
Differential Effects of Chemical Sympathectomy on Expression and Activity of Tyrosine Hydroxylase...
8
Neurochemical Research, Vol. 24, No. 1, 1999, pp. 25-32 Differential Effects of Chemical Sympathectomy on Expression and Activity of Tyrosine Hydroxylase and Levels of Catecholamines and DOPA in Peripheral Tissues of Rats Minoru Kawamura, 1,2 Joan P. Schwartz, 1 Takuo Nomura, 1 Irwin J. Kopin, 1 David S. Goldstein, 1 Thanh-True Huynh, 1 Douglas R. Hooper, 1 Judith Harvey-White, 1 and Graeme Eisenhofer 1,3 (Accepted June 6, 1998) Tyrosine hydroxylase (TH) mRNA and activity and concentrations of 3,4-dihydroxyphenylalanine (DOPA) and catecholamines were examined as markers of sympathetic innervation and catechol- amine synthesis in peripheral tissues of sympathectomized and intact rats. Chemical sympathec- tomy with 6-hydroxydopamine (6-OHDA) markedly decreased norepinephrine and to a generally lesser extent TH activities and dopamine in most peripheral tissues (stomach, lung, testis, duode- num, pancreas, salivary gland, spleen, heart, kidney, thymus). Superior cervical ganglia, adrenals and descending aorta were unaffected and vas deferens showed a large 92% decrease in norepi- nephrine, but only a small 38% decrease in TH activity after 6-OHDA. Presence of chromaffin cells or neuronal cell bodies in these latter tissues, indicated by consistent expression of TH mRNA, explained the relative resistance of these tissues to 6-OHDA. Stomach also showed consistent expression of TH mRNA before, but not after 6-OHDA, suggesting that catecholamine synthesizing cells in gastric tissue are sensitive to the toxic effects of 6-OHDA. Tissue concentrations of DOPA were mainly unaffected by 6-OHDA, indicating that much of the DOPA in peripheral tissues is synthesized independently of local TH or sympathetic innervation. The differential effects of chem- ical sympathectomy on tissue catecholamines, DOPA, TH mRNA and TH activity demonstrate that these variables are not simple markers of sympathetic innervation or catecholamine synthesis. Other factors, including presence of neuronal cell bodies, parenchymal chromaffin cells, non-neu- ronal sites of catecholamine synthesis and alternative sources of tissue DOPA, must also be con- sidered when tissue catecholamines, DOPA and TH are examined as markers of sympathetic innervation and local catecholamine synthesis. KEY WORDS: Tyrosine hydroxylase; sympathetic nervous system; 6-hydroxydopamine; norepinephrine; do- pamine; mRNA. INTRODUCTION The sympathetic nervous system plays an important role in the regulation of blood pressure, glucose, sodium 1 Clinical Neuroscience Branch, National Institute of Neurological Dis- orders and Stroke, National Institutes of Health, Bethesda MD 20892. 2 Institute of Bio-Active Science, Nippon Zoki Pharmaceutical Co., Ltd. Hyogo 673-14, Japan. and other key physiological and metabolic processes, and alterations in sympathoneural function occur in many disease states. Experimental and clinical evalua- 3 Address reprint requests to: Graeme Eisenhofer, Building 10, Room 4D20, National Institutes of Health, 10 Center Dr. MSC 1424, Be- thesda, MD 20892-1424. Ph (301) 496-8925; Fax (301) 402-0180; E-mail: [email protected] 25 0364-3190/98/0100-0025$16.00/0 C 1999 Plenum Publishing Corporation
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Differential Effects of Chemical Sympathectomy on Expression and Activity of Tyrosine Hydroxylase...

Neurochemical Research, Vol. 24, No. 1, 1999, pp. 25-32
Differential Effects of Chemical Sympathectomy onExpression and Activity of Tyrosine Hydroxylase and Levelsof Catecholamines and DOPA in Peripheral Tissues of Rats
Minoru Kawamura,1,2 Joan P. Schwartz,1 Takuo Nomura,1 Irwin J. Kopin,1 David S.Goldstein,1 Thanh-True Huynh,1 Douglas R. Hooper,1 Judith Harvey-White,1
and Graeme Eisenhofer1,3
(Accepted June 6, 1998)
Tyrosine hydroxylase (TH) mRNA and activity and concentrations of 3,4-dihydroxyphenylalanine(DOPA) and catecholamines were examined as markers of sympathetic innervation and catechol-amine synthesis in peripheral tissues of sympathectomized and intact rats. Chemical sympathec-tomy with 6-hydroxydopamine (6-OHDA) markedly decreased norepinephrine and to a generallylesser extent TH activities and dopamine in most peripheral tissues (stomach, lung, testis, duode-num, pancreas, salivary gland, spleen, heart, kidney, thymus). Superior cervical ganglia, adrenalsand descending aorta were unaffected and vas deferens showed a large 92% decrease in norepi-nephrine, but only a small 38% decrease in TH activity after 6-OHDA. Presence of chromaffincells or neuronal cell bodies in these latter tissues, indicated by consistent expression of TH mRNA,explained the relative resistance of these tissues to 6-OHDA. Stomach also showed consistentexpression of TH mRNA before, but not after 6-OHDA, suggesting that catecholamine synthesizingcells in gastric tissue are sensitive to the toxic effects of 6-OHDA. Tissue concentrations of DOPAwere mainly unaffected by 6-OHDA, indicating that much of the DOPA in peripheral tissues issynthesized independently of local TH or sympathetic innervation. The differential effects of chem-ical sympathectomy on tissue catecholamines, DOPA, TH mRNA and TH activity demonstratethat these variables are not simple markers of sympathetic innervation or catecholamine synthesis.Other factors, including presence of neuronal cell bodies, parenchymal chromaffin cells, non-neu-ronal sites of catecholamine synthesis and alternative sources of tissue DOPA, must also be con-sidered when tissue catecholamines, DOPA and TH are examined as markers of sympatheticinnervation and local catecholamine synthesis.
