
Oxidative stress-induced apoptosis of cochlear sensory cells: otoprotective strategies
Upload
independentCategory
view
3download
0
Thyroid Hormone Deficiency Before the Onset of Hearing CausesIrreversible Damage to Peripheral and Central Auditory Systems
MARLIES KNIPPER, CHRISTOPH ZINN, HANNES MAIER, MARK PRAETORIUS, KARIN ROHBOCK,IRIS KOPSCHALL, AND ULRIKE ZIMMERMANNDepartment of Oto-Rhino-Laryngology, University of Tu¨bingen, Tu¨bingen Centre for Hearing Research, D-72076 Tu¨bingen,Germany
Knipper, Marlies, Christoph Zinn, Hannes Maier, Mark Praeto-rius, Karin Rohbock, Iris Ko pschall, and Ulrike Zimmermann.Thyroid hormone deficiency before the onset of hearing causes irre-versible damage to peripheral and central auditory systems.J. Neu-rophysiol.83: 3101–3112, 2000. Both a genetic or acquired neonatalthyroid hormone (TH) deficiency may result in a profound mentaldisability that is often accompanied by deafness. The existence ofvarious TH-sensitive periods during inner ear development and gen-eral success of delayed, corrective TH treatment was investigated bytreating pregnant and lactating rats with the goitrogen methimazole(MMI). We observed that for the establishment of normal hearingability, maternal TH, before fetal thyroid gland function on estrusdays 17–18,is obviously not required. Within a crucial time betweenthe onset of fetal thyroid gland function and the onset of hearing atpostnatal day 12(P12), any postponement in the rise of TH-plasmalevels, as can be brought about by treating lactating mothers withMMI, leads to permanent hearing defects of the adult offspring. Theseverity of hearing defects that were measured in 3- to 9-mo-oldoffspring could be increased with each additional day of TH defi-ciency during this critical period. Unexpectedly, the active cochlearprocess, assayed by distortion product otoacoustic emissions(DPOAE) measurements, and speed of auditory brain stem responses,which both until now were not thought to be controlled by TH, provedto be TH-dependent processes that were damaged by a delay of THsupply within this critical time. In contrast, no significant differencesin the gross morphology and innervation of the organ of Corti ormyelin gene expression in the auditory system, detected as myelinbasic protein (MBP) and proteolipid protein (PLP) mRNA usingNorthern blot approach, were observed when TH supply was delayedfor few days. These classical TH-dependent processes, however, weredamaged when TH supply was delayed for several weeks. Thesesurprising results may suggest the existence of different TH-depen-dent processes in the auditory system: those that respond to correctiveTH supply (e.g., innervation and morphogenesis of the organ of Corti)and those that do not, but require T3 activity during a very tight timewindow (e.g., active cochlear process, central processes).
I N T R O D U C T I O N
One and one-half billion people worldwide potentially runthe risk of suffering from iodine-deficiency disorders, includ-ing more than 600 million with goiter. As a clinical disorder,iodine deficiency is most damaging during the critical period ofneural differentiation, causing severe, permanent brain abnor-malities (Bernal and Nunez 1995; Oppenheimer et al. 1994). Inaddition to mental retardation, profound deafness is one of the
noticeable features of congenital thyroid diseases, which maybe associated with neurological cretinism or congenital goiterand is observed in Pendred syndrome (Trotter 1960) or in aresistance to thyroid hormone (RTH), respectively (Refetoff etal. 1967).
The thyroid hormone (TH) is necessary for normal devel-opment of the auditory system (Meyerhoff 1980; Ritter 1967).In rats TH is so far presumed to be active during the first 4 wkpostnatal, when thyroxine deficiency leads to severe defectsboth of neuronal and morphogenetic cochlear processes (Uzielet al. 1983a, 1985a,b). Such defects include the abnormalpersistence of afferent dendrites and absence of efferent inner-vation (Uziel et al. 1983b) or the absence of inner sulcusformation (Uziel et al. 1983a), caused by lack of cell regressionin the great epithelial ridge (GER) of the immature Ko¨lliker‘sorgan (Kolliker 1863). In addition to the postnatal TH-depen-dent period, the expression of thyroid hormone receptors (TR)in the inner ear of rodents before fetal thyroid gland functionsuggests a dependency on maternal TH during earlier periodsof inner ear development (Bradley et al. 1994). Moreover,evidence indicates that TH is also active in the auditory systemin adults (Goldey and Crofton 1998; Parving 1990). The mech-anism by which the thyroid hormone exerts its manifold ac-tions, whether for construction of a normal neuronal network inthe developing brain (see for a review Bernal and Nunez 1995)or for inner ear development (Uziel 1985), has lost much of itsscientific interest due to the common practice in developedcountries of screening newborns for TH deficiencies, thusenabling TH replacement therapy to be carried out, if required.This has long been presumed to be a successful method forcorrecting mental retardation and hearing impairments (Kemp1907). To explore the possibility of various TH-sensitive pe-riods and their effect on inner ear development and to verifythe success of normal inner ear development on delayed TH-supply, we studied the consequences of transient TH-deficientperiods on the development of normal hearing in rodents.Pregnant and lactating rats were therefore treated with thegoitrogen methimazole (MMI). MMI can cross the placenta(Calvo et al. 1990, 1992; Marchant et al. 1977; Mortimeret al. 1997; Sack et al. 1995) and is safely used to suppressplasma-TH levels both in animals and humans (Becks andBurrow 1991; Lind 1997). The neuronal and morphogeneticdevelopment of the inner ear, as well as the auditory function,was analyzed in adult offspring of untreated (control) andMMI-treated dams. We observed that short periods of THdeficiency, when established within the critical developmental
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
31010022-3077/00 $5.00 Copyright © 2000 The American Physiological Society

period before the onset of auditory function atpostnatal day 12(P12), result in permanent hearing impairment. TH-dependentprocesses were elucidated to occur during this time, whichuntil now were not thought to be controlled by TH. On theother hand, well-known TH-dependent processes, such as theformation of the inner sulcus, synaptogenesis of efferent fibersof the organ of Corti, and myelin gene expression, remainedunaffected on retraction of TH during this short critical timeperiod, whereas they were damaged on prolonged periods ofTH deficiency. These results may not only reveal novel targetsfor TH but also give new insights to the participation of TH inthe development of the sensory system and brain.
M E T H O D S
Drug administration and experimental design
Wistar rats were purchased from Interfauna (Tuttlingen, Germany).The antithyroid drug methyl-mercapto-imidazol (MMI; 0.02%) wasadministered to the drinking water of pregnant females beforeembry-onic day 17(E17) to assure that TH levels were suppressed from theonset of fetal thyroid gland function onward, which in rats takes placeon estrusdays 17–18(Bernal and Nunez 1995; Calvo et al. 1990). Thetreatment was continued after birth and stopped betweenP3 and,maximum, P40. After discontinuation of MMI treatment, plasmalevels of thyroxine (T4) and triiodothyronine (T3) returned to normalwithin 2–4 days and pups grew up under standard conditions. Hearingmeasurements of offspring of untreated (control) and MMI-treateddams were determined at 3-, 6-, or 9-mo of age. The neuronal andmorphogenetic development of the inner ear was analyzed in tissuesections when animals were killed at the end of the experimentalperiod. To study the effects of maternal TH, various couples of ratswere paired for 3 or 8 days, separated and continuously treated withMMI either toE17(duringE3 to E17) or to specified days postnatally.The neuronal and morphogenetic inner ear development and thehearing function was analyzed in tissue sections of one to threeoffspring of three MMI-treated dams of each group (TH deficient forthe following times:E3 to E17; E3 to P12; E8 to P12; E17to P12),killed at 3, 6, and 9 mo of age. Hypothyroidism was assessed bydetermination of plasma thyroxine (T4) and triiodothyronine (T3)levels of individual pups, adult offspring, and dams (see next section).
T3/T4 determination
Blood from the pups was collected when the animals were killed forisolation of the cochlea, whereas blood from pregnant dams wascollected on retrobulbar puncture. The blood samples were allowed toclot and centrifuged at 1,000 rpm for 4 min. Serum was collected andfrozen at220°C for later use. Quantification of thyroxine (T4) ortriiodothyronine (3,5,39-L-triiodothyronine, T3) levels determined inrat serum was performed using a CIBA-Corning automated chemilu-minescence system containing ACS-T4 and ACS-T3 ReadyPack Testreagents and a ACS 180 Chemiluminometer. The measurement isbased on a competition of serum T3/T4 with T3, T4 analogues and T3,T4 antibodies.
Tissue preparation
Cochleae of postnatal or adult offspring of untreated (control) andMMI-treated dams were isolated and dissected as previously de-scribed (Knipper et al. 1997, 1998). Cochleae were fixated by immer-sion in 2% paraformaldehyde, 125 mM sucrose in 100 mM phosphate-buffered saline (PBS), pH 7.4, for 2 h, followed by overnightincubation in 25% sucrose, 1 mM protease inhibitor (Pefabloc, Boehr-inger, Mannheim, Germany) in PBS, pH 7.4. Cochleae of animalsolder thanP10 were decalcified after fixation for 15 min to 2 h in
Calex (Fisher Diagnostic, Fair Lawn, NJ). After overnight incubation,cochleae were embedded in O.C.T. compound (Miles Laboratories,Elkhart, IN). Tissues were then cryosectioned at 10mm thickness,mounted on SuperFrost/plus microscope slides, dried for 1 h, andstored at220°C before use.
Immunocytochemical staining
For immunohistochemistry, rat cochlea sections of offspring ofuntreated (control) or MMI-treated dams were thawed, permeabilizedwith 0.1% Triton-X-100 (Sigma) for 3 min at room temperature,preblocked with 1% bovine serum albumin in PBS, and incubatedovernight at 4°C with antibodies. For double labeling studies, bothantibodies were simultaneously incubated for identical time periods.We used antibodies to the 200 K neurofilament protein NF-200 asmarkers for afferent fibers (Berglund and Ryugo 1986; Knipper et al.1997) and antibodies to synaptophysin as markers for presynapses ininner hair cell (IHC) and efferent fibers (Knipper et al. 1995, 1997;Wiechers et al. 1999). Polyclonal antisera for neurofilament 200(N-4142) and monoclonal anti-synaptophysin (clone SVP-38) werepurchased from Sigma. Primary antibodies were detected with eitherCy3-conjugated secondary antibodies (0.35mg/ml; Jackson ImmunoResearch Laboratories, PA) or fluorescein isothiocyanate (FITC)–conjugated secondary antibodies (1:100, Sigma). Sections wererinsed, mounted, and photographed using an Olympus AX70 micro-scope equipped with epifluorescence illumination. Photomicrographswere taken on Kodak TMY-400 film with 400 ASA.
In vitro transcription of myelin basic protein (MBP) andproteolipid protein (PLP) cDNA
MBP cDNA, cloned in pBR322 by Roach et al. (1983) and PLP,cloned in pUC18 by Milner et al. (1985) were subcloned as describedby Schaeren-Wiemers and Gerfin-Moser (1993) and Lemke and Axel(1985). Subcloned cDNAs were kindly provided by M. Frank andM. E. Schwab, University Zurich, Switzerland. MBP cDNA encodeda 2.1 kb mRNA and PLP cDNAs encoded 3.2 and 1.6 kb mRNA. Thedigoxigenin-labeled RNA probes were transcribed from either T7 orT3 promoters for antisense, or sense transcripts in pBlueScript in thepresence of digoxigenin-UTP (Boehringer, Mannheim, Germany).
