
Endostatin/collagen XVIII—an inhibitor of angiogenesis—is expressed in cartilage and...
10
Endostatin/collagen XVIII—an inhibitor of angiogenesis—is expressed in cartilage and fibrocartilage Thomas Pufe a, * , Wolf J. Petersen b , Nicolai Miosge c , Mary B. Goldring d , Rolf Mentlein a , Deike J. Varoga e , Bernhard N. Tillmann a a Department of Anatomy, Christian-Albrechts-University, Olshausenstrasse 40, D-24098 Kiel, Germany b Department of Trauma, Hand and Reconstructive Surgery, University Hospital Mu ¨nster, Germany c Department of Anatomy Georg-August-University, Go ¨ttingen, Germany d Beth Israel Deaconess Medical Center, New England Baptist Bone and Joint Institute and Harvard Medical School, Boston, MA 02115, USA e Department of Orthopaedic Surgery, University Hospital Schleswig-Holstein, Campus Kiel, Germany Received 20 January 2004; received in revised form 8 June 2004; accepted 8 June 2004 Abstract Aim of the study was to get a deeper insight in the mechanisms regulating avascularity of cartilaginious tissues. In the center of our interest was the expression of the anti-angiogenic fragment of collagen XVIII and its potency to inhibit angiogenesis. We observed a strong endostatin/ collagen XVIII production in articular and fibrocartilage and an inhibitory potency concerning the VEGF-signalling pathway. Introduction: Cartilaginous tissue is mainly avascular and shows a limited intrinsic capacity for healing. Aim of this study was to investigate the expression of the antiangiogenic peptide endostatin/collagen XVIII in cartilage and fibrocartilage. Results: In fetal epiphyseal cartilage of humans high endostatin/collagen XVIII levels could be detected by ELISA whereas significantly lower levels were found in articular cartilage of adults. In the fibrocartilaginous tissue of the menisci, there was no significant difference in the endostatin/collagen XVIII concentrations between samples of fetuses and adults. But in the menisci of adults, endostatin/collagen XVIII concentrations were higher in the internal avascular two thirds of the meniscus whereas in the fetal menisci higher endostatin/collagen XVIII concentrations were found in the external third. Endostatin/collagen XVIII immunostaining of rat articular cartilage shows that endostatin/collagen XVIII downregulation starts soon after birth. In fetal cartilage and fibrocartilage of rats and humans, endostatin/ collagen XVIII could be immunostained in the extracellular matrix and in the pericellular matrix of endothelial cells, fibrochondrocytes and chondrocytes. In adult cells, weak endostatin/collagen XVIII immunostaining was restricted to the pericellular matrix of fibrochondrocytes and chondrocytes. The detection of endostatin/collagen XVIII could be verified by in situ hybridization. Chondrocytes in vitro released measurable amounts of endostatin/collagen XVIII into culture supernatants. Stimulation of chondrocytes with EGF, as an example of a growth factor, or dexamethasone had no influence on endostatin/collagen XVIII expression. Endostatin inhibited VEGF- induced phosphorylation of MAPK in chondrocytes. Conclusions: The spatial and temporal expression of endostatin/collagen XVIII in cartilaginous tissue and its potency regarding inactivation of VEGF signalling suggests that this antiangiogenic factor is important not only for the development but also for the maintenance of avascular zones in cartilage and fibrocartilage. Experimental procedures: We analyzed the spatial and temporal expression of endostatin/collagen XVIII—an endogenous angiogenesis inhibiting factor—in cartilage and fibrocartilage of humans and rats by immunohistochemical and biochemical (ELISA) methods and by in situ hybridization. To elucidate possible factors responsible for the induction or suppression of endostatin/collagen XVIII in cartilaginous tissues, chondrocytes (cell line C28/I2) were exposed to EGF and dexamethason. To study the possible interaction of endostatin/collagen XVIII with angiogenic factors, the immortalized human chondrocytes (C28/I2) have been incubated with VEGF and the phosphorylation of the MAPK Erk 1/2 (extracellular-regulated kinases), a known signal transduction pathway for VEGF has been determined under the influence of endostatin. D 2004 Elsevier B.V./International Society of Matrix Biology. All rights reserved. Keywords: Endostatin; Vascular endothelial growth factor (VEGF); Cartilage; Angiogenesis; Antiangiogenesis 0945-053X/$ - see front matter D 2004 Elsevier B.V./International Society of Matrix Biology. All rights reserved. doi:10.1016/j.matbio.2004.06.003 * Corresponding author. Tel.: +49-431-880-3087; fax: +49-431-880-1557. E-mail addresses: [email protected] (T. Pufe), [email protected] (W.J. Petersen), [email protected] (N. Miosge), [email protected] (M.B. Goldring), [email protected] (R. Mentlein), [email protected] (D.J. Varoga), [email protected] (B.N. Tillmann). www.elsevier.com/locate/matbio Matrix Biology 23 (2004) 267 – 276
-
Upload
sacklerinstitute -
Category
Documents
-
view
1 -
download
0
Transcript of Endostatin/collagen XVIII—an inhibitor of angiogenesis—is expressed in cartilage and...

www.elsevier.com/locate/matbio
Matrix Biology 23 (2004) 267–276
Endostatin/collagen XVIII—an inhibitor of angiogenesis—is expressed
in cartilage and fibrocartilage
Thomas Pufea,*, Wolf J. Petersenb, Nicolai Miosgec, Mary B. Goldringd,Rolf Mentleina, Deike J. Varogae, Bernhard N. Tillmanna
aDepartment of Anatomy, Christian-Albrechts-University, Olshausenstrasse 40, D-24098 Kiel, GermanybDepartment of Trauma, Hand and Reconstructive Surgery, University Hospital Munster, Germany
cDepartment of Anatomy Georg-August-University, Gottingen, GermanydBeth Israel Deaconess Medical Center, New England Baptist Bone and Joint Institute and Harvard Medical School,
Boston, MA 02115, USAeDepartment of Orthopaedic Surgery, University Hospital Schleswig-Holstein, Campus Kiel, Germany
Received 20 January 2004; received in revised form 8 June 2004; accepted 8 June 2004
Abstract
Aim of the study was to get a deeper insight in the mechanisms regulating avascularity of cartilaginious tissues. In the center of our interest
was the expression of the anti-angiogenic fragment of collagen XVIII and its potency to inhibit angiogenesis. We observed a strong endostatin/
collagen XVIII production in articular and fibrocartilage and an inhibitory potency concerning the VEGF-signalling pathway.
