
NTP-CERHR Expert Panel Report on the reproductive and developmental toxicity of soy formula

D
KKAa
b
c
d
U
a
ARR2AA
KPCDG
Prc
Prntf
Af
h0
Reproductive Toxicology 51 (2015) 133–144
Contents lists available at ScienceDirect
Reproductive Toxicology
j ourna l h o mepa ge: www.elsev ier .com/ locate / reprotox
evelopmental toxicity of perfluorononanoic acid in mice�
aberi P. Dasa, Brian E. Greya, Mitchell B. Rosenb, Carmen R. Wooda,atoria R. Tatum-Gibbsa, R. Daniel Zehrc, Mark J. Strynard,ndrew B. Lindstromd, Christopher Laua,∗
Toxicity Assessment DivisionIntegrated Systems Toxicology DivisionResearch Core Unit, National Health and Environmental Effects Research LaboratoryHuman Exposure and Atmospheric Sciences Division, National Exposure Research Laboratory, Office of Research and Development,.S. Environmental Protection Agency, Research Triangle Park, North Carolina 27711, United States
r t i c l e i n f o
rticle history:eceived 27 September 2014eceived in revised form2 November 2014ccepted 16 December 2014vailable online 25 December 2014
eywords:erfluorononanoic acidD-1 miceevelopmental toxicityene expression
a b s t r a c t
Perfluorononanoic acid (PFNA) is a ubiquitous and persistent environmental contaminant. Although itslevels in the environment and in humans are lower than those of perfluorooctane sulfonate (PFOS) orperfluorooctanoic acid (PFOA), a steady trend of increases in the general population in recent years hasdrawn considerable interest and concern. Previous studies with PFOS and PFOA have indicated develop-mental toxicity in laboratory rodent models. The current study extends the evaluation of these adverseoutcomes to PFNA in mice. PFNA was given to timed-pregnant CD-1 mice by oral gavage daily on ges-tational day 1–17 at 1, 3, 5 or 10 mg/kg; controls received water vehicle. Dams given 10 mg/kg PFNAcould not carry their pregnancy successfully and effects of this dose group were not followed. Similar toPFOS and PFOA, PFNA at 5 mg/kg or lower doses produced hepatomegaly in the pregnant dams, but didnot affect the number of implantations, fetal viability, or fetal weight. Mouse pups were born alive andpostnatal survival in the 1 and 3 mg/kg PFNA groups was not different from that in controls. In contrast,although most of the pups were also born alive in the 5 mg/kg PFNA group, 80% of these neonates diedin the first 10 days of life. The pattern of PFNA-induced neonatal death differed somewhat from thoseelicited by PFOS or PFOA. A majority of the PFNA-exposed pups survived a few days longer after birththan those exposed to PFOS or PFOA, which typically died within the first 2 days of postnatal life. Surviv-ing neonates exposed to PFNA exhibited dose-dependent delays in eye opening and onset of puberty. Inaddition, increased liver weight seen in PFNA-exposed offspring persisted into adulthood and was likely
related to the persistence of the chemical in the tissue. Evaluation of gene expression in fetal and neona-tal livers revealed robust activation of peroxisome proliferator-activated receptor-alpha (PPAR�) targetgenes by PFNA that resembled the responses of PFOA. Our results indicate that developmental toxicity ofPFNA in mice is comparable to that of PFOS and PFOA, and that these adverse effects are likely commonto perfluoroalkyl acids that peAbbreviations: PFAA, perfluoroalkyl acid; PFOS, perfluorooctane sulfonate;FOA, perfluorooctanoic acid; PFNA, perfluorononanoic acid; PFBA, perfluo-obutanoic acid; PPAR�, peroxisome proliferator activated receptor-alpha; CAR,onstitutive androstane receptor; PXR, pregnane X receptor; PND, postnatal day.� The information in this document has been funded by the U.S. Environmentalrotection Agency. It has been subjected to review by the National Health and Envi-onmental Effects Research Laboratory and approved for publication. Approval doesot signify that the contents reflect the views of the Agency, nor does mention ofrade names or commercial products constitute endorsement or recommendationor use.∗ Corresponding author at: Mail Drop B105-04, U.S. Environmental Protectiongency, Research Triangle Park, NC 27711, United States. Tel.: +1 919 541 5097;
ax: +1 919 541 4849.E-mail address: [email protected] (C. Lau).
ttp://dx.doi.org/10.1016/j.reprotox.2014.12.012890-6238/Published by Elsevier Inc.
rsist in the body.Published by Elsevier Inc.
1. Introduction
Perfluoroalkyl acids (PFAAs) and their derivatives are syn-thetic materials that have unique physico-chemical characteristicsof low surface free energy and surface active properties, whichmake them highly suitable for a wide range of industrialprocesses and consumer applications [1]. These may includemetal electroplating, emulsifiers for polymer production, fire-fighting foams, water- and stain-resistant coatings for textilesand carpets, and grease-proof paper products for food packag-
ing. These chemicals are non-biodegradable in the environment,many of which are ubiquitously distributed in all environmen-tal media, and detected worldwide in humans and wildlife[2–7].
1 e Toxi
c[tpirbsatmtrsbPastbtwst
ictlSPfd(vrr
P[dii[rraPibeprolvidmankmmd
34 K.P. Das et al. / Reproductiv
Over the past decade, a preponderance of evidence has indi-ated PFAA toxicities in laboratory animals and wildlife species4,8–10]; although a majority of studies has focused on thewo eight-carbon PFAAs, perfluorooctane sulfonate (PFOS) anderfluorooctanoate (PFOA). These include hepatotoxicity, tumor
nduction, developmental toxicity, immunotoxicity, endocrine dis-uption, and neurotoxicity. In humans, epidemiological findingsased on cross-sectional designs are equivocal; whereas sometudies have suggested associations between PFAA exposure anddverse effects on reproductive health, birth weight and puber-al and behavioral development [11–18], lipid and hormone
etabolism [19–22] and immune responses [23,24], others failedo detect such linkages [25–31]. The mechanisms of PFAA toxicityemain largely to be explored, although involvement of peroxi-ome proliferator-activated receptor-alpha (PPAR�) pathway haseen widely implicated [4,10,32]. The reported adverse effects ofFAAs in laboratory studies vary among chemicals with differentlkyl chain lengths (from 4 to14 carbons) and functional groups (e.g.ulfonates and carboxylates). Such variations are largely driven byhe pharmacokinetic (different rates of elimination from the bodyurden) and pharmacodynamic (different potencies at the targetissues) properties of an individual PFAA [10]. In general, PFAAsith longer alkyl chain lengths (>6 carbons) have much longer
erum half-lives and are much more potent in their actions thanhose with shorter chains.
Because of environmental concerns, production of PFOS ceasedn 2002 and emission of PFOA to the environment has been signifi-antly curtailed by the major manufacturers in the U.S. Since then, arend toward reduction of these chemicals in the U.S. general popu-ation was noted in the National Health and Nutrition Examinationurvey (NHANES) conducted by the Centers for Disease Control andrevention [5]. Indeed, the levels of PFOS and perfluorohexane sul-onate (PFHxS) have fallen by about half, and those of PFOA haveropped by 20%. In contrast, the levels of perfluorononanoic acidPFNA), though lowest among the four detectable PFAAs in this sur-ey, have been on a steady rise, tripling during the past decade. Theeason behind this unexpected rise is unclear, but the observationaises considerable concern.
Compared to PFOS and PFOA, the potential adverse effects ofFNA have not been vigorously investigated. Tatum-Gibbs et al.33] have shown that serum half-life of PFNA in the mouse (26–68ays) is much longer than that of PFOA (17–19 days) [34]. A recent
n vitro study using transfected COS-1 cells demonstrated that PFNAs more potent than PFOA for activating PPAR� [35]. Kinney et al.36] reported moderate acute inhalation toxicity of PFNA in maleats. Fang et al. [37–39] described the hepatotoxicity of PFNA inat; while Zhang et al. [40,41] further elaborated the transcriptionalnd proteomic changes in zebrafish liver after chronic exposure toFNA. Liu et al. [42] reported the thyroid-disrupting effects of PFNAn zebrafish. Immunotoxic effects of PFNA in rats and mice haveeen shown by Fang et al. [43,44] and Rockwell et al. [45]. Fengt al. [46,47] described PFNA-induced functional changes and apo-tosis in rat testes. Stump et al. [48] conducted a two-generationeproductive toxicity study in rats with S-111-S-WB, a mixturef perfluoro fatty acids that include PFNA (∼74%) and reported aack of effect on reproductive performance, litter size and pup sur-ival, although lower body weights and enlarged liver were notedn the exposed offspring. In contrast, Wolf et al. [49] evaluated theevelopmental effects of PFNA (at 0.8–2.0 mg/kg) with 129S1/SvlmJice and reported dose-dependent reductions of live pups at birth
nd survival of offspring to weaning; these adverse effects wereot observed in transgenic mice whose PPAR� was functionally
nocked out, suggesting the involvement of this nuclear receptor inediating the PFNA-induced developmental toxicity. In view of thearked persistence of PFNA, its relative high potency, and adverseevelopmental effects, the current study extends the findings from
cology 51 (2015) 133–144
Wolf et al. [49] to another mouse strain, CD-1 that is commonly usedin toxicological evaluations, and provides a full characterization ofPFNA developmental toxicity in this species.