Northern Blot analysis of MBP and PLP mRNA
Northern Blot analysis was performed as recently described(Gestwa et al. 1999; Knipper et al. 1998). The cartilaginous or bonycapsule was carefully dissected from the cochlea of postnatal orgrown pups of untreated (control) and MMI-treated dams. The openedspiral canals, including the modiolus around which they wind, werecollected, immediately frozen in liquid nitrogen, and stored at270°C.Brain stem, inferior colliculus, and auditory cortex at the level of thetemporal lobe were dissected and immediately frozen in liquid nitro-gen and stored at270°C. Detection of mRNAs was performed witha DIG System as recently described (Rueger et al. 1996). The effectof TH on mRNA levels was evaluated and semiquantified usingisolated mRNAs from a similar number of cochleae as previouslydescribed (Knipper et al. 1998).
Auditory brain stem responses
Auditory evoked brain stem responses (ABR) to clicks were re-corded in anesthetized rat pups or adult offspring (3–9 mo) of eitheruntreated (control) or MMI-treated dams. Anesthesia was performedby intraperitoneal injection of 100 mg/kg ketamin hydrochloride(Ketamin 50 Curamed, CuraMED Pharma, Karlsru¨he, Germany), 15mg/kg xylazin hydrochloride (Rompun 290, Bayer Leverkusen, Ger-many), and 0.5 mg/kg atropin sulfate (Atropinsulfat, Braun, Ger-many). Generation of acoustic stimuli and subsequent recording of
3102 KNIPPER ET AL.
evoked potentials were performed using a Phasis II System (Esaote-biomedica, Italy). Acoustic stimuli, consisting of clicks were deliv-ered monaurally at a rate of 11/s through a TDH-39 earphone (Tele-phonics) connected to a funnel fitted into the external auditory meatus.To record bioelectrical potentials, subdermal silver wire electrodeswere inserted at the vertex (reference), ventrolateral to the measuredear (active) and contralateral to the measured ear (ground). Afteramplification and band-pass filtering (200 Hz to 2 kHz), electricalsignals were averaged over 500 repetitions. The stimulus intensitiesused ranged from 120 dB peak equivalent SPL (peSPL) down to ABRthreshold (defined as the lowest intensity at which a response could berecorded) in 5-dB steps. Approaching threshold, three replicationswere conducted. The thresholds were confirmed afterward by anindependent observer and found not to differ by.5 dB.
Measurements of distortion product otoacoustic emissions(DPOAE)
For measurements of DPOAE, acoustic stimulation and measure-ment were performed by a PC-based system with an A/D converterboard using Labview software, which also performed fast Fouriertransformation (FFT) for data analysis. Pure tone continuous soundstimuli were generated and delivered on two separate channels usingdynamic speakers (Beyer DT48). Sound levels were measured with aprobe microphone (Bruel and Kjaer 4135) and a measuring amplifier(Bruel and Kjaer 2610). The speakers and microphone were coupledtogether, closed field into the animal’s ear canal, and the sound systemcalibrated with white noise (50 Hz to 20 kHz). DPOAEs were mea-sured as cubic distortion products at the frequency 2f1-f2. For eachfrequency f1 (7, 12, or 18 kHz), the individual best ratio (BR) f2/f1was determined using an intermediate sound level (60 dB SPL orhigher, if necessary), varying the ratio from 1.06 to 1.44 in steps of0.02. This ratio was used for the following input/output measurement.The sound level of f1 was kept at a constant 10 dB louder than the f2level. Sound was initially presented at the lowest level and increasedin steps of 5 dB. Recording windows were averaged so as to decreasethe background noise to at least 10 dB below the level of emission, butnot over more than 1,000 repetitions.
Experimental animals
The care and use of the animals reported in this study was approvedby the University’s “special animal care” following the guidelines ofthe Declaration of Helsinki.
R E S U L T S
Use of the MMI to control the plasma-TH level
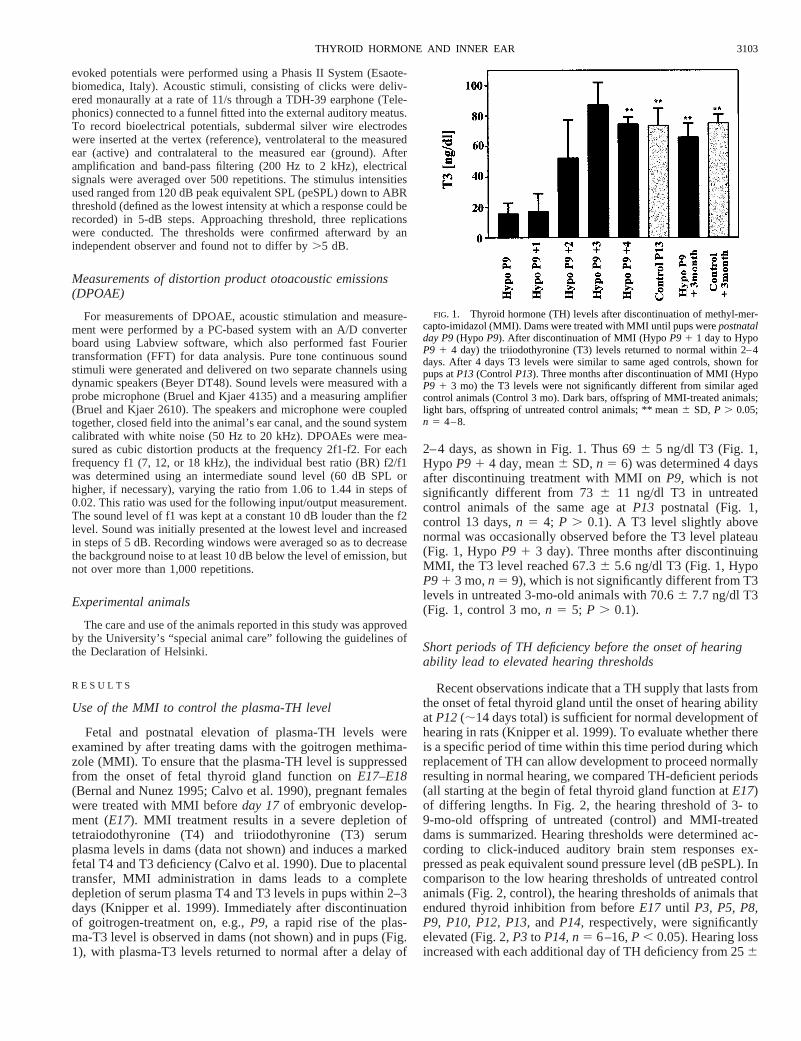
Fetal and postnatal elevation of plasma-TH levels wereexamined by after treating dams with the goitrogen methima-zole (MMI). To ensure that the plasma-TH level is suppressedfrom the onset of fetal thyroid gland function onE17–E18(Bernal and Nunez 1995; Calvo et al. 1990), pregnant femaleswere treated with MMI beforeday 17of embryonic develop-ment (E17). MMI treatment results in a severe depletion oftetraiodothyronine (T4) and triiodothyronine (T3) serumplasma levels in dams (data not shown) and induces a markedfetal T4 and T3 deficiency (Calvo et al. 1990). Due to placentaltransfer, MMI administration in dams leads to a completedepletion of serum plasma T4 and T3 levels in pups within 2–3days (Knipper et al. 1999). Immediately after discontinuationof goitrogen-treatment on, e.g.,P9, a rapid rise of the plas-ma-T3 level is observed in dams (not shown) and in pups (Fig.1), with plasma-T3 levels returned to normal after a delay of
2–4 days, as shown in Fig. 1. Thus 696 5 ng/dl T3 (Fig. 1,Hypo P9 1 4 day, mean6 SD,n 5 6) was determined 4 daysafter discontinuing treatment with MMI onP9, which is notsignificantly different from 736 11 ng/dl T3 in untreatedcontrol animals of the same age atP13 postnatal (Fig. 1,control 13 days,n 5 4; P . 0.1). A T3 level slightly abovenormal was occasionally observed before the T3 level plateau(Fig. 1, HypoP9 1 3 day). Three months after discontinuingMMI, the T3 level reached 67.36 5.6 ng/dl T3 (Fig. 1, HypoP91 3 mo,n 5 9), which is not significantly different from T3levels in untreated 3-mo-old animals with 70.66 7.7 ng/dl T3(Fig. 1, control 3 mo,n 5 5; P . 0.1).
Short periods of TH deficiency before the onset of hearingability lead to elevated hearing thresholds
Recent observations indicate that a TH supply that lasts fromthe onset of fetal thyroid gland until the onset of hearing abilityatP12(;14 days total) is sufficient for normal development ofhearing in rats (Knipper et al. 1999). To evaluate whether thereis a specific period of time within this time period during whichreplacement of TH can allow development to proceed normallyresulting in normal hearing, we compared TH-deficient periods(all starting at the begin of fetal thyroid gland function atE17)of differing lengths. In Fig. 2, the hearing threshold of 3- to9-mo-old offspring of untreated (control) and MMI-treateddams is summarized. Hearing thresholds were determined ac-cording to click-induced auditory brain stem responses ex-pressed as peak equivalent sound pressure level (dB peSPL). Incomparison to the low hearing thresholds of untreated controlanimals (Fig. 2, control), the hearing thresholds of animals thatendured thyroid inhibition from beforeE17 until P3, P5, P8,P9, P10, P12, P13,and P14, respectively, were significantlyelevated (Fig. 2,P3 to P14, n5 6–16,P , 0.05). Hearing lossincreased with each additional day of TH deficiency from 256
FIG. 1. Thyroid hormone (TH) levels after discontinuation of methyl-mer-capto-imidazol (MMI). Dams were treated with MMI until pups werepostnatalday P9(Hypo P9). After discontinuation of MMI (HypoP9 1 1 day to HypoP9 1 4 day) the triiodothyronine (T3) levels returned to normal within 2–4days. After 4 days T3 levels were similar to same aged controls, shown forpups atP13(ControlP13). Three months after discontinuation of MMI (HypoP9 1 3 mo) the T3 levels were not significantly different from similar agedcontrol animals (Control 3 mo). Dark bars, offspring of MMI-treated animals;light bars, offspring of untreated control animals; ** mean6 SD, P . 0.05;n 5 4–8.
