Blocking EphA4 upregulation after spinal cord injury results in enhanced chronic pain
13
Blocking EphA4 upregulation after spinal cord injury results in enhanced chronic pain Lillian Cruz-Orengo a , Johnny D. Figueroa a , Ixane Velázquez a , Aranza Torrado a , Cristina Ortíz a , Carmen Hernández a , Anabel Puig a , Annabell C. Segarra a , Scott R. Whittemore b,c,d , Jorge D. Miranda a, ⁎ a Department of Physiology, University of Puerto Rico School of Medicine, P.O. Box 365067 San Juan, PR 00936-5067, USA b Kentucky Spinal Cord Injury Research Center, University of Louisville School of Medicine, Louisville, KY 40202, USA c Department of Neurological Surgery, University of Louisville School of Medicine, Louisville, KY 40202, USA d Department of Anatomical Sciences and Neurobiology, University of Louisville School of Medicine, Louisville, KY 40202, USA Received 10 October 2005; revised 1 June 2006; accepted 10 July 2006 Available online 7 September 2006 Abstract Spinal cord injury (SCI) is characterized by a total or partial loss of motor and sensory functions due to the inability of neurons to regenerate. This lack of axonal regenerative response has been associated with the induction of inhibitory proteins for regeneration, such as the Eph receptor tyrosine kinases. One member of this family, the EphA4 receptor, coordinates appropriate corticospinal fibers projections during early development and is expressed in spinal commissural interneurons. Its mechanism of action is mediated by repulsive activity after ligand binding, but its role after trauma is unknown. We examined the temporal expression profile of this receptor after spinal cord contusion in adult rats by RT- PCR and immunohistochemistry. SCI induced a biphasic gene expression profile with an initial downregulation at 2 and 4 days post-injury (DPI) followed by a subsequent upregulation. Double labeling studies localized EphA4 immunoreactivity in neurons from the gray matter and astrocytes of the white matter. To test the role of this receptor, we reduced gene upregulation by intrathecal/subdural infusion of EphA4–antisense oligodeoxynucleotide (ODN) and subsequently assessed behavioral outcomes. No locomotor recovery was observed in the rats treated with the EphA4–antisense ODN. Interestingly, reducing EphA4 expression increased mechanical allodynia, as observed by the Von Frey test and decreased exploratory locomotor activity. These results indicate that upregulation of EphA4 receptor after trauma may prevent the development of abnormal pain syndromes and could potentially be exploited as a preventive analgesic mediator to chronic neuropathic pain. © 2006 Elsevier Inc. All rights reserved. Keywords: Allodynia; Antisense; Exploratory behavior; Plasticity; Trauma Introduction Following central nervous system (CNS) injury, the micro- environment at the lesion site prevents regrowth of axotomized neurons, despite their intrinsic capacity to regenerate (Aguayo et al., 1981; Kwon and Tetzlaff, 2001). Inhibitory molecular cues of myelin origin, NOGO, MAG and OMgp, are the best known and most intensively studied neurite outgrowth inhibitors (Schnell and Schwab, 1990; McKerracher et al., 1994; Mukhopadyay et al., 1994; Wang et al., 2002). However, other repulsive cues play relevant roles after SCI as evidenced by the lack of significant spinal regeneration in Nogo-deficient mice (Zheng et al., 2003a,b). Among those signals are proteins expressed by astrocytes, including proteoglycans, semaphorin- 3, Slit proteins, and Eph/ephrins (Irizarry-Ramírez et al., 2005; Silver and Miller, 2004). Some reports support the idea that SCI recapitulates molecular cues used in early stages of neural morphogenesis (Emery et al., 2003). The Eph receptor tyrosine kinase (RTK) family is the largest RTK family known. Their main characteristic is their ability to mediate cell–cell repulsion after binding of their ligand to an adjacent cell surface (Pasquale, 2005). Eph RTKs and their Experimental Neurology 202 (2006) 421 – 433 www.elsevier.com/locate/yexnr ⁎ Corresponding author. Fax: +1 787 753 0120. E-mail address: [email protected] (J.D. Miranda). 0014-4886/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.expneurol.2006.07.005
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Blocking EphA4 upregulation after spinal cord injury results in enhanced chronic pain

02 (2006) 421–433www.elsevier.com/locate/yexnr
Experimental Neurology 2
Blocking EphA4 upregulation after spinal cord injury results inenhanced chronic pain
Lillian Cruz-Orengo a, Johnny D. Figueroa a, Ixane Velázquez a, Aranza Torrado a,Cristina Ortíz a, Carmen Hernández a, Anabel Puig a, Annabell C. Segarra a,
Scott R. Whittemore b,c,d, Jorge D. Miranda a,⁎
a Department of Physiology, University of Puerto Rico School of Medicine, P.O. Box 365067 San Juan, PR 00936-5067, USAb Kentucky Spinal Cord Injury Research Center, University of Louisville School of Medicine, Louisville, KY 40202, USA
c Department of Neurological Surgery, University of Louisville School of Medicine, Louisville, KY 40202, USAd Department of Anatomical Sciences and Neurobiology, University of Louisville School of Medicine, Louisville, KY 40202, USA
Received 10 October 2005; revised 1 June 2006; accepted 10 July 2006Available online 7 September 2006
Abstract
Spinal cord injury (SCI) is characterized by a total or partial loss of motor and sensory functions due to the inability of neurons to regenerate.This lack of axonal regenerative response has been associated with the induction of inhibitory proteins for regeneration, such as the Eph receptortyrosine kinases. One member of this family, the EphA4 receptor, coordinates appropriate corticospinal fibers projections during earlydevelopment and is expressed in spinal commissural interneurons. Its mechanism of action is mediated by repulsive activity after ligand binding,but its role after trauma is unknown. We examined the temporal expression profile of this receptor after spinal cord contusion in adult rats by RT-PCR and immunohistochemistry. SCI induced a biphasic gene expression profile with an initial downregulation at 2 and 4 days post-injury (DPI)followed by a subsequent upregulation. Double labeling studies localized EphA4 immunoreactivity in neurons from the gray matter and astrocytesof the white matter. To test the role of this receptor, we reduced gene upregulation by intrathecal/subdural infusion of EphA4–antisenseoligodeoxynucleotide (ODN) and subsequently assessed behavioral outcomes. No locomotor recovery was observed in the rats treated with theEphA4–antisense ODN. Interestingly, reducing EphA4 expression increased mechanical allodynia, as observed by the Von Frey test and decreasedexploratory locomotor activity. These results indicate that upregulation of EphA4 receptor after trauma may prevent the development of abnormalpain syndromes and could potentially be exploited as a preventive analgesic mediator to chronic neuropathic pain.© 2006 Elsevier Inc. All rights reserved.
Keywords: Allodynia; Antisense; Exploratory behavior; Plasticity; Trauma
Introduction
Following central nervous system (CNS) injury, the micro-environment at the lesion site prevents regrowth of axotomizedneurons, despite their intrinsic capacity to regenerate (Aguayoet al., 1981; Kwon and Tetzlaff, 2001). Inhibitory molecularcues of myelin origin, NOGO, MAG and OMgp, are the bestknown and most intensively studied neurite outgrowthinhibitors (Schnell and Schwab, 1990; McKerracher et al.,
⁎ Corresponding author. Fax: +1 787 753 0120.E-mail address: [email protected] (J.D. Miranda).
0014-4886/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.expneurol.2006.07.005
1994; Mukhopadyay et al., 1994; Wang et al., 2002). However,other repulsive cues play relevant roles after SCI as evidencedby the lack of significant spinal regeneration in Nogo-deficientmice (Zheng et al., 2003a,b). Among those signals are proteinsexpressed by astrocytes, including proteoglycans, semaphorin-3, Slit proteins, and Eph/ephrins (Irizarry-Ramírez et al., 2005;Silver and Miller, 2004). Some reports support the idea that SCIrecapitulates molecular cues used in early stages of neuralmorphogenesis (Emery et al., 2003).
The Eph receptor tyrosine kinase (RTK) family is the largestRTK family known. Their main characteristic is their ability tomediate cell–cell repulsion after binding of their ligand to anadjacent cell surface (Pasquale, 2005). Eph RTKs and their

422 L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
ligands, ephrins, have been implicated in the establishment offunctional circuits, cell migration, and excitatory synapseregulation (Klein, 2004), actions mediated mainly by repulsiveactivity. However, their expression is not limited to embryonicstages. Recent evidence highlights the contribution of EphA4 inthe adult nervous system maintenance (Martone et al., 1997;Conover et al., 2000), in adult synaptic plasticity and learning inthe hippocampus (Murai and Pasquale, 2002; Murai et al.,2003a), and the central pattern generators of the spinal cord(Kullander et al., 2003). Moreover, EphA4 has been implicatedin corticospinal tract formation (Dottori et al., 1998; Kullanderet al., 2001a, b; Greferath et al., 2002), neuromuscular junction(Lai et al., 2001), and motor neuron pathfinding (Eberhart et al.,2000; Helmbacher et al., 2000). All these contribute to broadenthe scope of EphA4 as a relevant molecular cue for locomotordevelopment and maintenance. While the function of severalEphs and ephrins during spinal cord development has beendescribed (Coonan et al., 2001), their putative role after injuryhas not been fully elucidated.
