specificbinding is the peptide that remains bound after inhibitionwith the same non-labeled-peptide. Specific binding is thedifference between total and non-specific binding. If theratio of the slope between the quantity of 125 I peptide thatbinds specifically and the quantity of 125I peptide addedinitially rendered values above 2% (or 0.02 pmol bound perpmol added), the peptide was considered to have high bind-ing activity, based on previously described criteria [19,22].
Fig. 1 shows the sequence of each one of tested peptidesand its position in the Duffy binding protein. For each oneof the peptides, the slope of the specific binding activity toreticulocytes is represented by the black bar and to eryth-rocytes by the gray bars. The dotted line represents 2% ofbinding and is the cut off for selection of high bindingactivity peptides (HABPs).
Ten HARBPs Pv-DBP were identified; nine of thesepeptides clustered between residues 158 to 557, which rep-resents mainly from region II. Only one of them, 1625, alsobinds to erythrocytes. The other HARBP, 1660 (LNSNNNL-SNGKLDIKEYKYR) located in region VI, presents someidentities with the 1623 (SNGQPAGTLDNVLEFVTGHE)HARBP. No peptide bound exclusively to erythrocytes.
3.2. Determination of affinity constants
For selected HARBPs, saturation binding assays weremade using higher radiolabeled peptide concentrations thanin screening binding assay. Scatchard and Hill analysisallowed calculation of affinity constants (Kd), Hill coeffi-cients for each HARBPs and the approximate number ofbinding sites per cell (Fig. 2). All HARBPs present affinity
constants of around 100 nM, the highest affinity being thatof HARBP 1623 variable (SNGQPAGTLDNVLEFVT-GHE) with a Kd of 54 nM.
Peptide 1623 from region I, as well as all the peptides fromregion II: 1625 conserved (TISSAIINHAFLQNTVMKNC),1627 semiconserved (IPDRRYQLCMKELTNLVNNT), 1631variable (DMEGIGYSKVVENNLRSIFG), 1635 variable(RDYVSELPTEVQKLKEKCDG), 1637 variable (SYDQWITRKKNQWDVLSNKF), 1638 variable (ISVKNAEKVQTAGIVTPYDI) 1639 conserved (LKQELDEFNEVAFENEINKR), display positive cooperativity as well aspeptide 1642 semiconserved (SQPVDSSKAEKVPGDSTHGN), located in region III, and peptide 1660 semiconserved(LNSNNNLSNGKLDIKEYKYR) located in region VI.The number of binding sites per cell for these high bindingactivity peptides, calculated from the highest bound peptidevalue on the saturation curve (Bmax in Fig. 2) and thenumber of cells used in this assay, is between 2,000 and37,000.
3.3. Enzyme treatment
HABP binding was performed with the enzymaticallytreated reticulocytes (Table 1). The non-treated reticulo-cytes were used as a control to determine the effect thatreticulocyte enzyme treatment has on the binding activity ofeach one of the HARBPs. Only HARBPs 1631 and 1637binding depended on the presence of sialic acid, since bind-ing to neuraminidase treated reticulocytes dropped to lessthan 30%. But, in general, sialic acid removal facilitatesbinding (HARBPs 1627, 1625, 1635, 1642 and 1660) toreticulocytes. For the other HARBPs, such as 1623, 1638and 1639, there is no effect. Modification of reticulocytesurface with trypsin increases HABP 1637 binding anddecreases binding for HARBPs 1625, 1631, 1635, 1637 and1642 but did not have any effect on the binding of the otherpeptides. Half of the HARBPs reduced binding to chymo-trypsin treated reticulocytes.
3.4. Analogous assay
Fig. 3 shows the critical amino acids for the interactionbetween the 5 conserved HABPs and the reticulocytes.When these amino acids are replaced by glycine, a decreasegreater than 50% in peptide binding activity is observed.The change of any one of the other residues does not alterthe peptide binding specificity. HARBP 1642 show onlytwo critical residues and HARBP 1660 presented the high-est number of critical residues (six). HARBPs 1625 and1660 present a charged critical residue (K215 and K908). Thecritical residues are 53% hydrophobic, 12% charged (onlyK) and 35% polar. The HABP 1627 and 1637 criticalresidues of are mainly hydrophobic.