3103THYROID HORMONE AND INNER EAR
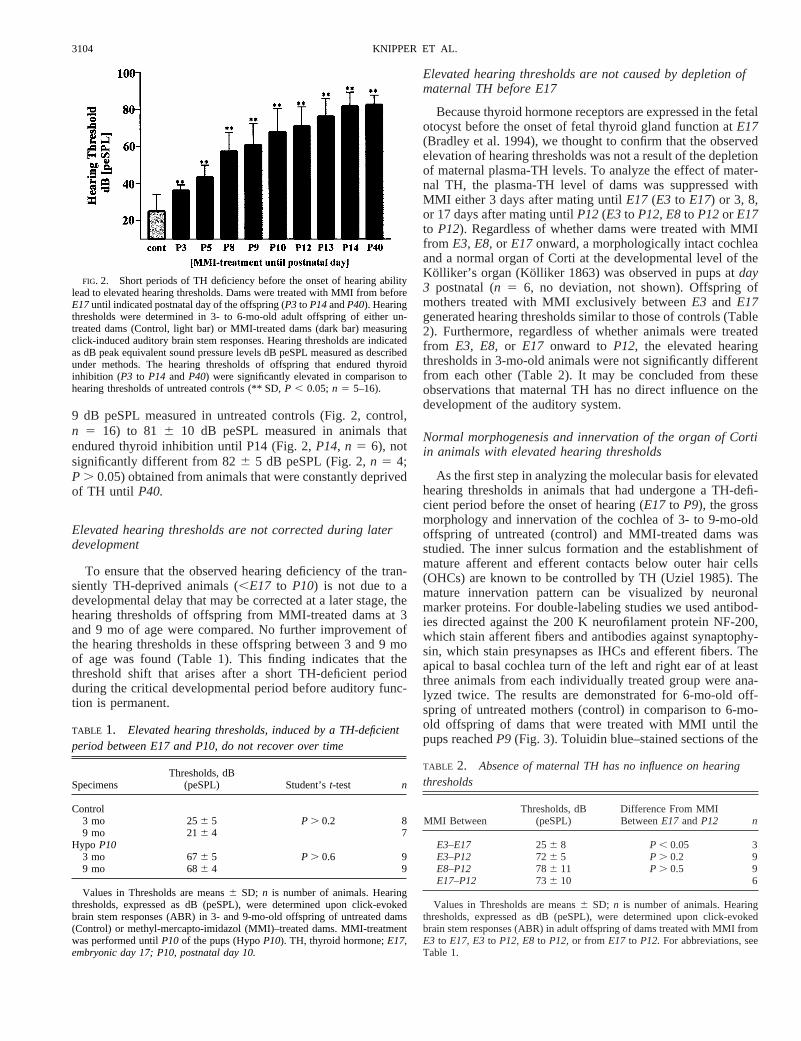
9 dB peSPL measured in untreated controls (Fig. 2, control,n 5 16) to 81 6 10 dB peSPL measured in animals thatendured thyroid inhibition until P14 (Fig. 2,P14, n5 6), notsignificantly different from 826 5 dB peSPL (Fig. 2,n 5 4;P . 0.05) obtained from animals that were constantly deprivedof TH until P40.
Elevated hearing thresholds are not corrected during laterdevelopment
To ensure that the observed hearing deficiency of the tran-siently TH-deprived animals (,E17 to P10) is not due to adevelopmental delay that may be corrected at a later stage, thehearing thresholds of offspring from MMI-treated dams at 3and 9 mo of age were compared. No further improvement ofthe hearing thresholds in these offspring between 3 and 9 moof age was found (Table 1). This finding indicates that thethreshold shift that arises after a short TH-deficient periodduring the critical developmental period before auditory func-tion is permanent.
Elevated hearing thresholds are not caused by depletion ofmaternal TH before E17
Because thyroid hormone receptors are expressed in the fetalotocyst before the onset of fetal thyroid gland function atE17(Bradley et al. 1994), we thought to confirm that the observedelevation of hearing thresholds was not a result of the depletionof maternal plasma-TH levels. To analyze the effect of mater-nal TH, the plasma-TH level of dams was suppressed withMMI either 3 days after mating untilE17 (E3 to E17) or 3, 8,or 17 days after mating untilP12(E3 to P12, E8to P12or E17to P12). Regardless of whether dams were treated with MMIfrom E3, E8,or E17onward, a morphologically intact cochleaand a normal organ of Corti at the developmental level of theKolliker’s organ (Kolliker 1863) was observed in pups atday3 postnatal (n 5 6, no deviation, not shown). Offspring ofmothers treated with MMI exclusively betweenE3 and E17generated hearing thresholds similar to those of controls (Table2). Furthermore, regardless of whether animals were treatedfrom E3, E8, or E17 onward to P12, the elevated hearingthresholds in 3-mo-old animals were not significantly differentfrom each other (Table 2). It may be concluded from theseobservations that maternal TH has no direct influence on thedevelopment of the auditory system.
Normal morphogenesis and innervation of the organ of Cortiin animals with elevated hearing thresholds
As the first step in analyzing the molecular basis for elevatedhearing thresholds in animals that had undergone a TH-defi-cient period before the onset of hearing (E17 to P9), the grossmorphology and innervation of the cochlea of 3- to 9-mo-oldoffspring of untreated (control) and MMI-treated dams wasstudied. The inner sulcus formation and the establishment ofmature afferent and efferent contacts below outer hair cells(OHCs) are known to be controlled by TH (Uziel 1985). Themature innervation pattern can be visualized by neuronalmarker proteins. For double-labeling studies we used antibod-ies directed against the 200 K neurofilament protein NF-200,which stain afferent fibers and antibodies against synaptophy-sin, which stain presynapses as IHCs and efferent fibers. Theapical to basal cochlea turn of the left and right ear of at leastthree animals from each individually treated group were ana-lyzed twice. The results are demonstrated for 6-mo-old off-spring of untreated mothers (control) in comparison to 6-mo-old offspring of dams that were treated with MMI until thepups reachedP9 (Fig. 3). Toluidin blue–stained sections of the
TABLE 2. Absence of maternal TH has no influence on hearingthresholds
MMI BetweenThresholds, dB
(peSPL)Difference From MMIBetweenE17 andP12 n
E3–E17 256 8 P , 0.05 3E3–P12 726 5 P . 0.2 9E8–P12 786 11 P . 0.5 9E17–P12 736 10 6
Values in Thresholds are means6 SD; n is number of animals. Hearingthresholds, expressed as dB (peSPL), were determined upon click-evokedbrain stem responses (ABR) in adult offspring of dams treated with MMI fromE3 to E17, E3to P12, E8to P12,or from E17 to P12.For abbreviations, seeTable 1.
FIG. 2. Short periods of TH deficiency before the onset of hearing abilitylead to elevated hearing thresholds. Dams were treated with MMI from beforeE17until indicated postnatal day of the offspring (P3 to P14andP40). Hearingthresholds were determined in 3- to 6-mo-old adult offspring of either un-treated dams (Control, light bar) or MMI-treated dams (dark bar) measuringclick-induced auditory brain stem responses. Hearing thresholds are indicatedas dB peak equivalent sound pressure levels dB peSPL measured as describedunder methods. The hearing thresholds of offspring that endured thyroidinhibition (P3 to P14 andP40) were significantly elevated in comparison tohearing thresholds of untreated controls (** SD,P , 0.05;n 5 5–16).
TABLE 1. Elevated hearing thresholds, induced by a TH-deficientperiod between E17 and P10, do not recover over time
SpecimensThresholds, dB
(peSPL) Student’st-test n
Control3 mo 256 5 P . 0.2 89 mo 216 4 7
Hypo P103 mo 676 5 P . 0.6 99 mo 686 4 9
Values in Thresholds are means6 SD; n is number of animals. Hearingthresholds, expressed as dB (peSPL), were determined upon click-evokedbrain stem responses (ABR) in 3- and 9-mo-old offspring of untreated dams(Control) or methyl-mercapto-imidazol (MMI)–treated dams. MMI-treatmentwas performed untilP10of the pups (HypoP10). TH, thyroid hormone;E17,embryonic day 17; P10, postnatal day 10.
3104 KNIPPER ET AL.
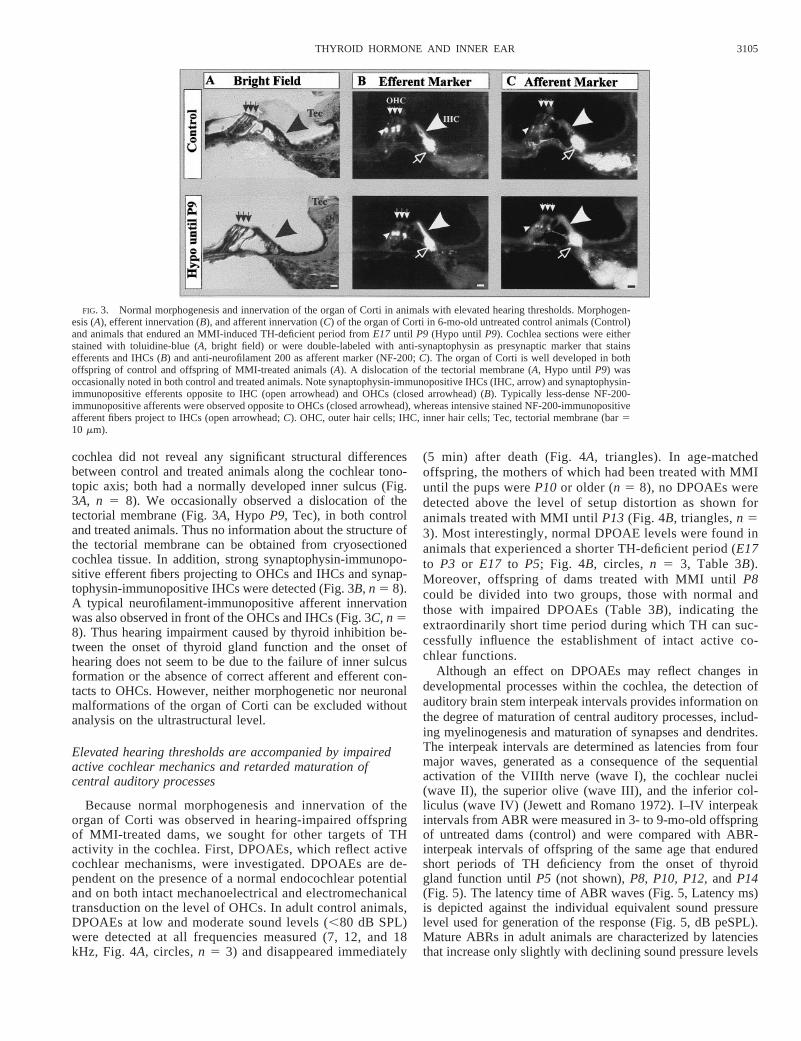
cochlea did not reveal any significant structural differencesbetween control and treated animals along the cochlear tono-topic axis; both had a normally developed inner sulcus (Fig.3A, n 5 8). We occasionally observed a dislocation of thetectorial membrane (Fig. 3A, Hypo P9, Tec), in both controland treated animals. Thus no information about the structure ofthe tectorial membrane can be obtained from cryosectionedcochlea tissue. In addition, strong synaptophysin-immunopo-sitive efferent fibers projecting to OHCs and IHCs and synap-tophysin-immunopositive IHCs were detected (Fig. 3B, n 5 8).A typical neurofilament-immunopositive afferent innervationwas also observed in front of the OHCs and IHCs (Fig. 3C, n 58). Thus hearing impairment caused by thyroid inhibition be-tween the onset of thyroid gland function and the onset ofhearing does not seem to be due to the failure of inner sulcusformation or the absence of correct afferent and efferent con-tacts to OHCs. However, neither morphogenetic nor neuronalmalformations of the organ of Corti can be excluded withoutanalysis on the ultrastructural level.
