
Detailed alignment of Saccharum and Sorghum chromosomes: comparative organization of closely related...
20
Copyright 1998 by the Genetics Society of America Detailed Alignment of Saccharum and Sorghum Chromosomes: Comparative Organization of Closely Related Diploid and Polyploid Genomes R. Ming,* S.-C. Liu,* Y.-R. Lin,* J. da Silva, ²,‡ W. Wilson, ² D. Braga, ²,‡ A. van Deynze, ² T. F. Wenslaff, § K. K. Wu, § P. H. Moore,** W. Burnquist, ‡ M. E. Sorrells, ² J. E. Irvine ²² and A. H. Paterson* ,1 *Plant Genome Mapping Laboratory, Department of Soil and Crop Science, Texas A&M University, College Station, Texas 77843, ² Department of Plant Breeding and Biometry, Cornell University, Ithaca, New York 14853, ‡ Copersucar Technology Center, Caixa Postal 162, 13.400 Piracicaba, Sao Paulo, Brazil, § Hawaii Agriculture Research Center, formerly Hawaiian Sugar Planters’ Association, Aiea, Hawaii 96701, **USDA-ARS, Hawaii Agriculture Research Center, formerly Hawaiian Sugar Planters’ Association, Aiea, Hawaii 96701 and ²² Texas A&M Agricultural Research and Extension Center, Weslaco, Texas 78596 Manuscript received February 5, 1998 Accepted for publication September 10, 1998 ABSTRACT The complex polyploid genomes of three Saccharum species have been aligned with the compact diploid genome of Sorghum (2n 5 2x 5 20). A set of 428 DNA probes from different Poaceae (grasses) detected 2460 loci in F 1 progeny of the crosses Saccharum officinarum Green German 3 S. spontaneum IND 81-146, and S. spontaneum PIN 84-1 3 S. officinarum Muntok Java. Thirty-one DNA probes detected 226 loci in S. officinarum LA Purple 3 S. robustum Molokai 5829. Genetic maps of the six Saccharum genotypes, including up to 72 linkage groups, were assembled into “homologous groups” based on parallel arrange- ments of duplicated loci. About 84% of the loci mapped by 242 common probes were homologous between Saccharum and Sorghum. Only one interchromosomal and two intrachromosomal rearrangements differ- entiated both S. officinarum and S. spontaneum from Sorghum, but 11 additional cases of chromosome structural polymorphism were found within Saccharum. Diploidization was advanced in S. robustum, incipi- ent in S. officinarum, and absent in S. spontaneum, consistent with biogeographic data suggesting that S. robustum is the ancestor of S. officinarum, but raising new questions about the antiquity of S. spontaneum. The densely mapped Sorghum genome will be a valuable tool in ongoing molecular analysis of the complex Saccharum genome. T HE consequences of polyploid formation in plants S. officinarum (Sreenivasan et al. 1987; Roach 1995; are exemplified by comparison of the closely re- J. E. Irvine, unpublished data). Molecular analyses lated genera Saccharum (sugarcane) and Sorghum. In of chloroplasts (Sobral et al. 1994), mitochondria as little as 5 million years since Saccharum and Sorghum (D’Hont et al. 1993), and ribosomal DNA (Glaszmann diverged from a common ancestor (Al-Janabi et al. et al. 1990) support the concept of S. spontaneum as 1994b), Saccharum species have reached gametic chro- a separate species. The aborted inflorescence species mosome numbers ranging from 18 to 85 or more. Two S. edule (2n 5 60, 70, 80, and some aneuploids) may be wild Saccharum species are recognized: Saccharum spon- a hybrid of S. officinarum or S. robustum with Miscanthus taneum (2n 5 36–128), with a putative center of origin sp. (Daniels and Roach 1987). in India, and S. robustum (2n 5 60–170), putatively from Modern sugarcane varieties derived from hybrids be- New Guinea. The cultivated species S. officinarum (2n 5 tween S. officinarum and S. spontaneum are among the 70–140) is thought to be derived from S. robustum, and world’s most valuable crops, at about $143 billion an- also has a center of origin and diversity in New Guinea. nually (Christou et al. 1992). High sugar content is While sugarcane breeders regard S. officinarum as a sepa- thought to be contributed by S. officinarum while vegeta- rate species, plant taxonomists generally view it as a tive vigor and resistance to biotic/abiotic stresses are cultivated derivative of wild S. robustum. Cultivated taxa attributed to S. spontaneum. referred to as S. barberi in India and S. sinense in China A high level of chromosome duplication and autoga- are believed to be natural hybrids of S. spontaneum and mous chromosome pairing in sugarcane preclude ge- netic mapping based on codominant “alleles.” DNA markers showing simplex or “single-dose” segregation Corresponding author: A. H. Paterson, Department of Soil and Crop (Wu et al. 1992) have been used to construct genetic Science, Texas A&M University, College Station, TX 77843-2474. linkage maps of S. spontaneum SES208 (2n 5 64) and E-mail: [email protected] its doubled haploid ADP068 (Al-Janabi et al. 1993; da 1 Present address: Department of Crop and Soil Science, University of Georgia, Athens, GA 30602. Silva et al. 1993). This map consisted of 64 linkage Genetics 150: 1663–1682 (December 1998)
-
Upload
independent -
Category
Documents
-
view
7 -
download
0
Transcript of Detailed alignment of Saccharum and Sorghum chromosomes: comparative organization of closely related...

Copyright 1998 by the Genetics Society of America
Detailed Alignment of Saccharum and Sorghum Chromosomes: ComparativeOrganization of Closely Related Diploid and Polyploid Genomes
R. Ming,* S.-C. Liu,* Y.-R. Lin,* J. da Silva,†,‡ W. Wilson,† D. Braga,†,‡ A. van Deynze,†
T. F. Wenslaff,§ K. K. Wu,§ P. H. Moore,** W. Burnquist,‡ M. E. Sorrells,†
J. E. Irvine†† and A. H. Paterson*,1
*Plant Genome Mapping Laboratory, Department of Soil and Crop Science, Texas A&M University, College Station, Texas 77843, †Departmentof Plant Breeding and Biometry, Cornell University, Ithaca, New York 14853, ‡Copersucar Technology Center, Caixa Postal 162, 13.400
Piracicaba, Sao Paulo, Brazil, §Hawaii Agriculture Research Center, formerly Hawaiian Sugar Planters’ Association, Aiea, Hawaii96701, **USDA-ARS, Hawaii Agriculture Research Center, formerly Hawaiian Sugar Planters’ Association, Aiea, Hawaii 96701
and ††Texas A&M Agricultural Research and Extension Center, Weslaco, Texas 78596
Manuscript received February 5, 1998Accepted for publication September 10, 1998
ABSTRACTThe complex polyploid genomes of three Saccharum species have been aligned with the compact diploid
genome of Sorghum (2n 5 2x 5 20). A set of 428 DNA probes from different Poaceae (grasses) detected2460 loci in F1 progeny of the crosses Saccharum officinarum Green German 3 S. spontaneum IND 81-146,and S. spontaneum PIN 84-1 3 S. officinarum Muntok Java. Thirty-one DNA probes detected 226 loci inS. officinarum LA Purple 3 S. robustum Molokai 5829. Genetic maps of the six Saccharum genotypes,including up to 72 linkage groups, were assembled into “homologous groups” based on parallel arrange-ments of duplicated loci. About 84% of the loci mapped by 242 common probes were homologous betweenSaccharum and Sorghum. Only one interchromosomal and two intrachromosomal rearrangements differ-entiated both S. officinarum and S. spontaneum from Sorghum, but 11 additional cases of chromosomestructural polymorphism were found within Saccharum. Diploidization was advanced in S. robustum, incipi-ent in S. officinarum, and absent in S. spontaneum, consistent with biogeographic data suggesting that S.robustum is the ancestor of S. officinarum, but raising new questions about the antiquity of S. spontaneum.The densely mapped Sorghum genome will be a valuable tool in ongoing molecular analysis of the complexSaccharum genome.
THE consequences of polyploid formation in plants S. officinarum (Sreenivasan et al. 1987; Roach 1995;are exemplified by comparison of the closely re- J. E. Irvine, unpublished data). Molecular analyses
lated genera Saccharum (sugarcane) and Sorghum. In of chloroplasts (Sobral et al. 1994), mitochondriaas little as 5 million years since Saccharum and Sorghum (D’Hont et al. 1993), and ribosomal DNA (Glaszmanndiverged from a common ancestor (Al-Janabi et al. et al. 1990) support the concept of S. spontaneum as1994b), Saccharum species have reached gametic chro- a separate species. The aborted inflorescence speciesmosome numbers ranging from 18 to 85 or more. Two S. edule (2n 5 60, 70, 80, and some aneuploids) may bewild Saccharum species are recognized: Saccharum spon- a hybrid of S. officinarum or S. robustum with Miscanthustaneum (2n 5 36–128), with a putative center of origin sp. (Daniels and Roach 1987).in India, and S. robustum (2n 5 60–170), putatively from Modern sugarcane varieties derived from hybrids be-New Guinea. The cultivated species S. officinarum (2n 5 tween S. officinarum and S. spontaneum are among the70–140) is thought to be derived from S. robustum, and world’s most valuable crops, at about $143 billion an-also has a center of origin and diversity in New Guinea. nually (Christou et al. 1992). High sugar content isWhile sugarcane breeders regard S. officinarum as a sepa- thought to be contributed by S. officinarum while vegeta-rate species, plant taxonomists generally view it as a tive vigor and resistance to biotic/abiotic stresses arecultivated derivative of wild S. robustum. Cultivated taxa attributed to S. spontaneum.referred to as S. barberi in India and S. sinense in China A high level of chromosome duplication and autoga-are believed to be natural hybrids of S. spontaneum and mous chromosome pairing in sugarcane preclude ge-
netic mapping based on codominant “alleles.” DNAmarkers showing simplex or “single-dose” segregation
Corresponding author: A. H. Paterson, Department of Soil and Crop (Wu et al. 1992) have been used to construct geneticScience, Texas A&M University, College Station, TX 77843-2474. linkage maps of S. spontaneum SES208 (2n 5 64) andE-mail: [email protected]
its doubled haploid ADP068 (Al-Janabi et al. 1993; da1 Present address: Department of Crop and Soil Science, Universityof Georgia, Athens, GA 30602. Silva et al. 1993). This map consisted of 64 linkage
Genetics 150: 1663–1682 (December 1998)

