
l-Dopa-induced locomotor-like activity in ankle flexor and extensor nerves of chronic and acute...
12
EXPERIMENTAL NEUROLOGY 86, 5 15-526 (1984) L-Dopa-Induced Locomotor-like Activity in Ankle Flexor and Extensor Nerves of Chronic and Acute Spinal Cats LUCINDA L. BAKER, SCOTT H. CHANDLER, AND LOUIS J. GOLDBERG’ Department of Kinesiology and the Brain Research Institute; Department of Oral Biology, School of Dentistry; and Department of Anatomy, School of Medicine, University of California. Los Angeles, Cahyornia 90024 Received October 12, 1983; revision received July 9, 1984 The ability of L-DOPA and nialamide to produce locomotor-like rhythmic discharges (fictive locomotion) in hind limb nerves of acute and chronically spinalized and paralyzed cats was exmained. Ankle flexor and extensor nerves of chronic cats exhibited pharmacologically induced alternate bursts of activity that had significantly shorter cycle times and burst durations than those produced in ankle flexor and extensor nerves of acutely prepared cats. Furthermore, prior to pharmacologic activation, both ankle flexor and extensor nerves of chronic preparations frequently exhibited spontaneous alternate bursts of activity. Neuronal discharges from nerves of chronic preparations, both prior to and after pharmacologic activation, exhibited much greater variability in both cycle time and burst duration compared with those observed in acute preparations. cl 1984 Academic PXS. 1~. INTRODUCTION Locomotor-like bursting discharges (fictive locomotion) can be recorded from hind limb flexor and extensor nerves of acutely spinalized and paralysed cats after the systemic administration of monoamine agonists or their precursors (7, 10, 13). These burst patterns, although slower in frequency, are similar to the patterns observed in normal cats during treadmill locomotion (16). In the unanesthetized, chronically spinalized cat Abbreviations: TA-tibialis anterior: MG. LG-medial, lateral gastrocnemius: S-soleus; LGSR, MGR-right LG-S. MG. ’ We thank V. R. Edgerton for assistance in research design and M. Foumier for his assistance during data collection and reduction. This work was supported by the National Institute of Neurological and Communicative Disorders and Stroke, research grant NS 6333. Please send all correspondence to Dr. Chandler. Department of Kinesiology. 515 OOl4-4886/84 $3.00 Copyright 0 1984 by Academic Press. Inc. All ngbts of reproduction in any form reserved
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of l-Dopa-induced locomotor-like activity in ankle flexor and extensor nerves of chronic and acute...

EXPERIMENTAL NEUROLOGY 86, 5 15-526 (1984)
L-Dopa-Induced Locomotor-like Activity in Ankle Flexor and Extensor Nerves of Chronic and Acute Spinal Cats
LUCINDA L. BAKER, SCOTT H. CHANDLER, AND LOUIS J. GOLDBERG’
Department of Kinesiology and the Brain Research Institute; Department of Oral Biology, School of Dentistry; and Department of Anatomy, School of Medicine, University
of California. Los Angeles, Cahyornia 90024
Received October 12, 1983; revision received July 9, 1984
The ability of L-DOPA and nialamide to produce locomotor-like rhythmic discharges (fictive locomotion) in hind limb nerves of acute and chronically spinalized and paralyzed cats was exmained. Ankle flexor and extensor nerves of chronic cats exhibited pharmacologically induced alternate bursts of activity that had significantly shorter cycle times and burst durations than those produced in ankle flexor and extensor nerves of acutely prepared cats. Furthermore, prior to pharmacologic activation, both ankle flexor and extensor nerves of chronic preparations frequently exhibited spontaneous alternate bursts of activity. Neuronal discharges from nerves of chronic preparations, both prior to and after pharmacologic activation, exhibited much greater variability in both cycle time and burst duration compared with those observed in acute preparations. cl 1984 Academic PXS. 1~.
INTRODUCTION
Locomotor-like bursting discharges (fictive locomotion) can be recorded from hind limb flexor and extensor nerves of acutely spinalized and paralysed cats after the systemic administration of monoamine agonists or their precursors (7, 10, 13). These burst patterns, although slower in frequency, are similar to the patterns observed in normal cats during treadmill locomotion (16). In the unanesthetized, chronically spinalized cat
Abbreviations: TA-tibialis anterior: MG. LG-medial, lateral gastrocnemius: S-soleus; LGSR, MGR-right LG-S. MG.