KEY WORDS: Tyrosine hydroxylase; sympathetic nervous system; 6-hydroxydopamine; norepinephrine; do-pamine; mRNA.
INTRODUCTION
The sympathetic nervous system plays an importantrole in the regulation of blood pressure, glucose, sodium
1 Clinical Neuroscience Branch, National Institute of Neurological Dis-orders and Stroke, National Institutes of Health, Bethesda MD 20892.
2 Institute of Bio-Active Science, Nippon Zoki Pharmaceutical Co.,Ltd. Hyogo 673-14, Japan.
and other key physiological and metabolic processes,and alterations in sympathoneural function occur inmany disease states. Experimental and clinical evalua-
3 Address reprint requests to: Graeme Eisenhofer, Building 10, Room4D20, National Institutes of Health, 10 Center Dr. MSC 1424, Be-thesda, MD 20892-1424. Ph (301) 496-8925; Fax (301) 402-0180;E-mail: [email protected]
250364-3190/98/0100-0025$16.00/0 C 1999 Plenum Publishing Corporation

26 Kawamura et al.
tion of sympathetic function and innervation rely im-portantly on assessments of synthesis, storage or releaseof norepinephrine, the sympathetic neurotransmitter (1).
With the exceptions of adrenomedullary productionof epinephrine and renal production of dopamine fromcirculating 3,4-dihydroxyphenylalanine (DOPA), mostof the catecholamines in peripheral tissues are generallythought to be synthesized in sympathetic nerves. Con-centrations of norepinephrine in extra-adrenal peripheraltissues therefore are commonly used to indicate thestatus of local sympathetic innervation. Tyrosine hy-droxylase (TH), the rate-limiting enzyme in catechola-mine synthesis, is also widely considered to be localizedin the periphery to catecholamine-producing sympatheticnerves and the adrenal medulla. The presence of TH inextra-adrenal peripheral tissues therefore provides an-other potential marker of sympathetic innervation; how-ever, due to lack of sufficiently sensitive assays of THactivity, such analyses are typically restricted to non-quantitative immunohistochemical procedures.
Recent reports have demonstrated catecholamineproduction in a variety of cell types in the peripheryother than chromaffin cells and sympathetic nerves, withTH more widely distributed than previously believed (1-6). These findings raise new questions about the mean-ing of peripheral tissue catecholamines and TH asindices of sympathetic innervation. To examine this is-sue, we applied a sensitive assay developed recently formeasurements of TH activity in peripheral tissues (7), toassess the influence of chemical sympathectomy with 6-hydroxydopamine (6-OHDA) on tissue TH. Changes intissue concentrations of DOPA, the immediate productof tyrosine hydroxylation, were also examined as an-other index of TH activity and compared with changesin concentrations of catecholamines. Additionally, sincetranscription and translation to gene product take placein cell bodies of sympathetic nerves (i.e., ganglia in thesympathetic chain), away from nerve endings, we alsoexamined peripheral tissues for the presence of THmRNA as a possible marker of TH in non-sympatho-neuronal cells.
EXPERIMENTAL PROCEDURE
Animals. Twenty-six male Sprague-Dawley rats (from Taconic Ger-mantown, NY) weighing 245-325 g were used in this study which wasapproved by the MINDS Animal Care and Use Committee. Animalswere provided food and water ad libitum before and during studies.
Materials. 6-OHDA hydrobromide was from Sigma ChemicalCompany, (St. Louis, MO). L-tyrosine hydrochloride, dihydropteridinereductase (55 U/mg protein, EC 1.6.99.7), NADPH, DL-6-methylte-trahydropterine dihydrochloride, m-hydroxybenzylhydrazine dihy-
drochloride (Sigma Chemical Company), 3-iodo-L-tyrosine (AldrichChemical Co., Milwaukee, WI) and bovine liver catalase (65,000U/mg protein, EC 1.11.1.6, Boehringer Mannheim, Indianapolis, IN)were used for TH assays. Norepinephrine bitartrate, dopamine hydro-chloride, DOPA, A-methyl DOPA, 3,4-dihydroxybenzylamine hydrob-romide and octanesulfonic acid (Sigma Chemical Company) were usedfor HPLC assays. Guanidine hydrochloride (ICN Biomedicals Inc.,Aurora, OH) was used for extraction of RNA. GeneAmp ThermostablerTth Reverse Transcriptase RNA PCR kit (Perkin Elmer Cetus, Nor-walk, CT) was used in the reverse transcription polymerase chain re-action (RT-PCR) for TH mRNA. Other reagents were analytical grade.
Treatment with 6-OHDA. 6-OHDA was dissolved in saline con-taining ascorbic acid and EDTA (0.1 mg/ml each) immediately beforeinjection. Ten rats were treated with 6-OHDA to destroy sympatheticnerve terminals and 16 other rats received an equivalent volume ofvehicle. 6-OHDA or vehicle was injected intravenously into animalsvia the tail vein with 3 consecutive injections of 10 mg/kg, 60 mg/kgand 60 mg/kg made at 2-day intervals.
Isolation of Peripheral Tissues. Adrenal glands, superior cervicalganglia, vas deferens, salivary gland, spleen, heart, duodenum, pan-creas, stomach, kidney, lung, thymus, testis and abdominal aorta wereharvested from animals under sodium pentobarbital anesthesia, 2 daysafter the final dose of 6-OHDA or vehicle. Isolated tissues were frozenimmediately on dry ice and stored at -80°C.