1664 R. Ming et al.
MATERIALS AND METHODSgroups and 8 homologous groups based on 276 restric-tion fragment length polymorphism (RFLP) and 208 Plant materials: Three interspecific F1 populations weresingle-dose (SD) arbitrarily primed polymerase chain used, each derived from crosses between heterozygous par-reaction (PCR) loci (da Silva et al. 1995). Mapping of ents: (1) 85 F1 plants from S. officinarum Green German (GG,
2n 5 97–117) 3 S. spontaneum IND 81-146 (IND, 2n 5 52–56);408 RFLP loci in a cultivated S. officinarum 3 S. sponta-(2) 85 F1 plants from S. officinarum Muntok Java (MJ, 2n 5neum hybrid R570 (2n 5 107–115) revealed 96 linkage140) 3 S. spontaneum PIN 84-1 (PIN, 2n 5 96); (3) 80 F1 plantsgroups and 10 putative homologous groups (Grivet etfrom S. officinarum LA Purple (LP, 2n 5 80) 3 S. robustum
al. 1996). Fifty-one linkage groups of S. officinarum LA Molokai 5829 (MOL, 2n 5 80). The SES 208 population wasPurple were mapped on the basis of 160 randomly am- previously described (da Silva et al. 1995). Populations Green
German 3 IND 81-146 (G 3 I) and PIN 84-1 3 Muntok Javaplified polymorphic DNA markers and 1 morphological(P 3 M) were genotyped at Texas A&M, LA Purple 3 Molokaimarker, in a cross with S. robustum (Mudge et al. 1996).5829 (L 3 MOL) was genotyped at the Hawaiian Sugar Plant-Molecular analysis of the complex autopolyploid ge-ers’ Association, and SES 208 was genotyped at Cornell. Newnome of Saccharum might be simplified by exploiting DNA probes were added to a previously described sorghum
its close relationship with the compact genome of Sor- population (Chittenden et al. 1994) at Texas A&M.ghum. The chromosome number of Saccharum ranges Genotyping: Genomic DNA extraction was slightly revised
from Li et al. (1995). About 7.5 mg sugarcane DNA per lanefrom 2n 5 36 to 170 (Daniels and Roach 1987; J. E.was digested with EcoRI, HindIII, or XbaI, according to theIrvine, unpublished data), while chromosome numbermanufacturer’s instructions. Southern blotting, radioactive la-of Sorghum ranges from 2n 5 10 to 40 (Doggett andbeling, and autoradiography were as described, except that 3
Prasada Rao 1995). A high-density sorghum map has ml 32P per reaction was used (Chittenden et al. 1994).been made in a cross between Sorghum bicolor and S. A total of 829 DNA probes were surveyed on G 3 I andpropinquum (Chittenden et al. 1994; A. H. Paterson P 3 M, including 132 cDNA and 18 genomic clones from
published sugarcane maps (da Silva et al. 1993, 1995): 201et al., unpublished data), both 2n 5 20. The range 2547–hypomethylated (PstI) Sorghum genomic clones (pSB, SHO);4183 Mbp/1C in DNA content of several Saccharum spp.199 random maize cDNA probes (CSU); 40 maize cDNAwith unspecified chromosome number (Arumugana-(CSU) and 45 genomic (BNL and UMC) clones previously
than and Earle 1991) is approximately consistent with mapped in sorghum; 89 rice cDNA (RZ) and 41 rice genomica three- to sixfold duplication of the genome of S. bicolor (RG) clones; 19 barley cDNAs (BCD) and 61 oat cDNAs
(CDO). Probe-enzyme combinations that generated the(2n 5 20; 760 Mbp/1C). This suggests that the differ-largest numbers of polymorphic bands were chosen for map-ence in DNA content of the Saccharum and Sorghumping. For L 3 MOL, 132 sugarcane cDNA probes, 25 sugarcanegenomes may largely be attributable to the larger num-genomic probes, and some additional probes from rice, maize,ber of chromosomes in the Saccharum nucleus, rather and oats were surveyed. Data for the SES 208 population are
than a higher percentage of repetitive DNA. as previously described (da Silva et al. 1995).Two prior studies have suggested parallels in gene Data analysis: Polymorphic restriction fragments from each
parent were scored present vs. absent in the mapping progeny.order between Saccharum and Sorghum, but had avail-Each marker was tested against the expected ratios for SD orable only sparsely distributed DNA markers. Dufour etsimplex (1:1), double-dose (DD) or duplex (11:3 for x 5 8,al. (1997) evaluated colinearity on the basis of 84 anchoror 7:2 for x 5 10), and “triple-dose” (TD) or triplex segregationprobes distributed at intervals of z20 cM across the (13:1 for x 5 8, or 11:1 for x 5 10; da Silva 1993; da Silva
Sorghum genome of .1500 cM (Chittenden et al. and Sorrells 1996). For determining significance thresholds,1994). Guimaraes et al. (1997) compared Saccharum we assumed that S. spontaneum is x 5 8 but S. officinarum and
S. robustum are x 5 10, on the basis of the periodicity observedand Sorghum on the basis of only 68 probes, and 63among known accessions ( J. E. Irvine, unpublished data). A(69%) of their Saccharum linkage groups containedchi-square threshold of 2% was used to keep both type I andtwo or fewer loci, making it impossible to test whethertype II errors below 5% (Wu et al. 1992). There is variationor not intrachromosomal rearrangements are present. in the chromosome counts available for GG (97–117) and
A long-term goal of our work is to study the basis, and IND (52–56). When analysis involved chromosome number,the consequences, of the rapid increase in chromosome the middle values of 107 for GG and 54 for IND were used.
SD markers were mapped using Mapmaker v2.0 for thenumber of Saccharum. The specific objectives of thisMacintosh (Lander et al. 1987). Linkage groups were built atstudy were as follows: (1) to construct detailed linkagethreshold LOD 5 5.0 and u 5 0.25. Genetic distances (inmaps for the three Saccharum species using a high-centimorgans) were calculated using the Kosambi mappingdensity sorghum linkage map as a template, (2) to assess function. DD and TD markers were superimposed on the SD
in detail the levels and patterns of chromosome struc- map by the method of da Silva (1993). Homologous groupstural rearrangement among Saccharum species and (HGs) were assembled on the basis of at least two probes
common to two linkage groups. The nomenclature of HGsbetween Saccharum and Sorghum, (3) to examine1–8 was consistent with the previously published SES 208 mapgenome coverage of the Saccharum maps through com-(da Silva et al. 1995). New HGs that shared no homologousparative genome analysis, (4) to compare the extent ofloci with SES 208 HGs 1–8 were denoted HGs 9–17 and arepreferential chromosome pairing in different Sac- consistent across the six maps from the three new crosses
charum species, and (5) to reexamine the elusive described herein. Inverted segregation data of each genotypequestion of the basal chromosome number(s) for Sac- were used to investigate repulsion phase linkages (Al-Janabi
et al 1994a; Grivet et al. 1996).charum.
1665Sugarcane-Sorghum Comparative Mapping
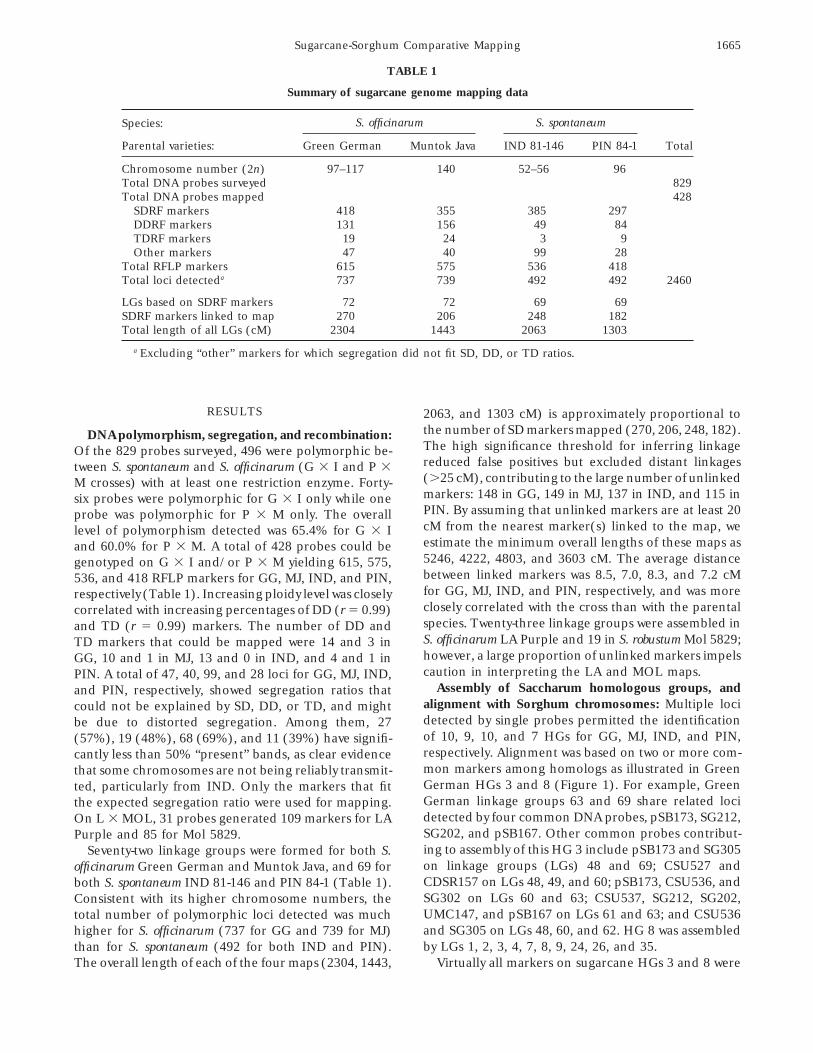
TABLE 1
Summary of sugarcane genome mapping data
S. officinarum S. spontaneumSpecies:
Parental varieties: Green German Muntok Java IND 81-146 PIN 84-1 Total
Chromosome number (2n) 97–117 140 52–56 96Total DNA probes surveyed 829Total DNA probes mapped 428
SDRF markers 418 355 385 297DDRF markers 131 156 49 84TDRF markers 19 24 3 9Other markers 47 40 99 28
Total RFLP markers 615 575 536 418Total loci detecteda 737 739 492 492 2460
LGs based on SDRF markers 72 72 69 69SDRF markers linked to map 270 206 248 182Total length of all LGs (cM) 2304 1443 2063 1303
a Excluding “other” markers for which segregation did not fit SD, DD, or TD ratios.
RESULTS 2063, and 1303 cM) is approximately proportional tothe number of SD markers mapped (270, 206, 248, 182).DNA polymorphism, segregation, and recombination:The high significance threshold for inferring linkageOf the 829 probes surveyed, 496 were polymorphic be-reduced false positives but excluded distant linkagestween S. spontaneum and S. officinarum (G 3 I and P 3(.25 cM), contributing to the large number of unlinkedM crosses) with at least one restriction enzyme. Forty-markers: 148 in GG, 149 in MJ, 137 in IND, and 115 insix probes were polymorphic for G 3 I only while onePIN. By assuming that unlinked markers are at least 20probe was polymorphic for P 3 M only. The overallcM from the nearest marker(s) linked to the map, welevel of polymorphism detected was 65.4% for G 3 Iestimate the minimum overall lengths of these maps asand 60.0% for P 3 M. A total of 428 probes could be5246, 4222, 4803, and 3603 cM. The average distancegenotyped on G 3 I and/or P 3 M yielding 615, 575,between linked markers was 8.5, 7.0, 8.3, and 7.2 cM536, and 418 RFLP markers for GG, MJ, IND, and PIN,for GG, MJ, IND, and PIN, respectively, and was morerespectively (Table 1). Increasing ploidy level was closelyclosely correlated with the cross than with the parentalcorrelated with increasing percentages of DD (r 5 0.99)species. Twenty-three linkage groups were assembled inand TD (r 5 0.99) markers. The number of DD andS. officinarum LA Purple and 19 in S. robustum Mol 5829;TD markers that could be mapped were 14 and 3 inhowever, a large proportion of unlinked markers impelsGG, 10 and 1 in MJ, 13 and 0 in IND, and 4 and 1 incaution in interpreting the LA and MOL maps.PIN. A total of 47, 40, 99, and 28 loci for GG, MJ, IND,
Assembly of Saccharum homologous groups, andand PIN, respectively, showed segregation ratios thatalignment with Sorghum chromosomes: Multiple locicould not be explained by SD, DD, or TD, and mightdetected by single probes permitted the identificationbe due to distorted segregation. Among them, 27of 10, 9, 10, and 7 HGs for GG, MJ, IND, and PIN,(57%), 19 (48%), 68 (69%), and 11 (39%) have signifi-respectively. Alignment was based on two or more com-cantly less than 50% “present” bands, as clear evidencemon markers among homologs as illustrated in Greenthat some chromosomes are not being reliably transmit-German HGs 3 and 8 (Figure 1). For example, Greented, particularly from IND. Only the markers that fitGerman linkage groups 63 and 69 share related locithe expected segregation ratio were used for mapping.detected by four common DNA probes, pSB173, SG212,On L 3 MOL, 31 probes generated 109 markers for LASG202, and pSB167. Other common probes contribut-Purple and 85 for Mol 5829.ing to assembly of this HG 3 include pSB173 and SG305Seventy-two linkage groups were formed for both S.on linkage groups (LGs) 48 and 69; CSU527 andofficinarum Green German and Muntok Java, and 69 forCDSR157 on LGs 48, 49, and 60; pSB173, CSU536, andboth S. spontaneum IND 81-146 and PIN 84-1 (Table 1).SG302 on LGs 60 and 63; CSU537, SG212, SG202,Consistent with its higher chromosome numbers, theUMC147, and pSB167 on LGs 61 and 63; and CSU536total number of polymorphic loci detected was muchand SG305 on LGs 48, 60, and 62. HG 8 was assembledhigher for S. officinarum (737 for GG and 739 for MJ)by LGs 1, 2, 3, 4, 7, 8, 9, 24, 26, and 35.than for S. spontaneum (492 for both IND and PIN).
The overall length of each of the four maps (2304, 1443, Virtually all markers on sugarcane HGs 3 and 8 were
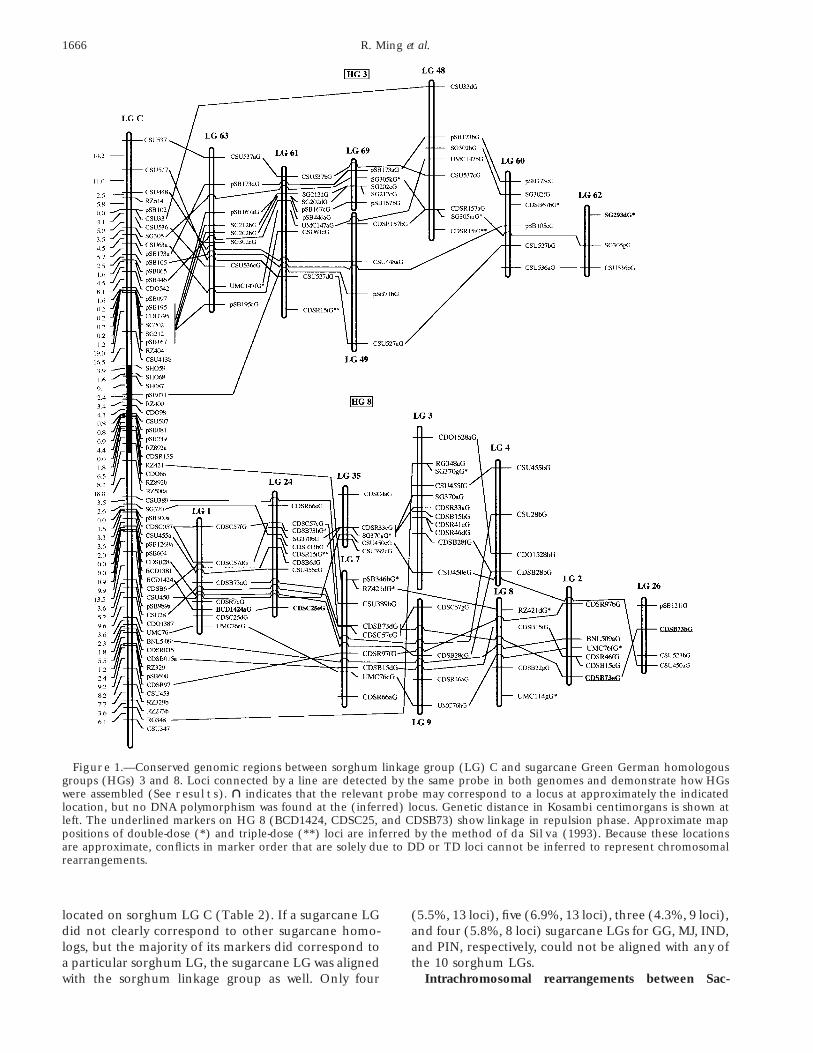
1666 R. Ming et al.
Figure 1.—Conserved genomic regions between sorghum linkage group (LG) C and sugarcane Green German homologousgroups (HGs) 3 and 8. Loci connected by a line are detected by the same probe in both genomes and demonstrate how HGswere assembled (See results). " indicates that the relevant probe may correspond to a locus at approximately the indicatedlocation, but no DNA polymorphism was found at the (inferred) locus. Genetic distance in Kosambi centimorgans is shown atleft. The underlined markers on HG 8 (BCD1424, CDSC25, and CDSB73) show linkage in repulsion phase. Approximate mappositions of double-dose (*) and triple-dose (**) loci are inferred by the method of da Silva (1993). Because these locationsare approximate, conflicts in marker order that are solely due to DD or TD loci cannot be inferred to represent chromosomalrearrangements.
located on sorghum LG C (Table 2). If a sugarcane LG (5.5%, 13 loci), five (6.9%, 13 loci), three (4.3%, 9 loci),and four (5.8%, 8 loci) sugarcane LGs for GG, MJ, IND,did not clearly correspond to other sugarcane homo-
logs, but the majority of its markers did correspond to and PIN, respectively, could not be aligned with any ofthe 10 sorghum LGs.a particular sorghum LG, the sugarcane LG was aligned
with the sorghum linkage group as well. Only four Intrachromosomal rearrangements between Sac-
1667Sugarcane-Sorghum Comparative Mapping
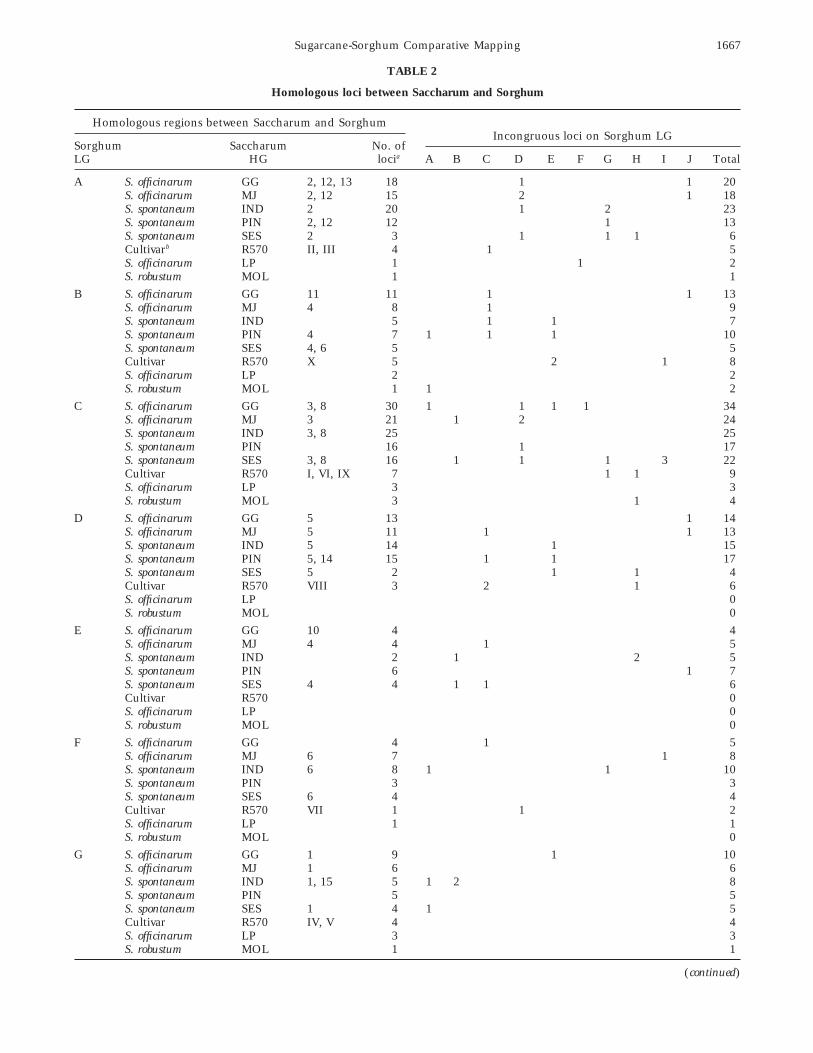
TABLE 2
Homologous loci between Saccharum and Sorghum
Homologous regions between Saccharum and SorghumIncongruous loci on Sorghum LG
Sorghum Saccharum No. ofLG HG locia A B C D E F G H I J Total
A S. officinarum GG 2, 12, 13 18 1 1 20S. officinarum MJ 2, 12 15 2 1 18S. spontaneum IND 2 20 1 2 23S. spontaneum PIN 2, 12 12 1 13S. spontaneum SES 2 3 1 1 1 6Cultivarb R570 II, III 4 1 5S. officinarum LP 1 1 2S. robustum MOL 1 1
B S. officinarum GG 11 11 1 1 13S. officinarum MJ 4 8 1 9S. spontaneum IND 5 1 1 7S. spontaneum PIN 4 7 1 1 1 10S. spontaneum SES 4, 6 5 5Cultivar R570 X 5 2 1 8S. officinarum LP 2 2S. robustum MOL 1 1 2
C S. officinarum GG 3, 8 30 1 1 1 1 34S. officinarum MJ 3 21 1 2 24S. spontaneum IND 3, 8 25 25S. spontaneum PIN 16 1 17S. spontaneum SES 3, 8 16 1 1 1 3 22Cultivar R570 I, VI, IX 7 1 1 9S. officinarum LP 3 3S. robustum MOL 3 1 4
D S. officinarum GG 5 13 1 14S. officinarum MJ 5 11 1 1 13S. spontaneum IND 5 14 1 15S. spontaneum PIN 5, 14 15 1 1 17S. spontaneum SES 5 2 1 1 4Cultivar R570 VIII 3 2 1 6S. officinarum LP 0S. robustum MOL 0
E S. officinarum GG 10 4 4S. officinarum MJ 4 4 1 5S. spontaneum IND 2 1 2 5S. spontaneum PIN 6 1 7S. spontaneum SES 4 4 1 1 6Cultivar R570 0S. officinarum LP 0S. robustum MOL 0
F S. officinarum GG 4 1 5S. officinarum MJ 6 7 1 8S. spontaneum IND 6 8 1 1 10S. spontaneum PIN 3 3S. spontaneum SES 6 4 4Cultivar R570 VII 1 1 2S. officinarum LP 1 1S. robustum MOL 0
G S. officinarum GG 1 9 1 10S. officinarum MJ 1 6 6S. spontaneum IND 1, 15 5 1 2 8S. spontaneum PIN 5 5S. spontaneum SES 1 4 1 5Cultivar R570 IV, V 4 4S. officinarum LP 3 3S. robustum MOL 1 1
(continued)

