
Novel nucleoside analogues with bases modified with (β-halo ...
Upload
independentCategory
view
0download
0
Purine Restriction Induces Pronounced TranslationalUpregulation of the NT1 Adenosine/Pyrimidine NucleosideTransporter in Leishmania major
Diana Ortiz1,*, Raquel Valdés1,*, Marco A. Sanchez1, Johanna Hayenga1, Carolyn Elya1,Siegfried Detke2, and Scott M. Landfear1,*
1Department of Molecular Microbiology and Immunology, Oregon Health & Science University,Portland, Oregon 972392Department of Biochemistry and Molecular Biology, University of North Dakota, Grand Forks,North Dakota 58202
SummaryLeishmania and other parasitic protozoa are unable to synthesize purines de novo and are reliantupon purine nucleoside and nucleobase transporters to import preformed purines from their hosts.To study the roles of the four purine permeases NT1-NT4 in Leishmania major, null mutants ineach transporter gene were prepared and the effect of each gene deletion on purine uptake wasmonitored. Deletion of the NT3 purine nucleobase transporter gene or both NT3 and the NT2nucleoside transporter gene resulted in pronounced upregulation of adenosine and uridine uptakemediated by the NT1 permease and also induced up to a 200-fold enhancement in the level of theNT1 protein but not mRNA. A similar level of upregulation of NT1 was achieved in wild typepromastigotes that were transferred to medium deficient in purines. Pulse labeling and treatment ofcells with the translation inhibitor cycloheximide revealed that control of NT1 expression occursprimarily at the level of translation and not protein turnover. These observations imply theexistence of a translational control mechanism that enhances the ability of Leishmania parasites toimport essential purines when they are present at limiting concentrations.
KeywordsPurines; transport; nutritional limitation; Leishmania major; translational control
IntroductionParasitic protozoa such as Leishmania, Trypanosoma and Plasmodium are unable tosynthesize purines de novo and are completely reliant upon their hosts to provide them withpreformed purines (Hammond and Gutteridge, 1984). Since most cells in vertebrates arecapable of purine biosynthesis, the requirement of these protozoa for host-derived purines isone of the principal biochemical distinctions between the host and parasite and represents apotential target for therapeutic intervention. To scavenge purines, these parasites haveevolved a unique set of purine transporters and salvage enzymes that allow them to take up avariety of purine nucleosides and nucleobases and convert them into essential purinemetabolites (Carter et al., 2008). The purine nucleoside and nucleobase transporters ofLeishmania parasites (Landfear et al., 2004) are high affinity concentrative proton-coupled
*For correspondence. [email protected]; Tel. (+1) 503 494 2426; Fax (+1) 503 494 6862.*These two authors contributed equally to this work and are both designated first authors.
NIH Public AccessAuthor ManuscriptMol Microbiol. Author manuscript; available in PMC 2011 October 1.
Published in final edited form as:Mol Microbiol. 2010 October ; 78(1): 108–118. doi:10.1111/j.1365-2958.2010.07328.x.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

permeases (Ortiz et al., 2009a; Stein et al., 2003) that allow the parasites to efficientlycompete with the host for these critical nutrients
Leishmania major and related Leishmania species express two purine nucleosidetransporters, NT1 (encompassing two closely related isoforms NT1.1 and NT1.2) thatmediates the uptake of adenosine (Ado) and the pyrimidine nucleosides (Vasudevan et al.,1998) and NT2 that takes up guanosine (Guo), inosine (Ino), and xanthosine (Xao) (Carter etal., 2000). In addition NT3 and NT4 are purine nucleobase transporters that importhypoxanthine (Hyp), xanthine (Xan), adenine (Ade), and guanine (Gua). NT3 functionsoptimally at neutral pH and appears to be the major nucleobase transporter in thepromastigote stage of the life cycle that lives in the gut of the sandfly vector (Ortiz et al.,2007). In contrast, NT4 has very low transport activity at neutral pH but is functional in theacidic pH range to which these parasites are exposed in the amastigote stage that lives insideacidified phagolysomal vesicles of vertebrate host macrophages (Ortiz et al., 2009a).Collectively these permeases provide conduits for the efficient import of essential purines inboth insect stage promastigotes and vertebrate stage amastigotes.
To understand the role of each permease in the Leishmania life cycle, null mutants for eachNT (nucleoside/nucleobase transporter) gene in L. major have been generated ((Ortiz et al.,2007) and this paper). In the process of characterizing such mutants, we observed thatdeletion of the NT3 genes (Δnt3 mutant) or both the NT2 and NT3 genes (Δnt2Δnt3 mutant)promoted a pronounced enhancement of Ado uptake activity (up to 60-fold) and of NT1protein expression (up to 200-fold) but only a small (~3-fold) increase in NT1 mRNA.Consistent with parallel studies of our colleagues on purine starvation in L. donovani (seeaccompanying manuscript), removal of purines from the medium of wild type L. majorpromastigotes induced a similar upregulation in expression of the NT1 transporters. Uptakeassociated with NT2, NT3, and NT4 were also enhanced by purine starvation but to a lesserdegree than for NT1.These results indicate the existence of a robust regulatory mechanism,operating primarily at the level of mRNA translation, that increases the purine salvagecapacity of these parasites in the face of purine limitation. Regulation of purine transporterexpression probably promotes the ability of Leishmania parasites to survive fluctuations inthe concentrations of available purines during their life cycle by increasing their capacity toimport these essential nutrients when they are present at limiting levels.
ResultsNull mutants in purine transporter genes upregulate the expression of the NT1 adenosine-pyrimidine transporter
Purines are essential nutrients that Leishmania import from their hosts using nucleoside(NT1 and NT2) and nucleobase (NT3 and NT4) permeases. To determine the biologicalfunction of different NTs in L. major, null mutants for each NT gene were systematicallygenerated and characterized. Examination of the Δnt3 mutant, which is deficient in themajor purine nucleobase transporter NT3 (Ortiz et al., 2007), revealed a significantly higherrate of uptake for the nucleosides Ado and Urd (11.4 ± 2.9-fold, n = 5, for Ado; 12.3 ± 2.0-fold, n = 2, for Urd), both substrates for the NT1 transporter (Vasudevan et al., 1998),compared to the uptake mediated by wild type parasites (Fig. 1A shows one suchexperiment). Furthermore, a further increase in uptake of Ado and Urd (18.6 ± 6.7-fold, n =5, for Ado; 19.8 ± 4.3-fold, n = 2, for Urd) was observed when the NT2 gene was alsodeleted in the Δnt3 background to generate the Δnt2Δnt3 double null mutant.Complementation of the Δnt3 or Δnt2Δnt3 mutants with the NT3 gene expressed from anepisome greatly reduced the rate of uptake mediated by NT1 observed in theuncomplemented null mutants. This observation confirms that the induction of Ado and Urd
Ortiz et al. Page 2
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

uptake observed in the null mutants is due to deletion of the purine transporter genes and notto some unrelated genetic modification that had occurred in each mutant line.