Preparation of Tissue Samples for TH mRNA. Extraction of RNAfrom peripheral tissues was performed using guanidine hydrochloride(8). Tissues were homogenized in 2.25 ml (or 300 M1 for tissues lessthan 100 mg wet weight) of 6 M guanidine hydrochloride, 20 mMsodium acetate, 1 mM dithiothreitol (pH 5.0). Homogenates were cen-trifuged (10,000 rpm X 10 min) at 4°C, and 1.25 ml (or 165 Ml) ofethanol was added to the supernatant and RNA was precipitated for60 min at -20°C. The precipitation was repeated once. RNA wasdissolved in 1 ml (or 400 Ml) of 200 Mg/ml proteinase K in 30 mMTris (pH 7.4), 100 mM sodium chloride, 1 mM EDTA, 0.5% sodiumdodecyl sulfate, and incubated for 1 hour at 37°C. After incubation,the RNA was extracted with phenolxhloroform (1:1) and precipitatedwith ethanol at -20°C.
Preparation of Tissue Samples for Catechol Content or TH Ac-tivity. Tissues were weighed frozen and homogenized in at least 4volumes of ice-cold 0.05 M sodium acetate buffer (pH 6.0) for assaysof TH activity or 0.4 M perchloric acid for assays of catechol contents.Homogenates were centrifuged (3000 rpm X 10 min) at 4°C and su-pernatants were stored at -80°C.
Reverse Transcription (RT) Polymerase Chain Reaction (PCR)for TH mRNA. The specific primers for TH were Upstream:GGCTGTGGTATTTGAGGAGA (nucleotides 221-240 of exon 2)and Downstream: AGGTGGTGACACTTGTCCA (nucleotides 532-550 of exon 3), with the expected product 330 bp. The RNA reversetranscription was performed at 70°C for 15 min using a reaction mix-ture I (20 Ml) that consisted of 10 mM Tris-HCl (pH 8.3), 90 mMKC1, 1.0 mM MnCl2, 0.2 mM dNTPs, 5 U of rTth DNA polymerase,15 pmol of Downstream primer and 250 ng of total RNA sample.After the RNA reverse transcription, the reaction mixture I was cooledto 4°C. The reaction mixture II (80 M1), consisting of 5% (v/v) glycerol,10 mM Tris-HCl (pH 8.3), 100 mM KC1, 0.05% (w/v) Tween 20, 0.75mM EGTA, 1.88 mM MgCl2 and 15 pmol of Upstream primer, wasadded to the reaction mixture I (20 M1) for the amplification of cDNA.The PCR reaction was performed with the first step (2 min at 95°C),followed by 40 cycles of melting step (1 min at 95°C) and anneal-ing/extending step (1 min at 60°C), and by the final step (7 min at60°C) using the total reaction mixture (I+11: 100 Ml). The final productwas separated on a 2% agarose gel by electrophoresis.

Tyrosine Hydroxylase and Sympathectomy 27
RT PCR for Cyclophillin. The specific primers for cyclophillinwere Upstream: TGGACCAAACACAAATGGTT (nucleotides 351-370 of exon 4) and Downstream: TGATCTTCTTGCTGGTCTT (nu-cleotides 493-551 of exon 5). The RNA reverse transcription and PCRreaction was performed using the same samples, reaction mixtures andmethod as those of TH, except for the primers. The expected productwas 161 bp.
TH Activity Assay. Assay of TH activity was by the method ofHooper et al. (7). The incubation mixture (200 M1) consisted of 100Ml of appropriately diluted tissue sample preparation and 100 M1 of 0.5M acetate buffer (pH 6.0), 200 MM L-tyrosine, 1.25 mM m-hydrox-ybenzylhydrazine, 1 mM DL-6-methyltetrahydropterine dihydrochlor-ide, 19.5 X 103 U/ml catalase, 3.8 U/ml dihydropteridine reductase, 1mM NADPH and 0.4 M glycerol. Blanks included the addition of theTH inhibitor, 3-iodo-L-tyrosine (0.45 mM). The mixture was incubatedat 37°C for 10 min and the incubation was terminated by adding 100M1 of 0.4 M perchloric acid. Dihydroxybenzylamine and A-methylDOPA (40 M1 = 12 pmol each, as an internal standard), 500 Ml ofTris-EDTA buffer (pH 8.6) and 10 mg of alumina were added to theincubates (300 Ml) and mixed for 30 min to extract the DOPA. Afterwashing the alumina, catechols were eluted with 100 Ml of a mixture of0.16 M acetic acid-0.04 M phosphoric acid and 90 Ml of the eluent wasinjected onto the HPLC system. TH activity (fmol/mg wet weight tis-sue/min) was calculated using the formula; TH activity = (DOPAsample
- DOPAblank)/(Weight of tissue • Time of incubation)Tissue Catechol Assay. The catechols were extracted from super-
natants of tissue homogenates onto alumina by a previously describedmethod (9). Dihydroxybenzylamine (20 Ml = 12 pmol, as a internalstandard), Tris-EDTA buffer (pH 8.6, 500-900 Ml) and 10 mg of alu-mina were added to the appropriate amount of sample (10-400 Ml) orto catechol standards (5.07 pmol DOPA, 1.48 pmol norepinephrine,1.63 pmol dopamine), and mixed for 30 min to extract catechols. Afterwashing the alumina, catechols were eluted with 100 Ml of a mixtureof 0.18 M acetic acid-0.02 M phosphoric acid and 90 M1 of the eluentwas injected onto the HPLC apparatus.