’ We thank V. R. Edgerton for assistance in research design and M. Foumier for his assistance during data collection and reduction. This work was supported by the National Institute of Neurological and Communicative Disorders and Stroke, research grant NS 6333. Please send all correspondence to Dr. Chandler. Department of Kinesiology.
515
OOl4-4886/84 $3.00 Copyright 0 1984 by Academic Press. Inc. All ngbts of reproduction in any form reserved

516 BAKER, CHANDLER, AND GOLDBERG
or rat with intact dorsal roots, locomotor-like movements in the hind limbs have also been demonstrated under various conditions (5, 6, 15, 18, 19). The locomotor capability of the spinal cord of a chronically spinalized and paralyzed cat deprived of phasic sensory input has not been reported in detail (10, 11). The purpose of this study was to determine whether or not the spinal cord of a cat transected at an early age and subsequently deprived of phasic sensory feedback is capable of producing locomotor-like patterns of burst discharges in hind limb flexor and extensor nerves (fictive loco- motion) prior to and after pharmacologic stimulation with L-DOPA and nialamide. It was also of interest to compare drug-induced burst discharge patterns and cycle characteristics in chronically transected cats with those induced in acutely spinalized and paralysed preparations.
METHODS
General Procedures. The experiments were carried out on 13 adult cats that were not colony raised (acute cats), and nine cats that were 6 to 9 months old (chronic cats). The spinal cord of the chronic cats were transected at the level of T12 when the cats here 2 weeks of age. They were then raised with normal littermates in large social cages.
At the time of the terminal experiment, all cats were anesthesized with general halothane anesthesia, tracheotomized, and the carotid and jugular vessels cannulated. The animals were anemically decapitated, artifically ventilated and paralyzed with gallamine triethiodide (approximately 0.5 mg/kg, i.v.). For anemic decapitation the common carotid arteries were ligated bilaterally, then the vertebral arteries were clamped by compression between the first and second cervical vetebrae. An interruption of sponta- neous respiration resulted from this procedure, together with dilated and nonreactive pupils. These signs were taken as an indication of a complete decapitation. The anesthesia was discontinued after the decapitation was completed. The spinal cords of the acute animals were transected at the T 11 -T 12 level. End-tidal CO2 concentration was monitored and maintained at 3 to 5% by adjusting the oxygen content of the inspired air and/or by adjusting the frequency of ventilation. Arterial blood pressure was monitored by a catheter inserted into the common carotid artery. If blood pressure decreased below 90 mm Hg, a single dose of a peripherally acting stimulant (Metaraminol) was administered. No animal required repeated stimulant injections. Rectal temperature was maintained at approximately 37°C by radiant heat.
Preparation. The left hind limb nerves innervating the lateral gastrocne- mius (LG), medial gastrocnemius (MG), soleus (S), and tibialis anterior (TA) were identified, dissected free, and subsequently cut. All nerves were

SPONTANEOUS NEURAL BURSTING IN CHRONIC SPINAL CATS 517
prepared for mounting on recording or stimulating electrodes. In some experiments the contralateral (right) lateral gastrocneumius-soleus (LGSR) and medial gastronemius (MGR) nerves were identified and transected. Nerve recordings were obtained from the LG, MG, S, TA, LGSR, and/or MGR nerves in all experiments. Four channels of the tape recorder were available for recording nerve activity, therefore, the nerves which exhibited the most rhythmic activity were chosen for recording. If more than four nerves were active, various combinations of nerve recordings were made. The nerves were mounted on silver bipolar electrodes within an oil pool, which was maintained at 37°C by radiant heat. Ventral and dorsal roots were maintained in continuity throughout all experiments.
Recordings. Nerve activity was recorded on an FM tape recorder. Injections (iv.) of 100 mg/kg nialamide (Sigma) and 100 mg/kg L-DOPA (Sigma) were made. Spontaneous activity in the nerves was recorded for as long as 5 h after administration of the drugs.
Data Analysis. The neurograms of both raw and integrated recordings were played back on an ink pen polygraph (Grass). Burst durations and cycle times were measured from the polygraph record. Data from recordings made between 1 and 4 h after the administration of the drugs were evaluated.