Introduction: Cartilaginous tissue is mainly avascular and shows a limited intrinsic capacity for healing. Aim of this study was to
investigate the expression of the antiangiogenic peptide endostatin/collagen XVIII in cartilage and fibrocartilage. Results: In fetal
epiphyseal cartilage of humans high endostatin/collagen XVIII levels could be detected by ELISA whereas significantly lower levels
were found in articular cartilage of adults. In the fibrocartilaginous tissue of the menisci, there was no significant difference in the
endostatin/collagen XVIII concentrations between samples of fetuses and adults. But in the menisci of adults, endostatin/collagen XVIII
concentrations were higher in the internal avascular two thirds of the meniscus whereas in the fetal menisci higher endostatin/collagen
XVIII concentrations were found in the external third. Endostatin/collagen XVIII immunostaining of rat articular cartilage shows that
endostatin/collagen XVIII downregulation starts soon after birth. In fetal cartilage and fibrocartilage of rats and humans, endostatin/
collagen XVIII could be immunostained in the extracellular matrix and in the pericellular matrix of endothelial cells, fibrochondrocytes
and chondrocytes. In adult cells, weak endostatin/collagen XVIII immunostaining was restricted to the pericellular matrix of
fibrochondrocytes and chondrocytes. The detection of endostatin/collagen XVIII could be verified by in situ hybridization. Chondrocytes
in vitro released measurable amounts of endostatin/collagen XVIII into culture supernatants. Stimulation of chondrocytes with EGF, as
an example of a growth factor, or dexamethasone had no influence on endostatin/collagen XVIII expression. Endostatin inhibited VEGF-
induced phosphorylation of MAPK in chondrocytes. Conclusions: The spatial and temporal expression of endostatin/collagen XVIII in
cartilaginous tissue and its potency regarding inactivation of VEGF signalling suggests that this antiangiogenic factor is important not
only for the development but also for the maintenance of avascular zones in cartilage and fibrocartilage. Experimental procedures: We
analyzed the spatial and temporal expression of endostatin/collagen XVIII—an endogenous angiogenesis inhibiting factor—in cartilage
and fibrocartilage of humans and rats by immunohistochemical and biochemical (ELISA) methods and by in situ hybridization. To
elucidate possible factors responsible for the induction or suppression of endostatin/collagen XVIII in cartilaginous tissues, chondrocytes
(cell line C28/I2) were exposed to EGF and dexamethason. To study the possible interaction of endostatin/collagen XVIII with
angiogenic factors, the immortalized human chondrocytes (C28/I2) have been incubated with VEGF and the phosphorylation of the
MAPK Erk 1/2 (extracellular-regulated kinases), a known signal transduction pathway for VEGF has been determined under the
influence of endostatin.
D 2004 Elsevier B.V./International Society of Matrix Biology. All rights reserved.
Keywords: Endostatin; Vascular endothelial growth factor (VEGF); Cartilage; Angiogenesis; Antiangiogenesis
* Corresponding author. Tel.: +49-431-880-3087; fax: +49-431-880-1557.
0945-053X/$ - see front matter D 2004 Elsevier B.V./International Society of Matrix Biology. All rights reserved.
doi:10.1016/j.matbio.2004.06.003
E-mail addresses: [email protected] (T. Pufe), [email protected] (W.J. Petersen), [email protected] (N. Miosge),
[email protected] (M.B. Goldring), [email protected] (R. Mentlein), [email protected] (D.J. Varoga), [email protected]
(B.N. Tillmann).

T. Pufe et al. / Matrix Biology 23 (2004) 267–276268
1. Introduction 1990), interferon-alpha (Angionello et al., 1995), thrombo-
Hyaline cartilage of adults is an avascular tissue with a
poor repair potential. In contrast, certain tunnels containing
blood vessels and known as cartilage canals are generally
found in the cartilaginous epiphyses of the fetal skeleton.
The significance of these vessels in cartilage nutrition and in
initiation of centers of ossification has still to be accurately
defined and the mechanism controlling the development and
regression of these structures are largely unknown (Brookes
and Revell, 1998).
In contrast to articular cartilage, these canals are lacking
during the fetal development of fibrocartilaginous structures
such as in the menisci of the knee (Petersen and Tillmann,
1995). Nevertheless, microvascular studies have shown that
in the late fetal stage and at the time of birth the complete
knee joint meniscus is vascularized. This vascular network
regresses soon after birth and in the adult the vascular supply
of the meniscus is restricted to a small rim close to the joint
capsule (Petersen and Tillmann, 1995). This specific vascu-
lar supply has clinical relevance regarding the healing of
meniscus tears. Lesions in the vascularized external part
have a good repair response, whereas lesions in the avascular
inner two thirds frequently fail to heal (Petersen and Till-
mann, 1995). Since the vascular status of cartilage and
fibrocartilage is closely related to its repair abilities, an
understanding of the mechanism regulating the tissue-spe-
cific vascularity might give further insights into the mecha-
nisms of healing and degeneration.
Angiogenesis—the formation of new blood vessels from
preexisting capillaries—is balanced controlled by a variety
of stimulating (angiogenic) and inhibiting (antiangiogenic)
peptides that act on invading endothelial and smooth muscle
cells (Mentlein and Held-Feindt, 2003). Endogenous inhi-
bition of angiogenesis is necessary for the development and
maturation of tissues which are largely avascular. This
might be caused either by expression of inhibitory factors
for vascular endothelial cells or by an intrinsic insufficiency
of cartilage cells to express stimulatory peptides (Kim et al.,
2000).
One of the most important angiogenetic factors is the
vascular endothelial growth factor (VEGF) (Ferrara, 1999).
VEGF has been identified in the zone of hypertrophic
cartilage in the growth plate (Gerber et al., 1999). VEGF
plays an integral role in endochondral ossification and for
the longitudinal growth of the skeleton. In adult articular
cartilage, VEGF is largely downregulated (Pufe et al.,
2001a) but in the disease state such as in osteoarthritis, this
angiogenic factor is reexpressed by chondrocytes. In the
knee joint menisci, high VEGF levels have been found
during the healing of meniscus lesions (Becker et al., in
press). These studies show that angiogenesis plays an
important role not only for physiological but also for
pathological events of the skeleton.
Several endogenous inhibitors of angiogenesis have been
identified. These include platelet factor 4 (Maione et al.,
spondin (Good et al., 1990), metastatin (Liu et al., 2001),
troponin-1 (Moses et al., 1999) or angiostatin (O’Reilly et
al., 1994), or endostatin (O’Reilly et al., 1997).
Endostatin is a 20-kDa antiangiogenic factor originally
identified from murine hemangioendothelioma cells and
seems to counteract many VEGF induced effects (O’Reilly
et al., 1997). Endostatin inhibits VEGF-induced endothelial
cell migration (Yamaguchi et al., 1999), VEGF-mediated
neovascularization (Takahashi et al., 2003) and VEGF-
induced vascular permeability (Takahashi et al., 2003).
Under physiologic conditions, endostatin is critical for the
regression of blood vessels in the vitreous of the eye (Fukai
et al., 2002). Despite some structural homologies of the
vitreous and cartilage, there are no information about endo-
statin/collagen XVIII expression in articular cartilage or the
menisci in the literature. However, in a recent study we
could identify endostatin/collagen XVIII in the fibrocarti-
lage of a ‘‘wrap around tendon’’ (retromalleolar region of
the posterior tibial tendon) (Pufe et al., 2003).
The aim of this study was to examine the spatial and
temporal expression of endostatin/collagen XVIII in carti-
lage and fibrocartilage, to elucidate possible factors respon-
sible for the induction or suppression of endostatin/collagen
XVIII in chondrocytes, and to study the possible interaction
of endostatin with a well-known signal transduction path-
way for VEGF (MAPK Erk 1/2 [extracellular-regulated
kinases]).
2. Results
2.1. Spatial and temporal expression of endostatin/collagen
XVIII in cartilage and fibrocartilage
Endostatin/collagen XVIII could be intensively immu-
nostained in fetal menisci and in epiphyseal cartilage (Fig. 1).
In the fetal meniscus, most cells had a fibroblastic
phenotype and type II collagen immunostaining was nega-
tive. Endostatin/collagen XVIII was present throughout the
basement membranes of vessels and in the pericellular
matrix of meniscus cells (Fig. 1). Sections that were immu-
nostained with antibodies against the angiogenic peptide
vascular endothelial growth factor (VEGF) showed that the
majority of cells in the fetal menisci express this peptide as
well. Factor VIII immunostaining reveled that the fetal
meniscus was nearly completely vascularized (not shown).