In several models of CNS trauma, upregulation of Ephreceptors has been observed, suggesting a possible role in therepulsive environment generated after injury (Moreno-Floresand Wandosell, 1999; Biervert et al., 2001; Knöll et al., 2001).More specifically, some reports showed an increase in theexpression of several members of the Eph RTK family after SCI(Miranda et al., 1999; Bundesen et al., 2003; Willson et al.,2003; Liu et al., 2006; Fabes et al., 2006). During development,the roles of Eph receptors are predominately to mediate cellmigration, axonal retraction, target recognition, as well asaxonal fasciculation. However, after CNS trauma, their over-expression may contribute to the non-permissive environmentthat blocks axonal outgrowth. Indeed, Benson et al. (2005)identified EphB3 as an additional myelin-based inhibitorymolecule of axonal regeneration. In addition, expression ofthese genes after injury may modulate the formation of sensorysynapses involved in pain.
Chronic neuropathic pain (CNP) is a common and devastat-ing sequelae that many spinally injured patients endure(Christensen and Hulsebosch, 1997; Finnerup et al., 2003;Hulsebosch, 2005). Some reports attribute CNP to sprouting ofnociceptive fibers and to inappropriate synapse formation afterSCI (Christensen and Hulsebosch, 1997; Ondarza et al., 2003).Battaglia et al. (2003) showed that the interaction of EphB1/ephrin-B2 functions to modulate nociceptive synapses asso-ciated with sensory abnormalities in persistent pain states. Theyfurther proposed that ephrin-B2 accumulation after peripheraltissue damage may lead to receptor redistribution that ultimatelytriggers hyperalgesia. Since EphA4 is a synapse modulatorinvolved in the formation of specific synapses and enhancesplasticity in structures such as the adult hippocampus (Muraiet al., 2003A,B) and in the neuromuscular junction (Lai et al.,2001), we hypothesized that changes in EphA4 expression afterSCI may also modulate synapse formation and/or maintenance.In this study, we examined the possible involvement of EphA4RTK as a nociceptive synaptic modulator after SCI analyzingbehavioral changes and nociceptive thresholds after receptorgene suppression.
Materials and methods
Surgery
Spinal cord injuryAdult female Sprague–Dawley rats (225 g) from Hilltop Lab
(Scottdale, PA) were anesthetized with a cocktail of 40 mg/kgKetamine, 4 mg/kg Xylazine, and 0.9 mg/kg Acepromazineadministered by intraperitoneal injection. A dorsal incision wasmade to expose T10 vertebra and a laminectomy wasperformed, leaving the spinal segment exposed. After exposureof the T10 segment by laminectomy, animals received amoderate contusion using the NYU impactor that provides acontusion of 12.5 g cm as previously described (Gruner, 1992;Miranda et al., 1999; Irizarry-Ramírez et al., 2005). Shamcontrol rats remained uncontused and were sutured afterlaminectomy.
Intrathecal/subdural infusionAntisense oligonucleotides were infused with Alzet® os-
motic minipumps Model 2004 (DURECT Co., Cupertino,CA) that were primed 48 h prior to surgery and connected to a28–32 gauge catheter (Rethaco, Allison Park, PA). Primingwas done with 10 nM (higher doses were toxic) oligodeox-ynucleotides. EphA4–antisense, or control sequences, number2.04812 from Biognostik (Göttingen, Germany) were used.Briefly, after lesion to the spinal cord at the T10 segment, asecond laminectomy was performed at T12. The dura wasopened just enough to allow the catheter to be inserted withthe opening rostral to lesion site. Infusions proceeded for 28days post-injury (DPI).
Surgical and postoperative animal careWhile the rats were under anesthesia, they were kept at 37°C
on a heating blanket to prevent hypothermia. Both treatmentgroups received two daily doses of the antibiotic Cefazolin(25 mg/kg, Bristol Myers Squibb) subcutaneously for 7consecutive days, beginning at the day of surgery. In addition,analgesia was provided by two daily doses of Buprenex for3 days. Animals were hydrated with 10 cm3 lactated Ringer'ssolution, administered subcutaneously after surgery and duringpostoperative care when needed (≈1 cm3 per 5 g weight loss).Bladders of injured rats were expressed three times daily untilspontaneous voiding returned, usually after 5 days. Animalswere housed individually on absorbent bedding cages. Theseexperiments were approved by the University of Puerto RicoIACUC and followed NIH guidelines for the safe use and careof laboratory animals.
Reverse transcriptase polymerase chain reaction
Four sets of sham and injured rats were sacrificed at 2, 4, 7,14, and 28 DPI as described by Irizarry-Ramírez et al. (2005).Briefly, the animals were anesthetized by intraperitonealadministration of Pentobarbital (40–50 mg/kg) and transcar-dially perfused with chilled 0.01 M phosphate-buffered saline,pH 7.4 (PBS). The T10 lesion epicenter (5 mm) was dissected

423L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
and total RNA extracted using Trizol (Invitrogen, Carlsbad,CA) following the manufacturer's instructions. To avoidgenomic contamination, the RNAs were treated with DNAse I(Ambion Inc., Austin, TX).
Heterogeneous reverse transcription was performed using1 μg of total RNA, 250 ng random hexamers (Promega), and0.2 U SuperScript II MMLV reverse transcriptase (Invitrogen).Reverse transcription with SuperScriptII was done accordingto the manufacturer's protocol. Mock cDNAwas prepared usingall reagents for reverse transcription except reverse transcriptaseenzyme and amplified as negative control to assess the possi-bility of genomic contamination. RT-PCR for the β-actin genewas used as a housekeeping gene to demonstrate the speci-ficity of the changes that are taking place in the injured spinalcord.
cDNA amplification was performed with 2 μM of specificEphA4 primers (forward: 5′-AGCGCTTCATCAGAGA-GAGCC-3′; reverse: 5′-GGTCCGGGCTAGGGTTATACT-3′)and β-actin as a housekeeping gene using the forward: 5′-CACACTTTCTACAATGAGCTGCGT-3′ and reverse: 5′-GGTGAGGATCTTCATGAGGTAGTC-3′ primers. Amplifica-tion reactions included 1.5 mM MgCl2, 250 μM dNTPs, and2.5 U Taq polymerase (Promega, Madison, WI). PCR productswithin the linear range of amplification were analyzed on a 1%agarose gel containing ethidium bromide as described (Irizarry-Ramírez et al., 2005; Willson et al., 2002). Densitometricanalysis of the RT-PCR products was performed using the Bio-Rad Gel Doc 2000 andMolecular Analysis Software after globalbackground subtraction and standardized relative to the expres-sion of β-actin.
The identity of the single band products (EphA4 and β-actin)obtained from RT-PCR assays was confirmed by sequencing. Inbrief, PCR products were purified from the agarose gel usingQIAquick Gel Extraction kit (Qiagen, Valencia, CA) and clonedinto EcoRV site in Bluescript+/− SK (Stratagene, La Jolla, CA).Following standardized protocols, recombinant plasmids wereused to transform competent DH5α cells (Sambrook andRussell, 2001). Plasmids were isolated using the Maxi-prepQIAquick Isolation kit (Qiagen) and sequenced with the T7Sequenase sequencing kit (Amersham Pharmacia Biotech,Piscataway, NJ) following the manufacturer's instructions.The sequenced clones were analyzed in a 6% polyacrylamidedenaturing sequencing gel and by autoradiography.
Immunohistochemistry
Perfusion and tissue preparationPerfusion and sectioning followed previously published
procedures (Miranda et al., 1999; Willson et al., 2002; Irizarry-Ramírez et al., 2005). Briefly, 7 DPI rats were anesthetized andperfused intracardially with PBS and then with 4% paraformal-dehyde (PFA) in saline. Spinal cords were removed and post-fixed in 4%PFA/PBS at 4°C for 2 h then incubated in 30%sucrose/PBS and allowed to sink at 4°C, overnight. Onecentimeter spinal cord segments spanning the lesion epicenterwere submerged in Tissue-Tek O.C.T. embedding media (MilesInc., Elkhart, IN) and placed on dry ice. They were then
transversely sectioned at 16 μm using a Leica CM3050 cryostat(Leica Microsystems, Nussloch, Germany) and mounted on gel-coated Superfrost microscope slides (Fisher Scientific, Pitts-burgh, PA).