To investigate whether the upregulation of uptake mediated by NT1 corresponded to anincrease in the level of NT1 protein, immunoblots (Fig. 1B) were performed using an anti-NT1 antibody directed against the large intracellular hydrophilic loop betweentransmembrane domains 6 and 7 of the L. mexicana NT1 transporter (Detke, 2007). Thelevel of NT1 protein in isolated membranes of wild type parasites was barely detectable. Incontrast, for Δnt3 two bands with apparent molecular weights between 43 and 45 kDa wereobserved. The identity of the ~43–45 kDa bands as NT1 permeases, and hence thespecificity of the anti-NT1 antibody, was confirmed by the observation that these bandswere barely detectable in isolated membranes from wild type L. major promastigotes, butthey were distinctly expressed in wild type parasites that had been complemented byexpressing NT1 from an episomal copy of the gene (WT[pNT1], Fig. 1B). The level ofexpression of these bands was further increased in the Δnt2Δnt3 null mutant. Moreover,complementation of both the Δnt3 and Δnt2Δnt3 null mutants with the NT3 gene expressedepisomally dramatically downregulated the expression of NT1. These results indicate thatupregulation of Ado and Urd uptake in both null mutants involves an increase in the level ofthe NT1 protein and does not result simply from upregulation of intracellular nucleosidemetabolism.
Quantitative analysis of NT1 overexpression in null mutantsTo quantify the overexpression of NT1 in the Δnt3 and Δnt2Δnt3 null mutants, serialdilutions of the membrane preparation from these mutants were performed and the level ofthe NT1 protein was analyzed by immunoblot. Starting with an initial 20 µg of Δnt3 orΔnt2Δnt3 membrane proteins, dilutions of each membrane preparation were made until thesignal representing the ~43–45 kDa bands could barely be detected and was similar to thesignal detected in the corresponding blot representing 20 µg of wild type membraneproteins. This comparison provides a minimum estimate for the upregulation of NT1 proteinexpression in each null mutant. The results revealed that the level of expression of NT1 inΔnt3 cells was upregulated ~20-fold (Fig. 2A), similar to the increase in Ado uptake (~10-fold increase). In Δnt2Δnt3, the level of NT1 was upregulated by ~200-fold when comparedto the barely detectable level of NT1 in wild type membranes (Fig. 2B). This estimated~200-fold increase in NT1 protein is larger than the ~20-fold increase of Ado uptakeobserved in this mutant. This discrepancy is likely due to the fact that the two experimentsmeasure different entities. Thus some of the overexpressed NT1 protein, detected in totalcellular membranes, may not traffic to the cell surface and thus may not contribute to uptakeof Ado.
Supplementation with guanosine attenuates NT1 upregulation in the Δnt3 null mutantA likely explanation for upregulation of the NT1 transporter in Δnt3 null mutants is thatremoval of the major purine nucleobase transporter results in limitation of intracellularpurines. These parasites may then respond to purine limitation by upregulating expression ofremaining purine permeases so that homeostasis with regard to essential purines may beachieved. The further upregulation of NT1 expression in the Δnt2Δnt3 double null mutantmight result from additional purine limitation in parasites that are also impaired in theirability to import the substrates of NT2, Guo, Ino, and Xao. Although purine nucleobases arepresent at µM levels in M199 medium and the medium was also supplemented with 100 µMXan, purine nucleosides that could be imported through the NT2 permease may also bepresent in the serum supplement employed for growth of the parasites, thus resulting inincreased purine limitation when the NT2 gene is deleted. If this explanation for the relativelevels of NT1 expression in Δnt3 and Δnt2Δnt3 null mutants is correct, then supplementing
Ortiz et al. Page 3
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

the medium with an NT2 ligand such as Guo might downregulate the level of expression ofNT1 in the Δnt3 mutant. To assess this possibility, both null mutants were grown with orwithout Guo and the level of NT1 expression was monitored by Ado uptake andimmunoblot over a range of time points (Fig. 3A, B). Indeed, the rate of Ado uptake and thelevel of NT1 expression in the Δnt3 null mutant were strongly downregulated (~12-fold)after 24 h of supplementation with Guo. In contrast, cells growing in medium supplementedwith Xan did not change the rate of uptake, and only a small reduction in the level of NT1protein was observed. This modest reduction in NT1 expression may be due to the increasein cell density that occurred during the course of this experiment, since NT1 expression isdownregulated at higher cell densities (data not shown). In contrast for the Δnt2Δnt3 cellline, growth in medium containing Guo or Xan for 24 h resulted in essentially the sameincrease in the rate of Ado uptake (~15-fold versus ~19-fold, Fig. 3C) or the level ofexpression of NT1 (Fig. 3D) when compared with wild type parasites grown under the sameconditions. Thus upregulation of NT1 expression in the Δnt3 null mutant can be reversed byGuo, but this reversal is not observed in the Δnt2Δnt3 null mutant that cannot transport Guo.This result is consistent with the notion that upregulation of NT1 expression in purinetransporter null mutants results from limitation of intracellular purine levels.
Expression of NT1 in other purine transporter null mutantsThe previous results reveal that expression of the NT1 permease is upregulated when theNT3 or both the NT2 and NT3 genes are deleted by homologous gene replacement. Todetermine whether expression of the NT1 transporter is also modulated by other purinepermeases, the level of NT1 protein was monitored in the Δnt2 and Δnt4 null mutants.Immunoblots of membranes (data not shown) indicated no significant induction of NT1 ineither the Δnt2 or Δnt4 mutants. The failure of these mutants to upregulate NT1 protein maybe due to the fact that, similar to the experiments in Figs. 1 and 2, these parasites werecultured in medium containing purine nucleobases (M199 medium complemented with 15%FBS and 100 µM Xan). Since these null mutants still express the major purine nucleobasetransporter NT3, they probably do not experience purine deprivation under these growthconditions.
The preceding observations suggested that NT1 might be upregulated in Δnt2 null mutants ifthese parasites were grown in medium that contained Guo as the sole purine source, as theywould then experience purine deprivation compared to wild type parasites. In the experimentshown in Fig. 4A, cells were grown in RPMI 1640 medium, which contains no purines,supplemented with 20% dialyzed FBS and 100 µM Guo. Under these conditions, Adouptake was upregulated ~8-fold for the Δnt2 null mutant compared to wild type cells. Forthe Δnt2Δnt3 null mutant, there was an even greater (~45-fold) upregulation of Ado uptakecompared to wild type cells. When these null mutants were complemented with NT2expressed episomally, the uptake of Ado was considerably downregulated. The substantiallygreater enhancement of Ado uptake in Δnt2Δnt3 null mutants compared to the Δnt2 line maybe the consequence of hydrolysis of Guo to the nucleobase Gua during growth of cells inculture, mediated by nucleoside hydrolases and nucleoside phosphorylases (Hassan andCoombs, 1986;Shi et al., 1999) expressed by these parasites and released into the mediumby dying cells. Consequently, although Guo is the only purine provided in the medium, theparasites are exposed to both Guo and Gua during growth. Under these conditions,intracellular purine levels would be modulated by expression of both the NT2 and NT3permeases.
