
Coordinated and Spatial Upregulation of Arc in Striatonigral Neurons Correlates With L-Dopa-Induced...
12
J Neuropathol Exp Neurol Copyright Ó 2005 by the American Association of Neuropathologists, Inc. Vol. 64, No. 11 November 2005 pp. 936–947 ORIGINAL ARTICLE Coordinated and Spatial Upregulation of Arc in Striatonigral Neurons Correlates With L-Dopa-Induced Behavioral Sensitization in Dyskinetic Rats Ve ´ronique Sgambato-Faure, PhD, Virginie Buggia, MSc, Franc xois Gilbert, MSc, Daniel Le ´vesque, PhD, Alim-Louis Benabid, MD, PhD, and Franc xois Berger, MD, PhD Abstract Although oral administration of L-Dopa remains the best therapy for Parkinson disease, its long-term administration causes the appear- ance of abnormal involuntary movements such as dyskinesia. Although persistent striatal induction of some genes has already been asso- ciated with such pathologic profiles in hemiparkinsonian rats, molec- ular and cellular mechanisms underlying such long-term adaptations remain to be elucidated. In this study, using a rat model of L-Dopa- induced dyskinesia, we report that activity regulated cytoskeletal (Arc)- associated protein is strongly upregulated in the lesioned striatum and that the extent of its induction further varies according to the occurrence or absence of locomotor sensitization. Moreover, Arc is preferentially induced, along with FosB, nur77, and homer-1a, in striatonigral neurons, which express mRNA encoding the precursor of dynorphin. Given the likely importance of Arc in the regulation of cytoskeleton during synaptic plasticity, its upregulation supports the hypothesis that a relationship exists between cytoskeletal modifica- tions and the longlasting action of chronically administrated L-Dopa. Key Words: Arc, Dynorphin, Dyskinesia, homer-1a, L-Dopa, Locomotor sensitization, nur77. INTRODUCTION Parkinson disease (PD) is a neurologic disorder characterized by an extensive loss of dopaminergic neurons in the substantia nigra pars compacta (1, 2). The loss of dopamine within the striatum disturbs the functional organi- zation of the basal ganglia networks by causing an imbalance between the 2 efferent striatal output pathways (direct: striatonigral; indirect: striatopallido–subthalamonigral) (3). Dopamine deficiency causes the motor symptoms of PD such as akinesia, rigidity, and tremor (3, 4). Despite the intense development of new therapeutic approaches (5), administra- tion of the natural dopamine precursor levodopa (L-Dopa) still remains the best symptomatic treatment (6). However, its long- term use is commonly associated with side effects such as motor fluctuations and abnormal involuntary movements, namely L-Dopa-induced dyskinesia (LID) (7–10; for review, [11]). The genesis of LID is not fully understood, but major factors include degree of presynaptic nigral denervation (12– 15); onset, dose, and manner of administration of L-Dopa (16); sensitization of dopaminergic receptors and their pulsatile stimulation (17, 18); imbalance between the output structures of basal ganglia (3, 4); as well as alteration of activity and discharge pattern of neurons of basal ganglia (19, 20). However, the precise mechanisms by which chronic L-Dopa leads to such long-term motor alterations require elucidation. From experimental studies using animal models of LID (i.e. dopaminergic-lesioned animals chronically treated with L-Dopa given intermittently to simulate dosing schedules in parkinsonian patients), it has been shown that abnormal in- formation storage in the corticostriatal pathways correlates with LID (21). Moreover, it has been shown that in the striatum, persistent molecular alterations noticeably related to glutamate signaling are involved in LID (22, 23). Many of these gene modulations are observed in striatonigral-projecting cells of the striatum, suggesting that dynorphin-containing cells may be associated with the development of behavioral sensitization that occurs in animal models of PD after L-Dopa treatment, although the involvement of opioid receptors is still a matter of debate (24–29). Often compared with an aberrant form of motor learning (30, 31), LID might also involve synaptic re- modeling. Among the cytoskeleton-related genes, the activity- regulated cytoskeletal (Arc)-associated gene is of particular interest. Initially identified in differential screens for seizure- stimulated hippocampal mRNA (32, 33), Arc mRNA rapidly localizes to discrete dendritic regions that have received direct synaptic stimulation (34, 35). Moreover, Arc is required for long-term potentiation as well as memory in the hippocampus (36–38). In the present study, we asked whether chronic L-Dopa increased the expression of Arc and whether its L-Dopa-driven induction could be correlated with dyskinesia or behavioral sensitization. Using hemiparkinsonian rats that were chron- ically treated with L-Dopa and developed dyskinetic profiles, we observed that a strong upregulation of Arc at both gene and protein levels, in response to L-Dopa administration in the dopaminergic-depleted striatum, was spatially coincident with From Inserm U318 (VS-F, VB, A-LB, FB), Laboratoire de Neurosciences Pre ´cliniques, CHU Pavillon B, Grenoble, France; and Centre de recherche en Neuroscience (FG, DL), CHUL, Sainte Foy, Que ´bec, Canada. Send correspondence and reprint requests to: Ve ´ronique Sgambato-Faure, PhD, INSERM U704, Dynamique des Re ´seaux Neuronaux, 2280 Rue de la Piscine, BP53, 38041 Grenoble cedex 9, France; E-mail: veronique. [email protected] This work was supported by grants from Institut National de la Sante ´ et de la Recherche Me ´dicale. 936 J Neuropathol Exp Neurol Volume 64, Number 11, November 2005
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Coordinated and Spatial Upregulation of Arc in Striatonigral Neurons Correlates With L-Dopa-Induced...

J Neuropathol Exp NeurolCopyright � 2005 by the American Association of Neuropathologists, Inc.
Vol. 64, No. 11November 2005
pp. 936–947
ORIGINAL ARTICLE
Coordinated and Spatial Upregulation of Arc in StriatonigralNeurons Correlates With L-Dopa-Induced Behavioral
Sensitization in Dyskinetic Rats
Veronique Sgambato-Faure, PhD, Virginie Buggia, MSc, Francxois Gilbert, MSc, Daniel Levesque, PhD,
Alim-Louis Benabid, MD, PhD, and Francxois Berger, MD, PhD
AbstractAlthough oral administration of L-Dopa remains the best therapy
for Parkinson disease, its long-term administration causes the appear-
ance of abnormal involuntary movements such as dyskinesia. Although
persistent striatal induction of some genes has already been asso-
ciated with such pathologic profiles in hemiparkinsonian rats, molec-
ular and cellular mechanisms underlying such long-term adaptations
remain to be elucidated. In this study, using a rat model of L-Dopa-
induced dyskinesia, we report that activity regulated cytoskeletal (Arc)-
associated protein is strongly upregulated in the lesioned striatum and
that the extent of its induction further varies according to the
occurrence or absence of locomotor sensitization. Moreover, Arc is
preferentially induced, along with FosB, nur77, and homer-1a, in
striatonigral neurons, which express mRNA encoding the precursor
of dynorphin. Given the likely importance of Arc in the regulation of
cytoskeleton during synaptic plasticity, its upregulation supports the
hypothesis that a relationship exists between cytoskeletal modifica-
tions and the longlasting action of chronically administrated L-Dopa.
Key Words: Arc, Dynorphin, Dyskinesia, homer-1a, L-Dopa,
Locomotor sensitization, nur77.
INTRODUCTIONParkinson disease (PD) is a neurologic disorder
characterized by an extensive loss of dopaminergic neuronsin the substantia nigra pars compacta (1, 2). The loss ofdopamine within the striatum disturbs the functional organi-zation of the basal ganglia networks by causing an imbalancebetween the 2 efferent striatal output pathways (direct:striatonigral; indirect: striatopallido–subthalamonigral) (3).Dopamine deficiency causes the motor symptoms of PD suchas akinesia, rigidity, and tremor (3, 4). Despite the intensedevelopment of new therapeutic approaches (5), administra-tion of the natural dopamine precursor levodopa (L-Dopa) still
remains the best symptomatic treatment (6). However, its long-term use is commonly associated with side effects such asmotor fluctuations and abnormal involuntary movements,namely L-Dopa-induced dyskinesia (LID) (7–10; for review,[11]). The genesis of LID is not fully understood, but majorfactors include degree of presynaptic nigral denervation (12–15); onset, dose, and manner of administration of L-Dopa (16);sensitization of dopaminergic receptors and their pulsatilestimulation (17, 18); imbalance between the output structuresof basal ganglia (3, 4); as well as alteration of activity anddischarge pattern of neurons of basal ganglia (19, 20).However, the precise mechanisms by which chronic L-Dopaleads to such long-term motor alterations require elucidation.
From experimental studies using animal models of LID(i.e. dopaminergic-lesioned animals chronically treated withL-Dopa given intermittently to simulate dosing schedules inparkinsonian patients), it has been shown that abnormal in-formation storage in the corticostriatal pathways correlates withLID (21). Moreover, it has been shown that in the striatum,persistent molecular alterations noticeably related to glutamatesignaling are involved in LID (22, 23). Many of these genemodulations are observed in striatonigral-projecting cells ofthe striatum, suggesting that dynorphin-containing cells maybe associated with the development of behavioral sensitizationthat occurs in animal models of PD after L-Dopa treatment,although the involvement of opioid receptors is still a matterof debate (24–29). Often compared with an aberrant form ofmotor learning (30, 31), LID might also involve synaptic re-modeling. Among the cytoskeleton-related genes, the activity-regulated cytoskeletal (Arc)-associated gene is of particularinterest. Initially identified in differential screens for seizure-stimulated hippocampal mRNA (32, 33), Arc mRNA rapidlylocalizes to discrete dendritic regions that have received directsynaptic stimulation (34, 35). Moreover, Arc is required forlong-term potentiation as well as memory in the hippocampus(36–38).
In the present study, we asked whether chronic L-Dopaincreased the expression of Arc and whether its L-Dopa-driveninduction could be correlated with dyskinesia or behavioralsensitization. Using hemiparkinsonian rats that were chron-ically treated with L-Dopa and developed dyskinetic profiles,we observed that a strong upregulation of Arc at both gene andprotein levels, in response to L-Dopa administration in thedopaminergic-depleted striatum, was spatially coincident with
From Inserm U318 (VS-F, VB, A-LB, FB), Laboratoire de NeurosciencesPrecliniques, CHU Pavillon B, Grenoble, France; and Centre de rechercheen Neuroscience (FG, DL), CHUL, Sainte Foy, Quebec, Canada.
