
Estrogen Receptor-α Knockout Mice Exhibit Resistance to the Developmental Effects of Neonatal...
15
Estrogen Receptor-a Knockout Mice Exhibit Resistance to the Developmental Effects of Neonatal Diethylstilbestrol Exposure on the Female Reproductive Tract John F. Couse,* , ² Darlene Dixon,‡ Mariana Yates,* Alicia B. Moore,‡ Liang Ma,§ ,1 Richard Maas,§ and Kenneth S. Korach* ,2 *Receptor Biology Section, Laboratory of Reproductive and Developmental Toxicology, National Institute of Environmental Health Sciences, National Institutes of Health, Research Triangle Park, North Carolina 27709; ²Department of Environmental and Molecular Toxicology, North Carolina State University, Raleigh, North Carolina 27695; ‡Comparative Pathobiology Section, Laboratory of Experimental Pathology, National Institute of Environmental Health Sciences, National Institutes of Health, Research Triangle Park, North Carolina 27709; and §Division of Genetics, Department of Medicine, Brigham and Women’s Hospital, Harvard Medical School and Howard Hughes Medical Institute, Boston, Massachusetts 02115 Data indicate that estrogen-dependent and -independent pathways are involved in the teratogenic/carcinogenic syndrome that follows developmental exposure to 17b-estradiol or diethylstilbestrol (DES), a synthetic estrogen. However, the exact role and extent to which each pathway contributes to the resulting pathology remain unknown. We employed the aERKO mouse, which lacks estrogen receptor-a (ERa), to discern the role of ERa and estrogen signaling in mediating the effects of neonatal DES exposure. The aERKO provides the potential to expose DES actions mediated by the second known ER, ERb, and those that are ER-independent. Wild-type and aERKO females were treated with vehicle or DES (2 mg/pup/day for Days 1–5) and terminated after 5 days and 2, 4, 8, 12, and 20 months for biochemical and histomorphological analyses. Assays for uterine expression of the genes Hoxa10, Hoxa11, and Wnt7a shortly after treatment indicated significant decreases in DES-treated wild-type but no effect in the aERKO. In contrast, the DES effect on uterine expression of Wnt4 and Wnt5a was preserved in both genotypes, suggesting a developmental role for ERb. Adult aERKO mice exhibited complete resistance to the chronic effects of neonatal DES exposure exhibited in treated wild-type animals, including atrophy, decreased weight, smooth muscle disorganization, and epithelial squamous metaplasia in the uterus; proliferative lesions of the oviduct; and persistent vaginal cornification. Therefore, the lack of DES effects on gene expression and tissue differentiation in the aERKO provides unequivocal evidence of an obligatory role for ERa in mediating the detrimental actions of neonatal DES exposure in the murine reproductive tract. © 2001 Academic Press Key Words: estrogen receptor; diethylstilbestrol; teratogenesis; uterus; Hox; Wnt. INTRODUCTION Laboratory and epidemiological studies have clearly im- plicated 17b-estradiol (E 2 ) and diethylstilbestrol (DES), en- dogenous and synthetic estrogens, respectively, as carcino- gens and teratogens in humans and laboratory animals (IARC, 1979, 1987; Marselos and Tomatis, 1992, 1993). Beginning in the 1940s, DES was prescribed during preg- nancy to maintain placental steroid production and thereby 1 Present address: Tulane University, Department of Cell and Molecular Biology, 2000 Percival Stern Hall, 6400 Freret Street, New Orleans, LA 70118. 2 To whom correspondence should be addressed at National Institutes of Health, National Institute of Environmental Health Sciences, Laboratory of Reproductive and Developmental Toxicol- ogy, Receptor Biology Section, MD B3-02, P.O. Box 12233, Re- search Triangle Park, NC 27709. Fax: (919) 541-0696. E-mail: [email protected]. Developmental Biology 238, 224 –238 (2001) doi:10.1006/dbio.2001.0413, available online at http://www.idealibrary.com on 0012-1606/01 $35.00 Copyright © 2001 by Academic Press All rights of reproduction in any form reserved. 224
Transcript of Estrogen Receptor-α Knockout Mice Exhibit Resistance to the Developmental Effects of Neonatal...

L
NRDS
Df
l
agiaeso
Developmental Biology 238, 224–238 (2001)doi:10.1006/dbio.2001.0413, available online at http://www.idealibrary.com on
Estrogen Receptor-a Knockout Mice ExhibitResistance to the Developmental Effectsof Neonatal Diethylstilbestrol Exposureon the Female Reproductive Tract
John F. Couse,* ,† Darlene Dixon,‡ Mariana Yates,* Alicia B. Moore,‡iang Ma,§ ,1 Richard Maas,§ and Kenneth S. Korach* ,2
*Receptor Biology Section, Laboratory of Reproductive and Developmental Toxicology,National Institute of Environmental Health Sciences, National Institutes of Health,Research Triangle Park, North Carolina 27709; †Department of Environmental andMolecular Toxicology, North Carolina State University, Raleigh, North Carolina27695; ‡Comparative Pathobiology Section, Laboratory of Experimental Pathology,
ational Institute of Environmental Health Sciences, National Institutes of Health,esearch Triangle Park, North Carolina 27709; and §Division of Genetics,epartment of Medicine, Brigham and Women’s Hospital, Harvard Medicalchool and Howard Hughes Medical Institute, Boston, Massachusetts 02115
ata indicate that estrogen-dependent and -independent pathways are involved in the teratogenic/carcinogenic syndrome thatollows developmental exposure to 17b-estradiol or diethylstilbestrol (DES), a synthetic estrogen. However, the exact role and
extent to which each pathway contributes to the resulting pathology remain unknown. We employed the aERKO mouse, whichacks estrogen receptor-a (ERa), to discern the role of ERa and estrogen signaling in mediating the effects of neonatal DES
exposure. The aERKO provides the potential to expose DES actions mediated by the second known ER, ERb, and those that areER-independent. Wild-type and aERKO females were treated with vehicle or DES (2 mg/pup/day for Days 1–5) and terminatedfter 5 days and 2, 4, 8, 12, and 20 months for biochemical and histomorphological analyses. Assays for uterine expression of theenes Hoxa10, Hoxa11, and Wnt7a shortly after treatment indicated significant decreases in DES-treated wild-type but no effectn the aERKO. In contrast, the DES effect on uterine expression of Wnt4 and Wnt5a was preserved in both genotypes, suggestingdevelopmental role for ERb. Adult aERKO mice exhibited complete resistance to the chronic effects of neonatal DES exposurexhibited in treated wild-type animals, including atrophy, decreased weight, smooth muscle disorganization, and epithelialquamous metaplasia in the uterus; proliferative lesions of the oviduct; and persistent vaginal cornification. Therefore, the lackf DES effects on gene expression and tissue differentiation in the aERKO provides unequivocal evidence of an obligatory role
for ERa in mediating the detrimental actions of neonatal DES exposure in the murine reproductive tract. © 2001 Academic Press
Key Words: estrogen receptor; diethylstilbestrol; teratogenesis; uterus; Hox; Wnt.
dg(Bn
INTRODUCTION
Laboratory and epidemiological studies have clearly im-plicated 17b-estradiol (E2) and diethylstilbestrol (DES), en-
1 Present address: Tulane University, Department of Cell andMolecular Biology, 2000 Percival Stern Hall, 6400 Freret Street,New Orleans, LA 70118.
2 To whom correspondence should be addressed at NationalInstitutes of Health, National Institute of Environmental HealthSciences, Laboratory of Reproductive and Developmental Toxicol-
224
ogenous and synthetic estrogens, respectively, as carcino-ens and teratogens in humans and laboratory animalsIARC, 1979, 1987; Marselos and Tomatis, 1992, 1993).eginning in the 1940s, DES was prescribed during preg-ancy to maintain placental steroid production and thereby
ogy, Receptor Biology Section, MD B3-02, P.O. Box 12233, Re-search Triangle Park, NC 27709. Fax: (919) 541-0696. E-mail:[email protected].
0012-1606/01 $35.00Copyright © 2001 by Academic Press
All rights of reproduction in any form reserved.

(rm(
a
ctaE
mms(ala(tlstdtmeenf
Zm
tmmgaivtmEtiad(
amd
ioInmw
225Effects of DES in the aERKO Female Mouse
lessen the risk of spontaneous abortion or preterm parturi-tion (Marselos and Tomatis, 1992). Despite a 1953 report ofthe ineffectiveness of DES for this indication (Dieckmannet al., 1953), clinical use increased in the years following(Marselos and Tomatis, 1992). In 1971, following two re-ports of a causal link between vaginal clear-cell adenocar-cinoma, a rare cancer, and in utero DES exposure (Green-wald et al., 1971; Herbst et al., 1971), the U.S. FDAproscribed the use of DES for pregnancy support (Herbst,2000). To date, DES remains the only drug in whichtransplacental carcinogenesis in man has been proven(Marselos and Tomatis, 1992). More prevalent than cancerare the multiple teratogenic effects attributed to in uteroDES exposure, including vaginal/cervical epithelial adeno-sis and squamous metaplasia, transverse vaginal ridges, andstructural malformations of the cervix and uterus (Herbst,2000; Marselos and Tomatis, 1992; Mittendorf, 1995). Inmales, in utero DES exposure is associated with increasedrisk of testicular cancer, epididymal cysts, cryptorchidism,and testicular hypoplasia (Herbst, 2000; Marselos andTomatis, 1992).
Over the past 30 years, rodent models that effectivelyassimilate the effects of developmental DES exposure inhumans have been thoroughly characterized, yet the under-lying mechanisms remain poorly understood (Marselos andTomatis, 1992, 1993). It is widely accepted that E2 or DESexposure during critical developmental periods elicits dis-tinct and permanent alterations in the Mullerian-derivedstructures of the female reproductive tract (oviduct, uterus,cervix, upper vagina) (Cunha et al., 1991). Interestingly, nullmouse models have illustrated that a lack of estrogensignaling has little effect on reproductive tract development(Couse et al., 1999; Couse and Korach, 1999; Fisher et al.,1998), whereas aberrant estrogen exposure during develop-ment is detrimental (Cunha et al., 1991). Hence, the under-lying mechanisms of developmental DES exposure likelyinvolve its ability to bind the nuclear estrogen receptor (ER)and mimic the broad spectrum of E2 actions (Korach et al.,1978, 1989). This aberrant stimulation of estrogen signal-ing by DES during development may disrupt the properexpression of estrogen-regulated genes and the ultimatedifferentiation/proliferation profile of cell populations. Re-ported alterations in the expression of Hoxa9, Hoxa10,Hoxa11 (Block et al., 2000; Ma et al., 1998), and Wnt7aMiller et al., 1998a) in the developing murine femaleeproductive tract following DES exposure support such aechanism. Furthermore, null mouse models of the Hox
Dolle et al., 1991; Hsieh-Li et al., 1995; Satokata et al.,1995) and Wnt (Miller et al., 1998a; Parr and McMahon,1998) gene families exhibit reproductive tract phenotypesthat are comparable to those elicited by developmental DESexposure. These data support disruption of developmentalgene expression as a mechanism of DES; however, it re-mains unknown whether this action is dependent on ER.Further complexity has been introduced by two recentdiscoveries: (1) a second nuclear ER (ERb) that exhibits E2-nd DES-induced in vitro activity comparable to that of the
Copyright © 2001 by Academic Press. All right
lassical ER (now termed ERa) (Giguere et al., 1998); and (2)he unique ability of DES to suppress the transactivationalctivity of the orphan nuclear receptors ERRa, ERRb, andRRg (Tremblay et al., 2001).Laboratory studies have also indicated that epigeneticechanisms, independent of direct interaction with geneticaterial but leading to permanent changes in gene expres-
ion, may also contribute to the ultimate effects of DESLiehr, 2000). Perinatal DES exposure in rodents causesttenuated uterine responses to subsequent estrogen stimu-ation during adulthood, characterized as decreased growthnd secretory activity and altered cellular differentiationMaier et al., 1985; Medlock et al., 1988, 1992). Similar DESreatments in mice lead to constitutive up-regulation ofactoferrin expression in adult uteri (Nelson et al., 1994),eminal vesicle (Beckman et al., 1994), and the urethropros-atic complex (Salo et al., 1997). Moreover, Li et al. (1997)emonstrated distinct alterations in the methylation pat-ern of the lactoferrin gene promoter in uterine tissue ofice neonatally exposed to DES, providing support for an
pigenetic pathway by which DES permanently alters genexpression. Similar changes in gene expression followingeonatal DES exposure are reported for epidermal growthactor (EGF), EGF-receptor, c-fos, c-jun, c-myc, bax, and
bcl-2 in the reproductive tract of sexually mature mice andhamsters (Falck and Forsberg, 1996; Nelson et al., 1994;
heng and Hendry, 1997). Once again, the role of the ER inediating this effect of DES remains unknown.In contrast to the hormonal and epigenetic mechanisms,
here is evidence of direct genotoxic actions of DES and itsetabolites, presumably independent of ER action. Theseechanisms are more often applied to explain the carcino-
enic properties of DES and include the induction of DNAdducts, microsatellite instability, sequence deletions ornsertions, and single-strand breaks in both in vitro and inivo systems (Liehr, 2000). Furthermore, DES and its me-abolites are reported to elicit aneuploidy and cell transfor-ation in Syrian hamster embryo cells lacking measurable
R levels, possibly via interfering with microtubule func-ion (Tsutsui et al., 1983). Supporting evidence of DES-nduced genetic instability is provided by descriptions ofneuploid DNA in premalignant vaginal lesions in “DESaughters” (Welch et al., 1983) and DES-exposed miceHajek et al., 1993).