For comparison, parasites were grown on the same medium except that 100 µM Xan wasemployed as the sole purine. In this medium, the Δnt2 null mutant did not upregulate Adouptake (Fig. 4B). Furthermore, complementation of both the Δnt2 and Δnt2Δnt3 nullmutants with the NT2 gene resulted in downregulation of Ado uptake when parasites were
Ortiz et al. Page 4
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

grown in Guo-replete medium (Fig. 4A) but not when they were grown with Xan as thepurine source (Fig 4B). These results indicate that deletion of the NT2 gene results inupregulation of Ado uptake only when ligands for NT2, such as Guo, are major sources ofpurines.
To investigate the effect of deleting the NT1 genes (both NT1.1 and NT1.2) on uptake ofpurines, the Δnt1 and the Δnt1Δnt3 null mutants were generated and characterized. Neitheruptake of Ino nor Ade was upregulated in the Δnt1 null mutant compared to wild typeparasites when promastigotes were grown on medium containing either 100 µM Xan or Ado(data not shown). Similarly, no increase in Ino or Ade uptake was noted for Δnt1Δnt3 nullmutants compared to Δnt3 null mutants for parasites grown on 100 µM Xan. Hence, deletionof the NT1 genes is not sufficient to induce the upregulation of purine transport observed foreither Δnt3 or Δnt2Δnt3 null mutants. It is possible that breakdown of Ado to Ade bynucleoside hydrolases released into the medium provides sufficient purine nucleobase tomitigate the effect of abrogating Ado uptake in these mutants.
Modulation of the level of NT1 mRNA in purine transporter null mutantsTo determine whether NT1 overexpression was the result of upregulation of NT1 mRNA,quantitative RT-PCR (qRT-PCR) was performed using primers specific for NT1 mRNA. Asshown in Table 1, there was no increase of NT1 mRNA in the Δnt3 null mutant compared towild type parasites, even though NT1 protein was upregulated at least 20-fold. In contrast,the Δnt2Δnt3 null mutant exhibited an increase in NT1 mRNA of ~3–4-fold compared towild type parasites. Furthermore, when this double null mutant was complemented with theNT3 gene, NT1 mRNA was reduced compared to the uncomplemented null mutant. Similarresults were obtained when NT1 mRNA was quantified employing Northern blots ratherthan qRT-PCR (Table 1). In summary, the NT1 protein is upregulated ~20-fold without acorresponding increase of NT1 mRNA in the Δnt3 null mutant, but the ~200-foldupregulation of NT1 protein in the Δnt2Δnt3 null mutant is accompanied by a small butreproducible increase of NT1 mRNA in this more heavily purine-restricted cell line.
Purine starvation induces upregulation of NT1 in wild type parasitesThe previous results indicated that deleting the NT2 and NT3 genes caused a pronouncedupregulation of NT1 protein and transport, likely due to purine restriction when thecorresponding purine permeases were not expressed. If this explanation is correct, it is likelythat wild type parasites exposed to purine starvation would also upregulate the NT1transporter. Hence, parasites were grown for 48 h in RPMI plus 20% dialyzed FBSsupplemented with 100 µM Ado, then washed and resuspended in fresh medium with orwithout 100 µM Ado, and allowed to grow for an additional 48 h (Fig. 5A,C). Under purinestarvation conditions (0 µM Ado), Ado uptake increased from 0–24 h reaching a maximum~60-fold increase by 24 h. When parasites starved for purines for 48 h were subsequentlytransferred into purine-replete medium (100 µM Ado), uptake of Ado was reduced ~47-foldwithin the first 24 h (Fig. 5B). Consistent with the uptake results, the level of NT1 proteinwas markedly increased in wild type parasites grown for 24 h under purine starvation (Fig.5C). These purine-starved parasites expressed a somewhat higher level of NT1 compared toΔnt3 parasites grown in 100 µM Xan. The increased level of NT1 was stronglydownregulated when parasites were grown in a purine-replete medium. Quantification ofNT1 mRNA by qRT-PCR indicated an upregulation of ~4-fold in cells exposed to purinestarvation versus cells grown in the presence of 100 µM Ado (Table 1). Hence, both purine-starved wild type parasites and Δnt2Δnt3 null mutants experience a modest upregulation ofNT1 mRNA but a considerably larger upregulation of NT1 protein.
Ortiz et al. Page 5
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

In order to understand whether purine-starved wild type cells were able to upregulate othertransporters, uptake of other purines was also evaluated (Fig. 5D). Following purinestarvation, uptake of Ino (a substrate for NT2) and Ade (a substrate for NT3 and NT4)increased ~15-fold and ~10-fold respectively. In contrast, Ado uptake was increased ~60-fold. These results confirm that parasites exert an adaptive mechanism that allows theparasite to salvage purines more efficiently by upregulating expression of purine transporterswhen their extracellular concentrations of purines decrease.
Upregulation of NT1 is due to an increased translation of NT1 mRNARegulation of NT1 protein by purines could be achieved either at the level of proteinsynthesis, i.e. translation, or protein turnover. To examine control at the level of proteinsynthesis, parasites were grown in the presence or absence of 100 µM Ado with or withoutthe translation inhibitor cycloheximide (Fig. 6A,B). Whereas both Ado uptake and NT1protein were strongly induced by purine starvation in the absence of cycloheximide, additionof the translation inhibitor completely abrogated this induction, suggesting that regulationoccurs at the level of translation of NT1 mRNA. Since cycloheximide globally inhibitsprotein synthesis and can result in artifacts that are due to secondary effects, synthesis ofNT1 protein was also measured directly by pulse labeling. Parasites were labeled with amixture of [35S]methionine and [35S]cysteine for 2 h, and membranes were isolated andemployed for immunoprecipitation using the anti-NT1 antibody. Labeled NT1 protein wasdetected in immunoprecipitates from Δnt2Δnt3 parasites and from wild type parasitesoverexpressing NT1 from an episomal expression vector, but no labeled NT1 was observedfor wild type parasites (Fig. 6C). A band of labeled protein of ~55 kDa, probablyrepresenting abundant tubulins that were non-specifically precipitated by the antibody, wasalso observed in immunoprecipitates and serves as a positive control for labeling. Similarly,when wild type parasites were pulse labeled under conditions of purine starvation, the NT1band was also detected (results not shown). These observations demonstrate directly thatsynthesis of NT1 is induced in the Δnt2Δnt3 null mutants and in purine starved parasites.
To determine whether purine limitation might control NT1 expression at the level of proteindegradation, protein synthesis in wild type and Δnt2Δnt3 null mutants was inhibited withcycloheximide and the level of Ado uptake was monitored as a function of time (Fig. 6D).Only a small difference in the rate of decay of Ado uptake was observed, and the Δnt2Δnt3parasites exhibited, if anything, a slightly more rapid rate of decay compared to wild typeparasites. Hence expression of the NT1 permease is not regulated at the level of proteinturnover.