Send correspondence and reprint requests to: Veronique Sgambato-Faure,PhD, INSERM U704, Dynamique des Reseaux Neuronaux, 2280 Rue dela Piscine, BP53, 38041 Grenoble cedex 9, France; E-mail: [email protected]
This work was supported by grants from Institut National de la Sante et de laRecherche Medicale.
936 J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005

that of the FosB-like proteins, homer-1a, nur77 (also known asnerve growth factor-inducible B [NGFI-B]), and preprodynor-phin (PPDyn) mRNA levels, in the striatal subregion accordingto the subtype of motor behavior displayed. Moreover, weobserved that Arc is colocalized with PPDyn mRNA in neuronsof the direct striatonigral pathway. Finally, we found thatL-Dopa-induced motor behavioral response and Arc upregulationare not affected by a modulation of opioid receptor activity.Altogether, these findings highlight a possible involvement ofArc in the development or maintenance of LID and locomotorsensitization and further support a role for cytoskeletal remod-eling in the regulation of such long-term motor alterations.
MATERIALS AND METHODSThe animal experimental protocols performed in this study
strictly conformed to the guidelines of the French Agricultureand Forestry Ministry and were approved by INSERM.
Lesion SurgeryMale adult Wistar rats, weighing 260 to 300 g, were
maintained on a 12-hour light/dark cycle with food and tapwater available ad libitum. All animals in this study wereanesthetized with chloral hydrate (400 mg/kg) and subjected tounilateral 6-OHDA lesion in the left substantia nigra parscompacta (SNc) using a conventional stereotaxic instrument(David Kopf, Phymep, Paris, France). The lesions were per-formed according to a standard procedure. Briefly, 12 mg of6-OHDA-HCl (Sigma, Saint Quentin Fallavier, France) dis-solved in 0.1% ascorbate saline were injected in the left SNc atthe following coordinates (according to the atlas of Paxinosand Watson (39)) in millimeters and relative to Bregma andcortical surface: A = 5.3; L = 2.1; H = 7.8. To preserve norad-renergic fibers, all animals were intraperitoneally (IP) injectedwith desipramine-HCl (25 mg/kg; Sigma) at least 30 minutesbefore 6-OHDA injection.
Amphetamine-Induced Rotational BehaviorTen to 15 days after lesion, animals were tested for
amphetamine-induced rotation. Rats received amphetaminesulfate salt (2.5 mg/kg, IP; Sigma) and were placed in a verticalcylinder. After a period of 30 to 45 minutes, circling behaviorwas observed for 15 minutes. Only animals showing individ-ual means $5 full turns/minute in the direction ipsilateral tothe lesion were selected for the study. This score has beenshown to correspond to at least 90% of DA depletion in thestriatum (14). The effectiveness of the lesion was furtherconfirmed postmortem by means of tyrosine hydroxylase (TH)immunohistochemistry and preproenkephalin (PPE) or pre-prodynorphin (PPDyn) in situ hybridization.
Drug TreatmentsAll the drugs were purchased from Sigma. Four to
5 weeks after lesion, rats were treated with chronic (twice dailyfor 10 days) IP injection of physiological saline (lesioned-onlyanimals, n = 3) or L-Dopa (L-3,4-Dihydroxyphenylalaninemethyl ester hydrochloride) (50 mg/kg) combined withbenserazide hydrochloride (10 mg/kg) (L-Dopa-treated ani-mals, n = 12). Benserazide was given to prevent decarbox-ylation of L-Dopa in the periphery, as is common in clinical
practice. Rats were observed individually after each injection.Another small group of hemiparkinsonian rats (n = 3) weretreated for a 3-week period with lower doses of L-Dopa(10 mg/kg) and benserazide (2 mg/kg). For pharmacologicstudies, 4 groups of hemiparkinsonian rats (n = 12) were treatedwith L-Dopa (at 50 mg/kg plus benserazide at 10 mg/kg)twice daily for 10 days as follows: administration of L-Dopaalone, coadministration of L-Dopa plus naloxone dihydro-chloride (at 10 mg/kg), coadministration of L-Dopa plusU-50,488H (trans-3,4-di-chloro-N-methyl-N[2]-[(1-pyrrolidinyl)cyclohexyl] benzeneacetamide methane-sulfonate salt) (at1 mg/kg), and administration of L-Dopa plus an acuteinjection of nor-binaltorphimine di-hydrochloride (20 mg/kg)1 hour before L-Dopa on the last day of treatment.
Motor Behavior AnalysisBriefly, individual rats were observed daily in their home
cage 1 hour after the injection, when the response to L-Dopawas maximal. Movements were considered as dyskinetic whenboth of the following criteria were fulfilled: induction byL-Dopa of repetitive and abnormal movements of the bodycontralateral to the lesion. Abnormal involuntary movementswere subdivided into 3 subtypes: forelimb dyskinesia, orolin-gual dyskinesia, and axial dyskinesia (40). The severity of eachdyskinesia subtype was evaluated for each animal by assigninga rating scale (0–4) (14, 41). Locomotor sensitization was alsoevaluated by counting the number of contralateral rotations perminute. Finally, animals were videotaped during the treatmentand before death.
Tissue PreparationTwo or 24 hours after the last L-Dopa injection, animals
received a lethal dose of chloral hydrate and were killed byintracardiac perfusion of Ringer�s solution followed by 4%paraformaldehyde in 0.1 M Na2HPO4/NaH2PO4 buffer, pH 7.5(phosphate buffer). Brains were removed and postfixed in thesame fixative solution for 2 hours, washed overnight in 0.1 Mphosphate buffer containing 15% sucrose, and then frozen.Rostrocaudal series of coronal sections (20 mm) were cut ona cryostat and then kept in a solution containing 30% ethyleneglycol, 30% glycerol, 0.1 M phosphate buffer, and 0.1% diethylpyrocarbonate (DEPC) at 20�C until they were processed foreither in situ hybridization or immunohistochemistry.
In Situ HybridizationThe antisense probes used in this study were a[35-S]UTP
radiolabeled riboprobes. Preproenkephalin and homer-1acDNAs were linearized after Apa I digestion and transcribedwith SP6 RNA polymerase. The preprodynorphin and Arcantisense probes were linearized after EcoR I digestion andtranscribed with T7 RNA polymerase. The nur77 (NGFI-B)antisense probe was linearized with BamH I and transcribedwith T3 (42). Transcription reaction contained 6 mL a[35S]-UTP (20 mCi/mL; Amersham Biosciences, Orsay, France),250 mM adenosine triphosphate (ATP), cytidine triphosphate(CTP), and guanosine triphosphate (GTP), and unlabeleduridine triphosphate (UTP) (10.5 mM); the reaction wasincubated at 37�C for 2 hours. After DNase I digestion for
q 2005 American Association of Neuropathologists, Inc. 937
J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005 Arc Correlates With L-Dopa-Induced Motor Behavior
15 minutes, the labeled RNA was purified on microspin G50columns (Amersham Biosciences). Free-floating sections weremounted on SuperFrost Plus slides in RNase-free conditions.Once the sections were dried, in situ hybridization was per-formed at 60�C in humid chambers for 16 hours according tothe previous protocol (43). After coverslips were removed in4 3 SSC and 10 mM DTT, the slides were washed in the samesolution for 1 hour at room temperature and then in 50%formamide, 10 mM Tris-HCl, pH 8, 75 mM NaCl, and 2.5 mMEDTA. Sections were treated with RNase A (20 mg/mL;Sigma) in 400 mM NaCl, 10 mM Tris-HCl, pH 7.5, and50 mM EDTA for 30 minutes at 37�C and then were rinsed for15 minutes at 60�C in 2 3 SSC followed by 0.1 3 SSC. Afterdehydration, the sections were air-dried and exposed withBiomax-MR films (Kodak).
ImmunohistochemistryFree-floating sections were rinsed in Tris-buffered saline
(TBS; 0.25M Tris and 0.25M NaCl, pH 7.5), incubated5 minutes in TBS containing 3% H2O2 and 10% methanol, andthen rinsed 3 times for 10 minutes each in TBS. After15-minute incubation in 0.2% triton X-100 in TBS, the sectionswere rinsed 3 times in TBS. These were incubated with theprimary antibody (anti-FosB 1/400 rabbit polyclonal H-75[SC-7203] from Tebu-Bio, Le Parray en Yvelines, France),anti-TH 1/2000 rabbit polyclonal from Jacques Boy (Reims,France), or anti-Arc 1/300 mouse monoclonal from BDBiosciences (Pont de Claix, France) for 72 hours at 4�C. After3 rinses in TBS, the sections were then incubated 2 hours at4�C with the corresponding secondary biotinylated antibodyin TBS. After being washed, the sections were incubated90 minutes at room temperature in avidin-biotin-peroxidasecomplex (ABC) solution (final dilution, 1/50; Vector Labora-
tories; ABCYS, Paris, France). The sections were then rinsed2 times in TBS and 2 times in TB (0.25M Tris, pH 7.5) for10 minutes each, placed in a solution of TB containing 0.1%3,3#-diaminobenzidine (DAB; 50 mg/100 mL), and developedby H2O2 addition (0.02%).
Double Labeling CombiningImmunohistochemistry andIn Situ Hybridization
Free-floating sections were processed for FosB or Arcimmunohistochemistry as described previously, except that allbuffers used were autoclaved. Sections were mounted onSuperFrost Plus slides in RNase-free conditions and air-driedfor at least 60 minutes followed by the indicated in situhybridization. The slides were then coated with LM-1 emulsion(Amersham Biosciences) and exposed. After development,slides were mounted with Eukitt.