ImmunofluorescenceEphA4 double labeling was performed with antibodies
against GFAP (Pharmingen, San Diego, CA) and NeuN(Chemicon International Inc., Temecula, CA) to identifyastrocytes and neurons, respectively. Spinal cord sectionswere post-fixed in 4% PFA/PBS for 10 min at 4°C and washed3 times in 0.1% Tween 20/PBS for 15 min. After blocking for2 h with 10% donkey serum in PBS, the sections wereincubated overnight in primary antibodies 1:200 sc-921 rabbitanti-EphA4 (Santa Cruz Biotechnology, Santa Cruz, CA), and1:100 mouse anti-NeuN or 1:200 mouse anti-GFAP at 4°C.After rinsing in 0.1% Tween 20/PBS, sections were incubatedwith 1:200 goat anti-rabbit FITC for EphA4 and 1:100 donkeyanti-mouse Rhodamine for both cellular markers (both fromJackson ImmunoResearch, West Grove, PA) for 2 h at roomtemperature. Sections were rinsed six times in PBS andcoverslipped with Prolong Antifade (Molecular Probes Invitro-gen Detection Technologies, Eugene, OR). Images werevisualized with a Zeiss LSM 5 PASCAL Confocal MicroscopeSystem (Carl Zeiss MicroImaging Inc., Thornwood, NY).Controls to demonstrate the specificity of the antibody againstthe EphA4 receptor during the immunohistochemistry assayswere performed with preabsorbed antibody (with the peptide[sc-921P] used to generate it).
Antibody specificityThe specificity of the sc-921 EphA4-antibody was assessed
by immunoblot analysis with the peptides sc-921P (the targetepitope of EphA4), sc-922P (encode another epitope ofEphA4), sc-919P, and sc-1015P (homologous epitopes ofEphA3 and A7) (1.0 μg each). These peptides were generatedby Santa Cruz Biotechnology, and all of them correspond to theC-terminus of the named proteins. A single immunoreactiveband was detected with sc-921P that disappeared with 0.5 μgblocking peptide preabsorption. No cross-reactivity wasobserved between the sc-921 anti-EphA4 antibody and theother peptides, confirming the specificity of the antibody. Allimmunofluorescence assays were controlled by incubating insecondary FITC-conjugated or Rhodamine-conjugated antibo-dies without primary antibody incubation.
Behavioral assays
BBB open field, grid walking, and beam crossing testsEphA4–antisense and control infused animals were evalu-
ated by Basso–Beattie–Bresnahan (BBB) open field test,horizontal grid ladder, and round/square horizontal beamcross (Basso et al., 1995; Behrmann et al., 1992; Merkler etal., 2001). Days selected for these tasks were 7, 14, 21 and 28DPI. The rats were trained in three sessions for 3 days prior tosurgery. Animals unable to train were not used for intrathecal/subdural infusion surgery or further behavioral tests.

Fig. 1. Expression of EphA4 mRNA in the adult rat after SCI. (A) EphA4 PCRproducts were analyzed in 1% agarose/ethidium bromide gels. RNA wasextracted from the lesion epicenter segment (5 mm) at 2, 4, 7, 14, and 28 dayspost-injury and standardized RT-PCR performed with specific primers. (B)Densitometric analysis of the PCR products revealed a biphasic temporal patternof EphA4 gene expression, while β-actin remained steady. The samples from 2,4, and 7 DPI showed a significant difference relative to the control values. Datarepresent mean relative to control±SEM, n=4, *p<0.05 and **p<0.01(ANOVA followed by Bonferroni post hoc test).
424 L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
Spontaneous exploratory behaviorSpontaneous locomotor activity was assessed in treated
animals at 14 and 28 DPI, as previously published (Mills et al.,2001). The rats were placed in the activity chambers monitoredby a Versamax analyzer (AccuScan, Columbus, OH). Trialswere performed in a dark room for 20 min, without priorhabituation time. Assay time was decreased from 30 to 20 minbecause, after 15 min, they habituated to their cages and nochanges in exploratory behavior were monitored after that time(Mills et al., 2001). The behaviors analyzed were horizontalactivity, total distance, number of movements, time of move-ment, and time spent at margin and center of the cage, amongothers. Vertical activity, which relies on hindlimb locomotionand weight support, was assessed as negative control.
Sensory evoked testSomatic allodynia was evaluated at 35 DPI using Semmes-
Weinstein Monofilaments sensory evaluators (Stoelting Co.,Wood Dale, IL) that utilize Von Frey methodology. Themanufacturer's operation manual was followed with modifica-tions to be able to observe a wider scope of somatosensoryabnormalities all through the body, not only on the inner paws.A rat body map scheme was designed with Adobe Illustrator®7.0 software (Adobe Systems Inc., San Jose, CA) with ten bodyparts for evaluation: left and right torso, flank, leg and paw,upper and lower axis. The left and right forelimbs were used asnegative controls. Each body part was divided in quadrants ofapproximately 1 cm2, except for the paws that were divided bythe skin between toes and ankle. Targeted forces applied were1.0, 2.0, 4.0, 8.0, 10.0, 15.0, 26.0, and 60.0 g, observed assubthreshold stimuli in naive animals. Responses were scaled as0, 1, 2, and 3 for no reaction, slight responsiveness, moderatealertness, and body withdrawal, respectively. The animals wereheld unrestrained, therefore, once they showed a response(slight, moderate, or severe) in a particular quadrant, no furtherstimulus (increase in targeted force) was applied to it to avoidawareness or excessive stress.
Data analysis
Densitometric analysis of the RT-PCR single band productsobserved in the agarose gel electrophoresis was performedusing the Bio-Rad Gel Doc 2000 and Molecular AnalysisSoftware (Bio-Rad Laboratories, Hercules, CA). After globalbackground subtraction, the level of EphA4 PCR product wasstandardized relative to the expression of β-actin. Injury-induced changes in the level of EphA4 expression werenormalized relative to control samples for each time pointstudied.
Images of double labeled sections were visualized with aZeiss LSM 5 PASCAL Confocal Microscope System (CarlZeiss MicroImaging Inc., Thornwood, NY) and photomicro-graphs assembled with Adobe PhotoShop® 5.0 software(Adobe Systems Inc., San Jose, CA).
The rat body map data during the Von Frey sensoryassessment were generated using Adobe Illustrator® 7.0 soft-ware. After data analysis, the same layout was used followed by
Adobe PhotoShop® 5.0 software for picture coloring andediting.
Statistical analysis comparing several groups (between shamand injured groups; between antisense and control groups) wasperformed by ANOVA followed by Bonferroni post hoc testusing InStat version 3.0 software (GraphPad Software Inc., SanDiego, CA). For comparing only two groups, Student's t testwas used or Mann–Whitney test. Both were performed asunpaired and two-tailed using InStat version 3.0 software.Statistical significance was set at p<0.05.
Results
SCI induced a biphasic pattern of EphA4 mRNA expression
Standardized RT-PCR studies generated unique PCR pro-ducts that migrated to the expected position: EphA4 to 889 bpand β-actin to 319 bp (Fig. 1A). The pattern of EphA4 mRNAexpression at the lesion epicenter was biphasic, while β-actinmRNA levels remained steady after SCI. Densitometric analysisdemonstrated an initial decrease of 60% and 62% in the PCRproduct at 2 and 4 days post-injury (DPI), respectively (Fig. 1B).Subsequently, EphA4 mRNA expression was increased 76%above control levels at 7 DPI, but returned to a basal expressionat 14 and 28 DPI (F=12.902; df=5, 20; p<0.0001).

425L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
EphA4 protein expression is observed in astrocytes andneurons of injured cords
Basal levels of EphA4 receptor expression in sham animalswere observed in neurons of the gray matter, and lessprominently in fibers of the white matter (Supplementary Fig.1). This expression pattern may be necessary to stabilize thesynapses and avoid aberrant sprouting during normal develop-ment or after trauma. Injury to the spinal cord produced changesin the EphA4 immunoreactivity in cells of the gray and whitematter (Supplementary Fig. 1).
Double label immunostaining for EphA4 (Fig. 2A) and NeuN(Fig. 2B) demonstrated that motor neurons expressed EphA4(Fig. 2C) at regions caudal (0.5 cm) and rostral (data not shown)to the lesion epicenter at 7 DPI. In addition, robust EphA4immunoreactivity (Fig. 2D) was observed in reactive astrocytes(Fig. 2E) in both gray and white matter of the contused spinalcord (Fig. 2F). Co-localization of EphA4 with oligodendrocytesand OX-42 was not observed (data not shown).