Cycle time and burst duration means and standard errors of the means for each trail and for each animal were calculated based on 10 to 50 consecutive cycles within the trial. Data from only one trial for each nerve in each animal were used for analysis of the correlation between cycle times and burst durations. Differences between mean cycle times or burst dura- tions for acute and chronic animals prior to and after the administration of L-DOPA were determined statistically by analysis of variance (ANOVA) and Tukey’s post hoc test for comparisons of means. Activities from the flexor and extensor nerves were evaluated separately. Statistical significance was accepted at the P < 0.05 level.
Linear regression analyses were computed to determine the relationship between cycle times and burst durations. The scatter of the data points was deemed to fit the regression line when the correlation coefficient (r) exceeded the 0.05 level of probability in Pearson’s product moment correlation coefficient table. Comparison of slopes of the regression lines was restricted to those linear regression lines that significantly fit the scatter of the data points.
RESULTS
Spontaneous Bursting Patterns in Hind Limb Nerves after the Administra- tion of L-DOPA. The systemic administration of L-DOPA and nialamide to acute and chronically spinalized and paralyzed cats produced, in all
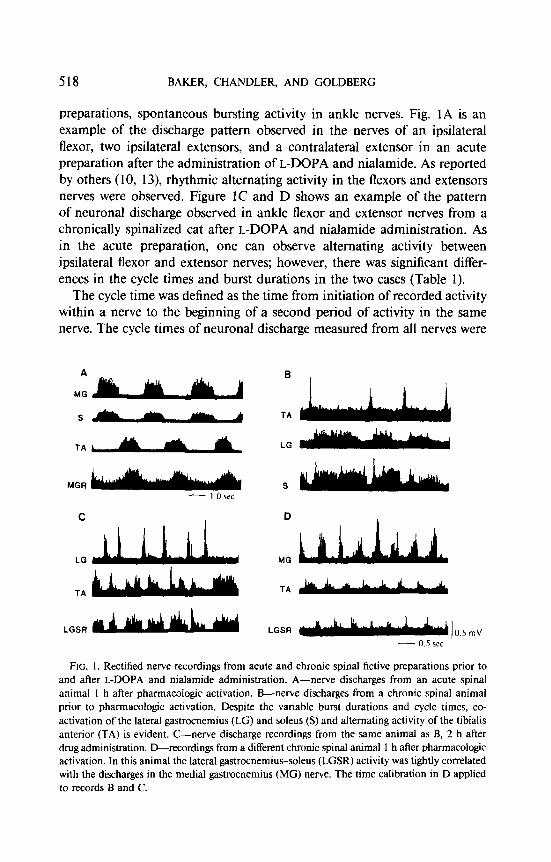
518 BARER, CHANDLER, AND GOLDBERG
preparations, spontaneous bursting activity in ankle nerves. Fig. 1A is an example of the discharge pattern observed in the nerves of an ipsilateral flexor, two ipsilateral extensors, and a contralateral extensor in an acute preparation after the administration of L-DOPA and nialamide. As reported by others (10, 13) rhythmic alternating activity in the flexors and extensors nerves were observed. Figure IC and D shows an example of the pattern of neuronal discharge observed in ankle flexor and extensor nerves from a chronically spinalized cat after L-DOPA and nialamide administration. As in the acute preparation, one can observe alternating activity between ipsilateral flexor and extensor nerves; however, there was significant differ- ences in the cycle times and burst durations in the two cases (Table 1).
The cycle time was defined as the time from initiation of recorded activity within a nerve to the beginning of a second period of activity in the same nerve. The cycle times of neuronal discharge measured from all nerves were
TA
LGSR
B
TA
LG
TA
LGSR
FIG. 1. Rectified nerve recordings from acute and chronic spinal fictive preparations prior to and after L-DOPA and nialamide administration. A-nerve discharges from an acute spinal animal I h after pharmacologic activation. B-nerve discharges from a chronic spinal animal prior to pharmacologic activation. Despite the variable burst durations and cycle times, co- activation of the lateral gastrocnemius (LG) and soleus (S) and alternating activity of the tibialis anterior (TA) is evident. C-nerve discharge recordings from the same animal as B, 2 h after drug administration. D-recordings from a different chronic spinal animal 1 h aher pharmacologic activation. In this animal the lateral gastrocnemius-soleus (LGSR) activity was tightly correlated with the discharges in the medial gastrocnemius (MC) nerve. The time calibration in D applied to records B and C.