Double In Situ HybridizationThe double in situ hybridization procedure was
performed as previously described (42). Briefly, the proportionof nur77 colocalization with PPE or PPDyn mRNA wasevaluated using double in situ hybridization with a [35S]UTP-labeled nur77 probe and a nonradioactive digoxigenin (Dig)-labeled PPE or PPDyn riboprobe. PPE and PPDyn riboprobeswere labeled with using the Riboprobe System of Promega(Madison, WI) with the Dig-RNA labeling mix (RocheDiagnostics, Laval, Canada). Double in situ hybridization wasperformed in the same conditions as for single in situ pro-cedure. The Dig-cRNA probe (approximately 50 ng) wassimply added in the same hybridization solution with theradioactive (2 3 106 cpm) cRNA probe for nur77. Anadditional step using a 50% formamide solution in 2 3 SSC
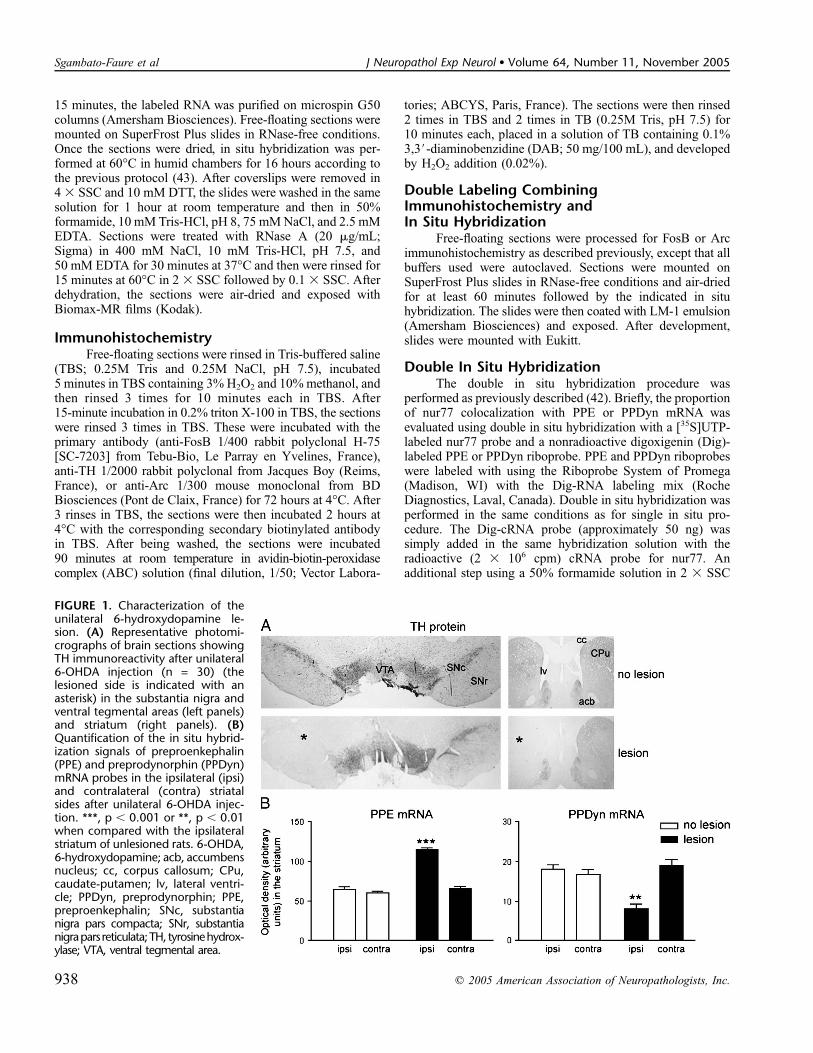
FIGURE 1. Characterization of theunilateral 6-hydroxydopamine le-sion. (A) Representative photomi-crographs of brain sections showingTH immunoreactivity after unilateral6-OHDA injection (n = 30) (thelesioned side is indicated with anasterisk) in the substantia nigra andventral tegmental areas (left panels)and striatum (right panels). (B)Quantification of the in situ hybrid-ization signals of preproenkephalin(PPE) and preprodynorphin (PPDyn)mRNA probes in the ipsilateral (ipsi)and contralateral (contra) striatalsides after unilateral 6-OHDA injec-tion. ***, p , 0.001 or **, p , 0.01when compared with the ipsilateralstriatum of unlesioned rats. 6-OHDA,6-hydroxydopamine; acb, accumbensnucleus; cc, corpus callosum; CPu,caudate-putamen; lv, lateral ventri-cle; PPDyn, preprodynorphin; PPE,preproenkephalin; SNc, substantianigra pars compacta; SNr, substantianigrapars reticulata;TH, tyrosinehydrox-ylase; VTA, ventral tegmental area.
938 q 2005 American Association of Neuropathologists, Inc.
Sgambato-Faure et al J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005
buffer after hybridization was performed to reduce nonspecificDig labeling. Detection of Dig-labeled RNA was performedwith an anti-Dig antibody conjugated to alkaline phosphatase(Boehringer Mannheim, Laval, Canada) visualized usinga nitroblue tetrazolium chloride, 5-bromo-4-chloro-3-indolylphosphate, and levamisole chromogen solution. Slides werethen dipped in LM-1 photographic emulsion (Amersham,Oakville, Ontario, Canada) melted at 42�C, air-dried, andstored in the dark for 12 days at 4�C. The emulsion wasdeveloped in D-19 developer and fixed (Kodak, New Haven,CT). Slides were coverslipped using a water-soluble mountingmedium (Permafluor; Lipshaw Immunon, Pittsburgh, PA).Single- or double-labeled cells were visualized and manuallycounted under bright-field illumination with a Zeiss photo-microscope at a magnification of 4003. Cell counts wereperformed in the lateral part of the striatum on 4 differentsections (3 fields per section).
Data AnalysisTissue sections were examined under light microscope.
TH, FosB, and Arc-positive neurons were digitalized atvarious magnifications with a computerized image analyzer(Analysis; Soft Imaging System, Munster, Germany). Cellcounts were performed in the lateral part of the striatumipsilateral to the SNc lesion (3 fields per section). Cells labeledwith PPE, PPDyn, and Arc probes were visualized by theaccumulation of silver grains over the soma. Densitometricmeasurements of mRNA signals were obtained from auto-radiograms using the NIH Image J 1.29 software (WayneRasband, NIMH) after subtraction of background value. Theywere performed in the lateral part of the striatum on 4 differentsections per rat. Histogram shows mean 6 standard error ofmean. Statistical analyses were assessed by analysis ofvariance with multiple comparisons (Newman-Keuls multiplecomparison test).
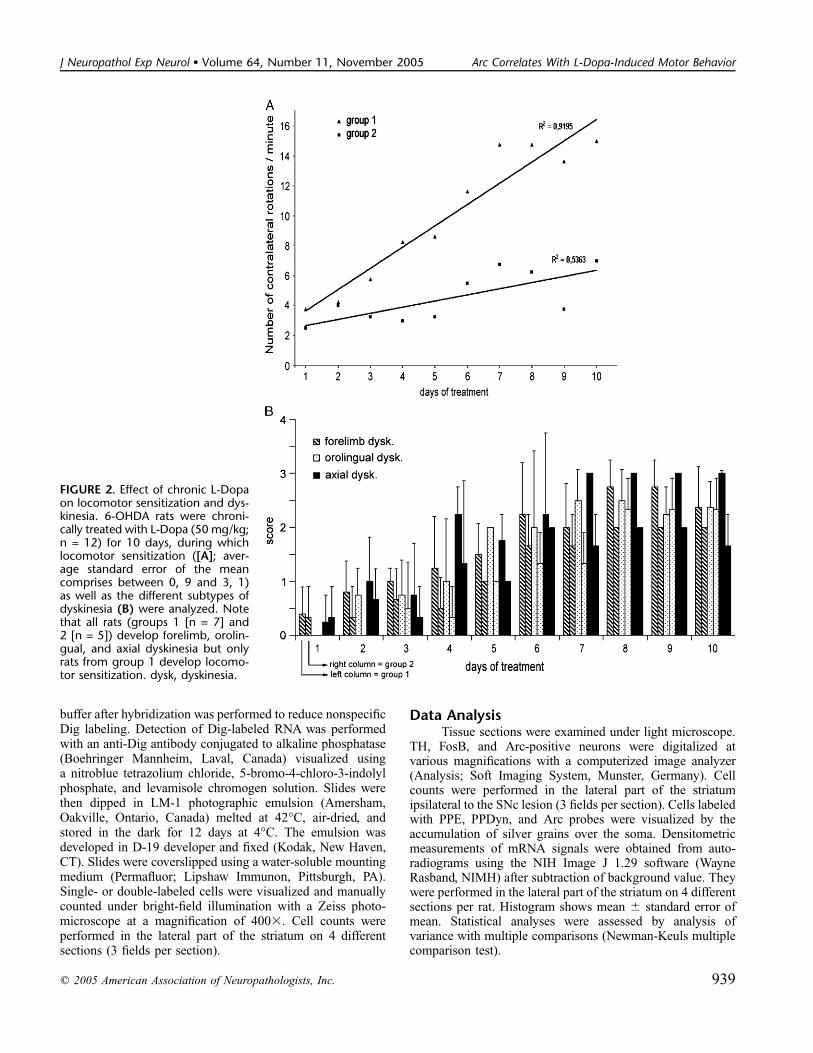
FIGURE 2. Effect of chronic L-Dopaon locomotor sensitization and dys-kinesia. 6-OHDA rats were chroni-cally treated with L-Dopa (50 mg/kg;n = 12) for 10 days, during whichlocomotor sensitization ([A]; aver-age standard error of the meancomprises between 0, 9 and 3, 1)as well as the different subtypes ofdyskinesia (B) were analyzed. Notethat all rats (groups 1 [n = 7] and2 [n = 5]) develop forelimb, orolin-gual, and axial dyskinesia but onlyrats from group 1 develop locomo-tor sensitization. dysk, dyskinesia.
q 2005 American Association of Neuropathologists, Inc. 939
J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005 Arc Correlates With L-Dopa-Induced Motor Behavior
RESULTS
Characterization of the Unilateral6-hydroxydopamine Lesion
The dopaminergic lesion was evaluated postmortem byhistochemical analysis (Fig. 1). A complete loss of TH im-munostaining was observed ipsilaterally in both the SNc andthe striatum of all animals used in the present study (Fig. 1A).The levels of denervation were the same in all groups of
animals. An analysis of the densitometric measurements of THimmunostaining in both SNc and ventral tegmental area (VTA)did not produce a statistical difference between the 2 groups(data not shown). At the striatal level, mRNA levels encodingPPE (p , 0.001) and PPDyn (p , 0.01) were respectivelyinduced or repressed by the lesion (Fig. 1B).