16 M. Ocampo et al. / Peptides 23 (2002) 13–22
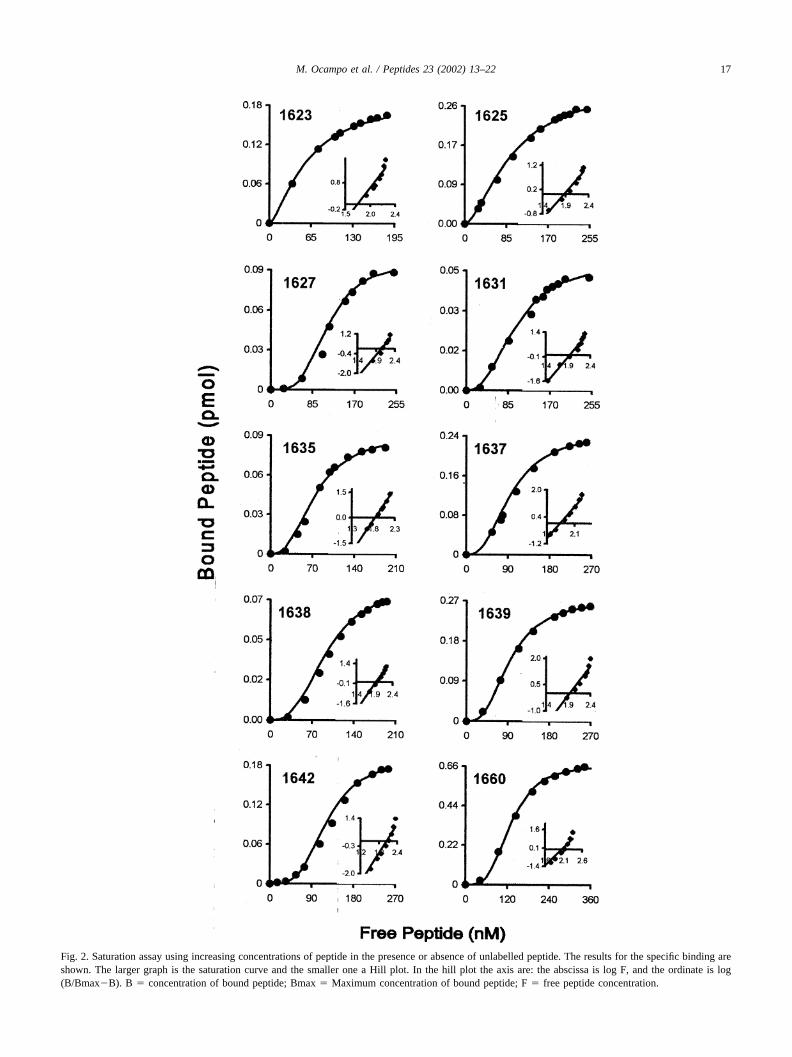
Fig. 2. Saturation assay using increasing concentrations of peptide in the presence or absence of unlabelled peptide. The results for the specific binding areshown. The larger graph is the saturation curve and the smaller one a Hill plot. In the hill plot the axis are: the abscissa is log F, and the ordinate is log(B/Bmax�B). B � concentration of bound peptide; Bmax � Maximum concentration of bound peptide; F � free peptide concentration.
17M. Ocampo et al. / Peptides 23 (2002) 13–22
3.5. Cross-linking assay
As shown in Fig. 4, the 1625, 1637 and 1639 high-binding peptides bind to proteins located on the reticulocytemembrane. Reticulocyte membrane electrophoretical pat-terns are presented in Fig. 4. Peptides bound strongly andspecifically to the 40 KDa band on reticulocyte membrane.The radiolabeled peptide interaction with this band wasinhibited when the binding was performed in the presenceof unlabelled peptide, indicating that a specific binding hadtaken place.