SPONTANEOUS NEURAL BURSTING IN CHRONIC SPINAL CATS 519
TABLE 1
Burst Durations and Cycle Times of Acute and Chronic Spinal Preparations”
Gastrocnemius
Tibia& anterior Medial Lateral Soleus
Acute, L-DOPA Cycle time 4.80 f 0.6 3.98 f 0.8 4.05 f 0.5 6.4 + 1.95
(8) (3) (5) (3) Burst duration 1.67 + 0.3 2.49 f 0.6 2.1 f 0.4 2.62 f 0.8
Chronic, after L-DOPA Cycle time 1.29 f 0.1 I.16 k 0.1 1.44 + 0.4 1.12 * 0.2
16) (4) (41 (5) Burst duration 0.52 + 0.2 0.74 + 0.1 0.44 f 0.1 0.21 + 0.04
Chronic, before L-DOPA Cycle time 1.45 k 0.4 1.38 + 0.2 1.74 Ii 0.2 0.64 + 0.3
(4) (3) (3) (3) Burst duration 0.5 * 0.1 0.52 * 0.1 0.53 * 0.1 0.44 f 0.1
’ All values are seconds, X + SE. Numbers in parentheses denote number of animals
much shorter in the chronic preparations compared with those in the acute preparations (Table 1, Fig. 1). The cycle times for extensor nerve activity in the acute preparations for all animals ranged from a mean of 2.5 to 8.6 s, and the mean cycle times of the flexors nerves ranged from 2.7 to 8.1 s. In the chronic preparations the range of mean cycle times for extensor nerve discharges after drug administration were 0.6 to 2.3 s, and those observed for flexor filaments was 0.9 to 1.9 s.
Burst durations in the acute preparations were significantly longer than those observed in the chronic preparation (Table 1, Fig. 1). The mean burst duration range for the acute and chronic preparations were as follows: acute extensors-l. 1 to 4.0 s; acute flexors-l.8 to 3.5 s; chronic extensors-O.2 to 0.8 s; chronic flexors-O.3 to 0.8 s.
The relationship between cycle times and burst durations of flexor and extensor nerve activity in the acute and chronic preparations was examined. These results are shown in Fig. 2. A significant linear relationship between cycle times and burst durations for both flexor and extensor nerve activity was found in both the acute and chronic preparations in more than 60% of the trials analyzed for all preparations. This result was obtained 2 to 4 h postinjection when nerve activity was at its highest level. Furthermore, there was no statistical difference observed between the slopes of the lines relating cycle time and burst duration for the extensor and flexor nerves in either the acute or chronic preparations.
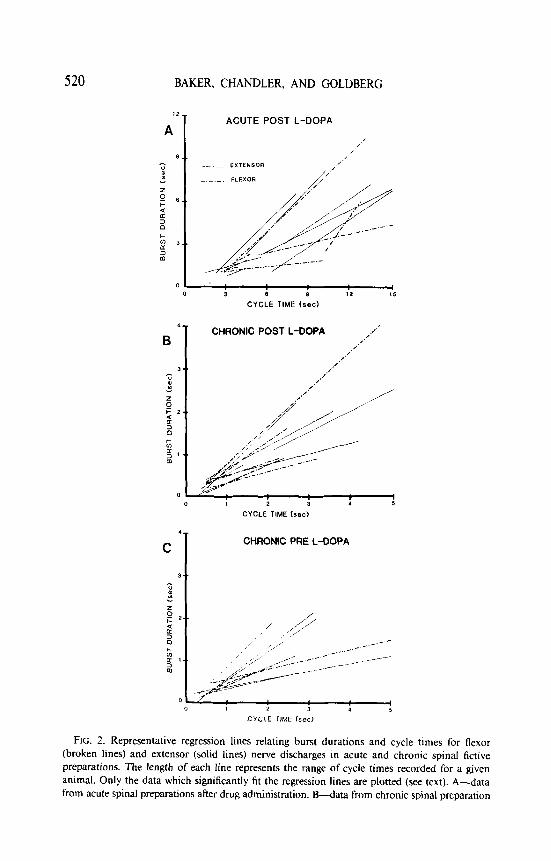
520 BAKER, CHANDLER, AND GOLDBERG
ACUTE POST L-DOPA
,’ /
__ EXTENSOA
-.-._. FLEXOR