1668 R. Ming et al.
TABLE 2
(Continued)
Homologous regions between Saccharum and SorghumIncongruous loci on Sorghum LG
Sorghum Saccharum No. ofLG HG locia A B C D E F G H I J Total
H S. officinarum GG 9 4 4S. officinarum MJ 9 5 1 6S. spontaneum IND 9, 16 5 1 1 7S. spontaneum PIN 2 2S. spontaneum SES 0Cultivar R570 0S. officinarum LP 0S. robustum MOL 0
I S. officinarum GG 4 4S. officinarum MJ 0 0S. spontaneum IND 6 1 1 8S. spontaneum PIN 7 5 5S. spontaneum SES 7 5 1 6Cultivar R570 0S. officinarum LP 1 1S. robustum MOL 0
J S. officinarum GG 10 3 3S. officinarum MJ 4 1 5S. spontaneum IND 10 6 1 1 8S. spontaneum PIN 10 7 1 8S. spontaneum SES 5 2 2Cultivar R570 0S. officinarum LP 0S. robustum MOL 0
Total 439 7 6 13 12 13 3 8 7 5 7 520
a Number of loci listed in the table including unassigned LGs corresponding to the Sorghum LG, detail in text.b Grivet et al. 1996.
charum and Sorghum chromosomes: To test possible On the basis of present data, we conclude that at least13 chromosomal rearrangements appear to differenti-intrachromosomal rearrangements between Saccharum
and Sorghum, the markers involved in the possible re- ate sorghum from sugarcane. These rearrangementsinvolved parts of sugarcane HGs 2, 3, 8, and 13 thatarrangement, plus two flanking markers when available,
were used to calculate LOD scores for the alternative corresponded to sorghum LGs A, B, C, and H. Two of13 rearrangements were supported by evidence fromorders. Only if the LOD difference between alternative
orders exceeded 2.0 in both Saccharum and Sorghum two sugarcane species, S. officinarum and S. spontaneum.Among the 11 additional rearrangements found, 5was a rearrangement inferred. Because apparent intra-
chromosomal translocations might actually result from (45%) were detected on more than one LG within S.officinarum or S. spontaneum, while the remaining 6 weremultiple inversions, we referred to both simply as “re-
arrangements.” For example, in GG HG 3 (Figure 1), detected on one LG only (Figure 2, Table 3). No evi-dence that contradicted these rearrangements wasLG 49 showed pSB071 mapping between CSU448 to
CSU527, a major rearrangement relative to sorghum. found within or among Saccharum species; however, inmany cases there were insufficient loci to test the re-On LGs 61 and 63, pSB167, SG212, and SG202 mapped
to the region between CSU536 and CSU537. These two arrangements on other homologs.Interchromosomal rearrangements between Sac-differences in gene order between Saccharum and Sor-
ghum may reflect a single chromosomal rearrangement; charum and Sorghum chromosomes: Among the totalof 10, 9, 10, and 7 putative HGs from GG, MJ, IND, andhowever, we do not yet have DNA markers necessary to
prove this. An inversion on LG 7 of HG 8 was detected PIN, respectively, some were found to be segments ofa single HG when aligned with the sorghum linkageby CDSR97, CDSB15, and UMC76. This inversion was
supported by the positions of CDSR97 and CDSB15 on map (Figure 2, Table 4). For example, sugarcane HGs3 and 8 were likely a single HG corresponding to sor-LG 2, although the position of the DD locus UMC76
was less certain. ghum LG C. One sorghum LG, F, corresponded to only