3.6. Competition binding assay with Pv-RII
38 nM radiolabeled recombinant Pv-RII (which includespeptides 1625, 1627, 1631, 1635, 1637, 1638 and 1639)bound to Duffy positive reticulocytes, in the previouslydescribed assay. This binding was inhibited by the samerecombinant protein (400 nM) by around 50%. Competitionbinding assays between the radiolabeled recombinant pro-tein and the HARBPs from this region showed that peptides1625, 1635, 1631 and 1639 specifically and significantlyinhibit reticulocyte protein binding at 40 nM concentrations.However, HARBPs 1627, 1637, 1638 and peptide controlsdid not present significant inhibition (see Fig. 5).
4. Discussion
It is a well established fact that P. vivax merozoitesinvade Duffy positive and not Duffy negative reticulocytes[15]. One protein has been identified from the P. vivaxmerozoite that specifically binds to the erythrocyte Duffyantigen [9]. A binding domain has been reported in thisprotein located in the an area labeled region II [18,7]. Therewas specific interest in the identification and recognition ofDuffy binding protein reticulocyte binding sequences. Ingeneral, peptides have greater binding activity to reticulo-cytes than to erythrocytes (see Fig. 1); according to recentlypublished results, Fy has greater expression on reticulocytesthan on mature RBCs [24].
According to previously established criteria, ten highactivity reticulocytes binding peptides were identified (Fig.1) in Plasmodium vivax Duffy binding protein. Only oneHABP was found [1623] in Duffy binding protein region I(N-terminal). Its binding activity diminished when chymo-trypsin treated reticulocytes were used. Peptides 1625 con-served (TISSAIINHAFLQNTVMKNC), 1627 semiconserved(IPDRRYQLCMKELTNLVNNT), 1631 variable (DMEGIGYSKVVENNLRSIFG), 1635 variable (RDYVSELPTEVQKLKEKCDG), 1637 variable (SYDQWITRKKNQWDVLSNKF), 1638 variable (ISVKNAEKVQTAGIVT-PYDI) and 1639 conserved (LKQELDEFNEVAFENEINKR) are located within the conserved region II (5�Cys-rich region), considered to be important for the bindingto human Duffy positive erythrocytes and reported as beinga Duffy Binding Domain for the Plasmodium vivax andPlasmodium knowlesi ligands. Moreover, binding betweenerythrocytes and this domain (EBP), expressed on the sur-face of COS cells, has been recently studied and it wasfound that binding residues are located in the central part ofthis region [18]. Data from this work supports this hypoth-esis and adds more information concerning relevant bindingsequences involved in this process. Polymorphism studies,including Colombian wild isolates, have identified com-pletely conserved sequences in two peptides (1625 TISSAI-INHAFLQNTVMKNC and 1639 KQELDEFNEVA-FENEINKYR) [4,21] and restricted variability in the N aswell as the C terminal regions in two more (1627 IPDR-RYQLCMKELTNL/FVNNT and 1660 L/SNSNNNL-SNGKLDIKEYK/EYR). The significant variability in theseregions did not seem to affect their binding activity toreticulocytes. These changes could be attributed to immunepressure.
Based on Hill analysis, all Pv-DBP peptides from regionII show positive cooperativity, suggesting that the bindingof each peptide molecule to its possible receptor site, on thereticulocyte membrane, facilitates the binding of the nextmolecule. In this region, there are between 2,000 and 37,000binding sites for each peptide, a range in which the Duffyantigen number per erythrocyte has been reported. Thecompletely conserved 1625 peptide (TISSAIIN-HAFLQNTVMKNC), located in region II, is the only onethat also has high specific erythrocyte binding activity. Thispeptide is probably very important in reported erythrocytebinding studies, in which the Pv-DBP erythrocyte bindingdomain was identified [2,7,18].