Since other EphA receptors had been shown to beupregulated after trauma to the central nervous system and in
Fig. 2. EphA4 receptor expression in reactive astrocytes and neurons of the spinacaudal to the lesion epicenter, was observed in adult injured animals. (D–F) Doublmatter in regions caudal and rostral (not shown) to the lesion epicenter. Boxed insmotor neuron (A–C) or a reactive astrocyte surrounding a motor neuron (D–F). (GEphA7 antibodies (sc-919P and sc-1015P, respectively) did not cross-react with sc-9when the antibody was preabsorbed with the sc-921P peptide. Scale bar=20 μm.
other neuropathologies (Moreno-Flores and Wandosell, 1999;Biervert et al., 2001;Knöll et al., 2001;Sobel, 2005), weconfirmed the specificity of the EphA4 antibody sc-921 (SantaCruz Biotechnology). Immunoblots (Fig. 2G) were performedagainst peptides that express similar epitopes from EphAreceptors known to increase after injury (EphA3 [sc-919P],EphA4 [sc-921P], EphA7 [sc-1015P]). Additionally, anotherpeptide (sc-922P) from an epitope close to the EphA4 sc-921Pwas used to confirm the specificity of our results. As observed inFig. 2G, the sc-921 antibody only showed positive immunor-eactivity to sc-921P. In addition, immunoreactivity was blockedwith sc-921P antibody preabsorption (Fig. 2G). Moreover,immunohistochemical studies with preabsorbed antibody didnot present any immunoreactivity (data not shown) in thesections studied confirming the antibody specificity.
EphA4–antisense oligodeoxynucleotide suppress geneexpression
To assess the specific contribution of EphA4 receptors aftertrauma to the spinal cord, we infused EphA4–antisense
l cord. (A–C) Co-localization of EphA4 in NeuN immunoreactivities, 0.5 cme labeling confirmed the presence of EphA4 in reactive astrocytes of the whiteets are high magnification pictures that showed the expression of EphA4 in a) Epitopes for another EphA4 antibody (sc-922P) and epitopes for EphA3 and21 antibody. Immunoreactivity to the same receptor epitope almost disappeared

426 L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
oligodeoxynucleotide (ODN) intrathecally using an osmoticminipump. To ensure that tissue disruption and edema did notprevent ODN diffusion and cellular uptake, a random sequenceof ODN-FITC conjugated was infused in sham and injured ratsfor 7 days. We observed diffusion of labeled oligonucleotide insham (Fig. 3A) and injured (Figs. 3B–D) rat spinal cords.Staining was observed several spinal segments rostral (Fig. 3C)and caudal (Fig. 3D) to lesion epicenter, demonstrating theability to widely deliver the ODN by intrathecal infusion.Cellular uptake was observed in neurons, white matter glialfibers, and within the cyst scar. A relevant point is that this ODNexpression was observed at the lumbar enlargement where spinalcentral pattern generators (CPGs) for locomotive rhythm arelocated (Fig. 3D). Excitatory interneurons of these CPGs areknown to express EphA4 (Kullander et al., 2003).
Blockade of EphA4 gene expression after ODN infusion wasdetermined by standardized RT-PCR at 7 DPI because, as shownin Fig. 1, the PCR products at earlier time points were lower thancontrol, and at 14 and 28 days, the differences in EphA4
Fig. 3. Blockade of EphA4 receptor expression by intrathecal/subdural infusion of ant(A) and injured (B–D) rats for 7 DPI. (B) Labeled ODN diffused throughout the lesion(C) and caudal (D) to the lesion site. (E–F) RT-PCR products from three antisenseAntisense oligonucleotide sequence infusion blocked the robust upregulation obserexpression. β-actin remained steady, confirming that gene blockade was specific for Etwo-tailed t test).
expression between control and injured animals were notsignificant. EphA4–antisense reduced gene expression 4-fold,relative to control-treated animals, after 7 days of infusion (Figs.3E–F). Meanwhile, β-actin expression remained unaffected byODN treatment. These results suggest that antisense ODNdiffuses, recognizes, and blocks, specifically and significantly,EphA4 gene expression in the injured spinal cord. Analysis ofspinal cord extracts at different time points after injury did notproduce an immunoreactive band in Western blot studies.Multiple antibodies from several companies were tested, and noprotein expression was observed. Probably, the preparation ofthe extract or the SDS-PAGE analysis induced a conformationalchange in the EphA4 protein, reducing its affinity for theantibody. Therefore, we assumed that the blockade of the proteinexpression would take place after the constant infusion of theantisense oligonucleotide since identical strategies have beenperformed to knock-down gene expression in vivo after traumaand monitored behavioral changes at later time points (Fundytuset al., 2002; Honore et al., 2002).
isense oligonucleotide. RandomODNs conjugated to FITC were infused to shamcavity formed after SCI. Diffusion and uptake were observed in segments rostral- and control-treated rats were analyzed in 1% agarose/ethidium bromide gels.ved at 7 DPI, while control oligonucleotides did not decrease EphA4 mRNAphA4 receptor. Data represent the mean±SEM, n=3, and **p<0.01 (Student's

427L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
EphA4 RTK does not play a role in locomotor recovery afterSCI
As shown in Figs. 4A–C, no changes were observedbetween EphA4–antisense infused rats and controls in BBBscore, grid walking, and beam crossing tests during the 28 daysof ODN administration. These results suggest that this receptordoes not play a role in locomotor recovery after SCI.
Spontaneous locomotor activity and mechanical allodynia areaffected with EphA4 gene suppression therapy
Changes in exploratory behavior had been described as areliable spontaneous measure of pain behavior in rats (Mills et
Fig. 4. EphA4–antisense ODN infusion has no effect in locomotor behavior. Nostatistical significance was observed between antisense- and control-treatedanimals for 28 days with respect to the locomotive assessments: (A) BBB score,(B) grid walking test, and (C) total sum of the square and round beam scores.Squares/dotted lines are antisense-treated rats; triangles/straight lines arecontrol-treated rats. Data represent the mean percent relative to control±SEM,n=8 (ANOVA followed by Bonferroni post hoc test; BBB: F=5.25; df=7,48;p>0.05; grid walking: F=4.281; df=7, 48; p>0.05; beam crossing: F=1.722;df=7, 48; p>0.05).
al., 2001). We evaluated horizontal activity (Fig. 5A), totaldistance (Fig. 5B), and number of movements (Fig. 5C) incontrol and antisense-treated rats. At 14 DPI, horizontalactivity and total distance are diminished in EphA4–antisenseinfused rats, even when the movements executed were similar(Fig. 5C), after the first 4 min (horizontal activity: F=14.125;df=9, 120; p<0.0001; total distance: F=18.058; df=9, 120;p<0.0001; number of movements: F=8.725; df=9, 120;p<0.0001). However, no differences in vertical activity,which depends on body weight support and hindlimblocomotion (white squares and triangles), were observedbetween groups, consistent with the results stated previously(Fig. 4). Antisense-treated rats moved for less time (Fig. 5D)and spent less time at the center of the cage (Fig. 5E) and moretime at the perimeter (Fig. 5F), suggesting a decreasedtendency to explore a novel environment (time of movement:F=16.844; df=9, 120; p<0.0001; time spent at center:F=2.212; df=9, 120; p<0.05; time spent at margin:F=2.005; df=9, 120; p<0.05). At 28 DPI, no significantchanges were observed between groups (data not shown), aresult we attribute to the habituation to the activity cageresulting in decreased exploratory behavior. In addition, shamrats were treated with either control (n=3) or antisense (n=3)oligonucleotides, and no significant differences were observedin the spontaneous behaviors of these animals (SupplementaryFig. 2). This result suggested that infusion of antisenseoligonucleotide was not the responsible factor to produce thechanges in spontaneous behavior since the sham-treatedanimals behave as naive-sham rats.
To determine if the changes in exploratory behavior wereattributed to abnormal nociception, we evaluated the rats'pain-evoked behavior. Using Semmes-Weinstein filaments, wemeasured somatic allodynia at 35 DPI. We could extrapolatethese results with the behavioral tests even when they weremeasured at different time points because central neuropathyis characterized by being highly persistent once it has beenestablished (Finnerup et al., 2003; Yoon et al., 2004).Knowing that chronic pain after SCI does not occur solelyin the paws, as usually measured in other paradigms, wemonitored pain responses mapping the anatomical areas testedafter a thoracic contusion (Fig. 6A). We observed thatEphA4–antisense-treated rats had an augmented response tonormally non-noxious stimuli when compared to controls(Fig. 6B). On the other hand, both groups were similar intheir response to subthreshold stimuli delivered to theforelimbs (Fig. 6B). These data served as negative controlbecause the forelimbs are not affected by the T10 contusionor the infused ODN. In addition, somatic allodynia wasmeasured in sham non-treated rats (n=8) with a 60 g targetforce in several quadrants and no response was obtained (datanot shown). Moreover, sham rats were treated with eithercontrol (n=3) or antisense (n=3) oligonucleotides, and noeffect was observed in the pain-evoked behaviors of theseanimals (data not shown). This result confirmed the role ofEphA4 expression in the injured spinal cord and demonstratedthat the infusion of the oligonucleotides was not responsiblefor increasing pain sensation. Since pain responses were

Fig. 5. EphA4–antisense ODN infusion for 28 days decreased spontaneous locomotor activity. Several exploratory parameters were analyzed for 20 min in theVersamax activity chamber. (A) The antisense-treated rats showed less horizontal activity than control-treated animals. This behavior is significantly differentwhen compared to the vertical activity (open squares for antisense- and open triangles for control-treated animals vertical). (B) The number of movements in bothgroups was similar, but the (C) total distance and (D) movement time were diminished after treatment with the antisense ODN. (E) Control-treated animals spentmore time at the center of the activity chamber compared to antisense-treated rats that preferred the (F) margins of the cage. Squares/dotted lines are antisense-treated rats; triangles/straight lines are control-treated rats. Data represent the mean±SEM, n=13, *p<0.05 and **p<0.01 (ANOVA followed by Bonferroni posthoc test).