DiscussionPurines are essential nutrients that Leishmania and other parasitic protozoa must scavengefrom their hosts. As such, these parasites have evolved mechanisms for maximizing theimport of purines when they are present in limiting concentrations. The landmark work ofGottlieb revealed that Crithidia fasciculata (Gottlieb, 1985) and C. luciliae (Alleman andGottlieb, 1996) upregulate expression of the purine salvage enzyme 3’-nucleotidase by asmuch as 1000-fold when parasites are starved for purines, and they also strongly enhanceexpression of purine nucleoside and nucleobase uptake activities (Cohn and Gottlieb, 1997).Related work by Gero and colleagues (Hall et al., 1996; Hall et al., 1998) confirmed thisupregulation in C. luciliae, and experiments employing Xenopus oocytes suggested that theenhancement of transport activity upon purine starvation was not accompanied by acorresponding increase in mRNAs encoding these transporters. Additional studies extendedupregulation by purine starvation to 3’-nucleotidase in L. donovani (Sacci et al., 1990) andto purine transporters in Trypanosoma brucei (de Koning et al., 2000). Hence, both transport
Ortiz et al. Page 6
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

and salvage enzymes appear to be regulated by extracellular purine levels in multiplekinetoplastid protozoa to maximize import of these nutrients under limiting conditions.
A previous study from our laboratory revealed that adenosine and uridine uptake, mediatedby NT1, were increased 8–10-fold in L. donovani parasites in which adenosine has beenremoved from the medium but xanthine was provided as a purine source to support growth(Seyfang and Landfear, 1999). In contrast, guanosine transport by NT2 was not affected byremoval of adenosine in a xanthine-replete background. These results imply that the purinetransporters also respond to the availability of their own specific substrates in a medium thatis not restricted overall for purines.
In the current study, molecular reagents including cloned purine transporter genes, nullmutants, and an antibody against the NT1 protein have been employed to investigate indetail the regulation of purine transporters by global purine limitation. Purine restriction wasapplied by knocking out genes for the purine nucleobase transporter NT3 and the purinenucleoside transporter NT2 and examining the response of the adenosine/pyrimidinenucleoside transporter NT1. The NT1 protein was strongly upregulated in the Δnt3 (~20-fold) and Δnt2Δnt3 (~200-fold) null mutants by the increasingly limited ability to importpurines. In contrast, the level of NT1 mRNA was not enhanced in the Δnt3 null mutant andwas only ~4-fold increased in the Δnt2Δnt3 null mutant, or in purine-starved wild typeparasites, revealing that regulation is exerted primarily at the level of protein abundance andnot mRNA accumulation. Experiments employing pulse labeling followed byimmunoprecipitation or blockade of protein synthesis with cycloheximide revealed thatcontrol of NT1 protein abundance operates at the level of mRNA translation and not proteinhalf-life. In the accompanying paper, similar conclusions have been reached by ourcolleagues examining the upregulation of purine transport and purine salvage enzymes in L.donovani following purine starvation of wild type parasites. Thus complementary studiesemploying distinct methods in two species of Leishmania both lead to the conclusion thatmultiple purine transporters are regulated by purine availability at the level of translation.
Kinetoplastid protozoa such as Leishmania and Trypanosoma exhibit very little regulation ofgene expression at the transcriptional level (Clayton and Shapira, 2007; Clayton, 2002).Examples of translational control have emerged from a variety of studies (Clayton andShapira, 2007), and sequences within the 5’- or 3’-untranslated regions of mRNAs havebeen implicated in translational control in several cases. Examination of potential regulatoryregions within the mRNAs of purine permeases is a compelling direction for future studiesand might ultimately lead to identification of regulatory proteins that interact with theseregions (Walrad et al., 2009). Another currently unanswered question concerns themechanism whereby parasites sense purines and ultimately respond by enhancing translationof mRNAs encoding purine transporters or other components of the salvage pathway.Kinetoplastid protozoa do not express G-protein coupled receptors or receptor tyrosinekinases (El-Sayed et al., 2005), thus excluding many of the signal transduction pathwaysthat eukaryotic organisms from yeast to human employ for sensing environmentalconditions. It is possible that global examination of alterations in protein expression or post-translational modifications induced by purine limitation could begin to illuminate thesensory apparatus employed to regulate expression of purine salvage components.
Experimental proceduresMaterials
Radiolabeled [2,8-3H]-adenine (50 Ci/mmol), [2,8-3H]-adenosine (36.8 Ci mmol−1),[2,8-3H]-inosine (39 Ci/mmol) and [5,6-3H]-uridine (32.3 Ci/mmol) were purchased from
Ortiz et al. Page 7
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Moravek Biochemicals, Inc. (Brea, CA). All other chemicals and regents were of the highestcommercial quality available.
Growth of parasitesL. major strain Friedlin VI (MHOM/IL/80Friedlin) parasites were cultured as described in(Ortiz et al., 2007). Briefly, parasites were propagated at 26°C in M199 containing 15% heatinactivated fetal bovine serum (FBS, Gibco® Invitrogen, Carlsbad, CA) supplemented with5 µg ml−1 of hemin and 100 µM Xan, unless is indicated.
Growth in defined purine mediumL. major parasites growing initially in M199 were washed 2X with PBS and resuspended atthe indicated density in RPMI (Sigma, Aldrich®, St. Louis, MO) supplemented with 20%dialyzed FBS, 5 µg ml−1 hemin, and either no purine or purine at the indicatedconcentration.
Nucleic acid purification, blotting and hybridizationFor isolation of genomic DNA and total RNA, DNAzol and TRIzol reagents (Invitrogen)were used according to manufacturer’s instructions. Southern and Northern blots wereperformed by standard methods (Sambrook et al., 1989).
Construction of vectorsTo generate the episomal expression constructs, individual open reading frames (ORFs)were amplified by PCR. The NT1.1 ORF was inserted into the EcoRI restriction site withinthe pX63NeoRI vector (Valdes et al., 2004) and the NT2 and NT3 ORFs were inserted intothe BamHI restriction site of the pXGBSD (Goyard and Beverley, 2000) expression vector.
Generation of null mutants by homologous gene replacementThe Δnt3 and Δnt4 null mutants have been reported previously (Ortiz et al., 2007). Forgeneration of single null mutants encompassing the Δnt1 and Δnt2 deletions, recombinanthomologous gene replacement constructs were prepared containing the appropriate drugselectable marker (puromycin resistance for NT1 and neomycin resistance for NT2) flankedby genomic DNA segments of 0.5–1 kb representing sequences upstream and downstreamof the relevant ORF. Linearized gene replacement constructs were gel purified andtransfected into wild type L. major promastigotes by electroporation (Ortiz et al., 2007), andafter 1 day parasites were plated onto M199 agar containing either puromycin (25 µg ml−1)or G418 (10–25 µg ml−1). Following growth at 26°C, colonies were picked and expanded inliquid medium, genomic DNA was prepared, and diagnostic PCRs were performed to ensurethat the intended integration had occurred. The resulting heterozygous knockout lines werethen plated onto agarose containing either the toxic adenosine analog tubercidin (10 µM) forthe Δnt1 null mutant or the toxic inosine analog formycin B (1–3 µM) for the Δnt2 nullmutant. Surviving colonies were tested by PCR and Southern blotting of genomic DNA toensure that they had undergone loss-of-heterozygosity (Gueiros-Filho and Beverley, 1994)to generate the null mutant. The double null mutants Δnt1Δnt3 and Δnt2Δnt3 were generatedin the same way, employing the Δnt3 null mutant as the starting cell line.