Conserved HABPs 1642 (SQPVDSSKAEKVPGD-STHGN) and 1660 (LNSNNNLSNGKLDIKEYKYR) arelocated in region II and region VI (3�Cys-rich region); todate, these regions have not been involved in the bindinghuman Duffy positive erythrocytes process.
In glycine peptide analog competition binding assays, thecritical residues for the binding process were found for eachconserved HABP, as shown in Fig. 3 for peptides 1625,1627, 1639, 1642 and 1660. It is likely that these residuesmay not only correspond to the amino acids directly in-
Table 1Relative binding to enzyme-treated reticulocytes
Peptide Neuraminidase Trypsin Chymotrypsin Control
1623 70 � 5 85 � 9 5 � 1 100 � 141625 140 � 10 5 � 1 5 � 1 100 � 111627 300 � 25 111 � 12 111 � 10 100 � 71631 30 � 3 26 � 3 52 � 5 100 � 271635 240 � 22 34 � 5 5 � 1 100 � 51637 5 � 1 15 � 2 100 � 12 100 � 101638 83 � 9 61 � 7 72 � 6 100 � 81639 111 � 10 80 � 8 120 � 15 100 � 81642 150 � 17 50 � 4 5 � 1 100 � 11660 400 � 45 167 � 15 100 � 12 100 � 17
18 M. Ocampo et al. / Peptides 23 (2002) 13–22

Fig. 3. Competition assay with 1625, 1627, 1639, 1642 and 1660 analogous peptides. The height of the black bars is proportional to the binding inhibitionof the original radiolabeled peptide (100 nM) by the original unlabeled peptide (*) (10 nM and 100 nM) or unlabeled analog peptides. The letter at the bottomrepresents the amino acid replaced by glycine in the analog peptide.
19M. Ocampo et al. / Peptides 23 (2002) 13–22

volved in binding to receptor on the reticulocyte, but mayalso be responsible for keeping the tridimensional peptidestructure which is necessary for binding. According to poly-morphism studies, most of these critical residues do nothave genetic variations in the different isolates [4]. Takinginto account that 53% of critical residues are hydrophobic,the interaction between the HABPs and reticulocytes is, in
part, mediated by hydrophobic interactions in spite ofHABPs being highly polar. The change of the same closeresidue affects binding: e.g. I203 and I204, HABP-1625; S543
and S544, HABP-1642 or N901 and N902, HABP-1660; un-derlined critical residues present a different effect on reticu-locyte binding activity. These indicate that the HABP bind-ing to reticulocytes depends on peptide sequence and not onamino acid composition. In general, the change of chargedresidues (R242, R243, K249, E250, K480, E481, D483, D542 andothers) does not affect the reticulocyte binding activity ofthese HABPs. These results, together with the HABP-bind-ing to neuraminidase-treated reticulocytes, indicate that theinteraction between HABPs and reticulocytes is indepen-dent of the peptide and reticulocyte charge and more de-pendent on peptide amino acid sequence.
Neuraminidase treatment of reticulocytes only decreasedthe binding activity of peptides 1631 and 1637. For the otherpeptides, such as 1627, 1635, 1642 and 1660, sialic acidremoval with neuraminidase increased their binding activ-ity, indicating that the binding is sialic acid independent andprobably the binding sites for these peptides are hindered bysialic acid residues. Interestingly, it has been reported thatneuraminidase treatment of human Fy (a�b�) erythrocytesdoes not affect Plasmodium vivax invasion [14]. Similarresults have been obtained for Plasmodium knowlesi [14,6],in spite of neuraminidase treatment possibly removing theDuffy blood group sialic acid.