428 L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
determined at subthreshold levels to keep the animals calm,the magnitude of any potential allodynic responses wasreduced or masked in contused animals treated with controloligonucleotides.
We considered the fact that we scored as positive only whenthe animal first manifested a response, regardless of how slightit was. This was done to ease the evaluation avoidingunnecessary discomfort and stress in the tested animals.Therefore, we decided to determine the number of animalsthat elicited a response to subthreshold stimulus, per quadrant,to assess more accurately the extent of abnormal pain sensitivityand to visualize the spread of allodynia. More antisense-treatedrats showed a response per quadrants affected within body partsthan control animals (Fig. 7). The quadrants more affected atsubthreshold levels of pain sensation were located in the upperand lower axis, as well as the left flank where 6 or 7 rats exerteda positive response in most quadrants (Fig. 7, antisense). Theseresults were different than control-treated animals in which thesame quadrants (at subthreshold levels) were responsive only in2 or 3 rats (Fig. 7, control). To determine the body part thatexhibited higher number of changes in mechanical allodynia,we calculated a sensitivity percent [total number of responsivequadrants / total number of quadrants within body part×100] for
both groups (Table 1). All body parts showed a higher percentof responsive quadrants within each body part in the antisense-treated rats than in control animals.
The pain threshold was estimated by comparing the mode ofthe targeted forces applied, ranking from 1.0 to 60.0 g. Incontrast to other paw withdrawal reflex studies (Christensen andHulsebosch, 1997), our rats were free and unrestrained. In thisstudy, any slight response was considered positive and noadditional force was applied in that precise quadrant in order tokeep the animals calm during evaluation. This approachallowed analysis of multiple regions of the body (not only thepaws), consistent with what is done in clinical studies (Finnerupet al., 2003). The most frequent (mode) targeted force at whichEphA4–antisense-treated animals showed a painful responsewas 4.0 g, while for controls it was 15.0 g (Table 1). Thisdecrease in mode of targeted force is consistent with a decreasein pain threshold observed in chronic pain in spinally injurypatients (Christensen and Hulsebosch, 1997; Finnerup et al.,2003).
To determine if enhanced aberrant sensory sprouting wasresponsible for the observed allodynia, we assessed the dorsalhorn for the expression of Substance P, CGRP, and serotonin.We could not find significant differences in immunoreactivity
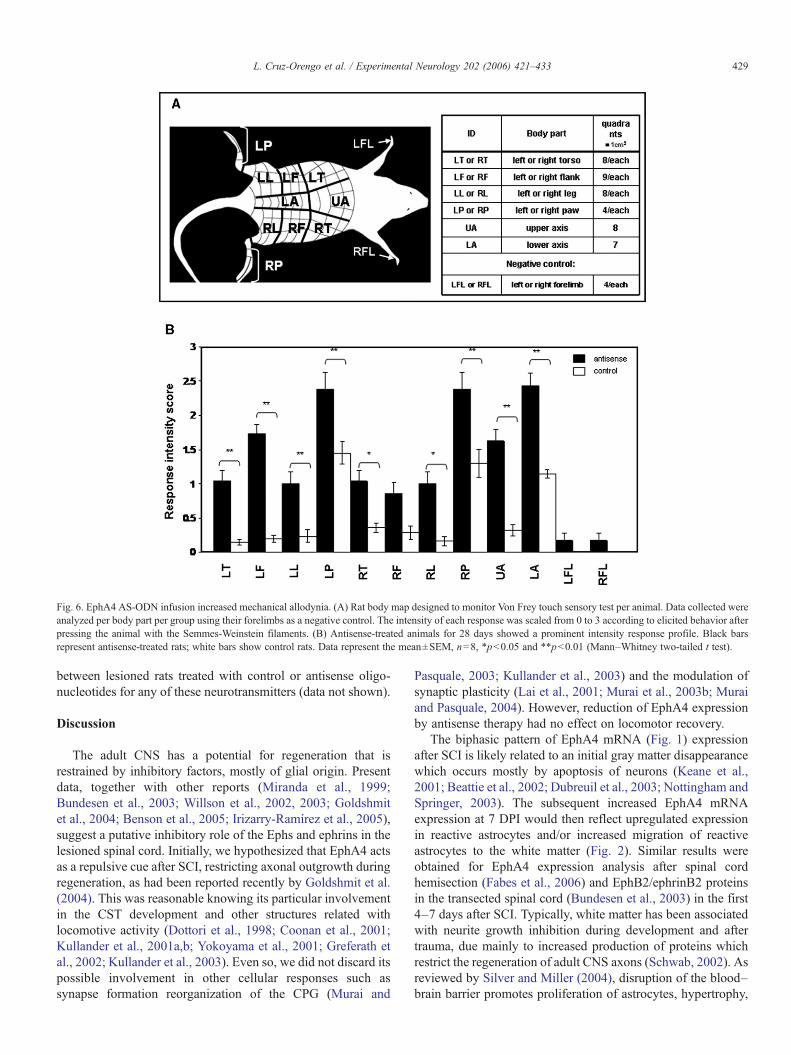
Fig. 6. EphA4 AS-ODN infusion increased mechanical allodynia. (A) Rat body map designed to monitor Von Frey touch sensory test per animal. Data collected wereanalyzed per body part per group using their forelimbs as a negative control. The intensity of each response was scaled from 0 to 3 according to elicited behavior afterpressing the animal with the Semmes-Weinstein filaments. (B) Antisense-treated animals for 28 days showed a prominent intensity response profile. Black barsrepresent antisense-treated rats; white bars show control rats. Data represent the mean±SEM, n=8, *p<0.05 and **p<0.01 (Mann–Whitney two-tailed t test).
429L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
between lesioned rats treated with control or antisense oligo-nucleotides for any of these neurotransmitters (data not shown).
Discussion
The adult CNS has a potential for regeneration that isrestrained by inhibitory factors, mostly of glial origin. Presentdata, together with other reports (Miranda et al., 1999;Bundesen et al., 2003; Willson et al., 2002, 2003; Goldshmitet al., 2004; Benson et al., 2005; Irizarry-Ramírez et al., 2005),suggest a putative inhibitory role of the Ephs and ephrins in thelesioned spinal cord. Initially, we hypothesized that EphA4 actsas a repulsive cue after SCI, restricting axonal outgrowth duringregeneration, as had been reported recently by Goldshmit et al.(2004). This was reasonable knowing its particular involvementin the CST development and other structures related withlocomotive activity (Dottori et al., 1998; Coonan et al., 2001;Kullander et al., 2001a,b; Yokoyama et al., 2001; Greferath etal., 2002; Kullander et al., 2003). Even so, we did not discard itspossible involvement in other cellular responses such assynapse formation reorganization of the CPG (Murai and
Pasquale, 2003; Kullander et al., 2003) and the modulation ofsynaptic plasticity (Lai et al., 2001; Murai et al., 2003b; Muraiand Pasquale, 2004). However, reduction of EphA4 expressionby antisense therapy had no effect on locomotor recovery.
The biphasic pattern of EphA4 mRNA (Fig. 1) expressionafter SCI is likely related to an initial gray matter disappearancewhich occurs mostly by apoptosis of neurons (Keane et al.,2001; Beattie et al., 2002; Dubreuil et al., 2003; Nottingham andSpringer, 2003). The subsequent increased EphA4 mRNAexpression at 7 DPI would then reflect upregulated expressionin reactive astrocytes and/or increased migration of reactiveastrocytes to the white matter (Fig. 2). Similar results wereobtained for EphA4 expression analysis after spinal cordhemisection (Fabes et al., 2006) and EphB2/ephrinB2 proteinsin the transected spinal cord (Bundesen et al., 2003) in the first4–7 days after SCI. Typically, white matter has been associatedwith neurite growth inhibition during development and aftertrauma, due mainly to increased production of proteins whichrestrict the regeneration of adult CNS axons (Schwab, 2002). Asreviewed by Silver and Miller (2004), disruption of the blood–brain barrier promotes proliferation of astrocytes, hypertrophy,

Table 1Sensitivity ratio and mode of targeted force
Body part Sensitivity percent
Antisense Control
Left torso 48.21% 14.29%Left flank 80.95% 19.05%Left leg 39.29% 10.71%Left paw 57.14% 21.43%Right torso 44.64% 35.71%Right flank 33.33% 17.46%Right leg 53.57% 10.71%Right paw 53.57% 21.43%Upper axis 67.86% 25.00%Lower axis 57.14% 12.24%Mode of targeted force 4.0 g 15.0 g
The sensitivity ratio was determined for each body part by adding the number ofquadrants from which was elicited any type of response as a ratio of the totalnumber of quadrants of n and multiplied by 100 or Sensitivity Ratio %=[∑n
responsive quadrants within body parts /body part quadrantsn]×100.EphA4-AS ODN infused rats showed a higher sensitivity in all body partscompared to controls. The mode of targeted force was determined as the mostcommon value in which a group elicits a response because we did not applystimuli near or above threshold. A lower targeted force was observed in theantisense-treated group than in control rats.