2. Materials and methods
2.1. Chemicals
Perfluorononanoic acid (PFNA, free acid, 97% pure, primarily lin-ear isomer according to supplier) and other chemicals used in thisstudy were purchased from Sigma–Aldrich (St. Louis, MO) unlessindicated otherwise. All other chemicals and reagents used for thisstudy were of analytical grade purchased from VWR (West Chester,PA). Stable isotope 1,2-13C9-labeled PFNA (13C9F17O2H, 99% pure)was supplied as free acid in methanol (50 �g/ml) by Cambridge Iso-tope Laboratories (Andover, MA) and used as internal standard forliquid chromatography/tandem mass spectrometric (LC–MS/MS)analysis of PFNA. For all studies, PFNA was dissolved in deionizedwater at 60 ◦C with sonication, and prepared fresh daily.
2.2. Animals
All animal studies were conducted in accordance with the Insti-tutional Animal Care and Use Committee guidelines established bythe U.S. Environmental Protection Agency’s Office of Research andDevelopment/National Health and Environmental Effects ResearchLaboratory. Procedures and facilities were consistent with the rec-ommendations of the 1996 National Research Council’s “Guide forthe Care and Use of Laboratory Animals,” the Animal Welfare Act,and Public Health Service Policy on the Humane Care and Use ofLaboratory Animals. Animal facilities were controlled for tempera-ture (20–24 ◦C), relative humidity (40–60%) and kept under a 12-hlight–dark cycle.
Timed-pregnant CD-1 mice were obtained from Charles RiverLaboratories (Raleigh, NC), where females were bred overnight,and the sperm-positive females, considered to be at gestationalday (GD) 0, were shipped to our facility on the same day. Uponarrival, mice were housed individually in polypropylene cages withAlpha-dri (Shepherd Specialty Papers, Kalamazoo, MI) bedding andprovided pellet feed (LabDiet 5008, PMI Nutrition International LLC,Brentwood, MO) and tap water ad libitum.
2.3. Treatment of animals
The study was performed in three separate blocks, with approx-imately 40 mice per block. Upon arrival at the animal facility, micewere weighed and randomly assigned to five treatment groups:vehicle control received deionized water, and other groups received1, 3, 5, or 10 mg PFNA/kg body weight per day on GD 1–17. Micewere weighed and dosed by oral gavage at a volume of 10 ml/kgbody weight; pH of PFNA dosing solution was approximately 3.2for 1 mg/ml (highest dose) and 3.4 for 0.5 mg/ml, compared to a pHof 5.4 for vehicle (deionized water). These dose levels were cho-sen to approximate the effective doses of PFOA previously used inour laboratory [50], taking into consideration the half-life of PFNA(estimated at 40 days in female mice).
Maternal weights and signs of toxicity were monitored daily.At GD 17, mice from each dose group were assigned to one of twogroups: some dams were sacrificed for maternal and fetal examina-tions (received PFNA GD 1–16); the rest (received PFNA GD 1–17)were allowed to give birth. For animals sacrificed by decapitation
on GD 17, trunk blood from each dam was collected and serumstored at −20 ◦C for subsequent analysis of PFNA. The gravid uteruswas removed, weighed and examined, and individual live fetuseswere weighed and prepared for teratological evaluation. Half of the
e Toxi
fnsoMTaaiott
2
wsbFs
2
att
2
caPhmmaw
fawUsm
rawont
2
v(Fwfoww
Pxmp4 (Mm00480657 m1), Pax6 (Mm00443072 m1),
K.P. Das et al. / Reproductiv
etuses from each litter were randomly assigned for skeletal exami-ation and the other half for visceral evaluation. Maternal liver andome fetal livers were removed, weighed, and immediately frozenn dry ice and stored at −80 ◦C for PCR studies or PFNA analysis.ice without live or dead fetuses were considered as non-pregnant.
o confirm non-pregnancy, uteri were removed and stained withmmonium sulfide to visualize implantation sites [51]. For damsllowed to deliver, mice were monitored on GD 18–19, at frequentntervals until parturition when the date and time of birth, numberf live and dead pups, and birth weights were recorded. Postna-al survival, growth, and development of the pups were monitoredhereafter.
.4. Skeletal evaluation
An overdose of pentobarbital was used to kill fetuses. Fetusesere then eviscerated, and immediately fixed in 95% ethanol. Sub-
equently all specimens were stained with alizarin red S and Alcianlue for visual examination of bone and cartilage, respectively [52].inally skeletal morphology was evaluated by the method of Narot-ky and Rogers [52].
.5. Visceral evaluation
Fetuses were fixed in Bodian’s solution (2% formaldehyde, 5%cetic acid, 72% ethanol, 21% water), and a free-hand razor sec-ioning and dissection technique was used to examine the head,horacic and abdominal viscera [53].
.6. Postnatal observation, necropsy and sample collection
All litters were carefully observed daily for any morphologicalhange and weighed on postnatal day (PND) 1, 2, 3, 4, 7, 10, 14, 17,nd 21. Eye opening was monitored daily beginning on PND 10. OnND 24, pups were weighed, weaned, and males and females wereoused separately. Vaginal opening and preputial separation wereonitored beginning on PND 25. For post-weaning growth deter-ination, four male and four female pups were selected randomly
t PND 25 from each litter for biweekly weighing, and body weightsere recorded up to PND 287.
After weaning, dams were weighed and necropsies were per-ormed on the 28th day postpartum. Blood samples were collected,nd serum was stored frozen at −20 ◦C for PFNA analysis. Liversere removed, weighed, and stored at −80 ◦C for PFNA analysis.teri were removed and stained with 2% ammonium sulfide, and
ubsequently the implantation sites were counted according to theethod of Narotsky and Kavlock [51].One male and one female from each litter were weighed and sac-
ificed at PND 1, 10, 24, 42 and 70. Blood samples were collected,nd serum was stored frozen at −20 ◦C for PFNA analysis. Liversere removed, weighed, and about 200 mg (based on availability)
f fresh liver tissue was stored in RNA-Later® (AM7021 Life Tech-ologies, Grand Island, NY) and processed for specific gene analysis;he rest of the liver was stored at −80 ◦C for PFNA analysis.
.7. Determination of PFNA in serum and liver
The procedures for chemical analysis have been described pre-iously [33]. In brief, mouse livers were thawed, homogenizedpolytron) in six volumes of picopure water, and stored at −80 ◦C.or sample preparation, 25 �l of serum or 50 �l of liver homogenateere aliquoted into polypropylene tubes, to which 100 �l of 0.1 M
ormic acid and 1 ml of acetonitrile (ACN) fortified with 12.5 ng/mlf internal standard (13C9-labeled PFNA) was added sequentially,ith vigorous mixing (vortex) between each addition. Samplesere then capped, mixed for 20 min, and centrifuged (×3500 rpm)
cology 51 (2015) 133–144 135
for 5 min at room temperature. Two-hundred �l of the supernatantwas transferred to a HPLC auto sampling vial, followed by the addi-tion of 200 �l of 2 mM ammonium acetate.
Analysis of samples was performed using a LC/MS/MS sys-tem consisting of an Agilent 1100 HPLC (Agilent Technologies,Palo Alto, CA) interfaced with a Sciex 3000 triple quadrupolemass spectrometer (Applied Biosystems/MDS Sciex, Foster City,CA). The HPLC method was a 3.5 min isocratic run at a flowrate of 200 �l/min consisting of 70% ACN and 30% 2 mM ammo-nium acetate. Samples (10 �l) were injected onto a PhenomenexLuna C18 (3 mm × 50 mm) HPLC analytical column fitted with a4 × 30 mm Phenomenex Security Guard column (Torrance, CA). Thecolumn temperature was kept constant at 25 ◦C in a temperature-controlled column compartment. Electrospray negative ionization(−1500 V) was used in the mass spectrometer source, withturbo-dryer gas maintained at 375 ◦C. Mass transitions for PFNA(analyte) and 13C9-PFNA (internal standard) were monitored usingmultiple-reaction-monitoring (MRM) and analyte-specific massspectrometer parameters were optimized for both compounds.Two mass transitions were monitored for quantification of theanalyte (PFNA). A quantification ion (463.0 m/z → 418.8 m/z) anda confirmation ion (463.0 m/z → 218.9 m/z), were monitored forPFNA in order to ensure proper analyte identification and quan-tification. The mass transition monitored for 13C9-PFNA (internalstandard) was 472.0 m/z → 427.0 m/z. The limit of quantification(LOQ) for analysis was set to the lowest calibration point on thestandard curve. For samples below the LOQ (primarily control ani-mals), we assigned a value equal to the LOQ divided by the squareroot of 2 [54]. Analyst 1.4.2 software was used for quantification ofall compounds (Applied Biosystems/MDS Sciex, Foster, CA).
2.8. Analysis and evaluation of gene expression by real-time PCR
Transcripts evaluated by real-time RT-PCR included genesknown to be regulated by PPAR� in addition to several cytochromeP450 genes commonly used to assess the activity of various othernuclear receptors. Additional genes included those associated withglucose and cholesterol metabolism, two functional categoriespotentially regulated by PPAR�. Pax6 was evaluated as a nega-tive control since changes in the expression of this gene were notexpected in the fetal liver based on historical data from our labora-tory.