HPLC with Electrochemical Detection. HPLC systems includeda Model 510 solvent delivery system and a refrigerated autosampler(Waters Associates, Milford, MA). A 5-Mm particle size Axxi-chromC18 reversed-phase column (250 X 4.6 mm I.D., Axxiom Chroma-tography, Moorpark, CA) was used for chromatographic separations.Column temperature was maintained at 18°C using a model 1166 re-frigerated water circulator (PolyScience, Nile, IL). A Model 5100Acoulometric detector with a triple electrode system (EnvironmentalScience Associates, Bedford, MA) was used to quantify the catecholseluted from the column. The first electrode in the Model 5021 con-ditioning cell was set at a potential of +0.35 V. The second andthird electrodes in the Model 5011 analytical cell were set at poten-tials of +0.15 V and -0.35 V. The output from the third electrodewas recorded with a Macintosh SE computer (Apple Computer, Cu-pertino, CA) and Dynamax Method Manager hardware and softwarepackage (version 1.4; Rainin Instruments, Woburn, MA). The mobilephase (0.1 M NaH2PO4, 0.13 M EDTA, 0.23 mM octane sulfonate,0.7-1.0% acetonitrile, pH 3.29 for catechol assay and 0.1 MNaH2PO4, 0.13 M EDTA, 0.22 mM octane sulfonate, 1.5% aceton-itrile, pH 3.30 for TH activity assay) was filtered through a 0.22 Mmpore size filter, and pumped through the chromatographic system ata rate of 1.0 ml/min.
Statistical Analysis. Results are presented as means ± SEM. Dif-ferences in tissue concentrations of catechols or TH activities betweenvehicle- and 6-OHDA-treated groups were analyzed using Student's ttest. Differences in changes of tissue concentrations of catechols andactivities of TH among individual tissues of 6-OHDA-treated animals
compared to mean values in vehicle-treated animals were analyzed byANOVA. Post hoc tests were by the Scheffe F-test.
RESULTS
TH mRNA. TH mRNA was consistently detected asa bright signal by RT-PCR in superior cervical gangliaand adrenals, and was also more faintly but still consis-tently detected in abdominal aorta and vas deferens ofboth vehicle-treated and 6-OHDA-treated rats (Fig. 1,Table I). TH mRNA was also consistently detected instomach of vehicle-treated rats, but not in 6-OHDA-treated animals. TH mRNA was inconsistently detectedin lung, testis and duodenum and was undetectable insalivary gland, spleen, heart, kidney and thymus of ve-hicle-treated animals. Thus, in these latter tissues no at-tempt was made to examine for the presence of THmRNA in 6-OHDA-treated animals. Poor quality of ex-tracted mRNA in pancreas (presumably due to the highconcentration RNAase in this tissue) made reliable de-tection of TH mRNA in this tissue difficult.
Tissue Norepinephrine Concentrations. Norepi-nephrine concentrations in tissues of vehicle-treated ratsvaried widely from 39 ± 10 pmol/g wet tissue weightin abdominal aorta to 1,580,000 ± 310,000 pmol/g inadrenal gland (Table II). 6-OHDA-treated animals hadmarkedly decreased tissue norepinephrine concentrationsin all tissues except for the adrenal gland, superior cer-vical ganglia and abdominal aorta. Tissue norepineph-rine concentrations were maintained at 92% of theconcentrations in vehicle-treated animals in adrenalgland, at 97% in superior cervical ganglia and at 74%in abdominal aorta. Decreased norepinephrine concen-trations in all other tissues varied from 2% those of con-trol concentrations in testis to 23% in kidneys.
Tissue Dopamine Concentrations. Dopamine con-centrations in tissues of vehicle-treated rats varied from5 ± 1 pmol/g wet tissue weight in testis to 94,600 ±18,300 pmol/g in adrenal gland (Table II). After 6-OHDA, significant decreases in dopamine concentra-tions were observed in vas deferens, stomach, lung, du-odenum, pancreas, salivary gland, spleen, heart, kidneyand thymus. In contrast, dopamine concentrations in ad-renal gland, aorta, superior cervical ganglia and testistended to increase compared to those of vehicle-treatedanimals. Tissue dopamine concentrations after 6-OHDAtreatment (as a % of values in vehicle-treated animals)were maintained at relatively higher (P < 0.05) concen-trations than those of norepinephrine in stomach (56%vs 8%), lung (17% vs 6%), testis (203% vs 2%), duo-denum (24% vs 5%), pancreas (66% vs 15%), kidney