In the epiphysis of the distal femur-intensive endostatin/
collagen XVIII, immunostaining was observed not only in
the pericellular matrix of chondrocytes but also in the
extracellular matrix of the cartilage (Fig. 1a). Endostatin/
collagen XVIII immunostaining was negative in chondro-
cytes and in the extracellular matrix around cartilage
channels (Fig. 1a). The chondrocytes around the cartilage
channels showed a strong immunoreactivity for VEGF
(Fig. 1b). In the growth plate, endostatin/collagen XVIII

Fig. 1. Immunostaining of endostatin/collagen XVIII in human cartilage and menisci. Immunostaining of endostatin/collagen XVIII (a, c– f) and of VEGF (b)
in fetal epiphyseal (a) and adult (c) articular cartilage and in growth plate (e) and in fetal (b) and adult menisci (d). In adult cartilage (f), no immunoreactions
were detectable after adsorption of the endostatin/collagen XVIII antibody to recombinant endostatin. Nuclei in the sections were counterstained with Meyer’s
hemalum. Bar = 10 Am (c–e) or 100 Am (a, b, e); original magnification: 175-fol
Fig. 2. (A) Light microscopic in situ-hybridisation with the anti-sense probe
for endostatin of collagen type XVIII in human osteoarthritic cartilage. The
mRNA (asterisk) is found in the chondrocytes adjacent to the tidemark
(arrows). (B) The corresponding sense probe shows no staining, bar = 70 Am.
T. Pufe et al. / Matrix Biology 23 (2004) 267–276 269
could be immunostained in the proliferation zone but not
in the zone of hypertrophic cartilage (Fig. 1e). This zone
was strongly immunopositive for VEGF (not shown).
In the adult, immunolabeling for endostatin/collagen
XVIII was positive in the pericellular matrix of articular
chondrocytes (Fig. 1d) and meniscus fibrochondrocytes (not
shown). The extracellular matrix of cartilage and menisci
was endostatin/collagen XVIII negative.
2.2. Expression of collagen XVIII mRNA in human cartilage
Collagen-type XVIII mRNA could be detected in fetal,
normal (not shown) and osteoarthritic cartilage (Fig. 2A).
The mRNA (asterisk) is found in the chondrocytes adjacent
to the tidemark (arrows) (Fig. 2B). The corresponding sense
probe shows no staining.
2.3. Endostatin/collagen XVIII expression is age-related
in rat articular cartilage and menisci
To study endostatin/collagen XVIII concentrations dur-
ing childhood, a rat model was used. Endostatin/collagen
XVIII could be intensively immunostained in fetal menisci
and in epiphyseal cartilage (Fig. 3a and b). The expression
of endostatin/collagen XVIII decreased shortly after birth
(Fig. 3c,d,f,g). The immunostaining in the articular cartilage
of 1- and 2-day-old rats was markedly weaker than in the
fetal animal. In the articular cartilage and menisci of adult
animals, endostatin/collagen XVIII was only slightly detect-
able (Fig. 3h and i).
2.4. Endostatin/collagen XVIII concentrations in
cartilage and menisci
The endostatin/collagen XVIII concentrations in fetal and
adult menisci samples were homogenized and endostatin/
collagen XVIII was measured by a sensitive ELISA (Fig. 4).
The highest average endostatin/collagen XVIII concen-
trations were found in epiphyseal cartilage (mean: 964 pg/
d (a, b, e); 750-fold (c, d, e).

Fig. 3. (a– i) Immunostaining of endostatin/collagen XVIII in fetal (a and b), 1-day-old (c–e), 2-day-old (f, g) and 5-month-old rat joints (h, i). In articular
cartilage, immunoreactions were relatively high in fetal and less intense in normal adult cartilage and meniscus tissues. No immunoreaction was detectable after
adsorption of the endostatin/collagen XVIII antibody to recombinant endostatin (e). Nuclei in the sections were counterstained with Meyer’s hemalum.
Bar = 100 Am (a, c–h); Bar = 10 Am (b, i); original magnification: 45-fold (a, c); 90-fold (f, h); 175-fold (d, e, g); 350-fold (b); 560-fold (i).
T. Pufe et al. / Matrix Biology 23 (2004) 267–276270
mg fresh weight). In the articular cartilage of adults, endo-
statin/collagen XVIII concentrations were significantly low-
er ( p < 0.05), but average endostatin/collagen XVIII
concentrations of 228 pg/mg fresh weight could be still
measured. The lowest endostatin/collagen XVIII concentra-
tions were measured in cartilage samples from patients with
osteoarthritis (147 pg/mg fresh weight).
In the fetal menisci, the average endostatin/collagen
XVIII concentrations (mean: 569 pg/mg fresh weight) were
significantly lower than in fetal epiphyseal cartilage but still
higher than in adult articular cartilage. In the adult, there
was no significant change in the overall endostatin/collagen
XVIII concentrations of the menisci as compared to fetal
tissue. However in the adult, endostatin/collagen XVIII
concentrations increased significantly ( p < 0.05) in the in-
ternal two thirds (ITT) and decreased significantly in the
external third (ET) of the menisci ( p < 0.05) (Fig. 4).
These results could also be verified in Western blotting
experiments, further proving the specificity of the immuno-
reaction. One band of 20 kDa that corresponds to endostatin
protein was stained in menisci and in traction tendons
(Fig. 5).
2.5. Endostatin/collagen XVIII is expressed in cultured
chondrocytes (C28/I2) but is not affected by growth factors
To elucidate if endostatin/collagen XVIII is also ex-
pressed by chondrocytes under in vitro conditions, we cul-
tured immortalized chondrocytes and measured endostatin/
collagen XVIII concentrations by a sensitive ELISA.