Two micrograms of total RNA were digested using 2 unitsDNaseI (#M6101, Promega Corporation, Madison, WI) accordingto the manufacturer’s protocol. The RNA was then quantifiedusing a Quant-iT RiboGreen RNA assay kit according to themanufacturer’s protocol (#R11490, Life Technologies, GrandIsland, NY) and approximately 1.5 �g RNA reverse transcribedusing a High Capacity cDNA Archive Kit according to the pro-vided protocol (#4322171, Life Technologies). Amplificationwas performed on an Applied Biosystems model 7900HT FastReal-Time PCR System in duplicate using 25 ng cDNA and Taq-Man Universal PCR Master Mix (#4304437, Life Technologies)in a total volume of 12 �l. Beta-2-microglobulin (B2m, Entrez#12010) was used as the endogenous reference gene. The fol-lowing TaqMan assays (Life Technologies) were included inthe study: B2m (Mm00437762 m1), A2m (Mm00558642 m1),Acox1 (Mm00443579 m1), Amy1 (Mm00651524 m1),Cyp17a1 (Mm00484040 m1), Gck (Mm00439129 m1),Hmgcs1 (Mm00524111 m1), Pdk4 (Mm00443325 m1),
Ehhadh (Mm00470091 s1), Cyp3all (Mm00731567 m1),Cyp1a1 (Mm00487218 m1), Lss (Mm00461312 m1), Pexlla(Mm00478137 m1), Mbl2 (Mm 00487623 m1), Cyp7a1(Mm00484152 m1), and Cyp2b10 (Mm00456591 m1).

1 e Toxicology 51 (2015) 133–144
2
ueedpTsp(cawmTwwwltWmow[
m(qomssawtalaothpoa
3
3
bs1sdgisgoh5
Maternal Weight Gain
Gestational Age (days)
181614121086420
weig
ht (g
ram
s)
0
5
10
15
20
25
30Control1 mg/k g3 mg/k g 5 mg/k g
**
*
*
*
*
*
Fig. 1. Effects of gestational exposure to PFNA (GD 1–17) on maternal weight gain in
36 K.P. Das et al. / Reproductiv
.9. Statistical analysis
Data are presented as means and standard errors. Litter wassed as the statistical unit. Statistical significance of treatmentffects was determined for each outcome variable using mainffects analysis of variance (ANOVA) that included block andose as factors. When a significant treatment effect was detected,airwise differences among groups were tested post hoc usingukey–Kramer adjustments for multiple comparisons. For mea-urements taken across time, repeated measures analyses wereerformed with mixed effects linear models using SAS Proc MixedSAS/STAT 9.3, Cary, NC). The models included an autoregressiveovariance structure for data across time points. Fixed effects werege, treatment and the treatment by age interaction, and blockas included as a random effect. Pairwise tests among treat-ent groups within each age/time point were calculated with
ukey–Kramer adjustments. For maternal weight gain the damas considered the subject for the repeated measures; for pupeight, the pup (nested within dam) was the subject and damas included as a random factor. Gene expression data were ana-
yzed using one-way ANOVA by evaluating the change in cyclehreshold for each gene relative to the reference gene (�CT).
here a significant main effect was observed, individual treat-ent contrasts were evaluated for significance by a Student’s t-test
f the least squares means (SAS JMP 7.0, Cary, NC). Fold changeas calculated using the 2−��CT method of Livak and Schmittgen
55].The U.S. Environmental Protection Agency now uses the bench-
ark dose (BMD) approach [56,57] for non-cancer risk assessmentU.S. EPA, 1995). This approach is designed to provide a moreuantitative alternative to dose-response assessment than the no-bserved-adverse-effect-level (NOAEL) process, by constructingathematical models to fit all data points in the dose-response
tudy, and to take data variance into consideration. In the currenttudy, BMD5 and BMDL5 values were calculated for the maternalnd developmental toxicity of PFNA using Benchmark Dose Soft-are 2.2 (U.S. EPA, 2011). BMD5 refers to the central estimate of
he administered dose predicted to cause a 5% increase in responsebove background, BMDL5 is defined as the corresponding lowerimit of the 95% confidence interval on the BMD5 [58]. Selection of
specific curve-fitting model for the BMD determination was basedn the Akaike’s Information Criterion (AIC) value. The AIC is equalo −2L + 2p, where L is the log-likelihood at the maximum likeli-ood estimates for the parameters, and p is the number of modelarameters estimated. The model that demonstrates “goodness-f-fit” with the lowest AIC value is presumed to be the mostppropriate.
. Results
.1. Maternal body weight
Gestationally exposed mice from each PFNA dose group gainedody weight as expected (Fig. 1). Maternal weight gains showed noignificant dose-dependent changes except for increases from GD1 to GD 17 for the 3 and 5 mg/kg groups. Pregnant mice did nothow overt toxicity except at the highest dose of 10 mg/kg, whereams did not carry their pregnancies to term (Table 1). In fact, damsiven 10 mg/kg PFNA began to lose weight (instead of gaining asn controls) significantly at GD 8 and thereafter. These mice were
acrificed on GD 13. At that stage, body weight of dams in 10 mg/kgroup was 26.9 ± 2.2 g, compared to 38.2 ± 0.7 in controls. Becausevert toxicity was clearly indicated in the 10 mg/kg dose group, theighest PFNA dose evaluated for the remainder of this study wasmg/kg.
mice. ANOVA with repeated measure and Bonferroni adjustment indicated a signifi-cant dose x age interaction. Points represent means ± S.E. of 19–27 animals. Asterisksdenote significant differences (p < 0.05) from controls.
3.2. Non-pregnant and maternal liver weight
Exposure of pregnant and non-pregnant mice to 1, 3, or 5 mg/kgPFNA resulted in increased absolute and relative liver weight atterm (mice examined on GD 17) (Fig. 2). The increase in both abso-lute and relative liver weight was dose-dependent in pregnantmice. In non-pregnant mice, the dose-dependent increases werealso seen in relative liver weight, although these increases in abso-lute liver weight were indistinguishable among the PFNA dosegroups. At 28 days postpartum (4 days after weaning), the increasedabsolute and relative liver weights were maintained in all of thetreatment groups (Fig. 2).
3.3. Pregnancy outcome
In utero PFNA exposure up to 5 mg/kg had no detrimental effectson pregnancy outcome at term (Table 1). There were no changes innumber of implants, number of live fetuses, or fetal weights follow-ing exposure to 1, 3, or 5 mg/kg PFNA. The absolute and relative fetalliver weights were significantly increased in the treated groups,but no dose-dependency was apparent. There was no evidence oftreatment effects from either skeletal or visceral examination offetuses (data not shown). Prenatal PFNA exposure produced signif-icant increases in full-litter resorptions only at 10 mg/kg dose andthis observation was noted on GD 13 where metrial gland only,metrial gland plus placenta, macerated fetuses or dead fetus werenoted.
3.4. Serum and liver PFNA levels in dams and fetuses
The mean serum concentrations of PFNA in pregnant mice atterm showed a dose-dependent increase following administrationof 1, 3, or 5 mg/kg PFNA (Fig. 3A). After a similar 17 days of chemicaltreatment, the serum concentrations of PFNA in non-pregnant micewere approximately twice as high as those in pregnant mice. At28 days after parturition, the serum concentrations of PFNA in thenursing dams of 1 or 3 mg/kg groups fell below the levels detectedat term. The mean liver concentration of PFNA in dams at term, in
non-pregnant mice, and in dams post weaning mirrored the profileof serum levels (Fig. 3B). The degree of PFNA accumulation in mouseliver was highest in non-pregnant, next higher in pregnant, andlowest in post-weaning (PND 28) mice. The hepatic levels of PFNA
K.P. Das et al. / Reproductive Toxicology 51 (2015) 133–144 137
Table 1Mouse reproductive outcome examined at GD 17. Data represent mean ± SE per litter examined where n = 8–10. Significance between control and treatment groups areindicated by *(P < 0.05). FLR = full-litter resorptions. *For the 10 mg/kg group, dams began to lose weight by GD 8, and were sacrificed on GD 13; body weight of controls atGD 13 was 38.2 ± 0.7.
PFNA (mg/kg) 0 1 3 5 10Non-pregnant examined (#) 4 6 5 2 1Pregnant dams examined (#) 8 8 8 10 7Pregnant dams with FLR (#) 2 0 1 1 7Maternal body weight (g) 47.6 ± 1.9 51.4 ± 1.3 52.4 ± 2.4 51.2 ± 2.6 26.9 ± 2.2 at GD 13*Maternal body weight minus gravid uterus weight (g) 31.5 ± 1.0 33.7 ± 0.7 35.3 ± 1.4 34.8 ± 1.2 –Implants (# per litter) 12.0 ± 1.5 11.8 ± 0.8 11.0 ± 1.1 11.4 ± 1.3 –Live fetuses (# per litter) 11.3 ± 1.3 11.5 ± 0.7 10.8 ± 1.0 10.0 ± 1.6 –% Live fetus per litter 94.8 ± 2.7 98.3 ± 1.7 98.1 ± 1.2 92.3 ± 2.4 –% Prenatal loss per litter 5.2 ± 2.7 1.7 ± 1.7 1.9 ± 1.2 7.7 ± 2.4 –
1.00.08.1
waibt
Fid
Fetal weight (g) 1.02 ± 0.04
Absolute fetal liver weight (g) 0.07 ± 0.01
Relative fetal liver weight (%) 6.67 ± 0.35
ere consistently about 5–10 times higher than the serum levels
t each corresponding administered dose. The PFNA concentrationsn fetal liver also increased in a dose-dependent manner (Fig. 3C),ut their levels were much lower (at 11–20%) than those seen inhe maternal liver.A. At term
PFNA Dose(mg/kg)
531Control
wei
ght (
gram
s)
0
1
2
3
4
5
6
7 PregnantNonpregnant
C. At term
PFNA Dose (mg/kg)
531Control
perc
ent
0
2
4
6
8
10
12
14
16
18
perc
ent
b
c
d
cc
b
c
d
cc
b
b
Absolute
Relative M
a
a
a
a
ig. 2. Liver weight of mice exposed to various doses of PFNA. Panels (A) and (C) illustrndicated a significant treatment effect. Bars represent the means ± S.E. of 5–15 animalsetermined by Tukey–Kramer test.