Therefore, existing evidence supports both ER-dependentnd -independent mechanisms as mediators of the develop-ental and carcinogenic actions of DES. In this report, we
escribe our use of the estrogen receptor-a knockout mouse(aERKO), which lacks functional ERa, to gain furthernsight into the role of this receptor in mediating the effectsf neonatal DES exposure in the female reproductive tract.n brief, our results demonstrate that adult wild-type miceeonatally exposed to DES exhibit the previously describedorphological defects throughout the reproductive tract,hereas their aERKO littermates do not. Furthermore, the
DES-elicited suppression of Hoxa10, Hoxa11, and Wnt7aexpression exhibited in the uteri of treated wild-type mice
s of reproduction in any form reserved.

h
De
Faru2
CG
CbB
tmc
omCc
2amebta
wa
226 Couse et al.
shortly after treatment was absent in the aERKO uteri.These data provide unequivocal evidence of an obligatoryrole for ERa in mediating the effects of neonatal DESexposure in the murine female reproductive tract.
MATERIALS AND METHODS
Generation and Treatment of Animals
All procedures were performed in accordance with the NIHGuide for the Care and Use of Laboratory Animals and werepreapproved by the NIEHS ICACU. All animals were maintained inplastic cages under a 12-h light/12-h dark schedule in atemperature-controlled room (21–22°C), fed NIH 31 mouse chowand fresh water ad libutum. Mice were generated by breeding
eterozygous (ERa1/2) animals in a continuous mating scheme.Pregnant mice were single-housed and litters adjusted to 4–8 pupsper dam upon delivery. Pups were treated with DES (Sigma Chemi-cals, St. Louis, MO) dissolved in corn oil at 2 mg/pup/day (s.c.) for
ays 1–5 (day of birth 5 1); controls were similarly treated with anqual volume (20 ml) of corn oil. Mice were weaned at 21–22 days
of age and housed 2–4 per cage as described above. Prior to 60 days,mice were implanted with an implantable microidentification unit(IMI-1000; BioMedic Data Systems, Seaford, DE) and a tail biopsycollected for genomic DNA extraction as previously described(Couse et al., 1994). Mice were selected and terminated at fourpredetermined ages designated as I (4–5 months), II (8–9 months),III (12–13 months), and IV (18–22.7 months). On termination,whole blood was collected and heparinized and the plasma wasstored at 270°C; all tissues were fixed in 10% buffered formalin.or gene expression analyses, animals were terminated at 5 days ofge (6 h after the final injection), and at 2–3 months of age, theeproductive tract was removed quickly and the ovary, oviduct,terus, and vagina were separated, snap-frozen, and stored at70°C. Animals were ERa-genotyped via polymerase chain reac-
tion (PCR) on genomic DNA using the following primers fordetection of an untargeted ERa gene (forward) 59-CGGTCTA-
GGCCAGTCGGGCATC, (reverse) 59-CAGGCCTTACACAGC-GCCACCC (281-bp amplimer from wild-type ERa exon 2); and
for detection of the gene disruption (forward) 59-GTGTTCC-GGCTGTCAGCGCA, (reverse) 59-GTCCTGATAGCGGTCCG-CCA (555-bp amplimer).
Cyto- and Immunohistochemistry
Formalin-fixed tissues were paraffin-embedded and 6 mm sec-tions mounted on ProbeOn Plus slides (Fisher Scientific, Pitts-burgh, PA). Sections were stained with hematoxylin and eosin(H&E) for routine light microscope evaluation. Immunohistochem-istry for ERa was performed as follows. Following standard depar-affinization and hydration protocol, endogenous peroxidase wasquenched by incubation in 3% H2O2 and antigen retrieval carriedout in 13 citrate buffer in a decloaking chamber (BioCare Medical,Walnut Creek, CA) according to the manufacturer’s protocol. Allwashes were carried out in 13 Automation Buffer (AB) (BiomedaCorporation, Foster City, CA). Sections were blocked with 5%normal horse serum (Jackson ImmunoResearch Laboratories, WestGrove, PA) followed by incubation with a 1:50 dilution of mono-clonal anti-ERa antibody (ER1D5) for 1 h (Beckman Coulter, Brea,
A), washed in 13 AB, and incubated with a 1:500 dilution ofiotinylated horse anti-mouse IgG antibody (Vector Laboratories,urlingame, CA). Specific immunoreactivity was detected using
Copyright © 2001 by Academic Press. All right
he Vectastain ABC Kit (Vector Laboratories) according to theanufacturer’s protocol using 3,39-diaminobenzidine tetrahydro-
hloride as a chromogen, followed by counterstain with hematoxylin.
Cloning of the Wnt4 and Wnt5a cDNAProbe Templates
Riboprobe templates for the mouse Wnt4 and Wnt5a consistedf a reverse transcription–PCR generated (RT-PCR) cDNA frag-ent cloned into the pCRII-TOPO vector (Invitrogen, Carlsbad,A) according to the manufacturer’s protocol. The Wnt4 templateonsisted of 426 bp of the mouse Wnt4 cDNA (bp 230–655,
GenBank Accession Number M89797) amplified from C57BL/6ovary RNA using the following primers: (forward) 59-AGG-TGATGGACTCAGTGCGC, (reverse) 59-CCCGCATGTGTGTC-AAGATG. The Wnt5a template consisted of 253 bp of the mouseWnt5a cDNA (bp 609–861, GenBank Accession Number M89798)amplified from C57BL/6 lung RNA using the following primers:(forward) 59-CCATGTCTTCCAAGTTCTTCC, (reverse) 59-CAT-TCCTTGATGCCTGTCTTC. All clones were confirmed by se-quence analysis.
Total RNA Isolation, RNase Protection Assay,and in Situ Hybridization
Total RNA was isolated using Trizol reagent (Life Technologies,Rockville, MD) according to the manufacturer’s protocol. Concen-trations of the final preparations were calculated from an A260
reading (Beckman DU-7 spectrophotometer) and an aliquot ana-lyzed by gel electrophoresis to ensure integrity. Radiolabeledantisense riboprobes for ribonuclease protection assays (RPAs)were generated from linearized vectors using the Ambion Maxi-Script Kit (Ambion, Austin, TX) and [32P]CTP (Amersham Pharma-cia Biotech, Piscataway, NJ) according to the manufacturer’s pro-tocol. Wnt4 and Wnt5a antisense riboprobes were generated fromthe templates described herein; Hoxa10 and Hoxa11 antisenseriboprobes were described previously (Benson et al., 1995; Hsieh-Liet al., 1995; Ma et al., 1998). The Hoxa10 antisense riboprobedifferentially detects the splice variants Hoxa10-1 and Hoxa10-2(Benson et al., 1995). A cyclophilin antisense riboprobe was gener-ated from the pTRI–cyclophilin template (Ambion) and included inall assays for normalization. Sizes (nt) of the respective protectedfragments were as follows: cyclophilin 5 103, Wnt4 5 426,Wnt5a 5 253, Hoxa10-1 5 316, Hoxa10-2 5 146, and Hoxa 11 537. RPAs were carried out on total RNA from tissue of individualnimals using Hybspeed RPA reagents (Ambion) according to theanufacturer’s protocol. All RPA samples were fractionated by
lectrophoresis on precast 6% bis-acrylamide/7 M urea/13 Tris–orate gels (Invitrogen), which were then fixed, dried, and exposedo a phosphorimager screen followed by analysis with a Storm 860nd ImageQuant Software (Molecular Dynamics, Sunnyvale, CA).
In situ hybridization for Hoxa10 mRNA in the neonatal uterias carried out as previously described (Ma et al., 1998), using the
bove-described antisense riboprobe labeled with digoxigenin.
Semiquantitative RT-PCR for Wnt7a
Repeated assays indicated the levels of Wnt7a mRNA in theneonatal uterus to be below the levels of reproducible detection byRPA. Therefore, the following semiquantitative RT-PCR protocolwas carried out for these analyses. PCR for Wnt7a and b-actintranscripts was simultaneously carried out on cDNA preparationsfrom total RNA from tissue of individual animals. Primers for
s of reproduction in any form reserved.

ppn
ptcaBc
SM(cqsfg5A5Co
mF6wy(
W
WcwDtdcHtat
227Effects of DES in the aERKO Female Mouse
Wnt7a mRNA detection amplified a 302-bp fragment (bp 539–840,GenBank Accession Number M89801): (forward) 59-TGAACTT-ACACAATAACGAGGC, (reverse) 59-CTCTTCACAGTAATT-GGGTGAC. Primers for detection of the mouse b-actin mRNAwere purchased from Clontech (Palo Alto, CA). RT generation ofcDNA was carried in a reaction of 2.5 mM random hexamers(Applied Biosystems, Foster City, CA), 0.2 mM (d)-NTPs each(Invitrogen), 25 U RNase inhibitor (Applied Biosystems), 62.5 UMuLV reverse transcriptase (Applied Biosystems), and 0.5 mg oftotal RNA in a volume of 25 ml. Semiquantitative PCR reactionsfor each cDNA preparation were prepared as follows: 60 ml reactionconsisting of 13 Optimized PCR buffer F (Invitrogen), 8.0 ml cDNAreparation, 50 pmol of each Wnt7a primer, 5 pmol of each b-actinrimer, 0.2 mM each deoxy (d)-NTPs (Invitrogen), and 3 U Plati-umTaq DNA polymerase (Life Technologies). This was then
aliquoted at 15 ml per reaction to generate four identical reactionser cDNA preparation, one for each of the four PCR terminationime points (20, 25, 30, and 35 cycles). Thermal cycling conditionsonsisted of an initial 95°C/2 min, 95°C/30 s, 58°C/45 s, 72°C/30 s,nd 72°C/7 min in a GeneAmp 9600 Thermal Cycler (Appliediosystems). This process was carried out in duplicate for eachDNA preparation.