Chondrocytes cultured under normal oxygen pressure
released measurable amounts of endostatin/collagen XVIII
into their culture supernatants. Application of EGF, as an
example of a growth factor, or dexamethasone, as example
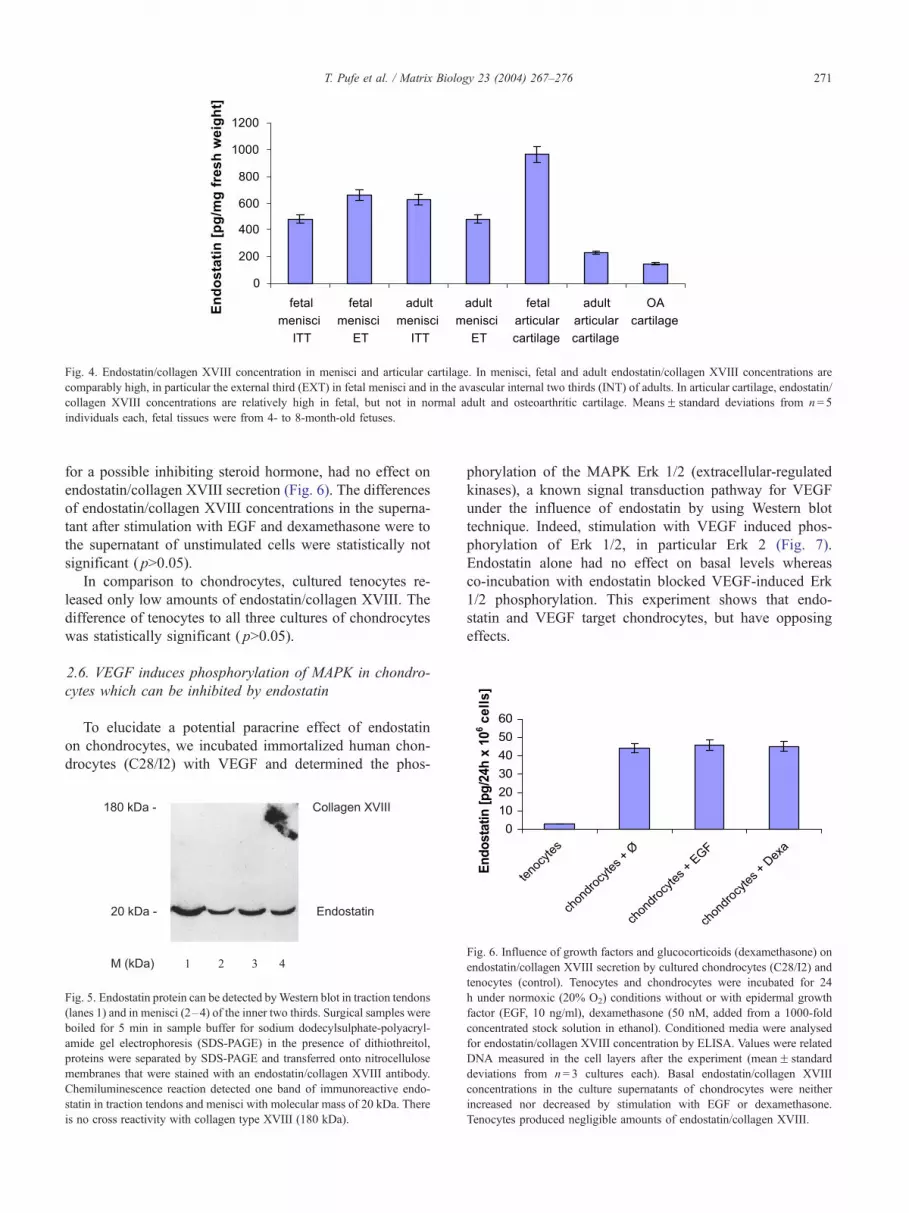
Fig. 4. Endostatin/collagen XVIII concentration in menisci and articular cartilage. In menisci, fetal and adult endostatin/collagen XVIII concentrations are
comparably high, in particular the external third (EXT) in fetal menisci and in the avascular internal two thirds (INT) of adults. In articular cartilage, endostatin/
collagen XVIII concentrations are relatively high in fetal, but not in normal adult and osteoarthritic cartilage. MeansF standard deviations from n= 5
individuals each, fetal tissues were from 4- to 8-month-old fetuses.
T. Pufe et al. / Matrix Biology 23 (2004) 267–276 271
for a possible inhibiting steroid hormone, had no effect on
endostatin/collagen XVIII secretion (Fig. 6). The differences
of endostatin/collagen XVIII concentrations in the superna-
tant after stimulation with EGF and dexamethasone were to
the supernatant of unstimulated cells were statistically not
significant ( p>0.05).
In comparison to chondrocytes, cultured tenocytes re-
leased only low amounts of endostatin/collagen XVIII. The
difference of tenocytes to all three cultures of chondrocytes
was statistically significant ( p>0.05).
2.6. VEGF induces phosphorylation of MAPK in chondro-
cytes which can be inhibited by endostatin
To elucidate a potential paracrine effect of endostatin
on chondrocytes, we incubated immortalized human chon-
drocytes (C28/I2) with VEGF and determined the phos-
Fig. 5. Endostatin protein can be detected byWestern blot in traction tendons
(lanes 1) and in menisci (2–4) of the inner two thirds. Surgical samples were
boiled for 5 min in sample buffer for sodium dodecylsulphate-polyacryl-
amide gel electrophoresis (SDS-PAGE) in the presence of dithiothreitol,
proteins were separated by SDS-PAGE and transferred onto nitrocellulose
membranes that were stained with an endostatin/collagen XVIII antibody.
Chemiluminescence reaction detected one band of immunoreactive endo-
statin in traction tendons and menisci with molecular mass of 20 kDa. There
is no cross reactivity with collagen type XVIII (180 kDa).
phorylation of the MAPK Erk 1/2 (extracellular-regulated
kinases), a known signal transduction pathway for VEGF
under the influence of endostatin by using Western blot
technique. Indeed, stimulation with VEGF induced phos-
phorylation of Erk 1/2, in particular Erk 2 (Fig. 7).
Endostatin alone had no effect on basal levels whereas
co-incubation with endostatin blocked VEGF-induced Erk
1/2 phosphorylation. This experiment shows that endo-
statin and VEGF target chondrocytes, but have opposing
effects.
Fig. 6. Influence of growth factors and glucocorticoids (dexamethasone) on
endostatin/collagen XVIII secretion by cultured chondrocytes (C28/I2) and
tenocytes (control). Tenocytes and chondrocytes were incubated for 24
h under normoxic (20% O2) conditions without or with epidermal growth
factor (EGF, 10 ng/ml), dexamethasone (50 nM, added from a 1000-fold
concentrated stock solution in ethanol). Conditioned media were analysed
for endostatin/collagen XVIII concentration by ELISA. Values were related
DNA measured in the cell layers after the experiment (meanF standard
deviations from n= 3 cultures each). Basal endostatin/collagen XVIII
concentrations in the culture supernatants of chondrocytes were neither
increased nor decreased by stimulation with EGF or dexamethasone.
Tenocytes produced negligible amounts of endostatin/collagen XVIII.

Fig. 7. VEGF induces the phosphorylation of the MAPK Erk 1/2 which is
suppressed by co-incubation with endostatin. Human chondrocytes (C-28/
I2) were stimulated for 20 min with VEGF and endostatin (both at 10 ng/
ml) alone or with a combination, lysed and analysed for Tyr-204
phosphorylated Erk 1 (44 kDa) and Erk 2 (42 kDa) by Western blotting
using a specific antibody. Equal amounts of proteins were used in the
Western blot, as shown by incubating membranes with an antibody against
non-activated Erk2 (lower panel).
T. Pufe et al. / Matrix Biology 23 (2004) 267–276272
2.7. Endostatin decreases NO production in C-28/I2
chondrocytes
Since OA is associated with an increased synthesis of
NO, we measured the influence of endostatin on NO
production in the C-28/I2 cell line (Fig. 8). In fact, as
measured with the Griess reaction, endostatin reduces the
NO release into the culture medium after 24 h.
Fig. 8. Endostatin increases NO production of chondrocytes moderately.
C28/I2 cells were exposed to endostatin (10 ng/ml) for 24 h, and aliquots of
the conditioned medium assayed for nitrate/nitrite (NO) by the Griess
reaction; values are the mean and S.D. of n= 6 experiments; p= 0.01 versus
control.
3. Discussion
A mircosequence analysis of endostatin revealed its
identity to the C-terminal fragment of the NC1 domain
of collagen XVIII (O’Reilly et al., 1997). Collagen XVIII/
endostatin is expressed in the developing and postnatal
eye, along with collagens II and IX (Fukai et al., 2002).
Mice lacking collagen XVIII and its proteolytically derived
product endostatin show delayed regression of blood
vessels in the vitreous along the surface of the retina after
birth (Fukai et al., 2002). These results suggest that
collagen XVIII/endostatin is critical for normal blood
vessel formation of the eye and might be involved in the
development of other avascular tissues. In some aspects,
the structure of cartilage is similar to the vitreous, which
contains cartilage-specific forms of collagen II and IX
(Fukai et al., 2002).