Elevated hearing thresholds are accompanied by impairedactive cochlear mechanics and retarded maturation ofcentral auditory processes
Because normal morphogenesis and innervation of theorgan of Corti was observed in hearing-impaired offspringof MMI-treated dams, we sought for other targets of THactivity in the cochlea. First, DPOAEs, which reflect activecochlear mechanisms, were investigated. DPOAEs are de-pendent on the presence of a normal endocochlear potentialand on both intact mechanoelectrical and electromechanicaltransduction on the level of OHCs. In adult control animals,DPOAEs at low and moderate sound levels (,80 dB SPL)were detected at all frequencies measured (7, 12, and 18kHz, Fig. 4A, circles,n 5 3) and disappeared immediately
(5 min) after death (Fig. 4A, triangles). In age-matchedoffspring, the mothers of which had been treated with MMIuntil the pups wereP10 or older (n 5 8), no DPOAEs weredetected above the level of setup distortion as shown foranimals treated with MMI untilP13 (Fig. 4B, triangles,n 53). Most interestingly, normal DPOAE levels were found inanimals that experienced a shorter TH-deficient period (E17to P3 or E17 to P5; Fig. 4B, circles, n 5 3, Table 3B).Moreover, offspring of dams treated with MMI untilP8could be divided into two groups, those with normal andthose with impaired DPOAEs (Table 3B), indicating theextraordinarily short time period during which TH can suc-cessfully influence the establishment of intact active co-chlear functions.
Although an effect on DPOAEs may reflect changes indevelopmental processes within the cochlea, the detection ofauditory brain stem interpeak intervals provides information onthe degree of maturation of central auditory processes, includ-ing myelinogenesis and maturation of synapses and dendrites.The interpeak intervals are determined as latencies from fourmajor waves, generated as a consequence of the sequentialactivation of the VIIIth nerve (wave I), the cochlear nuclei(wave II), the superior olive (wave III), and the inferior col-liculus (wave IV) (Jewett and Romano 1972). I–IV interpeakintervals from ABR were measured in 3- to 9-mo-old offspringof untreated dams (control) and were compared with ABR-interpeak intervals of offspring of the same age that enduredshort periods of TH deficiency from the onset of thyroidgland function untilP5 (not shown),P8, P10, P12,and P14(Fig. 5). The latency time of ABR waves (Fig. 5, Latency ms)is depicted against the individual equivalent sound pressurelevel used for generation of the response (Fig. 5, dB peSPL).Mature ABRs in adult animals are characterized by latenciesthat increase only slightly with declining sound pressure levels
FIG. 3. Normal morphogenesis and innervation of the organ of Corti in animals with elevated hearing thresholds. Morphogen-esis (A), efferent innervation (B), and afferent innervation (C) of the organ of Corti in 6-mo-old untreated control animals (Control)and animals that endured an MMI-induced TH-deficient period fromE17 until P9 (Hypo until P9). Cochlea sections were eitherstained with toluidine-blue (A, bright field) or were double-labeled with anti-synaptophysin as presynaptic marker that stainsefferents and IHCs (B) and anti-neurofilament 200 as afferent marker (NF-200;C). The organ of Corti is well developed in bothoffspring of control and offspring of MMI-treated animals (A). A dislocation of the tectorial membrane (A, Hypo until P9) wasoccasionally noted in both control and treated animals. Note synaptophysin-immunopositive IHCs (IHC, arrow) and synaptophysin-immunopositive efferents opposite to IHC (open arrowhead) and OHCs (closed arrowhead) (B). Typically less-dense NF-200-immunopositive afferents were observed opposite to OHCs (closed arrowhead), whereas intensive stained NF-200-immunopositiveafferent fibers project to IHCs (open arrowhead;C). OHC, outer hair cells; IHC, inner hair cells; Tec, tectorial membrane (bar510 mm).
3105THYROID HORMONE AND INNER EAR
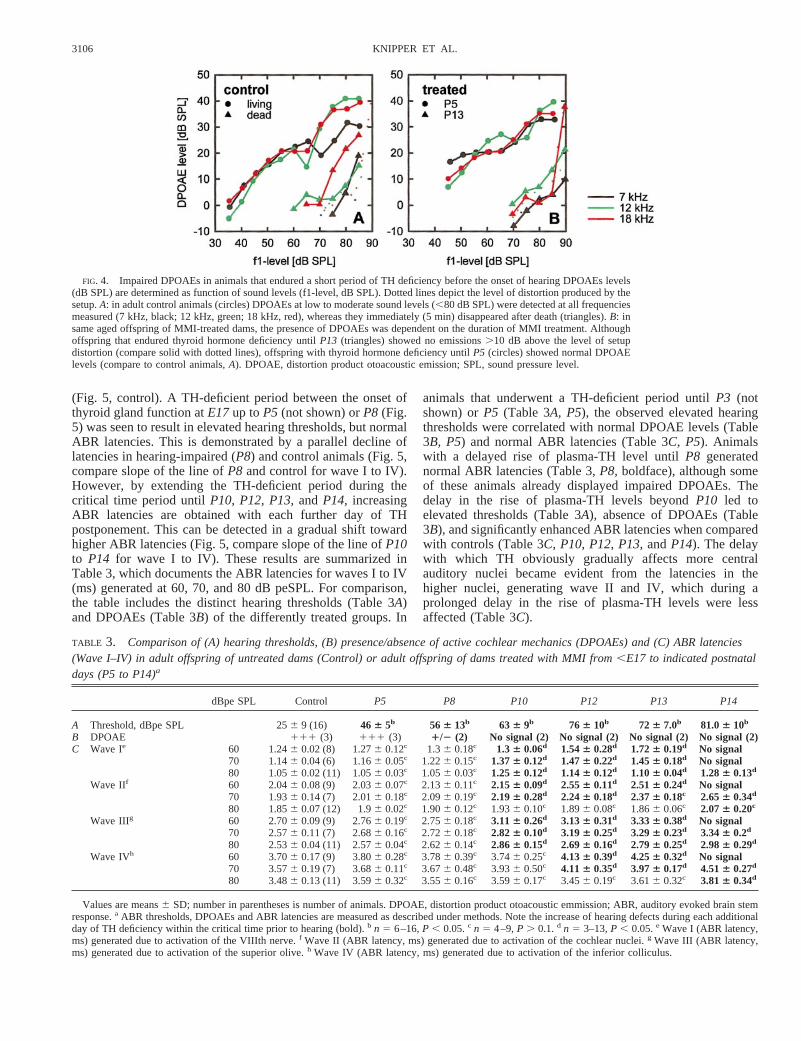
(Fig. 5, control). A TH-deficient period between the onset ofthyroid gland function atE17up toP5 (not shown) orP8 (Fig.5) was seen to result in elevated hearing thresholds, but normalABR latencies. This is demonstrated by a parallel decline oflatencies in hearing-impaired (P8) and control animals (Fig. 5,compare slope of the line ofP8 and control for wave I to IV).However, by extending the TH-deficient period during thecritical time period untilP10, P12, P13,andP14, increasingABR latencies are obtained with each further day of THpostponement. This can be detected in a gradual shift towardhigher ABR latencies (Fig. 5, compare slope of the line ofP10to P14 for wave I to IV). These results are summarized inTable 3, which documents the ABR latencies for waves I to IV(ms) generated at 60, 70, and 80 dB peSPL. For comparison,the table includes the distinct hearing thresholds (Table 3A)and DPOAEs (Table 3B) of the differently treated groups. In
animals that underwent a TH-deficient period untilP3 (notshown) orP5 (Table 3A, P5), the observed elevated hearingthresholds were correlated with normal DPOAE levels (Table3B, P5) and normal ABR latencies (Table 3C, P5). Animalswith a delayed rise of plasma-TH level untilP8 generatednormal ABR latencies (Table 3,P8, boldface), although someof these animals already displayed impaired DPOAEs. Thedelay in the rise of plasma-TH levels beyondP10 led toelevated thresholds (Table 3A), absence of DPOAEs (Table3B), and significantly enhanced ABR latencies when comparedwith controls (Table 3C, P10, P12, P13,andP14). The delaywith which TH obviously gradually affects more centralauditory nuclei became evident from the latencies in thehigher nuclei, generating wave II and IV, which during aprolonged delay in the rise of plasma-TH levels were lessaffected (Table 3C).
FIG. 4. Impaired DPOAEs in animals that endured a short period of TH deficiency before the onset of hearing DPOAEs levels(dB SPL) are determined as function of sound levels (f1-level, dB SPL). Dotted lines depict the level of distortion produced by thesetup.A: in adult control animals (circles) DPOAEs at low to moderate sound levels (,80 dB SPL) were detected at all frequenciesmeasured (7 kHz, black; 12 kHz, green; 18 kHz, red), whereas they immediately (5 min) disappeared after death (triangles).B: insame aged offspring of MMI-treated dams, the presence of DPOAEs was dependent on the duration of MMI treatment. Althoughoffspring that endured thyroid hormone deficiency untilP13 (triangles) showed no emissions.10 dB above the level of setupdistortion (compare solid with dotted lines), offspring with thyroid hormone deficiency untilP5 (circles) showed normal DPOAElevels (compare to control animals,A). DPOAE, distortion product otoacoustic emission; SPL, sound pressure level.
TABLE 3. Comparison of (A) hearing thresholds, (B) presence/absence of active cochlear mechanics (DPOAEs) and (C) ABR latencies(Wave I–IV) in adult offspring of untreated dams (Control) or adult offspring of dams treated with MMI from,E17 to indicated postnataldays (P5 to P14)a
dBpe SPL Control P5 P8 P10 P12 P13 P14
A Threshold, dBpe SPL 256 9 (16) 46 6 5b 56 6 13b 63 6 9b 76 6 10b 72 6 7.0b 81.06 10b
B DPOAE 111 (3) 111 (3) 1/2 (2) No signal (2) No signal (2) No signal (2) No signal (2)C Wave Ie 60 1.246 0.02 (8) 1.276 0.12c 1.36 0.18c 1.36 0.06d 1.546 0.28d 1.726 0.19d No signal
70 1.146 0.04 (6) 1.166 0.05c 1.226 0.15c 1.376 0.12d 1.476 0.22d 1.456 0.18d No signal80 1.056 0.02 (11) 1.056 0.03c 1.056 0.03c 1.256 0.12d 1.146 0.12d 1.106 0.04d 1.286 0.13d
Wave IIf 60 2.046 0.08 (9) 2.036 0.07c 2.136 0.11c 2.156 0.09d 2.556 0.11d 2.516 0.24d No signal70 1.936 0.14 (7) 2.016 0.18c 2.096 0.19c 2.196 0.28d 2.246 0.18d 2.376 0.18c 2.656 0.34d
80 1.856 0.07 (12) 1.96 0.02c 1.906 0.12c 1.936 0.10c 1.896 0.08c 1.866 0.06c 2.076 0.20c
Wave IIIg 60 2.706 0.09 (9) 2.766 0.19c 2.756 0.18c 3.116 0.26d 3.136 0.31d 3.336 0.38d No signal70 2.576 0.11 (7) 2.686 0.16c 2.726 0.18c 2.826 0.10d 3.196 0.25d 3.296 0.23d 3.346 0.2d
80 2.536 0.04 (11) 2.576 0.04c 2.626 0.14c 2.866 0.15d 2.696 0.16d 2.796 0.25d 2.986 0.29d
Wave IVh 60 3.706 0.17 (9) 3.806 0.28c 3.786 0.39c 3.746 0.25c 4.136 0.39d 4.256 0.32d No signal70 3.576 0.19 (7) 3.686 0.11c 3.676 0.48c 3.936 0.50c 4.116 0.35d 3.976 0.17d 4.516 0.27d
80 3.486 0.13 (11) 3.596 0.32c 3.556 0.16c 3.596 0.17c 3.456 0.19c 3.616 0.32c 3.816 0.34d
Values are means6 SD; number in parentheses is number of animals. DPOAE, distortion product otoacoustic emmission; ABR, auditory evoked brain stemresponse.a ABR thresholds, DPOAEs and ABR latencies are measured as described under methods. Note the increase of hearing defects during each additionalday of TH deficiency within the critical time prior to hearing (bold).b n 5 6–16,P , 0.05.c n 5 4–9,P . 0.1.d n 5 3–13,P , 0.05.e Wave I (ABR latency,ms) generated due to activation of the VIIIth nerve.f Wave II (ABR latency, ms) generated due to activation of the cochlear nuclei.g Wave III (ABR latency,ms) generated due to activation of the superior olive.h Wave IV (ABR latency, ms) generated due to activation of the inferior colliculus.
