
Thrust Restraint Design for Ductile Iron Pipe- Seventh Edition
Upload
khangminh22Category
view
1download
0
BIOLOGY OF REPRODUCTION (2013) 89(3):64, 1–12Published online before print 24 July 2013.DOI 10.1095/biolreprod.113.110619
Restraint Stress Impairs Oocyte Developmental Potential in Mice: Role of CRH-Induced Apoptosis of Ovarian Cells1
Bo Liang,3 De-Li Wei,3 Ya-Nan Cheng,3 Hong-Jie Yuan,3 Juan Lin, Xiang-Zhong Cui, Ming-Jiu Luo, andJing-He Tan2
College of Animal Science and Veterinary Medicine, Shandong Agricultural University, Tai-an City, People’s Republic ofChina
ABSTRACT
This study examined the role of CRH-induced ovarian cellapoptosis in the restraint stress (RS)-induced impairment ofoocyte competence. Oocyte percentages of apoptotic cumuluscells (CCs) did not differ between stressed and control micebefore in vitro maturation (IVM) but became significantly higherin stressed mice after IVM without serum, growth factor, andhormone. The level of Bcl2 mRNA decreased significantly inmural granulosa cells (MGCs) and ovarian homogenates afterRS. Whereas ovarian estradiol, testosterone, and IGF1 de-creased, cortisol and progesterone increased significantlyfollowing RS. RS increased the level of CRH in serum, ovary,and oocyte while enhancing the expression of CRHR1 in CCs,MGCs, and thecal cells. RS down-regulated ovarian expressionof glucocorticoid receptor and brain-derived neurotrophicfactor. Furthermore, CRH supplementation to IVM mediumimpaired oocyte developmental potential while increasingapoptotic CCs, an effect that was completely overcome byaddition of the CRHR1 antagonist antalarmin. Results suggestthat RS impaired oocyte competence by increasing CRH but notglucocorticoids. Increased CRH initiated a latent apoptoticprogram in CCs and oocytes during their intraovarian develop-ment, which was executed later during IVM to impair oocytecompetence. Thus, elevated CRH interacted with increasedCRHR1 on thecal cells and MGCs, reducing the production oftestosterone, estrogen, and IGF1 while increasing the level ofprogesterone. The imbalance between estrogen and progester-one and the decreased availability of growth factors triggeredapoptosis of MGCs and facilitated CC expression of CRHR1,which interacted with the oocyte-derived CRH later during IVMto induce CC apoptosis and reduce oocyte competence.
apoptosis, corticotrophin-releasing hormone (CRH), mouse,oocyte, restraint stress
INTRODUCTION
Studies suggest that psychological stress can exert detri-mental effects on reproduction in women [1–6]. Restraint of
animals is an experimental procedure developed for studies ofpsychogenic stress [7, 8]. Mice and rats exposed to restraintstress (RS) during pregnancy show impaired function ofcorpora lutea and reduced pregnancy rates and litter size [9–11]. Adverse effects of psychological stress on reproductionhave also been reported in sows [12, 13] and ewes [14].
A prospective study indicates that stressful life events mayreduce the chances of a successful outcome following in vitrofertilization (IVF), possibly through psychobiological mecha-nisms affecting medical end points such as oocyte retrievaloutcome [15]. Furthermore, our recent studies have unequiv-ocally demonstrated that RS applied during final folliculargrowth and maturation significantly diminished the develop-mental potential and induced chromosome aneuploidy ofmouse oocytes [16, 17]. However, the mechanism by whichpsychological stress damages the oocyte is largely unknown.
Studies on the mechanisms by which stress affects femalereproduction have demonstrated that stress activates thehypothalamo-pituitary-adrenocortical (HPA) axis and thusenhances production of corticotrophin-releasing hormone(CRH) by the hypothalamus, adrenocorticotropic hormone(ACTH) and b-endorphin by the pituitary, and glucocorticoidsby the adrenal cortex. Some studies suggest that CRH,glucocorticoids, and ACTH affect ovarian function at thehypothalamus and the pituitary gland levels to decrease thesynthesis and release of gonadotropin-releasing hormone,luteinizing hormone (LH), and follicle-stimulating hormone(FSH), whereas others suggest that they act on the ovary tomodulate steroidogenesis and/or oogenesis directly [18–20].However, evidence regarding the direct effect of the HPAproducts on the oocyte is limited and conflicting. For example,whereas RS of mice increased the level of cortisol butdiminished oocyte developmental competence significantly,exposure of mouse oocytes in vitro to stress-like concentrationsof cortisol did not affect their maturation and development tothe blastocyst stage [16, 21].
Early studies showed that oocytes from atretic follicles withmore apoptotic cells are less developmentally competent thanoocytes from healthy follicles with fewer apoptotic cells [22–24]. Stressors of different types [25–28] or treatments withglucocorticoids [29–32] induce apoptosis in different tissues,including testicular germ cells [33]. Fluid from follicles ofwomen whose oocytes were not fertilized contained cortisollevels significantly higher than in the fluid from folliclescontaining successfully fertilized oocytes [34]. Peripheral CRHhas been identified in the reproductive system, including theovary and testis [18]. CRH suppressed estrogen release fromcultured rat granulosa cells by inhibiting release of insulin-likegrowth factor 1 (IGF1) [35, 36], and both estrogen and IGF1inhibited apoptosis in different cells, including granulosa cells[37–39]. Furthermore, recent findings support the idea that
1Supported by grants from the National Basic Research Program ofChina (2012CB944403 and 2007CB947403), the China NationalNatural Science Foundation (31272444, 30972096, and 30771556),and the China National Project of Transgenics (2009ZX08008-006B).2Correspondence: Jing-He Tan, College of Animal Science andVeterinary Medicine, Shandong Agricultural University, Tai-an City,Shandong Province 271018, P. R. China. E-mail: [email protected] authors contributed equally to this work.
Received: 8 May 2013.First decision: 30 May 2013.Accepted: 1 July 2013.� 2013 by the Society for the Study of Reproduction, Inc.eISSN: 1529-7268 http://www.biolreprod.orgISSN: 0006-3363
1 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022

CRH participates in the stress-induced apoptosis of hippocam-pal neurons [40].
We therefore hypothesize that stress, and thus activation, ofthe HPA axis may impair oocyte potential by an indirectmechanism that induces apoptosis first in ovarian cells and thenin the oocyte itself. In other words, stress increases CRH and/orglucocorticoids that impair oocyte developmental potential bytriggering apoptosis of follicular cells. The objective of thepresent study was to test this hypothesis. Female mice weresubjected to RS for 24 h starting 24 h after equine chorionicgonadotropin (eCG) injection to mimic the human IVF-associated distress taking place sometime during the FSH-stimulation period in the follicular phase. At the end of the RSperiod, mice were killed to examine the role of CRH-inducedapoptosis of ovarian cells in the RS-induced impairment ofoocyte competence. Results suggested that RS impaired oocytecompetence by increasing CRH but not glucocorticoids.Increased CRH initiated a latent apoptotic program in cumuluscells (CCs) and oocytes during their intraovarian development,which was executed later during in vitro maturation (IVM) toimpair oocyte competence.
MATERIALS AND METHODS
Unless otherwise specified, all chemicals and reagents used in the presentstudy were purchased from Sigma Chemical Co.
Mice and Treatments
Mice of the Kunming strain were kept in a room with a 14L:10Dphotoperiod, with lights-off at 2000 h. The animals were handled according tothe rules stipulated by the Animal Care and Use Committee of ShandongAgricultural University. Female mice at the age of 6–8 wk were injected witheCG (10 IU i.p.), and at 24 h after eCG injection, the mice were subjected to RSfor 24 h. For restraint treatment, an individual mouse was put in a microcageconstructed by the authors [16], which was placed in an ordinary home cage.The microcage offered the same photoperiod and controlled temperature (22–258C) as in the large home cage for the unstressed animals. While in themicrocage, mice could move back and forth to some extent and could take foodand water freely, but they could not turn around. Antalarmin was dissolved firstin ethanol and then in saline. Immediately before restraint treatment, mice wereinjected i.p. with antalarmin (20 mg/kg in a volume of 0.1 ml) or an equivalentvolume of vehicle, and serum was recovered for cortisol measurement atdifferent times after injection or restraint treatment.
Recovery of Ovaries, Oocytes, and Mural Granulosa Cells
Both stressed and unstressed control mice were killed at 48 h after eCGinjection by decollation to collect ovaries for the recovery of oocytes and muralgranulosa cells (MGCs). The large follicles on the ovary were ruptured in M2medium [41] to release cumulus-oocyte complexes (COCs). Only COCs withmore than three layers of unexpanded CCs, containing oocytes larger than 70lm in diameter, and with a homogenous cytoplasm were used for experiments.The MGC sheets released into M2 medium at puncture of follicles werecollected and pelleted by centrifugation at 200 3 g for 5 min at roomtemperature. The pellets were then resuspended in TRIzol (Invitrogen,Australia Pty. Ltd.) or in the sample buffer for use in quantitative real-timePCR or Western blot analysis, respectively.
IVM of Oocytes
The COCs were cultured in groups of approximately 30 in 100-ll drops ofmaturation medium at 37.58C in a humidified atmosphere of 5% CO
2in air.