All samples were electrophoresed in a gel of 13 TBE/2% Nu-eive Agarose/0.7% SeaKem Agarose (FMC Bioproducts, Rockland,E), which was then transferred to BrightStar Nylon membrane
Ambion) according the manufacturer’s protocol. All blots were UVrosslinked (UV Stratalinker 1800; Stratagene, La Jolla, CA). Foruantitation, blots were probed with oligos complementary toequences nested within the RT-PCR primers and consisted of theollowing oligos and their complements, which when annealedenerated flanking 10-bp overlaps (indicated by “/”): Wnt7a,9-GTCGAAACAA/GCGGCCCACCTTTCTGAAGA/TCAAGA-GCC (bp 731–770, GenBank Accession Number M89801); b-actin,9-GCCCATCTAC/GAGGGCTATGCTCTCCCTCA/CGCCATC-TG (bp 411–450, GenBank Accession Number M12481). Theverlapping sequences were filled-in in the presence of [32P]dCTP
(Amersham Pharmacia Biotech) using Klenow DNA polymerase(Life Technologies) according the manufacturer’s protocol. Allhybridizations were carried out for 12–18 h at 55°C in a solution of13 SSC, 0.2 M NaH2PO4, 53 Denhardt’s solution, 0.1% SDS, 0.5
g/ml salmon-sperm DNA, and .1.0 3 108 cpm of each probe.ollowing hybridization, blots were washed at room temperature in3 SCC followed by 0.5% SDS/0.13 SSC at 55°C at 30 min perash; then exposed to a phosphorimager screen followed by anal-sis with a Storm 860 and accompanying ImageQuant SoftwareMolecular Dynamics).
Plasma E2 Radioimmunoassay (RIA)
Serum E2 was assayed on 200 ml aliquots per animal using theultrasensitive 17b-estradiol double-antibody [125I]RIA (DiagnosticSystems Laboratories, Webster, TX) according to the manufactur-er’s protocol and counted on a Packard Multi-Prias 2 GammaCounter.
Statistical Analysis
All data sets were first tested for homoscedasticity of varianceusing the Levine’s test. In cases where data were found to lackhomoscedasticity of variance (P , 0.05), all data were log-transformedprior to further statistical analysis. In all cases, data sets were analyzedby two-way ANOVA followed by individual post hoc comparisons.Statistical significance was assigned at P , 0.05.
Copyright © 2001 by Academic Press. All right
RESULTS
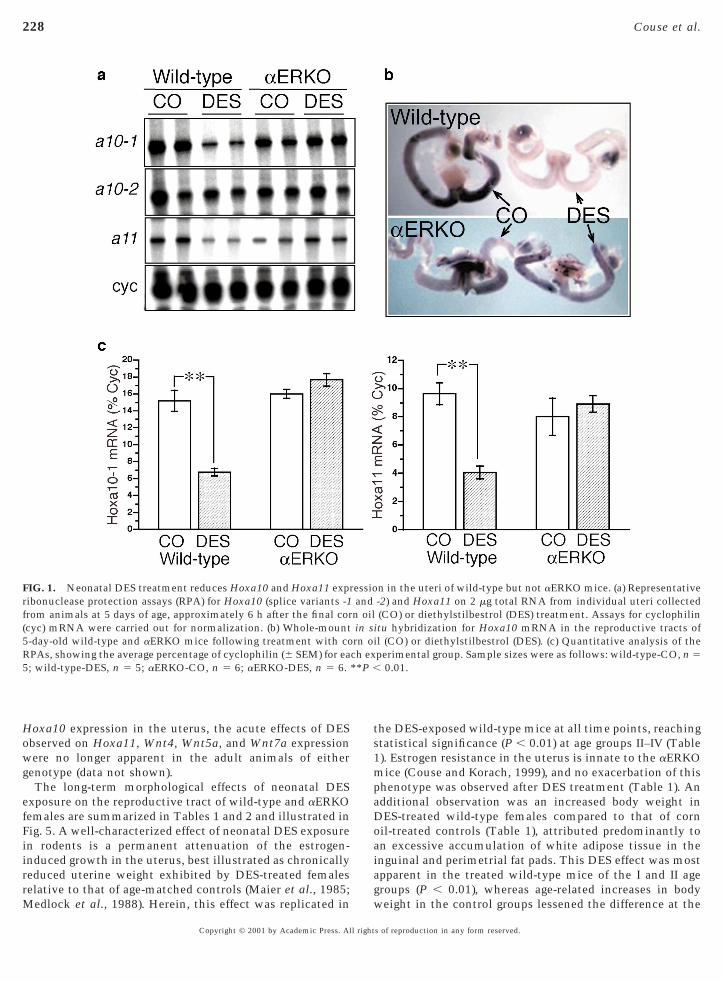
The acute effects of DES exposure on developmental geneexpression were assessed via analysis of mRNA levels forHoxa10, Hoxa11, Wnt4, Wnt5a, and Wnt7a in uterinetissue collected on neonatal Day 5, approximately 6–8 hafter the final treatment. As shown in Fig. 1, DES exposureelicited a significant decrease in the uterine levels ofmRNAs encoding Hoxa10-1 (P , 0.01) and Hoxa11 (P ,0.01) in the wild-type neonate when compared to that ofcorn oil-treated controls. In contrast, uterine tissues fromaERKO neonates failed to exhibit this effect of DES expo-sure and possessed Hoxa10-1 and Hoxa11 mRNA levels inboth treatment groups that were comparable to that of thecorn oil-treated wild-type. These results were corroboratedby whole-mount in situ hybridization for Hoxa10, as shownin Fig. 1b. There was no effect of genotype or treatment onthe levels of Hoxa10-2 transcripts, a previously describedsplice variant of the mouse Hoxa10 gene with unknownfunction (Benson et al., 1995) (Fig. 1).
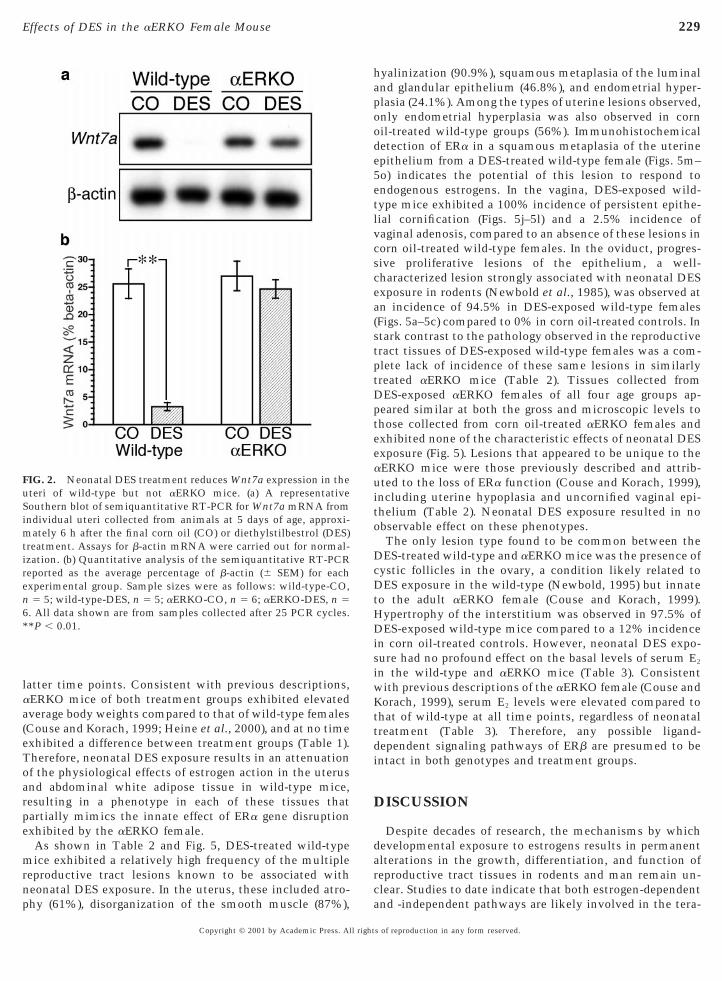
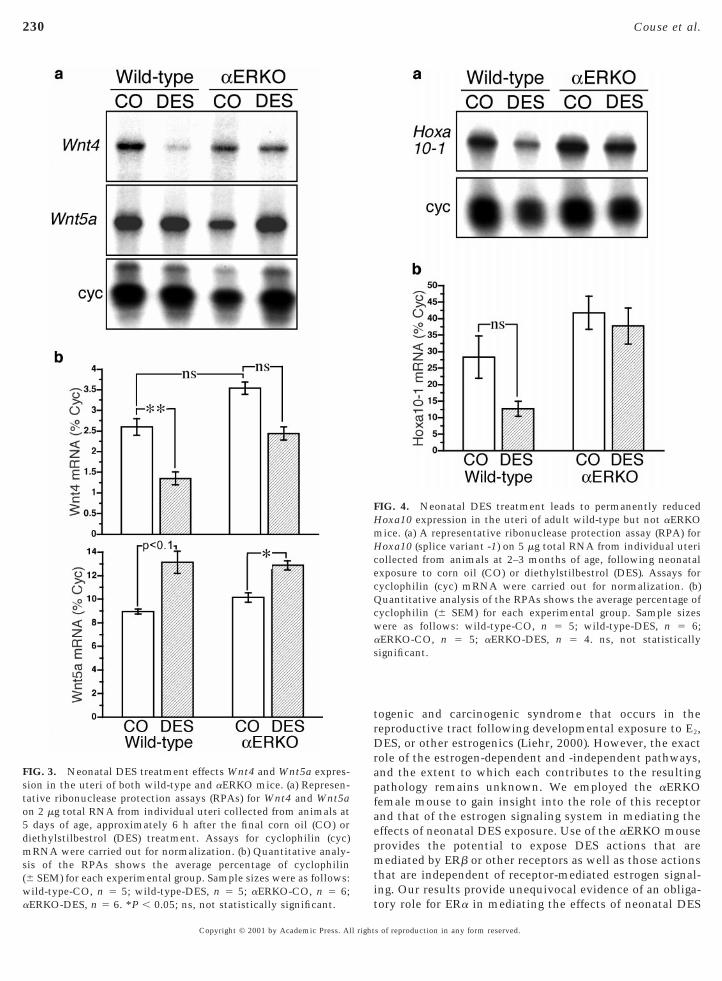
The results of similar assays for Wnt7a, Wnt4, and Wnt5atranscripts in neonatal uterine tissue following corn oil orDES treatment are shown in Figs. 2 and 3, respectively. Astriking effect of DES exposure in the neonatal uterus wasan over 80% decrease in Wnt7a mRNA levels compared tothat of corn oil-treated controls (P , 0.01) (Fig. 2). However,once again this effect of DES was exhibited only in thewild-type neonates and completely lacking in the DES-treated aERKO. In contrast to Wnt7a, the observed effectsof DES on uterine expression of the Wnt4 and Wnt5a geneswere observed in both wild-type and aERKO mice (Fig. 3).DES treatment elicited a significant decrease in uterineWnt4 mRNA levels that exceeded 50% (P , 0.01) in thewild-type, while a less-robust decrease was observed in theDES-treated aERKO mice. Interestingly, the basal levels of
nt4 transcripts in the uteri of the neonatal controlaERKO mice were increased compared to that of thewild-type, and although reduced following DES treatment,the ultimate levels of Wnt4 mRNA in the DES-treatedaERKO uteri remained similar to that of the corn oil-treated wild-type. In contrast to the reducing effect of DESon Wnt4 expression, a trend toward increased uterineexpression of Wnt5a was observed in the DES-treatedwild-type (P , 0.1) and aERKO neonates (P , 0.05).