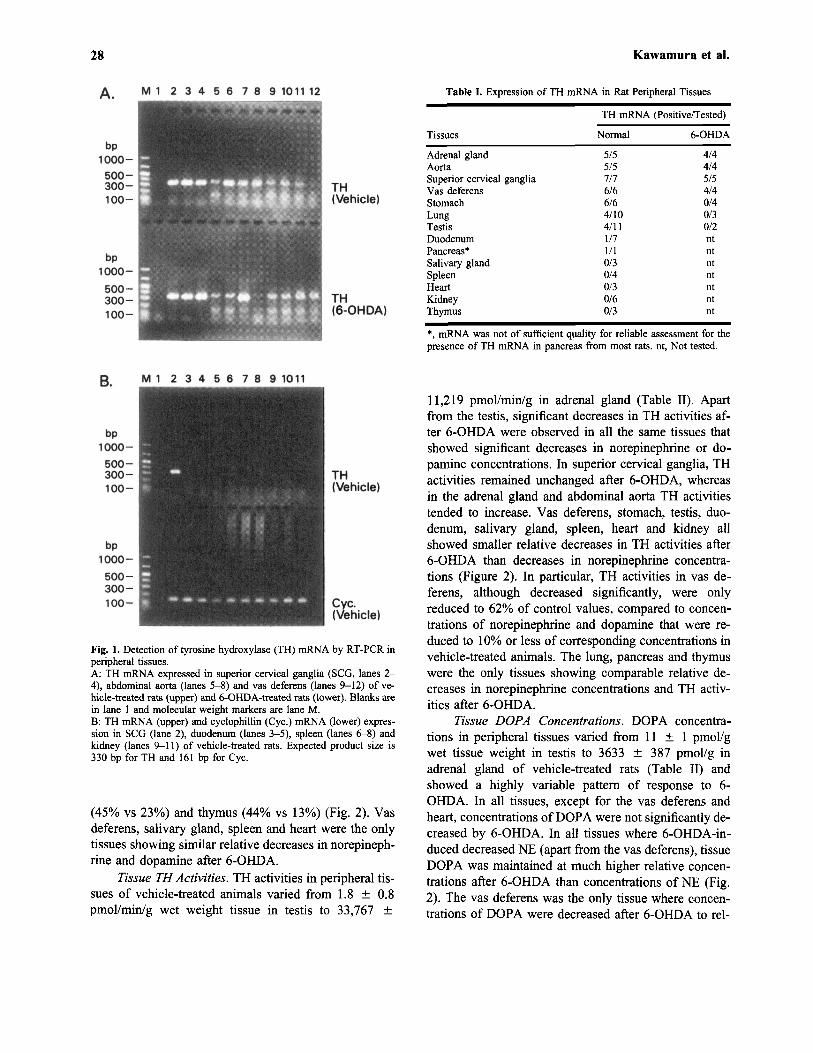
28 Kawamura et al.
Table I. Expression of TH mRNA in Rat Peripheral Tissues
Tissues
Adrenal glandAortaSuperior cervical gangliaVas deferensStomachLungTestisDuodenumPancreas*Salivary glandSpleenHeartKidneyThymus
TH mRNA (Positive/Tested)
Normal
5/55/57/76/66/64/104/111/71/10/30/40/30/60/3
6-OHDA
4/44/45/54/40/40/30/2ntntntntntntnt
*, mRNA was not of sufficient quality for reliable assessment for thepresence of TH mRNA in pancreas from most rats, nt, Not tested.
Fig. 1. Detection of tyrosine hydroxylase (TH) mRNA by RT-PCR inperipheral tissues.A: TH mRNA expressed in superior cervical ganglia (SCG, lanes 2-4), abdominal aorta (lanes 5-8) and vas deferens (lanes 9-12) of ve-hicle-treated rats (upper) and 6-OHDA-treated rats (lower). Blanks arein lane 1 and molecular weight markers are lane M.B: TH mRNA (upper) and cyclophillin (Cyc.) mRNA (lower) expres-sion in SCG (lane 2), duodenum (lanes 3-5), spleen (lanes 6-8) andkidney (lanes 9-11) of vehicle-treated rats. Expected product size is330 bp for TH and 161 bp for Cyc.
(45% vs 23%) and thymus (44% vs 13%) (Fig. 2). Vasdeferens, salivary gland, spleen and heart were the onlytissues showing similar relative decreases in norepineph-rine and dopamine after 6-OHDA.
Tissue TH Activities. TH activities in peripheral tis-sues of vehicle-treated animals varied from 1.8 ± 0.8pmol/min/g wet weight tissue in testis to 33,767 ±
11,219 pmol/min/g in adrenal gland (Table II). Apartfrom the testis, significant decreases in TH activities af-ter 6-OHDA were observed in all the same tissues thatshowed significant decreases in norepinephrine or do-pamine concentrations. In superior cervical ganglia, THactivities remained unchanged after 6-OHDA, whereasin the adrenal gland and abdominal aorta TH activitiestended to increase. Vas deferens, stomach, testis, duo-denum, salivary gland, spleen, heart and kidney allshowed smaller relative decreases in TH activities after6-OHDA than decreases in norepinephrine concentra-tions (Figure 2). In particular, TH activities in vas de-ferens, although decreased significantly, were onlyreduced to 62% of control values, compared to concen-trations of norepinephrine and dopamine that were re-duced to 10% or less of corresponding concentrations invehicle-treated animals. The lung, pancreas and thymuswere the only tissues showing comparable relative de-creases in norepinephrine concentrations and TH activ-ities after 6-OHDA.
Tissue DOPA Concentrations. DOPA concentra-tions in peripheral tissues varied from 11 ± 1 pmol/gwet tissue weight in testis to 3633 ± 387 pmol/g inadrenal gland of vehicle-treated rats (Table II) andshowed a highly variable pattern of response to 6-OHDA. In all tissues, except for the vas deferens andheart, concentrations of DOPA were not significantly de-creased by 6-OHDA. In all tissues where 6-OHDA-in-duced decreased NE (apart from the vas deferens), tissueDOPA was maintained at much higher relative concen-trations after 6-OHDA than concentrations of NE (Fig.2). The vas deferens was the only tissue where concen-trations of DOPA were decreased after 6-OHDA to rel-

Tyrosine Hydroxylase and Sympathectomy 29
atively lower concentrations than observed for the6-OHDA-induced decrease in TH activity. In lung, du-odenum, spleen and kidney, tissue concentrations ofDOPA in 6-OHDA-treated animals were maintained atmuch higher relative concentrations than those of THactivity and concentrations of dopamine and norepi-nephrine.
DISCUSSION
This study combined measurements of tissue THmRNA and catechol concentrations with a new sensitiveassay of TH activity to show that these variables aredifferentially affected by chemical sympathectomy andthat tissue concentrations of norepinephrine, dopamineand DOPA particularly, are not always simple markersof sympathetic innervation or TH activity. Examinationof the relationship of TH to sympathetic innervation andtissue catecholamines has previously been hindered bylack of sufficiently sensitive assays of TH enzyme activ-ity.
The neurotoxic actions of 6-OHDA are dependenton uptake of the toxin by the norepinephrine transporter,which is localized mainly to sympathetic nerve terminalsrather than cell bodies or outer membranes of chromaffincells (10-12). This explains the decreased norepineph-rine concentrations after chemical sympathectomy insympathetically innervated tissues and lack of effect inthe adrenal gland, aorta and superior cervical ganglia.The unchanged or increased TH activities and dopamineconcentrations in adrenal gland and superior cervicalganglia after 6-OHDA are explained similarly, consistentwith findings of previous reports (11,13,14). The similarresponse pattern of catecholamine contents and TH ac-tivity in abdominal aorta and adrenal gland presumablyreflects the existence of chromaffin tissue (e.g. organs ofZuckerkandl) around the abdominal aorta (15,16).