In this study, it was observed that binding activity de-creased for peptides 1625, 1631, 1635 and 1642 when thereticulocytes were trypsin or chymotrypsin treated and for1623 when reticulocytes were chymotrypsin treated. It hasbeen reported that the chymotrypsin treatment of Duffypositive erythrocytes removes the Duffy determinants Fya,Fyb and Fy6 but not Fy3, Fy4 and Fy5, and that treatmentinhibits invasion by Plasmodium vivax [13,16]. Moreoveranti-Fya, Fyb and Fy6 antibodies inhibit reticulocyte inva-sion.
The receptor sites for these peptides seem to be mem-brane proteins susceptible to trypsin or chymotrypsin treat-ment.
Peptide 1637 binding sites seem to be on a sialoglycop-rotein, according to neuraminidase or trypsin-treated reticu-locyte binding results. In studies with trypsin-treated eryth-rocytes, there is an increase in Fy3 and Fy6 reactivity andslight reduction of Fyb activity and non inhibition of Plas-modium vivax invasion of erythrocytes [14,16]. This couldindicate that the 1637 peptide binding sites are not involvedin critical steps during P. vivax merozoite invasion, or thatpeptide 1637 carries an alternative reticulocyte binding se-quence for Duffy binding protein, since the reticulocyte-binding sites are chymotrypsin resistant. These results thusshow that there are different binding sites on reticulocytesand some of them could reside on Duffy antigen.
After analyzing enzymatic treatment results, we suggestthat the 1637 and 1639 peptides do specifically bind toreticulocytes, but to different binding sites (see above) on
Fig. 4. Crosslinking assay for peptides 1625, 1637 and 1639. A) Reticu-locyte membrane patterns (lanes 1 and 2) and autoradiographs for reticu-locyte proteins which were specifically cross-linked with radiolabeledpeptide 1625 (Lanes 3 and 4). Lanes 1 and 3 represent total binding andlanes 2 and 4 show inhibited binding. B) For peptides 1637 and 1639SDS-PAGE (lanes 1 to 4) autoradiographs (lanes 5 to 8): peptide 1637(lanes 1, 2, 5, 6), total binding (lanes 1 and 5) and inhibited binding (lanes2 and 6) are presented; peptide 1639 (lanes 3, 4, 7, 8), total binding (lanes3 and 7) and inhibited binding (lanes 4 and 8) are presented.
Fig. 5. Inhibition of Pv-RII protein binding to reticulocytes. Each one of thepeptides is pre-incubated with reticulocytes and then incubated with radio-labeled protein. The percentage of Pv-RII protein binding to reticulocytesis shown at 40 �M peptide concentration for each one. C1 is a peptidewhich does not belong to the Pv-DBP protein region II. C2 is a peptidewhich does not bind to reticulocytes. C3 corresponds to protein binding inthe presence of non-radiolabeled Pv-RII. T represents the total value ofradiolabeled protein binding to reticulocytes.
20 M. Ocampo et al. / Peptides 23 (2002) 13–22

the same molecule, since they bind to the same 40 KDaprotein (Fig. 4). These binding sites are chymotrypsin-re-sistant; in spite of this (and considering that these peptidesbind to the reticulocyte membrane 40 KDa band) Coomasiedid not stain this band. The glycophorin Duffy Blood grouphas been reported as being a 35 to 43 KDa erythrocytesurface protein. Therefore we suggest that the 40 KDabinding molecule on the reticulocytes identified by cross-linking is the Duffy antigen.
It should be emphasized that ten Duffy binding proteinpeptides having high binding activity to reticulocytes, lo-cated in regions I, II and VI, were found (two of themconserved, three semiconserved and five variable) and thataffinity constants calculated were between 50 and 130nM(representing high cell affinity). Hill coefficients demon-strate that the high binding activity peptides have positivecooperativity.
The protein was gently radiolabeled with 125I by theIodogen method, since it was suggested by Dr. Chitnis [20]that strong radio labeling could abolish binding to reticulo-cytes. Peptides 1625, 1631, 1635 and 1639 are able toinhibit recombinant protein binding to Duffy positive re-ticulocytes. It has been reported that denatured recombinantprotein is not capable of binding to Duffy positive erythro-cytes, suggesting that this protein’s 3D structure is veryimportant to these proteins’ binding activity [20].