The permanent nature of the acute effects of neonatalDES exposure on Hoxa10, Hoxa11, Wnt4, Wnt5a, and
nt7a expression in the uterus was assessed on tissueollected from sexually mature females (2–3 months) thatere exposed to either corn oil or DES as neonates. TheES-elicited reduction in Hoxa10–1 expression observed in
he uteri of neonatal wild-type mice appeared to persisturing sexual maturity, although it was no longer statisti-ally significant (Fig. 4). Once again the uterine levels ofoxa10–1 mRNA in the adult aERKO females of both
reatment groups were within the wild-type control rangend showed no effect of neonatal DES exposure. In contrasto the apparent permanent effect of DES exposure on
s of reproduction in any form reserved.
wg
e
228 Couse et al.
Hoxa10 expression in the uterus, the acute effects of DESobserved on Hoxa11, Wnt4, Wnt5a, and Wnt7a expression
ere no longer apparent in the adult animals of eitherenotype (data not shown).The long-term morphological effects of neonatal DES
xposure on the reproductive tract of wild-type and aERKOfemales are summarized in Tables 1 and 2 and illustrated inFig. 5. A well-characterized effect of neonatal DES exposurein rodents is a permanent attenuation of the estrogen-induced growth in the uterus, best illustrated as chronicallyreduced uterine weight exhibited by DES-treated femalesrelative to that of age-matched controls (Maier et al., 1985;Medlock et al., 1988). Herein, this effect was replicated in
FIG. 1. Neonatal DES treatment reduces Hoxa10 and Hoxa11 exprribonuclease protection assays (RPA) for Hoxa10 (splice variants -1from animals at 5 days of age, approximately 6 h after the final cor(cyc) mRNA were carried out for normalization. (b) Whole-mount5-day-old wild-type and aERKO mice following treatment with coRPAs, showing the average percentage of cyclophilin (6 SEM) for ea5; wild-type-DES, n 5 5; aERKO-CO, n 5 6; aERKO-DES, n 5 6.
Copyright © 2001 by Academic Press. All right
the DES-exposed wild-type mice at all time points, reachingstatistical significance (P , 0.01) at age groups II–IV (Table1). Estrogen resistance in the uterus is innate to the aERKOmice (Couse and Korach, 1999), and no exacerbation of thisphenotype was observed after DES treatment (Table 1). Anadditional observation was an increased body weight inDES-treated wild-type females compared to that of cornoil-treated controls (Table 1), attributed predominantly toan excessive accumulation of white adipose tissue in theinguinal and perimetrial fat pads. This DES effect was mostapparent in the treated wild-type mice of the I and II agegroups (P , 0.01), whereas age-related increases in bodyweight in the control groups lessened the difference at the
n in the uteri of wild-type but not aERKO mice. (a) Representative-2) and Hoxa11 on 2 mg total RNA from individual uteri collected(CO) or diethylstilbestrol (DES) treatment. Assays for cyclophilin
itu hybridization for Hoxa10 mRNA in the reproductive tracts ofl (CO) or diethylstilbestrol (DES). (c) Quantitative analysis of theperimental group. Sample sizes were as follows: wild-type-CO, n 5
0.01.
essioandn oilin srn oich ex**P ,
s of reproduction in any form reserved.
e
mrnp
hapood
D
ee
HDis
ttd
229Effects of DES in the aERKO Female Mouse
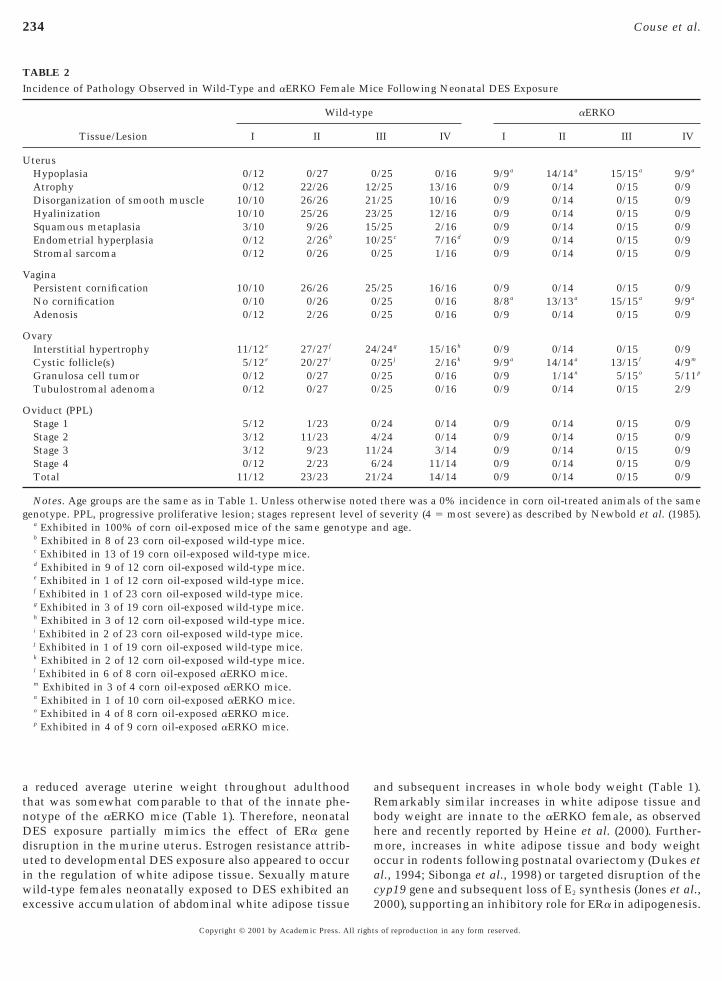
latter time points. Consistent with previous descriptions,aERKO mice of both treatment groups exhibited elevatedaverage body weights compared to that of wild-type females(Couse and Korach, 1999; Heine et al., 2000), and at no timeexhibited a difference between treatment groups (Table 1).Therefore, neonatal DES exposure results in an attenuationof the physiological effects of estrogen action in the uterusand abdominal white adipose tissue in wild-type mice,resulting in a phenotype in each of these tissues thatpartially mimics the innate effect of ERa gene disruptionxhibited by the aERKO female.As shown in Table 2 and Fig. 5, DES-treated wild-typeice exhibited a relatively high frequency of the multiple
eproductive tract lesions known to be associated witheonatal DES exposure. In the uterus, these included atro-hy (61%), disorganization of the smooth muscle (87%),
FIG. 2. Neonatal DES treatment reduces Wnt7a expression in theuteri of wild-type but not aERKO mice. (a) A representativeSouthern blot of semiquantitative RT-PCR for Wnt7a mRNA fromindividual uteri collected from animals at 5 days of age, approxi-mately 6 h after the final corn oil (CO) or diethylstilbestrol (DES)treatment. Assays for b-actin mRNA were carried out for normal-ization. (b) Quantitative analysis of the semiquantitative RT-PCRreported as the average percentage of b-actin (6 SEM) for eachexperimental group. Sample sizes were as follows: wild-type-CO,n 5 5; wild-type-DES, n 5 5; aERKO-CO, n 5 6; aERKO-DES, n 56. All data shown are from samples collected after 25 PCR cycles.**P , 0.01.
Copyright © 2001 by Academic Press. All right
yalinization (90.9%), squamous metaplasia of the luminalnd glandular epithelium (46.8%), and endometrial hyper-lasia (24.1%). Among the types of uterine lesions observed,nly endometrial hyperplasia was also observed in cornil-treated wild-type groups (56%). Immunohistochemicaletection of ERa in a squamous metaplasia of the uterine
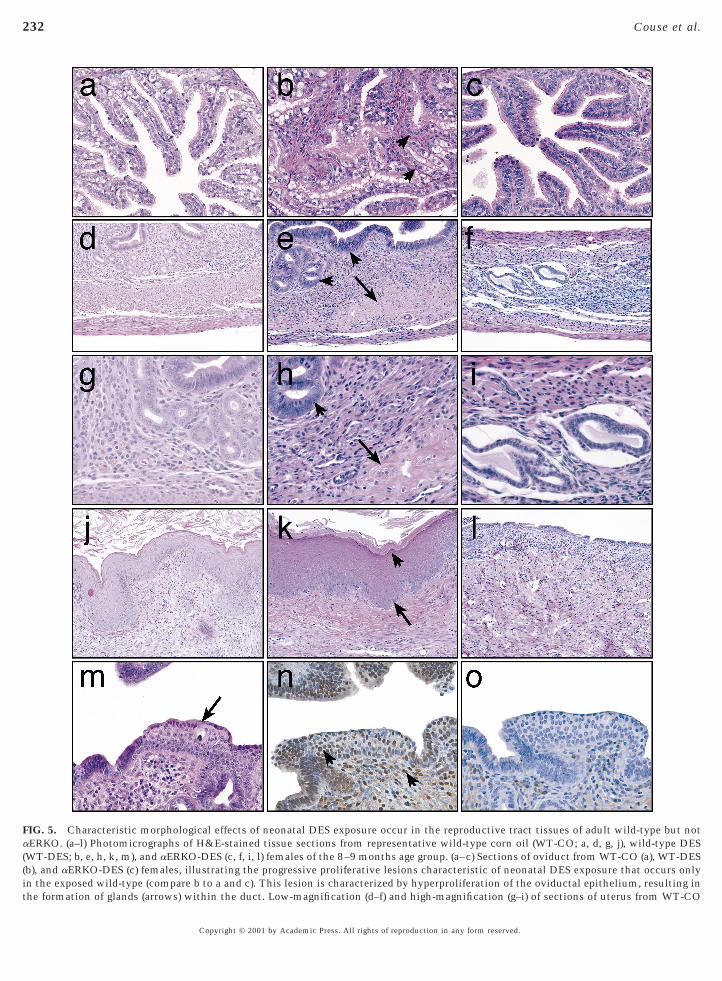
epithelium from a DES-treated wild-type female (Figs. 5m–5o) indicates the potential of this lesion to respond toendogenous estrogens. In the vagina, DES-exposed wild-type mice exhibited a 100% incidence of persistent epithe-lial cornification (Figs. 5j–5l) and a 2.5% incidence ofvaginal adenosis, compared to an absence of these lesions incorn oil-treated wild-type females. In the oviduct, progres-sive proliferative lesions of the epithelium, a well-characterized lesion strongly associated with neonatal DESexposure in rodents (Newbold et al., 1985), was observed atan incidence of 94.5% in DES-exposed wild-type females(Figs. 5a–5c) compared to 0% in corn oil-treated controls. Instark contrast to the pathology observed in the reproductivetract tissues of DES-exposed wild-type females was a com-plete lack of incidence of these same lesions in similarlytreated aERKO mice (Table 2). Tissues collected from
ES-exposed aERKO females of all four age groups ap-peared similar at both the gross and microscopic levels tothose collected from corn oil-treated aERKO females andxhibited none of the characteristic effects of neonatal DESxposure (Fig. 5). Lesions that appeared to be unique to the
aERKO mice were those previously described and attrib-uted to the loss of ERa function (Couse and Korach, 1999),including uterine hypoplasia and uncornified vaginal epi-thelium (Table 2). Neonatal DES exposure resulted in noobservable effect on these phenotypes.