Two media were prepared for oocyte maturation based on TCM-199 (Gibco).Whereas medium with serum, growth factor, and hormone (SGH) wassupplemented with 10% (v/v) fetal bovine serum (Gibco), 1 lg/ml of 17b-estradiol, 24.2 mg/ml of sodium pyruvate, 0.05 IU/ml of FSH, 0.05 IU/ml ofLH, and 10 ng/ml of epidermal growth factor, medium without SGH wassupplemented with only 24.2 mg/ml of sodium pyruvate and 0.3 mg/ml ofpolyvinyl alcohol.
Antalarmin and/or CRH were added to maturation medium to observe theireffect on oocyte maturation. For preparation, CRH (2 310�4 M) and antalarmin
(2 3 10�4 M) were dissolved in water and dimethyl sulfoxide, respectively. Thestock solutions were stored at �208C until use.
Oocyte Activation and Embryo Culture
At 24 h of maturation culture, oocytes were stripped of their CCs bypipetting with a thin pipette in M2 medium containing 0.1% hyaluronidase.Oocytes were then incubated in the activating medium (Ca2þ-free CZB mediumsupplemented with 10 mM SrCl
2and 5 lg/ml of cytochalasin B) for 6 h at
37.58C in a humidified atmosphere with 5% CO2
in air. At the end of treatment,oocytes were examined with a Nikon inverted microscope for activation.Oocytes were considered to be activated when each contained one or two well-developed pronuclei. Activated oocytes were cultured for 4 days in regularCZB medium (30–35 oocytes per 100-ll drop). Glucose (5.5 mM) was addedto the CZB medium when embryos were beyond the 3- or 4-cell stage.
Assessment of CC Apoptosis
The CCs freed from 60–80 COCs were collected into a 0.5-ml tube andseparated from medium by centrifugation (200 3 g, 5 min, room temperature).The CC pellets were resuspended in 50 ll of M2 medium supplemented with0.01 mg/ml of Hoechst 33342 and stained in the dark for 5 min. The stainedcells were then centrifuged (200 3 g, 5 min, room temperature) again toconcentrate cells. After removal of approximately half the supernatant, a 5-lldrop of suspension was smeared on the slide and observed under a LeicaDMLB fluorescence microscope (4003). Six to eight fields were randomlyexamined on each smear, and percentages of apoptotic cells were calculatedfrom 60–80 cells observed in each field.
Blood Serum Preparation and Ovarian Homogenization
Mice were killed by decollation, and trunk blood (;1 ml) was collectedinto ice-cooled centrifugal tubes and centrifuged (1700 3 g, 10 min, 48C) toseparate serum. The serum collected was stored at�808C until hormone assay.For ovarian homogenization, the ovaries were snap-frozen in liquid nitrogenimmediately after removal from the females. The frozen ovaries were weighedand transferred to an electrical homogenizer (ULTRA TURRAX IKA T18basic) with the proper amount of homogenization solutions. Homogenizationwas performed while cooling on ice. Following homogenization, thehomogenates were centrifuged (15 000 3 g, 10 min, 48C), and the supernatantwas collected for immediate use or stored at �808C until use.
Quantitative Real-Time PCR
Ovarian homogenization was performed using TRIzol reagent (1 ml per 75mg of ovarian tissue). More than 5 3 105 MGCs from four mice orapproximately 500 COCs from each treatment were treated with TRIzol reagentfor RNA isolation. The RNA isolated was resuspended in diethyl pyrocar-bonate-treated MilliQ water (DEPC-dH
2O) and digested with RNase-free
DNase I (Takara Biotechniques). The purified RNA was dissolved in DEPC-dH
2O and spectroscopically quantified at 260 nm. Purity and integrity of the
RNA was assessed by determination of the A260
:A280
ratio (1.8–2.0) andelectrophoresis in 1% agarose.
TABLE 1. Oligonucleotide primer sequences used for real-time PCR.
cDNA Oligonucleotide sequences (50– 30)*Amplified
product size (bp)
Bcl2 F: TTCGGGATGGAGTAAACTGG 157R: TGGATCCAAGGCTCTAGGTG
Bax F: TGCAGAGGATGATTGCTGAC 183R: GATCAGCTCGGGCACTTTAG
Casp3 F: GGGCCTGTTGAACTGAAAAA 242R: CCGTCCTTTGAATTTCTCCA
Igf1 F: GGACCAGAGACCCTTTGCGGGG 210R: GGCTGCTTTTGTAGGCTTCAGTGG
Bdnf F: GCCTCCTCTACTCTTTCTG 255R: GGATTACACTTGGTCTCGT
Nr3c1 F: AGTCAAGGTTTCTGCGT 233R: CCATCACTTTTGTTTCG
Gapdh F: AAGGTGGTGAAGCAGGCAT 244R: GGTCCAGGGTTTCTTACTCCT
* F, forward; R, reverse.
LIANG ET AL.
2 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022

Reverse transcription was performed in a total volume of 20 ll usingSuperscript III Reverse Transcriptase (Invitrogen Australia Pty., Ltd). Briefly, 2ll of each RNA sample were mixed in a 0.2-ml reaction tube with 4 ll ofdNTP, 1.5 ll of Oligo dT
18(Takara), and 6 ll of DEPC-dH
2O, and the mixture
was incubated in a PCR instrument at 658C for 5 min. As soon as the incubationended, the reaction tube was cooled on ice for 2 min and then centrifuged (2003 g for 10 sec at 48C) for a few seconds. Then, 4 ll of 53 RT buffer, 0.5 ll ofRNase inhibitor, and 0.5 ll of Superscript III Reverse Transcriptase were addedto the reaction tube. The mixture was then incubated at 508C for 1 h, followedby incubation at 708C for 15 min before storage at �208C until use.
Gene-specific primers for real-time RT-PCR are listed in Table 1.Quantification of mRNA was conducted using the Mx3005P Real-Time PCRSystem (Stratagene). Amplification reactions were performed in a 10-llreaction volume containing 1 ll of cDNA, 5 ll of 23 SYBR Green Master Mix(Stratagene), 0.15 ll of 500-fold diluted reference dye, 3.05 ll of RNase-freewater, and 0.4 ll each of forward and reverse gene-specific primers (10 lM).Cycle amplification conditions comprised an initial denaturation step at 958Cfor 10 min followed by 40 cycles at 958C for 5 sec and 608C for 20 sec.Immediately after amplification, PCR products were analyzed by sequencing,dissociation-curve analysis, and gel electrophoresis to determine specificity ofthe reaction. Gene expression was normalized to the gapdh internal control. Allvalues were then expressed relative to calibrator samples using the 2�(DDCT)
method [42].
Hormone Assays
The same group of stressed or control mice were killed to collect serum andovaries for hormone assay. Ovaries were homogenized in PBS (800 ll per 100mg of ovarian tissue). Each treatment was repeated three times, with eachreplicate containing ovarian homogenates or serum from three mice. Whereasserum from each mouse was assayed individually, ovarian homogenates fromthree mice were pooled before assays.
Radioimmunoassay of steroid hormones. Radioimmunoassay wasconducted by the Central Hospital of Tai-An City using commercial kits fromJiuding Biomedical Techniques Co. Ltd. The minimum levels of detection forassays of estradiol, progesterone, testosterone, and cortisol were 1 pg/ml, 1 ng/ml, 10 pg/ml, and 10 ng/ml, respectively. The intra- and interassay coefficientsof variation were, respectively, 7.7% and 8.9% for estradiol, 7.2% and 8.9% forprogesterone, 7.4% and 9.8% for testosterone, and less than 10% and less than15% for cortisol.
ELISA of IGF1 and CRH. ELISA was conducted using a Mouse CRHELISA kit and Mouse IGF-1 ELISA kit (USA R&D Co.) purchased fromShanghai Quiqi Biological Technology Co., Ltd. Briefly, 50 ll of standards orsamples were added in duplicate to wells of a microtiter plate precoated withmouse monoclonal antibodies and incubated for 30 min at 378C. After the platewas washed using the wash solution and blot dried using paper towels, 50 ll ofEnzyme Conjugate (horseradish peroxidase-conjugated antibodies) were addedto each well, mixed thoroughly, and incubated for 30 min at 378C. Then, afterthe microtiter plate was washed and blot dried, 50 ll of Substrates A and Bwere added to each well and incubated for 15 min at 378C. The optical densitywas read at 450 nm using a plate reader (BioTek-ELx808, BioTek Instruments,Inc.) within 15 min after the reaction was terminated by adding 50 ll of theStop Solution. The concentrations of IGF1 and CRH in ovarian homogenatesand serum were calculated according to their respective standard curves.
Western Blot Analysis
Ovarian homogenization was performed using radioimmunoprecipitationassay (RIPA) buffer (150 mM NaCl, 1.0% Triton X-100, 0.5% sodiumdeoxycholate, 0.1% SDS, 1 mM phenylmethyl sulfonyl fluoride, and 50 mMTris; pH 8.0). For 75 mg of ovarian tissue, 1 ml of RIPA buffer was added.After the total protein concentration was determined with a BCA Protein AssayKit (P0012; Beyotime Institute of Biotechnology) and adjusted to 1 lg/ll, thesamples were subpackaged and frozen at �808C. CCs from approximately 60COCs or MGCs from approximately 30 follicles were placed in a 1.5-mlmicrofuge tube containing 20 ll of sample buffer (20 mM Hepes, 100 mMKCl, 5 mM MgCl
2, 2 mM dithiothreitol, 0.3 mM phenylmethyl sulfonyl
fluoride, and 3 lg/ml leupeptin; pH 7.5) and frozen at �808C. For proteinextraction, 5 ll of 53 SDS-PAGE loading buffer were added to each tube, andthe tubes were heated to 1008C for 5 min.