Fig. 7. Higher numbers of antisense-treated animals elicit a nociceptiveresponse. Color scale depicts the number of animals within a group that elicitany kind of response, from 0 to 3 as described in Fig. 6. Light gray is 0 rats anddark red is 7 rats. EphA4–antisense infusion results in a higher number ofanimals that were responsive to a given stimulus than control rats, as observedby the higher number of red colored quadrants of the (antisense) rat versus thelight and blue colored (control) rats.
430 L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
and rearrangement of their processes forming a barrier between“normal” and damaged tissue. Furthermore, they increase theproduction of proteins inhibitory for the regeneration of adultCNS axons such as tenascin, semaphorin-3, Slit proteins, andchondroitin sulfate proteoglycan (Raineteau and Schwab, 2001;Krautstrunk et al., 2002; Silver and Miller, 2004). Theseproteins, together with the resultant glial scar, assemble achemical and physical barrier for axonal regrowth. Theobserved expression of EphA4 after SCI in reactive astrocytessuggests that it may function as one of the inhibitory cues thatplay a part in the pathophysiology mediated by gliosis at thelesion site.
The ventrolateral funiculus at thoracic levels of the spinal cordcarries axons that are highly relevant to locomotor function (Loyet al., 2002a,b; Schucht et al., 2002). Therefore, upregulation ofEphA4 receptors coupled with reactive gliosis in these regionswas considered an interesting candidate as post-injury axonalinhibitor. An alternative role for EphA4 expression after SCI inthe adult spinal cord, but also related to locomotor behavior, wasthrough the reorganization of the CPG circuit (Raineteau andSchwab, 2001). Kullander and Klein (2002) identified glutama-tergic, EphA4-positive neurons as key components of the CPGcircuitry that helps in the local rewiring for development ofnormal locomotor behavior. However, our data suggest that theinhibitory effect of EphA4 expression may have a desirable andbeneficial response after trauma, unrelated to locomotivebehavior. As mentioned above, Eph/ephrins interactions areessential in CNS development for target recognition and specificsynapse formation (Klein, 2004). Therefore, if trauma triggers there-expression of molecular cues silenced during development(Emery et al., 2003), it is reasonable to propose that EphA4 ismediating the prevention of sensory fiber sprouting to avoidinappropriate synapse formation.
Sprouting is a common phenomenon after CNS injury andcan lead to plasticity that could be either beneficial (Weidner etal., 2001; Schwab, 2002) or detrimental (Ondarza et al., 2003;Abrams and Widenfalk, 2005). Spinally injured patients oftendevelop CNP which may result from inappropriate plasticitydue to sprouting (Christensen and Hulsebosch, 1997). Ifplasticity could be controlled or fine tuned, functional recoverymay be achieved without a concomitant development ofallodynia or other neuropathic conditions. Our results suggestthat endogenous repulsive molecular cues are able to attenuatenociceptive abnormalities by modulating plasticity. As demon-strated, antisense mediated reduction in EphA4 expressionexacerbated mechanical allodynia (Fig. 6 and Table 1), resultingin decreased locomotor activity (Fig. 5). These results aresignificant as the locomotor activity is not diminished by a lackof motility (Fig. 4) but could be related to an increase in pain-attributed discomfort.
In apparent contrast to the present study, Goldshmit et al.(2004) showed that EphA4−/− mice exhibited enhanced CSTand rubrospinal sprouting after SCI. This was mediated by adiminished astrocytic gliosis. While there were differences inspecies, rat vs. mouse; type of lesion, contusion vs. hemi-transection; and different behavioral paradigms, it is likely thatEphA4 differentially affects sensory and motor axonal sprout-ing. However, we did not observe large differences in GFAPimmunoreactivity in the antisense ODN treated spinal cords(data not shown), suggesting a possible effect of EphA4 onsensory axons. As described by Faulkner et al. (2004), reactiveastrocytes also provide tissue protection and resultant functionalpreservation. Therefore, the pattern of EphA4 expression inGFAP-positive cells after SCI and its possible involvement inlimiting pain-like behaviors suggest a putative role of thisreceptor and astrocytes as nociceptic synaptic modulators. The

431L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
astrocyte's participation in pain syndromes after peripheraltissue damage and other neuropathic pain models has beenpreviously described (McMahon et al., 2005), but not after CNStrauma itself. Our findings add a new dimension to theastrocyte's involvement, implicating their participation inmediating central changes.
However, we did not find changes in immunoreactivity ofneurotransmitter and modulators associated with nociceptiveinputs in the dorsal horn of treated animals (Hulsebosch, 2005).EphA4 could be regulating other nociceptive modulators sincechanges in CGRP immunoreactivity are not always associatedto mechanical hyperalgesia and/or inflammatory neuropathicpain (Barclay et al., 2002). The opioid system (Kohno et al.,2005; Mabuchi et al., 2003), peripheral benzodiazepinereceptors (Karchewski et al., 2004), non-classical pain media-tors recently described as the imidazole receptor [with agmatineas its ligand] (Fairbanks et al., 2000), and the GABA R/channelρ1 subunit sensitive to gabapentin (Zheng et al., 2003a,b) haveall been implicated in modulating pain responses.
We did observe in antisense-treated animals an increase in 5-HT expression in cells of the ventral horn and intermediate zoneof the gray matter, but not in lamina II (data not shown). Thisexpression is consistent with the sprouting of descendingpropriospinal tracts but not with a sensory sprouting (Oatway etal., 2005). This upregulation of 5-HT immunoreactivity wouldbe predicted to improve locomotor activity, as observed byOatway et al. (2005). We propose that they do not increase dueto their discomfort arising from the increased allodynia. Ourdata suggest that the increase in EphA4 expression after SCI isblocking aberrant sprouting that could promote the formation ofunwanted synapses and the development of allodynia. Under-standing the molecular mechanisms that regulate aberrantsprouting after SCI may suggest new therapeutic approachesto better control the onset of chronic neuropathic pain.
Acknowledgments
This work was supported by the NIH-SNRP (NS39405),MBRS-SCORE (S06-GM008224), MBRS-RISE (GM-68138),PR-EPSCOR (EPS-9874782), and the Associated Deanship ofBiomedical Sciences and Graduate Studies of the UPR Schoolof Medicine. Special thanks to Ioannis M. Torres for herexcellent technical support and to the RCMI Image AnalysisResource Center (G12RR03051), the Experimental SurgicalResearch Laboratory and the Animal Resource Center at theUPR Medical Sciences Campus for the use of their facilities.
Appendix A. Supplementary data
Supplementary data associated with this article can be found,in the online version, at doi:10.1016/j.expneurol.2006.07.005.
References
Abrams, M., Widenfalk, J., 2005. Emerging strategies to promote improvedfunctional outcome after peripheral nerve injury. Restor. Neurol. Neurosci.23 (5–6), 367–382.
Aguayo, A.J., David, S., Bray, G.M., 1981. Influences of the glial environmenton the elongation of axons after injury: transplantation studies in adultrodents. J. Exp. Biol. 95, 231–240.
Barclay, J., Patel, S., Dorn, G., Wotherspoon, G., Moffatt, S., Eunson, L.,Abdel'al, S., Natt, F., Hall, J., Winter, J., Bevan, S., Wishart, W., Fox, A.,Ganju, P., 2002. Functional downregulation of P2X3 receptor subunit in ratsensory neurons reveals a significant role in chronic neuropathic andinflammatory pain. J. Neurosci. 22 (18), 8139–8147.
Basso, D.M., Beattie, M.S., Bresnahan, J.C., 1995. A sensitive and reliablelocomotor rating scale for open field testing in rats. J. Neurotrauma 12 (1),1–21.
Battaglia, A.A., Sehayek, K., Grist, J., McMahon, S.B., Gavazzi, I., 2003. EphBreceptors and ephrin-B ligands regulate spinal sensory connectivity andmodulate pain processing. Nat. Neurosci. 6 (4), 339–340.
Beattie, M.S., Hermann, G.E., Rogers, R.C., Bresnahan, J.C., 2002. Cell deathin models of spinal cord injury. Prog. Brain Res. 137, 37–47.
Behrmann, D.L., Bresnahan, J.C., Beattie, M.S., Shah, B.R., 1992. Spinal cordinjury produced by consistent mechanical displacement of the cord in rats:behavioral and histologic analysis. J. Neurotrauma 9 (3), 197–217.