4- CHRONIC POST L-DOPA .I-’
B .x .f .’ .x .fl 3.. .N
2
CYCLE TIME bed
FIG. 2. Representative regression lines relating burst durations and cycle times for flexor (broken lines) and extensor (solid lines) nerve discharges in acute and chronic spinal fictive preparations. The length of each line represents the range of cycle times recorded for a given animal. Only the data which significantly fit the regression lines are plotted (see text). A-data from acute spinal preparations after drug administration. B-data from chronic spinal preparation
SPONTANEOUS NEURAL BURSTING IN CHRONIC SPINAL CATS
A NORMAL WALK
LG
-20 0 20 40 I)0 aa ,m 120
PERCENTAGE OF LG CYCLE
B ACUTE POST L-DOPA
LG
s
PERCENTAGE OF LG CYCLE
C
MG
TA
LGSR
D
CHRONIC POST L-DOPA
+I+
+
+ +
20 40 BO 80
PERCENTAGE OF MG CYCLE
CHRONIC PRE L-DOPA
PERCENTAGE OF MG CYCLE
521
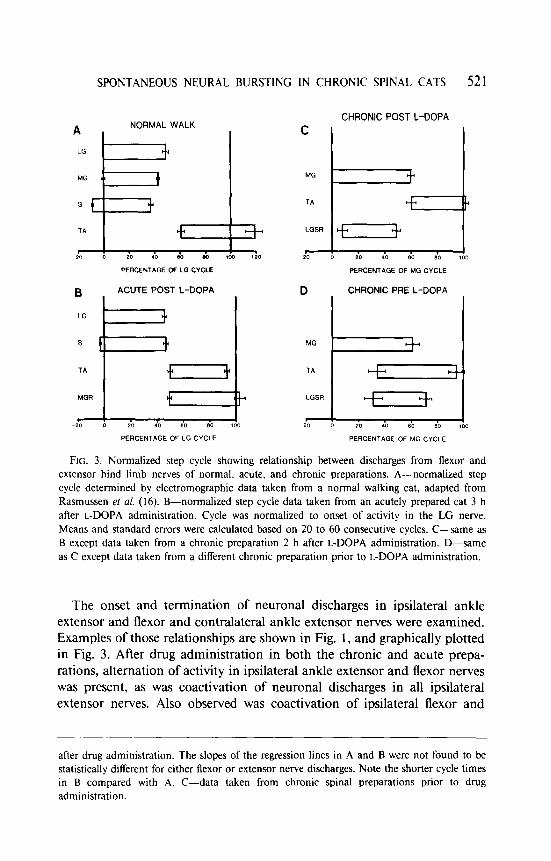
FIG. 3. Normalized step cycle showing relationship between discharges from flexor and extensor hind limb nerves of normal. acute, and chronic preparations. A-normalized step cycle determined by electromographic data taken from a normal walking cat, adapted from Rasmussen ef ni. (16). B-normalized step cycle data taken from an acutely prepared cat 3 h after L-DOPA administration. Cycle was normalized to onset of activity in the LG nerve. Means and standard errors were calculated based on 20 to 60 consecutive cycles. C-same as B except data taken from a chronic preparation 2 h after L-DOPA administration. D-same
as C except data taken from a different chronic preparation prior to L-DOPA administration,
The onset and termination of neuronal discharges in ipsilateral ankle extensor and flexor and contralateral ankle extensor nerves were examined. Examples of those relationships are shown in Fig. 1, and graphically plotted in Fig. 3. After drug administration in both the chronic and acute prepa- rations, alternation of activity in ipsilateral ankle extensor and flexor nerves was present, as was coactivation of neuronal discharges in all ipsilateral extensor nerves. Also observed was coactivation of ipsilateral flexor and
after drug administration. The slopes of the regression lines in A and B were not found to be statistically different for either flexor or extensor nerve discharges. Note the shorter cycle times in B compared with A. C-data taken from chronic spinal preparations prior to drug administration.