Total proteins were separated on a 12% polyacrylamide gel by SDS-PAGEand transferred electrophoretically onto polyvinylidene fluoride membranes.After being washed in TBST (150 mM NaCl, 2 mM KCl, 25 mM Tris, and0.05% Tween 20; pH 7.4) and blocked with TBST containing 3% bovine serumalbumin (BSA) for 2 h at 378C, the membranes were incubated at 48C overnightwith goat anti-CRH receptor 1 (CRHR1) polyclonal antibodies (1:400,ab59023; Abcam Co., Ltd.) and mouse anti-glyceraldehyde phosphatedehydrogenase (GAPDH) monoclonal antibodies (1:1000, CW0100; Kang-weishiji Biotechnology Co., Ltd.). Then, the membranes were washed in TBSTand incubated for 1.5 h at 378C with alkaline phosphatase-conjugated rabbit
FIG. 1. Mouse CC smears stained with Hoechst 33342 and observed under a fluorescence microscope. The heterochromatin is heavily stained with theHoechst dye and gives bright fluorescence. Whereas the apoptotic cells show pyknotic nuclei full of heterochromatin (red arrows), healthy cells shownormal nuclei with sparse heterochromatin spots. Smears of CCs collected from stressed mice with approximately 1% (A), 10% (B), and 30% (C) apoptoticcells before or after culture of COCs with or without SGH are shown. Original magnification 3400.
TABLE 2. Percentages of apoptotic CCs, metaphase II (MII) oocytes, and blastocysts after COCs from stressed and unstressed mice were cultured inmaturation medium with (þ) or without (�) SGH supplementation.
Mice
Apoptotic cumulus cells (%)* MII oocytes (%)� Blastocysts (%)z
Before culture þSGH �SGH þSGH �SGH þSGH �SGH
Control 0.6 6 0.3a 10.6 6 0.7a 22.7 6 0.4a 97.6 6 0.8a 95.5 6 0.7a 46.3 6 1.1a 25.3 6 1.3a
Stressed 1.4 6 0.5a 12.2 6 0.9a 31.7 6 0.5b 97.2 6 1.0a 94.4 6 1.0a 25.3 6 0.5b 11.9 6 0.8b
* Each treatment was repeated three times, with each replicate containing pooled CCs released from approximately 60 COCs.� Each treatment was repeated five or six times, with each replicate containing approximately 30 oocytes. Percentages of metaphase II oocytes werecalculated from COCs cultured.z Percentages of blastocysts were calculated from oocytes activated.a,b Values in the same column without a common letter differ significantly (P , 0.05).
RESTRAINT STRESS, CRH, AND OOCYTE COMPETENCE
3 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022
anti-goat immunoglobulin (Ig) G (1:1000, ZB-2311; Zhongshan Golden BridgeBiotechnology Co., Ltd.) and goat anti-mouse IgG (1:4000, CW0110;Kangweishiji Biotechnology Co., Ltd.). Finally, signals were detected by a5-bromo-4-chloro-3-indolyl phosphate/tetranitroblue tetrazolium chloride alka-line phosphatase color development kit (Beyotime Institute of Biotechnology).Relative quantities of proteins were determined with Image-Pro Plus (IPP)software (Media Cybernetics, Inc.) by analyzing the sum density of eachprotein band image. The relative quantities of CRHR1 in unstressed controlmice were arbitrarily set as one, and the values in stressed mice were expressedrelative to this quantity.
Immunohistochemistry
Detection of CRHR1 in ovarian sections. Ovaries were embedded inparaffin and sectioned serially at a thickness of 5 lm. Immunoperoxidasestaining was performed on deparaffinized sections using the Polink-2 plusPolymer HRP Detection System for Goat Primary Antibody (PV-9003;Zhongshan Golden Bridge Biotechnology Co., Ltd.). Sections were incubatedwith 3% peroxide for 15 min to exhaust endogenous peroxidase activity andwith 0.1% trypsin for 15 min at 378C for antigen retrieval. After being washedin PBS, the sections were incubated with goat polyclonal antibody to CRHR1(1:150, ab59023; Abcam Co., Ltd.) or PBS (for negative control) overnight at48C, Polymer Helper (Zhongshan Golden Bridge Biotechnology Co., Ltd.) for15 min at 378C, and then poly-horseradish peroxidase anti-goat IgG for 30 minat 378C. Sections were then immersed in diaminobenzidine solution for 2 minto develop color and counterstained with hematoxylin. Positive staining wasrevealed as brown spots and nonspecific staining of all cell nuclei as light blue.The relative quantity of CRHR1 on thecal cells was determined with the IPP
software by analyzing the sum density of the positive spots. Optical density andthe area were measured on three thecal segments of each antral follicle, and anaverage density:area ratio was then calculated to represent the amount ofCRHR1 in the follicle. The relative quantities of CRHR1 in unstressed controlmice were arbitrarily set as one, and the values in stressed mice were expressedrelative to this quantity.
Detection of CRH in oocytes. Oocytes were washed in M2 mediumbetween treatments. Cumulus-free oocytes were fixed with 4% paraformalde-hyde in PHEM buffer [43] for 30 min and then blocked in PHEM containing1% BSA and 100 mM glycine for 1 h. Blocked oocytes were incubated withrabbit anti-CRH polyclonal antibody (1:40, sc-6329; Santa Cruz Biotechnol-ogy, Inc.) at 48C overnight, Cy3-conjugated goat anti-rabbit IgG (1:600;Jackson ImmunoResearch) in 3% BSA in M2 medium for 1 h, and then 10 lg/ml of Hoechst 33342 in M2 medium for 10 min. Negative-control samples inwhich the primary antibody was omitted were also evaluated. The stainedoocytes were observed with a Leica laser-scanning confocal microscope (TCS,SP2; Leica Microsystems). Blue diode (405 nm) and helium/neon (543 nm)lasers were used to excite the Hoechst 33342 and Cy3, respectively.Fluorescence was detected with bandpass emission filters (Hoechst 33342,420–480 nm; Cy3, 560–605 nm). The relative content of CRH was quantifiedby measuring fluorescence intensities. For each experimental series, all imageswere acquired with identical settings. The relative intensities were measured onthe raw images using the IPP software under fixed thresholds across all slides.The average relative fluorescence of unstressed control oocytes was set to one,and the averages of stressed oocytes were expressed relative to this value.
Data Analysis
At least three replicates were performed for each treatment. Percentage datawere arc sine transformed and analyzed with ANOVA; a Duncan multiple-comparison test was used to locate differences. The Statistical Package forSocial Science software (version 11.5; SPSS, Inc.) was used. Data areexpressed as the mean 6 SEM, with P , 0.05 considered to be statisticallysignificant.
RESULTS
RS Triggered Apoptosis of CCs While Reducing OocyteDevelopmental Potential
When smears stained with Hoechst 33342 were observedunder a fluorescence microscope, healthy CCs showed normalnuclei with sparse heterochromatin spots, but apoptotic CCsshowed pyknotic nuclei full of heterochromatin (Fig. 1).Percentages of apoptotic CCs did not differ between controland stressed mice before or after culture with SGH butincreased significantly in stressed mice after culture withoutSGH (Table 2). Although percentages of metaphase II oocytesdid not differ, percentages of blastocysts were significantly
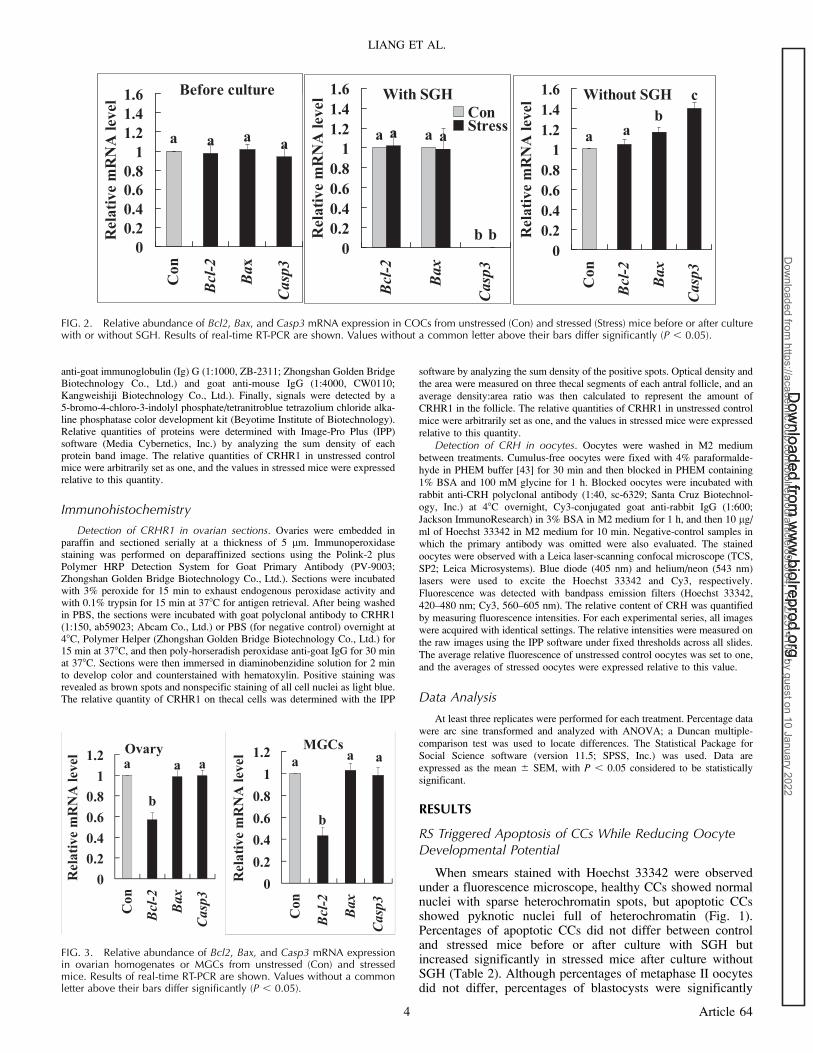
FIG. 2. Relative abundance of Bcl2, Bax, and Casp3 mRNA expression in COCs from unstressed (Con) and stressed (Stress) mice before or after culturewith or without SGH. Results of real-time RT-PCR are shown. Values without a common letter above their bars differ significantly (P , 0.05).