This study has shown that endostatin/collagen XVIII an
endogenous inhibitor of angiogenesis is expressed in human
cartilage and fibrocartilage. In the articular cartilage of
adults, endostatin/collagen XVIII concentrations were sig-
nificantly higher than in the fetal epiphysis. Since material
of young human donors is rare, we studied the temporal
expression of endostatin/collagen XVIII during childhood in
a rat model. In the rat, downregulation of endostatin/
collagen XVIII occurred soon after birth. Since in this stage
the distal femur was already ossified with a well-developed
articular cartilage, this time point might correspond to the
end of the first decade in humans.
In contrast to adult articular cartilage, there was no
significant decrease in the endostatin/collagen XVIII con-
centrations of the menisci. From adult articular cartilage,
other antiangiogenic factors such as thrombospondin (Good
et al., 1990), metastatin (Liu et al., 2001), and troponin-1
(Moses et al., 1999) have been identified. The presence of
these molecules might explain why lower endostatin con-
centrations are necessary to maintain avascularity in artic-
ular cartilage compared to meniscus fibrocartilage. Another
explanation might be that the menisci are not completely
avascular. In the external part adjacent to the vascularized
joint capsule, there are some vessels in the meniscus tissue.
A strong physiologic inhibitory stimulus might be necessary
to prevent ingrowth of these vessels in the inner two thirds
of the menisci.
Avascularity is an important feature of adult articular
cartilage and fibrocartilage. The extracellular matrix (ECM)
of both tissues is functionally adapted to bear high com-
pressive and tensile forces. Ingrowing vascular elements
would impair the material properties of this tissue-specific
ECM, therefore a protection by angiogenesis inhibiting
molecules might be necessary to maintain the tissue homeo-
stasis of cartilage and fibrocartilage.
High endostatin/collagen XVIII levels in developing
cartilage and fibrocartilage might reflect the angiogenic
activity of fetal tissue where angiogenesis is balanced by
inhibiting and stimulatory peptides. This leads to the
question why angiogenesis inhibitors should be present
in tissues that are angiogenic. One possibility is that the
proteolytic activity that accompanies fetal growth, may
also mobilize circulating angiogenesis inhibitors from
precursor proteins that are not antiangiogenic them-
selves—a mechanism that has been postulated for tumor
angiogenesis (Mentlein and Held-Feindt, 2003). A second
possibility is that endostatin has a physiological function in
fetal development to inhibit vascular overgrowth which

T. Pufe et al. / Matrix Biology 23 (2004) 267–276 273
might be induced by high levels of angiogenic factors such
as VEGF. In both cartilage and fibrocartilage of fetuses,
we observed also strong immunostaining for VEGF. In
epiphyseal and growth plate cartilage, endostatin/collagen
XVIII and VEGF expression was spatially separated but
no such separation was found in the menisci. In the adult
articular cartilage and in the menisci, VEGF was down-
regulated but endostatin could still be measured.
The mechanism by which endostatin exerts its effects is
unknown, but several possibilities are emerging (Takahashi
et al., 2003). Three types of interactions have been sug-
gested by which endostatin may alter the behavior of
endothelial cells.
First, it may interact with specific endostatin receptors
located on endothelial cells, which would result in activation
of intracellular signalling pathways and altered gene expres-
sion. Cell surface glypicans are low-affinity endostatin
receptors (Karumanchi et al., 2001).
Second, endostatin may bind to extracellular proteins
and alter their ability to influence endothelial cell behavior.
For example, endostatin inhibits adhesion of endothelial
cells to collagen I via alpha(2)beta(1) integrin (Furumatsu
et al., 2002).
Third, endostatin may bind to other cell surface receptors
and thereby modulate their signaling. A recent study has
shown that endostatin blocks VEGF-mediated signalling via
direct interaction with the VEGF receptor 2 (KDR) (Kim et
al., 2002). Recently, we and others have identified the
VEGFR-2 on the surface of hypertrophic and osteoarthritic
chondrocytes (Petersen et al., 2002; Pufe et al., 2001a,b;
Pfander et al., 2001; Enomoto et al., 2003). In human
umbilical vein endothelial cells endostatin-inhibited VEGF
induced ERK and p38 MAPK activation which is a down-
stream event of the VEGFR-2/KDR signaling (Kim et al.,
2002). The results of the present study show that endostatin
interferes with the same signaling cascade in human chon-
drocytes. In tumor cells and endothelial cells, endostatin
inhibits activation of metalloproteinase (MMP) 1 and 2
(Kim et al., 2000). In cartilage and in C28/I-2 chondrocytes,
VEGF induces MMP-1 and -3 and -13 (Pufe et al., 2004a,b).
MMPs are known to be crucial for degrading extracellular
matrix components (Westermarck and Kahari, 1999). In
C28/I-2 chondrocytes, endostatin is able to reduce NO
release. This is remarkable concerning the inhibitory effect
of NO on matrix synthesis in articular cartilage (Studer et
al., 1999). Since endostatin/collagen XVIII is downregu-
lated in osteoarthritic cartilage, this molecule might play a
role for the pathogenesis of this degenerative disease. More
research is needed to elucidate possible effects of endostatin
on chondrocytes.
In situ hybridization and immunostaining experiments
using fetal and adult tissue samples demonstrated that
collagen XVIII, the precursor for endostatin, is ubiquitous-
ly located in basement membrane zones (Halfter et al.,
1998; Fukai et al., 2002; Miosge et al., 2003), its expres-
sion patterns almost identical to that of laminin. Interest-
ingly, typical integral components of basement membranes
such as laminin have been identified and immunolocalized
in cartilage (Durr et al., 1996) and in fibrocartilage (Hayes
et al., 2001).
Since formation of cartilaginous and fibrocartilaginous
tissue is a functional adaptation to compressive and shear-
ing forces (Pauwels, 1960; Altmann, 1964), it seemed
likely that the avascular nature of fibrocartilage may also
be influenced by mechanical stimuli. In a previous in vitro
study, we analyzed tendon fibroblast cultures which were
incubated with intermittent hydrostatic pressure to show
that mechanical factors are able to stimulate fibroblasts to
release factors that inhibit endothelial cell proliferation
(Pufe et al., 2003). Those experiments demonstrated clear-
ly that mechanical factors influence endothelial growth not
only in a direct way but also via humoral factors in a
paracrine manner. Increased endostatin/collagen XVIII
concentrations in the supernatant of fibroblasts cultured
under the influence of intermittent hydrostatic pressure
might explain this observation. In contrast to the influence
of mechanical factors, the expression of endostatin/colla-
gen XVIII is unaffected by inflammatory growth factors or
antiinflammatory glucocorticoids.
In conclusion, our study gives evidence that the differ-
ential spatial expression of endostatin expression plays an
important role in the organization of blood vessels in
cartilage. This peptide appears to be one of the antiangio-
genic factors regulating the vascular status of cartilage by
counteracting effects of VEGF. More research is needed to
elucidate the effects of endostatin on chondrocytes.
4. Experimental procedures
4.1. Human tissues
The knee joints of human fetuses (18, 23, 26, 32, and 34
weeks post gestation) were harvested during routine autop-
sy, the femora and the menisci were dissected, fixed in 4%
formalin and prepared for immunohistochemistry. The knee
of the contralateral side was obtained for the biochemical
analysis. The menisci and the epiphyseal cartilage were
immediately frozen in liquid nitrogen.
Articular cartilage and menisci from adults were obtained
from body donors (23–76 years). The material was divided;
one part was fixed in 4% formalin, the other part was frozen
in liquid nitrogen. Before freezing, the menisci were sharply
divided into two parts (the internal two thirds = avascular
zone, the external one third = vascular zone). The study was
approved by the ethical commission of the Medical Faculty
of the Christian-Albrechts-University of Kiel.