Evaluation of L-Dopa-Induced Motor BehaviorThe motor behavior of hemiparkinsonian rats was
observed and evaluated as described in the ‘‘Materials andMethods’’ section. Although rats treated chronically with thevehicle did not show any abnormal motor activity (data notshown), rats treated with high doses of L-Dopa could besubdivided into 2 groups according to their rotational behavior(Fig. 2A). The first group (group 1, n = 7) developed severelocomotor sensitization over time (linear regression coefficientR2 = 0.9195, p , 0.001), whereas the second group (group 2,n = 5) displayed almost no locomotor sensitization (R2 =0.5363, p = 0.057) despite the fact that they received the sameL-dopa regimen and displayed the same levels of denervation.The 3 subtypes of dyskinesia (i.e. forelimb, orolingual, andaxial) were also evaluated (Fig. 2B). For each, a gradualincrease of the scores was observed over time (Fig. 2B), but nofurther difference was noted between the first and secondgroups (compare left and right columns for each dyskinesiasubtype). The dispersion of the results reflects the variability inthe severity of the dyskinesia displayed by each animal. Nodyskinesia was observed for vehicle-treated hemiparkinsonianrats (data not shown).
Effect of Chronic L-Dopa Treatment on FosBExpression in the Depleted Striatum
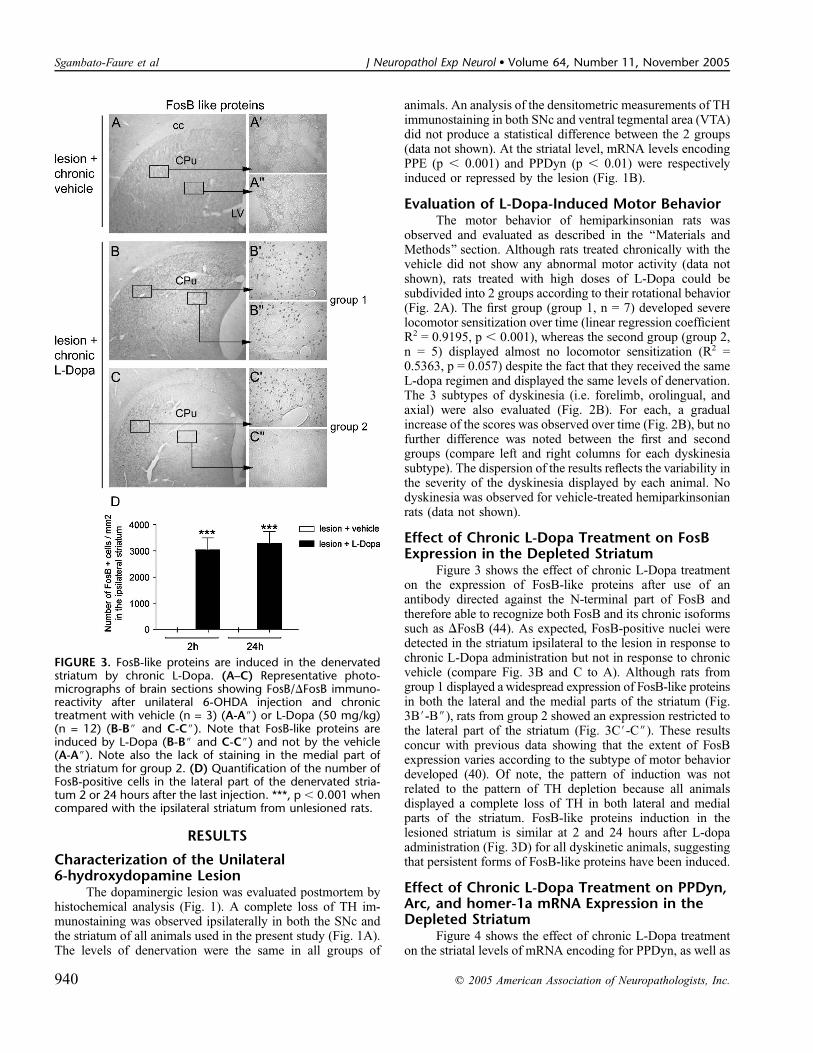
Figure 3 shows the effect of chronic L-Dopa treatmenton the expression of FosB-like proteins after use of anantibody directed against the N-terminal part of FosB andtherefore able to recognize both FosB and its chronic isoformssuch as DFosB (44). As expected, FosB-positive nuclei weredetected in the striatum ipsilateral to the lesion in response tochronic L-Dopa administration but not in response to chronicvehicle (compare Fig. 3B and C to A). Although rats fromgroup 1 displayed a widespread expression of FosB-like proteinsin both the lateral and the medial parts of the striatum (Fig.3B#-B$), rats from group 2 showed an expression restricted tothe lateral part of the striatum (Fig. 3C#-C$). These resultsconcur with previous data showing that the extent of FosBexpression varies according to the subtype of motor behaviordeveloped (40). Of note, the pattern of induction was notrelated to the pattern of TH depletion because all animalsdisplayed a complete loss of TH in both lateral and medialparts of the striatum. FosB-like proteins induction in thelesioned striatum is similar at 2 and 24 hours after L-dopaadministration (Fig. 3D) for all dyskinetic animals, suggestingthat persistent forms of FosB-like proteins have been induced.
Effect of Chronic L-Dopa Treatment on PPDyn,Arc, and homer-1a mRNA Expression in theDepleted Striatum
Figure 4 shows the effect of chronic L-Dopa treatmenton the striatal levels of mRNA encoding for PPDyn, as well as
FIGURE 3. FosB-like proteins are induced in the denervatedstriatum by chronic L-Dopa. (A–C) Representative photo-micrographs of brain sections showing FosB/DFosB immuno-reactivity after unilateral 6-OHDA injection and chronictreatment with vehicle (n = 3) (A-A$) or L-Dopa (50 mg/kg)(n = 12) (B-B$ and C-C$). Note that FosB-like proteins areinduced by L-Dopa (B-B$ and C-C$) and not by the vehicle(A-A$). Note also the lack of staining in the medial part ofthe striatum for group 2. (D) Quantification of the number ofFosB-positive cells in the lateral part of the denervated stria-tum 2 or 24 hours after the last injection. ***, p , 0.001 whencompared with the ipsilateral striatum from unlesioned rats.
940 q 2005 American Association of Neuropathologists, Inc.
Sgambato-Faure et al J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005
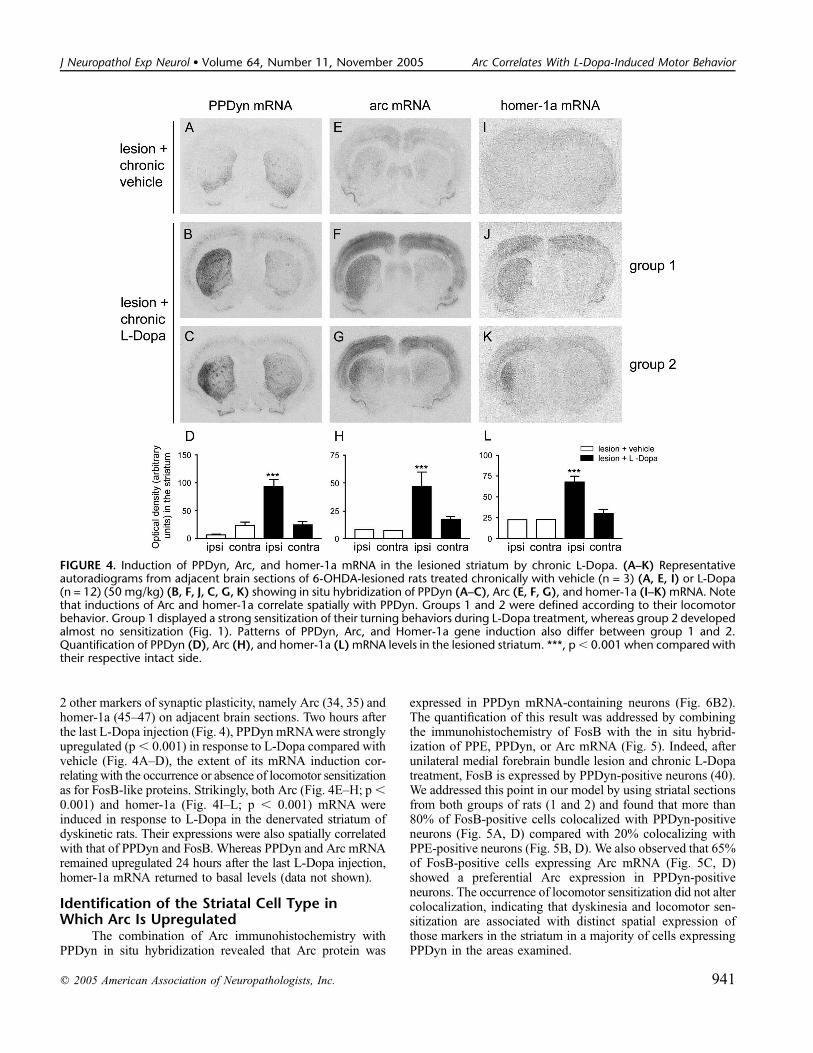
2 other markers of synaptic plasticity, namely Arc (34, 35) andhomer-1a (45–47) on adjacent brain sections. Two hours afterthe last L-Dopa injection (Fig. 4), PPDyn mRNAwere stronglyupregulated (p , 0.001) in response to L-Dopa compared withvehicle (Fig. 4A–D), the extent of its mRNA induction cor-relating with the occurrence or absence of locomotor sensitizationas for FosB-like proteins. Strikingly, both Arc (Fig. 4E–H; p ,0.001) and homer-1a (Fig. 4I–L; p , 0.001) mRNA wereinduced in response to L-Dopa in the denervated striatum ofdyskinetic rats. Their expressions were also spatially correlatedwith that of PPDyn and FosB. Whereas PPDyn and Arc mRNAremained upregulated 24 hours after the last L-Dopa injection,homer-1a mRNA returned to basal levels (data not shown).