522 BAKER, CHANDLER, AND GOLDBERG
contralateral extensor nerve activity in all acute preparations (Figs. 1A and 3B) and one of the four chronic preparations (Fig. 1C). In three chronic preparations the contralateral extensor was coactive with the ipsilateral extensor nerve (Figs. 1D and 3C).
In Fig. 3B, coactivation is seen between the ipsilateral extensor nerves. The soleus nerve discharge onset preceded the onset of activity in the LG by approximately lo%, and the termination of activity occurred simulta- neously at about 50% of the cycle. After cessation of the extensor bursts, activity began in both the ipsilateral flexor and contralateral extensor nerves. This pattern of neuronal discharge was consistently observed in all acute preparations.
For the chronic animal shown in Fig. 3C, the ipsilateral extensor nerve was active for approximately 60% of the cycle duration. Similar to that observed for acute preparations, the ipsilateral flexor nerve became active at the termination of the extensor burst and its duration was slightly longer and extended into the onset of the next extensor burst (Fig. 3C). In general, the relationship between onset and termination of neuronal activity in the nerves of the chronic animals was more variable than that observed in the acute preparation.
Spontaneous Discharge Patterns in Hind limb Nerves prior to the Admin- istration of L-DOPA. Neuronal bursting activity was never observed in acute, spinalized cats prior to the administration of L-DOPA or nialamide. This is in contrast to the seven chronic spinal cats, all of which demonstrated some spontaneous neural activity in ankle nerves prior to the administration of the drugs.
Four chronic animals demonstrated spontaneous activity in the flexor (TA) nerve and six showed activity in at least one extensor nerve. The cycle times and burst durations observed prior to drug administration were similar to those which occurred after the administration of nialamide and L-DOPA (Table 1). For cycle times and burst durations there was no statistical difference between the values before and after administration of L-DOPA.
Prior to administration of L-DOPA, flexor and extensor nerves demon- strated a significant linear relationship between burst duration and cycle times in 75 and 40%, respectively, of the recordings from all preparations (Fig. 2C). The slopes of the regression lines prior to the administration of L-DOPA were similar to those after the drug administration. Furthermore, there was no significant difference between the slopes of the regression lines for the flexor nerve recordings compared with those for the extensor nerves in the chronic spinal animals prior to the drug-induced activity.
The onset of activity in the ipsilateral flexor nerve with respect to an ipsilateral extensor nerve was much more variable prior to L-DOPA

SPONTANEOUS NEURAL BURSTING IN CHRONIC SPINAL CATS 523
administration. In some instances strong coactivation with the ipsilateral extensors occurred (Fig. 3D). A most consistent observation in neuronal discharge prior to L-DOPA administration was seen in the strong correlation between the onset of activity in the ipsilateral flexor and contralateral extensor nerves (Fig. 3D).
DISCUSSION
The results presented here confirm and extend the findings of others (10, 11) that the lumbosacral spinal cord isolated from supraspinal centers by low thoracic transection of 2 weeks of age and deprived of phasic sensory feedback can be stimulated by L-DOPA and nialamide to produce bursting patterns, some aspects of which resemble fictive locomotion in the acutely transected and paralysed cat. Although these discharge patterns can hardly be called locomotion, our data suggest that there exists in the chronic spinal animal a neuronal network(s) capable of generating some aspects of locomotor-like discharge patterns in flexor and extensor nerves in the absence of phasic peripheral feedback.
The two strongest similarities between spontaneous burst patterns in the acute and chronic spinal preparations were seen in the relationship between burst duration and cycle time, and in the reciprocal discharge pattern that occurred in the ipsilateral flexor and extensor nerves. The linear relationship between burst duration and cycle times for both extensor and flexor nerves were similar in the chronic and acute spinal animals. The central network(s) in the spinal cord of the chronic animals was still capable of maintaining the relationship between burst durations and cycle times to a degree similar to that which occurred in the acute animals. However, the absolute cycle times and burst durations were significantly shorter in the chronic prepa- rations suggesting that the excitability of the central pattern generating network(s) had been altered.