FIG. 3. Relative abundance of Bcl2, Bax, and Casp3 mRNA expressionin ovarian homogenates or MGCs from unstressed (Con) and stressedmice. Results of real-time RT-PCR are shown. Values without a commonletter above their bars differ significantly (P , 0.05).
LIANG ET AL.
4 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022

lower in stressed than in control mice after maturation with orwithout SGH.
The relative abundance of Casp3, Bax, and Bcl2 mRNA infreshly collected COCs did not differ between control andstressed mice (Fig. 2). After maturation with SGH, theabundance of Bax and Bcl2 mRNA did not differ betweencontrol and stressed mice, but Casp3 mRNA was undetectable.After maturation without SGH, the level of Bax and Casp3mRNA increased significantly in stressed mice. These resultssuggested that RS initiated a latent apoptotic program in CCsand oocytes that was executed later during IVM and impairedoocyte developmental potential and that SGH in culturemedium prevented apoptosis of CCs and oocytes by inhibitingexpression of CASP3 and BAX.
RS Induced Apoptosis in Ovarian Cells, Particularlyin MGCs
The relative abundance of Bax and Casp3 mRNA did notdiffer between control and stressed mice, but Bcl2 mRNAdecreased significantly in both ovarian homogenates andMGCs of stressed mice (Fig. 3). Furthermore, the decrease inBcl2 mRNA was more obvious in MGCs (43%) than in ovarianhomogenates (57%), suggesting that RS induced apoptosis firstin MGCs.
Effects of RS on Steroid Hormone Levels in Serum
and Ovaries
Following RS, concentrations of cortisol and progesteroneincreased but that of estradiol decreased significantly in bothserum and ovarian homogenates (Fig. 4). Interestingly, whereasthe level of testosterone in serum increased, that in the ovarydecreased significantly following RS.
Effects of RS on Ovarian Expression of IGF1 and Brain-
Derived Neurotrophic Factor
Concentrations of IGF1 in both serum and ovary decreasedsignificantly following RS (Fig. 5). Although the levels of Igf1and Bdnf mRNA in COCs were unchanged, those in MGCsdecreased significantly following RS (Fig. 5), suggesting thatRS decreased ovarian IGF1 and brain-derived neurotrophicfactor (BDNF) expression mainly by inhibiting MGCs.
RS Increased the CRH Level in Serum, Ovary, and Oocyte
Concentrations of CRH increased significantly in bothserum and ovary following RS (Fig. 6). Oocyte CRH occurredmainly as large spots distributed beneath the plasma membraneor near the germinal vesicle (Fig. 6). Quantification showedthat the relative level of CRH was significantly higher instressed than in control oocytes.
FIG. 4. Concentrations of cortisol, progesterone, estradiol, and testosterone in serum and ovarian homogenates from unstressed (Con) and stressed(Stress) mice. Values without a common letter above their bars differ significantly (P , 0.05).
RESTRAINT STRESS, CRH, AND OOCYTE COMPETENCE
5 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022

Localization and Quantification of CRHR1 in DifferentOvarian Tissues of Control and Stressed Mice
Positive signals for CRHR1 were highly detected in stromaland thecal cells of the antral follicles but were barely detectablein MGCs and CCs (Fig. 7). Quantification indicated that therelative level of CRHR1 in thecal cells increased significantlyfollowing RS. Western blot analysis showed that the relativeCRHR1 levels in CCs, MGCs, and ovarian homogenates wereall significantly higher in stressed than in control mice (Fig. 8).
CRH Impaired Oocyte Cytoplasmic Maturation whileIncreasing Apoptosis of CCs In Vitro
Oocytes from control and stressed mice were cultured for 24h in TCM-199 containing SGH in the presence or absence ofCRH and/or the CRHR1 antagonist antalarmin before beingactivated with SrCl
2for embryo development. In both control
and stressed mice, CRH reduced blastocyst rates significantlybut had no effect on cell number per blastocyst (Table 3).Supplementation of antalarmin eliminated the detrimentaleffect of CRH on oocyte developmental potential. CRH didnot show any effect on oocyte cytoplasmic maturation ofcumulus-denuded oocytes, indicating that its effect wasmediated by CCs. Antalarmin did not affect oocyte cytoplas-mic maturation in the absence of CRH, excluding its toxicityon oocytes. To observe the effect of CRH on apoptosis of CCs,oocytes were matured without SGH. The results (Table 3)showed that in both control and stressed mice, CRH increasedapoptotic percentages of CCs significantly and that antalarminovercame the proapoptotic effect of CRH completely.
Restraint stress elevated glucocorticoid levels by increasingCRH while decreasing ovarian expression of glucocorticoidreceptors (nuclear receptor subfamily 3, group C, member 1;NR3C1).
When stressed mice were injected with antalarmin, no effectwas observed at 24 h after injection, but cortisol decreased tothe level in the control mice at 3 h after injection (Table 4). Ourquantitative real-time PCR, however, showed that the relativelevels of ovarian Nr3c1 mRNA were lower significantly instressed (0.51 6 0.07) than in control mice (1.00 6 0.00).Results suggest that although RS elevated glucocorticoid levelsby increasing CRH, glucocorticoids may not act on the ovarydirectly.
DISCUSSION
The present results suggest that RS initiated a latentapoptotic program in CCs and oocytes during their develop-ment in the ovary, which was executed later during IVM toinduce apoptosis and impair oocyte developmental potential.Thus, although percentages of apoptotic CCs and the mRNAlevels of the proapoptotic Bax and Casp3 did not differbetween control and stressed mice before or after culture ofCOCs with SGH, they were significantly higher in stressedmice after culture without SGH. Further observations indicatedthat Bcl2 mRNA decreased significantly in both ovarianhomogenates and MGCs of stressed mice, and the decrease inBcl2 mRNA was more obvious in MGCs than in ovarianhomogenates. Although the levels of Igf1 and Bdnf mRNAs inCOCs were unchanged, those in MGCs decreased significantlyfollowing RS, suggesting that RS decreased ovarian growthfactor concentrations mainly by inhibiting MGCs. Becauseboth IGF1 and BDNF are antiapoptotic in the ovary [44], thepresent results suggest that RS induced apoptosis earlier inMGCs than in COCs. It is known that the first signs for atresiaof antral follicles are the presence of pyknotic nuclei indicativeof apoptosis in the membrana granulosa and antrum [45–49].The whole membrana granulosa and cumulus are thendestroyed. Finally, the follicular fluid is reabsorbed, and theoocyte is affected at the advanced stage of atresia [50].Furthermore, targeted expression of Bcl2 in mouse oocytesinhibits ovarian follicular atresia and prevents spontaneous andchemotherapy-induced oocyte apoptosis in vitro [51].
The present study demonstrated that RS increased concen-trations of cortisol and progesterone but decreased levels ofestradiol and testosterone significantly in the mouse ovary.
FIG. 5. Concentrations of IGF1 in ovaries or serum and relative mRNAlevels of Igf1 and Bdnf in COCs and MGCs from unstressed (Contl) andstressed (Stres) mice. Values without a common letter above their barsdiffer significantly (P , 0.05).
LIANG ET AL.