In keeping with the localization of transcriptionprocesses and the resistance of sympathetic ganglia andadrenal and extra-adrenal chromaffin tissue to the toxicactions of 6-OHDA, TH mRNA was consistently de-tected in adrenal gland, superior cervical ganglia andaorta of both vehicle-treated and 6-OHDA-treated ani-mals. The vas deferens was the only other peripheraltissue where TH mRNA was consistently detected inboth vehicle-treated and 6-OHDA-treated animals. Inthis tissue, TH activity was also maintained at a rela-tively high level after 6-OHDA treatment (62% of ve-hicle-treated animals) despite larger than 90% loss oftissue norepinephrine and dopamine. These results canbe explained by findings that the adrenergic innervation
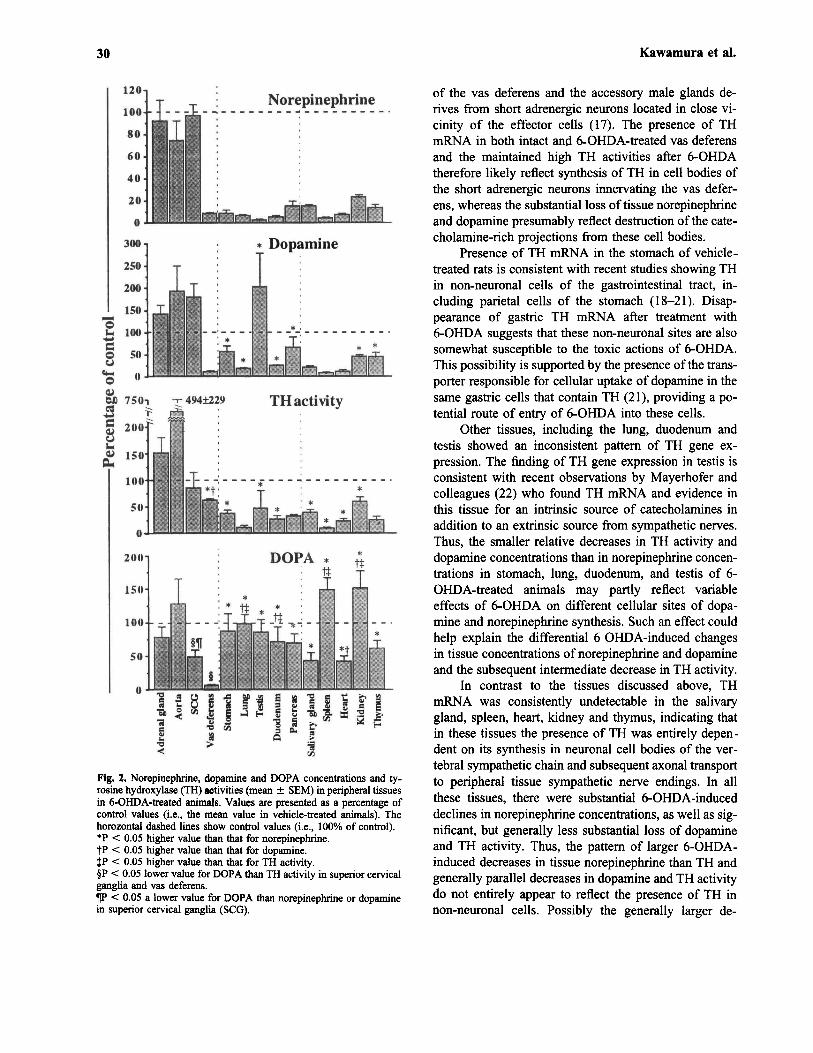
30 Kawamura et al.
Fig. 2. Norepinephrine, dopamine and DOPA concentrations and ty-rosine hydroxylase (TH) activities (mean ± SEM) in peripheral tissuesin 6-OHDA-treated animals. Values are presented as a percentage ofcontrol values (i.e., the mean value in vehicle-treated animals). Thehorozontal dashed lines show control values (i.e., 100% of control).*P < 0.05 higher value than that for norepinephrine.+P < 0.05 higher value than that for dopamine.+P < 0.05 higher value than that for TH activity.§P < 0.05 lower value for DOPA than TH activity in superior cervicalganglia and vas deferens.PP < 0.05 a lower value for DOPA than norepinephrine or dopaminein superior cervical ganglia (SCG).
of the vas deferens and the accessory male glands de-rives from short adrenergic neurons located in close vi-cinity of the effector cells (17). The presence of THmRNA in both intact and 6-OHDA-treated vas deferensand the maintained high TH activities after 6-OHDAtherefore likely reflect synthesis of TH in cell bodies ofthe short adrenergic neurons innervating the vas defer-ens, whereas the substantial loss of tissue norepinephrineand dopamine presumably reflect destruction of the cate-cholamine-rich projections from these cell bodies.
Presence of TH mRNA in the stomach of vehicle-treated rats is consistent with recent studies showing THin non-neuronal cells of the gastrointestinal tract, in-cluding parietal cells of the stomach (18-21). Disap-pearance of gastric TH mRNA after treatment with6-OHDA suggests that these non-neuronal sites are alsosomewhat susceptible to the toxic actions of 6-OHDA.This possibility is supported by the presence of the trans-porter responsible for cellular uptake of dopamine in thesame gastric cells that contain TH (21), providing a po-tential route of entry of 6-OHDA into these cells.