1669Sugarcane-Sorghum Comparative Mapping
Figure 2.—RFLP linkage maps of sorghum generated from S. bicolor 3 S. propinquum F2 population and corresponding genomicregions from five sugarcane maps (see materials and methods). The filled region in the sorghum map indicates approximatelocation of the centromere based on maize (LGs A, B, C, and F) and wheat (LG D; Paterson et al. 1995) genetic maps. Thesefive sugarcane maps each correspond to the genome of individual parental varieties: Green German (G); Muntok Java (M); IND81-146 (I); PIN 84-1 (P); SES 208 (S). Within each cell, the first number in parentheses is the number of loci detected by thecorresponding DNA probe. The second number is the number of these loci that could be assigned to LGs; zero indicates thatall loci were unlinked to the map. After the letter indicating the parent, the next number is the corresponding HG(s). HGs 1–8are as previously described (Da Silva et al. 1995), and HGs 9–17 are newly identified. The letter A–J followed by a number inthe data column indicates a subset of incongruous (not colinear between Saccharum and Sorghum) loci detected by an otherwisecongruent DNA probe. If it is at the end of a cell, it indicates duplication in the Saccharum genome; if it is at the beginningof a cell, then it is not mapped in this part of the Saccharum genome. For example, the probe CDSR125 on sorghum LG D wasfound in two IND LGs corresponding to sorghum LGs A and D, respectively, hence it is designated [(2/2)I 5: A2]; the sameprobe was found in a GG LG corresponding to sorghum LG A, but not LG D, and designated [A1: (6/1)G 13]. These incongruousloci are also listed on the bottom of each linkage map with the probe name and the LG where the probe maps in sorghum.The approximate position of each incongruous locus in sugarcane is shown to the right of the individual map (data column)that each locus belongs to. Differences in the order of loci between sorghum and sugarcane (possible chromosomal rearrangement)are indicated by a line with arrows at both ends for a possible inversion, or a line with an arrow on one end for a possibletranslocation. For those DNA probes that could not be mapped in sugarcane (far right column), gb indicates backgroundhybridization was prohibitively high; np, no polymorphism; ns, no signal; ls, light signal; d, segregation distortion—markersshowed ,50% present band.

1670 R. Ming et al.
Figure 2.—Continued.
one sugarcane HG. Seven sorghum LGs, C, D, E, G, H, as three chromosomal breaks (one each in LGs B, E,and J).I, and J, each corresponded to 2 sugarcane HGs. The
remaining two sorghum LGs, A and B, each corre- Only one of the possible interchromosomal re-arrangements (HG 4 with LGs B, E) is found in bothsponded to 3 sugarcane HGs. The 17 sugarcane HGs
could be reduced to 10 nonoverlapping sets that corre- S. officinarum and S. spontaneum.Details of correspondence between sugarcane andsponded to different regions of the sorghum ge-
nome—9 of these sets were found in S. officinarum and sorghum maps: A total of 439 loci (84%) detected by 242probes (Figure 2, Table 2) could be compared between9 in S. spontaneum. The missing sets were different in
each species. Saccharum and Sorghum. Their corresponding arrange-ments are as follows:Cases where 1 sugarcane HG corresponds to parts of
two or more sorghum LGs are likely to reflect interchro- Sorghum LG A: This group corresponded to HGs 2,12, and 13 of GG, HGs 2 and 12 of MJ, HG 2 of IND,mosomal rearrangements. Because the sorghum map is
complete, with the number of LGs corresponding to the HGs 2 and 12 of PIN, HG 2 of SES, and HGs II and IIIof R570 (Grivet et al. 1996; Figure 2, Table 2). Thenumber of chromosomes, we can rule out the possibility
that such overlap means that the sorghum LGs are part numbers (and percentages) of loci in the respectivesugarcane maps that support this correspondence areof a single chromosome. Four of the 10 sugarcane HGs
(4, 5, 6, and 10) show correspondence to two sorghum 18 (90%) on 8 LGs in GG, 15 (88%) on 13 LGs in MJ,20 (95%) on 11 LGs in IND, 12 (92%) on 10 LGs inLGs each (B and E, D and J, B and F, and E and J).
These rearrangements could be accounted for by as few PIN, 3 (50%) in SES, and 4 (80%) in R570 (Table 2).

1671Sugarcane-Sorghum Comparative Mapping
Figure 2.—Continued.