4.2. Rat model
To investigate endostatin/collagen XVIII expression in
cartilage during childhood, a rat model was used. Eighteen

T. Pufe et al. / Matrix Biology 23 (2004) 267–276274
fetal, postnatal (1 and 2 days old) and adult Wistar rats were
killed by CO2. Then the knee joints were harvested, fixed in
4% formaline and prepared for immunohistochemistry.
These procedures are in accordance with the National
Institute of Health guidelines for the use of laboratory
animals. A permission of the local government animal rights
protection authorities was not required.
4.3. Cell cultures
The immortalized human chondrocyte cell line, C-28/I2
(Goldring et al., 1994), was cultured in Dulbecco’s modified
Eagle’s medium (DMEM)/Ham’s F12 (1/1, v/v) containing
10% fetal calf serum (FCS) and passaged using trypsin-
EDTA solution at >95% confluency every 5 to 6 days.
These cells express type II collagen, aggrecan, and other
markers of the differentiated phenotype and have been used
to study the regulation of gene expression and signalling in
response to cytokines and other factors (Loeser et al., 2000;
Osaki et al., 2003; Parker et al., 2003; Tan et al., 2003; Grall
et al., 2003).
Culture and stimulation of rat tenocytes: Achilles tendons
were dissected from postnatal (2–5 days old) rats, cut into
small pieces of less than 0.5 mm, transferred into a small
volume (2 ml per 10 cm2 culture flasks; TPP, Switzerland)
of Dulbecco’s modified Eagle’s medium (DMEM) with
50% foetal calf serum (FCS), and left for 24 h at 37 jCwithout enzymatic digestion. Then 3 ml DMEM plus 50%
FCS were added, and tissue pieces left for another 24 h at 37
jC. During this time, tenocytes migrate out of the tissue and
adhere to the bottom of the culture dish. After removing the
medium with the remaining tissue pieces floating on top,
fresh DMEM plus 10% FCS was added, and the cells were
cultivated for 96 h at 37 jC. For subculture, cells were
detached by a short treatment (about 20 s) with trypsin
(0.1%; 1:250, Gibco, Paisley, UK). 106 cells were seeded
into fresh dishes, and cultivated for 24 h in DMEM plus
10% FCS.
4.4. Immunohistochemistry
For immunohistochemistry, the samples were embedded
in paraffin, sectioned in a longitudinal direction (5 Am),
irradiated at 750 W in a microwave oven with 3% hydrogen
peroxide in 0.01 M sodium citrate buffer, pH 6.0 (twice for
5 min), dewaxed, immunostained with anti-endostatin/col-
lagen XVIII (1:40 in Tris-buffered saline, 60 min; AB 1878
anti human endostatin/collagen XVIII polyclonal antibody,
Chemicon, USA or for rat 1:20 in Tris-buffered saline, code
number 1092, courtesy of Prof. Dr. Rupert Timpl), anti-
VEGF (1:40 in Tris-buffered saline, 60 min; sc7269 mouse
monoclonal IgG2a, Santa Cruz Biotechnology, CA, USA),
anti-collagen type II (1:10; CIIC1mouse monoclonal anti-
body; Developmental studies hybridoma bank) or for dem-
onstration of blood vessels by anti-factor VIII (1:200;
DAKO rabbit polyclonal antibody) followed by biotinylated
secondary antibodies and a peroxidase-labeled streptavi-
din–biotin staining technique; nuclei were counterstained
with hemalum.
4.5. In situ-hybridization
A 205-bp sac/hinc II fragment corresponding to the c-
terminal endostatin domain of collagen type XVIII was
used for hybridization (Sasaki et al., 1998). Non-radioac-
tively labeled sense and anti-sense RNAs were produced
in vitro with the digoxigenin (DIG) labeling kit (Roche
Diagnostics, Mannheim, Germany). Paraffin sections were
deparaffinized, rehydrated and pre-treated with proteinase
K, then incubated with the hybridization solution (50%
formamide, 5� SSC, 0.1 mg/Al yeast-tRNA) for 1 h at
37 jC. The RNA concentration for collagen type XVIII
was 50 ng digoxigenin labeled anti-sense probe in 50
Al hybridization solution per section. In control sections,
an equivalently labeled amount of sense-RNA was used
instead of anti-sense-RNA. Post-hybridization treatment
included washes with 1� SSC (2� 15 min at 50 jC),1� SSC (2� 15 min), 0.1� SSC (4� 15 min) both at
60 jC and finally PBS for 15 min at RT. Specimens
were subsequently incubated with a 1:60 dilution of anti-
DIG peroxidase-labeled antibody (Dakopats) in PBS for 1
h at RT.
4.6. Enzyme-linked immunosorbent assay (ELISA) and
Western blots
For ELISA, frozen tissue samples were crushed in an
achate mortar under liquid nitrogen, homogenized in 150
mM NaCl 20 mM Tris/HCl-buffer, pH 7.4; a soluble
fraction was obtained by centrifugation (48000� g, 60
min), and aliquots (100 Al) were analyzed by a sandwich
ELISA for endostatin/collagen XVIII (Chemicon, USA
Cyt274). Human recombinant endostatin (Chemicon,
USA) served as standard. For statistical analysis of the
ELISA results, the Dunnett’s test was used.
For Western blots, samples were reduced in the presence
of 10 mM dithiothreitol, proteins separated by sodium
dodecylsulphate-polyacrylamide gel electrophoresis (SDS-
PAGE; 10% gels), transferred onto nitrocellulose membranes
that were blocked and incubated with antibodies according to
standard techniques as described (Held-Feindt et al., 1999).
Signals were detected by chemoluminescence reaction
(ECL-Pus; Amersham-Pharmacia, Uppsala, Sweden).
4.7. Stimulation of chondrocytes
The immortalized human chondrocyte cell line, C-28/I2
(20,21) was used for in vitro examinations. 106 cells were
seeded into fresh dishes, and cultivated for 24 h in Dulbec-
co’s modified Eagle’s medium (DMEM/ Ham’s F12 (1:1))
plus 10% fetal calf serum (FCS). Then, the medium was
replaced by DMEM/ Ham’s F12 (1:1) (without FCS), and

T. Pufe et al. / Matrix Biology 23 (2004) 267–276 275
the cells exposed to different stimulators for 24 h. Condi-
tioned medium was withdrawn, and aliquots assayed for
endostatin/collagen XVIII content. The cells were washed
with phosphate-buffered saline, lysed, the DNA content was
measured fluorometrically with the CyQuant reagent (Mo-
lecular Probes, Eugene, OR, USA), and related to a standard
number of cells (counted with trypsinised cells (Pufe et al.,
2001b)).
Statistical significance was evaluated by the Dunnett’s
multiple comparisons test or the t-test.
4.8. Analysis of kinase phosphorylation
The C28/I2 chondrocytes were seeded in fresh dishes and
cultured for 48 h in DMEM/ HAM F12 (1:1) plus 10% FCS.