Other tissues, including the lung, duodenum andtestis showed an inconsistent pattern of TH gene ex-pression. The finding of TH gene expression in testis isconsistent with recent observations by Mayerhofer andcolleagues (22) who found TH mRNA and evidence inthis tissue for an intrinsic source of catecholamines inaddition to an extrinsic source from sympathetic nerves.Thus, the smaller relative decreases in TH activity anddopamine concentrations than in norepinephrine concen-trations in stomach, lung, duodenum, and testis of 6-OHDA-treated animals may partly reflect variableeffects of 6-OHDA on different cellular sites of dopa-mine and norepinephrine synthesis. Such an effect couldhelp explain the differential 6 OHDA-induced changesin tissue concentrations of norepinephrine and dopamineand the subsequent intermediate decrease in TH activity.
In contrast to the tissues discussed above, THmRNA was consistently undetectable in the salivarygland, spleen, heart, kidney and thymus, indicating thatin these tissues the presence of TH was entirely depen-dent on its synthesis in neuronal cell bodies of the ver-tebral sympathetic chain and subsequent axonal transportto peripheral tissue sympathetic nerve endings. In allthese tissues, there were substantial 6-OHDA-induceddeclines in norepinephrine concentrations, as well as sig-nificant, but generally less substantial loss of dopamineand TH activity. Thus, the pattern of larger 6-OHDA-induced decreases in tissue norepinephrine than TH andgenerally parallel decreases in dopamine and TH activitydo not entirely appear to reflect the presence of TH innon-neuronal cells. Possibly the generally larger de-

Tyrosine Hydroxylase and Sympathectomy 31
creases in tissue norepinephrine contents than THactivity may reflect differential effects of 6-OHDA, firstto deplete vesicular stores of transmitter and second todestroy sympathetic nerve endings or non-neuronal cellswhere 6-OHDA is concentrated by monoamine trans-porters. Decreases in TH activity that closely mirroredthose in tissue dopamine, but not the larger decreases innorepinephrine, might then be expected from residualTH activity and synthesis of dopamine in remainingsympathetic nerves where vesicular stores of norepi-nephrine are depleted.
The above outlined explanations for the differentialeffects of 6-OHDA on TH activity and tissue norepi-nephrine and dopamine do not, however, account for theextremely variable 6-OHDA-induced changes in tissueconcentrations of DOPA. These concentrations weregenerally maintained above or close to normal concen-trations, with significant decreases only in the heart andvas deferens. Only in the vas deferens did the decreasein DOPA match the decrease in tissue norepinephrineand dopamine, but this tissue also showed only a smallreduction in TH activity. Thus, in most tissues, concen-trations of DOPA do not reflect activity of TH and areunlikely to be derived from local synthesis by TH. Thisconclusion is in agreement with that reached by Andenand colleagues (23) who surmised that DOPA in severalperipheral tissues was derived from circulating DOPA.Eldrup and colleagues (24) also found that surgical sym-pathectomy had no influence on DOPA concentrationsin muscle tissue, concluding that DOPA in muscle islocated outside sympathetic nerves and may act as a res-ervoir for DOPA throughout the body. Our findings in-dicate that the independence of tissue DOPAconcentrations on sympathetic innervation is much moregeneral than indicated by Eldrup et al. (24). Other find-ings that tissue concentrations of DOPA are unaffectedor only marginally reduced in mutant mice lacking THsuggests that most tissue DOPA is formed by an alter-native pathway to that responsible for synthesis ofDOPA and catecholamines in aminergic neurons andchromaffin tissue (our unpublished observations). Sucha pathway has previously been implicated by Kobayashiand colleagues (25) to account for relatively high re-maining dopamine concentrations in a previously re-ported strain of mutant mice lacking TH.
In conclusion, the heterogenous effects of chemicalsympathectomy on catechol contents and tyrosine hy-droxylase activity and gene expression among differentorgans and tissues demonstrate that these variables donot provide simple indices of sympathetic innervation orcatecholamine synthesis. Tissue differences in presenceof neuronal cell bodies, parenchymal chromaffin cells
and other non-neuronal sites of catecholamine synthesisas well as alternative sources of supply of tissue DOPArepresent other factors that should be considered whenperipheral tissue concentrations of norepinephrine, dopa-mine, TH and DOPA are considered as markers of sym-pathetic innervation and local catecholamine synthesis.
REFERENCES
1. Goldstein, D. S. 1992. Clinical uses of catechols in the assessmentof sympathoadrenal activity in stress and disease, pages 893-913,in Kvetnansky, R., McCarty, R., Axelrod, J. (eds), Stress: Neu-roendocrine and Molecular Approaches. New York: Gordon andBreach Science Publishers.
2. Oomori, Y., Iuchi, H., Ishikawa, K., Satoh, Y., and Ono, K. 1994.Immunocytochemical study of tyrosine hydroxylase and dopamineB-hydroxylase immunoreactivities in the rat pancreas. Histochem -istry 101:313-323.
3. Dees, W. L., Hiney, J. K., Schultea, T. D., Mayerhofer, A., Dan-ilchik, M, Dissen, G. A., and Ojeda, S. R. 1995. The primateovary contains a population of catecholaminergic neuron-like cellsexpressing nerve growth factor receptors. Endocrinology 136:5760-5768.
4. Huang, M.-H., Friend, D. S., Sunday, M. E., Singh, K., Haley, K.,Austen, K. F., Kelly, R. A., and Smith, T. W. 1996. An Intrinsicadrenergic system in mammalian heart. J. Clin. Invest. 98:1298-1303.
5. Josefsson, E., Bergquist, J., Ekman, R., and Tarkowski, A. 1996.Catecholamines are synthesized by mouse lymphocytes and reg-ulate function of these cells by induction of apoptosis. Immunol-ogy 88:140-146.