1672 R. Ming et al.
Figure 2.—Continued.
New HGs 12 and 13 may be terminal segments of SES arrangement was supported by two LGs in the IND map(S. spontaneum) and one LG in the GG map (S. offici-HG 2. A cluster of 4 loci—CSU382 and SG442 of IND
HG 2, CDSR94 of PIN HG 2, and CDSR153 of SES HG narum), and not contradicted by any LGs, indicatingthat it likely happened before the divergence of the2—were located on sorghum LG G. A second cluster of
4 loci, CDSR125 of GG HG 12 and IND HG 2, and Saccharum species from each other. MJ and PIN did notsegregate at appropriate loci to detect this rearrange-CDSC5 and SHO74 of MJ HG 12 were located on sor-
ghum LG D. CDSB29 of GG and MJ HG 2, and CDSC24 ment.Two other rearrangements were suggested by individ-of SES HG 2 were located on sorghum LG J and LG H,
respectively. Seven (19%) of 36 LG A probes surveyed ual GG and IND linkage groups (respectively), in re-gions for which other LGs did not segregate at thewere not polymorphic in sugarcane. Based on data from
five sugarcane maps, about 90% of sorghum LG A was affected loci. In GG, pSB581 was relocated to the seg-ment between pSB1368 and CDSB62, while in IND, anmapped in sugarcane (Table 5).
An intrachromosomal rearrangement differentiates inversion occurred in the segment between SG293 andBNL9.11 (Figure 2). The corresponding regions of MJsugarcane HG 2 from sorghum LG A, marked by reloca-
tion of the pSB079–CDSC056 interval to a region be- and PIN were too sparsely mapped to test these re-arrangements.tween CSU456 and CDSR067 in sugarcane. This re-

1673Sugarcane-Sorghum Comparative Mapping
Figure 2.—Continued.
Sorghum LG B: This group corresponded to HG 11 of further test this rearrangement did not segregate forpolymorphisms.GG, part of HG 4 of MJ, HG 4 of PIN, parts of HGs 4
and 6 of SES, and HG X of R570. Although about 65% Sorghum LG C: This group corresponded to HGs 3and 8 of GG, HG 8 of MJ, HGs 3 and 8 of IND and SES,of this sorghum LG was collectively covered by the five
sugarcane maps, only small HGs could be formed in and HGs I, VI, and IX of R570. The numbers (andpercentages) of loci supporting correspondence to sor-individual GG, PIN, and SES maps. The numbers (and
percentages) of loci supporting correspondence to sor- ghum LG C are 30 (86%) on 16 LGs in GG, 21 (88%)on 12 LGs in MJ, 25 (100%) on 12 LGs in IND, 16ghum LG B are 11 (85%) from nine LGs in GG, 8 (89%)
from six LGs in MJ, 5 (71%) from five LGs in IND, 7 (94%) on 10 LGs in PIN, 16 (73%) in SES, 7 (78%) inR570. Among 15 exceptions (12%) of the total 122 loci(70%) from seven LGs in PIN, 5 (36%) on SES HGs 4
and 6, and 5 (63%) on R570 HG X. Four contiguous mapped on HGs 3 and 8 in these five sugarcane maps,6 loci detected by 4 probes were located on sorghumloci, CDO686, RZ567, RZ166, CDO1380, of the seven
in SES HG 6 were located on a clustered region of LG D, 4 loci on sorghum LG I, 2 on LG B, and 1 eachon LGs A, E, F, and G (Figure 2; Table 2), respectively.sorghum LG F. Since only half of the HG 4 loci of SES
and MJ, and half of the HG 6 loci of SES were located Six (11%) of 53 probes on LG C surveyed were notpolymorphic in sugarcane, with no marked clustering.on sorghum LG B, this LG may not be well conserved
in sugarcane. Two incongruous loci, CDSR91 in PIN About 90% of sorghum LG C was collectively coveredby the sugarcane maps.and CDSR49 in IND, were located on sorghum LG E.
Four more exceptions, CDSR155 in GG, pSB604 in MJ, An important feature is that two HGs in GG, IND,and SES corresponded to sorghum LG C, with “break-CDSC57 in IND, and CSU536 in PIN mapped to sor-
ghum LG C. CDSR126 in PIN and pSB204 in GG were points” at similar positions. However, HG 8 of MJ ex-tended over these break points. HGs 3 and 8 in INDlocated on sorghum LGs A and J, respectively. Seven
(26%) of 27 probes surveyed on LG B were not polymor- overlapped at locus RZ421. The pooled data from thesefive sugarcane maps indicates that sorghum LG C isphic in sugarcane.
An inversion involving CSU81 and CSU422 was de- conserved as a single HG in sugarcane, which is com-prised of both HGs 3 and 8. These HGs were previouslytected in one LG of GG. In other GG linkage groups,
as well as in MJ, IND, and PIN, the loci needed to thought to be independent (da Silva et al. 1995).

1674 R. Ming et al.
Figure 2.—Continued.
Extensive intrachromosomal rearrangement appears sive intrachromosomal rearrangements are apparent inthis part of the sugarcane genome, only the inversionsto have occurred between sorghum LG C and sugarcane
HGs 3 and 8. An inversion involving loci CSU450, in MJ and IND can be asserted to be consistent acrosstaxa.CSU28, and UMC76 occurred in two LGs of MJ and
two LGs of IND. Two other inversions involving UMC76, Sorghum LG D: This group corresponded to HG 5 ofGG, HGs 5 and 14 of MJ, HG 5 of IND and PIN, partCDSB15, and CDSR97, and SG370 and CDSC57 of GG
(two and one LGs, respectively), were also observed. A of HG 5 of SES, and HG VIII of R570. The numbers(and percentages) of loci supporting correspondencerearrangement in three LGs of GG involving loci
CDSB6, CSU450, and CSU28 was detected in a nearby to sorghum LG D are 13 (93%) loci on 10 LGs in GG,11 (85%) loci on 9 LGs in MJ, 14 (100%) loci on 8 LGs(but different) region to a rearrangement in IND involv-
ing CDSB15, pSB600, and CDSR97. No LG in MJ and of IND, 15 (88%) loci on 14 LGs in PIN, and 3 (50%)loci in R570. HGs 5 and 14 of MJ overlapped on thePIN covered these rearranged regions. Two additional
rearrangements were detected in GG (one and two LGs, same region of sorghum LG D, suggesting that they area single HG. HG 5 of SES covered part of sorghum LGsrespectively), but could not be tested with available loci
in MJ, IND, and PIN. A rearrangement was also observed D and J with only two markers on each LG. Amongexceptions, CDSB29 in GG and MJ were on sorghumin SES from SG212 to the region between RZ421 and
CDO66. This is the same region as one of the GG rear- LG J; CDSR35 in MJ and PIN were on LG C; CSU539in IND, CDSR95 in PIN, and RZ143 in SES were on LGrangements, but in the opposite direction. While exten-

1675Sugarcane-Sorghum Comparative Mapping
Figure 2.—Continued.
E. CDO365 of SES was on LG H. Six out of 32 (19%) made it impossible to detect rearrangements in this partof the sugarcane genome.probes on LG D surveyed were not polymorphic in sug-
arcane. About 75% of sorghum LG D was mapped in Sorghum LG F: This group corresponded largely toHG 6 of MJ, IND, and SES. The numbers (and percent-sugarcane, and no rearrangement could be detected.
Sorghum LG E: This group corresponded to part of ages) of loci supporting correspondence to sorghumLG F are four (80%) on four LGs in GG, seven (88%)HG 10 of GG, part of HG 4 of MJ and SES, and HG X
of R570. A putative recently discovered translocation on six LGs in MJ, eight (80%) on seven LGs in IND,three (100%) on four LGs in PIN, and four (57%) inbetween S. bicolor and S. propinquum (A. H. Paterson,
unpublished data) complicated the analysis of this re- SES. On R570 HG VII, only 2 of the 5 probes weremapped in sorghum; one was on LG D and the othergion. The numbers (and percentages) of loci support-
ing correspondence to sorghum LG E are four (100%) was on LG F. A small HG 6 was assembled in MJ andIND, but there were not enough common markers inon four LGs in GG, four (80%) on four LGs in MJ, two
(67%) on three LGs in IND, six (86%) on seven LGs GG and PIN to form a HG. Among the exceptions, threeloci on SES HG6 were located on sorghum LG B asin PIN, and four (80%) in SES. In R570, the only two
probes corresponding to LG E were located on HG X noted previously. Others are CDSC30 of MJ on LG A,SG305 of GG on LG C, CSU549 of IND on LG G, andthat also partly corresponded to sorghum LG B. HG 4
of MJ and SES also spanned parts of LGs B and E, and CDO87 of MJ on LG I. Seven (25%) of the 28 probessurveyed on LG F were not polymorphic in sugarcane.HG 10 of GG covered segments of LG E and J. Among
four incongruous loci, one was on LG B, two on LG C, About 90% of sorghum LG F was mapped in sugarcane,and no rearrangement could be detected.and one on LG J. Ten (53%) of the 19 LG E probes
surveyed were not polymorphic in sugarcane. About Sorghum LG G: This group corresponded to HG 1 ofGG and MJ, HGs 1 and 15 of IND, HG 1 of SES, and40% of sorghum LG E was mapped in sugarcane. The
short LGs in sugarcane maps corresponding to LG E HGs IV and V of R570. The numbers (and percentages)

1676 R. Ming et al.
Figure 2.—Continued.
of loci supporting correspondence to sorghum LG G mapped between pSB419 and pSB300, supported bytwo LGs. Eleven (55%) of the 20 LG H probes surveyedare nine (82%) on 10 LGs in GG, six (86%) on 6 LGs
in MJ, five (63%) on 5 LGs in IND, five (100%) on 3 showed no polymorphism in G 3 I or P 3 M. Seven ofthe 11 nonpolymorphic loci were clustered in a smallLGs in PIN, four (67%) in SES, and four (100%) in
R570. HG 1 of SES spanned two segmental HGs 1 and genomic region and homologous loci were detectedwithin a very short genetic distance. About 60% of sor-15 of IND, suggesting that they are different parts of
the same HG. Incongruous loci CDSR67 in IND and ghum LG H was mapped in sugarcane.Sorghum LG I: This group corresponded to HGs 7 andSG293 in SES were located on LG A, UMC4 and pSB521
in IND on LG B, and CDSR49 in GG on LG E. Mapped 17 of PIN, and HG 7 of SES. Although nine LG I lociwere genotyped in S. officinarum MJ, none were linked.loci on sugarcane linkage groups varied from two to
eight. Six (29%) of 21 LG G probes surveyed were not Only three LGs were formed in GG. In the other speciesS. spontaneum, seven and five LGs were formed in INDpolymorphic. About 60% of sorghum LG D was mapped
in sugarcane, and no rearrangement could be detected. and PIN, respectively. The numbers (and percentages)of loci supporting correspondence to sorghum LG ISorghum LG H: This group corresponded to HG 9 of
GG and MJ, and HGs 9 and 16 of IND. No HG in SES were four (100%) on three LGs in GG, six (75%) onseven LGs in IND, five (100%) on five LGs in PIN, andcorresponded clearly to sorghum LG H—only CDSC24
on HG 2 and CDO365 on HG 5 were located on sor- five (83%) in SES. Incongruous locus CDSB15 in INDwas on LG C, CDSR49 in SES on LG E, and CDSC42ghum LG H. No probe mapped in R570 corresponded
to LG H. The numbers (and percentages) of loci sup- in IND on LG F. Fourteen (54%) of the 26 LG I probeswere not polymorphic. About 55% of sorghum LG I wasporting correspondence to sorghum LG H are four
(100%) on three LGs in GG, five (83%) on six LGs in mapped in sugarcane, and no rearrangement could bedetected.MJ, five (71%) on five LGs in IND, and two (100%) on
three LGs in PIN. With the limited number of homolo- Sorghum LG J: This group corresponded to part ofHG 10 of GG, HG 10 of IND and PIN, and HG 5 ofgous loci, HG 9 of GG and MJ covered HGs 9 and 16
of IND, suggesting that they are parts of the same HG. SES. The numbers (and percentages) of loci supportingcorrespondence to sorghum LG J were three (100%)Within these HGs, pSB302 in MJ and CDSR95 in IND
were located on sorghum LG E; pSB1108 in IND was on two LGs in GG, four (80%) on four LGs in MJ, six(75%) on seven LGs in IND, and seven (87%) on fiveon LG J.
A rearrangement was observed in GG where pSB1248 LGs in PIN. Incongruous loci pSB289 and pSB95 of MJ