However, we found that linear peptides are capable ofpartly inhibiting this protein’s binding, suggesting that thesepeptides are exposed on the protein surface and are probablynot involved in disulfide bridge formation. Besides that,peptide 1625 is located at the N terminal fragment. Al-though results suggest that the peptides have lesser affinityfor reticulocytes, since the peptide concentration needed toinhibit such protein binding must be in the micromolarrange, these results strongly suggest that these sequences areinvolved in protein recognition of the reticulocyte. This isalso supported by the 1625 1635 and 1639 HARBP se-quences being highly conserved among the different para-site strains reported to date.
Evidence for HARBP role during merozoite invasion isbased on the fact that the majority of these sequences arelocated in region II, for which there is reported experimentalevidence showing its importance as ligand during merozoiteinvasion. There are different reticulocyte binding sites forthese sequences; further studies could show that this shouldbe an advantage for protein function during merozoite in-vasion.
Acknowledgments
We are extremely grateful to Dr Chetan Chitnis, from theInternational Centre for Genetic Engineering and Biotech-nology, New Delhi, for his kind and invaluable gift ofrecombinant PvRII and his suggestion for using it in mildconditions, which led to our successful results. This re-
search project was supported by the Presidency of the Re-public of Colombia, and the Ministry of Public Health. Wethank Jason Garry for painstakingly reviewing the manu-script. The collaboration of Aurora Cortes B and our chem-istry section is greatly appreciated.
5. References
[1] Adams JH, Hudson DE, Torit M, Ward GE, Wellems TE, Aikawa M,Miller LH. The Duffy receptor family of Plasmodium knowlesi islocated within the micronemes of invasive malaria merozoites. Cell1990;63:143–53.
[2] Adams JH, Sim LK, Dolan SA, Fang X, Kaslow DC, Miller LH. Afamily of erythrocyte binding proteins of malaria parasites. Proc NatlAcad Sci USA 1992;89:7085–9.
[3] Aikawa M, Miller LH, Jonhson J, Rabbege JR. Erythrocyte entry bymalaria parasites: A moving junction between erythrocyte and para-site. J Cell Biol 1978;77:72–82.
[4] Ampudia E, Patarroyo MA, Patarroyo ME, Murillo LA. Geneticpolymorphism of the Duffy receptor binding domain of Plasmodiumvivax in Colombian wild isolates. Mol Biochem Parasitol 1996;78:269–72.
[5] Bannister LH, Butcher GA, Dennis ED, Mitchell GH. Structure andinvasive behavior of Plasmodium knowlesi merozoites in vitro. Par-asitology 1975;71:483–91.
[6] Barnwell JW, Nichols M, Rubinstein P. In vitro evaluation of the roleof the Duffy blood group in erythrocyte invasion by Plasmodiumvivax. J Exp Med 1989;169:1795–802.
[7] Dutta S, Daugherty JR, Ware LA, Lanar DE, Ockenhouse CF. Ex-pression, purification and characterization of a functional region ofthe Plasmodium vivax Duffy binding protein. Mol Biochem Parasitol2000;109:179–84.
[8] Dvorak JA, Miller LH, Whitehouse WC, Shiroishi T. Invasion oferythrocytes of malaria merozoites. Science 1975;187:748–9.
[9] Fang X, Kaslow DC, Adams JH, Miller LH. Cloning of the Plasmo-dium vivax Duffy receptor. Mol Biochem Parasitol 1991;44:125–32.
[10] Fraser T, Michon P, Barnwell JW, Noe AR, Al-Yaman F, KaslowDC, Adams JH. Expression and serologic activity of a soluble re-combinant Plasmodium vivax Duffy binding protein. Infect Immun1997;65:2772–7.