Then the medium was replaced with DMEM/ Ham’s F12
without FCS and the cells exposed to the stimulators for 1
h (VEGF and endostatin each 10ng/ml). For detection of
phosphorylated MAPK, cells were rinsed once in cold
phosphate-buffered saline (PBS), lysed with Triton-lysis-
buffer (50 mM Tris–HCl, pH 7.8, 100 mM NaCl, 2 mM
EDTA, 1% Triton X-100, 2 mM Na3VO4) and scraped off
by a rubber policeman. The lysates were mixed vigorously
(vortex mixer) and clarified in an Eppendorf centrifuge (15
min, 14,000� g, 4 jC). After protein determination from an
aliquot, samples with equal amounts were boiled in 50–200
Al SDS-PAGE sample buffer, separated by SDS-PAGE
(10%), transferred onto a polyvinylidene difluoride (PVDF)
membrane that was blocked with 5% BSA overnight or for
1 h. The blots were incubated in 1% BSA with anti-
phosphorylated extracellular-signal related kinases ERK 1/
2 (1:200, mouse monoclonal reacting with Tyr-204 phos-
phorylated ERK 1/2; Santa Cruz sc-7383, CA, USA),
followed (after washings) by horseradish peroxidase-labeled
anti-mouse or anti-rabbit IgG (1:30,000; DAKO, Glostrup,
Denmark) and visualized by enhanced chemiluminescence
(ECL system; Amersham). In some experiments, the blots
were stripped by washing with methanol (18 h 4 jC) and re-
probed with an antibody to ERK 2 (1:200; Santa Cruz sc-
1647).
4.9. Determination of nitrite/nitrate
Subconfluent C28/I2 chondrocytes (5� 105 cells) were
stimulated for 24 h in medium with 10% FCS, and nitrite /
nitrate oxidized from nitric oxide (NO) was measured
spectrophotometrically by the Griess reaction (Sigma-
Aldrich G-4410).
Acknowledgements
We wish to thank Inka Kronenbitter, Frank Lichte,
Marion Lorenzen, Miriam Lemmer, Sonja Seiter, Karin
Stengel and Regine Worm for their expert technical
assistance.
Part of this work was supported by a grant of the
Deutsche Forschungsgemeinschaft (DFG Pu 214/3-1 and Pe
873/2-1) and by a grant from the ‘‘Stiftung zur Forderung
der Medizinischen Forschung’’ of the Medical Faculty of
the University of Kiel. Dr. Goldring’s research is supported
by NIH grants AR-45378 and AG-22021.
References
Altmann, K., 1964. Zur kausalen Histiogenese des Knorpels. W. Roux’s
Theorie und experimentelle Wirklichkeit. Z. Anat. Entwickel. Ges. 37,
1–167.
Angionello, A.L., Sgadari, C., Taub, D.D., Liao, F., Farber, J.M., Miahesh-
wari, S., Kleinman, H.K., Reaman, G.H., Tosato, G., 1995. Human
recombinant interferone-inducible protein 10 is a potent inhibitor of
angiogenesis. J. Exp. Med. 182, 155–162.
Becker, R., Pufe, T., Kulow, S., Giessmann, N., Neumann, W., Mentlein,
R., Petersen, W., 2004. Expression of the vascular endothelial growth
factor (VEGF) during meniscus healing in a rabbit model. J. Bone Jt.
Surg., Br. (in press).
Brookes, M., Revell, W.J., 1998. Blood Supply of Bone. Springer, New
York.
Durr, J., Lammi, P., Goodman, S.L., Aigner, T., von der Mark, K., 1996.
Identification and immunolocalization of laminin in cartilage. Exp. Cell
Res. 222, 225–233.
Enomoto, H., Inoki, I., Komiya, K., Shiomi, T., Ikeda, E., Obata, K.,
Matsumoto, H., Toyama, Y., Okada, Y., 2003. Vascular endothelial
growth factor isoforms and their receptors are expressed in human
osteoarthritic cartilage. Am. J. Pathol. 162, 171–181.
Ferrara, N., 1999. Molecular and biological properties of vascular endo-
thelial growth factor. J. Mol. Med. 77, 527–543.
Fukai, N., Eklund, L., Marneros, A.G., Oh, S.P., Keene, D.R., Tamarkin,
L., Niemela, M., Ilves, M., Li, E., Pihlajaniemi, T., Olsen, B., 2002.
Lack of collagen XVII/endostatin results in eye abnormalities. EMBO J.
21, 1535–1544.
Furumatsu, T., Yamaguchi, N., Nishida, K., Kawai, A., Kunisada, T.,
Namba, M., Inoue, H., Ninomiya, Y., 2002. Endostatin inhibits adhe-
sion of endothelial cells to collagen I via alpha(2)beta(1) integrin, a
possible cause of prevention of chondrosarcoma growth. J. Biochem.
(Tokyo) 131, 619–626.
Gerber, H.P., Vu, T.H., Ryan, A.M., Kowalski, J., Werb, Z., Ferrara, N.,
1999. VEGF couples hypertrophic cartilage remodeling, ossification
and angiogenesis during endochondral bone formation. Nat. Med. 5,
623–628.
Goldring, M.B., Birkhead, J.R., Suen, L.-F., Yamin, R., Mizuno, S., Glo-
wacki, J., Arbiser, J.L., Apperley, J.F., 1994. Interleukin-1-h-modulated
gene expression in immortalized human chondrocytes. J. Clin. Invest.
94, 2307–2316.
Good, D.J., Polverini, P.J., Rastinejad, F., Le Beau, M.M., Lemons, R.S.,
Frazier, W.A., Bouck, N.P., 1990. A tumor suppressor inhibitor of
angiogenesis is immunologically and functionally indistinguishable
from a fragment of thrombospondin. Proc. Natl. Acad. Sci. U. S. A.
87, 6624–6628.
Grall, F., Gu, X., Tan, L., Cho, J.-Y., Inan, M.S., Pettit, A., Thamrongsak,
U., Choy, B.K., Manning, C., Akbarali, Y., Zerbini, L., Rudders, S.,
Goldring, S.R., Gravallese, E.M., Oettgen, P., Goldring, M.B., Lib-
ermann, T.A., 2003. Responses to the pro-inflammatory cytokines
interleukin-1 and tumor necrosis factor-a in cells derived from rheu-
matoid synovium and other joint tissues involve NF-nB-mediated
induction of the Ets transcription factor ESE-1. Arthritis Rheum.
48, 1249–1260.
Halfter, W., Dong, S., Schurer, B., Cole, G.J., 1998. Collagen XVIII is a
basement membrane heparan sulfate proteoglycan. J. Biol. Chem. 273,
25404–25412.

T. Pufe et al. / Matrix Biology 23 (2004) 267–276276
Hayes, A.J., Benjamin, M., Ralphs, J.R., 2001. Extracellular matrix in
development of the intervertebral disc. Matrix Biol. 20, 107–121.
Held-Feindt, J., Krisch, B., Mentlein, R., 1999. Molecular and functional
analysis of the somatostatin receptor subtype 2 (sst2) in human glioma
cells. Mol. Brain. Res. 64, 101–107.
Karumanchi, S.A., Jha, V., Ramchandran, R., Karihaloo, A., Tsiokas, L.,
Chan, B., Dhanabal, M., Hanai, J.I., Venkataraman, G., Shriver, Z.,
Keiser, N., Kalluri, R., Zeng, H., Mukhopadhyay, D., Chen, R.L., Land-
er, A.D., Hagihara, K., Yamaguchi, Y., Sasisekharan, R., Cantley, L.,
Sukhatme, V.P., 2001. Cell surface glypicans are low-affinity endostatin
receptors. Mol. Cell 7, 811–822.
Kim, Y.M., Jang, J.W., Lee, O.H., Yeon, J., Choi, E.Y., Kim, K.W., Lee,
S.T., Kwon, Y.G., 2000. Endostatin inhibits endothelial and tumor cel-
lular invasion by blocking the activation and catalytic activity of matrix
metalloproteinase. Cancer Res. 60, 5410–5413.