6 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022

Furthermore, RS also decreased IGF1 concentrations in theovary by inhibiting mRNA expression of Igf1 in MGCs anddown-regulated mRNA expression of Bdnf. In cattle, two of theearliest detectable differences between the healthy dominantfollicle selected for ovulation and the subordinate folliclesdestined to undergo atresia are the greater availability of IGFand the greater capacity to produce estradiol in the dominantfollicle [52]. In goats, the levels of estradiol and IGF1 weresignificantly lower in atretic follicles than in healthy follicles[38]. The level of progesterone, on the other hand, was higherin atretic follicles. Furthermore, another study showed thatIGF1 significantly reduced the apoptotic percentage of goatgranulosa cells cultured in vitro [53]. BDNF is a member of thenerve growth factor family that is important for neuronalsurvival and plasticity. Recently, BDNF has been found to beexpressed in ovarian tissues [54, 55], and evidence for a rolefor ovarian BDNF in oocyte development and maturation isincreasing [56–58]. Furthermore, a recent study has shown thatchronic unpredictable stress decreased the expression of BDNFin mouse ovaries [59]. Taken together, the data suggest that RSinduced apoptosis of follicular cells by upsetting the balancebetween estrogen and progesterone and by decreasing the
availability of IGF1 and BDNF. In addition, a decrease intestosterone may also induce apoptosis by activating caspases.It has been shown that testosterone inhibits apoptosis in theventral prostate by potentially targeting the transcriptionalactivity of effector caspase 3 and 6 genes as well as thecleavage of procaspase 3 and 6 into active enzymes [60]. Inutero exposure to the antiandrogen compound (flutamide)induced in rat testes a chronic apoptotic germ cell deathassociated with a long-term increase in the expression andactivation in germ cells of caspases 3 and 6 [61].
To study the mechanisms by which RS changes the balancebetween estrogen and progesterone and reduces the availabilityof IGF1, we observed that RS elevated the level of CRH inboth serum and ovarian homogenates and increased theexpression of CRHR1 in thecal cells and MGCs. CytoplasmicCRH has been detected in thecal and stromal cells and in cellswithin the corpora lutea in both rat [62] and human [63]ovaries. Our Western blot analysis showed marked expressionof CRHR1 proteins in both CCs and MGCs, but ourimmunohistochemistry of ovarian sections detected rarepositive signals of CRHR1 in these cells. Previous studiesalso showed that CRHR was not detectable in MGCs but was
FIG. 6. Concentrations of CRH in serum, ovarian homogenates, and oocytes from unstressed (Contl) and stressed (Stres) mice. Whereas CRHconcentrations in serum and ovarian homogenates were measured by ELISA, relative CRH levels in oocytes were determined by measuring fluorescenceintensities on laser-scanning confocal micrographs showing CRH distribution in control (A) and stressed (B) oocytes. Chromatin and CRH were pseudo-colored blue and red, respectively. Values without a common letter above their bars differ significantly (P , 0.05).
FIG. 7. Immunohistochemical localization of CRHR1 in the mouse ovary and its quantification in thecal cells. Positive CRHR1 staining appears as brownspots and nonspecific staining of nuclei as light blue. Positive signals for CRHR1 were highly detected in stromal cells (A) and in the thecal cellssurrounding the antral follicles (B) but were barely detectable in mural and cumulus granulosa cells (B). A graph showing the relative level of CRHR1 inthecal cells in control and stressed mice is also shown (C). Means without a common letter above their bars differ significantly (P , 0.05).
RESTRAINT STRESS, CRH, AND OOCYTE COMPETENCE
7 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022

detectable in ovarian stromal cells, thecal cells, and CCs [18,62, 64]. However, evidence suggests the occurrence of CRHRin granulosa cells. For example, the suppressive effects of CRHon estrogen and IGF1 release from rat granulosa and humangranulosa-luteal cells were antagonized by a selective CRHreceptor antagonist [35, 36].
Immobilization stress induced a marked expression ofCrhr1 mRNA in the stromal cells of rat ovaries [64]. CRHhas been shown to restrain the production of estrogen bycultured rat and human granulosa cells [35, 65]. It is knownthat estrogen is synthesized by granulosa cells from androgenproduced by thecal cells. Studies in rats demonstrated thatCRH acted via functional Leydig cell receptors to exertautocrine inhibitory actions on Leydig cell steroidogenesis [66,67]. Because the ovarian thecal cell is considered to be theembryological and functional equivalent of the testicularLeydig cell [68], it is reasonable to hypothesize that CRHplays a similar role in the androgen biosynthesis in the ovary.The reason for the increase in progesterone after RS is unclear,but a decrease in activity of an enzyme that degrades
progesterone (e.g., 20a- or 20b-hydroxysteroid dehydroge-nase) may increase progesterone concentrations [69]. In goats,although expression of mRNA encoding CYP11A1 (P450scc)was unaffected, that of mRNA encoding CYP17A1 (P450c17)decreased to the lowest level and that of mRNA encodingCYP19A1 (P450arom) became undetectable following atresia[70]. Follicular atresia in cattle [71] and sheep [72] was alsocharacterized by a loss of CYP19A1 in granulosa cells and adecrease in levels of CYP17A1 in the theca interna. Therefore,the present results showing that testosterone production wassignificantly decreased with increased expression of CRH andits receptors in thecal cells of the mouse ovary after RS supporthypotheses that CRH plays an inhibitory role in the androgenbiosynthesis in the ovary and that decreased activity ofenzymes that consume progesterone may lead to progesteroneaccumulation.
The present results indicate that IGF1 in mouse ovaries wasproduced mainly by MGCs. In the rat, IGF1 is produced byMGCs of the developing follicle [73]. In murine ovaries,besides a potent stimulatory effect on granulosa cell replication
FIG. 8. Relative levels of CRHR1 in CCs, MGCs, and ovarian homogenates (Ovary) of control (Contl) and stressed (Stres) mice. Results of Western blotanalysis are shown. The values of control mice were set as one, and the values of stressed mice were expressed relative to the control value. Meanswithout a common letter above their bars differ significantly (P , 0.05).
TABLE 3. Blastocyst development in vitro after COCs or denuded oocytes (DOs) from control or stressed mice were activated with Sr2þ followingmaturation in the presence or absence of CRH and/or antalarmin.
Mice CRH (M)* Antalarmin (M)* Blastocysts/activated oocytes (%) Cells per blastocyst (n) Apoptotic cumulus cells (%)
ControlCOCs 0 0 46.6 6 4.3d 45.7 6 1.5a 18.8 6 1.2a
10�6 0 23.6 6 5.9b 45.7 6 1.1a 29.2 6 1.9d
10�9 0 35.1 6 5.8c 45.8 6 1.0a —10�6 10�6 42.0 6 5.2cd 44.0 6 0.9a 16.5 6 1.4a
0 10�6 46.0 6 1.6d 43.9 6 1.1a 19.7 6 1.6ab
DOs 0 0 12.4 6 2.1a 33.2 6 0.6b —10�6 0 13.8 6 1.0a 34.9 6 1.2b —
StressedCOCs 0 0 23.4 6 2.1b 36.9 6 0.8b 27.6 6 1.7cd
10�6 0 6.9 6 1.2a 36.2 6 0.9b 36.5 6 2.8e
10�9 0 15.9 6 1.9ab 37.1 6 0.6b —10�6 10�6 19.8 6 1.3b 36.8 6 1.2b 26.0 6 1.2cd
0 10�6 27.9 6 4.6b 35.7 6 0.6b 23.6 6 1.5bc
* Each treatment was repeated four or five times, with each replicate containing approximately 30 oocytes. Neither maturation rates (ranging between92% and 93%) nor activation rates (ranging from 92% to 96%) of oocytes differed between treatments.a–d Values in the same column without a common letter differ significantly (P , 0.05).
LIANG ET AL.
8 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022

[74], IGF1 also exerts a stimulatory effect on aromatase
activity [75]. In addition, IGF1 exerts a synergistic role with
FSH on steroidogenesis of cultured rat granulosa cells [76].
Furthermore, CRH suppressed the release of both estrogen and
IGF1 in cultured rat granulosa cells, and this suppressive effect
was overcome by the addition of low doses of exogenous IGF1
[36]. Taken together, the data suggest that the inhibitory effect
of CRH on estrogen release was mediated by a suppression of
the autocrine/paracrine action of IGF1.
The present results also showed that the cortisol level inboth serum and ovary increased significantly after RS.Glucocorticoids were found to enhance FSH-stimulatedprogesterone synthesis in cultured granulosa cells of rats andcattle [77–79] and to suppress P450 aromatase activity anddecrease the number of LH receptors in rats [78, 80], cattle[79], and pigs [81]. Furthermore, porcine studies havedemonstrated that increased glucocorticoid concentrations candisrupt ovarian IGF1 synthesis and IGF action both in vitro andin vivo [82, 83]. On the other hand, glucocorticoids were foundto promote androgen production by cultured thecal cells incattle [84]. In the present study, CRH was higher in the ovarythan in the serum, whereas cortisol was lower in the ovary thanin the serum. This may help to explain why testosterone wasdecreased in the ovary while cortisol was increased after RS,because the effect of increased CRH might override the effectof cortisol, which was reported to promote androgenproduction by cultured thecal cells [84]. Furthermore, thepresent results show that RS decreased Nr3c1 mRNAexpression in the ovary. Taken together, the data suggest thatRS caused apoptosis of ovarian cells and impaired oocytecompetence mainly by increasing CRH but not glucocorticoids.
The present study also found that whereas the level oftestosterone decreased in the ovary, it increased significantly inserum following RS of female mice. Because this was differentfrom the situation reported for male rats in which bloodtestosterone was decreased after stress [85], we examinedtestosterone concentrations in male mice and found asignificant decrease following RS (data not shown). Studiesof testosterone levels under stress in women have reportedconflicting results; whereas some found general increases [86],others observed both significant increases and decreases [87].
TABLE 4. Serum cortisol concentrations of control and stressed mice atdifferent times after injection with or without antalarmin.