Furthermore, the slopes of the burst duration-cycle time relationship were found to be the same for flexors and extensors (Fig. 2). This is in contrast to that reported for fictive locomotion in the acute spinal cat (10). mesencephalic cat (12). and in freely walking and normal cats (9, 2 1). In those studies, the duration of extensor activity was shown to vary linearly with cycle duration whereas the burst duration of the flexor nerve or flexor muscle electromyographic activity was shown to be independent to cycle duration. Except for differences in experimental protocol, we can offer no explanation for the fact that previous investigators showed no significant relationship between burst duration and cycle time for flexor neuronal discharge.
A major difference observed between acute and chronic cats was in the duration of both the cycle times and burst durations. The observation that

524 BAKER, CHANDLER, AND GOLDBERG
burst durations and cycle times of the neuronal discharges were markedly shorter in chronic cats may indicate that the excitability of the neuronal networks generating these discharge patterns had increased in the chronic vs the acute preparation (3). It has been shown that chronic spinalization produces degeneration of descending noradrenergic and dopaminergic ter- minals within the spinal cords of both rats and cats (1, 2, 4, 14). Therefore, network excitability changes could be due to a receptor supersensitivity to L-DOPA or its metabolites (8) in the chronically transected spinal cord. This, however, could not be the sole explanation for the relative increase in excitability in the present experiments because in the chronic cats spontaneous nerve discharges were observed prior to L-DOPA administration. On the other hand, the cycle times of nerve discharges of the acute spinal cats are generally much longer than any reported by investigators for freely walking animals whether over ground (21) or on a treadmill (16, 21). This may indicate that the neuronal networks within the spinal cord of the acutely prepared cat are significantly depressed at the time of recording (2 to 4 h posttransection). This suggests that the level of excitability of the neuronal networks within the chronic cat’s spinal cord more nearly approx- imates the normal condition than it does in the acutely spinalized cat.
Although receptor supersensitivity is most likely not solely responsible for the increased excitability of the chronically isolated spinal cord, L- DOPA does affect the bursting activity seen in the nerves of chronic preparations. A significant linear relationship between burst duration and cycle times and a precise relationship between the onset of ipsilateral flexor and extensor nerve activity during a given cycle were observed less frequently in the chronicly prepared animals prior to the administration of the drugs. Unfortunately, the mechanism of L-DOPA action on the chronicly transected spinal cord is not well understood. Nevertheless, the administration of L-
DOPA does promote the production of coordinated discharge patterns in flexor and extensor nerves.
Rhythmicity of neuronal activity was another major difference in the acute and chronic preparations. Although the chronic preparations after L-DOPA demonstrated coordinated discharge patterns between nerves which were similar in some respects to locomotor-like patterns, burst duration and cycle times of neuronal activity were quite variable from cycle to cycle (Fig. 1 B). This large variability in cycle times and burst durations was never observed in the acute preparations (Fig. 1A). When the typical coordinated bursting pattern began in the acute preparation it was maintained for many hours with little variation in either cycle time or burst duration. This rhythmic activity was all but absent in the recordings from the chronic preparations. Whereas the burst durations were highly correlated with cycle

SPONTANEOUS NEURAL BURSTING IN CHRONIC SPINAL CATS 525
times the mechanism for maintaining the near periodicity of bursting activity was virtually inoperative in the chronic preparations.
It has been shown that the adult spinalized cat, in which the spinal cord was transected at an early age, is capable of treadmill locomotion and can produce rhythmic EMG patterns of activity in hind limb muscles that resemble normal treadmill locomotor patterns in many respects (18). The absence of these stable rhythmic patterns of activity in the chronic preparation deprived of phasic peripheral feedback after L-DOPA administration suggests that phasic peripheral feedback is quite important for the production of stable, rhythmic discharge patterns in a cat deprived of normal spinal cord development. This is not the case in acutely spinalized cats in which L-DOPA or nialamide does produce stable rhythmic patterns of activity. In the 2-week-old kitten, the age at which our chronicly spinalized cats were operated, adult locomotor activities have not yet fully developed ( 17, 20). However, the results presented here have shown that the immature spinal cord, isolated from all supraspinal input, is capable of developing the ability to produce rudimentary alternating patterns of flexor and extensor activity in the absence of phasic peripheral feedback.
REFERENCES
1. ANDERSON, E. G., AND L. 0. HALGERSON. 1966. The distribution of 5-hydroxytryptamine and norepinepherine in cat spinal cord. J. Neurochem. 13: 479-485.