The only lesion type found to be common between theDES-treated wild-type and aERKO mice was the presence ofcystic follicles in the ovary, a condition likely related toDES exposure in the wild-type (Newbold, 1995) but innateto the adult aERKO female (Couse and Korach, 1999).
ypertrophy of the interstitium was observed in 97.5% ofES-exposed wild-type mice compared to a 12% incidence
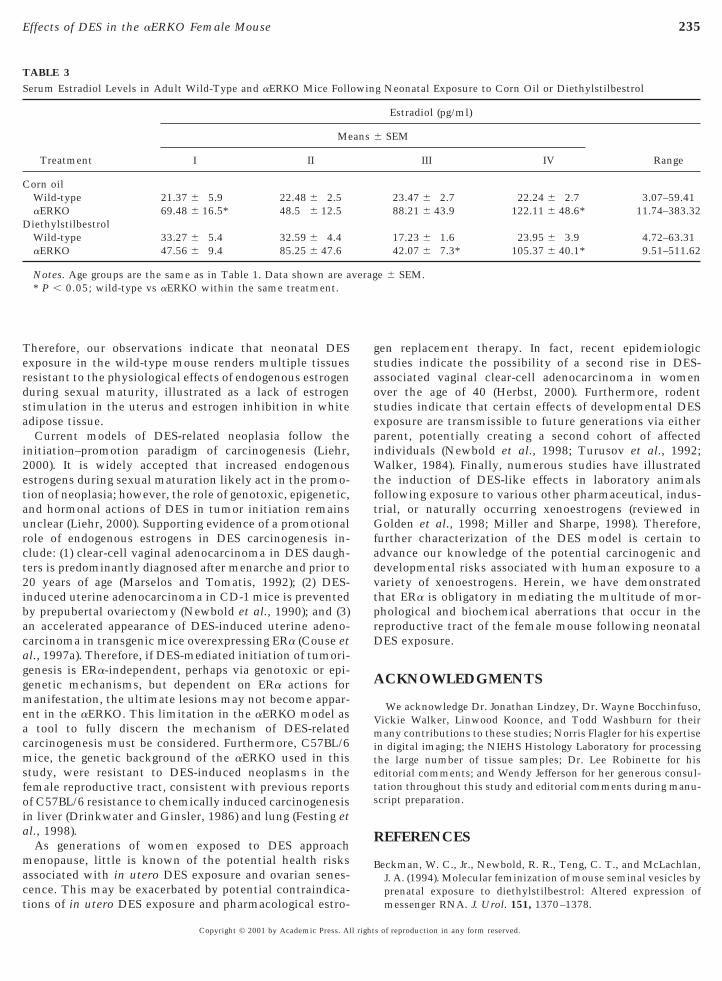
n corn oil-treated controls. However, neonatal DES expo-ure had no profound effect on the basal levels of serum E2
in the wild-type and aERKO mice (Table 3). Consistentwith previous descriptions of the aERKO female (Couse andKorach, 1999), serum E2 levels were elevated compared tohat of wild-type at all time points, regardless of neonatalreatment (Table 3). Therefore, any possible ligand-ependent signaling pathways of ERb are presumed to be
intact in both genotypes and treatment groups.
DISCUSSION
Despite decades of research, the mechanisms by whichdevelopmental exposure to estrogens results in permanentalterations in the growth, differentiation, and function ofreproductive tract tissues in rodents and man remain un-clear. Studies to date indicate that both estrogen-dependentand -independent pathways are likely involved in the tera-
s of reproduction in any form reserved.
fae
s
230 Couse et al.
Copyright © 2001 by Academic Press. All right
togenic and carcinogenic syndrome that occurs in thereproductive tract following developmental exposure to E2,DES, or other estrogenics (Liehr, 2000). However, the exactrole of the estrogen-dependent and -independent pathways,and the extent to which each contributes to the resultingpathology remains unknown. We employed the aERKOemale mouse to gain insight into the role of this receptornd that of the estrogen signaling system in mediating theffects of neonatal DES exposure. Use of the aERKO mouse
provides the potential to expose DES actions that aremediated by ERb or other receptors as well as those actionsthat are independent of receptor-mediated estrogen signal-ing. Our results provide unequivocal evidence of an obliga-tory role for ERa in mediating the effects of neonatal DES
FIG. 4. Neonatal DES treatment leads to permanently reducedHoxa10 expression in the uteri of adult wild-type but not aERKOmice. (a) A representative ribonuclease protection assay (RPA) forHoxa10 (splice variant -1) on 5 mg total RNA from individual utericollected from animals at 2–3 months of age, following neonatalexposure to corn oil (CO) or diethylstilbestrol (DES). Assays forcyclophilin (cyc) mRNA were carried out for normalization. (b)Quantitative analysis of the RPAs shows the average percentage ofcyclophilin (6 SEM) for each experimental group. Sample sizeswere as follows: wild-type-CO, n 5 5; wild-type-DES, n 5 6;aERKO-CO, n 5 5; aERKO-DES, n 5 4. ns, not statisticallysignificant.
FIG. 3. Neonatal DES treatment effects Wnt4 and Wnt5a expres-ion in the uteri of both wild-type and aERKO mice. (a) Represen-
tative ribonuclease protection assays (RPAs) for Wnt4 and Wnt5aon 2 mg total RNA from individual uteri collected from animals at5 days of age, approximately 6 h after the final corn oil (CO) ordiethylstilbestrol (DES) treatment. Assays for cyclophilin (cyc)mRNA were carried out for normalization. (b) Quantitative analy-sis of the RPAs shows the average percentage of cyclophilin(6 SEM) for each experimental group. Sample sizes were as follows:wild-type-CO, n 5 5; wild-type-DES, n 5 5; aERKO-CO, n 5 6;aERKO-DES, n 5 6. *P , 0.05; ns, not statistically significant.
s of reproduction in any form reserved.

s
1aafapdabeD
nlc
treat
231Effects of DES in the aERKO Female Mouse
exposure in the developing murine female reproductivetract.
The sensitivity of the developing female reproductivetract to the detrimental effects of DES has been welldocumented (Marselos and Tomatis, 1993). The completelack of DES effects in the aERKO female indicate thisensitivity is the result of the high levels of ERa present in
these tissues during late fetal and neonatal development(Greco et al., 1993; Korach et al., 1988; Yamashita et al.,989). None of the DES-exposed aERKO females, includingnimals exceeding 22 months of age, exhibited the distinctbnormalities that were apparent in DES-treated wild-typeemales. This included the absence of several lesions char-cteristic of neonatal DES exposure, such as progressiveroliferative lesion of the oviduct (Newbold et al., 1985);ecreased weight, atrophy, smooth muscle disorganization,nd epithelial squamous metaplasia in the uterus (New-old, 1995); and persistent cornification of the vaginalpithelium (Takasugi, 1976). Therefore, the absence ofES-induced abnormalities in the reproductive tract of the
aERKO supports a predominant role of ERa in mediatingthe detrimental effects of DES in female reproductive tractdevelopment. The lack of apparent ERb-mediated effects of
eonatal DES exposure, despite the presence of elevatedevels of endogenous E2, is consistent with previous diffi-ulty in detecting ERb mRNA or protein in uterine, ovi-
duct, and vaginal tissue of newborn and adult mice (Couseet al., 1997b; Jefferson et al., 2000).
Recent molecular studies have begun to provide clues asto the mechanisms by which developmental DES exposure
TABLE 1Sample Number and Whole Body and Uterine Weights in Adult WFollowing Neonatal Exposure to Corn Oil or Diethylstilbestrol (D
Treatment group I
Sample numberWT—corn oil 12WT—DES 12aERKO—corn oil 11aERKO—DES 9
Body weightb (g)WT—corn oil 23.3 (0.4) 2WT—DES 32.8 (2.1)** 3aERKO—corn oil 28.7 (0.9)† 3aERKO—DES 30.5 (1.2) 3
Uterine weightb,c
WT—corn oil 0.42 (0.04) 0WT–DES 0.30 (0.04) 0aERKO—corn oil 0.26 (0.05) 0aERKO—DES 0.28 (0.03) 0
a Age groups (months): I 5 4–5, II 5 8–9, III 5 12–13, IV 5 18–2b Data shown for body weight and uterine weight are average (Sc Uterine weight reported as percentage of body weight.* P , 0.05, **P , 0.01; corn oil vs DES within the same geno† P , 0.05, ††P , 0.01; wild-type vs aERKO within the same
Copyright © 2001 by Academic Press. All right
leads to its syndrome of defects in the female reproductivetract. Considerable efforts have focused on the clusteredhomeobox genes Hoxa10 and Hoxa11, both of which areexpressed in the paramesonephric ductal segments of theMullerian duct that ultimately differentiate into the uterusand cranial cervix (Kitajewski and Sassoon, 2000; Taylor etal., 1997). The developmental expression pattern of Hoxa10and Hoxa11 is highly conserved in mice and humans(Taylor et al., 1997), the importance of which has beeneffectively illustrated by studies of null mice. Hoxa10 nullmice exhibit a partial anterior homeotic transformation inwhich the anterior 25% of the uterus resembles the struc-tural and molecular characteristics of the oviduct (Bensonet al., 1996); Hoxa11 null mice exhibit thinner, shorteruterine horns with a paucity of glands (Gendron et al.,1997). These phenotypes in Hoxa10 and Hoxa11 null miceare somewhat similar to those described in mice followingdevelopmental DES exposure (Kitajewski and Sassoon,2000). Therefore, a potential mechanism of DES may in-volve alterations in the developmental program required forproper uterine differentiation (Block et al., 2000; Kitajewskiand Sassoon, 2000; Ma et al., 1998). By 6 h after the finalneonatal DES treatment, a significant reduction in Hoxa10and Hoxa11 expression of .50% was observed in the uteriof wild-type mice, consistent with previous reports (Blocket al., 2000; Ma et al., 1998). Based on reported gene–dosageeffects following targeted disruption of the Hoxa11 andHoxa13 genes (Hsieh-Li et al., 1995; Warot et al., 1997), thislevel of reduction in gene expression could ostensibly elicitan aberrant phenotype. More important, our studies illus-
ype (WT) and aERKO Mice
Age groupa
I III IV
19 1827 209 9
15 10
.5) 29.1 (1.2) 31.6 (1.2)
.2)** 34.1 (1.0) 36.3 (2.1)
.2)†† 31.1 (1.7) 32.1 (2.0)
.4) 34.6 (2.0) 30.4 (1.9)
.05) 0.65 (0.08) 0.93 (0.14)
.02)** 0.26 (0.02)** 0.44 (0.09)**
.01)†† 0.20 (0.06)†† 0.33 (0.07)†
.03) 0.20 (0.06) 0.20 (0.04)
.ment.
ild-TES)
I
23271014
5.1 (02.3 (13.8 (11.6 (1
.46 (0
.23 (0
.09 (0
.15 (0
2.7.EM).
type
s of reproduction in any form reserved.
(
232 Couse et al.
FIG. 5. Characteristic morphological effects of neonatal DES exposure occur in the reproductive tract tissues of adult wild-type but notaERKO. (a–l) Photomicrographs of H&E-stained tissue sections from representative wild-type corn oil (WT-CO; a, d, g, j), wild-type DES(WT-DES; b, e, h, k, m), and aERKO-DES (c, f, i, l) females of the 8–9 months age group. (a–c) Sections of oviduct from WT-CO (a), WT-DESb), and aERKO-DES (c) females, illustrating the progressive proliferative lesions characteristic of neonatal DES exposure that occurs onlyin the exposed wild-type (compare b to a and c). This lesion is characterized by hyperproliferation of the oviductal epithelium, resulting inthe formation of glands (arrows) within the duct. Low-magnification (d–f) and high-magnification (g–i) of sections of uterus from WT-CO
Copyright © 2001 by Academic Press. All rights of reproduction in any form reserved.