Treatment* Injection�Time after
injection/restraint (h) Cortisol (ng/ml)
Control — — 12.4 6 1.9a
Vehicle 24 13.2 6 1.7a
Vehicle 1 21.4 6 1.8b
Stressed — 24 40.7 6 4.3b
Vehicle 24 45.7 6 2.3b
Antalarmin 24 37.7 6 3.5b
Vehicle 1 38.3 6 0.8c
Antalarmin 1 26.6 6 1.0b
Antalarmin 3 14.9 6 0.7a
* Each treatment was repeated six to eight times, with each replicatecontaining a single mouse. Immediately before restraint treatment, micewere injected with antalarmin or an equivalent volume of vehicle, andserum was recovered for cortisol measurement at different times afterinjection or restraint treatment.� Antalarmin was dissolved first in ethanol and then in saline and wasadministered i.p. (20 mg/kg in a volume of 0.1 ml).a–c Values in the same column without a common letter differ significantly(P , 0.05).
FIG. 9. The possible pathways by which RS diminishes oocyte developmental competence by increasing expression of CRH and CRHR1 in differentovarian compartments, including thecal cells (THCs), MGCs, follicular fluid (FF), CCs, and the oocytes (OOs). E2, estradiol; G, glucocorticoid; P4,progesterone.
RESTRAINT STRESS, CRH, AND OOCYTE COMPETENCE
9 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022

In females, both the adrenal gland and the ovary contain thebiosynthetic pathways necessary for androgen synthesis andsecretion [88], so the present results suggest that the increasedserum testosterone after stress of female mice came from theadrenal gland. In both men and women, significantly elevatedlevels of testosterone, estradiol, androstenedione, and sexhormone-binding globulin were observed along with signifi-cantly increased ACTH and serum cortisol as a response toacute psychosocial stress [89]. In humans, the secretion ofandrogens and androgenic precursors is regulated by ACTH,analogous to the control of cortisol [90]. Concentrations ofdehydroepiandrosterone were significantly increased in womenbut not in men after performing bench presses [91].Furthermore, citalopram administration, which significantlyincreased overall levels of corticosterone, resulted in a greatercorticosterone response and greater adrenal weights in femalethan in male mice [92]. Taken together, these data suggest thatRS of female mice caused a hyperfunction of the adrenal gland,which increased the production of both cortisol and testoster-one.
We showed in the present study that RS increased both thelevel of CRH in mouse oocytes and the expression of CRHR1in CCs. Both the presence of CRH in oocytes and thelocalization of CRHR on CCs have been reported in rats andhumans [62, 63]. A recent study showed that apoptotic CCs, inwhich extra-long BCL-2 interacting mediator of cell death(BIM
EL) was up-regulated, accelerated oocyte aging and
degeneration in vitro via a paracrine action [93]. Furthermore,our supplementation of CRH and/or the CRHR1 antagonistantalarmin to IVM medium showed that in both control andstressed mice, CRH reduced blastocyst rates while increasingapoptotic percentages of CCs significantly and that supple-mentation of antalarmin eliminated the detrimental effect ofCRH on CCs and oocyte developmental potential. This furtherconfirms that CRH induces CC apoptosis and impairs oocytecompetence by acting on CRHR in CCs.
In the present study, when RS mice were injected withantalarmin, no effect was observed at 24 h after injection, butthe cortisol level decreased to that in the control mice whenobserved at 3 h after injection. This result further confirmedthat RS elevated glucocorticoid levels by increasing CRHsecretion, and it suggested that antalarmin was a short-timeantagonist for CRHR1. In fact, the in vivo effect of antalarminon glucocorticoid secretion has usually been measured shortlyafter injection [94–96], and to our knowledge, its effect at 24 hafter administration has not been reported.
To summarize, the possible pathways by which RSdiminishes oocyte developmental competence have beendepicted in Figure 9. RS impaired oocyte competence mainlyby increasing CRH but not glucocorticoids. Increased CRHinitiated a latent apoptotic program in CCs and oocytes duringtheir intraovarian development, which was executed laterduring IVM to impair oocyte competence. Thus, elevated CRHinteracted with increased CRHR on thecal cells and MGCs,reducing the production of testosterone, estrogen, and IGF1while increasing the level of progesterone. The imbalancebetween estrogen and progesterone and the decreased avail-ability of growth factors triggered apoptosis of MGCs andfacilitated CC expression of CRHR, which interacted with theoocyte-derived CRH later during IVM to induce CC apoptosisand reduce oocyte competence.
REFERENCES
1. Copper RL, Goldenberg RL, Das A, Elder N, Swain M, Norman G,Ramsey R, Cotroneo P, Collins BA, Johnson F, Jones P, Meier AM. Thepreterm prediction study: maternal stress is associated with spontaneous
preterm birth at less than thirty-five weeks’ gestation. Am J ObstetGynecol 1996; 175:1286–1292.
2. Csemiczky G, Landgren BM, Collins A. The influence of stress and stateanxiety on the outcome of IVF-treatment: psychological and endocrino-logical assessment of Swedish women entering IVF-treatment. Acta ObstetGynecol Scand 2000; 79:113–118.
3. Kee BS, Jung BJ, Lee SH. A study on psychological strain in IVF patients.J Assist Reprod Genet 2000; 17:445–448.
4. Klonoff-Cohen H, Chu E, Natarajan L, Sieber W. A prospective study ofstress among women undergoing in vitro fertilization or gameteintrafallopian transfer. Fertil Steril 2001; 76:675–687.
5. Schroder AK, Katalinic A, Diedrich K, Ludwig M. Cumulative pregnancyrates and drop-out rates in a German IVF programme: 4102 cycles in 2130patients. Reprod Biomed Online 2004; 8:600–606.
6. Neggers Y, Goldenberg R, Cliver S, Hauth J. The relationship betweenpsychosocial profile, health practices, and pregnancy outcomes. ActaObstet Gynecol Scand 2006; 85:277–285.
7. Pare WP, Glavin GB. Restraint stress in biomedical research: a review.Neurosci Biobehav Rev 1986; 10:339–370.
8. Glavin GB, Pare WP, Sandbak T, Bakke HK, Murison R. Restraint stressin biomedical research: an update. Neurosci Biobehav Rev 1994; 18:223–249.
9. Wiebold JL, Stanfield PH, Becker WC, Hillers JK. The effect of restraintstress in early pregnancy in mice. J Reprod Fertil 1986; 78:185–192.
10. Sugino N, Nakamura Y, Okuno N, Shimamura K, Teyama T, IshimatsuM, Kato H. Effects of restraint stress on luteal function in rats during mid-pregnancy. J Reprod Fertil 1994; 101:23–26.
11. Mairesse J, Lesage J, Breton C, Breant B, Hahn T, Darnaudery M,Dickson SL, Seckl J, Blondeau B, Vieau D, Maccari S, Viltart O. Maternalstress alters endocrine function of the feto-placental unit in rats. Am JPhysiol Endocrinol Metab 2007; 292:E1526–E1533.
12. Tsuma VT, Einarsson S, Madej A, Kindahl H, Lundeheim N, RojkittikhunT. Endocrine changes during group housing of primiparous sows in earlypregnancy. Acta Vet Scand 1996; 37:481–489.
13. Peltoniemi OA, Love RJ, Heinonen M, Tuovinen V, Saloniemi H.Seasonal and management effects on fertility of the sow: a descriptivestudy. Anim Reprod Sci 1999; 55:47–61.
14. Smith J, Ferguson D, Jauregui G, Panarace M, Medina M, Lehnert S, HillJR. Short-term maternal psychological stress in the post-conception periodin ewes affects fetal growth and gestation length. Reproduction 2008; 136:259–265.
15. Ebbesen SM, Zachariae R, Mehlsen MY, Thomsen D, Højgaard A,Ottosen L, Petersen T, Ingerslev HJ. Stressful life events are associatedwith a poor in-vitro fertilization (IVF) outcome: a prospective study. HumReprod 2009; 24:2173–2182.
16. Zhang SY, Wang JZ, Li JJ, Wei DL, Sui HS, Zhang ZH, Zhou P, Tan JH.Maternal restraint stress diminishes the developmental potential ofoocytes. Biol Reprod 2011; 84:672–681.
17. Zhou P, Lian HY, Cui W, Wei DL, Li Q, Liu YX, Liu XY, Tan JH.Maternal-restraint stress increases oocyte aneuploidy by impairingmetaphase I spindle assembly and reducing spindle assembly checkpointproteins in mice. Biol Reprod 2012; 86(3):83,1–14.
18. Kiapekou E, Zapanti E, Mastorakos G, Loutradis D. Update on the role ofovarian corticotropin-releasing hormone. Ann N Y Acad Sci 2010; 1205:225–229.
19. Whirledge S, Cidlowski JA. Glucocorticoids, stress, and fertility. MinervaEndocrinol 2010; 35:109–125.
20. Guelfi G, Zerani M, Brecchia G, Parillo F, Dall’Aglio C, Maranesi M,Boiti C. Direct actions of ACTH on ovarian function of pseudopregnantrabbits. Mol Cell Endocrinol 2011; 339:63–71.
21. Andersen CY. Effect of glucocorticoids on spontaneous and follicle-stimulating hormone induced oocyte maturation in mouse oocytes duringculture. J Steroid Biochem Mol Biol 2003; 85:423–427.