2. CARLSSON, A., B. FALCK, K. FUXE, AND N. A. HILLARP. 1964. Cellular localization of monamines in the spinal cord. Acta Physiol. Stand. 60: I 12-119.
3. CHANDLER, S. H., L. L. BAKER. AND L. J. GOLDBERG. 1984. Characterization of synaptic potentials in hindlimb extensor motoneurons during L-DOPA induced fictive locomotion in acute and chronic spinal cats. Brain Res. 303: 91-100.
4. DAHLSTROM, A., AND K. FUXE. 1965. Evidence for the existence of monoamine neurons in the central nervous system. Acla Phgsiol. Scund. 64(Suppl. 247): I-86.
5. FORSSBERG, H., S. GRILLNER, AND J. HALBERTSMA. 1980. The locomotion of the low spinal cat. 1. Coordination within a hindlimb. .4&z Ph,vsiol. Stand. 108: 269-28 1.
6. FORSSBERG, H.. S. GRILLNER. J. HALBERTSMA, AND S. ROSSIGNOL. 1980. The locomotion of the low spinal cat 2: Interlimb coordination. Acfa Physiol. Stand 108: 283-295.
7. FORSSBERG, H.. AND S. GRILLNER. 1973. The locomotion of the acute spinal cat injected with clonidine i.v. Brain Rex 50: 184- 186.
8. GENTLEMAN, S., M. PARENTI, J. W. COMMISSIONG, AND N. H. NEFF. Dopamine-activated adenylate cyclase of spinal cord: supersensitivity following transection of the cord. Bruin Rex 210: 271-275.
9. GOSLOW, G. E., R. M. REINKING, AND D. G. STUART. 1973. The cat step cycle: hind limb joint angles and muscle lengths during unrestrained locomotion. J. Morphol. 141: l-41.
10. GRILLNER, S., AND P ZANGGER. 1979. On the central generation of locomotion in the low spinal cat. Exp. Brain Rex 34: 24 1-26 I.
I I. GRILLNER, S., AND P. ZANCCER. 1974. Locomotor movements generated by the deafferented spinal cord. Am Physiol. Scund. 91: 38A-39A.

526 BAKER, CHANDLER, AND GOLDBERG
12. GRILLNER, S. 1975. Locomotion in vertebrates: central mechanisms and reflex interaction. Physiol. Rrv. 55: 247-304.
13. JANKOWSKA, E., M. G. M. JUKES, S. LUND, AND A. LUNDBERG. 1967. The effect of DOPA on the spinal cord. 5. Reciprocal organization of pathways transmitting excitatory action to alpha motoneurons of flexors and extensors. Acfa Physiol. Scat& 70: 369- 388.
14. MAGNUSSON, T., AND E. ROSENGREN. 1963. Catecholamines of the spinal cord normally and after transection. Experientiu 19: 229-230.
15. MEISEL, R. L.. AND B. RAKERD. 1982. Induction of hindlimb stepping movements in rats spinally transected as adults or as neonates. Bruin Res. 240: 353-356.
16. RASMUSSEN, S., A. K. CHAN, ANDG. E. GOSLOW. 1978. The cat step cycle: electromyographic patterns for hindlimb muscles during posture and unrestrained locomotion. J. Morphol. 155: 253-270.
17. SCHEIBEL, M. E., AND A. B. SCHEIBEL. 1970. Developmental relationship between spinal motoneuron dendrite bundles and patterned activity in the hind limb of cats. E.up. Neural. 29: 328-335.
18. SMITH, J. L., L. A. SMITH, R. F. ZERNICKE, AND M. HOY. 1982. Locomotion in exercised and nonexercised cats cordotomized at two or twelve weeks of age. Ex~. Neural. 76: 393-413.
19. TEN CATE, J. 1962. Innervation of locomotor movements by the lumbosacral cord in birds and mammals. J. Exp. Biol. 39: 239-242.
20. VILLABLANCA, J. R.. AND C. E. OLMSTEAD. 1979. Neurological development of kittens. Dev. Psychobiol. 12: 10 I- 127.
2 1. WETZEL, M. C., A. E. ATWATER, J. V. WAIT, AND D. G. STUART. 1977. Neural implications of different profiles between treadmill and overground locomotion timing in cats. J. Neurophysiol. 38: 492-501.




