Identification of the Striatal Cell Type inWhich Arc Is Upregulated
The combination of Arc immunohistochemistry withPPDyn in situ hybridization revealed that Arc protein was
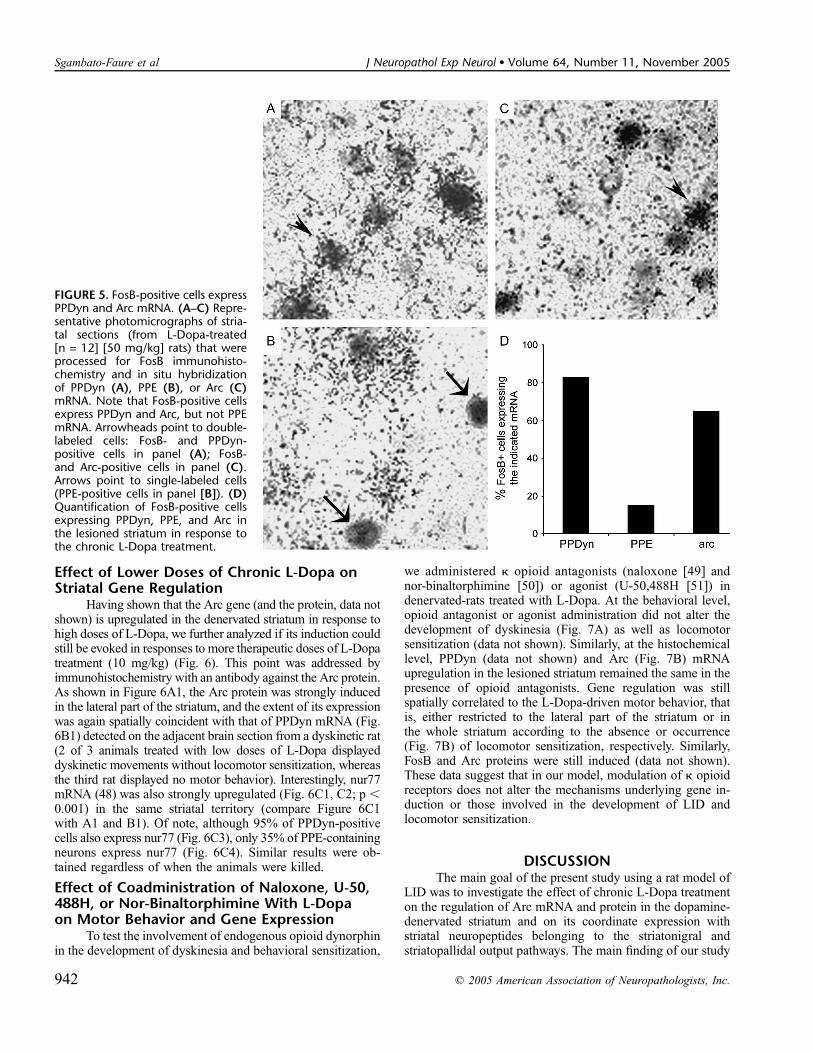
expressed in PPDyn mRNA-containing neurons (Fig. 6B2).The quantification of this result was addressed by combiningthe immunohistochemistry of FosB with the in situ hybrid-ization of PPE, PPDyn, or Arc mRNA (Fig. 5). Indeed, afterunilateral medial forebrain bundle lesion and chronic L-Dopatreatment, FosB is expressed by PPDyn-positive neurons (40).We addressed this point in our model by using striatal sectionsfrom both groups of rats (1 and 2) and found that more than80% of FosB-positive cells colocalized with PPDyn-positiveneurons (Fig. 5A, D) compared with 20% colocalizing withPPE-positive neurons (Fig. 5B, D). We also observed that 65%of FosB-positive cells expressing Arc mRNA (Fig. 5C, D)showed a preferential Arc expression in PPDyn-positiveneurons. The occurrence of locomotor sensitization did not altercolocalization, indicating that dyskinesia and locomotor sen-sitization are associated with distinct spatial expression ofthose markers in the striatum in a majority of cells expressingPPDyn in the areas examined.
FIGURE 4. Induction of PPDyn, Arc, and homer-1a mRNA in the lesioned striatum by chronic L-Dopa. (A–K) Representativeautoradiograms from adjacent brain sections of 6-OHDA-lesioned rats treated chronically with vehicle (n = 3) (A, E, I) or L-Dopa(n = 12) (50 mg/kg) (B, F, J, C, G, K) showing in situ hybridization of PPDyn (A–C), Arc (E, F, G), and homer-1a (I–K) mRNA. Notethat inductions of Arc and homer-1a correlate spatially with PPDyn. Groups 1 and 2 were defined according to their locomotorbehavior. Group 1 displayed a strong sensitization of their turning behaviors during L-Dopa treatment, whereas group 2 developedalmost no sensitization (Fig. 1). Patterns of PPDyn, Arc, and Homer-1a gene induction also differ between group 1 and 2.Quantification of PPDyn (D), Arc (H), and homer-1a (L) mRNA levels in the lesioned striatum. ***, p, 0.001 when compared withtheir respective intact side.
q 2005 American Association of Neuropathologists, Inc. 941
J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005 Arc Correlates With L-Dopa-Induced Motor Behavior
Effect of Lower Doses of Chronic L-Dopa onStriatal Gene Regulation
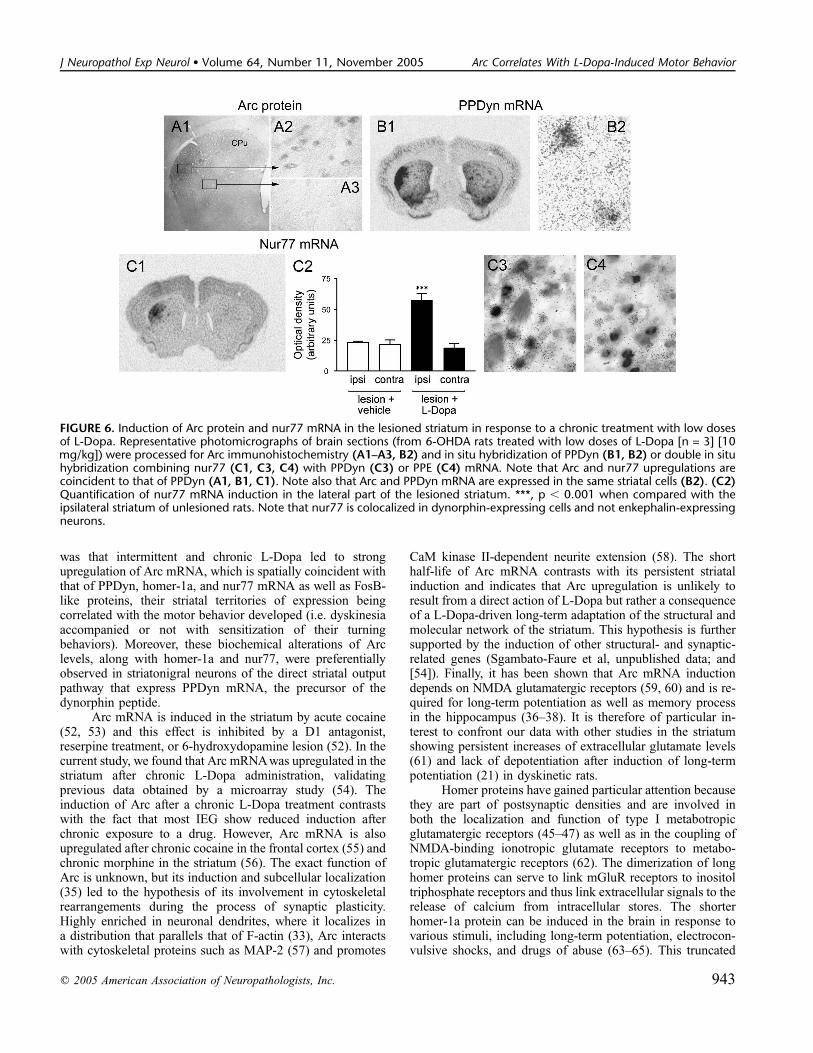
Having shown that the Arc gene (and the protein, data notshown) is upregulated in the denervated striatum in response tohigh doses of L-Dopa, we further analyzed if its induction couldstill be evoked in responses to more therapeutic doses of L-Dopatreatment (10 mg/kg) (Fig. 6). This point was addressed byimmunohistochemistry with an antibody against the Arc protein.As shown in Figure 6A1, the Arc protein was strongly inducedin the lateral part of the striatum, and the extent of its expressionwas again spatially coincident with that of PPDyn mRNA (Fig.6B1) detected on the adjacent brain section from a dyskinetic rat(2 of 3 animals treated with low doses of L-Dopa displayeddyskinetic movements without locomotor sensitization, whereasthe third rat displayed no motor behavior). Interestingly, nur77mRNA (48) was also strongly upregulated (Fig. 6C1, C2; p ,0.001) in the same striatal territory (compare Figure 6C1with A1 and B1). Of note, although 95% of PPDyn-positivecells also express nur77 (Fig. 6C3), only 35% of PPE-containingneurons express nur77 (Fig. 6C4). Similar results were ob-tained regardless of when the animals were killed.