6 ± 0.02 1.07 ± 0.02 0.98 ± 0.1 –9 ± 0.01* 0.09 ± 0.01* 0.08 ± 0.01 –4 ± 0.3* 8.2 ± 0.4* 8.0 ± 0.23* –
3.5. Postnatal survival
Postnatal survival was monitored until weaning at PND 24and was significantly affected only at the 5 mg/kg dose of PFNA(Fig. 4). The pup deaths in the 5 mg/kg group started on PND 2 and
B. Post-weaning
PFNA Dose (mg/kg)
531Control
wei
ght (
gram
s)
0
1
2
3
4
5
6
7
D. Post-weaning
PFNA Dose (mg/kg)
531Control0
2
4
6
8
10
12
14
16
18
c
d
d
c
Maternal Liver Weights
aternal Liver Weights
ab
a
b
ate data at term, panels (B) and (D) illustrate data at 28 days postpartum. ANOVA. Different letters denote significant differences (p < 0.05) among exposure groups
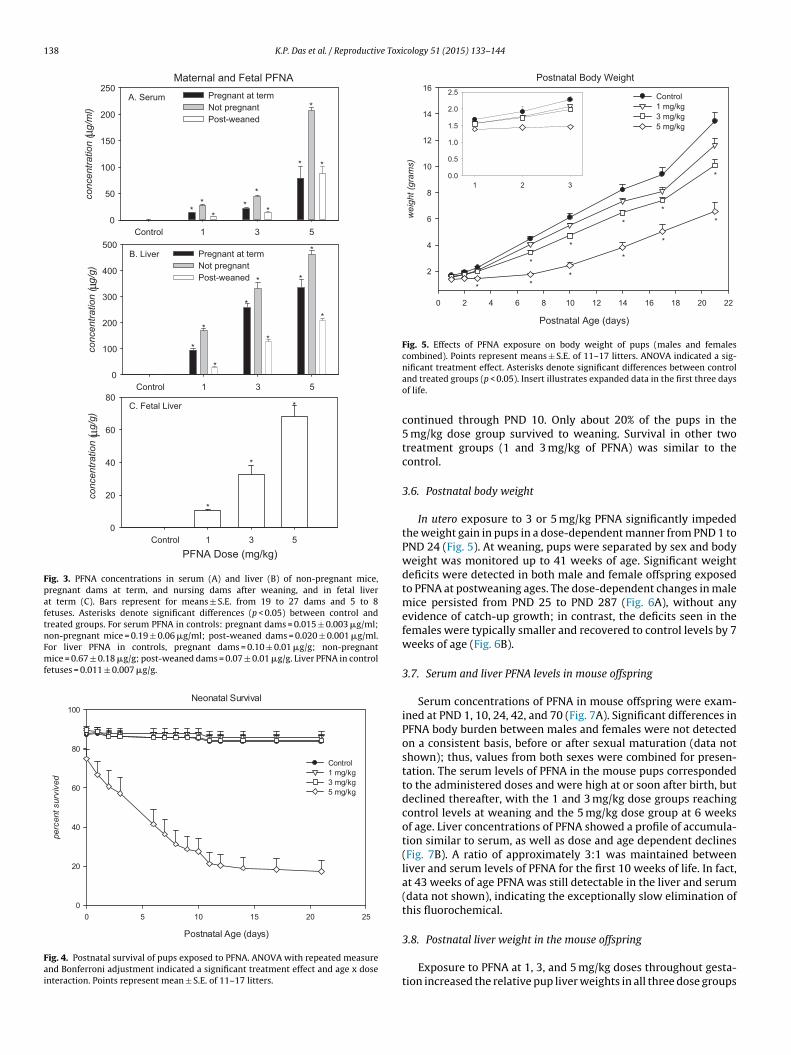
138 K.P. Das et al. / Reproductive Toxicology 51 (2015) 133–144
Maternal and Fetal PFNA
531Control
conc
entra
tion
(g/
ml)
0
50
100
150
200
250Pregnant at termNot pregnant Post-weaned
B. Liver
531Control
conc
entra
tion
(g/
g)
0
100
200
300
400
500Pregnant at termNot pregnantPost-weaned
C. Fetal Liver
PFNA Dose (mg/kg)531Control
conc
entra
tion
(g/
g)
0
20
40
60
80
*
*
*
*
*
*
*
*
*
*
*
*
**
**
*
*
*
*
*
A. Serumµ
µµ
Fig. 3. PFNA concentrations in serum (A) and liver (B) of non-pregnant mice,pregnant dams at term, and nursing dams after weaning, and in fetal liverat term (C). Bars represent for means ± S.E. from 19 to 27 dams and 5 to 8fetuses. Asterisks denote significant differences (p < 0.05) between control andtreated groups. For serum PFNA in controls: pregnant dams = 0.015 ± 0.003 �g/ml;non-pregnant mice = 0.19 ± 0.06 �g/ml; post-weaned dams = 0.020 ± 0.001 �g/ml.For liver PFNA in controls, pregnant dams = 0.10 ± 0.01 �g/g; non-pregnantmice = 0.67 ± 0.18 �g/g; post-weaned dams = 0.07 ± 0.01 �g/g. Liver PFNA in controlfetuses = 0.011 ± 0.007 �g/g.
Neo natal Survival
Postnatal Age (days)
2520151050
perc
ent s
urvi
ved
0
20
40
60
80
100
Control 1 mg/kg 3 mg/k g 5 mg/k g
Fig. 4. Postnatal survival of pups exposed to PFNA. ANOVA with repeated measureand Bonferroni adjustment indicated a significant treatment effect and age x doseinteraction. Points represent mean ± S.E. of 11–17 litters.
Postnatal Body Weight
Postnatal Age (days)
2220181614121086420
wei
ght (
gram
s)
2
4
6
8
10
12
14
16Control1 mg/kg3 mg/kg5 mg/kg
3210.0
0.5
1.0
1.5
2.0
2.5
**
*
*
*
*
*
*
**
*
Fig. 5. Effects of PFNA exposure on body weight of pups (males and femalescombined). Points represent means ± S.E. of 11–17 litters. ANOVA indicated a sig-
nificant treatment effect. Asterisks denote significant differences between controland treated groups (p < 0.05). Insert illustrates expanded data in the first three daysof life.continued through PND 10. Only about 20% of the pups in the5 mg/kg dose group survived to weaning. Survival in other twotreatment groups (1 and 3 mg/kg of PFNA) was similar to thecontrol.
3.6. Postnatal body weight
In utero exposure to 3 or 5 mg/kg PFNA significantly impededthe weight gain in pups in a dose-dependent manner from PND 1 toPND 24 (Fig. 5). At weaning, pups were separated by sex and bodyweight was monitored up to 41 weeks of age. Significant weightdeficits were detected in both male and female offspring exposedto PFNA at postweaning ages. The dose-dependent changes in malemice persisted from PND 25 to PND 287 (Fig. 6A), without anyevidence of catch-up growth; in contrast, the deficits seen in thefemales were typically smaller and recovered to control levels by 7weeks of age (Fig. 6B).
3.7. Serum and liver PFNA levels in mouse offspring
Serum concentrations of PFNA in mouse offspring were exam-ined at PND 1, 10, 24, 42, and 70 (Fig. 7A). Significant differences inPFNA body burden between males and females were not detectedon a consistent basis, before or after sexual maturation (data notshown); thus, values from both sexes were combined for presen-tation. The serum levels of PFNA in the mouse pups correspondedto the administered doses and were high at or soon after birth, butdeclined thereafter, with the 1 and 3 mg/kg dose groups reachingcontrol levels at weaning and the 5 mg/kg dose group at 6 weeksof age. Liver concentrations of PFNA showed a profile of accumula-tion similar to serum, as well as dose and age dependent declines(Fig. 7B). A ratio of approximately 3:1 was maintained betweenliver and serum levels of PFNA for the first 10 weeks of life. In fact,at 43 weeks of age PFNA was still detectable in the liver and serum(data not shown), indicating the exceptionally slow elimination ofthis fluorochemical.
3.8. Postnatal liver weight in the mouse offspring
Exposure to PFNA at 1, 3, and 5 mg/kg doses throughout gesta-tion increased the relative pup liver weights in all three dose groups

K.P. Das et al. / Reproductive Toxicology 51 (2015) 133–144 139
Post-weaning Body Weight
Postnatal Age (days)
3002752502252001751501251007550250
wei
ght (
gram
s)
0
10
20
30
40
50
60
70
Control 1 mg/kg3 mg/kg5 mg/kg
B. Female
Postnatal Age (days)
3002752502252001751501251007550250
wei
ght (
gram
s)
0
10
20
30
40
50
60
70
Control1 mg/kg3 mg/kg5 mg/kg
**
**
* **
* ****
* * * * * * * * * *
A. Male
**
**
Fig. 6. Post-weaning body weight of male (A) and female (B) offspring exposedto PFNA. ANOVA with repeated measure and Bonferroni adjustment indicated asignificant overall treatment effect and significant dose x age x sex interaction. Step-del
a4tr
3
o(pn1tdps
3
FsbMsPic
PFNA i n Offspring
Postnatal Age (days)
706050403020100
conc
entra
tion
(µg/
ml)
0
50
100
150
200
250Con tro l1 mg/k g 3 mg/k g 5 mg/kg
B. Liver
Postnatal Age (days)
706050403020100
conc
entra
tion
(µg/
g)
0
50
100
150
200
250Con tro l1 mg/k g 3 mg/k g 5 mg/k g
A. Serum
**
*
*
**
*
*
*
* *
*
* *
*
* *
* *
Fig. 7. PFNA concentrations in serum and liver of mouse offspring exposed toPFNA. Points represent means ± S.E. of 6–13 litters (data combined from malesand females). Asterisks denote significant differences from controls. In controls,
own analysis indicated a significant treatment effect for males, and a treatmentffect and dose x age interaction for females. Points represent means ± S.E. of 6–13itters. Asterisks denote significant differences (p < 0.05) from controls.
t PND 1, 10, and 24 in a dose-dependent manner, whereas at PND2 significant elevations of relative liver weight were seen only inhe two higher dose groups and at PND 70 no significant effectsemained (Fig. 8).