3106 KNIPPER ET AL.
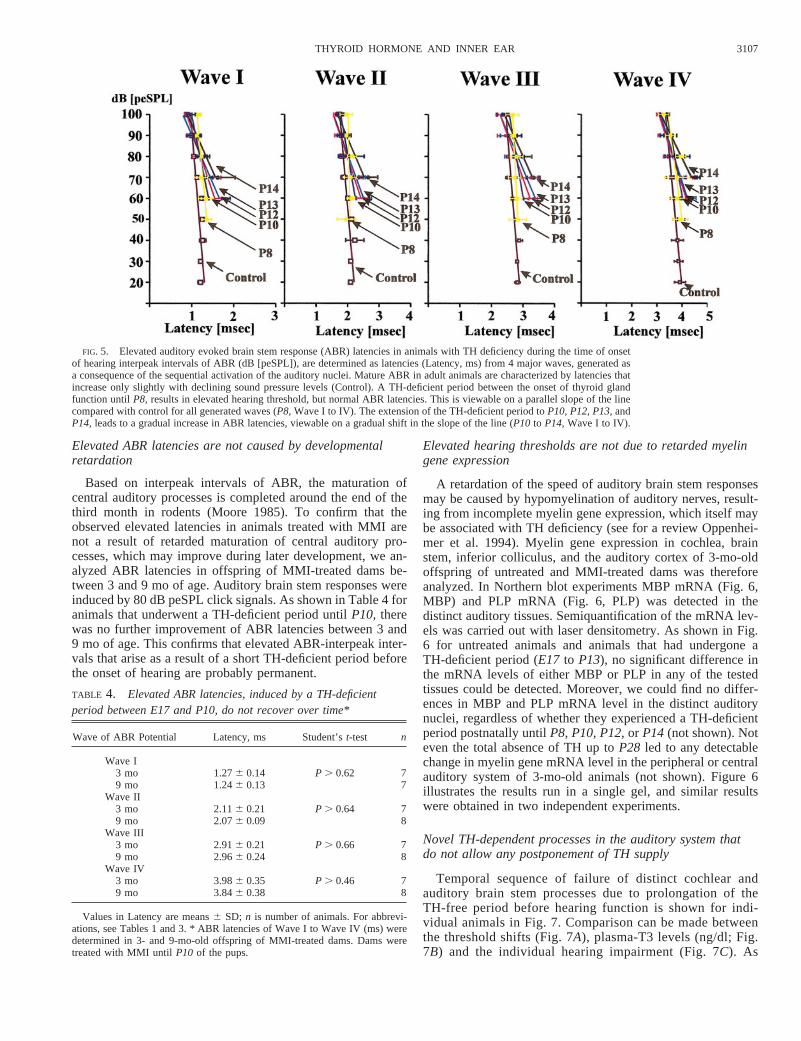
Elevated ABR latencies are not caused by developmentalretardation
Based on interpeak intervals of ABR, the maturation ofcentral auditory processes is completed around the end of thethird month in rodents (Moore 1985). To confirm that theobserved elevated latencies in animals treated with MMI arenot a result of retarded maturation of central auditory pro-cesses, which may improve during later development, we an-alyzed ABR latencies in offspring of MMI-treated dams be-tween 3 and 9 mo of age. Auditory brain stem responses wereinduced by 80 dB peSPL click signals. As shown in Table 4 foranimals that underwent a TH-deficient period untilP10, therewas no further improvement of ABR latencies between 3 and9 mo of age. This confirms that elevated ABR-interpeak inter-vals that arise as a result of a short TH-deficient period beforethe onset of hearing are probably permanent.
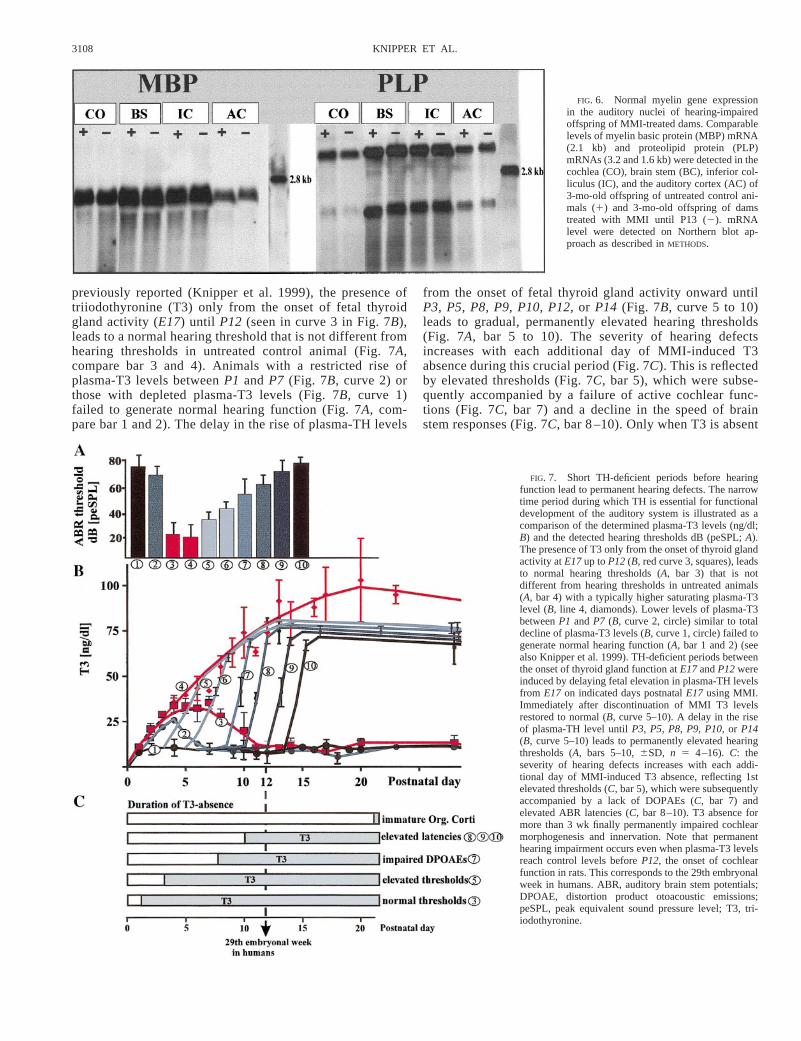
Elevated hearing thresholds are not due to retarded myelingene expression
A retardation of the speed of auditory brain stem responsesmay be caused by hypomyelination of auditory nerves, result-ing from incomplete myelin gene expression, which itself maybe associated with TH deficiency (see for a review Oppenhei-mer et al. 1994). Myelin gene expression in cochlea, brainstem, inferior colliculus, and the auditory cortex of 3-mo-oldoffspring of untreated and MMI-treated dams was thereforeanalyzed. In Northern blot experiments MBP mRNA (Fig. 6,MBP) and PLP mRNA (Fig. 6, PLP) was detected in thedistinct auditory tissues. Semiquantification of the mRNA lev-els was carried out with laser densitometry. As shown in Fig.6 for untreated animals and animals that had undergone aTH-deficient period (E17 to P13), no significant difference inthe mRNA levels of either MBP or PLP in any of the testedtissues could be detected. Moreover, we could find no differ-ences in MBP and PLP mRNA level in the distinct auditorynuclei, regardless of whether they experienced a TH-deficientperiod postnatally untilP8, P10, P12,or P14(not shown). Noteven the total absence of TH up toP28 led to any detectablechange in myelin gene mRNA level in the peripheral or centralauditory system of 3-mo-old animals (not shown). Figure 6illustrates the results run in a single gel, and similar resultswere obtained in two independent experiments.
Novel TH-dependent processes in the auditory system thatdo not allow any postponement of TH supply
Temporal sequence of failure of distinct cochlear andauditory brain stem processes due to prolongation of theTH-free period before hearing function is shown for indi-vidual animals in Fig. 7. Comparison can be made betweenthe threshold shifts (Fig. 7A), plasma-T3 levels (ng/dl; Fig.7B) and the individual hearing impairment (Fig. 7C). As
FIG. 5. Elevated auditory evoked brain stem response (ABR) latencies in animals with TH deficiency during the time of onsetof hearing interpeak intervals of ABR (dB [peSPL]), are determined as latencies (Latency, ms) from 4 major waves, generated asa consequence of the sequential activation of the auditory nuclei. Mature ABR in adult animals are characterized by latencies thatincrease only slightly with declining sound pressure levels (Control). A TH-deficient period between the onset of thyroid glandfunction untilP8, results in elevated hearing threshold, but normal ABR latencies. This is viewable on a parallel slope of the linecompared with control for all generated waves (P8,Wave I to IV). The extension of the TH-deficient period toP10, P12, P13,andP14, leads to a gradual increase in ABR latencies, viewable on a gradual shift in the slope of the line (P10 to P14,Wave I to IV).
TABLE 4. Elevated ABR latencies, induced by a TH-deficientperiod between E17 and P10, do not recover over time*
Wave of ABR Potential Latency, ms Student’st-test n
Wave I3 mo 1.276 0.14 P . 0.62 79 mo 1.246 0.13 7
Wave II3 mo 2.116 0.21 P . 0.64 79 mo 2.076 0.09 8
Wave III3 mo 2.916 0.21 P . 0.66 79 mo 2.966 0.24 8
Wave IV3 mo 3.986 0.35 P . 0.46 79 mo 3.846 0.38 8
Values in Latency are means6 SD; n is number of animals. For abbrevi-ations, see Tables 1 and 3. * ABR latencies of Wave I to Wave IV (ms) weredetermined in 3- and 9-mo-old offspring of MMI-treated dams. Dams weretreated with MMI untilP10 of the pups.