e2mooMHcs
i
ot
io
D
car(tdtwH
lwhvn
233Effects of DES in the aERKO Female Mouse
trate that the action of DES on Hoxa10 and Hoxa11expression in the neonatal uterus is dependent on thepresence of a functional ERa.
The reducing effect of neonatal DES exposure on uterineHoxa10 expression remained detectable during adulthood(Fig. 4) and is possibly significant, considering the criticalrole of Hoxa gene expression in implantation and pregnancy(Benson et al., 1996; Hsieh-Li et al., 1995; Lim et al., 1999).Furthermore, aberrant uterine Hoxa10 and Hoxa11 expres-sion is reported in women with endometriosis (Taylor et al.,1999). Although an association between in utero DESexposure and reduced fertility or endometriosis in humansremains controversial (Golden et al., 1998), these datawarrant further study of uterine HOX gene expression inDES daughters.
Previous studies have provided indirect evidence of ste-roid receptor-mediated regulation of Hoxa10 and Hoxa11xpression in the mouse and human uterus (Block et al.,000; Ma et al., 1998; Taylor et al., 1998). Furthermore,ultiple consensus binding sites for the ER and progester-
ne receptor are present in the known genomic sequencesf Hoxa10 and Hoxa11 (Ma et al., 1998). However, whereasa et al. (1998) reported that E2 continues to reduceoxa10 expression in the adult mouse uterus, Taylor and
olleagues (Block et al., 2000; Taylor et al., 1998) report atimulatory effect of DES and E2 in multiple in vitro
systems but no discernable effect in human endometrialtissue during periods of elevated serum estrogens (Taylor etal., 1998). Herein, no misregulation of Hoxa10 expressionin the uteri of adult aERKO mice was observed, despite thepresence of elevated plasma E2. Therefore, our data indicatethat estrogen regulation of Hoxa10 and Hoxa11 expressionin both the neonatal and adult uterus is predominantlydependent on the presence of functional ERa.
In addition to the homeobox genes, Wnt gene productsalso play critical roles in the regulation of tissue patterningduring embryogenesis and adulthood (Kitajewski and Sas-soon, 2000). At 6 days of age in the mouse uterus, Wnt4expression is limited to the mesenchyme nearest the lu-men, Wnt5a is limited to the stroma, and Wnt7a appears inthe luminal and glandular epithelium (Miller et al., 1998b).Targeted deletion of the Wnt7a gene in mice results in amultitude of uterine phenotypes that resemble the effectsof neonatal DES exposure (Kitajewski and Sassoon, 2000),including a disorganized myometrium, reduced uterine
(d, g), WT-DES (e, h), and aERKO-DES (f, i) females, illustrating the euminal and glandular epithelium (short arrows) and disorganizatioere apparent only in the WT-DES (e and h). Uterine tissue from aEypoplasia of the smooth muscle, endometrial stroma, and glanduagina from WT-CO (j), WT-DES (k), and aERKO-DES (l) femaleeonatal DES exposure that occurs only in the exposed wild-type (c
exhibited an uncornified vaginal epithelium, a phenotype innate touterine epithelium of a WT-DES female, stained with H&E (mimmunoreactivity in the cells of the epithelial lesion and underlyiwhich the ERa antibody was excluded during immunohistochemi
Copyright © 2001 by Academic Press. All right
diameter, absence of glands, and a stratified vaginal-likeepithelium rather than the simple columnar epitheliumcharacteristic of the normal uterus (Parr and McMahon,1998). As with the Hoxa genes, DES elicits an acutereduction in Wnt7a expression of almost 90% in the neo-natal wild-type mouse uterus (Fig. 2). These data are con-sistent with a previously reported effect of prenatal DESexposure on Wnt7a expression (Miller et al., 1998a). Moreimportant, the lack of a DES-elicited reduction in Wnt7aexpression in the aERKO uteri provides definitive evidenceof the requirement for ERa in this mechanism. Therefore, its likely that the loss of ERa function in the aERKO female
prevents the aberrant developmental defects of DES in thereproductive tract by allowing for normal expression ofHoxa10, Hoxa11, and Wnt7a.
The preservation of two immediate effects of DES in theneonatal aERKO uterus, decreased Wnt4 and increasedWnt5a expression, suggests the existence of ERa-independent DES-activated pathways. A likely candidatefor the maintenance of certain hormonal actions of DES inthe aERKO uterus is ERb, although it remains difficult todetect ERb mRNA or protein in uterine tissue of newbornmice (Jefferson et al., 2000). Also possible is the activationf apparent ER-independent pathways in the uterus, such ashose reported in ovariectomized aERKO females following
treatment with 4-hydroxy-E2, methoxychlor, or kepone(Das et al., 1997; Ghosh et al., 1999). The recent report byTremblay et al. (2001) that DES, and not E2, specificallynteracts with and suppresses the transactivational activityf the estrogen-related receptors ERRa, -b, and -g presents
the possibility that these receptors may also be involved inWnt4 and Wnt5a regulation. Nonetheless, the effects of
ES on the uterine Wnt4 and Wnt5a expression weretransient, given that normal mRNA levels were detected inthe adult wild-type and aERKO mice.
Decreased uterine weight in sexually mature females is aharacteristic effect of neonatal DES exposure in the rodentnd is attributed to an attenuation of the uterine estrogenesponse resulting from constitutive reduction in ER levelsMaier et al., 1985; Medlock et al., 1988, 1992). Similarly,he prostate of the neonatal estrogenized rat exhibits re-uced androgen receptor levels and an attenuated responseo endogenous androgens, leading to reduced prostateeight in the adult (Prins, 1992; Prins and Birch, 1995).erein, neonatal DES-exposed wild-type females exhibited
s of neonatal DES exposure in the uterus, including an estrogenizedalinization of the inner circular smooth muscle (long arrows) that-DES (f, i) exhibited no effects of neonatal DES exposure, but only
pithelium, a phenotype innate to the loss of ERa. (j–l) Sections oflustrating the excessive epithelial cornification characteristic ofre k to j and l). This effect was absent in the aERKO-DES (l), whichoss of ERa. (m–o) Illustration of early squamous metaplasia in theimmunostained for ERa (n), illustrating specific nuclear ERa
roma (indicated by arrows). (o) Represention of a serial section innd therefore serves as a negative control.
ffectn/hyRKO
lar es, il
ompathe l) or
ng ststry a
s of reproduction in any form reserved.
aRb
2
U
234 Couse et al.
a reduced average uterine weight throughout adulthoodthat was somewhat comparable to that of the innate phe-notype of the aERKO mice (Table 1). Therefore, neonatalDES exposure partially mimics the effect of ERa genedisruption in the murine uterus. Estrogen resistance attrib-uted to developmental DES exposure also appeared to occurin the regulation of white adipose tissue. Sexually maturewild-type females neonatally exposed to DES exhibited anexcessive accumulation of abdominal white adipose tissue
TABLE 2Incidence of Pathology Observed in Wild-Type and aERKO Femal
Tissue/Lesion
Wild-
I II
terusHypoplasia 0/12 0/27Atrophy 0/12 22/26Disorganization of smooth muscle 10/10 26/26Hyalinization 10/10 25/26Squamous metaplasia 3/10 9/26Endometrial hyperplasia 0/12 2/26b
Stromal sarcoma 0/12 0/26
VaginaPersistent cornification 10/10 26/26No cornification 0/10 0/26Adenosis 0/12 2/26
OvaryInterstitial hypertrophy 11/12e 27/27f
Cystic follicle(s) 5/12e 20/27i
Granulosa cell tumor 0/12 0/27Tubulostromal adenoma 0/12 0/27
Oviduct (PPL)Stage 1 5/12 1/23Stage 2 3/12 11/23Stage 3 3/12 9/23Stage 4 0/12 2/23Total 11/12 23/23
Notes. Age groups are the same as in Table 1. Unless otherwise ngenotype. PPL, progressive proliferative lesion; stages represent lev
a Exhibited in 100% of corn oil-exposed mice of the same genotb Exhibited in 8 of 23 corn oil-exposed wild-type mice.c Exhibited in 13 of 19 corn oil-exposed wild-type mice.d Exhibited in 9 of 12 corn oil-exposed wild-type mice.e Exhibited in 1 of 12 corn oil-exposed wild-type mice.f Exhibited in 1 of 23 corn oil-exposed wild-type mice.g Exhibited in 3 of 19 corn oil-exposed wild-type mice.h Exhibited in 3 of 12 corn oil-exposed wild-type mice.i Exhibited in 2 of 23 corn oil-exposed wild-type mice.j Exhibited in 1 of 19 corn oil-exposed wild-type mice.k Exhibited in 2 of 12 corn oil-exposed wild-type mice.l Exhibited in 6 of 8 corn oil-exposed aERKO mice.m Exhibited in 3 of 4 corn oil-exposed aERKO mice.n Exhibited in 1 of 10 corn oil-exposed aERKO mice.o Exhibited in 4 of 8 corn oil-exposed aERKO mice.p Exhibited in 4 of 9 corn oil-exposed aERKO mice.
Copyright © 2001 by Academic Press. All right
nd subsequent increases in whole body weight (Table 1).emarkably similar increases in white adipose tissue andody weight are innate to the aERKO female, as observed
here and recently reported by Heine et al. (2000). Further-more, increases in white adipose tissue and body weightoccur in rodents following postnatal ovariectomy (Dukes etal., 1994; Sibonga et al., 1998) or targeted disruption of thecyp19 gene and subsequent loss of E2 synthesis (Jones et al.,000), supporting an inhibitory role for ERa in adipogenesis.
e Following Neonatal DES Exposure
aERKO
III IV I II III IV
/25 0/16 9/9a 14/14a 15/15a 9/9a
/25 13/16 0/9 0/14 0/15 0/9/25 10/16 0/9 0/14 0/15 0/9/25 12/16 0/9 0/14 0/15 0/9/25 2/16 0/9 0/14 0/15 0/9/25c 7/16d 0/9 0/14 0/15 0/9/25 1/16 0/9 0/14 0/15 0/9
/25 16/16 0/9 0/14 0/15 0/9/25 0/16 8/8a 13/13a 15/15a 9/9a
/25 0/16 0/9 0/14 0/15 0/9
/24g 15/16h 0/9 0/14 0/15 0/9/25j 2/16k 9/9a 14/14a 13/15l 4/9m
/25 0/16 0/9 1/14n 5/15o 5/11p
/25 0/16 0/9 0/14 0/15 2/9
/24 0/14 0/9 0/14 0/15 0/9/24 0/14 0/9 0/14 0/15 0/9/24 3/14 0/9 0/14 0/15 0/9/24 11/14 0/9 0/14 0/15 0/9/24 14/14 0/9 0/14 0/15 0/9
there was a 0% incidence in corn oil-treated animals of the sameseverity (4 5 most severe) as described by Newbold et al. (1985).
nd age.
e Mic
type
012212315100
2500
24000
04
116
21
otedel of
ype a
s of reproduction in any form reserved.
acag
meacm
235Effects of DES in the aERKO Female Mouse
Therefore, our observations indicate that neonatal DESexposure in the wild-type mouse renders multiple tissuesresistant to the physiological effects of endogenous estrogenduring sexual maturity, illustrated as a lack of estrogenstimulation in the uterus and estrogen inhibition in whiteadipose tissue.