Uptake assaysIn L. major, uptake of [3H]-substrates was assayed as described in (Valdes et al., 2009),under the following conditions: Ade (25 µM, 5 µCi ml−1), Ado (1 µM, 5 Ci ml−1), Ino (1µM, 5 µCi ml−1), Urd (10 µM, 5 µCi ml−1). Results were analyzed using Prism 4.0bsoftware (GraphPad Software, Inc., La Jolla, CA)
Ortiz et al. Page 8
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Membrane preparationLeishmania parasites in early logarithmic phase (3–8 × 106 cells ml−1, 100 ml) werecollected by centrifugation at 2,000 rpm (835 × g) for 10 min at room temperature andwashed 2X with PBS. The pellet was resuspended in 500 µl of 0.1 M KH2PO4 pH 7.4, 10%glycerol, and 5 mM EDTA, pH 8 (Lysis Buffer) supplemented with protease inhibitorcocktail (Complete, Hoffman-LaRoche, Ldt, Switzerland). All further steps were conductedat 4°C. Cells maintained on ice were disrupted by sonication (Branson Sonifier 150, outputpower setting of 10, Branson Ultrasonics Corp., Danbury, CT) two times for 30 sec eachwith intervals of 1 min. The homogenates were centrifuged for 15 min at 14,000 × g in amicrocentrifuge to remove cell debris and were then treated with 75 mM NaHCO3 plusprotease inhibitors for 30 min on ice followed by another centrifugation. The supernatantswere subjected to ultracentrifugation at 60,000 rpm (128,000 × g) for 45 min at 4°C in aTL-100.2 rotor (Beckman Coulter Inc., Brea, CA). The pellet was dissolved in 50 µl of 2%of SDS in PBS supplemented with protease inhibitors. Protein quantification wasdetermined using the Bio-Rad DC Protein Assay kit (Hercules, CA).
Immunoblot analysisTo determine the level of expression of NT1 protein in Leishmania parasites, 20 µg of cellmembrane were mixed with an equal volume of 2X Laemmli sample buffer and heated at70°C for 10 min. Samples were resolved by electrophoresis on 10% gradient NuPAGE®
Novex Bis-Tris gels (Invitrogen) under reducing conditions using the XCell SureLock™Mini-Cell system (Invitrogen). Subsequently, proteins were electro-transferred underdenaturing conditions onto nitrocellulose membranes (Protran, Whatman, GmbH, Germany)using the XCell II™ Blot Module (Invitrogen). Detection of NT1 by immunoblot wasperformed using the Odyssey® Infrared Imaging System (LI-COR® Biosciences. Lincoln,NE). Briefly, membrane was incubated with 8–10 ml LI-COR blocking buffer for 1 h atroom temperature with gentle agitation. Subsequently, the membrane was incubatedsimultaneously with rabbit polyclonal anti Leishmania mexicana NT1-loop VII (Gln236-Lys331) antibody (Detke, 2007) and mouse anti α-tubulin (Calbiochem-Novabiochem, SanDiego, CA) antibodies (dilutions 1:3,000 and 1:5,000, respectively) in 6–8 ml of LI-CORblocking buffer overnight at 4°C. The next day, the antibody solution was removed and themembrane was washed four times for 10 min each with 15 ml PBS with 0.1% Tween (PBS-T) before addition of secondary antibody conjugated to fluorescent dyes, IRDye® 800CWGoat Anti Rabbit IgG and IRDye® 680 Goat Anti Mouse IgG (dilution 1:20,000, Odyssey,LI-COR), in 6–8 ml LI-COR blocking buffer with gentle agitation for 45 min at roomtemperature. At the end of the incubation period, the membrane was washed as abovefollowed by one wash with 15 ml PBS only. The membrane was analyzed using theOdyssey® Infrared Imaging System, and fluorescence intensity was determined using LI-COR imaging software followed by exporting to Microsoft Excel for graphicalrepresentation as mean ± standard error.
Metabolic labeling and immunoprecipitationLeishmania parasites at early logarithmic phase (3–8 × 106 cells ml−1) were collected bycentrifugation at 835 × g for 10 min at room temperature, washed twice with sterile PBS,and resuspended at 4 × 107 cells ml−1 in RPMI Met-Cys-free medium (Sigma Aldrich)containing 5% dialyzed FBS (Invitrogen). 100 µCi of tran[35S]label (43.5 TBq/mmol, MP)were added per ml of cells and incubated at 26°C for 2 h. After labeling, 1 ml of cells (4 ×107) were transferred into an Eppendorf tube, washed twice with PBS, and resuspended in500 µl of cold Lysis Buffer supplemented with Halt Protease Inhibitor Cocktail (ThermoFisher Scientific, Walthan, MA). Subsequently, 500 µl of cold glass beads (200–500microns, Sigma Aldrich) were added and vortexed for 20 min at 5 min intervals. Betweenintervals cells were kept in ice for 2 min. Lysed cells were pelleted in a microcentrifuge at
Ortiz et al. Page 9
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

17,000 × g at 4°C for 15 min to separate beads and unbroken cells from lysates.Supernatants were then ultracentrifuged as described above to isolate [35S]-labeled totalmembrane proteins, and these preparations were resuspended with 50 µl of RIPA buffer(Sambrook et al., 1989) and incubated at 4°C for 1 h. Rabbit polyclonal anti-NT1 antibody(1:25 dilution) (Detke, 2007) was added to this sample and incubated overnight at 4°C withgentle mixing followed by addition of 50 µl protein-A sepharose (GE, Piscataway, NJ) andincubation for 1 h at 4°C with gentle mixing. Sepharose bead-immune complexes werecollected by centrifugation at 17,000 × g for 5 min at 4°C and washed twice with 0.5%Triton X-100 in PBS containing protease inhibitors. Beads were then washed twice withPBS containing protease inhibitors and finally resuspended in 50 µl of protein loadingsample buffer containing 5 mM DTT, heated at 70°C for 10 min, and loaded onto a 4–12%NUPAGE gel (Invitrogen) followed by electrophoresis for 1 h at 200 V. The gel was soakedin 30% methanol-5% glycerol for 30 min at room temperature and dried. [35S]-immunoprecipitated proteins were detected by exposure to phosphor-imaging screen thatwas developed using a PhosphorImager SI (Molecular Dynamics, Sunnyvale, CA).
Quantitative real-time PCR (qRT-PCR)Quantitative real-time PCR was performed as detailed previously (Ortiz et al., 2009b).NT1.1 primers were designed from the amino terminus and the mRNA spliced leadersequence using Applied Biosystems PRIMER EXPRESS 2.0 Software and were:CAGCTCTGCGCTACTGCCGCACAC (forward) and GATCGGATGCGGTGTCCATCTC(reverse), producing a 111 bp amplification product, while primers for α-tubulin were:TGTTGAGGATGACGCGTTCA (forward) and GCGGAACGTGCTTGCC (reverse),producing a 64 bp amplification product. Primers for amplification of α-tubulin RNA wereused as internal controls. Relative changes in target amplicons were determined by the Pfafflmethod (Pfaffl, 2001).