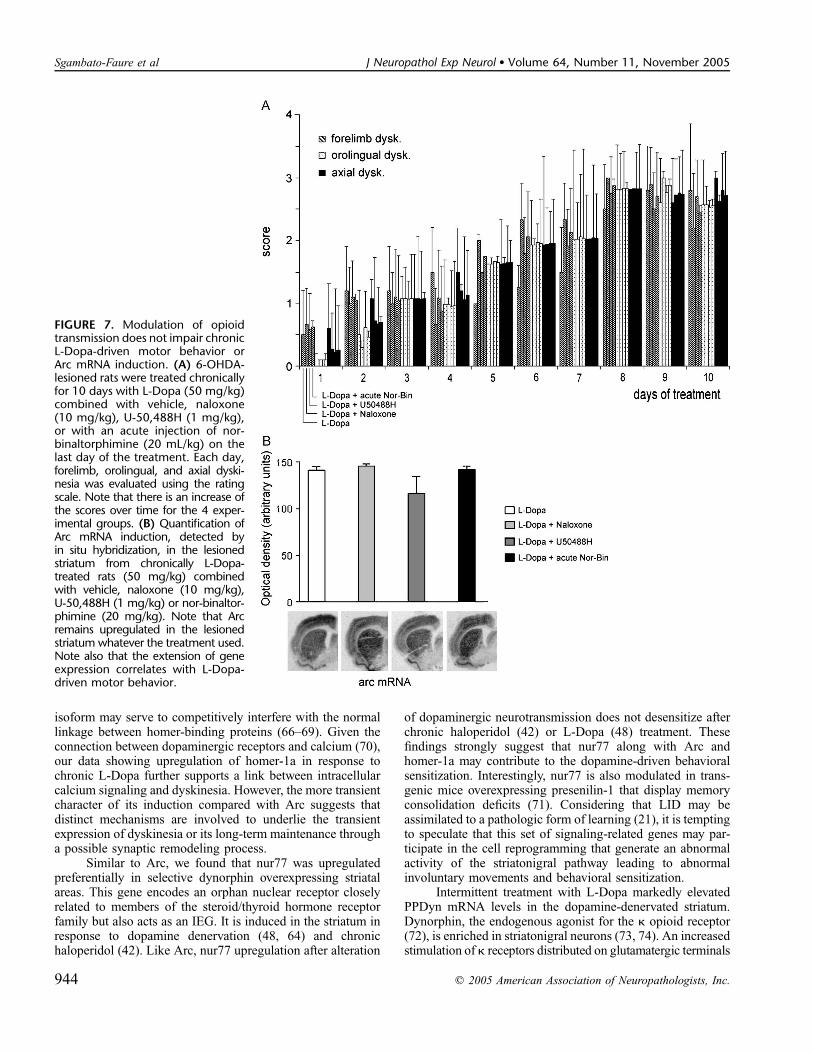
Effect of Coadministration of Naloxone, U-50,488H, or Nor-Binaltorphimine With L-Dopaon Motor Behavior and Gene Expression
To test the involvement of endogenous opioid dynorphinin the development of dyskinesia and behavioral sensitization,
we administered k opioid antagonists (naloxone [49] andnor-binaltorphimine [50]) or agonist (U-50,488H [51]) indenervated-rats treated with L-Dopa. At the behavioral level,opioid antagonist or agonist administration did not alter thedevelopment of dyskinesia (Fig. 7A) as well as locomotorsensitization (data not shown). Similarly, at the histochemicallevel, PPDyn (data not shown) and Arc (Fig. 7B) mRNAupregulation in the lesioned striatum remained the same in thepresence of opioid antagonists. Gene regulation was stillspatially correlated to the L-Dopa-driven motor behavior, thatis, either restricted to the lateral part of the striatum or inthe whole striatum according to the absence or occurrence(Fig. 7B) of locomotor sensitization, respectively. Similarly,FosB and Arc proteins were still induced (data not shown).These data suggest that in our model, modulation of k opioidreceptors does not alter the mechanisms underlying gene in-duction or those involved in the development of LID andlocomotor sensitization.
DISCUSSIONThe main goal of the present study using a rat model of
LID was to investigate the effect of chronic L-Dopa treatmenton the regulation of Arc mRNA and protein in the dopamine-denervated striatum and on its coordinate expression withstriatal neuropeptides belonging to the striatonigral andstriatopallidal output pathways. The main finding of our study
FIGURE 5. FosB-positive cells expressPPDyn and Arc mRNA. (A–C) Repre-sentative photomicrographs of stria-tal sections (from L-Dopa-treated[n = 12] [50 mg/kg] rats) that wereprocessed for FosB immunohisto-chemistry and in situ hybridizationof PPDyn (A), PPE (B), or Arc (C)mRNA. Note that FosB-positive cellsexpress PPDyn and Arc, but not PPEmRNA. Arrowheads point to double-labeled cells: FosB- and PPDyn-positive cells in panel (A); FosB-and Arc-positive cells in panel (C).Arrows point to single-labeled cells(PPE-positive cells in panel [B]). (D)Quantification of FosB-positive cellsexpressing PPDyn, PPE, and Arc inthe lesioned striatum in response tothe chronic L-Dopa treatment.
942 q 2005 American Association of Neuropathologists, Inc.
Sgambato-Faure et al J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005
was that intermittent and chronic L-Dopa led to strongupregulation of Arc mRNA, which is spatially coincident withthat of PPDyn, homer-1a, and nur77 mRNA as well as FosB-like proteins, their striatal territories of expression beingcorrelated with the motor behavior developed (i.e. dyskinesiaaccompanied or not with sensitization of their turningbehaviors). Moreover, these biochemical alterations of Arclevels, along with homer-1a and nur77, were preferentiallyobserved in striatonigral neurons of the direct striatal outputpathway that express PPDyn mRNA, the precursor of thedynorphin peptide.
Arc mRNA is induced in the striatum by acute cocaine(52, 53) and this effect is inhibited by a D1 antagonist,reserpine treatment, or 6-hydroxydopamine lesion (52). In thecurrent study, we found that Arc mRNA was upregulated in thestriatum after chronic L-Dopa administration, validatingprevious data obtained by a microarray study (54). Theinduction of Arc after a chronic L-Dopa treatment contrastswith the fact that most IEG show reduced induction afterchronic exposure to a drug. However, Arc mRNA is alsoupregulated after chronic cocaine in the frontal cortex (55) andchronic morphine in the striatum (56). The exact function ofArc is unknown, but its induction and subcellular localization(35) led to the hypothesis of its involvement in cytoskeletalrearrangements during the process of synaptic plasticity.Highly enriched in neuronal dendrites, where it localizes ina distribution that parallels that of F-actin (33), Arc interactswith cytoskeletal proteins such as MAP-2 (57) and promotes
CaM kinase II-dependent neurite extension (58). The shorthalf-life of Arc mRNA contrasts with its persistent striatalinduction and indicates that Arc upregulation is unlikely toresult from a direct action of L-Dopa but rather a consequenceof a L-Dopa-driven long-term adaptation of the structural andmolecular network of the striatum. This hypothesis is furthersupported by the induction of other structural- and synaptic-related genes (Sgambato-Faure et al, unpublished data; and[54]). Finally, it has been shown that Arc mRNA inductiondepends on NMDA glutamatergic receptors (59, 60) and is re-quired for long-term potentiation as well as memory processin the hippocampus (36–38). It is therefore of particular in-terest to confront our data with other studies in the striatumshowing persistent increases of extracellular glutamate levels(61) and lack of depotentiation after induction of long-termpotentiation (21) in dyskinetic rats.
Homer proteins have gained particular attention becausethey are part of postsynaptic densities and are involved inboth the localization and function of type I metabotropicglutamatergic receptors (45–47) as well as in the coupling ofNMDA-binding ionotropic glutamate receptors to metabo-tropic glutamatergic receptors (62). The dimerization of longhomer proteins can serve to link mGluR receptors to inositoltriphosphate receptors and thus link extracellular signals to therelease of calcium from intracellular stores. The shorterhomer-1a protein can be induced in the brain in response tovarious stimuli, including long-term potentiation, electrocon-vulsive shocks, and drugs of abuse (63–65). This truncated
FIGURE 6. Induction of Arc protein and nur77 mRNA in the lesioned striatum in response to a chronic treatment with low dosesof L-Dopa. Representative photomicrographs of brain sections (from 6-OHDA rats treated with low doses of L-Dopa [n = 3] [10mg/kg]) were processed for Arc immunohistochemistry (A1–A3, B2) and in situ hybridization of PPDyn (B1, B2) or double in situhybridization combining nur77 (C1, C3, C4) with PPDyn (C3) or PPE (C4) mRNA. Note that Arc and nur77 upregulations arecoincident to that of PPDyn (A1, B1, C1). Note also that Arc and PPDyn mRNA are expressed in the same striatal cells (B2). (C2)Quantification of nur77 mRNA induction in the lateral part of the lesioned striatum. ***, p , 0.001 when compared with theipsilateral striatum of unlesioned rats. Note that nur77 is colocalized in dynorphin-expressing cells and not enkephalin-expressingneurons.
q 2005 American Association of Neuropathologists, Inc. 943
J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005 Arc Correlates With L-Dopa-Induced Motor Behavior
isoform may serve to competitively interfere with the normallinkage between homer-binding proteins (66–69). Given theconnection between dopaminergic receptors and calcium (70),our data showing upregulation of homer-1a in response tochronic L-Dopa further supports a link between intracellularcalcium signaling and dyskinesia. However, the more transientcharacter of its induction compared with Arc suggests thatdistinct mechanisms are involved to underlie the transientexpression of dyskinesia or its long-term maintenance througha possible synaptic remodeling process.
Similar to Arc, we found that nur77 was upregulatedpreferentially in selective dynorphin overexpressing striatalareas. This gene encodes an orphan nuclear receptor closelyrelated to members of the steroid/thyroid hormone receptorfamily but also acts as an IEG. It is induced in the striatum inresponse to dopamine denervation (48, 64) and chronichaloperidol (42). Like Arc, nur77 upregulation after alteration
of dopaminergic neurotransmission does not desensitize afterchronic haloperidol (42) or L-Dopa (48) treatment. Thesefindings strongly suggest that nur77 along with Arc andhomer-1a may contribute to the dopamine-driven behavioralsensitization. Interestingly, nur77 is also modulated in trans-genic mice overexpressing presenilin-1 that display memoryconsolidation deficits (71). Considering that LID may beassimilated to a pathologic form of learning (21), it is temptingto speculate that this set of signaling-related genes may par-ticipate in the cell reprogramming that generate an abnormalactivity of the striatonigral pathway leading to abnormalinvoluntary movements and behavioral sensitization.
Intermittent treatment with L-Dopa markedly elevatedPPDyn mRNA levels in the dopamine-denervated striatum.Dynorphin, the endogenous agonist for the k opioid receptor(72), is enriched in striatonigral neurons (73, 74). An increasedstimulation of k receptors distributed on glutamatergic terminals
FIGURE 7. Modulation of opioidtransmission does not impair chronicL-Dopa-driven motor behavior orArc mRNA induction. (A) 6-OHDA-lesioned rats were treated chronicallyfor 10 days with L-Dopa (50 mg/kg)combined with vehicle, naloxone(10 mg/kg), U-50,488H (1 mg/kg),or with an acute injection of nor-binaltorphimine (20 mL/kg) on thelast day of the treatment. Each day,forelimb, orolingual, and axial dyski-nesia was evaluated using the ratingscale. Note that there is an increase ofthe scores over time for the 4 exper-imental groups. (B) Quantification ofArc mRNA induction, detected byin situ hybridization, in the lesionedstriatum from chronically L-Dopa-treated rats (50 mg/kg) combinedwith vehicle, naloxone (10 mg/kg),U-50,488H (1 mg/kg) or nor-binaltor-phimine (20 mg/kg). Note that Arcremains upregulated in the lesionedstriatum whatever the treatment used.Note also that the extension of geneexpression correlates with L-Dopa-driven motor behavior.