Current models of DES-related neoplasia follow theinitiation–promotion paradigm of carcinogenesis (Liehr,2000). It is widely accepted that increased endogenousestrogens during sexual maturation likely act in the promo-tion of neoplasia; however, the role of genotoxic, epigenetic,and hormonal actions of DES in tumor initiation remainsunclear (Liehr, 2000). Supporting evidence of a promotionalrole of endogenous estrogens in DES carcinogenesis in-clude: (1) clear-cell vaginal adenocarcinoma in DES daugh-ters is predominantly diagnosed after menarche and prior to20 years of age (Marselos and Tomatis, 1992); (2) DES-induced uterine adenocarcinoma in CD-1 mice is preventedby prepubertal ovariectomy (Newbold et al., 1990); and (3)n accelerated appearance of DES-induced uterine adeno-arcinoma in transgenic mice overexpressing ERa (Couse etl., 1997a). Therefore, if DES-mediated initiation of tumori-enesis is ERa-independent, perhaps via genotoxic or epi-
genetic mechanisms, but dependent on ERa actions foranifestation, the ultimate lesions may not become appar-
nt in the aERKO. This limitation in the aERKO model astool to fully discern the mechanism of DES-related
arcinogenesis must be considered. Furthermore, C57BL/6ice, the genetic background of the aERKO used in this
study, were resistant to DES-induced neoplasms in thefemale reproductive tract, consistent with previous reportsof C57BL/6 resistance to chemically induced carcinogenesisin liver (Drinkwater and Ginsler, 1986) and lung (Festing etal., 1998).
As generations of women exposed to DES approachmenopause, little is known of the potential health risksassociated with in utero DES exposure and ovarian senes-cence. This may be exacerbated by potential contraindica-tions of in utero DES exposure and pharmacological estro-
TABLE 3Serum Estradiol Levels in Adult Wild-Type and aERKO Mice Foll
Treatment
Me
I II
Corn oilWild-type 21.37 6 5.9 22.48 6 2.5aERKO 69.48 6 16.5* 48.5 6 12.5
DiethylstilbestrolWild-type 33.27 6 5.4 32.59 6 4.4aERKO 47.56 6 9.4 85.25 6 47.6
Notes. Age groups are the same as in Table 1. Data shown are a* P , 0.05; wild-type vs aERKO within the same treatment.
Copyright © 2001 by Academic Press. All right
gen replacement therapy. In fact, recent epidemiologicstudies indicate the possibility of a second rise in DES-associated vaginal clear-cell adenocarcinoma in womenover the age of 40 (Herbst, 2000). Furthermore, rodentstudies indicate that certain effects of developmental DESexposure are transmissible to future generations via eitherparent, potentially creating a second cohort of affectedindividuals (Newbold et al., 1998; Turusov et al., 1992;Walker, 1984). Finally, numerous studies have illustratedthe induction of DES-like effects in laboratory animalsfollowing exposure to various other pharmaceutical, indus-trial, or naturally occurring xenoestrogens (reviewed inGolden et al., 1998; Miller and Sharpe, 1998). Therefore,further characterization of the DES model is certain toadvance our knowledge of the potential carcinogenic anddevelopmental risks associated with human exposure to avariety of xenoestrogens. Herein, we have demonstratedthat ERa is obligatory in mediating the multitude of mor-phological and biochemical aberrations that occur in thereproductive tract of the female mouse following neonatalDES exposure.
ACKNOWLEDGMENTS
We acknowledge Dr. Jonathan Lindzey, Dr. Wayne Bocchinfuso,Vickie Walker, Linwood Koonce, and Todd Washburn for theirmany contributions to these studies; Norris Flagler for his expertisein digital imaging; the NIEHS Histology Laboratory for processingthe large number of tissue samples; Dr. Lee Robinette for hiseditorial comments; and Wendy Jefferson for her generous consul-tation throughout this study and editorial comments during manu-script preparation.
REFERENCES
Beckman, W. C., Jr., Newbold, R. R., Teng, C. T., and McLachlan,J. A. (1994). Molecular feminization of mouse seminal vesicles byprenatal exposure to diethylstilbestrol: Altered expression ofmessenger RNA. J. Urol. 151, 1370–1378.
g Neonatal Exposure to Corn Oil or Diethylstilbestrol
Estradiol (pg/ml)
SEM
RangeIII IV
23.47 6 2.7 22.24 6 2.7 3.07–59.4188.21 6 43.9 122.11 6 48.6* 11.74–383.32
17.23 6 1.6 23.95 6 3.9 4.72–63.3142.07 6 7.3* 105.37 6 40.1* 9.51–511.62
e 6 SEM.
owin
ans 6
verag
s of reproduction in any form reserved.

B
B
C
C
D
D
D
D
F
F
F
G
G
G
G
G
G
H
H
236 Couse et al.
Benson, G. V., Lim, H., Paria, B. C., Satokata, I., Dey, S. K., andMaas, R. L. (1996). Mechanisms of reduced fertility in Hoxa-10mutant mice: Uterine homeosis and loss of maternal Hoxa-10expression. Development 122, 2687–2696.
enson, G. V., Nguyen, T. H., and Maas, R. L. (1995). Theexpression pattern of the murine Hoxa-10 gene and the sequencerecognition of its homeodomain reveal specific properties ofAbdominal B-like genes. Mol. Cell. Biol. 15, 1591–1601.
lock, K., Kardana, A., Igarashi, P., and Taylor, H. S. (2000). In uterodiethylstilbestrol (DES) exposure alters Hox gene expression inthe developing mullerian system. FASEB J. 14, 1101–1108.
Couse, J. F., Davis, V. L., Hanson, R. B., Jefferson, W. N.,McLachlan, J. A., Bullock, B. C., Newbold, R. R., and Korach,K. S. (1997a). Accelerated onset of uterine tumors in transgenicmice with aberrant expression of the estrogen receptor afterneonatal exposure to diethylstilbestrol. Mol. Carcinog. 19, 236–242.
Couse, J. F., Davis, V. L., Tally, W. C., and Korach, K. S. (1994). Animproved method of genomic DNA extraction for screeningtransgenic mice. Biotechniques 17, 1030–1032.
Couse, J. F., Hewitt, S. C., Bunch, D. O., Sar, M., Walker, V. R.,Davis, B. J., and Korach, K. S. (1999). Postnatal sex reversal of theovaries in mice lacking estrogen receptors a and b. Science 286,2328–2331.ouse, J. F., and Korach, K. S. (1999). Estrogen receptor null mice:What have we learned and where will they lead us? Endocr. Rev.20, 358–417.ouse, J. F., Lindzey, J., Grandien, K., Gustafsson, J. Å., and Korach,K. S. (1997b). Tissue distribution and quantitative analysis ofestrogen receptor-a (ERa) and estrogen receptor-b (ERb) messen-ger ribonucleic acid in the wild-type and ERa-knockout mouse.Endocrinology 138, 4613–4621.
Cunha, G. R., Cooke, P. S., Bigsby, R., and Brody, J. R. (1991).Ontogeny of sex steroid receptors in mammals. In “NuclearHormone Receptors: Molecular Mechanisms, Cellular Func-tions, Clinical Abnormalities” (M. G. Parker, Ed.), pp. 235–268.Academic Press, London.as, S. K., Taylor, J. A., Korach, K. S., Paria, B. C., Dey, S. K., andLubahn, D. B. (1997). Estrogenic responses in estrogen receptor-adeficient mice reveal a distinct estrogen signaling pathway. Proc.Natl. Acad. Sci. USA 94, 12786–12791.ieckmann, W. J., Davis, M. E., Rynkiewicz, L. M., and Pottinger,R. E. (1953). Does the administration of diethylstilbestrol duringpregnancy have therapeutic value? Am. J. Obstet. Gynecol. 66,1062–1081.
Dolle, P., Izpisua-Belmonte, J. C., Brown, J. M., Tickle, C., andDuboule, D. (1991). HOX-4 genes and the morphogenesis ofmammalian genitalia. Genes Dev. 5, 1767–1776.rinkwater, N. R., and Ginsler, J. J. (1986). Genetic control ofhepatocarcinogenesis in C57BL/6J and C3H/HeJ inbred mice.Carcinogenesis 7, 1701–1707.ukes, M., Chester, R., Yarwood, L., and Wakeling, A. E. (1994).Effects of a non-steroidal pure antioestrogen, ZM 189,154, onoestrogen target organs of the rat including bones. J. Endocrinol.141, 335–341.
alck, L., and Forsberg, J.-G. (1996). Immunohistochemical studieson the expression and estrogen dependency of EGF and itsreceptor and c-fos proto-oncogene in the uterus and vagina ofnormal and neonatally estrogen-treated mice. Anat. Rec. 245,459–471.
esting, M. F., Lin, L., Devereux, T. R., Gao, F., Yang, A., Anna,C. H., White, C. M., Malkinson, A. M., and You, M. (1998). Atleast four loci and gender are associated with susceptibility to the
Copyright © 2001 by Academic Press. All right
chemical induction of lung adenomas in A/J x BALB/c mice.Genomics 53, 129–136.
isher, C. R., Graves, K. H., Parlow, A. F., and Simpson, E. R. (1998).Characterization of mice deficient in aromatase (ArKO) becauseof targeted disruption of the cyp19 gene. Proc. Natl. Acad. Sci.USA 95, 6965–6970.endron, R. L., Paradis, H., Hsieh-Li, H. M., Lee, D. W., Potter,S. S., and Markoff, E. (1997). Abnormal uterine stromal andglandular function associated with maternal reproductive defectsin Hoxa-11 null mice. Biol. Reprod. 56, 1097–1105.hosh, D., Taylor, J. A., Green, J. A., and Lubahn, D. B. (1999).Methoxychlor stimulates estrogen-responsive messenger ribo-nucleic acids in mouse uterus through a non-estrogen receptor(non-ER) alpha and non-ER beta mechanism. Endocrinology 140,3526–3533.iguere, V., Tremblay, A., and Tremblay, G. B. (1998). Estrogenreceptor b: Re-evaluation of estrogen and antiestrogen signaling.Steroids 63, 335–339.olden, R. J., Noller, K. L., Titus-Ernstoff, L., Kaufman, R. H.,Mittendorf, R., Stillman, R., and Reese, E. A. (1998). Environ-mental endocrine modulators and human health: An assessmentof the biological evidence. Crit. Rev. Toxicol. 28, 109–227.reco, T. L., Duello, T. M., and Gorski, J. (1993). Estrogen recep-tors, estradiol, and diethylstilbestrol in early development: Themouse as a model for the study of estrogen receptors and estrogensensitivity in embryonic development of male and female repro-ductive tracts. Endocr. Rev. 14, 59–71.reenwald, P., Barlow, J. J., Nasca, P. C., and Burnett, W. S. (1971).Vaginal cancer after maternal treatment with synthetic estro-gens. N. Engl. J. Med. 285, 390–392.ajek, R. A., Nguyen, T. V., Johnston, D. A., and Jones, L. A. (1993).In vivo induction of increased DNA ploidy of mouse cervicovagi-nal epithelium by neonatal estrogen treatment. Biol. Reprod. 49,908–917.eine, P. A., Taylor, J. A., Iwamoto, G. A., Lubahn, D. B., andCooke, P. S. (2000). Increased adipose tissue in male and femaleestrogen receptor-a knockout mice. Proc. Natl. Acad. Sci. USA97, 12729–12734.