[11] Galinski MR, Corredor C, Ingravallo P, Barnwell J. A reticulocyte-binding protein complex of Plasmodium vivax merozoites. Cell 1992;69:1213–26.
[12] Hadley TJ, Klotz FW, Miller LH. Invasion of erythrocytes by malariaparasites: A cellular and molecular overview. Annu Rev Microbiol1986;40:451–77.
[13] Miller LH, Mason SJ, Dvorak JA, McGinniss MH, Rothman IK.Erythrocyte receptors for (Plasmodium knowlesi) malaria: Duffyblood group determinants. Science 1975;189:561–3.
[14] Miller LH, Shiroishi T, Dvorak JA, Durocher JR, Schrier BK. Enzy-matic modification of the erythrocyte membrane surface and its effecton malarial merozoite invasion. J Mol Med 1975;1:55.
[15] Miller LH, Mason SJ, Clyde DF, McGinniss MH. The resistancefactor to Plasmodium vivax in blacks: The Duffy blood group geno-type FyFy. New Engl J Med 1976;295:302–4.
[16] Nichols M, Rubinstein P, Barnwell J, Rodriguez S, Rosenfield RE. ANew Human Duffy Blood Group Specificity Defined by a MurineMonoclonal Antibody Immunogenetics and Association with Suscep-tibility to Plasmodium vivax. J Exp Med 1987;166:776–85.
[17] Ocampo M, Urquiza M, Guzman F, Rodrıguez LE, Suarez J, CurtidorH, Rosas J, Dıaz M, Patarroyo ME. Two MSA-2 peptides that bind tohuman red blood cells are relevant to Plasmodium falciparum mer-ozoite invasion. J Peptide Res 2000;55:216–23.
21M. Ocampo et al. / Peptides 23 (2002) 13–22

[18] Ranjan A, Chitnis Ch. Mapping regions containing binding residueswithin functional domains of Plasmodium vivax and Plasmodiumknowlesi erythrocyte-binding proteins. Proc Natl Acad Sci USA1999;96:14067–72.
[19] Rodriguez LE, Urquiza M, Ocampo M, Suarez JE, Curtidor H, Guz-man F, Vargas LE, Trivinos M, Rosas M, Patarroyo ME. Plasmodiumfalciparum EBA-175 KDa protein peptides which bind to human redblood cells. Parasitology 2000;120:225–35.
[20] Singh S, Pandey K, Chattopadhayay R, Yazdani SS, Lynn A, Bharad-waj A, Ranjan A, Chitnis C. Biochemical, biophysical, and functionalcharacterization of bacterially expressed and refolded receptor bind-ing domain of Plasmodium vivax Dufy-binding protein. J Biol Chem2001;276:17111–16.
[21] Tsuboi T, Kappe SHI, Al-Yaman F, Prickett MD, Alpers M, AdamsJH. Natural variation within the principal adhesion domain of the
Plasmodium vivax Duffy binding protein. Infect Immun 1994;62:5581–6.
[22] Urquiza M, Rodriguez LE, Suarez JE, Guzman F, Ocampo M, Curti-dor H, Segura C, Trujillo E, Patarroyo ME. Identification of Plasmo-dium falciparum MSP-1 peptides from the protein able to bind to redblood cells. Parasite Immunol 1996;18: 515–26.
[23] Wertheimer SP, Barnwell JW. Plasmodium vivax interaction with thehuman Duffy blood group glycoprotein: Identification of a parasitereceptor-like protein. Exp Parasitol 1989;69:340–50.
[24] Woolley IJ, Hotmire KA, Sramkoski RM, Zimmerman PA. Dif-ferential expression of the Duffy antigen receptor for chemokinesaccording to RBC age and FY genotype. Transfusion 2000;40:949 –53.
[25] Yamamura HY, Enna SJ, Kuhar MJ. Neurotransmitter ReceptorBinding. New York: Raven Press, 1978.
22 M. Ocampo et al. / Peptides 23 (2002) 13–22




