6. Mezey, E., Eisenhofer, G., Harta, G., Hansson, S., Gould, L., Hu-nyady, B., and Hoffman, B. J. 1996. A novel nonneuronal ca-techolaminergic system: Exocrine pancreas synthesizes andreleases dopamine. Proc. Natl. Acad. Sci. USA 93:10377-10382.
7. Hooper, D., Kawamura, M., Hoffman, B., Kopin, I. J., Hunyady,B., Mezey, E., and Eisenhofer, G. 1997. Tyrosine hydroxylaseassay for detection of low levels of enzyme activity in peripheraltissue. J. Chromatogr. 694:317-324.
8. Schwartz, J. P. 1988. Stimulation of nerve growth factor mRNAcontent in C6 glioma cells by a B-adrenergic receptor and bycyclic AMP. Glia 1:282-285.
9. Eisenhofer, G., Goldstein, D. S., Stall, R., Keiser, H. R., Sunder-land, T., Murphy, D. L., and Kopin, I. J. 1986. Simultaneous liq-uid-chromatographic determination of 3,4-dihydroxyphenylglycol,catecholamines and 3,4-dihydroxyphenylalanine in plasma andtheir responses to inhibition of monoamine oxidase. Clin. Chem.32:2030-2033.
10. Thoenen, H., and Tranzer, J. P. 1968. Chemical sympathectomyby selective destruction of adrenergic nerve endings with 6-hy-droxydopamine. Naunyn-Schmiedebergs Arch. Pharmacol. Exp.Path. 261:271-288.
11. Kostrzewa, R. M., and Jacobowitz, D. 1974. Pharmacological ac-tions of 6-hydroxydopamine. Pharmacol. Rev. 26:199-288.
12. Jonsson, G. 1983. Chemical lesioning techniques, pages pp 463-507, in Bjorklund, A,. Hokfelt, T. (eds): Handbook of chemicalneuroanatomy, Vol. 1: Methods in chemical neuroanatomy. Am-sterdam-New York- Oxford: Elsevier.
13. Thoenen, H., Mueller, R. A., and Axelrod, J. 1969. Trans-synapticinduction of adrenal tyrosine hydroxylase. J. Pharmacol. Exp.Ther. 169:249-254.
14. Fety, R., Misere, V., Lambas-Senas, L., and Renaud, B. 1986.Central and peripheral changes in catecholamine-synthesizing en-zyme activities after systemic administration of the neurotoxinDSP-4. Eur. J. Pharmacol. 124:197-202.

32 Kawamura et al.
15. West, G. B., Shepherd, D. M., Hunter, R. B., and Macgregor, A.R. 1953. The function of the organs of zuckerkandl. Clin. Sci. 12:317-325.
16. Gallardo, M. G. P. DE, Freire, F., and Tramezzani, J. H. 1974.The organ of zuckerkandl of the newborn rat and its postnatalinvolution. Acta Physiol. Lat. 24:290-304.
17. Sjostrand, N. O. 1965. The adrenergic innervation of the vas defer-ens and the accessory male genital glands: An experimental andcomparative study of its anatomical and functional organization insome mammals, including the presence of adrenaline and chromaf-fin cells in these organs. Acta Physiol. Scand. 65:Suppl 257:5-82.
18. Back, N., Ahonen, M., Soinila, S., Kivilaakso, E., and Kiviluoto,T. 1995. Catecholamine-synthesizing enzymes in the rat stomach.Histochem. Cell. Biol. 104:63-67.
19. Eisenhofer, G., Aneman, A., Friberg, P., Hooper, D., Fandrinks,L., Lonroth, H., Hunyady, B., and Mezey, E. 1997. Substantialproduction of dopamine in the human gastrointestinal tract. J.Clin. Endocrinol. Metab. 82:3864-3871.
20. Hunyady, B., Hoffman, B. J., Eisenhofer, G., Hansson, S. R.,Mezey, E. 1997. Dopaminergic characteristics of isolated parietalcells from rats. J. Physiol. (Paris) 91:247-256.
21. Mezey, E., Eisenhofer, G., Hansson, S., Hunyady, B., and Hoff-man, B. J. 1998. Dopamine produced by the stomach may act asa paracrine/autocrine hormone in the rat. Neuroendocrinology176:336-348.
22. Mayerhofer, A., Danilchik, M., Pau, K. Y., Lara, H. E., Russell,L. D., and Ojeda, S. R. 1996. Testis of prepubertal rhesus mon-keys receives a dual catecholaminergic input provided by the ex-trinsic innervation and an intragonadal source of catecholamines.Biol. Reprod. 55:509-518.
23. Anden, N. E., Grabowska-Anden, M., and Schwieler, J. 1989.Transfer of DOPA from sympatho-adrenal system to the pancreas,liver and kidney via the blood circulation. Acta Physiol. Scand.136:75-79.
24. Eldrup, E., Richter, E. A., and Christensen, N. J. 1989. DOPA,norepinephrine, and dopamine in rat tissues: No effect of sym-pathectomy on muscle DOPA. Am. J. Physiol. 256:E284-E287.
25. Kobayashi, K., Morita, S., Sawada, H., Mizuguchi, T., Yamada,K., Nagatsu, I., Hata, T., Watanabe, Y., Fujita, K., and Nagatsu,T. 1995. Targeted disruption of the tyrosine hydroxylase locusresults in severe catecholamine depletion and perinatal lethality inmice. J. Biol. Chem. 270:27235-27243.










![Usefulness of [18F]-DA and [18F]-DOPA for PET imaging in a mouse model of pheochromocytoma](https://static.fdokumen.com/doc/165x107/6325a7d9852a7313b70e9a7d/usefulness-of-18f-da-and-18f-dopa-for-pet-imaging-in-a-mouse-model-of-pheochromocytoma.jpg)