.9. Developmental landmarks
Consistent with the adverse effects on growth, postnatal devel-pment of pups exposed to PFNA in utero was significantly delayedFig. 9). In control mice, eye-opening was noted at 15 days ofostnatal age, preputial separation in males at 28 days and vagi-al opening in females at 30 days. Mouse offspring exposed to
mg/kg reached these developmental landmarks at approximatelyhe same age as controls. However, there were dose-dependentelays in the 3 and 5 mg/kg groups, with delays of eye-opening andreputial separation by 2 and 5 days, and delays of vaginal openingeparation by 3 and 7 days, respectively.
.10. Gene expression in fetal and neonatal mouse liver
Real-time PCR analysis of selected transcripts is shown inig. 10. PFNA induced a clear PPAR�-dependent gene expressionignature in both fetal and neonatal mouse liver as characterizedy altered expression of Acox1, Ehhadh, Pdk4, Pex11a, Pxmp4, andbl2 [59]. Cyp17a1, a cytochrome p450 gene associated with
teroid hormone synthesis and a gene potentially regulated byPAR� [60], was up-regulated primarily in fetal mice. Of particularnterest was Cyp2b10, a transcript related to activation of theonstitutive androstane receptor (CAR) and a gene previously
serum PFNA ranged from 0.008 to 0.024 �g/ml, and liver PFNA ranged from 0.050to 0.109 �g/g.
shown to be modified by other PFAAs [61–63]. Cyp2b10, however,was up-regulated only on PND 24, whereas, Cyp3a11, a transcriptoften associated with activation of the pregnane X receptor (PXR)but one which can also be regulated by CAR as well [64], wasup-regulated across several time points. In contrast, Cyp1a1 andCyp7a1, two commonly used markers of the aryl hydrocarbonreceptor (AhR) and the liver X receptor (LXR), respectively, werenot found to be up-regulated in the current study, although theexpression of Cyp7a1 was not evaluated until PND 42. It was clearfrom these data that activation of PPAR� by PFNA persisted intoyoung adulthood, as indicated by up-regulation of Acox1, Ehhadh,and Pex11a on PND 42. However, as a likely reflection of the PFNAbody burden in the mouse offspring, these transcriptional changesappeared to wane after PND 24. It should be noted that estimatesof gene expression on PND 42 and PND 70 were less certain thanthose at the earlier time points due to reduced numbers of liversamples in these groups. As expected, expression of the negativecontrol gene Pax6, was not detected at any time point in the study.
4. Discussion
The developmental toxicity of PFOS, PFOA, and perfluorobu-tanoic acid (PFBA) in the mouse has been described previously

140 K.P. Das et al. / Reproductive Toxicology 51 (2015) 133–144
Offspring Liver Weight
Postnatal age (days)706050403020100
weig
ht (g
ram
s)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Control1 mg/kg3 mg/k g5 mg/k g
B. Relative Liver weight
Postnatal age (da ys)706050403020100
perc
ent
0
2
4
6
8
10
12
Control1 mg/k g3 mg/k g5 mg/k g
*
*
*
*
*
* *
*
*
*
Fig. 8. Absolute (A) and relative (B) liver weight of offspring exposed to PFNA.ANOVA indicated a significant treatment effect. Points represent means ± S.E. of6t
bilPa5cwotllntwcide[ti
tpcdtb
C Vag inal ope ning
PFNA Dose (mg/kg)5310
Day
s
25
30
35
40
A Eye ope ning
PFNA Dose (mg/kg)5310
Day
s
5
10
15
20
25
B Preputial sepa ration
PFNA Dose (mg/kg )5310
Day
s
25
30
35
40
b
c
b
c
c
a a
b
a a
a,ba
Developmental Landmarks
Fig. 9. Developmental landmarks of mouse offspring exposed to PFNA. Panel (A)illustrates eye opening, panel (B) preputial separation in males and panel C vaginal
dose groups, which likely reflects the accumulation of PFNA in the
–13 litters. Asterisks denote significant differences (p < 0.05) between control andreatment groups.
y our laboratory [50,65–67]. Results from this study with PFNAllustrate a number of similarities and differences between theong-chain PFAAs. As noted with PFOS and PFOA, body burdens ofFNA were elevated with the increasing administered doses, with
preferential storage in the liver, reaching a liver-to-serum ratio of–8:1. Similar to PFAAs studied previously, PFNA crossed the pla-ental barrier readily, as a dose-dependent increase of the chemicalas seen in the fetal liver. Although the PFNA level in fetal liver was
nly a fraction of that in maternal liver, it was remarkably similar tohe serum levels in the pregnant dams at term, suggesting that theower chemical accumulation in the fetal hepatic compartment isimited by the placental transfer of PFNA. In addition, compared toon-pregnant mice at the corresponding doses and exposure dura-ion, the serum and hepatic levels of PFNA in the pregnant miceere always significantly lower, an observation consistent with the
hemical transfer from maternal to fetal compartment. It is alsonteresting to note the general fall of PFNA levels of the nursingams, perhaps reflecting clearance of the chemical through urinaryxcretion and lactational transfer, as previously shown with PFOA68]. Indeed, although PFNA in mouse milk was not determined inhe present study, this fluorochemical has been commonly detectedn human and cow milk, along with other PFAAs [69–71].
Neither PFOS, PFOA, nor PFNA produced any overt maternaloxicity (such as weight loss), except at very high doses whereregnancy loss was observed. In fact, for PFNA, small but signifi-ant increases of maternal weight were seen in the 3 and 5 mg/kg
ose groups during the last week of gestation, which likely reflectshe dose-related enlargement of maternal liver because the num-er of live fetuses and fetal weight were not influenced by PFNAopening in females. ANOVA indicated a significant treatment effect. Points representmeans ± S.E. of 6–13 litters. Different letters denote significant differences (p < 0.05)among exposure groups determined by Tukey–Kramer test.
treatment. The incidence of PFNA-induced hepatomegaly in bothpregnant and non-pregnant mice was not unexpected because thiseffect appears to be a common feature of PFAA exposure, whichhas been shown to be mediated largely by activation of the PPAR�signaling pathway. In fact, previous studies have indicated thatPFNA is equipotent, if not slightly more potent than PFOA in acti-vating the mouse PPAR� [35,72].
Similar to other PFAAs evaluated thus far, gestational exposureto PFNA did not produce any frank terata (skeletal or visceral) oraffect fetal viability or fetal weight. However, as noted with mater-nal liver, the fetal liver weights were increased significantly in all
fetal liver and activation of PPAR� (vide infra). Exposure to ourhighest dose (10 mg/kg) of PFNA led to full-litter resorptions thatwere detected by GD 13. As in the case of PFOA, the mechanism(s)

K.P. Das et al. / Reproductive Toxi
Fig. 10. Expression profile of target genes characterizing the responses of PPAR�,CAR or PXR in liver of mouse offspring exposed to PFNA. For each age point, the num-ber of controls is indicated in parentheses and the number of treated animals is givenin brackets. Red or green signals correspond to average up- or down-regulation,respectively. Non-significant values (NS) or genes not detected by RT-PCR ares3d
rbo
w3wigowoddIdpt
TB
hown in black. Missing data are indicated in gray. PFNA-1 = 1 mg/kg PFNA; PFNA- = 3 mg/kg PFNA; PFNA-5 = 5 mg/kg PFNA; GD = gestational day; PND = postnatalay. The full-color figure is available in the web version of this paper.
esponsible for the PFNA-induced early pregnancy loss is unknown,ut the perturbations apparently took place at the early stage ofrganogenesis.