3107THYROID HORMONE AND INNER EAR
previously reported (Knipper et al. 1999), the presence oftriiodothyronine (T3) only from the onset of fetal thyroidgland activity (E17) until P12 (seen in curve 3 in Fig. 7B),leads to a normal hearing threshold that is not different fromhearing thresholds in untreated control animal (Fig. 7A,compare bar 3 and 4). Animals with a restricted rise ofplasma-T3 levels betweenP1 and P7 (Fig. 7B, curve 2) orthose with depleted plasma-T3 levels (Fig. 7B, curve 1)failed to generate normal hearing function (Fig. 7A, com-pare bar 1 and 2). The delay in the rise of plasma-TH levels
from the onset of fetal thyroid gland activity onward untilP3, P5, P8, P9, P10, P12,or P14 (Fig. 7B, curve 5 to 10)leads to gradual, permanently elevated hearing thresholds(Fig. 7A, bar 5 to 10). The severity of hearing defectsincreases with each additional day of MMI-induced T3absence during this crucial period (Fig. 7C). This is reflectedby elevated thresholds (Fig. 7C, bar 5), which were subse-quently accompanied by a failure of active cochlear func-tions (Fig. 7C, bar 7) and a decline in the speed of brainstem responses (Fig. 7C, bar 8 –10). Only when T3 is absent
FIG. 6. Normal myelin gene expressionin the auditory nuclei of hearing-impairedoffspring of MMI-treated dams. Comparablelevels of myelin basic protein (MBP) mRNA(2.1 kb) and proteolipid protein (PLP)mRNAs (3.2 and 1.6 kb) were detected in thecochlea (CO), brain stem (BC), inferior col-liculus (IC), and the auditory cortex (AC) of3-mo-old offspring of untreated control ani-mals (1) and 3-mo-old offspring of damstreated with MMI until P13 (2). mRNAlevel were detected on Northern blot ap-proach as described inMETHODS.
FIG. 7. Short TH-deficient periods before hearingfunction lead to permanent hearing defects. The narrowtime period during which TH is essential for functionaldevelopment of the auditory system is illustrated as acomparison of the determined plasma-T3 levels (ng/dl;B) and the detected hearing thresholds dB (peSPL;A).The presence of T3 only from the onset of thyroid glandactivity atE17up toP12(B, red curve 3, squares), leadsto normal hearing thresholds (A, bar 3) that is notdifferent from hearing thresholds in untreated animals(A, bar 4) with a typically higher saturating plasma-T3level (B, line 4, diamonds). Lower levels of plasma-T3betweenP1 andP7 (B, curve 2, circle) similar to totaldecline of plasma-T3 levels (B, curve 1, circle) failed togenerate normal hearing function (A, bar 1 and 2) (seealso Knipper et al. 1999). TH-deficient periods betweenthe onset of thyroid gland function atE17andP12wereinduced by delaying fetal elevation in plasma-TH levelsfrom E17 on indicated days postnatalE17 using MMI.Immediately after discontinuation of MMI T3 levelsrestored to normal (B, curve 5–10). A delay in the riseof plasma-TH level untilP3, P5, P8, P9, P10,or P14(B, curve 5–10) leads to permanently elevated hearingthresholds (A, bars 5–10,6SD, n 5 4–16). C: theseverity of hearing defects increases with each addi-tional day of MMI-induced T3 absence, reflecting 1stelevated thresholds (C, bar 5), which were subsequentlyaccompanied by a lack of DOPAEs (C, bar 7) andelevated ABR latencies (C, bar 8–10). T3 absence formore than 3 wk finally permanently impaired cochlearmorphogenesis and innervation. Note that permanenthearing impairment occurs even when plasma-T3 levelsreach control levels beforeP12, the onset of cochlearfunction in rats. This corresponds to the 29th embryonalweek in humans. ABR, auditory brain stem potentials;DPOAE, distortion product otoacoustic emissions;peSPL, peak equivalent sound pressure level; T3, tri-iodothyronine.
3108 KNIPPER ET AL.

for more than 3 wk postnatally, cochlear morphogenesis andinnervation is permanently impaired (Fig. 7C, immatureOrg. Corti).
D I S C U S S I O N
The discovery of a thyroid extract therapy to cure mentaldiseases that are associated with cretinism by Murray and itsdescription by Sir William Osler dates back to 1891 (Murray1891; Osler 1889). The first indications that TH is also essen-tial for auditory functions were soon to follow, and replace-ment therapy with thyroid extracts was reported to have thecapability of improving hearing defects as well as the growthand mental development of infants suffering from cretinism(Kemp 1907). Approximately 60 yr later, this concept wasqueried and the proposal made that under distinct conditions,hearing dysfunction may be permanent despite early treatmentwith thyroid hormone (Deol 1973). However, later findingsdefined the corrective effects of thyroxine for various devel-opmental processes in further detail (Uziel et al. 1985a,b).Progressive irreversible morphogenetic or neuronal damage tothe cochlea was found in hypothyroid rats only after the firsttwo postnatal months (Uziel et al. 1985a). Because the devel-opmental stage of 1-yr-old infants corresponds approximatelyto that of 1- to 2-mo-old rats (Eggermont et al. 1991; Teas etal. 1982), TH-replacement therapy in humans, if begun before1 yr of age in infants, has been assumed to successfullycompensate hearing loss as a result of TH deficiency.
Specificity of MMI effects
Using methimazole (MMI) instead of propylthiouracil(PTU) (Uziel et al. 1983a,b) as goitrogen, we recently con-firmed the irreversible neuronal and morphogenetic damage ofthe cochlea when plasma-TH levels are suppressed from,E17until .P28 (Knipper et al. 1999). Here we document theirreversible elevation of hearing thresholds when shorter TH-deficient periods (using MMI) are induced between the onset offetal thyroid gland activity atE17–E18and the onset of hearingat P12. MMI and PTU are common goitrogens that inhibitthyroid peroxidase-catalyzed thyroid hormone formation. Bothdrugs have been used safely for decades to suppress plas-ma-TH levels in animals and humans (Becks and Burrow 1991;Lind 1997). Neither the administration of MMI beforeE17(present study) nor the administration of MMI afterP1 or P2(Knipper et al. 1999) had any effect on the development ofnormal hearing. Thus MMI induces hearing loss only when theMMI-induced T3-deficient period spans the critical develop-mental period before the onset of hearing. Within this criticalperiod we can also observe a deterioration of hearing, whichappears to reflect the subsequent failure of TH-dependent stepsalong the peripheral-centripetal axis of the auditory system.We presume from these observations that the effects on innerear development by MMI are mediated by a specific effect onT3 synthesis, rather than due to a cochlear sensitivity to apresumptive MMI toxicity especially during this limited criti-cal period.
Role of maternal TH for hearing function
Plasma-TH level in dams declines within 2–3 days afterMMI treatment, similar to that described for the pups (Knipper
et al. 1999). Because mating was limited to 3 days, the admin-istration of MMI to dams from the third day after mating ispresumed to lead to depletion of the plasma-TH level in damssometimes betweenE3 and E6. The absence of TH in damsbetweenE3 and E17 (Table 2) would therefore lead to aTH-free period at least betweenE6 and E17 (Table 2). Inanother group, administration of MMI to pregnant dams fromE3 to P12, E8to P12,andE17to P12(Table 2) guarantees thatTH is suppressed during the next developmental period, whenthe pups are able to produce TH via their own thyroid glandactivity from E17 onward. The analysis of both groups thuscovers the developmental period whena- and b-thyroid hor-mone receptor expression was observed in the neonatal otocystfrom aboutE14onward (Bradley et al. 1994), the time mater-nal TH is presumed to act. The decline of maternal TH betweenE3 and E17 is of no consequence for the development ofhearing, because adult rats have similar thresholds than controlrats (Table 2) and because hearing thresholds in offspring ofMMI-treated dams fromE3 to P12, E8to P12andE17 to P12are similar (Table 2). This implies that the neonatally ex-pressed TR receptors in the otocyst (Bradley et al. 1994) act inhormone-independent pathways, as described for other tissues(Helmer et al. 1996). This result is in good agreement withrecent findings describing the lack of effects of thyroid hor-mone on late fetal rat brain development (Schwartz et al.1997). Neither gene expression nor cerebellar morphogenesiswere altered in the absence of fetal TH (Schwartz et al. 1997).
Novel TH-dependent processes during the criticaldevelopmental time period before hearing function
Our data imply that the observed hearing impairment isobviously not due to a failure of the classical TH-dependentprocesses, the formation of the inner sulcus and maturation ofefferent and afferent innervation (Fig. 3), both of which ariseafter prolonged TH-deficient periods (Deol 1973; Uziel 1985).This suggests the existence of additional TH-dependent pro-cesses during auditory system development and indicates thatthe auditory system reacts different to a corrective TH supply.Although morphogenesis and innervation of the cochlea ap-parently do respond to corrective TH supply, other TH-depen-dent processes require T3 activity during a very tight timeperiod. The increase in hearing loss after prolongation of aTH-deficient period is reflected first in affected thresholds(Figs. 2 and 7A), then in affected thresholds and impairedactive cochlear function (Figs. 2 and 4), and finally in affectedthresholds, impaired active cochlear function and elevatedbrain stem latencies (Figs. 2 and 5, Table 3). This implies thatTH deficiency during this critical period of development af-fects individual sections along the centripetal axis of the audi-tory system step by step. This concept is supported by theobserved tendency of higher order brain stem nuclei to be lessaffected when T3-deficient periods are prolonged (Table 3C).These differences are probably connected to the time at whichTH usually acts in these areas during development. Thereforethe various grades of hearing impairment appear to reflectdifferent targets of TH in the auditory system, starting from theperiphery in a centripetal direction. Elevated hearing thresh-olds in animals that have undergone TH deficiency from theonset of thyroid function (E17) onward up toP3 andP5 (Fig.2, Table 3A) may reflect the earliest TH activities within the
3109THYROID HORMONE AND INNER EAR

organ of Corti. In this context, it will be interesting to definethe effects of TH on potassium channels in hair cells, whoseexpression was recently shown to be accelerated by TH(Ruesch et al. 1999). However, the possibility of TH-depen-dent changes affecting cochlea ultrastructure of the cytoskele-ton (Poddar et al. 1996) or the expression or posttranslationalmodification of functional important proteins (Tosic et al.1992) should also be considered.
In all animals that underwent a delay in the rise of TH untilP8 or thereafter, we also observed a loss of DPOAEs (Fig. 4B,Table 3B). DPOAEs are sound emissions from the ear, whichare produced by pairs of tones that interact nonlinearly toproduce extra tones (Patuzzi 1996). These tones are nowthought to be produced by an overlap of the actively amplifiedtraveling waves on the basilar membrane (Probst et al. 1991;Zureck 1985). The active mechanisms in the cochlea are drivenby hair cell electromotility, which is in turn dependent on afunctional mechanoelectrical transduction of acoustic stimuli,as well as on intact endocochlear and membrane potentials (fora review, see Dallos and Evans 1995). The observed impair-ment of DPOAEs may thus be due either to impaired mecha-noelectrical or electromechanical transduction or impaired en-docochlear potential (EP), one of the driving forces forelectromotility (for a review see, Patuzzi 1996). Althoughthyroid hormone receptors have been found to be expressed inOHCs (Knipper et al. 1998; Lautermann and Ten Cate 1997),so far neither of the processes that are the driving forces forelectromotility have been shown to be controlled by TH. Fu-ture studies should therefore be aimed at discovering TH-dependent steps that are responsible for the loss of DPOAEs inanimals that sustained TH-deficient periods before hearing.