22. Jewgenow K, Heerdegen B, Muller K. In vitro development ofindividually matured bovine oocytes in relation to follicular wall atresia.Theriogenology 1999; 51:745–756.
23. Han ZB, Lan GC, Wu YG, Han D, Feng WG, Wang JZ, Tan JH.Interactive effects of granulosa cell apoptosis, follicle size, cumulus-oocyte complex morphology, and cumulus expansion on the develop-mental competence of goat oocytes: a study using the well-in-drop culturesystem. Reproduction 2006; 132:749–758.
24. Feng WG, Sui HS, Han ZB, Chang ZL, Zhou P, Liu DJ, Bao S, Tan JH.Effects of follicular atresia and size on the developmental competence ofbovine oocytes: a study using the well-in-drop culture system.Theriogenology 2007; 67:1339–1350.
25. McConkey DJ, Orrenius S, Okret S, Jondal M. Cyclic AMP potentiates
LIANG ET AL.
10 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022

glucocorticoid-induced endogenous endonuclease activation in thymo-cytes. FASEB J 1993; 7:580–585.
26. Ercan S, Ozer C, Tasx M, Erdogan D, Babul A. Effects of leptin on stress-induced changes of caspases in rat gastric mucosa. J Gastroenterol 2007;42:461–468.
27. Haack D, Luu H, Cho J, Chen MJ, Russo-Neustadt A. Exercise reverseschronic stress-induced Bax oligomer formation in the cerebral cortex.Neurosci Lett 2008; 438:290–294.
28. Jalalvand E, Javan M, Haeri-Rohani A, Ahmadiani A. Stress- and non-stress-mediated mechanisms are involved in pain-induced apoptosis inhippocampus and dorsal lumbar spinal cord in rats. Neuroscience 2008;157:446–452.
29. Dorscheid DR, Wojcik KR, Sun S, Marroquin B, White SR. Apoptosis ofairway epithelial cells induced by corticosteroids. Am J Respir Crit CareMed 2001; 164(10 pt 1):1939–1947.
30. Hareramadas B, Rembhotkar GW, Rai U. Glucocorticoid-inducedthymocyte apoptosis in wall lizard Hemidactylus flaviviridis. Gen CompEndocrinol 2004; 135:293–299.
31. Plotkin LI, Manolagas SC, Bellido T. Glucocorticoids induce osteocyteapoptosis by blocking focal adhesion kinase-mediated survival. Evidencefor inside-out signaling leading to anoikis. J Biol Chem 2007; 282:24120–24130.
32. Schlossmacher GA, Stevens A, White A. Glucocorticoid receptormediated apoptosis: mechanisms of resistance in cancer cells. J Endocrinol2011; 211(1):17–25.
33. Sasagawa I, Yazawa H, Suzuki Y, Nakada T. Stress and testicular germcell apoptosis. Arch Androl 2001; 47:211–216.
34. Jimena P, Castilla JA, Peran F, Ramirez JP, Vergara F Jr, Molina R,Vergara F, Herruzo A. Adrenal hormones in human follicular fluid. ActaEndocrinol (Copenh) 1992; 127:403–406.
35. Calogero AE, Burrello N, Negri-Cesi P, Papale L, Palumbo MA, Cianci A,Sanfilippo S, D’Agata R. Effects of corticotropin-releasing hormone onovarian estrogen production in vitro. Endocrinology 1996; 137:4161–4166.
36. Calogero AE, Barreca A, Burrello N, Palermo I, Giordano G, D’Agata R,Vicari E. Corticotrophin-releasing hormone inhibits insulin-like growthfactor-I release from primary cultures of rat granulosa cells. J Endocrinol2002; 174:493–498.
37. Jolly PD, Tisdall DJ, Heath DA, Lun S, McNatty KP. Apoptosis in bovinegranulosa cells in relation to steroid synthesis, cyclic adenosine 30,50-monophosphate response to follicle-stimulating hormone and luteinizinghormone, and follicular atresia. Biol Reprod 1994; 51:934–944.
38. Yu YS, Sui HS, Han ZB, Li W, Luo MJ, Tan JH. Apoptosis in granulosacells during follicular atresia: relationship with steroids and insulin-likegrowth factors. Cell Res 2004; 14:341–346.
39. Bencomo E, Perez R, Arteaga MF, Acosta E, Pena O, Lopez L, Avila J,Palumbo A. Apoptosis of cultured granulosa-lutein cells is reduced byinsulin-like growth factor I and may correlate with embryo fragmentationand pregnancy rate. Fertil Steril 2006; 85:474–480.
40. Zhang Y, Liu W, Ma C, Geng J, Li Y, Li S, Yu F, Zhang X, Cong B.Endoplasmic reticulum stress contributes to CRH-induced hippocampalneuron apoptosis. Exp Cell Res 2012; 318:732–740.
41. Hogan B, Costantini F, Lacy E. Manipulating the Mouse Embryo: ALaboratory Manual. Cold Spring Harbor, NY: Cold Spring HarborLaboratory Press; 1986.
42. Livak KJ, Schmittgen TD. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2�(DDCT) method. Methods 2001; 25:402–408.
43. Cui W, Zhang J, Lian HY, Wang HL, Miao DQ, Zhang CX, Luo MJ, TanJH. Roles of MAPK and spindle assembly checkpoint in spontaneousactivation and MIII arrest of rat oocytes. PLoS ONE 2012; 7:e32044.
44. Hussein MR. Apoptosis in the ovary: molecular mechanisms. HumReprod Update 2005; 11:162–177.
45. Byskov AG. Follicular atresia. In: Jones RE (ed.), The Vertebrate Ovary.New York: Plenum Press; 1978:533–562.
46. Hirshfield AN, Midgley AR Jr. Morphometric analysis of folliculardevelopment in the rat. Biol Reprod 1978; 19:597–605.
47. Kruip TA, Dieleman SJ. Macroscopic classification of bovine follicles andits validation by micromorphological and steroid biochemical procedures.Reprod Nutr Dev 1982; 22:465–473.
48. Blondin P, Dufour M, Sirard MA. Analysis of atresia in bovine folliclesusing different methods: flow cytometry, enzyme-linked immunosorbentassay, and classic histology. Biol Reprod 1996; 54:631–637.
49. Irving-Rodgers HF, van Wezel IL, Mussard ML, Kinder JE, Rodgers RJ.Atresia revisited: two basic patterns of atresia of bovine antral follicles.Reproduction 2001; 122:761–775.
50. Driancourt M. Follicular dynamics in sheep and cattle. Theriogenology1991; 35:55–79.
51. Morita Y, Perez GI, Maravei DV, Tilly KI, Tilly JL. Targeted expressionof Bcl-2 in mouse oocytes inhibits ovarian follicle atresia and preventsspontaneous and chemotherapy-induced oocyte apoptosis in vitro. MolEndocrinol 1999; 13:841–850.
52. Quirk SM, Cowan RG, Harman RM, Hu CL, Porter DA. Ovarian folliculargrowth and atresia: the relationship between cell proliferation and survival.J Anim Sci 2004; 82(suppl):E40–E52.
53. Yu Y, Li W, Han Z, Luo M, Chang Z, Tan J. The effect of follicle-stimulating hormone on follicular development, granulosa cell apoptosisand steroidogenesis and its mediation by insulin-like growth factor-I in thegoat ovary. Theriogenology 2003; 60:1691–1704.
54. Jensen T, Johnson AL. Expression and function of brain-derivedneurotrophin factor and its receptor, TrkB, in ovarian follicles from thedomestic hen (Gallus gallus domesticus). J Exp Biol 2001; 204(pt 12):2087–2095.
55. Seifer DB, Feng B, Shelden RM, Chen S, Dreyfus CF. Brain-derivedneurotrophic factor: a novel human ovarian follicular protein. J ClinEndocrinol Metab 2002; 87:655–659.
56. Martins da Silva SJ, Gardner JO, Taylor JE, Springbett A, De Sousa PA,Anderson RA. BDNF promotes bovine oocyte cytoplasmic competencefor embryo development. Reproduction 2005; 129:423–434.
57. Kawamura K, Kawamura N, Mulders SM, Sollewijn Gelpke MD, HsuehAJ. Ovarian BDNF promotes the development of oocytes into preimplan-tation embryos. Proc Natl Acad Sci U S A 2005; 102:9206–9211.
58. Yu Y, Yan J, Li M, Yan L, Zhao Y, Lian Y, Li R, Liu P, Qiao J. Effects ofcombined epidermal growth factor, brain-derived neurotrophic factor andinsulin-like growth factor-1 on human oocyte maturation and earlyfertilized and cloned embryo development. Hum Reprod 2012; 27:2146–2159.
59. Wu LM, Hu MH, Tong XH, Han H, Shen N, Jin RT, Wang W, Zhou GX,He GP, Liu YS. Chronic unpredictable stress decreases expression ofbrain-derived neurotrophic factor (BDNF) in mouse ovaries: relationshipto oocytes developmental potential. PLoS ONE 2012; 7(12):e52331.
60. Omezzine A, Mauduit C, Tabone E, Nabli N, Bouslama A, Benahmed M.Caspase-3 and �6 expression and activation are targeted by hormoneaction in the rat ventral prostate during the apoptotic cell death process.Biol Reprod 2003; 69:752–760.