Kim, Y.M., Hwang, S., Kim, Y.M., Pyun, B.J., Kim, T.Y., Lee, S.T., Gho,
Y.S., Kwon, Y.G., 2002. Endostatin blocks vascular endothelial growth
factor-mediated signaling via direct interaction with KDR/Flk-1. J. Biol.
Chem. 277, 27872–27879.
Liu, N., Lapcevich, R.K., Underhill, C.B., Han, Z., Gao, F., Swart, G.,
Plum, S., Zhang, L., Green, S.J., 2001. Metastatin: a hyaluronan-bind-
ing complex from cartilage inhibits tumor growth. Cancer Res. 61,
1022–1028.
Loeser, R., Sadiev, S., Tan, L., Goldring, M.B., 2000. Integrin expression
by primary and immortalized human chondrocytes: evidence of a dif-
ferential role for a1h1 and a2h1 integrins mediating chondrocyte ad-
hesion to types II and VI collagen. Osteoarthr. Cartil. 8, 96–105.
Maione, T.E., Gray, G.S., Petro, J., Hunt, A.J., Donner, A.L., Bauer, S.I.,
Carson, H.F., Sharpe, R.J., 1990. Inhibition of angiogenesis by recom-
binant human platelet factor-4 and related peptides. Science 247, 77–79.
Mentlein, R., Held-Feindt, J., 2003. Angiogenesis factors in gliomas—a
new key to tumour therapy? Naturwissenschaften 90, 385–394.
Miosge, N., Simniok, T., Sprysch, P., Herken, R., 2003. The collagen type
XVIII endostatin domain is co-localized with perlecan in basement
membranes in vivo. J. Histochem. Cytochem. 51, 285–296.
Moses, M.A., Wiedeschain, D., Wu, I., Fernandez, C., Ghazizadh Lane,
W.S., Flynn, E., Sytkowski, A., Tao, T., Langer, R., 1999. Troponin I
is present in human cartilage and inhibits angiogenesis. Proc. Natl.
Acad. Sci. 96, 2645–2650.
O’Reilly, M.S., Holmgren, L., Shing, Y., Chen, C., Rosenthal, R.A., Moses,
M., Lane, W.S., Cao, Y., Sage, E.H., Folkman, J., 1994. Angiostatin: a
novel angiogenesis inhibitor that mediates the suppression of metastases
by a Lewis lung carcinoma. Cell 79, 315–328.
O’Reilly, M.S., Behm, T., Sing, Y., Fikai, N., Vasios, G., Lane, W.S.,
Flynn, E., Birkhead, J.R., Olsen, B.R., Folkman, J., 1997. Endostatin:
an endogenous inhibitor of angiogenesis and tumor growth. Cell 88,
277–285.
Osaki, M., Tan, L., Choy, B.K., Yoshida, Y., Auron, P.E., Cheah, K.S.E.,
Goldring, M.B., 2003. The TATA-containing core promoter of the type
II collagen gene (COL2A1) is the target of interferon-g-mediated inhi-
bition in human chondrocytes: requirement for Stat1a, Jak1, and Jak2.
Biochem. J. 369, 103–115.
Parker, W.L., Goldring, M.B., Philip, A., 2003. Endoglin is expressed on
human chondrocytes and forms a heteromeric complex with betaglycan
in a ligand and type II TGF-h receptor independent manner. J. Bone
Miner. Res. 18, 289–302.
Pauwels, F., 1960. Eine neue Theorie uber den Einfluß mechanischer Reize
auf die Differenzierung der Stutzgewebe. Zehnter Beitrag zur funktio-
nellen Anatomie und kausalen Morphologie des Stutzapparates. Z.
Anat. Entwicklungsgesch. 121, 478–515.
Petersen, W., Tillmann, B., 1995. Age-related blood and lymph supply of
the knee menisci. A cadaver study. Acta Orthop. Scand. 66, 308–312.
Petersen, W., Tsokos, M., Pufe, T., 2002. Expression of VEGF121 and
VEGF165 in hypertrophic chondrocytes of the human growth plate
and epiphyseal cartilage. J. Anat. 201, 153–157.
Pfander, D., Kortje, D., Zimmermann, R., Weseloh, G., Kirsch, T., Ges-
slein, M., Cramer, T., Swoboda, B., 2001. Vascular endothelial growth
factor in articular cartilage of healthy and osteoarthritic human knee
joints. Ann. Rheum. Dis. 60, 1070–1073.
Pufe, T., Petersen, W., Tillmann, B., Mentlein, R., 2001a. Expression of the
vascular endothelial growth factor in osteoarthritic cartilage. Arthritis
Rheum. 44, 1082–1088.
Pufe, T., Petersen, W., Tillmann, B., Mentlein, R., 2001b. The angiogenic
peptide vascular endothelial cell growth factor (VEGF) is expressed in
fetal and ruptured tendons. Virchows Arch. 439, 579–585.
Pufe, T., Petersen, W., Kurz, B., Tsokos, M., Tillmann, B., Mentlein,
R., 2003. Mechanical factors influence the expression of endosta-
tin—an inhibitor of angiogenesis—in tendons. J. Orthoptera Res. 21,
610–616.
Pufe, T., Lemke, A., Kurz, B., Petersen, W., Tillmann, B., Grodzinsky, A.J.,
Mentlein, R., 2004a. Mechanical overload induces VEGF in cartilage
discs via hypoxia-inducible factor. Am. J. Pathol. 164, 185–192.
Pufe, T., Harde, V., Petersen, W., Goldring, M.B.;, Tilmann, B., Mentlein,
R., 2004b. Vascular endothelial growth factor (VEGF) induces matrix
metalloproteinase expression in immortalized chondrocytes. J. Pathol.
202, 367–374.
Sasaki, T., Fukai, N., Mann, K., Gohring, W., Olsen, B.R., Timpl, R., 1998.
Structure, function and tissue forms of the C-terminal globular domain
of collagen XVIII containing the angiogenesis inhibitor endostatin.
EMBO J. 17, 4249–4256.
Studer, R., Jaffurs, D., Stefanovic-Racic, M., Robbins, P.D., Evans, C.H.,
1999. Nitric oxide in osteoarthritis. Osteoarthr. Cartil. 7, 377–379.
Takahashi, K., Saishin, Y., Saishin, Y., Silva, R.L., Oshima, Y., Oshima, S.,
Melia, M., Paszkiet, B., Zerby, D., Kadan, M.J., Liau, G., Kaleko, M.,
Connelly, S., Luo, T., Campochiaro, P.A., 2003. Intraocular expression
of endostatin reduces VEGF-induced retinal vascular permeability, neo-
vascularization, and retinal detachment. FASEB J. 17, 896–898.
Tan, L., Peng, H., Osaki, M., Choy, B.K., Auron, P.E., Sandell, L.J.,
Goldring, M.B., 2003. Egr-1 mediates transcriptional repression of
COL2A1 promoter activity by interleukin-1h. J. Biol. Chem. 278,
17688–17700.
Westermarck, J., Kahari, V.M., 1999. Regulation of matrix metalloprotei-
nase expression in tumor invasion. FASEB J. 13, 781–792.
Yamaguchi, N., Anaud-Apte, B., Lee, M., Sasaki, T., Fukai, N., Shapiro,
R., Que, I., Lowik, C., Timpl, R., Olsen, B.R., 1999. Endostatin inhibits
VEGF-induced endothelial cell migration and tumor growth indepen-
dently of zinc binding. EMBO J. 18, 4414–4423.




