AcknowledgmentsWe dedicate this paper to the memory of Dr. Siegfried Detke, a valued colleague without whom this work wouldnot have been possible. We thank our colleagues in the Ullman laboratory for extensive discussions concerningtheir related studies of purine starvation in L. donovani that appear in the accompanying paper. This work wassupported by Grant AI044138 from the National Institutes of Health.
ReferencesAlleman MM, Gottlieb M. Enhanced acquisition of purine nucleosides and nucleobases by purine-
starved Crithidia luciliae. Mol. Biochem. Parasitol. 1996; 76:279–287. [PubMed: 8920013]Carter NS, Drew ME, Sanchez M, Vasudevan G, Landfear SM, Ullman B. Cloning of a novel inosine-
guanosine transporter gene from Leishmania donovani by functional rescue of a transport-deficientmutant. J. Biol. Chem. 2000; 275:20935–20941. [PubMed: 10783393]
Carter NS, Yates P, Arendt CS, Boitz JM, Ullman B. Purine and pyrimidine metabolism inLeishmania. Adv Exp Med Biol. 2008; 625:141–154. [PubMed: 18365665]
Clayton C, Shapira M. Post-transcriptional regulation of gene expression in trypanosomes andleishmanias. Mol Biochem Parasitol. 2007; 156:93–101. [PubMed: 17765983]
Clayton CE. Life without transcriptional control? From fly to man and back again. Embo J. 2002;21:1881–1888. [PubMed: 11953307]
Cohn CS, Gottlieb M. The acquisition of purines by trypanosomatids. Parasitol. Today. 1997; 13:231–235. [PubMed: 15275076]
de Koning HP, Watson CJ, Sutcliffe L, Jarvis SM. Differential regulation of nucleoside andnucleobase transporters in Crithidia fasciculata and Trypanosoma brucei brucei. Mol BiochemParasitol. 2000; 106:93–107. [PubMed: 10743614]
Ortiz et al. Page 10
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Detke S. TOR-induced resistance to toxic adenosine analogs in Leishmania brought about by theinternalization and degradation of the adenosine permease. Exp Cell Res. 2007; 313:1963–1978.[PubMed: 17428463]
El-Sayed NM, Myler PJ, Bartholomeu DC, Nilsson D, Aggarwal G, Tran AN, Ghedin E, Worthey EA,Delcher AL, Blandin G, Westenberger SJ, Caler E, Cerqueira GC, Branche C, Haas B, Anupama A,Arner E, Aslund L, Attipoe P, Bontempi E, Bringaud F, Burton P, Cadag E, Campbell DA,Carrington M, Crabtree J, Darban H, da Silveira JF, de Jong P, Edwards K, Englund PT, FazelinaG, Feldblyum T, Ferella M, Frasch AC, Gull K, Horn D, Hou L, Huang Y, Kindlund E, KlingbeilM, Kluge S, Koo H, Lacerda D, Levin MJ, Lorenzi H, Louie T, Machado CR, McCulloch R,McKenna A, Mizuno Y, Mottram JC, Nelson S, Ochaya S, Osoegawa K, Pai G, Parsons M, PentonyM, Pettersson U, Pop M, Ramirez JL, Rinta J, Robertson L, Salzberg SL, Sanchez DO, Seyler A,Sharma R, Shetty J, Simpson AJ, Sisk E, Tammi MT, Tarleton R, Teixeira S, Van Aken S, Vogt C,Ward PN, Wickstead B, Wortman J, White O, Fraser CM, Stuart KD, Andersson B. The genomesequence of Trypanosoma cruzi, etiologic agent of Chagas disease. Science. 2005; 309:409–415.[PubMed: 16020725]
Gottlieb M. Enzyme regulation in a trypanosomatid: Effect of purine starvation on levels of 3'-nucleotidase activity. Science. 1985; 227:72–74. [PubMed: 2981117]
Goyard S, Beverley SM. Blasticidin resistance: a new independent marker for stable transfection ofLeishmania. Mol Biochem Parasitol. 2000; 108:249–252. [PubMed: 10838227]
Gueiros-Filho FJ, Beverley SM. Loss-of-heterozygosity in Leishmania major: DHFR-TS as a negativemarker. Woods Hole Molecular Parasitology Meeting Abstracts. 1994; 5:133C.
Hall ST, Hillier CJ, Gero AM. Crithidia luciliae: Regulation of purine nucleoside transport byextracellular purine concentrations. Exp. Parasitol. 1996; 83:314–321. [PubMed: 8823248]
Hall ST, JI P, Gero AM, Krishna S. Crithidia luciliae: functional expression of nucleoside andnucleobase transporters in Xenopus laevis oocytes. Exp. Parasitol. 1998; 90:181–188. [PubMed:9769248]
Hammond DJ, Gutteridge WE. Purine and pyrimidine metabolism in the Trypanosomatidae. MolBiochem Parasitol. 1984; 13:243–261. [PubMed: 6396514]
Hassan HF, Coombs GH. A comparative study of the purine- and pyrimidine-metabolising enzymes ofa range of trypanosomatids. Comp Biochem Physiol B. 1986; 84:219–223. [PubMed: 3015485]
Landfear SM, Ullman B, Carter N, Sanchez M. Nucleoside and nucleobase transporters in parasiticprotozoa. Eukaryotic Cell. 2004; 3:245–254. [PubMed: 15075255]
Ortiz D, Sanchez MA, Pierce S, Herrmann T, Kimblin N, Bouwer HG, Landfear SM. Moleculargenetic analysis of purine nucleobase transport in Leishmania major. Mol. Microbiol. 2007;64:1228–1243. [PubMed: 17542917]
Ortiz D, Sanchez MA, Koch HP, Larsson HP, Landfear SM. An acid-activated nucleobase transporterfrom Leishmania major. J Biol Chem. 2009a; 284:16164–16169. [PubMed: 19366701]
Ortiz D, Sanchez MA, Quecke P, Landfear SM. Two novel nucleobase/pentamidine transporters fromTrypanosoma brucei. Mol Biochem Parasitol. 2009b; 163:67–76. [PubMed: 18992774]
Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic AcidsRes. 2001; 29:e45. [PubMed: 11328886]
Sacci JB Jr, Campbell TA, Gottlieb M. Leishmania donovani: regulated changes in the level ofexpression of the surface 3'-nucleotidase/nuclease. Exp Parasitol. 1990; 71:158–168. [PubMed:2164952]
Sambrook, J.; Fritsch, EF.; Maniatis, T. Molecular Cloning: A Laboratory Manual. Cold SpringHarbor, N.Y.: Cold Spring Harbor Laboratory; 1989.