Most (about 80% or more) of the PFNA-exposed mouse pupsere born alive and postnatal survival was not compromised at
mg/kg or lower doses. However, profound neonatal mortalityas noted in the 5 mg/kg group, with only 20% of the pups surviv-
ng to weaning. Although neonatal death was also reported afterestational exposure to PFOS and PFOA [50,65,73–76], the profilef this adverse effect elicited by PFNA is different from those seenith the eight-carbon PFAAs. A majority of the newborns (rats
r mice) exposed prenatally to high doses (15–20 mg/kg) of PFOSied within the first day of life [65]; some of those exposed to highoses (10–20 mg/kg) of PFOA survived about a day longer [50].
n contrast, the rate of neonatal mortality evinced by the 5 mg/kgose of PFNA was more gradual. Although born alive, these mouseups were weak and failed to thrive in the ensuing days, withhe number of deaths increasing steadily over the first 10 days of
able 2enchmark dose estimates of PFNA on various parameters for maternal and developmen
BMD5 (m
Increased absolute maternal liver weight at GD 17 0.43
Increased relative maternal liver weight at GD 17 0.30
Elevated relative fetal liver weight at GD 17 0.36
Reduced pup survival at PND 24 3.84
Reduced pup body weight at PND 1 1.03
Reduced pup body weight at PND 7 0.40
Reduced pup body weight at PND 14 1.76
Reduced pup body weight at PND 21 1.71
Increased pup relative liver weight at PND 1 0.24
Increased pup relative liver weight at PND 10 0.65
Increased pup relative liver weight at PND 24 0.59
Increased pup relative liver weight at PND 70 1.86
Delayed eye opening in offspring 1.99
Delayed preputial separation in male offspring 1.84
Delayed vaginal opening in female offspring 1.36
cology 51 (2015) 133–144 141
postnatal life. The loss of litters did not appear to be associatedwith a lack of maternal care (personal observation). About 20%of pups in this dose group survived past weaning into adulthood.The pathophysiological mechanism(s) of PFNA-induced neonatalmortality is unclear, but does not likely involve pulmonary insuffi-ciency potentially related to partitioning of chemicals into the lipidbilayers of lung surfactant that was suggested for PFOS and PFOA[10], primarily due to a lack of death seen abruptly within the firsthours to days after birth (as in the case of PFOS and PFOA). Alterna-tively, Wolf et al. [49] have clearly shown that PPAR� is an essentialmediator of PFNA-induced neonatal mortality, as incidence of post-natal loss was markedly attenuated in the transgenic 129S1/Svlmmice whose PPAR� was functionally knocked-out. In addition,the apparent physical weakness of the PFNA-exposed neonatesand their failure to thrive may suggest complications of processesinvolved in intermediary metabolism, as PFNA is known to becapable of and in fact, more potent than PFOA or PFOS in interferingwith the functions of metabolic sensors such as PPAR�. Furtherinvestigation will be required to unveil the mechanistic eventsleading to the differential responses among PFOS, PFOA and PFNAregarding neonatal death caused by exposure to these chemicals.
Postnatal growth deficits and developmental delays in rats andmice have been reported previously with gestational and lacta-tional exposure to PFOS and PFOA [50,66]. PFNA produced similaradverse effects, albeit in a more exaggerated manner. Weightgain of mouse neonates was significantly impeded during thepre-weaning period, in a dose-dependent manner. By weaning, sur-viving mouse offspring in the 3 mg/kg and 5 mg/kg groups weighedless than 73% and 50% respectively, of the controls. This growthretardation was likely related to the significant body burdens ofPFNA in the pups, in part due to the long elimination half-life (26–69days) of this chemical in mice [33]. Indeed, the serum and hepaticlevels of PFNA in the weanlings almost matched those of the nurs-ing dams. The growth impairment in the PFNA-exposed offspringcontinued into adulthood, but a sex-selectivity of this effect wasapparent. Body weight deficits in the males persisted to 41 weeksof age, and were most apparent in the 5 mg/kg group, although onemust be cautioned about the interpretation of these data as theyonly represent the few survivors in this dose group; in contrast,body weight of female offspring recovered to control levels by 6weeks of postnatal age. The reason behind this sex-selective effect ispresently unknown; however, one can rule out a sex-related differ-ence in the rate of PFNA elimination [33], and indeed, no discerniblesex-related differences in the body burdens of PFNA in the mouse
offspring were noted in the current study. Significant delays in eye-opening and onset of puberty (as indicated by preputial separationin males and vaginal opening in females) were noted in the 3 and5 mg/kg groups. Although these effects were also seen with PFOStal toxicity in mouse.
g/kg) BMDL5 (mg/kg)
0.270.210.223.110.930.281.401.010.190.460.431.170.941.301.07

1 e Toxi
aa
stodtoPrfnlaii[gPPat[hwa[riCpssanl
mdcpCnaTmotPc[towft
C
T
f
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
42 K.P. Das et al. / Reproductiv
nd PFOA exposure [50,66], the extent of delay was much greaternd more consistent with PFNA exposure.
Hepatomegaly and associated histopathological changes are aensitive feature of PFAA exposure in rodents. PFNA is no excep-ion to this finding. Relative liver weight in PFNA-exposed mouseffspring was significantly elevated in all dose groups, in a dose-ependent manner. Interestingly, the liver enlargement persistedhrough 10 weeks of postnatal age, in part reflecting the high degreef PFNA accumulation in this tissue. It is noteworthy that decline ofFNA in circulation preceded that in liver, as serum PFNA alreadyeached background levels by 6 weeks of age. Changes in hepaticunctions induced by PFAAs can also be evaluated by toxicoge-omic query. A strong signature of PPAR� responses, and to a
esser extent, that of CAR has been reported previously for PFOAnd PFOS [62,63,77–79]. The PPAR� pathway plays a major rolen maintaining lipid and glucose homeostasis, as well as regulat-ng inflammatory response, cell proliferation, and differentiation80]. In the present study, the expression profiles of candidateenes indicated that perinatal exposure to PFNA led to activation ofPAR�, CAR and PXR in the liver of the mouse offspring. The effect ofFNA on PPAR�-dependent gene expression persisted into youngdulthood. Altered expression of Pdk4 and Gck may further reflecthe effects of PFNA on gluconeogenic activity mediated by PPAR�81,82]. Cyp17a1, a cytochrome P450 gene associated with steroidormone synthesis and a gene potentially regulated by PPAR� [60],as up-regulated primarily in fetal mice as well. Because PFAAs
re potential activators of CAR and/or PXR in adult and fetal mice61–63,84], it was not surprising that target genes of these nucleareceptors such as Cyp3a11 and Cyp2b10 [64,83] were up-regulatedn the current study. However, it was not clear why regulation ofyp2b10 was limited to a single age point (PND 24). It should beointed out that Cyp2b10 was found to be a low copy number tran-cript in this study (data not shown), which might have introducedome uncertainty in the data. While changes in the expression of
number of hepatic target genes were persistent during the post-atal period, the overall effects diminished as animals aged, which
ikely corresponded to the gradual decline of PFNA in the liver.Exposure to PFNA during pregnancy thus produced a host of
aternal and developmental changes in the mouse, in a dose-ependent manner. To facilitate health risk assessment of thishemical, benchmark dose (BMD5) estimates on a few selectedarameters evaluated in the current study are presented in Table 2.omparable low values of BMD5 and BMDL5 were consistentlyoted with the hepatic effects of PFNA (on relative maternal, fetalnd pup liver weights), followed by the growth retarding effects.hese findings agree with the notion that hepatic effects are theost sensitive apical endpoint to indicate the adverse outcomes
f PFAAs in general. In summary, the profile of PFNA developmen-al toxicity in mice generally resembles those of other long-chainFAAs such as PFOA and PFOS, but differs from those of shorter-hain chemicals such as PFBA and perfluorohexanoic acid (PFHxA)67,85]. Collectively, our results are consistent with the hypothesishat the longer the alkyl chain length, the more potent are the devel-pmental effects of PFAAs (i.e. PFNA > PFOA, PFOS > PFBA, PFHxA),hich is likely related to their different rates of elimination (there-
ore body burden of the chemical) and their different potencies onarget tissues [10].
onflict of interest
The authors declare that there are no conflicts of interest.
ransparency document
The Transparency document associated with this article can beound in the online version.
[
cology 51 (2015) 133–144
Acknowledgments
The authors wish to thank Ms. Judith Schmid for her expert assis-tance in the statistical analysis of longitudinal data with repeatedmeasures, and Dr. Michael Narotsky for providing the BenchmarkDose estimates.
References
[1] Kissa E. Fluorinated surfactants and repellents. 2nd ed. New York: MarcelDekker; 2001.
[2] Giesy JP, Kannan K. Global distribution of perfluorooctane sulfonate in wildlife.Environ Sci Technol 2001;35:1339–42.
[3] Houde M, Martin JW, Letcher RJ, Solomon KR, Muir DC. Biological monitoring ofpolyfluoroalkyl substances: a review. Environ Sci Technol 2006;40:3463–73.
[4] Lau C, Anitole K, Hodes C, Lai D, Pfahles-Hutchens A, Seed J. Perfluoroalkyl acids:a review of monitoring and toxicological findings. Toxicol Sci 2007;99:366–94.
[5] Kato K, Wong LY, Jia LT, Kuklenyik Z, Calafat AM. Trends in exposure to polyflu-oroalkyl chemicals in the U.S. Population: 1999–2008. Environ Sci Technol2011;45:8037–45.
[6] Gewurtz SB, Backus SM, De Silva AO, Ahrens L, Armellin A, Evans M, et al.Perfluoroalkyl acids in the Canadian environment: multi-media assessmentof current status and trends. Environ Int 2013;59:183–200.
[7] Houde M, De Silva AO, Muir DC, Letcher RJ. Monitoring of perfluori-nated compounds in aquatic biota: an updated review. Environ Sci Technol2011;45:7962–73.
[8] Lau C, Butenhoff JL, Rogers JM. The developmental toxicity of perfluoroalkylacids and their derivatives. Toxicol Appl Pharmacol 2004;198:231–41.
[9] Giesy JP, Naile JE, Khim JS, Jones PD, Newsted JL. Aquatic toxicology of perflu-orinated chemicals. Rev Environ Contam Toxicol 2010;202:1–52.