1677Sugarcane-Sorghum Comparative Mapping
Figure 2.—Continued.
were on LGs A and D, respectively, pSB302 of IND on significant difference (P , 0.0001 by paired t -test).Eleven of these intervals could be compared betweenLG E, and pSB240 of PIN on LG H. There were more
loci homologous to LG J in S. spontaneum than S. offici- S. officinarum (GG and MJ) and S. spontaneum (INDand PIN) with total map units of 92.7 and 106.9 cM,narum, a property shared only by LG I. However, homol-
ogous groups were established in both species. Fifteen respectively (data not shown), a nonsignificant differ-ence.(58%) of the 26 LG J probes showed no polymorphism
in populations G 3 I and P 3 M. About 75% of sorghum Preferential pairing: Chromosome pairing behaviorwas investigated by reanalysis of each genetic locus, withLG J was mapped in sugarcane, and no rearrangement
could be detected. segregation data recoded in the opposite linkage phase(Al-Janabi et al. 1994a; da Silva et al. 1995; Grivet etRecombination rate: Disregarding regions where
chromosomal rearrangements were found, 25 sets of al. 1996). Pairwise comparisons were conducted at eachlocus with all linked or unlinked SD markers that aresyntenic chromosomal intervals could be compared for
recombination rate in sorghum and sugarcane (Table putatively borne by corresponding homologous chro-mosomes using a LOD threshold of 3.0.6). Among these, 22 pairs showed more recombination
in sugarcane. Over all 25 intervals, recombination was In GG, 8 (12.5%) of the 72 LG had markers showingevidence of preferential pairing, including one pair of311 cM in sugarcane and 105 cM in sorghum, a highly