Benson, M.D., Romero, M.I., Lush, M.E., Lu, Q.R., Henkemeyer, M., Parada,L.F., 2005. Ephrin-B3 is a myelin-based inhibitor of neurite outgrowth. Proc.Natl. Acad. Sci. U. S. A. 102 (30), 10694–10699.
Biervert, C., Horvath, E., Fahrig, T., 2001. Semiquantitative expression analysisof ephrin-receptor tyrosine kinase mRNA's in a rat model of traumatic braininjury. Neurosci. Lett. 315, 25–28.
Bundesen, L.Q., Scheel, T.A., Bregman, B.S., Kromer, L.F., 2003. Ephrin-B2and EphB2 regulation of astrocyte–meningeal fibroblast interactions inresponse to spinal cord lesions in adult rats. J. Neurosci. 23 (21), 7789–7800.
Christensen, M.D., Hulsebosch, C.E., 1997. Chronic central pain after spinalcord injury. J. Neurotrauma 14 (8), 517–537.
Conover, J.C., Doetsch, F., Garcia-Verdugo, J.M., Gale, N.W., Yancopoulos,G.D., Alvarez-Buylla, A., 2000. Disruption of Eph/ephrin signaling affectsmigration and proliferation in the adult subventricular zone. Nat. Neurosci.3 (11), 1091–1097.
Coonan, J.R., Greferath, U., Messenger, J., Hartley, L., Murphy, M., Boyd,A.W., Dottori, M., Galea, M.P., Bartlett, P.F., 2001. Development ofreorganization of corticospinal projections in EphA4 deficient mice.J. Comp. Neurol. 436 (2), 248–262.
Dottori, M., Hartley, L., Galea, M.P., Paxinos, G., Palizzotto, M., Kilpatrick, T.,Bartlet, P.F., Murphy, M., Köntgen, F., Boyd, A.W., 1998. EphA4 receptorprotein tyrosine kinase is required for the development of the corticospinaltract. Proc. Natl. Acad. Sci. 95 (22), 13248–13253.
Dubreuil, C.I., Winton, M.J., McKerracher, L., 2003. Rho activation patternsafter spinal cord injury and the role of activated Rho in apoptosis in thecentral nervous system. J. Cell Biol. 162 (2), 233–243.
Eberhart, J., Swartz, M., Koblar, S.A., Pasquale, E.B., Tanaka, H., 2000.Expression of EphA4, ephrin-A2 and ephrin-A5 during axon outgrowth tothe hindlimb indicates potential roles in the pathfinding. Dev. Neurosci. 22(3), 237–250.
Emery, D.L., Royo, N.C., Fischer, I., Saatman, K.E., McIntosh, T.K., 2003.Plasticity following injury to the adult central nervous system: isrecapitulation of a developmental state worth promoting? J. Neurotrauma12, 1271–1292.
Fabes, J., Anderson, P., Yáñez-Muñoz, R.J., Thrasher, A., Brennan, C.,Bolsover, S., 2006. Accumulation of the inhibitory receptor EphA4 mayprevent regeneration of corticospinal tract axons following lesion. Eur. J.Neurosci. 23, 1721–1730.
Fairbanks, C.A., Schreiber, K.L., Brewer, K.L., Yu, C.G., Stone, L.S., Kitto,K.F., Nguyen, H.O., Grocholski, B.M., Shoeman, D.W., Kehl, L.J.,Regunathan, S., Reis, D.J., Yezierski, R.P., Wilcox, G.L., 2000. Agmatinereverses pain induced by inflammation, neuropathy, and spinal cord injury.Proc. Natl. Acad. Sci. 97 (19), 10584–10589.
Faulkner, J.R., Herrmann, J.E., Woo, M.J., Tansey, K.E., Doan, N.B., Sofroviev,M., 2004. Reactive astrocytes protect tissue and preserve function afterspinal cord injury. J. Neurosci. 24 (9), 2143–2155.
Finnerup, N.B., Johannesen, I.L., Fuglsang-Frederiksen, A., Bach, F.W., Jensen,T.S., 2003. Sensory function in spinal cord injury patients with and withoutcentral pain. Brain 126, 57–70.

432 L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
Fundytus, M.E., Osborne, M.G., Henry, J.L., Coderre, T.J., Dray, A., 2002.Antisense oligonucleotide knockdown of mGluR1 alleviates hyperalgesiaand allodynia associated with chronic inflammation. Pharmacol. Biochem.Behav. 73 (2), 401–410.
Goldshmit, Y., Galea, M.P., Graham, W., Barlett, P.F., Turnley, A.M., 2004.Axonal regeneration and lack of astrocytic gliosis in EphA4-deficient mice.J. Neurosci. 24 (45), 10064–10075.
Greferath, U., Canty, A.J., Messenger, J., Murphy, M., 2002. Developmentalexpression of EphA4-tyrosine kinase receptor in the mouse brain and spinalcord. Gene Expression Pattern 2, 267–274.
Gruner, J.A., 1992. A monitored contusion model of spinal cord injury in the rat.J. Neurotrauma 9 (2), 123–126.
Helmbacher, F., Schneider-Maunoury, S., Topilko, P., Tiret, L., Charnay, P.,2000. Targeting of EphA4 tyrosine kinase receptors affects dorsal/ventralpathfinding of limbs motor axons. Development 127 (15), 3313–3324.
Honore, P., Kage, K., Mikusa, J., Watt, A.T., Johnston, J.F., Wyatt, J.R.,Faltynek, C.R., Jarvis, M.F., Lynch, K., 2002. Analgesic profile ofintrathecal P2X3 antisense oligonucleotide treatment in chronic inflamma-tory and neuropathic pain states in rats. Pain 99 (1–2), 11–19.
Hulsebosch, C.E., 2005. From discovery to clinical trials: treatment strategiesfor central neuropathic pain after spinal cord injury. Curr. Pharm. Des. 11(11), 1411–1420.
Irizarry-Ramírez, M., Willson, C.A., Cruz-Orengo, L., Figueroa, J., Velázquez,I., Jones, H., Foster, R.D., Whittemore, S.R., Miranda, J.D., 2005.Upregulation of EphA3 receptor after spinal cord injury. J. Neurotrauma22 (8), 929–935.
Karchewski, L.A., Bloechlinger, S., Woolf, C.J., 2004. Axonal injury-dependentinduction of the peripheral benzodiazepine receptor in small-diameter adultrat primary sensory neurons. Eur. J. Neurosci. 20 (3), 671–683.
Keane, R.W., Kraydieh, S., Lotocki, G., Bethea, J.R., Krajewski, S., Reed, J.C.,Dietrich, W.D., 2001. Apoptotic and anti-apoptotic mechanisms followingspinal cord injury. J. Neuropathol. Exp. Neurol. 60 (5), 422–429.
Klein, R., 2004. Eph/ephrin signaling in morphogenesis, neural developmentand plasticity. Curr. Opin. Cell Biol. 16 (5), 580–589.
Knöll, B., Isenmann, S., Kilic, E., Walkenhorst, J., Engel, S., Wehinger, J., Bähr,M., Drescher, U., 2001. Graded expression patterns of ephrin-As in thesuperior colliculus after lesion of the adult mouse optic nerve. Mech. Dev.106 (1–2), 119–127.
Kohno, T., Ji, R.R., Ito, N., Allchorne, A.J., Befort, K., Karchewski, L.A.,Woolf,C.J., 2005. Peripheral axonal injury results in reduced mu opioid receptorpre- and post-synaptic action in the spinal cord. Pain 117 (1–2), 77–87.
Krautstrunk, M., Scholtes, F., Martin, D., Schoenen, J., Schmitt, A.B., Plate, D.,Nacimiento, W., Noth, J., Brook, G.A., 2002. Increased expression of theputative axon growth-repulsive extracellular matrix molecule, keratansulphate proteoglycan, following traumatic injury of the adult rat spinalcord. Acta Neuropathol. 104 (6), 592–600.
Kullander, K., Klein, R., 2002. Mechanisms and functions of Eph and ephrinssignaling. Nat. Rev., Mol. Cell Biol. 3, 475–486.
Kullander, K., Croll, S.D., Zimmer, M., Pan, L., McClain, J., Hughes, V.,Zabski, S., De Chiara, T.M., Klein, R., Yancopoulos, G.D., Gale, N.W.,2001a. Ephrin-B3 is the midline barrier that prevents corticospinal tractaxons from recrossing, allowing for unilateral motor control. Genes Dev.15 (7), 877–888.
Kullander, K., Mather, N.K., Diella, F., Dottori, M., Boyd, A.W., Klein, R.,2001b. Kinase-dependent and kinase-independent functions of EphA4receptors in major axon tract formation in vivo. Neuron 29 (1), 73–84.
Kullander, K., Butt, S.J.B., Lebret, J.M., Lunfald, L., Restrepo, C.E., Rydström,A., Klein, R., Kiehn, O., 2003. Role of EphA4 and ephrinB3 in localneuronal circuits that control walking. Science 299 (5614), 1889–1892.