A decrease in speed of auditory brain stem responses may becaused by hypomyelination of the auditory nerve. There areseveral reports that underline the possibility that parts of themyelination process, including myelin gene expression, may beunder the control of TH (for a review see Oppenheimer et al.1994). We recently observed that the expression of myelingenes in the peripheral and central part of the intradural seg-ment of the VIIIth nerve began simultaneously and beforecochlea function; expression rapidly achieved its maximal lev-els at aroundP12, the day on which cochlea function com-mences (Knipper et al. 1998). It was shown that TH is respon-sible for the immediate saturation of myelin gene expression(Knipper et al. 1998). The part played by TH-dependent pro-cesses in improvement hearing sensitivity may thus include thetimely expression of myelin gene expression before the onsetof hearing atP12 (Knipper et al. 1998) and thereby may be aprerequisite for the nerve conduction and nerve impulse trans-mission along the auditory tract (Knipper et al. 1998), thedriving forces for the subsequent maturation steps of centralauditory projections (Eggermont et al. 1991; Moore et al.1995). Maturation processes in the central auditory systeminclude the increase in myelin sheaths and the maturation ofcentral synapses and dendrites (Eggermont et al. 1991; Mooreet al. 1995; Shah et al. 1978). Indeed, similar to the visualsystem, various studies support the notion that the timely onsetof auditory experience is essential for the development of intactmaturation of central auditory processes (Ptok and Ptok 1996;Walger et al. 1993). In the present study elevated latencieswere observed in animals that had sustained TH-deficient pe-riods until P10, but not in those with shorter TH-deficient
periods (Table 3). Elevated latencies were observed, althoughno reduction of myelin gene expression along the auditory tractin these animals was detected (Fig. 5). Considering that thedelayed activity of TH in these animals definitely leads toretarded myelin gene expression, as shown previously (Knip-per et al. 1998) and under the assumption that the saturation ofthe myelin gene expression may indeed be a prerequisite forauditory experience to promote central maturation steps, theobserved enhanced ABR latencies could be an indirect conse-quence of impaired cochlear function and retarded onset ofauditory experience during development. In the light of theseresults, the possible connection between the observed impair-ment of the cytoarchitecture of the neocortex and cerebellumand thyroid hormone deficiency should be reconsidered (Ber-bel et al. 1993; Eayrs 1971; Graves and Hawkes 1990; Legrand1967).
Comparison of the critical period of the inner ear in ratsand humans
After MMI treatment was stopped, T3 levels returned tonormal within 2–4 days (Fig. 1). After the delay in the rise ofplasma-T3 levels from the onset of fetal thyroid gland function(E17) up to P3, P5,and P8, T3-levels are therefore back tonormal beforeP12,which marks the end of the TH-dependentperiod (Knipper et al. 1999) and the onset of hearing function(Rubel 1978; Shah et al. 1978). Based on the degree of co-chlear maturation in rats (Lavigne-Rebillard and Pujol 1987)and humans (Johansson et al. 1964; Moore et al. 1995), as wellas the precise time at which brain stem responses and activecochlear functions can be registered in rats (Geal-Dor et al.1993; He et al. 1994; Jewett and Romano 1972) and humans(Birnholz and Benacerral 1983; Bonfils et al. 1988; Ponton etal. 1996; Starr et al. 1977),P12 in rats corresponds to thedevelopmental stage of the auditory system at approximatelythe 29th embryonal week in humans. Thus the observed hear-ing defects in transiently TH-deprived animals, which havebeen shown in the present study to return to normal plasma-T3levels beforeP12, confirm that the developmental stage atwhich a TH-replacement therapy in humans should be mostsuccessful in avoiding hearing impairment will be long beforebirth, even before the 29th embryonal week (see Fig. 7,B andC). The benefits of TH-replacement therapy for newborns toimprove hearing ability and its exact timing should therefore beurgently reconsidered. Current recommendations for sufficientTH and iodine levels during pregnancy may have to be alteredbecause even short, transient periods of TH and iodine defi-ciency during the critical period between the onset of fetalthyroid gland function (approximately the 10th embryonalweek) and the onset of the first auditory brain stem responseswithin the 29th embryonal week (Moore et al. 1995), may besuggested to be the cause of various degrees of congenitalhearing deficiencies.
Some genetic diseases related to thyroid dysfunction areaccompanied by hearing loss but, similar to findings demon-strated here, showed normal cochlear morphogenesis and in-nervation (Biebermann et al. 1997; Green et al. 1988;O’Malley et al. 1995; Utiger 1995). The various degrees ofhearing impairment observed in multiple forms of genetic andacquired diseases related to thyroid dysfunction (Bernal andNunez 1995; Biebermann et al. 1997; Meyerhoff 1980; Op-
3110 KNIPPER ET AL.

penheimer et al. 1994; Refetoff et al. 1967; Trotter 1960)should therefore be reconsidered in the context of the resultspresented here.
We thank C. Uysal for skillful photographic assistance and Dr. Besenthal forhelping us determine T3/T4 levels in the central clinical laboratories. We aregrateful to Drs. N. Schaeren-Wiemers and Marcus Frank from the BrainResearch Institute, Zurich, Switzerland, for the generous gift of PLP and MBPcDNA.
This work was supported by the Deutsche Forschungsgemeinschaft Ze149/6-1, Kni 316/2-1, and SFB 430, Kni-B3.
Address for reprint requests: M. Knipper, Centre for Hearing Research,Molecular Neurobiology, University of Tu¨bingen, Ro¨ntgenweg 11, D-72076Tubingen, Germany.
Received 3 August 1999; accepted in final form 7 January 2000.
REFERENCES
BECKS, G. P.AND BURROW, G. N. Thyroid disease and pregnancy.Med. Clin.North Am.75: 121–150, 1991.
BERBEL, P., GUAADANO-FERRAZ, A., MARTINEZ, M., QUILES, J. A., BALBOA, R.,AND INNOCENTI, G. M. Organization of auditory callosal connections inhypothyroid adult rats.Eur. J. Neurosci.5: 1465–1478, 1993.
BERGLUND, A. M. AND RYUGO, D. K. A monoclonal antibody labels TIIneurons of the spiral ganglion.Brain Res.383: 327–332, 1986.
BERNAL, J. AND NUNEZ, J. Thyroid hormones and brain development.Eur. J.Endocrinol.133: 390–398, 1995.
BIEBERMANN, H., GRUTERS, M., SCHONEBERG, T., AND GUDERMANN, T. Con-genital hypothyroidism caused by mutations in the thyrotropin-receptorgene.N. Engl. J. Med.336: 1390–1391, 1997.
BIRNHOLZ, J. C. AND BENACERRAL, B. R. The development of fetal hearing.Science222: 516–518, 1983.
BONFILS, P., UZIEL, A., AND PUJOL, R. Evoked otoacoustic emissions fromadults and infants: clinical applications.Acta Otolaryngol.105: 445–449,1988.
BRADLEY, D. J., TOWLE, H. C., AND YOUNG, W. S. Alpha and beta thyroidhormone receptor (TR) gene expression during auditory neurogenesis: evi-dence for TR isoform-specific transcriptional regulation in vivo.Proc. Natl.Acad. Sci. USA91: 439–443, 1994.
CALVO, R., OBREGON, M. J., ESCOBAR-DEL-REY, F., AND MORREALE-DE-ESCO-BAR, G. The rat placenta and the transfer of thyroid hormones from themother to the fetus.Endocrinology131: 357–365, 1992.
CALVO, R., OBREGON, M. J., RUIZ DE ONA, C., ESCOBAR DEL REY, F., AND
MORREALE DE ESCOBAR, G. Congenital hypothyroidism, as studied in rats.Crucial role of maternal thyroxine, but not of 3,5,39-triiodothyronine in theprotection of the fetal brain.J. Clin. Invest. 86: 889–899, 1990.
DALLOS, P. AND EVANS, B. N. High-frequency motility of outer hair cells andthe cochlear amplifier.Science267: 2006–2009, 1995.
DEOL, M. S. An experimental approach to the understanding and treatment ofhereditary syndromes with congenital deafness and hypothyroidism.J. Med.Gen.1: 235–242, 1973.
EAYRS, J. T. Thyroid and developing brain: anatomical and behavioural effects.In: Hormones and Development,edited by M. Hamburgh and E.J.W. Bar-rington. New York: Appleton Centruy Crofts, 1971, p. 345–355.
EGGERMONT, J. J., PONTON, C. W., COUPLAND, S. G., AND WINKELAAR , R.Maturation of the traveling-wave delay in the human cochlea.J. Acoust. Soc.Am.90: 288–298, 1991.
FREERKSEN, D. L., SCHROEDL, N. A., AND HARTZELL, C. R. Triiodothyroninedepresses the NAD-linked glycerol-3-phosphate dehydrogenase activity ofcultured neonatal rat heart cells. Arch. Biochem. Biophys.228: 474–479,1994.
GEAL-DOR, M., FREEMAN, S., LI, G., AND SOHMER, H. Development of hearingin neonatal rats: air and bone conducted ABR thresholds.Hear. Res.69:236–242, 1993.
GESTWA, G., WIECHERS, B., ZIMMERMANN , U., PRAETORIUS, M., ROHBOCK, K.,KOPSCHALL, I., ZENNER, H. P.,AND KNIPPER, M. Differential expression oftrkB.T1 and trkB.T2, truncated trkC and p75NGFR in the cochlea prior tohearing function.J. Comp. Neurol.414: 33–49, 1999.
GOLDEY, E. S.AND CROFTON, K. M. Thyroxine replacement attenuates hypo-thyroxinemia, hearing loss, and motor deficits following developmentalexposure to Aroclor 1254 in rats.Toxicol. Sci.45: 94–105, 1998.
GRAVES, C. AND HAWKES, R. Maturation of the corpus callosum of the rat. I.Influence of thyroid hormones on the topography of callosal projections.J. Comp. Neurol.291: 128–146, 1990.
GREEN, R. P., BIRKENMEIER, E. H., BEAMER, W. G., MALTAIS, L. J., AND
GORDON, J. I. The hypothyroid (hyt/hyt) mouse: a model system for studyingthe effects of thyroid hormone on developmental changes on gene expres-sion.Proc. Natl. Acad. Sci.85: 5592–5596, 1988.
HE, D. Z., EVANS, B. N., AND DALLOS, P. First appearance and development ofelectromotility in neonatal gerbil outer hair cells.Hear. Res.78: 77–90, 1994.
HELMER, E. B., RAAKA , B. M., AND SAMUELS, H. Hormone-dependent and-independent transcriptional activation by thyroid hormone receptors aremediated by different mechanism.Endocrinology137: 390–399, 1996.
JEWETT, D. L. AND ROMANO, M. N. Neonatal development of auditory systempotentials averaged from the scalp of rat and cat.Brain Res.36: 101–115,1972.
JOHANSSON, B., WEDENBERG, E., AND WESTIN, B. Measurement of tone re-sponse by the human fetus.Acta Otolaryngol. Stockh.57: 188–192, 1964.