10] Lau C. Perfluorinated compounds. In: Luch A, editor. Molecular clinical and envi-ronmental toxicology, vol. 3: Environmental toxicology. Basel, Switzerland:Birkhäuser-Verlag; 2012. p. 47–86.
11] Apelberg BJ, Witter FR, Herbstman JB, Calafat AM, Halden RU, Needham LL,et al. Cord serum concentrations of perfluorooctane sulfonate (PFOS) and per-fluorooctanoate (PFOA) in relation to weight and size at birth. Environ HealthPerspect 2007;115:1670–6.
12] Fei C, McLaughlin JK, Tarone RE, Olsen J. Fetal growth indicators and perfluori-nated chemicals: a study in the Danish National Birth Cohort. Am J Epidemiol2008;168:66–72.
13] Fei C, McLaughlin JK, Lipworth L, Olsen J. Maternal levels of perfluorinatedchemicals and subfecundity. Hum Reprod 2009;24:1200–5.
14] Washino N, Saijo Y, Sasaki S, Kato S, Ban S, Konishi K, et al. Correlations betweenprenatal exposure to perfluorinated chemicals and reduced fetal growth. Envi-ron Health Perspect 2009;117:660–7.
15] Lopez-Espinosa MJ, Fletcher T, Armstrong B, Genser B, Dhatariya K, MondalD, et al. Association of perfluorooctanoic acid (PFOA) and perfluorooctane sul-fonate (PFOS) with age of puberty among children living near a chemical plant.Environ Sci Technol 2011;45:8160–6.
16] Hoffman K, Webster TF, Weisskopf MG, Weinberg J, Vieira VM. Exposure topolyfluoroalkyl chemicals and attention deficit/hyperactivity disorder in U.S.children 12–15 years of age. Environ Health Perspect 2010;118:1762–7.
17] Vested A, Ramlau-Hansen CH, Olsen SF, Bonde JP, Kristensen SL, HalldorssonTI, et al. Associations of in utero exposure to perfluorinated alkyl acids withhuman semen quality and reproductive hormones in adult men. Environ HealthPerspect 2013;121:453–8.
18] Kristensen SL, Ramlau-Hansen CH, Ernst E, Olsen SF, Bonde JP, Vested A, et al.Long-term effects of prenatal exposure to perfluoroalkyl substances on femalereproduction. Hum Reprod 2013;28:3337–48.
19] Steenland K, Tinker S, Shankar A, Ducatman A. Association of perfluorooc-tanoic acid (PFOA) and perfluorooctane sulfonate (PFOS) with uric acid amongadults with elevated community exposure to PFOA. Environ Health Perspect2010;118:229–33.
20] Nelson JW, Hatch EE, Webster TF. Exposure to polyfluoroalkyl chemicals andcholesterol, body weight, and insulin resistance in the general US population.Environ Health Perspect 2010;118:197–202.
21] Melzer D, Rice N, Depledge MH, Henley WE, Galloway TS. Associationbetween serum perfluorooctanoic acid (PFOA) and thyroid disease in the USNational Health and Nutrition Examination Survey. Environ Health Perspect2010;118:686–92.
22] Knox SS, Jackson T, Frisbee SJ, Javins B, Ducatman AM. Perfluorocarbon expo-sure, gender and thyroid function in the C8 Health Project. J Toxicol Sci2011;36:403–10.
23] Grandjean P. Serum vaccine antibody concentrations in children exposed toperfluorinated compounds. J Am Med Assoc 2012;307:391–7.
24] Granum B, Haug LS, Namork E, Stolevik SB, Thomsen C, Aaberge IS, et al.Pre-natal exposure to perfluoroalkyl substances may be associated with altered
vaccine antibody levels and immune-related health outcomes in early child-hood. J Immunotoxicol 2013;10:373–9.25] Stein CR, Savitz DA. Serum perfluorinated compound concentration and atten-tion deficit/hyperactivity disorder in children 5–18 years of age. Environ HealthPerspect 2011;119:1466–71.

e Toxi
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
K.P. Das et al. / Reproductiv
26] Stein CR, Savitz DA, Bellinger DC. Perfluorooctanoate and neuropsychologicaloutcomes in children. Epidemiology 2013;24:590–9.
27] Vesterholm Jensen D, Christensen J, Virtanen HE, Skakkebaek NE, Main KM,Toppari J, et al. No association between exposure to perfluorinated compoundsand congenital cryptorchidism: a nested case-control study among 215 boysfrom Denmark and Finland. Reproduction 2014;147:411–7.
28] Whitworth KW, Haug LS, Baird DD, Becher G, Hoppin JA, Skjaerven R, et al. Per-fluorinated compounds and subfecundity in pregnant women. Epidemiology2012;23:257–63.
29] Olsen GW, Ehresman DJ, Buehrer BD, Gibson BA, Butenhoff JL, Zobel LR. Longi-tudinal assessment of lipid and hepatic clinical parameters in workers involvedwith the demolition of perfluoroalkyl manufacturing facilities. J Occup EnvironMed 2012;54:974–83.
30] Raymer JH, Michael LC, Studabaker WB, Olsen GW, Sloan CS, Wilcosky T, et al.Concentrations of perfluorooctane sulfonate (PFOS) and perfluorooctanoate(PFOA) and their associations with human semen quality measurements.Reprod Toxicol 2012;33:419–27.
31] Andersen CS, Fei CY, Gamborg M, Nohr EA, Sorensen TIA, Olsen J. Prenatal expo-sures to perfluorinated chemicals and anthropometry at 7 years of age. Am JEpidemiol 2013;178:921–7.
32] DeWitt JC, Shnyra A, Badr MZ, Loveless SE, Hoban D, Frame SR, et al. Immuno-toxicity of perfluorooctanoic acid and perfluorooctane sulfonate and therole of peroxisome proliferator-activated receptor alpha. Crit Rev Toxicol2009;39:76–94.
33] Tatum-Gibbs K, Wambaugh JF, Das KP, Zehr RD, Strynar MJ, Lindstrom AB, et al.Comparative pharmacokinetics of perfluorononanoic acid in rat and mouse.Toxicology 2011;281:48–55.
34] Lou IC, Wambaugh JF, Lau C, Hanson RG, Lindstrom AB, Strynar MJ, et al. Mod-eling single and repeated dose pharmacokinetics of PFOA in mice. Toxicol Sci2009;107:331–41.
35] Wolf CJ, Schmid JE, Lau C, Abbott BD. Activation of mouse and human per-oxisome proliferator-activated receptor-alpha (PPARalpha) by perfluoroalkylacids (PFAAs): further investigation of C4–C12 compounds. Reprod Toxicol2012;33:546–51.
36] Kinney LA, Chromey NC, Kennedy GL. Acute inhalation toxicity of ammoniumperfluorononanoate. Food Chem Toxicol 1989;27:465–8.
37] Fang XM, Zou SS, Zhao YY, Cui RN, Zhang W, Hu JY, et al. Kupf-fer cells suppress perfluorononanoic acid-induced hepatic peroxisomeproliferator-activated receptor alpha expression by releasing cytokines. ArchToxicol 2012;86:1515–25.
38] Fang XM, Gao GZ, Xue HY, Zhang XT, Wang HC. In vitro and in vivo studies ofthe toxic effects of perfluorononanoic acid on rat hepatocytes and Kupffer cells.Environ Toxicol Pharmacol 2012;34:484–94.
39] Fang XM, Gao GZ, Xue HY, Zhang XT, Wang HC. Exposure of perfluorononanoicacid suppresses the hepatic insulin signal pathway and increases serum glucosein rats. Toxicology 2012;294:109–15.
40] Zhang W, Liu Y, Zhang HX, Dai JY. Proteomic analysis of male zebrafishlivers chronically exposed to perfluorononanoic acid. Environ Int 2012;42:20–30.
41] Zhang W, Zhang YT, Zhang HX, Wang JS, Cui RN, Dai JY. Sex dif-ferences in transcriptional expression of FABPs in zebrafish liver afterchronic perfluorononanoic acid exposure. Environ Sci Technol 2012;46:5175–82.
42] Liu Y, Wang JS, Fang XM, Zhang HX, Dai JY. The thyroid-disrupting effects oflong-term perfluorononanoate exposure on zebrafish (Danio rerio). Ecotoxicol-ogy 2011;20:47–55.
43] Fang X, Zhang L, Feng Y, Zhao Y, Dai J. Immunotoxic effects of perfluorononanoicacid on BALB/c mice. Toxicol Sci 2008;105:312–21.
44] Fang X, Feng Y, Wang J, Dai J. Perfluorononanoic acid-induced apoptosis inrat spleen involves oxidative stress and the activation of caspase-independentdeath pathway. Toxicology 2010;267:54–9.
45] Rockwell CE, Turley AE, Cheng X, fields PE, Klaassen CD. Acute immunotoxiceffects of perfluorononanoic acid (PFNA) in C57BL/6 mice. Clin Exp Pharmacol2013;S4:002.
46] Feng YX, Shi ZM, Fang XM, Xu MQ, Dai JY. Perfluorononanoic acid induces apo-ptosis involving the Fas death receptor signaling pathway in rat testis. ToxicolLett 2009;190:224–30.
47] Feng YX, Fang XM, Shi ZM, Xu MQ, Dai JY. Effects of PFNA exposure on expres-sion of junction-associated molecules and secretory function in rat Sertoli cells.Reprod Toxicol 2010;30:429–37.
48] Stump DG, Holson JF, Murphy SR, Farr CH, Schmit B, Shinohara M. An oraltwo-generation reproductive toxicity study of S-111-S-WB in rats. Reprod Tox-icol 2008;25:7–20.