Seyfang A, Landfear SM. Substrate depletion upregulates uptake of myo-inositol, glucose andadenosine in Leishmania. Mol. Biochem. Parasitol. 1999; 104:121–130. [PubMed: 10589986]
Shi W, Schramm VL, Almo SC. Nucleoside hydrolase from Leishmania major. Cloning, expression,catalytic properties, transition state inhibitors, and the 2.5-a crystal structure. J Biol Chem. 1999;274:21114–21120. [PubMed: 10409664]
Stein A, Vasudevan G, Carter N, Ullman B, Landfear SM, Kavanaugh MP. Equilibrative nucleosidetransporter family members from Leishmania donovani are electrogenic proton symporters. J.Biol. Chem. 2003; 278:35127–35134. [PubMed: 12835315]
Ortiz et al. Page 11
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Valdes R, Vasudevan G, Conklin D, Landfear SM. Transmembrane domain 5 of the LdNT1.1nucleoside transporter is an amphipathic helix that forms part of the nucleoside translocationpathway. Biochemistry. 2004; 43:6793–6802. [PubMed: 15157113]
Valdes RM, Arastu-Kapur SA, Landfear SM, Shinde U. An Ab initio structural model of a nucleosidepermease predicts functionally important residues. J Biol Chem. 2009; 284:19067–19076.[PubMed: 19429678]
Vasudevan G, Carter NS, Drew ME, Beverley SM, Sanchez MA, Seyfang A, Ullman B, Landfear SM.Cloning of Leishmania nucleoside transporter genes by rescue of a transport-deficient mutant.Proc. Natl. Acad. Sci. U.S.A. 1998; 95:9873–9878. [PubMed: 9707568]
Walrad P, Paterou A, Acosta-Serrano A, Matthews KR. Differential trypanosome surface coatregulation by a CCCH protein that co-associates with procyclin mRNA cis-elements. PLoSPathog. 2009; 5:e1000317. [PubMed: 19247446]
Ortiz et al. Page 12
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 1.Upregulation of Ado and Urd uptake and of the NT1 adenosine/pyrimidine nucleosidetransporter in Δnt3 and Δnt2Δnt3 purine transporter null mutants. A. Uptake of 1 µM[3H]Ado and 10 µM [3H]Urd was measured (1 min time point), and the rates of uptake wereplotted for wild type parasites (WT), the Δnt3 and Δnt2Δnt3 purine transporter null mutants,and the null mutants complemented with the NT3 purine nucleobase transporter genemaintained on an episomal expression vector ([pNT3]). Uptake data in this and subsequentfigures represent the average and standard deviations of 3 replicate measurements. B.Immunoblot of membrane fractions (20 µg protein) probed with the anti-NT1 antibody(NT1) and anti-α-tubulin antibody (Tubulin). The left panel displays an immunoblot
Ortiz et al. Page 13
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

employing WT and WT[pNT1] cells to reveal the position of the ~43–45 kDa bandsrepresenting NT1, and the right panel displays an immunoblot of the same cell linesanalyzed in part A. The numbers at the left (43 and 55) represent the positions of molecularweight markers, designated in kilodaltons.
Ortiz et al. Page 14
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 2.Quantification of upregulation of the NT1 permease in the Δnt3 and Δnt2Δnt3 null mutants.Membrane fractions from wild type (20 µg), Δnt3 (A) and Δnt2Δnt3 (B) null mutants (0.1–20 µg) were separated by SDS-PAGE and analyzed by immunoblotting as in Fig. 1B.
Ortiz et al. Page 15
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 3.Effect on Ado uptake and NT1 expression of supplementing the growth medium with Guoor Xan. A. The Δnt3 null mutant grown in M199 medium supplemented with 100 µM Xanwere pelleted, washed, and transferred to fresh medium containing either 100 µM Guo orXan, and uptake of 1 µM Ado was measured over a 24 h time course. B. Experiment as inpart A, except that membrane fractions (20 µg) were monitored by immunoblotting. C. Wildtype parasites and the Δnt2Δnt3 null mutant were grown for 24 h in medium supplementedwith 100 µM Guo or Xan, and uptake of Ado was measured. D. Lysates from parasites inpart C were analyzed by immunoblotting.
Ortiz et al. Page 16
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 4.Effect on Ado uptake of growing the Δnt2 and Δnt2Δnt3 null mutants in mediumsupplemented with 100 µM Guo or Xan. Parasites were grown in medium containing 20%dialyzed FBS and supplemented with 100 µM Guo (A) or Xan (B) for 5 days, and uptake of1µM [3H]Ado was measured.
Ortiz et al. Page 17
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
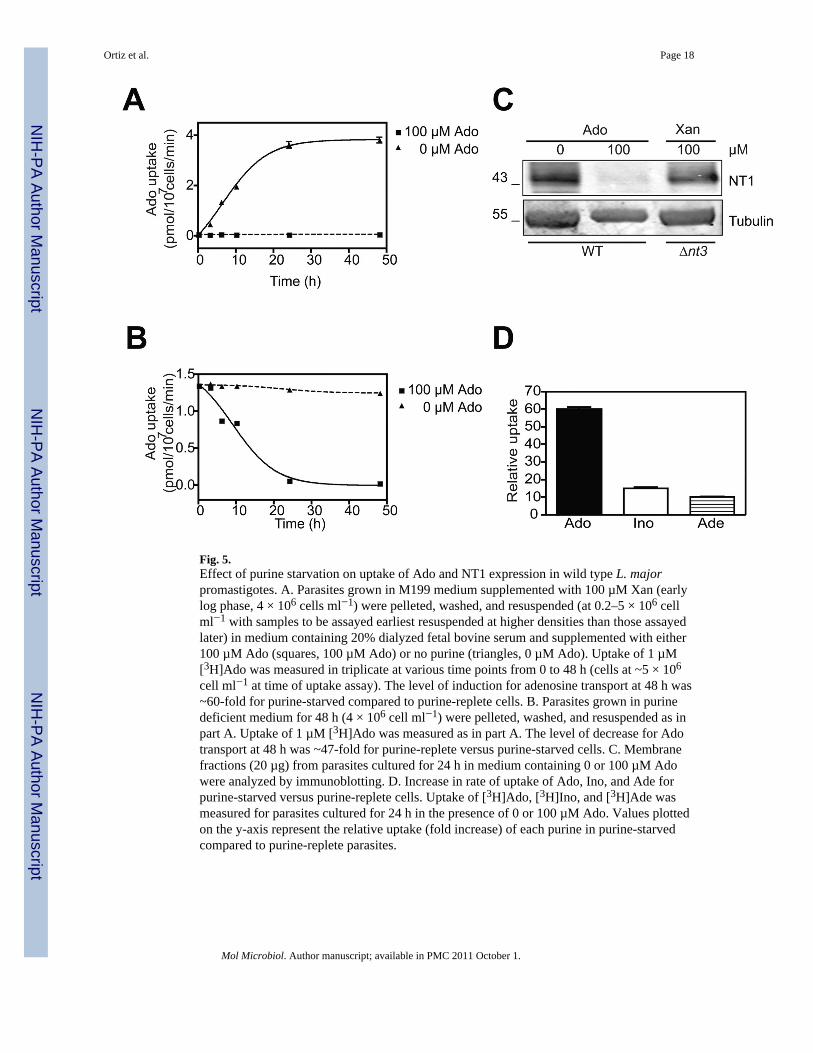
Fig. 5.Effect of purine starvation on uptake of Ado and NT1 expression in wild type L. majorpromastigotes. A. Parasites grown in M199 medium supplemented with 100 µM Xan (earlylog phase, 4 × 106 cells ml−1) were pelleted, washed, and resuspended (at 0.2–5 × 106 cellml−1 with samples to be assayed earliest resuspended at higher densities than those assayedlater) in medium containing 20% dialyzed fetal bovine serum and supplemented with either100 µM Ado (squares, 100 µM Ado) or no purine (triangles, 0 µM Ado). Uptake of 1 µM[3H]Ado was measured in triplicate at various time points from 0 to 48 h (cells at ~5 × 106
cell ml−1 at time of uptake assay). The level of induction for adenosine transport at 48 h was~60-fold for purine-starved compared to purine-replete cells. B. Parasites grown in purinedeficient medium for 48 h (4 × 106 cell ml−1) were pelleted, washed, and resuspended as inpart A. Uptake of 1 µM [3H]Ado was measured as in part A. The level of decrease for Adotransport at 48 h was ~47-fold for purine-replete versus purine-starved cells. C. Membranefractions (20 µg) from parasites cultured for 24 h in medium containing 0 or 100 µM Adowere analyzed by immunoblotting. D. Increase in rate of uptake of Ado, Ino, and Ade forpurine-starved versus purine-replete cells. Uptake of [3H]Ado, [3H]Ino, and [3H]Ade wasmeasured for parasites cultured for 24 h in the presence of 0 or 100 µM Ado. Values plottedon the y-axis represent the relative uptake (fold increase) of each purine in purine-starvedcompared to purine-replete parasites.