61. Omezzine A, Chater S, Mauduit C, Florin A, Tabone E, Chuzel F, Bars R,Benahmed M. Long-term apoptotic cell death process with increasedexpression and activation of caspase-3 and �6 in adult rat germ cellsexposed in utero to flutamide. Endocrinology 2003; 144:648–661.
62. Mastorakos G, Webster EL, Friedman TC, Chrousos GP. Immunoreactivecorticotropin-releasing hormone and its binding sites in the rat ovary. JClin Invest 1993; 92:961–968.
63. Mastorakos G, Scopa CD, Vryonidou A, Friedman TC, Kattis D,Phenekos C, Merino MJ, Chrousos GP. Presence of immunoreactivecorticotropin-releasing hormone in normal and polycystic human ovaries.J Clin Endocrinol Metab 1994; 79:1191–1197.
64. Nappi RE, Rivest S. Stress-induced genetic expression of a selectivecorticotropin-releasing factor-receptor subtype within the rat ovaries: aneffect dependent on the ovulatory cycle. Biol Reprod 1995; 53:1417–1428.
65. Ghizzoni L, Mastorakos G, Vottero A, Barreca A, Furlini M, Cesarone A,Ferrari B, Chrousos GP, Bernasconi S. Corticotropin-releasing hormone(CRH) inhibits steroid biosynthesis by cultured human granulosa-luteincells in a CRH and interleukin-1 receptor-mediated fashion. Endocrinol-ogy 1997; 138:4806–4811.
66. Ulisse S, Fabbri A, Dufau ML. Corticotropin-releasing factor receptorsand actions in rat Leydig cells. J Biol Chem 1989; 264:2156–2163.
67. Fabbri A, Tinajero JC, Dufau ML. Corticotropin-releasing factor isproduced by rat Leydig cells and has a major local anti-reproductive rolein the testis. Endocrinology 1990; 127:1541–1543.
68. Erickson GF, Magoffin DA, Dyer CA, Hofeditz C. The ovarian androgenproducing cells: a review of structure/function relationships. Endocr Rev1985; 6:371–399.
69. Hsueh AJ, Adashi EY, Jones PB, Welsh TH Jr. Hormonal regulation of thedifferentiation of cultured ovarian granulosa cells. Endocr Rev 1984; 5:76–127.
70. Yuan JH, Wang JZ, Lan GC, Sui HS, Yu JN, Tan JH. Expression ofsteroidogenic enzymes and synthesis of steroid hormones duringdevelopment of ovarian follicles in prepubertal goats. Domest AnimEndocrinol 2008; 34:451–460.
71. Xu Z, Garverick HA, Smith GW, Smith MF, Hamilton SA, YoungquistRS. Expression of messenger ribonucleic acid encoding cytochrome P450side-chain cleavage, cytochrome P450 17 alpha-hydroxylase, and
RESTRAINT STRESS, CRH, AND OOCYTE COMPETENCE
11 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022

cytochrome P450 aromatase in bovine follicles during the first follicularwave. Endocrinology 1995; 136:981–989.
72. Huet C, Monget P, Pisselet C, Monniaux D. Changes in extracellularmatrix components and steroidogenic enzymes during growth and atresiaof antral ovarian follicles in the sheep. Biol Reprod 1997; 56:1025–1034.
73. Oliver JE, Aitman TJ, Powell JF, Wilson CA, Clayton RN. Insulin-likegrowth factor I gene expression in the rat ovary is confined to thegranulosa cells of developing follicles. Endocrinology 1989; 124:2671–2679.
74. Zhou J, Chin E, Bondy C. Cellular pattern of insulin-like growth factor-I(IGF-I) and IGF-I receptor gene expression in the developing and matureovarian follicle. Endocrinology 1991; 129:3281–3288.
75. Adashi EY, Resnick CE, Brodie AM, Svoboda ME, Van Wyk JJ.Somatomedin-C-mediated potentiation of follicle-stimulating hormone-induced aromatase activity of cultured rat granulosa cells. Endocrinology1985; 117:2313–2320.
76. Adashi EY, Resnick CE, Svoboda ME, Van Wyk JJ. Somatomedin-Csynergizes with follicle-stimulating hormone in the acquisition ofprogestin biosynthetic capacity by cultured rat granulosa cells. Endocri-nology 1985; 116:2135–2142.
77. Adashi EY, Jones PB, Hsueh AJ. Synergistic effect of glucocorticoids onthe stimulation of progesterone production by follicle-stimulating hormonein cultured rat granulosa cells. Endocrinology 1981; 109:1888–1894.
78. Schoonmaker JN, Erickson GF. Glucocorticoid modulation of follicle-stimulating hormone-mediated granulosa cell differentiation. Endocrinol-ogy 1983; 113:1356–1363.
79. Kawate N, Inaba T, Mori J. Effects of cortisol on the amounts of estradiol-17b and progesterone secreted and the number of luteinizing hormonereceptors in cultured bovine granulosa cells. Anim Reprod Sci 1993; 32:15–25.
80. Hsueh AJ, Erickson GF. Glucocorticoid inhibition of FSH-inducedestrogen production in cultured rat granulosa cells. Steroids 1978; 32:639–648.
81. Danisova A, Sebokova E, Kolena J. Effect of corticosteroids on estradioland testosterone secretion by granulosa cells in culture. Exp ClinEndocrinol 1987; 89:165–173.
82. Viveiros MM, Liptrap RM. Glucocorticoid influence on porcine granulosacell IGF-I and steroid hormone production in vitro. Theriogenology 1999;51:1027–1043.
83. Viveiros MM, Liptrap RM. ACTH treatment disrupts ovarian IGF-I andsteroid hormone production. J Endocrinol 2000; 164:255–264.
84. Spicer LJ, Chamberlain CS. Influence of cortisol on insulin- and insulin-
like growth factor 1 (IGF-1)-induced steroid production and on IGF-1receptors in cultured bovine granulosa cells and thecal cells. Endocrine1998; 9:153–161.
85. Orr TE, Mann DR. Role of glucocorticoids in the stress-inducedsuppression of testicular steroidogenesis in adult male rats. Horm Behav1992; 26:350–363.
86. Powell LH, Lovallo WR, Matthews KA, Meyer P, Midgley AR, Baum A,Stone AA, Underwood L, McCann JJ, Janikula Herro K, Ory MG.Physiologic markers of chronic stress in premenopausal, middle-agedwomen. Psychosom Med 2002; 64:502–509.
87. Kunstmann A, Christiansen K. Testosterone levels and stress in women:the role of stress coping strategies, anxiety and sex role identification.Anthropol Anz 2004; 62:311–321.
88. Longcope C. Adrenal and gonadal androgen secretion in normal females.Clin Endocrinol Metab 1986; 15:213–228.
89. Lennartsson AK, Kushnir MM, Bergquist J, Billig H, Jonsdottir IH. Sexsteroid levels temporarily increase in response to acute psychosocial stressin healthy men and women. Int J Psychophysiol 2012; 84:246–253.
90. Parker LN. Control of adrenal androgen secretion. Endocrinol Metab ClinNorth Am 1991; 20:401–421.
91. Le Panse B, Vibarel-Rebot N, Parage G, Albrings D, Amiot V, DeCeaurriz J, Collomp K. Cortisol, DHEA, and testosterone concentrationsin saliva in response to an international powerlifting competition. Stress2010; 13:528–532.
92. Goel N, Bale TL. Sex differences in the serotonergic influence on thehypothalamic- pituitary-adrenal stress axis. Endocrinology 2010; 151:1784–1794.
93. Wu Y, Wang XL, Liu JH, Bao ZJ, Tang DW, Wu Y, Zeng SM. BIMEL
-mediated apoptosis in cumulus cells contributes to degenerative changes inaged porcine oocytes via a paracrine action. Theriogenology 2011; 76:1487–1495.
94. Traslavina GA, Franci CR. The CRH-R1
receptor mediates luteinizinghormone, prolactin, corticosterone and progesterone secretion induced byrestraint stress in estrogen-primed rats. Brain Res 2011; 1421:11–19.
95. Lutfy K, Aimiuwu O, Mangubat M, Shin CS, Nerio N, Gomez R, Liu Y,Friedman TC. Nicotine stimulates secretion of corticosterone via bothCRH and AVP receptors. J Neurochem 2012; 120:1108–1116.
96. Bornstein SR, Webster EL, Torpy DJ, Richman SJ, Mitsiades N, Igel M,Lewis DB, Rice KC, Joost HG, Tsokos M, Chousos GP. Chronic effects ofa nonpeptide corticotropin-releasing hormone type I receptor antagonist onpituitary-adrenal function, body weight, and metabolic regulation.Endocrinology 1998; 139:1546–1555.
LIANG ET AL.
12 Article 64
Dow
nloaded from w
ww
.biolreprod.org. D
ownloaded from
ww
w.biolreprod.org.
Dow
nloaded from https://academ
ic.oup.com/biolreprod/article/89/3/64, 1-12/2514107 by guest on 10 January 2022
Copyright © 2022 FDOKUMEN




















![[sp1] oocyte collection from superstimulated disease-free ...](https://static.fdokumen.com/doc/165x107/631dd3361aedb9cd850f788f/sp1-oocyte-collection-from-superstimulated-disease-free-.jpg)