Herbst, A. L. (2000). Behavior of estrogen-associated female genitaltract cancer and its relation to neoplasia following intrauterineexposure to diethylstilbestrol (DES). Gynecol. Oncol. 76, 147–156.
Herbst, A. L., Ulfelder, H., and Poskanzer, D. C. (1971). Adenocar-cinoma of the vagina. Association of maternal stilbestrol therapywith tumor appearance in young women. N. Engl. J. Med. 284,878–881.
Hsieh-Li, H. M., Witte, D. P., Weinstein, M., Branford, W., Li, H.,Small, K., and Potter, S. S. (1995). Hoxa 11 structure, extensiveantisense transcription, and function in male and female fertil-ity. Development 121, 1373–1385.
International Association for Research on Cancer (IARC). (1979).“IARC Monographs on the Evaluation of the Carcinogenic Riskof Chemicals to Humans: Sex Hormones (II).” IARC, Lyon,France.
IARC. (1987). “IARC Monographs on the Evaluation of the Carci-nogenic Risk of Chemicals to Humans: An Updating of IARCMonographs 1987, Vols. 1–42.” IARC, Lyon, France.
Jefferson, W. N., Couse, J. F., Banks, E. P., Korach, K. S., andNewbold, R. R. (2000). Expression of estrogen receptor beta isdevelopmentally regulated in reproductive tissues of male andfemale mice. Biol. Reprod. 62, 310–317.
Jones, M. E., Thorburn, A. W., Britt, K. L., Hewitt, K. N., Wreford,N. G., Proietto, J., Oz, O. K., Leury, B. J., Robertson, K. M., Yao,
s of reproduction in any form reserved.

K
L
L
L
M
M
M
M
M
M
M
M
N
N
N
S
S
T
T
T
T
T
237Effects of DES in the aERKO Female Mouse
S., and Simpson, E. R. (2000). Aromatase-deficient (ArKO) micehave a phenotype of increased adiposity. Proc. Natl. Acad. Sci.USA 97, 12735–12740.
Kitajewski, J., and Sassoon, D. (2000). The emergence of moleculargynecology: homeobox and wnt genes in the female reproductivetract. BioEssays 22, 902–910.
orach, K. S., Chae, K., Levy, L. A., Daux, W. L., and Sarver, P. J.(1989). Diethylstilbestrol metabolites and analogs: Stereochemi-cal probes for the estrogen receptor binding site. J. Biol. Chem.264, 5642–5647.
Korach, K. S., Horigome, T., Tomooka, Y., Yamashita, S., Newbold,R. R., and McLachlan, J. A. (1988). Immunodetection of estrogenreceptor in epithelial and stromal tissues of neonatal mouseuterus. Proc. Natl. Acad. Sci. USA 85, 3334–3337.
Korach, K. S., Metzler, M., and McLachlan, J. A. (1978). Estrogenicactivity in vivo and in vitro of some diethylstilbestrol metabo-lites and analogs. Proc. Natl. Acad. Sci. USA 75, 468–471.
i, S., Washburn, K. A., Moore, R., Uno, T., Teng, C., Newbold,R. R., McLachlan, J. A., and Negishi, M. (1997). Developmentalexposure to diethylstilbestrol elicits demethylation of estrogen-responsive lactoferrin gene in mouse uterus. Cancer Res. 57,4356–4359.
iehr, J. G. (2000). Is estradiol a genotoxic mutagenic carcinogen?Endocr. Rev. 21, 40–54.
im, H., Ma, L., Ma, W. G., Maas, R. L., and Dey, S. K. (1999).Hoxa-10 regulates uterine stromal cell responsiveness to proges-terone during implantation and decidualization in the mouse.Mol. Endocrinol. 13, 1005–1017.a, L., Benson, G. V., Lim, H. J., Dey, S. K., and Maas, R. L. (1998).Abdominal B (AbdB) Hoxa genes: Regulation in adult uterus byestrogen and progesterone and repression in Mullerian duct by thesynthetic estrogen diethylstilbestrol (DES). Dev. Biol. 197, 141–154.
Maier, D. B., Newbold, R. R., and McLachlan, J. A. (1985). Prenataldiethylstilbestrol exposure alters murine uterine responses to pre-pubertal estrogen stimulation. Endocrinology 116, 1878–1886.
Marselos, M., and Tomatis, L. (1992). Diethylstilboestrol: I. Phar-macology, toxicology and carcinogenicity in humans. Eur. J.Cancer 28A, 1182–1189.arselos, M., and Tomatis, L. (1993). Diethylstilboestrol: II. Phar-macology, toxicology and carcinogenicity in experimental ani-mals. Eur. J. Cancer 29A, 149–155.edlock, K. L., Branham, W. S., and Sheehan, D. M. (1992).Long-term effects of postnatal exposure to diethylstilbestrol onuterine estrogen receptor and growth. J. Steroid Biochem. Mol.Biol. 42, 23–28.edlock, K. L., Sheehan, D. M., Nelson, C. J., and Branham, W. S.(1988). Effects of postnatal DES treatment on uterine growth,development, and estrogen receptor levels. J. Steroid Biochem.Mol. Biol. 29, 527–532.iller, C., Degenhardt, K., and Sassoon, D. A. (1998a). Fetalexposure to DES results in de-regulation of Wnt7a during uterinemorphogenesis. Nat. Genet. 20, 228–230.iller, C., Pavlova, A., and Sassoon, D. A. (1998b). Differentialexpression patterns of Wnt genes in the murine female reproduc-tive tract during development and the estrous cycle. Mech. Dev.76, 91–99.iller, W. R., and Sharpe, R. M. (1998). Environmental oestrogensand human reproductive cancers. Endocr. Rel. Cancer 5, 69–96.ittendorf, R. (1995). Teratogen update: Carcinogenesis and tera-togenesis associated with exposure to diethylstilbestrol (DES) inutero. Teratology 51, 435–445.elson, K. G., Sakai, Y., Eitzman, B., Steed, T., and McLachlan, J.(1994). Exposure to diethylstilbestrol during a critical develop-
Copyright © 2001 by Academic Press. All right
mental period of the mouse reproductive tract leads to persistentinduction of two estrogen-regulated genes. Cell Growth Differ. 5,595–606.ewbold, R. (1995). Cellular and molecular effects of developmen-tal exposure to diethylstilbestrol: Implications for other environ-mental estrogens. Environ. Health Perspect. 103, 83–87.
Newbold, R. R., Bullock, B. C., and McLachlan, J. A. (1985).Progressive proliferative changes in the oviduct of mice follow-ing developmental exposure to diethylstilbestrol. Teratog. Car-cinog. Mutagen 5, 473–480.
Newbold, R. R., Bullock, B. C., and McLachlan, J. A. (1990). Uterineadenocarcinoma in mice following developmental treatmentwith estrogens: A model for hormonal carcinogenesis. CancerRes. 50, 7677–7681.ewbold, R. R., Hanson, R. B., Jefferson, W. N., Bullock, B. C.,Haseman, J., and McLachlan, J. A. (1998). Increased tumors butuncompromised fertility in the female descendants of miceexposed developmentally to diethylstilbestrol. Carcinogenesis19, 1655–1663.
Parr, B. A., and McMahon, A. P. (1998). Sexually dimorphicdevelopment of the mammalian reproductive tract requires Wnt-7a. Nature 395, 707–710.
Prins, G. S. (1992). Neonatal estrogen exposure induces lobe-specific alterations in adult rat prostate androgen receptor ex-pression. Endocrinology 130, 3703–3714.
Prins, G. S., and Birch, L. (1995). The developmental pattern ofandrogen receptor expression in rat prostate lobes is altered afterneonatal exposure to estrogen. Endocrinology 136, 1303–1314.
Salo, L. K., Makela, S. I., Stancel, G. M., and Santti, R. S. (1997).Neonatal exposure to diethylstilbestrol permanently alters thebasal and 17b-estradiol induced expression of c-fos proto-oncogene in mouse urethroprostatic complex. Mol. Cell. Endo-crinol. 126, 133–141.
atokata, I., Benson, G., and Maas, R. (1995). Sexually dimorphicsterility phenotypes in Hoxa10-deficient mice. Nature 374, 460–463.
ibonga, J. D., Dobnig, H., Harden, R. M., and Turner, R. T. (1998).Effect of the high-affinity estrogen receptor ligand ICI 182,780 onthe rat tibia. Endocrinology 139, 3736–3742.
akasugi, N. (1976). Cytological basis for permanent vaginalchanges in mice treated neonatally with steroid hormones. Int.Rev. Cytol. 44, 193–224.aylor, H. S., Arici, A., Olive, D., and Igarashi, P. (1998). HOXA10is expressed in response to sex steroids at the time of implanta-tion in the human endometrium. J. Clin. Invest. 101, 1379–1384.
aylor, H. S., Bagot, C., Kardana, A., Olive, D., and Arici, A. (1999).HOX gene expression is altered in the endometrium of womenwith endometriosis. Hum. Reprod. 14, 1328–1331.
aylor, H. S., Vanden Heuvel, G. B., and Igarashi, P. (1997). Aconserved Hox axis in the mouse and human female reproductivesystem: Late establishment and persistent adult expression of theHoxa cluster genes. Biol. Reprod. 57, 1338–1345.
remblay, G. B., Kunath, T., Bergeron, D., Lapointe, L., Champigny,C., Bader, J.-A., Rossant, J., and Giguere, V. (2001). Diethylstilbes-trol regulates trophoblast stem cell differentiation as a ligand oforphan nuclear receptor ERRb. Genes Dev. 15, 833–838.
Tsutsui, T., Maizumi, H., McLachlan, J. A., and Barrett, J. C. (1983).Aneuploidy induction and cell transformation by diethylstilbes-trol: A possible chromosomal mechanism in carcinogenesis.Cancer Res. 43, 3814–3821.
Turusov, V. S., Trukhanova, L. S., Parfenov, Yu. D., and Tomatis, L.(1992). Occurrence of tumours in the descendants of CBA male
s of reproduction in any form reserved.

W
238 Couse et al.
mice prenatally treated with diethylstilbestrol. Int. J. Cancer 50,131–135.
Walker, B. E. (1984). Tumors of female offspring of mice exposedprenatally to diethylstilbestrol. J. Natl. Cancer Inst. 73, 133–140.
Warot, X., Fromental-Ramain, C., Fraulob, V., Chambon, P., andDolle, P. (1997). Gene dosage-dependent effects of the Hoxa-13and Hoxd-13 mutations on morphogenesis of the terminal partsof the digestive and urogenital tracts. Development 124, 4781–4791.elch, W. R., Fu, Y. S., Robboy, S. J., and Herbst, A. L. (1983).Nuclear DNA content of clear cell adenocarcinoma of the vaginaand cervix and its relationship to prognosis. Gynecol. Oncol. 15,230–238.
Copyright © 2001 by Academic Press. All right
Yamashita, S., Newbold, R. R., McLachlan, J. A., and Korach, K. S.(1989). Developmental pattern of estrogen receptor expression infemale mouse genital tracts. Endocrinology 125, 2888–2896.
Zheng, X., and Hendry III, W. J. (1997). Neonatal diethylstilbestroltreatment alters the estrogen-regulated expression of both cellproliferation and apoptosis-related proto-oncogenes (c-jun, c-fos,c-myc, bax, bcl-2, and bcl-x) in the hamster uterus. Cell GrowthDiffer. 8, 425–434.
Received for publication May 1, 2001Revised July 18, 2001
Accepted July 20, 2001Published online September 7, 2001
s of reproduction in any form reserved.




