1678 R. Ming et al.
Figure 2.—Continued.
loci in HG 5, two pairs in HG 8, and five pairs not yet LP and MOL. There were another 6, 8, 3, 3, 2, and 9proximal duplications in GG, MJ, IND, PIN, MOL, andlinked to HGs (Table 7, Figure 1). Among eight pairs
of markers involved in preferential pairing in GG, three LP, respectively.pairs each were detected by the same probes (CSU393,CDSB6, and CDSB73).
DISCUSSIONPrior reports of preferential pairing in S. robustum
(Al-Janabi et al. 1994a) were strongly supported by pres- Levels and patterns of chromosome structural re-arrangement in Saccharum and Sorghum: Their closeent data. In MOL5829, 7 (50%) of 14 LG and two pairs
of unlinked markers showed evidence of preferential relationship, high degree of colinearity, and cross-hybridization of DNA probes, all impel use of the smallpairing. S. officinarum loci segregating in the same popu-
lation also showed a high degree of preferential pairing. genome of Sorghum to guide molecular mapping andpositional cloning in Saccharum (sugarcane), one ofIn LA Purple, 8 (35%) of 23 LGs and one pair of un-
linked markers showed preferential pairing. the world’s most valuable crops. At least five SaccharumHGs appear to correspond largely if not completely toNo preferential pairing was found in S. spontaneum.
Proximal duplication: A total of 47 proximal duplica- single Sorghum chromosomes (LGs A, C, G, H, and I).In no case does any HG correspond clearly to moretions of individual DNA clones to two or more loci on
the same linkage group (or homologous group) were than two LGs, suggesting that no more than four or fivemajor interchromosomal translocations have occurredobserved. Eight were shared by different Saccharum
species, detected by CDSB28 and BNL9.11 for MJ and since the Saccharum-Sorghum divergence (involvinghomologs of sorghum LGs B, E, and J).IND; CDSR133 for GG and IND; CDSR78 for GG and
PIN; and CDSB4, CDSB35, CDSB67, and CDSC45 for We found only 1 interchromosomal and 2 intrachro-
1679Sugarcane-Sorghum Comparative Mapping
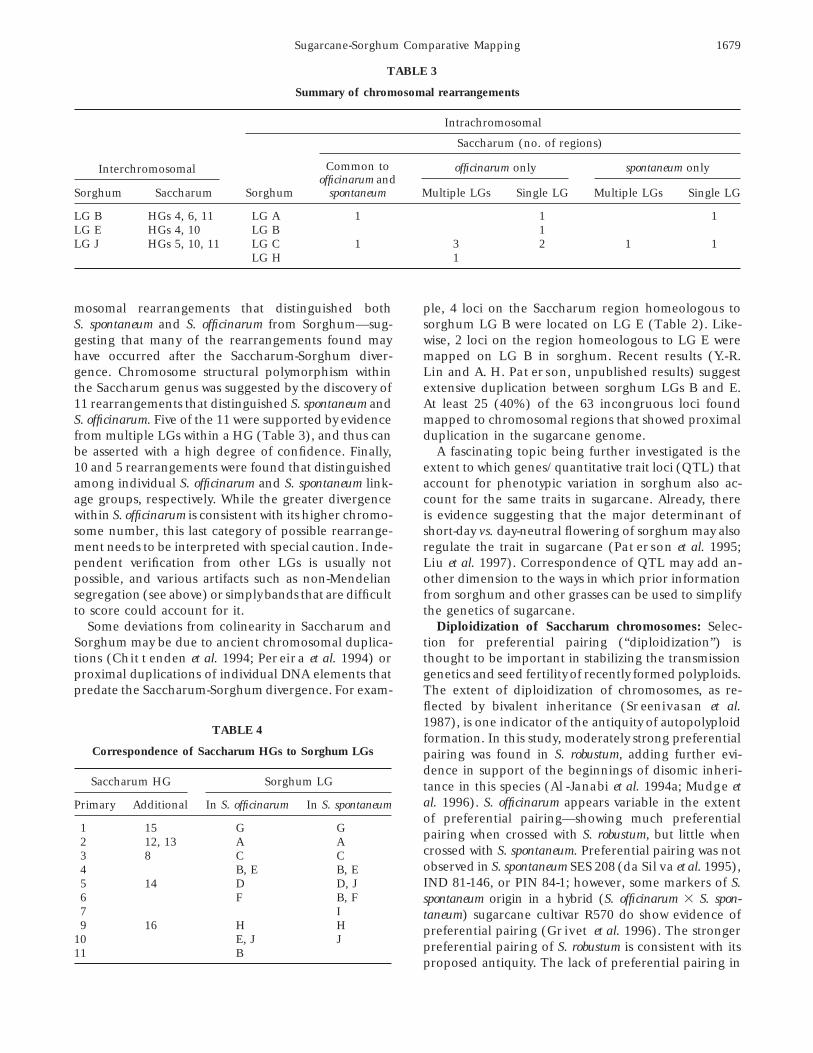
TABLE 3
Summary of chromosomal rearrangements
Interchromosomal
Intrachromosomal
Saccharum (no. of regions)
officinarum only spontaneum onlyCommon toofficinarum and
Sorghum Saccharum Sorghum spontaneum Multiple LGs Single LG Multiple LGs Single LG
LG B HGs 4, 6, 11 LG A 1 1 1LG E HGs 4, 10 LG B 1LG J HGs 5, 10, 11 LG C 1 3 2 1 1
LG H 1
mosomal rearrangements that distinguished both ple, 4 loci on the Saccharum region homeologous tosorghum LG B were located on LG E (Table 2). Like-S. spontaneum and S. officinarum from Sorghum—sug-
gesting that many of the rearrangements found may wise, 2 loci on the region homeologous to LG E weremapped on LG B in sorghum. Recent results (Y.-R.have occurred after the Saccharum-Sorghum diver-
gence. Chromosome structural polymorphism within Lin and A. H. Paterson, unpublished results) suggestextensive duplication between sorghum LGs B and E.the Saccharum genus was suggested by the discovery of
11 rearrangements that distinguished S. spontaneum and At least 25 (40%) of the 63 incongruous loci foundmapped to chromosomal regions that showed proximalS. officinarum. Five of the 11 were supported by evidence
from multiple LGs within a HG (Table 3), and thus can duplication in the sugarcane genome.A fascinating topic being further investigated is thebe asserted with a high degree of confidence. Finally,
10 and 5 rearrangements were found that distinguished extent to which genes/quantitative trait loci (QTL) thataccount for phenotypic variation in sorghum also ac-among individual S. officinarum and S. spontaneum link-
age groups, respectively. While the greater divergence count for the same traits in sugarcane. Already, thereis evidence suggesting that the major determinant ofwithin S. officinarum is consistent with its higher chromo-
some number, this last category of possible rearrange- short-day vs. day-neutral flowering of sorghum may alsoregulate the trait in sugarcane (Paterson et al. 1995;ment needs to be interpreted with special caution. Inde-
pendent verification from other LGs is usually not Liu et al. 1997). Correspondence of QTL may add an-other dimension to the ways in which prior informationpossible, and various artifacts such as non-Mendelian
segregation (see above) or simply bands that are difficult from sorghum and other grasses can be used to simplifythe genetics of sugarcane.to score could account for it.
Some deviations from colinearity in Saccharum and Diploidization of Saccharum chromosomes: Selec-tion for preferential pairing (“diploidization”) isSorghum may be due to ancient chromosomal duplica-
tions (Chittenden et al. 1994; Pereira et al. 1994) or thought to be important in stabilizing the transmissiongenetics and seed fertility of recently formed polyploids.proximal duplications of individual DNA elements that
predate the Saccharum-Sorghum divergence. For exam- The extent of diploidization of chromosomes, as re-flected by bivalent inheritance (Sreenivasan et al.1987), is one indicator of the antiquity of autopolyploid
TABLE 4formation. In this study, moderately strong preferential
Correspondence of Saccharum HGs to Sorghum LGs pairing was found in S. robustum, adding further evi-dence in support of the beginnings of disomic inheri-
Saccharum HG Sorghum LG tance in this species (Al-Janabi et al. 1994a; Mudge etal. 1996). S. officinarum appears variable in the extentPrimary Additional In S. officinarum In S. spontaneumof preferential pairing—showing much preferential
1 15 G G pairing when crossed with S. robustum, but little when2 12, 13 A Acrossed with S. spontaneum. Preferential pairing was not3 8 C Cobserved in S. spontaneum SES 208 (da Silva et al. 1995),4 B, E B, EIND 81-146, or PIN 84-1; however, some markers of S.5 14 D D, J
6 F B, F spontaneum origin in a hybrid (S. officinarum 3 S. spon-7 I taneum) sugarcane cultivar R570 do show evidence of9 16 H H preferential pairing (Grivet et al. 1996). The stronger
10 E, J Jpreferential pairing of S. robustum is consistent with its11 Bproposed antiquity. The lack of preferential pairing in
1680 R. Ming et al.
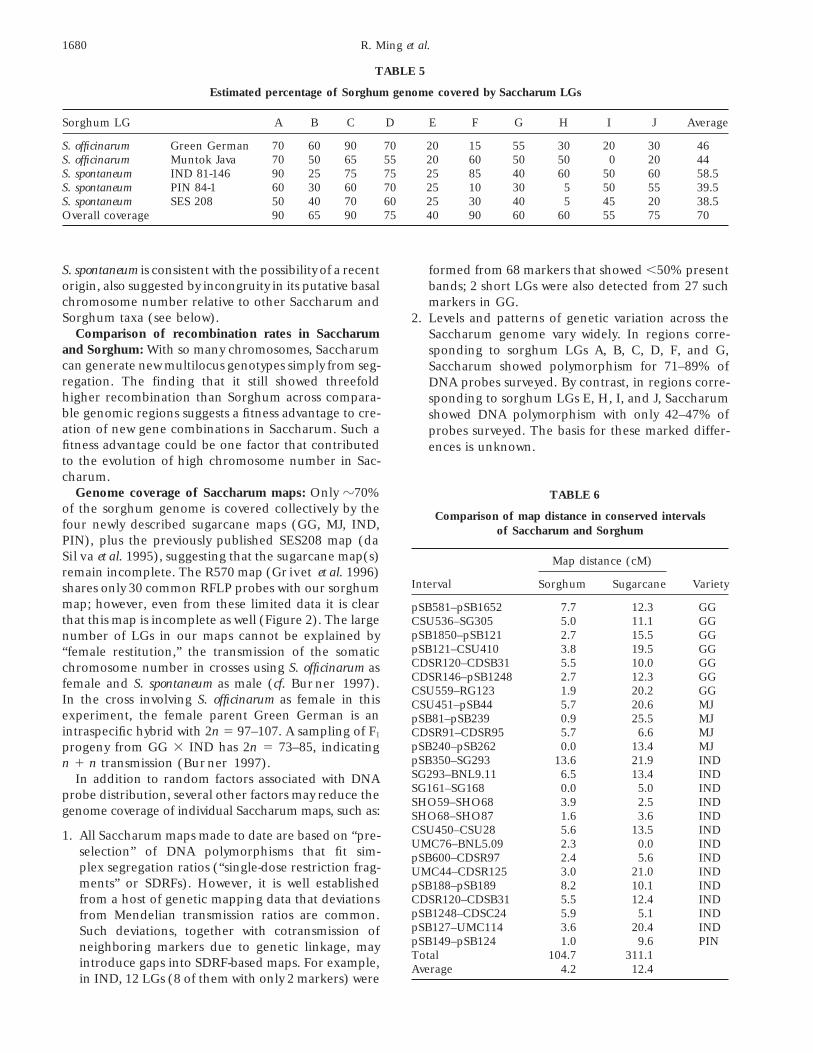
TABLE 5
Estimated percentage of Sorghum genome covered by Saccharum LGs
Sorghum LG A B C D E F G H I J Average
S. officinarum Green German 70 60 90 70 20 15 55 30 20 30 46S. officinarum Muntok Java 70 50 65 55 20 60 50 50 0 20 44S. spontaneum IND 81-146 90 25 75 75 25 85 40 60 50 60 58.5S. spontaneum PIN 84-1 60 30 60 70 25 10 30 5 50 55 39.5S. spontaneum SES 208 50 40 70 60 25 30 40 5 45 20 38.5Overall coverage 90 65 90 75 40 90 60 60 55 75 70
S. spontaneum is consistent with the possibility of a recent formed from 68 markers that showed ,50% presentorigin, also suggested by incongruity in its putative basal bands; 2 short LGs were also detected from 27 suchchromosome number relative to other Saccharum and markers in GG.Sorghum taxa (see below). 2. Levels and patterns of genetic variation across the
Comparison of recombination rates in Saccharum Saccharum genome vary widely. In regions corre-and Sorghum: With so many chromosomes, Saccharum sponding to sorghum LGs A, B, C, D, F, and G,can generate new multilocus genotypes simply from seg- Saccharum showed polymorphism for 71–89% ofregation. The finding that it still showed threefold DNA probes surveyed. By contrast, in regions corre-higher recombination than Sorghum across compara- sponding to sorghum LGs E, H, I, and J, Saccharumble genomic regions suggests a fitness advantage to cre- showed DNA polymorphism with only 42–47% ofation of new gene combinations in Saccharum. Such a probes surveyed. The basis for these marked differ-fitness advantage could be one factor that contributed ences is unknown.to the evolution of high chromosome number in Sac-charum.
Genome coverage of Saccharum maps: Only z70% TABLE 6of the sorghum genome is covered collectively by the
Comparison of map distance in conserved intervalsfour newly described sugarcane maps (GG, MJ, IND, of Saccharum and SorghumPIN), plus the previously published SES208 map (daSilva et al. 1995), suggesting that the sugarcane map(s) Map distance (cM)remain incomplete. The R570 map (Grivet et al. 1996)
Interval Sorghum Sugarcane Varietyshares only 30 common RFLP probes with our sorghummap; however, even from these limited data it is clear pSB581–pSB1652 7.7 12.3 GGthat this map is incomplete as well (Figure 2). The large CSU536–SG305 5.0 11.1 GG
pSB1850–pSB121 2.7 15.5 GGnumber of LGs in our maps cannot be explained bypSB121–CSU410 3.8 19.5 GG“female restitution,” the transmission of the somaticCDSR120–CDSB31 5.5 10.0 GGchromosome number in crosses using S. officinarum asCDSR146–pSB1248 2.7 12.3 GGfemale and S. spontaneum as male (cf. Burner 1997). CSU559–RG123 1.9 20.2 GG
In the cross involving S. officinarum as female in this CSU451–pSB44 5.7 20.6 MJexperiment, the female parent Green German is an pSB81–pSB239 0.9 25.5 MJintraspecific hybrid with 2n 5 97–107. A sampling of F1 CDSR91–CDSR95 5.7 6.6 MJ
pSB240–pSB262 0.0 13.4 MJprogeny from GG 3 IND has 2n 5 73–85, indicatingpSB350–SG293 13.6 21.9 INDn 1 n transmission (Burner 1997).SG293–BNL9.11 6.5 13.4 INDIn addition to random factors associated with DNASG161–SG168 0.0 5.0 INDprobe distribution, several other factors may reduce the SHO59–SHO68 3.9 2.5 IND
genome coverage of individual Saccharum maps, such as: SHO68–SHO87 1.6 3.6 INDCSU450–CSU28 5.6 13.5 IND1. All Saccharum maps made to date are based on “pre-UMC76–BNL5.09 2.3 0.0 IND
selection” of DNA polymorphisms that fit sim- pSB600–CDSR97 2.4 5.6 INDplex segregation ratios (“single-dose restriction frag- UMC44–CDSR125 3.0 21.0 INDments” or SDRFs). However, it is well established pSB188–pSB189 8.2 10.1 IND
CDSR120–CDSB31 5.5 12.4 INDfrom a host of genetic mapping data that deviationspSB1248–CDSC24 5.9 5.1 INDfrom Mendelian transmission ratios are common.pSB127–UMC114 3.6 20.4 INDSuch deviations, together with cotransmission ofpSB149–pSB124 1.0 9.6 PINneighboring markers due to genetic linkage, mayTotal 104.7 311.1
introduce gaps into SDRF-based maps. For example, Average 4.2 12.4in IND, 12 LGs (8 of them with only 2 markers) were