Ortiz et al. Page 18
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig. 6.Translational control of NT1 expression by purine limitation. A. Inhibition of translationwith cycloheximide (CHX) prevents upregulation of Ado uptake in purine-starved parasites.Promastigotes were incubated for 24 h in medium containing either 0 or 100 µM Ado in theabsence (−) or presence (+) of 10 µg ml−1 cycloheximide. Uptake of 1 µM [3H]Ado wasmeasured as in Fig. 1. B. Cycloheximide inhibits upregulation of the NT1 protein by purinestarvation. Parasites cultured as in part A were analyzed by immunoblotting. C. Pulselabeling demonstrates that biosynthesis of NT1 protein is induced in Δnt2Δnt3 parasites.Wild type (WT) promastigotes, parasites overexpressing the NT1 protein from an episomalexpression vector ([pNT1]), and Δnt2Δnt3 parasites were labeled with a mixture of[35S]methionine and [35S]cysteine for 2 h followed by immunoprecipitation with anti-NT1antibody. The band at ~43 kDa represents newly synthesized NT1 protein, and the band at~55 kD is a non-specifically precipitated protein, probably abundantly expressed tubulins.D. Turnover of adenosine transporters for wild type and Δnt2Δnt3 null mutants. Parasiteswere incubated in medium containing 10 µg ml−1 cycloheximide to inhibit protein synthesis,and the decay of Ado transport activity was measured by quantifying uptake of 1 µM[3H]Ado from 0–75 h after addition of drug. For each cell line, the level of uptake at t = 0was set to a value of 1.0, and the relative uptake values for later time points were normalizedto this value. The absolute uptake value at t = 0 was 3.58 ± 0.078 pmol min−1 per 107 cellsfor wild type and 115 ± 1.8 pmol min−1 per 107 cells for Δnt2Δnt3 cells (triplicatemeasurements).
Ortiz et al. Page 19
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Ortiz et al. Page 20
Tabl
e 1
Rel
ativ
e le
vels
of N
T1 m
RN
A in
wild
type
and
nul
l mut
ant p
aras
ites.
For s
ampl
es fr
om w
ild ty
pe a
nd m
utan
t par
asite
s in
the
left
pane
l of t
he ta
ble,
prom
astig
otes
wer
e gr
own
in M
199
med
ium
plu
s 15%
FB
S w
ith 1
00 µ
M X
an. F
or th
e w
ild ty
pe sa
mpl
es sh
own
in th
e rig
ht p
anel
, pro
mas
tigot
es w
ere
grow
n in
RPM
I plu
s 20%
dia
lyze
d FB
S w
ith (1
00 µ
M) o
r with
out (
0 µM
) Ado
. Tot
al R
NA
was
isol
ated
from
eac
h sa
mpl
e, a
nd th
e am
ount
of N
T1m
RN
A w
as d
eter
min
ed b
y qR
T-PC
R o
r Nor
ther
n bl
ot. T
he N
T1 m
RN
A w
as n
orm
aliz
ed to
the α-
tubu
lin m
RN
A in
eac
h sa
mpl
e. F
or e
ach
set o
fex
perim
ents
(lef
t or r
ight
pan
els)
, the
leve
l of N
T1 m
RN
A in
wild
type
cel
ls g
row
n in
pur
ine-
repl
ete
med
ium
was
des
igna
ted
as a
val
ue o
f 1.0
, and
the
amou
nt o
f NT1
mR
NA
in th
e ot
her s
ampl
es w
as c
alcu
late
d re
lativ
e to
that
val
ue. N
orth
ern
blot
s wer
e ex
pose
d to
a p
hosp
horim
ager
to q
uant
ify th
ein
tens
ity o
f the
NT1
and
α-tu
bulin
mR
NA
sign
als.
ND
indi
cate
s not
det
erm
ined
. For
eac
h ex
perim
ents
, tw
o bi
olog
ical
repl
icat
es w
ere
prep
ared
for e
ach
RN
A, a
nd th
e re
lativ
e N
T1 m
RN
A le
vel w
as d
eter
min
ed in
trip
licat
e fo
r eac
h R
NA
sam
ple.
The
mea
n re
lativ
e va
lue
of N
T1 m
RN
A w
as c
alcu
late
d fo
rea
ch o
f the
se tw
o R
NA
sam
ples
(n =
2),
and
the
mea
n an
d ra
nge
of v
alue
s for
thes
e tw
o sa
mpl
es a
re re
porte
d in
this
tabl
e.
Met
hod
WT
Δnt3
Δnt3
[pN
T3]
Δnt2Δn
t3Δn
t2Δn
t3[p
NT3
]W
T(1
00 µ
M)
WT
(0 µ
M)
qRT-
PCR
(n =
2)
1.0
0.85
± 0
.21
1.1
± 0.
713.
4 ±
2.0
1.7
± 0.
771.
03.
5 ±
1.4
Nor
ther
nbl
ot(n
= 2
)
1.0
0.97
± 0
.37
1.5
± 0.
843.
3 ±
1.1
1.7
± 0.
49N
DN
D
Mol Microbiol. Author manuscript; available in PMC 2011 October 1.
Copyright © 2022 FDOKUMEN




