944 q 2005 American Association of Neuropathologists, Inc.
Sgambato-Faure et al J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005

(75) and striatonigral neurons (76) may therefore contribute tothe sensitization of dopamine agonist-induced behavioralresponse. It has been shown that the blockade of opioidtransmission allows attenuation of LID in rats (25, 26).However, a more recent study indicated that opioid antagonisttreatment increased LID in a nonhuman primate model of PD(77). We found that blockade of opioid receptors did not alterthe development of dyskinesia or the gene modifications thatare associated with it in our rat LID model. This might be theresult of the downregulation of kappa opioid-binding sites inthe striatum and substantia nigra after chronic L-Dopa (78).Moreover, upregulation of endogenous dynorphin may triggerdesensitization and downregulation of opioid receptors, be-cause it is known that opioid receptors desensitize rapidly afteropioid administration (79).
Another striking observation made in this study (besidesthe perfect topographic superimposition between striatalregions of gene upregulations) is the spatial correlation ofgene expression with dyskinesia accompanied (whole stria-tum) or not (lateral part of the striatum) with locomotorsensitization. This particular pattern of expression have beenobserved in other studies (40) and is not the result of anincomplete denervation of the striatum because we did notreport any significant difference in TH immunoreactivitylevels between these experimental groups. Furthermore, theabsence of locomotor sensitization was not the result of thefact that rats were tested in their home cages, because placingthem in a cylinder did not increase the number of rotations,whatever the group involved. Besides dopaminergic input (80,81), the striatum receives major glutamatergic afferents fromthe cerebral cortex and the thalamus (82–85). The cortical andthalamic afferents are topographically organized along itsrostrocaudal axis and mainly target the lateral portion of thestriatum (86–88). Therefore, the lateral and medial parts of thestriatum differ in terms of afferent inputs. The similaritybetween the striatal territory targeted by cortical and thalamicafferents and the selective spatial induction of gene expressionin the group of dyskinetic animals that did not show behavioralsensitization suggests that the development of sensitizationmay be associated with corticothalamostriatal individualvulnerability.
Together, these findings demonstrate that Arc alongwith homer-1a and nur77 is upregulated in the dynorphin-containing striatonigral neurons from dyskinetic rats supporta role for cytoskeletal remodeling in the regulation of L-Dopa-induced long-term motor alterations. Further studies are requiredto elucidate if these molecular changes are responsible for orprotective against LID.
ACKNOWLEDGMENTSThe authors thank Drs. L. Kerkerian-Le Goff and P.
Salin for critical reading of the manuscript; Drs. R. Paletzkifor generously providing Arc, PPE, and homer-1a cDNAs; andDrs. S. Watson and H. Akil for the gift of PPDyn cDNA. Theauthors also thank M. El Atifi for her help with Excel graphs.
REFERENCES1. Ehringer H, Hornykiewicz O. Distribution of noradrenaline and dopamine
(3-hydroxytyramine) in the human brain and their behavior in diseases ofthe extrapyramidal system. Klin Wochenschr 1960;38:1236–39
2. Agid Y. Parkinson’s disease: Some physiopathological aspects and clinicalpractice. Rev Prat 1989;39:641–46
3. Obeso JA, Rodriguez-Oroz MC, Rodriguez M, et al. The basal gangliaand disorders of movement: Pathophysiological mechanisms. NewsPhysiol Sci 2002;17:51–55
4. Obeso JA, Rodriguez-Oroz M, Marin C, et al. The origin of motorfluctuations in Parkinson’s disease: Importance of dopaminergic inner-vation and basal ganglia circuits. Neurology 2004;62:S17–30
5. Johnston TH, Brotchie JM. Drugs in development for Parkinson’s disease.Curr Opin Investig Drugs 2004;5:720–26
6. Birkmayer W, Hornykiewicz O. Der L-Dioxyphenylalanin (4DOPA)-Effekt bei der Parkinson-Akinese. Wien Klin Wschr 1961;73:787–88
7. Barbeau A. L-Dopa therapy in Parkinsonvs disease: A critical review ofnine years experience. Can Med Assoc J 1969;101:791–800
8. Krack P, Pollak P, Limousin P, et al. From off-period dystonia to peak-dosechorea: The clinical spectrum of varying subthalamic nucleus activity.Brain 1999;122:1133–46
9. Nutt JG. Motor fluctuations and dyskinesia in Parkinson’s disease.Parkinsonism Relat Disord 2001;8:101–8
10. Rascol O, Fabre N. Dyskinesia: L-Dopa-induced and tardive dyskinesia.Clin Neuropharmacol 2001;24:313–23
11. Defebvre L. Motor complications in dopa treatment of Parkinson disease:Clinical description and evaluation. Therapie 2004;59:93–6
12. Di Monte DA, McCormack A, Petzinger G, et al. Relationship amongnigrostriatal denervation, parkinsonism, and dyskinesias in the MPTPprimate model. Mov Disord 2000;15:459–66
13. Baas H, Fuchs G, Gemende I, et al. Which factors influence therapeuticdecisions in Parkinson’s disease? J Neurol 2002;249:49–52
14. Winkler C, Kirik D, Bjorklund A, et al. L-DOPA-induced dyskinesia inthe intrastriatal 6-hydroxydopamine model of Parkinson�s disease: Rela-tion to motor and cellular parameters of nigrostriatal function. NeurobiolDis 2002;10:165–86
15. Paille V, Brachet P, Damier P. Role of nigral lesion in the genesis of dys-kinesias in a rat model of Parkinson’s disease. Neuroreport 2004;15:561–64
16. Jenner P. Avoidance of dyskinesia: Preclinical evidence for continuousdopaminergic stimulation. Neurology 2004;62:S47–55
17. Calon F, Tahar AH, Blanchet PJ, et al. Dopamine-receptor stimulation:Biobehavioral and biochemical consequences. Trends Neurosci 2000;23:S92–S100
18. Graybiel AM, Canales JJ, Capper-Loup C. Levodopa-induced dyskinesiasand dopamine-dependent stereotypies: A new hypothesis. TrendsNeurosci 2000;23:S71–77
19. Bevan MD, Magill PJ, Terman D, et al. Move to the rhythm: Oscillationsin the subthalamic nucleus–external globus pallidus network. TrendsNeurosci 2002;25:525–31
20. Hutchison WD, Dostrovsky JO, Walters JR, et al. Neuronal oscillations inthe basal ganglia and movement disorders: Evidence from whole animaland human recordings. J Neurosci 2004;24:9240–43
21. Picconi B, Centonze D, Hakansson K, et al. Loss of bidirectional striatalsynaptic plasticity in L-DOPA-induced dyskinesia. Nat Neurosci 2003;6:501–6
22. Chase TN. Levodopa therapy: Consequences of the nonphysiologicreplacement of dopamine. Neurology 1998;50:S17–25
23. Chase TN. Striatal plasticity and extrapyramidal motor dysfunction.Parkinsonism Relat Disord 2004;10:305–13
24. Matsumoto RR, Brinsfield KH, Patrick RL, et al. Rotational behaviormediated by dopaminergic and nondopaminergic mechanisms afterintranigral microinjection of specific mu, delta and kappa opioid agonists.J Pharmacol Exp Ther 1988;246:196–203
25. Carey RJ. Naloxone reverses L-Dopa induced overstimulation effects ina Parkinson’s disease animal model analogue. Life Sci 1991;48:1303–8
26. Henry B, Brotchie JM. Potential of opioid antagonists in the treatment oflevodopa-induced dyskinesias in Parkinson’s disease. Drugs Aging 1996;9:149–58
27. Marin C, Engber TM, Chaudhuri P, et al. Effects of kappa receptoragonists on D1 and D2 dopamine agonist and antagonist-induced be-haviors. Psychopharmacology 1996;123:215–21
q 2005 American Association of Neuropathologists, Inc. 945
J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005 Arc Correlates With L-Dopa-Induced Motor Behavior

28. Newman DD, Rajakumar N, Flumerfelt BA, et al. A kappa opioidantagonist blocks sensitization in a rodent model of Parkinson’s disease.Neuroreport 1997;8:669–72
29. Henry B, Fox SH, Crossman AR, et al. Mu- and delta-opioid receptorantagonists reduce levodopa-induced dyskinesia in the MPTP-lesionedprimate model of Parkinson’s disease. Exp Neurol 2001;171:139–46
30. Calabresi P, Giacomini P, Centonze D, et al. Levodopa-induced dyskinesia:A pathological form of striatal synaptic plasticity? Ann Neurol 2000;47:60–68
31. Chase TN, Oh JD. Striatal dopamine- and glutamate-mediated dysregu-lation in experimental parkinsonism. Trends Neurosci 2000;23:86–91
32. Link W, Konietzko U, Kauselmann G et al. Somatodendritic expression ofan immediate early gene is regulated by synaptic activity. Proc Natl AcadSci U S A 1995;92:5734–38
33. Lyford GL, Yamagata K, Kaufmann WE, et al. Arc, a growth factor andactivity-regulated gene, encodes a novel cytoskeleton-associated proteinthat is enriched in neuronal dendrites. Neuron 1995;14:433–45
34. Steward O, Wallace CS, Lyford GL, et al. Synaptic activation causes themRNA for the IEG Arc to localize selectively near activated postsynapticsites on dendrites. Neuron 1998;21:741–51
35. Steward O, Worley P. Local synthesis of proteins at synaptic sites ondendrites: Role in synaptic plasticity and memory consolidation?Neurobiol Learn Mem 2002;78:508–27
36. Guzowski JF, Lyford GL, Stevenson GD, et al. Inhibition of activity-dependent Arc protein expression in the rat hippocampus impairs themaintenance of long-term potentiation and the consolidation of long-termmemory. J Neurosci 2000;20:3993–4001