49] Wolf CJ, Zehr RD, Schmid JE, Lau C, Abbott BD. Developmental effectsof perfluorononanoic acid in the mouse are dependent on peroxisomeproliferator-activated receptor-alpha. PPAR Res 2010;2010, 11 pages [ArticleID 282896].
50] Lau C, Thibodeaux JR, Hanson RG, Narotsky MG, Rogers JM, Lindstrom AB, et al.Effects of perfluorooctanoic acid exposure during pregnancy in the mouse.Toxicol Sci 2006;90:510–8.
51] Narotsky MG, Kavlock RJ. In vivo assessment of prenatal developmental toxicity
in rodents. Curr Protoc Toxicol 2003 [ed. Mahin D Maines. Chapter 13: Unit135].52] Narotsky MG, Rogers JM. Examination of the axial skeleton of fetal rodents. In:Tuan RS, Lo CW, editors. Developmental biology protocols, vol. I. New Jersey:Humana; 2000. p. 139–50.
[
cology 51 (2015) 133–144 143
53] Wilson JG. Methods for administering agents and detecting malformations inexperimental animals. In: Wilson JG, Warkany J, editors. Teratology: principlesand techniques. Chicago: Univ Chicago Press; 1965. p. 262–77.
54] Calafat AM, Wong LY, Kuklenyik Z, Reidy JA, Needham LL. Polyfluoroalkylchemicals in the US population: Data from the National Health and Nutri-tion Examination Survey (NHANES) 2003–2004 and comparisons with NHANES1999–2000. Environ Health Perspect 2007;115:1596–602.
55] Livak KJ, Schmittgen TD. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2−��CT Method. Methods 2001;25:402–8.
56] Barnes DG, Daston GP, Evans JS, Jarabek AM, Kavlock RJ, Kimmel CA, et al.Benchmark dose workshop: criteria for use of a benchmark dose to estimate areference dose. Regul Toxicol Pharmacol 1995;21:296–306.
57] Crump KS. A new method for determining allowable daily intakes. FundamAppl Toxicol 1984;4:854–71.
58] Allen BC, Kavlock RJ, Kimmel CA, Faustman EM. Dose-response assessmentfor developmental toxicity. II. Comparison of generic benchmark dose esti-mates with no observed adverse effect levels. Fundam Appl Toxicol 1994;23:487–95.
59] Anderson SP, Dunn C, Laughter A, Yoon L, Swanson C, Stulnig TM, et al. Overlap-ping transcriptional programs regulated by the nuclear receptors peroxisomeproliferator-activated receptor alpha, retinoid X receptor, and liver X receptorin mouse liver. Mol Pharmacol 2004;66:1440–52.
60] Anderson SP, Howroyd P, Liu J, Qian X, Bahnemann R, Swanson C, KwakMK, Kensler TW, Corton JC. The transcriptional response to a peroxisomeproliferator-activated receptor alpha agonist includes increased expression ofproteome maintenance genes. J Biol Chem 2004;279:52390–8.
61] Cheng X, Klaassen CD. Perfluorocarboxylic acids induce cytochrome P450enzymes in mouse liver through activation of PPAR-alpha and CAR transcrip-tion factors. Toxicol Sci 2008;106:29–36.
62] Rosen MB, Schmid JE, Das KP, Wood CR, Zehr RD, Lau C. Gene expression profil-ing in the liver and lung of perfluorooctane sulfonate-exposed mouse fetuses:comparison to changes induced by exposure to perfluorooctanoic acid. ReprodToxicol 2009;27:278–88.
63] Rosen MB, Lee JS, Ren H, Vallanat B, Liu J, Waalkes MP, et al. Toxicogenomicdissection of the perfluorooctanoic acid transcript profile in mouse liver: evi-dence for the involvement of nuclear receptors PPAR alpha and CAR. ToxicolSci 2008;103:46–56.
64] Xie W, Barwick JL, Downes M, Blumberg B, Simon CM, Nelson MC, et al. Human-ized xenobiotic response in mice expressing nuclear receptor SXR. Nature2000;406:435–9.
65] Thibodeaux JR, Hanson RG, Rogers JM, Grey BE, Barbee BD, Richards JH, et al.Exposure to perfluorooctane sulfonate during pregnancy in rat and mouse. I.Maternal and prenatal evaluations. Toxicol Sci 2003;74:369–81.
66] Lau C, Thibodeaux JR, Hanson RG, Rogers JM, Grey BE, Stanton ME, et al.Exposure to perfluorooctane sulfonate during pregnancy in rat and mouse. II.Postnatal evaluation. Toxicol Sci 2003;74:382–92.
67] Das KP, Grey BE, Zehr RD, Wood CR, Butenhoff JL, Chang SC, et al. Effectsof perfluorobutyrate exposure during pregnancy in the mouse. Toxicol Sci2008;105:173–81.
68] Fenton SE, Reiner JL, Nakayama SF, Delinsky AD, Stanko JP, Hines EP, et al.Analysis of PFOA in dosed CD-1 mice. Part 2. Disposition of PFOA in tissuesand fluids from pregnant and lactating mice and their pups. Reprod Toxicol2009;27:365–72.
69] Karrman A, Ericson I, van Bavel B, Darnerud PO, Aune M, Glynn A, et al. Expo-sure of perfluorinated chemicals through lactation: levels of matched humanmilk and serum and a temporal trend, 1996–2004, in Sweden. Environ HealthPerspect 2007;115:226–30.
70] Capriotti AL, Cavaliere C, Cavazzini A, Foglia P, Lagana A, Piovesana S, et al.High performance liquid chromatography tandem mass spectrometry deter-mination of perfluorinated acids in cow milk. J Chromatogr A 2013;1319:72–9.
71] Mondal D, Weldon RH, Armstrong BG, Gibson LJ, Lopez-Espinosa MJ, ShinHM, et al. Breastfeeding: a potential excretion route for mothers and impli-cations for infant exposure to perfluoroalkyl acids. Environ Health Perspect2014;122:187–92.
72] Rosen MB, Das KP, Wood CR, Wolf CJ, Abbott BD, Lau C. Evaluation of perfluo-roalkyl acid activity using primary mouse and human hepatocytes. Toxicology2013;308:129–37.
73] Grasty RC, Grey BE, Lau CS, Rogers JM. Prenatal window of susceptibility toperfluorooctane sulfonate-induced neonatal mortality in the Sprague-Dawleyrat. Birth Defects Res B 2003;68:465–71.
74] Luebker DJ, York RG, Hansen KJ, Moore JA, Butenhoff JL. Neonatal mortalityfrom in utero exposure to perfluorooctanesulfonate (PFOS) in Sprague-Dawleyrats: dose–response, and biochemical and pharamacokinetic parameters. Tox-icology 2005;215:149–69.
75] Yahia D, Tsukuba C, Yoshida M, Sato I, Tsuda S. Neonatal death of mice treatedwith perfluorooctane sulfonate. J Toxicol Sci 2008;33:219–26.
76] Yahia D, Abd El-Nasser M, Abedel-Latif M, Tsukuba C, Yoshida M, Sato I, et al.Effects of perfluorooctanoic acid (PFOA) exposure to pregnant mice on repro-duction. J Toxicol Sci 2010;35:527–33.
77] Rosen MB, Abbott BD, Wolf DC, Corton JC, Wood CR, Schmid JE, et al. Geneprofiling in the livers of wild-type and PPAR{alpha}-null mice exposed to per-
fluorooctanoic acid (PFOA). Toxicol Pathol 2008;36:592–607.78] Rosen MB, Schmid JR, Corton JC, Zehr RD, Das KP, Abbott BD, et al. Geneexpression profiling in wild-type and PPARalpha-null mice exposed to perflu-orooctane sulfonate reveals pparalpha-independent effects. PPAR Res 2010:23[Article ID 794739].

1 e Toxi
[
[
[
[
[
[
44 K.P. Das et al. / Reproductiv
79] Oshida K, Vasani N, Carswell G, Ward W, Fostel J, Shah I, et al. Identificationof novel chemical and genetic effectors of the nuclear receptor peroxisomeproliferator-activated receptor-alpha (PPAR-a). PLoS ONE 2014 [in press].
80] Escher P, Wahli W. Peroxisome proliferator-activated receptors: insight into
multiple cellular functions. Mutat Res 2000;448:121–38.81] Desvergne B, Wahli W. Peroxisome proliferator-activated receptors: nuclearcontrol of metabolism. Endocr Rev 1999;20:649–88.
82] Patsouris D, Mandard S, Voshol PJ, Escher P, Tan NS, Havekes LM, et al. PPARal-pha governs glycerol metabolism. J Clin Invest 2004;114:94–103.
[
cology 51 (2015) 133–144
83] Xie W, Barwick JL, Simon CM, Pierce AM, Safe S, Blumberg B, et al. Reciprocalactivation of xenobiotic response genes by nuclear receptors SXR/PXR and CAR.Genes Dev 2000;14:3014–23.
84] Rosen MB, Thibodeaux JR, Wood CR, Zehr RD, Schmid JE, Lau C. Gene expres-
sion profiling in the lung and liver of PFOA-exposed mouse fetuses. Toxicology2007;239:15–33.85] Iwai H, Hoberman AM. Oral (gavage) combined developmental and perina-tal/postnatal reproduction toxicity study of ammonium salt of perfluorinatedhexanoic acid in mice. Int J Toxicol 2014;33:219–37.
Copyright © 2022 FDOKUMEN




