Kwon, B.K., Tetzlaff, W., 2001. Spinal cord regeneration: from gene totransplants. Spine 26 (Suppl. 24), S13–S22.
Lai, K.O., Ip, F.C., Cheung, J., Fu, A.K., Ip, N.Y., 2001. Expression of Ephreceptors in the skeletal muscle and their localization at the neuromuscularjunction. Mol. Cell. Neurosci. 17 (6), 1034–1047.
Liu, X., Hawkes, E., Ishimaru, T., Tran, T., Sretavan, D.W., 2006. EphB3: anendogenous mediator of adult axonal plasticity and regrowth after CNSinjury. J. Neurosci. 26 (12), 3087–3101.
Loy, D.N., Magnunson, D.K.S., Zhang, Y.P., Onifer, S.M., Mills, M.D., Cao, Q.,
Darnall, J.B., Fajardo, L.C., Burke, D.A., Whittemore, S.R., 2002a.Functional redundancy of ventral spinal locomotor pathways. J. Neurosci.22 (1), 315–323.
Loy, D.N., Talbott, J.F., Onifer, S.M., Mills, M.D., Burke, D.A., Dennison, J.B.,Fajardo, L.C., Magnunson, D.K.S., Whittemore, S.R., 2002b. Both dorsaland ventral spinal cord pathways contribute to overground locomotion in theadult rat. Exp. Neurol. 177 (2), 575–580.
Mabuchi, T., Matsumura, S., Okuda-Ashitaka, E., Kitano, T., Kojima, H.,Nagano, T., Minami, T., Ito, S., 2003. Attenuation of neuropathic pain by thenociceptin/orphanin FQ antagonist JTC-801 is mediated by inhibition ofnitric oxide production. Eur. J. Neurosci. 17, 1384–1392.
Martone, M.E., Holash, J.A., Bayardo, A., Pasquale, E.B., Ellisman, M.H.,1997. Immunolocalization of the receptor tyrosine kinase EphA4 in the adultrat central nervous system. Brain Res. 771 (2), 238–250.
McKerracher, L., David, S., Jackson, J.L., Kottis, V., Dunn, R., Braun, P.E.,1994. Identification of myelin-associated glycoprotein as a major myelin-derived inhibitor of neurite growth. Neuron 13 (4), 805–811.
McMahon, S.B., Cafferty, W.B.J., Marchand, F., 2005. Immune and glialcell factors as pain mediators and modulators. Exp. Neurol. 192,444–462.
Merkler, D., Metz, G.A.S., Raineteau, O., Dietz, V., Schwab, M.E., Fouad, K.,2001. Locomotor recovery in spinal cord-injured rats treated with anantibody neutralizing the myelin-associated neurite growth inhibitorNogo-A. J. Neurosci. 21 (10), 3665–3673.
Mills, C.D., Grady, J.J., Hulsebosch, C.E., 2001. Changes in exploratorybehavior as a measure of chronic central pain following spinal cord injury.J. Neurotrauma 8 (10), 1091–1105.
Miranda, J.D., White, L.A., Marcillo, A.E., Willson, C.A., Jagid, J.,Whittemore, S.R., 1999. Induction of Eph B3 after spinal cord injury.Exp. Neurol. 156 (1), 218–222.
Moreno-Flores, M.T., Wandosell, F., 1999. Upregulation of Eph receptors afterexcitotoxic injury in adult hippocampus. Neuroscience 91 (1), 193–201.
Mukhopadyay, G., Doherty, P., Walsh, F.S., Crocker, P.R., Filbin, M.T., 1994. Anovel role for myelin-associated glycoprotein as an inhibitor of axonalregeneration. Neuron 13 (3), 757–767.
Murai, K.K., Pasquale, E.B., 2002. Can Eph receptors stimulate the mind?Neuron 33, 159–162.
Murai, K.K., Pasquale, E.B., 2003. ‘Eph’ective signaling: forward, reverse andcrosstalk. J. Cell Sci. 116, 2823–2832.
Murai, K.K., Pasquale, E.B., 2004. Eph receptors, ephrins, and synapticfunction. The Neuroscientist 10 (4), 304–314.
Murai, K.K., Nguyen, L.N., Irie, F., Yamaguchi, Y., Pasquale, E.B., 2003a.Control of hippocampal dendritic spine morphology through ephrinA3/EphA4 signaling. Nat. Neurosci. 6 (2), 153–160.
Murai, K.K., Nguyen, L.N., Koolpe, M., McLennan, R., Krull, C.E.,Pasquale, E.B., 2003b. Targeting the EphA4 receptor in the nervoussystem with the biologically active peptides. Mol. Cell. Neurosci. 24 (4),1000–1011.
Nottingham, S.A., Springer, J.E., 2003. Temporal and spatial distribution ofactivated caspase3 after subdural kainic acid infusions in rat spinal cord.J. Comp. Neurol. 464 (4), 463–471.
Oatway, M.A., Chen, Y., Bruce, J.C., Dekaban, G.A., Weaver, L.C., 2005.Anti-CD11d integrin antibody treatment restores normal serotonergicprojections to the dorsal, intermediate, and ventral horns of the injuredspinal cord. J. Neurosci. 25 (3), 637–647.
Ondarza, A.B., Ye, Z., Hulsebosch, C.E., 2003. Direct evidence of primaryafferent sprouting in distant segments following spinal cord injury inthe rat: colocalization of GAP-43 and CGRP. Exp. Neurol. 184 (1),373–380.
Pasquale, E.B., 2005. Eph receptor signaling casts a wide net on cell behavior.Nat. Rev., Mol. Cell Biol. 6, 462–475.
Raineteau, O., Schwab, M.E., 2001. Plasticity of motor systems after incompletespinal injury. Nat. Rev., Neurosci. 2, 263–273.
Sambrook, J., Russell, D.W., 2001. Molecular Cloning, 3rd ed. Cold SpringHarbor Laboratory Press, Cold Spring Harbor, NY.
Schnell, L., Schwab, M.E., 1990. Axonal regeneration in the rat spinal cordproduced by an antibody against myelin-associated neurite growthinhibitors. Nature 343 (6252), 269–272.

433L. Cruz-Orengo et al. / Experimental Neurology 202 (2006) 421–433
Schucht, P., Raineteau, O., Schwab, M.E., Foauad, K., 2002. Anatomicalcorrelates of locomotor recovery following dorsal and ventral lesions of therat spinal cord. Exp. Neurol. 176 (1), 143–153.
Schwab, M.E., 2002. Increasing plasticity and functional recovery of thelesioned spinal cord. Prog. Brain Res. 137, 351–359.
Silver, J., Miller, J.H., 2004. Regeneration beyond the glial scar. Nat. Rev.,Neurosci. 5, 146–156.
Sobel, R., 2005. Ephrin A receptors and ligands in lesions and normal-appearingwhite matter in multiple sclerosis. Brain Pathol. 15, 35–45.
Wang, K.C., Kaprovica, V., Kim, J.A., Sivansankaran, R., Guo, Y., Neve,R.L., He, Z., 2002. Oligodendrocyte-myelin glycoprotein is a NOGOreceptor ligand that inhibits neurite outgrowth. Nature 417 (6892),941–944.
Weidner, N., Ner, A., Salimi, N., Tuszynski, M.H., 2001. Spontaneouscorticospinal axonal plasticity and functional recovery after adult centralnervous system injury. Proc. Natl. Acad. Sci. 98 (6), 3513–3518.
Willson, C.A., Irizarry-Ramírez, M., Gaskins, H.E., Cruz-Orengo, L.,Figueroa, J.D., Whittemore, S.R., Miranda, J.D., 2002. Up-regulation of
EphA receptor expression in the injured adult spinal cord. Cell Transplant11 (3), 229–239.
Willson, C.A., Miranda, J.D., Foster, R.D., Onifer, S.M., Whittemore, S.R.,2003. Transection of the adult rat spinal cord upregulates EphB3 receptorand ligand expression. Cell Transplant 12, 279–290.
Yokoyama, N., Romero, M.I., Cowan, C.A., Galvan, P., Helmbacher, F.,Charnay, P., Parada, L.F., Henkemeyer, M., 2001. Forward signaling ofephrin-B3 prevents contralateral corticospinal axons from recrossing thespinal cord midline. Neuron 29 (1), 85–97.
Yoon, Y.W., Dong, H., Arends, J.J.A., Jacquin, M.F., 2004. Mechanical and coldallodynia in a rat spinal cord contusion model. Somatosens. Motor Res. 21(1), 25–31.
Zheng, B., Ho, C., Li, S., Keirstead, H., Steward, O., Tessier-Lavigne, M.,2003a. Lack of enhanced spinal regeneration in nogo-deficient mice.Neuron 38, 213–224.
Zheng, W., Xie, W., Zhang, J., Strong, J.A., Wang, L., Yu, L., Xu, M., Lu, L.,2003b. Function of δ-aminobutyric acid receptor/channel ρI subunits inspinal cord. J Biol. Chem. 278 (8), 48321–48329.