37. Plath N, Ohana O, Dammermann B, et al. Aberrant LTP in arg3.1/ARCknockout animals. Soc Neurosci Abstr 2001;27:1598
38. Ying SW, Futter M, Rosenblum K, et al. Brain-derived neurotrophic factorinduces long-term potentiation in intact adult hippocampus: Requirementfor ERK activation coupled to CREB and up-regulation of Arc synthesis.J Neurosci 2002;22:1532–40
39. Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. NewYork: Academic Press, 1986
40. Andersson M, Hilbertson A, Cenci MA. Striatal fosB expression iscausally linked with l-DOPA-induced abnormal involuntary movementsand the associated upregulation of striatal prodynorphin mRNA in a ratmodel of Parkinson’s disease. Neurobiol Dis 1999;6:461–74
41. Cenci MA, Lee CS, Bjorklund A. L-DOPA-induced dyskinesia in the ratis associated with striatal overexpression of prodynorphin- and glutamicacid decarboxylase mRNA. Eur J Neurosci 1998;10:2694–2706
42. Beaudry G, Langlois MC, Weppe I, et al. Contrasting patterns and cellularspecificity of transcriptional regulation of the nuclear receptor nervegrowth factor-inducible B by haloperidol and clozapine in the ratforebrain. J Neurochem 2000;75:1694–1702
43. Sgambato V, Pages C, Rogard M, et al. Extracellular signal-regulatedkinase (ERK) controls immediate early gene induction on corticostriatalstimulation. J Neurosci 1998;18:8814–25
44. Nestler EJ, Kelz MB, Chen J. DFosB: A molecular mediator of long-termneural and behavioral plasticity. Brain Res 1999;835:10–17
45. Thomas U. Modulation of synaptic signalling complexes by homerproteins. J Neurochem 2002;81:407–13
46. Fagni L, Worley PF, Ango F. Homer as both a scaffold and transductionmolecule. Sci STKE 2002;137:RE8
47. Fagni L, Ango F, Perroy J, et al. Identification and functional roles ofmetabotropic glutamate receptor-interacting proteins. Semin Cell DevBiol 2004;15:289–98
48. St-Hilaire M, Landry E, Levesque D, et al. Denervation and repeated L-DOPA induce a coordinate expression of the transcription factor NGFI-Bin striatal projection pathways in hemi-parkinsonian rats. Neurobiol Dis2003;14:98–109
49. Hill RG. The status of naloxone in the identification of pain control mech-anisms operated by endogenous opioids. Neurosci Lett 1981;21:217–22
50. Portoghese PS, Lipkowski AW, Takemori AE. Binaltorphimine and nor-binaltorphimine, potent and selective kappa-opioid receptor antagonists.Life Sci 1987;40:1287–92
51. Piercey MF, Lahti RA, Schroeder LA, et al. U-50,488H, a pure kappareceptor agonist with spinal analgesic loci in the mouse. Life Sci 1982;31:1197–1200
52. Fosnaugh JS, Bhat RV, Yamagata K, et al. Activation of Arc, a putative�effector� immediate early gene, by cocaine in rat brain. J Neurochem1995;64:2377–80
53. Tan A, Moratalla R, Lyford GL, et al. The activity-regulated cytoskeletal-associated protein Arc is expressed in different striosome-matrix patternsfollowing exposure to amphetamine and cocaine. J Neurochem 2000;74:2074–78
54. Konradi C, Westin JE, Carta M, et al. Transcriptome analysis in a ratmodel of L-DOPA-induced dyskinesia. Neurobiol Dis 2004;17:219–36
55. Freeman WM, Brebner K, Lynch WJ, et al. Changes in rat frontal cortexgene expression following chronic cocaine. Brain Res Mol Brain Res2002;104:11–20
56. Marie-Claire C, Courtin C, Roques BP, et al. Cytoskeletal genes regu-lation by chronic morphine treatment in rat striatum. Neuropsychophar-macology 2004;29:2208–15
57. Fujimoto T, Tanaka H, Kumamaru E, et al. Arc interacts withmicrotubules/microtubule-associated protein 2 and attenuates microtu-bule-associated protein 2 immunoreactivity in the dendrites. J NeurosciRes 2004;76:51–63
58. Donai H, Sugiura H, Ara D, et al. Interaction of Arc with CaM kinase IIand stimulation of neurite extension by Arc in neuroblastoma cellsexpressing CaM kinase II. Neurosci Res 2003;47:399–408
59. Steward O, Worley PF. Selective targeting of newly synthesized ArcmRNA to active synapses requires NMDA receptor activation. Neuron2001;30:227–40
60. Martin KC. Arc mRNA dynamics: Return to sender—the NMDA receptorprovides the targeting address for Arc mRNA. Trends Neurosci 2001;24:621–23
61. Robelet S, Melon C, Guillet B, et al. Chronic L-DOPA treatment increasesextracellular glutamate levels and GLT1 expression in the basal ganglia ina rat model of Parkinson’s disease. Eur J Neurosci 2004;20:1255–66
62. Sheng M, Kim E. The Shank family of scaffold proteins. J Cell Sci2000;113:1851–56
63. Brakeman PR, Lanahan AA, O’Brian R, et al. Homer: A protein thatselectively binds to metabotropic glutamate receptors. Nature 1997;386:284–88
64. Berke JD, Paletzki RF, Aronson GJ, et al. A complex program of stria-tal gene expression induced by dopaminergic stimulation. J Neurosci1998;18:5301–10
65. Bottai D, Guzowski JF, Schwarz MK, et al. Synaptic activity-inducedconversation of intronic to exonic sequence in Homer 1 immediate earlygene expression. J Neurosci 2002;22:167–75
66. Xiao B, Tu JC, Petralia RS, et al. Homer regulates the association of group1 metabotropic glutamate receptors with multivalent complexes of homer-related, synaptic proteins. Neuron 1998;21:707–16
67. Tu JC, Xiao B, Yuan JP, et al. Homer binds a novel proline-rich motif andlinks group 1 metabotropic glutamate receptors with IP3 receptors.Neuron 1998;21:717–26
68. Ango F, Prezeau L, Muller T, et al. Agonist-independent activation ofmetabotropic glutamate receptors by the intracellular protein Homer.Nature 2001;411:962–65
69. Ango F, Robbe D, Tu JC, et al. Homer-dependent cell surface expressionof metabotropic glutamate receptor type 5 in neurons. Mol Cell Neurosci2002;20:323–29
70. Bergson C, Levenson R, Goldman-Rakic PS, et al. Dopamine receptor-interacting proteins: The Ca(21) connection in dopamine signaling.Trends Pharmacol Sci 2003;24:486–92
71. Dickey CA, Loring JF, Montgomery J, et al. Selectively reducedexpression of synaptic plasticity-related genes in amyloid precursorprotein 1 presenilin-1 transgenic mice. J Neurosci 2003;23:5219–26
72. Chavkin C, James IF, Goldstein A. Dynorphin is a specific endogenousligand of the kappa opioid receptor. Science 1982;215:413–15
73. Vincent S, Hokfelt T, Christensson I, et al. Immunohistochemicalevidence for a dynorphin immunoreactive striato-nigral pathway. Eur JPharmacol 1982;85:251–52
74. Gerfen CR, Young WS. 3rd Distribution of striatonigral and striatopallidalpeptidergic neurons in both patch and matrix compartments: An in situhybridization histochemistry and fluorescent retrograde tracing study.Brain Res 1988;460:161–67
75. Meshul CK, McGinty JF. Kappa opioid receptor immunoreactivity in thenucleus accumbens and caudate–putamen is primarily associated withsynaptic vesicles in axons. Neuroscience 2000;96:91–99
76. Abou-Khalil B, Young AB, Penney JB. Evidence for the presynapticlocalization of opiate binding sites on striatal efferent fibers. Brain Res1984;323:21–29
946 q 2005 American Association of Neuropathologists, Inc.
Sgambato-Faure et al J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005

77. Samadi P, Gregoire L, Bedard PJ. Opioid antagonists increase the dyskineticresponse to dopaminergic agents in parkinsonian monkeys: Interaction be-tween dopamine and opioid systems. Neuropharmacology 2003;45:954–63
78. Johansson PA, Andersson M, Andersson KE, et al. Alterations in corticaland basal ganglia levels of opioid receptor binding in a rat model ofl-DOPA-induced dyskinesia. Neurobiol Dis 2001;8:220–39
79. Morris BJ, Herz A. Control of opiate receptor number in vivo:Simultaneous kappa-receptor down-regulation and mu-receptor up-regulation following chronic agonist/antagonist treatment. Neuroscience1989;29:433–42
80. Moore RY, Bhatnagar RK, Heller A. Anatomical and chemical studies ofa nigro-neostriatal projection in the cat. Brain Res 1971;30:119–35
81. Doucet G, Descarries L, Garcia S. Quantification of the dopamineinnervation in adult rat neostriatum. Neuroscience 1986;19:427–45
82. Parent A, Mackey A, De Bellefeuille L. The subcortical afferents tocaudate nucleus and putamen in primate: A fluorescence retrogradedouble labeling study. Neuroscience 1983;10:1137–50
83. Parent A. Extrinsic connections of the basal ganglia. Trends Neurosci1990;13:254–58
84. Smith AD, Bolam JP. The neural network of the basal ganglia as revealedby the study of synaptic connections of identified neurones. TrendsNeurosci 1990;13:259–65
85. Smith Y, Raju DV, Pare JF, et al. The thalamostriatal system: A highlyspecific network of the basal ganglia circuitry. Trends Neurosci 2004;27:520–27
86. McGeorge AJ, Faull RL. The organization of the projection from thecerebral cortex to the striatum in the rat. Neuroscience 1989;29:503–37
87. Berendse HW, Galis-de Graaf Y, Groenewegen HJ. Topographicalorganization and relationship with ventral striatal compartments ofprefrontal corticostriatal projections in the rat. J Comp Neurol 1992;316:314–47
88. Van der Kooy D. The organization of the thalamic, nigral and raphe cellsprojecting to the medial vs lateral caudate-putamen in rat. A fluorescentretrograde double labeling study. Brain Res 1979;169:381–87
q 2005 American Association of Neuropathologists, Inc. 947
J Neuropathol Exp Neurol � Volume 64, Number 11, November 2005 Arc Correlates With L-Dopa-Induced Motor Behavior




