KEMP, W. R. Deafness in myxodema.Br. Med. J.1: 355, 1907.KHVOLES, R., FREEMAN, S.,AND SOHMER, H. Development of transient evoked
otoacoustic emissions in the neonatal rat.Auditol. Neurootol.3: 40–53,1998.
KNIPPER, M., BANDTLOW, C., GESTWA, L., KOPSCHALL, I., ROHBOCK, K.,WIECHERS, B., ZENNER, H. P., AND ZIMMERMANN , U. Thyroid hormoneaffects Schwann cell and oligodendrocyte gene expression at the glialtransition zone of the VIIIth nerve prior to cochlea function.Development125: 3709–3718, 1998.
KNIPPER, M., KOPSCHALL, I., ROHBOCK, K., KOPKE, A.K.E., BONK, I., ZIMMER-MANN, U., AND ZENNER, H. P. Transient expression of NMDA receptorsduring rearrangement of AMPA receptor expressing fibers in the developinginner ear.Cell Tissue Res.1: 23–42, 1997.
KNIPPER, M., ROHBOCK, K., KOPSCHALL, I., GESTWA, L., WIECHERS, B., BRUG-GER, H., MAIER, H., TEN CATE, W.-J., LAUTERMANN, J., ZENNER, H. P.,AND
ZIMMERMANN , U. Distinct thyroid hormone-dependent expression of trkBand p75NGFR in nonneuronal cells during the critical TH-dependent periodof the cochlea.J. Neurobiol.38: 338–356, 1999.
KNIPPER, M., ZIMMERMANN , U., ROHBOCK, K., KOPSCHALL, I., AND ZENNER,H. P. Synaptophysin and GAP-43 proteins in efferent fibers of the inner earduring postnatal development.Dev. Brain Res.89: 73–86, 1995.
KOLLIKER, A. Kollikers Handbuch der Gewebelehre des Menschen. Bau derSchnecke.Leipzig: 1863, vol. 4.
LAUTERMANN, J. AND TEN CATE, W.J.F. Postnatal expression of thea-thyroidhormone receptor in the rat cochlea.Hear. Res.107: 23–28, 1997.
LAVIGNE-REBILLARD, M. AND PUJOL, R. Surface aspects of the developinghuman organ of Corti.Acta Otolaryngol. Stockh. Suppl.436: 43–50, 1987.
LEGRAND, J. Variations en fonction de l’age de la response du cervelet al’action morphogenetique de la thyroide chez le rat.Arch. Anat. Microsc.Morphol. Exp.56: 291–307, 1967.
LEMKE, G.AND AXEL, R. Isolation and sequence of a cDNA encoding the majorstructural protein of peripheral myelin.Cell 40: 501–508, 1985.
LIND, P. Therapy of hypo- and hyperthyroidism in pregnancy.Acta Med.Austriaca24: 157–158, 1997.
MARCHANT, B., BROWNLIE, B. E., HART, D. M., HORTON, P. W., AND ALEX-ANDER, W. D. The placental transfer of propylthiouracil, methimazole andcarbimazole.J. Clin. Endocrinol. Metab.45: 1187–1193, 1977.
MEYERHOFF, W. L. Hypothyroidism and the ear: electrophysiological, mor-phological and chemical considerations.Laryngoscope89: Suppl.19, 1980.
MILNER, R. J., LAI, C., NAVE, K. A., LENOIR, D., OGATA, J., AND SUTCLIFFE,J. G. Nucleotide sequence of two mRNAs for rat brain myelin proteolipidprotein.Cell 42: 931–939, 1985.
MOORE, D. R. Postnatal development of the mammalian central auditoy systemand the neural consequences of auditory deprivation.Acta Otolaryngol.Suppl.421: 19–30, 1985.
MOORE, J. K., PERAZZO, L. M., AND BRAUN, A. Time course of axonalmyelination in the human brainstem auditory pathway.Hear. Res.87:21–31, 1995.
MORTIMER, R. H., CANNELL, G. R., ADDISON, R. S., JOHNSON, L. P., ROBERTS,M. S., AND BERNUS, I. Methimazole and propylthiouracil equally cross theperfused human term placental lobule.J. Clin. Endocrinol. Metab.82:3099–3102, 1997.
MURRAY, G. R. Note on the treatment of myxoedema by hypodermic injectionsof an extract of the thyroid gland of a sheep.Br. Med. J.2: 796–797, 1891.
O’MALLEY , B. W., LI, D., AND TURNER, D. S. Hearing loss and cochlearabnormalities in the congenital hypthyroid (hyt/hyt) mouse.Hear. Res.88:181–189, 1995.
3111THYROID HORMONE AND INNER EAR

OPPENHEIMER, J. H., SCHWARTZ, H. L., AND STRAIT, K. A. Thyroid hormoneaction 1994: the plot thickens.Eur. J. Endocrinol.130: 15–24, 1994.
OSLER, W. The Principles and Practice of Medicine(3rd ed.). New York:Appleton, 1889, p. 840–843.
PARVING, A. Hearing problems and hormonal disturbances in the elderly.ActaOtolaryngol. Suppl. Stockh.476: 44–53, 1990.
PATUZZI, R. Cochlear micromechanics and macromechanics. In:The Cochlea,edited by P. Dallos, P. A. Popper, and R. R. Fay. New York: SpringerVerlag, 1996, p. 186–258.
PODDAR, R., PAUL, S., CHAUDHURY, S.,AND SARKAR, P. K. Regulation of actinand tubulin gene expression by thyroid hormone during rat brain develop-ment.Mol. Brain Res.35: 111–118, 1996.
PONTON, C. W., MOORE, J. K., AND EGGERMONT, J. J. Auditory brain stemresponse generation by parallel pathways: differential maturation of axonalconduction time and synaptic transmission.Ear Hear.17: 402–410, 1996.
PROBST, R., LONSBURY-MARTIN, B. L., AND MARTIN, G. K. A review ofotoacoustic emissions. J. Acoust. Soc. Am.89: 2027–2067, 1991.
PTOK, J. AND PTOK, A. Die Entwicklung des Horens.Sprache Stimme Gehor.1: 6–10, 1996.
REFETOFF, S., DE WIND, L. T., AND DE GROTT, L. J. Familiar syndromecombining deaf-mutism, stippled epiphyses, goiter and abnormally highPBT: possible target organ refractoriness to thyroid hormones.J. Clin.Endocrinol. Metab.27: 279–294, 1967.
RITTER, F. N. The effects of hypothyroidism upon the ear, nose and throat. Aclinical and experimental study.Laryngoscope77: 1427–1479, 1967.
ROACH, A., BOYLAN, K., HORVATH, S., PRUSINER, S. B.,AND HOOD, L. E. Char-acterization of cloned cDNA representing rat myelin basic protein: absence ofexpression in brain of shiver mutant mice.Cell 34: 799–806, 1983.
RUBEL, E. W. Ontogeny of structure and function in the vertebrate’s auditorysystem. In:Handbook of Sensory Physiology. Development of SensorySystems,edited by M. Jacobsen. New York: Springer Verlag, 1978, vol. IX,p. 135–247.
RUEGER, B., THALHAMMER , J., OBERMAIER, I., AND GRUENEWALD-JANHO, S.Experimental procedure for the detection of rare human mRNA with theDIG system.Boehringer Mannheim Information99: 30–33, 1996.
RUESCH, A., ERWRY, L. C., OLIVER, D., VENSTROM, B., AND FORREST, D.Thyroid hormone receptorb-dependent expression of potassium conduc-tance in inner hair cells at the onset of hearing. Proc. Natl. Acad. USA95:15758–15762, 1999.
SACK, J., WELLER, A., RIGLER, O.,AND ROZIN, A. A simple model for studyingthe correction of in utero hypothyroidsm in the rat.Pediatr. Res.37:497–501, 1995.
SCHAEREN-WIEMERS, N. AND GERFIN-MOSER, A. Single protocol to detecttranscripts of various types and expression levels in neural tissue and
cultured cells; in situ hybridization using digoxigenin-labeled cRNA probes.Histochemistry100: 431–440, 1993.
SCHWARTZ, H. L., ROSS, M. E., AND OPPENHEIMER, J. H. Lack of effect ofthyroid hormone on late fetal rat brain development.Endocrinology138:3119–3124, 1997.
SHAH, S. N., BHARGAVA, V. K., AND MCKEAN, C. M. Maturation changes inearly auditory evoked potentials and myelination of the inferior colliculus inrats.Neuroscience3: 561–653, 1978.
STARR, A., AMILI , R. N., MARTIN, W. H., AND SANDERS, S. Development ofauditory function in newborn infants revealed by auditory brainstem poten-tials. Pediatrics60: 831–839, 1977.
TEAS, D. C., KLEIN, A. J.,AND KRAMER, S. J. An analysis of auditory brainstemresponses in infants.Hear. Res.7: 19–54, 1982.
TOSIC, M., TORCH, S., COMTE, V., DOLIVO, M., HONEGGER, P.,AND MATTHIEU,J. M. Triiodothyronine has diverse and multiple stimulating effects onexpression of the major myelin protein genes.J. Neurochem.59: 1770–1777, 1992.
TROTTER, N. R. The association of deafness with thyroid dysfunction.Br. Med.Bull. 16: 92–98, 1960.
UTIGER, R. D. Thyrotrophin-receptor mutations and thyroid dysfunction.New.Engl. 332: 183–195, 1995.
UZIEL, A. Non-genetic factors affecting dearing development.Acta Otolaryn-gol. 421: 57–61, 1985.
UZIEL, A., LEGRAND, C., OHRESSER, M., AND MAROT, M. Maturational anddegenerative processes in the organ of Corti after neonatal hypothyroidism.Hear. Res.11: 203–218, 1983a.
UZIEL, A., LEGRAND, C., AND RABIE, A. Corrective effects of thyroxine oncochlear abnormalities induced by congenital hypothyroidism in the rat. I.Morphogical study.Dev. Brain Res.19: 111–122, 1985a.
UZIEL, A., MAROT, M., AND RABIE, A. Corrective effects of thyroxine oncochlear abnormalities induced by congenital hypothyroidism in the rat. II.Electrophysiological study.Dev. Brain Res.19: 123–127, 1985b.
UZIEL, A., PUJOL, R., LEGRAND, C., AND LEGRAND, J. Cochlear synaptogenesisin the hypothyroid rat.Brain Dev.283: 295–301, 1983b.
WALGER, M., LASKA, M., SCHNEIDER, I., DIEKMANN , H., AND VON WEDEL,H. Maturation of auditory evoked potentials in young guinea pigs withbinaural conductive hearing loss.Eur. Arch. Otorhinolaryngol.250: 362–365, 1993.
WIECHERS, B., GESTWA, G., MACK, A., ZENNER, H. P., AND KNIPPER, M.Changing patterns of BDNF expression correlate with rearrangement offibres during cochlear development of rats and mice.J. Neurosci.19:3033–3042, 1999.
ZUREK, P. M. Acoustic emissions from the ear: a summary of results fromhumans and animals. J. Acoust. Soc. Am.78: 340–344, 1985.
3112 KNIPPER ET AL.
Copyright © 2022 FDOKUMEN




