1681Sugarcane-Sorghum Comparative Mapping
Gaps in sugarcane HGs often correspond to locations 92% are 2n 5 80, providing no clue for a basic numberof x 5 8 or x 5 10. Among 95 samples from S. edule,of centromeric DNA in other taxa: Although the sor-
ghum centromeres have not been mapped directly, S. barberi, and S. sinense, 28% can be accounted for bya series involving multiples of 10 (2n 5 60, 70, 80,their possible locations on five LGs could be inferred
from centromeres of maize (LGs A, B, C, and F) and 90, 110), but none by a series involving multiples of 8(excluding 6 samples of 2n 5 80).wheat (LG D) (Paterson et al. 1995). Curiously, many
sugarcane HGs show prominent gaps in the inferred Due to the surprisingly incomplete genome coverageof the Saccharum maps available to date, firm conclu-centromeric regions, in marked contrast to the cluster-
ing of markers near centromeric regions in most plants. sions regarding basal chromosome numbers still remainpremature. Da Silva et al. (1995) reported a linkageSuch gaps separated sugarcane HGs 2 and 12 (MJ), 4
and 11 (GG, MJ, and PIN), and 3 and 8 (GG and IND), map with 8 homologous groups in S. spontaneum, appar-ently supporting the series of eight (above); however,corresponding to sorghum LGs A, B, and C, respectively
(Figures 1 and 2). We have found DNA markers that we show that HGs 3 and 8 reported by da Silva et al.(1995) appear to join up into a single group. Similarly,showed overlap between sugarcane HGs 2 and 12 (MJ),
and 3 and 8 (GG and IND). It remains unclear why Grivet et al. (1996) reported a map with 10 HGs inhybrid R570 (90% S. officinarum), but our comparativesugarcane maps would be marker deficient near the
centromere, in contrast to most other organisms that data reveal that this map covered no more than 6 ofthe previously established sugarcane HGs. Our mapshave been mapped.
Basic chromosome number of Saccharum: The basic appear to suggest a total of 10 HGs for Saccharum. Themaps constructed from G 3 I and P 3 M detect all thechromosome number (x) for Saccharum remains uncer-
tain. In the Andropogoneae tribe, x 5 10 is common previously known HGs plus 3 new HGs (9, 10, and 11).HG 7 was not detected in S. officinarum, while HG 11(Whalen 1991) but not without exceptions, such as
2n 5 6 and 8 for Iseilema (Clayton and Renvoize was not found in S. spontaneum (Table 4).If x 5 10 were the ancestral chromosome number for1986). An ancestral chromosome number of x 5 5 has
been suggested for the Andropogoneae (Celarier Saccharum, as it appears to be for Sorghum, the mostparsimonious explanation of the possible x 5 8 set of1956; Mehra and Sharma 1975; Clayton and Ren-
voize 1986) and is strongly supported by evidence from S. spontaneum would be a recent origin. Saccharum HGs5 and 6 in S. spontaneum each corresponded to parts ofboth isozyme (Wendel et al. 1985) and DNA markers
(Helentjaris et al. 1988) in maize (n 5 10). A growing two sorghum LGs (D and J, and B and F, respectively)while the corresponding regions of S. officinarum corre-body of evidence also supports ancient duplication of
much of the (n 5 10) sorghum genome (Chittenden sponded only to a single sorghum LG (D and F, respec-tively). Fusion of ancestral chromosomes would be oneet al. 1994; Pereira et al. 1994; Lin et al. 1995; Paterson
et al. 1995, 1996a,b). The basic chromosome number possible explanation of both this observation and then 5 8 series of S. spontaneum. A recent origin of S. spon-of Erianthus, a close relative of sugarcane, was suggested
as x 5 10 (D’Hont et al. 1995). taneum might also explain the fewer rearrangementsthat distinguished its chromosomes from one anotherBased on data from 10 sources (J. E. Irvine, unpub-
lished results), chromosome numbers ranging from and the absence of preferential pairing of its chromo-somes. However, the recent origin of S. spontaneum is2n 5 36 to 170 are known in Saccharum. Among 1086
S. spontaneum samples for which chromosome counts contraindicated by its geographic distribution (muchwider than S. robustum) and its generally lower levelswere available, a series involving multiples of 8 (2n 5
40, 48, 56, 64, 72, 80, 88, 96, 112, 120, and 128) accounts of polyploidy. Further characterization of the complexgenome of Saccharum, with the simple genome of Sor-for 77% of the data reported. Among 96 S. robustum
samples, 72% are multiples of 10 (2n 5 60, 70, 80, 90, ghum as a guide, will help to clarify these and otherquestions.100, 110, 140, and 170). Of the 497 S. officinarum samples
We thank Ryan Dalrymple, Melanie Marine, and Linghua Zhu fortechnical assistance, and Zhikang Li, Peter Morrell, and Mark Burow
TABLE 7 for valuable comments. We appreciate funding from the InternationalConsortium for Sugarcane Biotechnology (A.H.P., J.E.I., M.E.S.), U.S.Markers linked in repulsion phase in Green GermanDepartment of Agriculture-Agricultural Research Service and Hawai-ian Sugar Planters’ Association (P.H.M., K.K.W., T.F.W.), CopersucarPair of markers LOD LG (HG)Technologies Corporation (D.B., J.D.S., W.B.), U.S. Department ofAgriculture National Research Initiative (M.E.S.), and Texas (A.H.P.,pSB520fG-pSB1379dG 11.9 LG 6-LG 40J.E.I.) and Cornell (M.E.S.) Agricultural Experiment Stations.CSU393dG-CSU393eG 8.4 LG 50-LG 12 (HG 5)
CDSB6aG-CDSB6eG 6.1 Unlinked-LG 25CDSB73bG-VDSB73eG 5.3 LG 26-LG 2 (HG 8)CDSB54aG-CSU507bG 5.0 Unlinked-unlinked LITERATURE CITEDBCD142aG-CDSC25eG 4.5 LG 1-LG 24 (HG 8)CDSC56bG-CSU553bG 4.0 Unlinked-unlinked Al-Janabi, S. M., R. J. Honeycutt, M. McClelland and B. W. S.
Sobral, 1993 A genetic linkage map of Saccharum spontaneumCDSR85gG-CSU542bG 3.8 Unlinked-unlinkedL. ‘SES 208’. Genetics 134: 1249–1260.

1682 R. Ming et al.
Al-Janabi, S. M., R. J. Honeycutt and B. W. S. Sobral, 1994a Chro- tive mapping of Andropogoneae: Saccharum L. (sugarcane) andits relation to sorghum and maize. Proc. Natl. Acad. Sci. USAmosome assortment in Saccharum. Theor. Appl. Genet. 89: 959–
963. 94: 14261–14266.Helentjaris, T., D. Weber and S. Wright, 1988 Identification ofAl-Janabi, S. M., R. J. Honeycutt, C. Peterson and B. W. S. Sobral,
1994b Phylogenetic analysis of organellar DNA sequences in the genomic locations of duplicate nucleotide sequences in maizeby analysis of restriction fragment length polymorphisms. Genet-the Andropogoneae: Saccharum. Theor. Appl. Genet. 88: 933–944.
Arumuganathan, K., and E. D. Earle, 1991 Nuclear DNA content ics 118: 353–363.Lander, E. S., P. Green, J. Abrahamson, A. Barlow, M. J. Dalyof some important plant species. Plant Mol. Biol. Rep. 9: 208–219.
Burner, D. M., 1997 Chromosome transmission and meiotic behav- et al., 1987 MAPMAKER: an interactive computer package forconstructing primary genetic linkage maps of experimental andior in various sugarcane crosses. J. Am. Soc. Sugar Cane Technol.
17: 38–50. natural populations. Genomics 1: 174–181.Li, Z. K., S. R. M. Pinson, J. W. Stansel and W. D. Park, 1995 Identi-Celarier, R. P., 1956 Cytotaxonomy of the Andropogoneae. 1. Sub-
tribes Dimeriinae and Saccharinae. Cytologia 21: 272–291. fication of QTL for heading date and plant height in rice usingRFLP markers. Theor. Appl. Genet. 91: 374–381.Chittenden, L. M., K. F. Schertz, Y.-R. Lin, R. A. Wing and A. H.
Paterson, 1994 A detailed RFLP map of Sorghum bicolor 3 S. Lin, Y.-R., K. F. Schertz and A. H. Paterson, 1995 Comparativeanalysis of QTLs affecting plant height and maturity across thepropinquum, suitable for high-density mapping, suggests ancestral
duplication of Sorghum chromosomes or chromosomal segments. Poaceae, in reference to an interspecific sorghum population.Genetics 141: 391–411.Theor. Appl. Genet. 87: 925–933.
Christou, P., T. L. Ford and M. Kofron, 1992 The development Liu, S.-J., Y.-R. Lin, J. E. Irvine and A. H. Paterson, 1997 MappingQTLs in autopolyploids, pp. 95–102 in Molecular Dissection of Com-of a variety-independent gene-transfer method for rice. Trends
Biotechnol. 10: 239–246. plex Traits, edited by A. H. Paterson. CRC Press, Boca Raton,FL.Clayton, W. D., and S. A. Renvoize, 1986 Genera Graminum: Grasses
of the World (Kew Bulletin Additional Series XIII). Lubrecht & Mehra, P. N., and M. L. Sharma, 1975 Cytological studies in somecentral and eastern Himalayan grasses I. The Andropogoneae. Cyto-Cramer, London.
da Silva, J., 1993 A methodology for genome mapping of autopoly- logia 40: 61–74.Mudge, J., W. R. Anderson, R. L. Kehrer and D. J. Fairbanks, 1996ploids and its application to sugarcane (Saccharum spp.). Ph.D.
Thesis, Cornell University, Ithaca, NY. A RAPD genetic map of Saccharum officinarum. Crop Sci. 36:1362–1366.da Silva, J., and M. E. Sorrells, 1996 Linkage analysis in polyploids
Paterson, A. H., Y.-R. Lin, Z. K. Li, K. F. Schertz, J. Doebley et al.,using molecular markers, pp. 211–228 in Methods of Genome Analy-1995 Convergent domestication of cereal crops by independentsis in Plants, edited by P. R. Jauhar. CRC Press, Boca Raton, FL.mutations at corresponding genetic loci. Science 269: 1714–1718.da Silva, J., M. E. Sorrells, W. Burnquist and S. D. Tanksley, 1993
Paterson, A. H., T.-H. Lan, K. P. Reischmann, C. Chang, Y.-R. LinRFLP linkage map and genome analysis of Saccharum spontaneum.et al., 1996a Toward a unified genetic map of higher plants,Genome 36: 782–791.transcending the monocot-dicot divergence. Nat. Genet 14: 380–da Silva, J., R. J. Honeycutt, W. Burnquist, S. M. Al-Janabi, M. E.382.Sorrells et al., 1995 Saccharum spontaneum L. ‘SES 208’ genetic
Paterson, A. H., S.-C. Liu, Y.-R. Lin and C. Chang, 1996b Statuslinkage map combining RFLP- and PCR-based markers. Mol.of genome mapping in the tropical grains and grasses (Poaceae),Breed. 1: 165–179.pp. 267–179 in Genome Mapping in Plants, edited by A. H. Pater-Daniels, J., and B. T. Roach, 1987 Taxonomy and evolution inson. Academic Press, San Diego.sugarcane, pp. 7–84 in Sugarcane Improvement Through Breeding,
Pereira, M. G., M. Lee, P. Bramel-Cox, W. Woodman, J. Doebleyedited by D. J. Heinz. Elsevier Press, Amsterdam.et al., 1994 Construction of an RFLP map in sorghum and com-D’Hont, A., Y. H. Lu, P. Feldmann and J. C. Glaszmann, 1993 Cyto-parative mapping in maize. Genome 37: 236–243.plasmic diversity in sugarcane revealed by heterologous probes.
Roach, B. T., 1995 Sugar canes, pp. 160–166 in Evolution of CropSugar Cane 1: 12–15.Plants, edited by J. Smartt and N. W. Simmonds. LongmanD’Hont, A., P. S. Rao, P. Feldmann, L. Grivet, N. Islam-Faridi etGroup, Burnt Mill, England.al., 1995 Identification and characterization of sugarcane inter-
Sobral, B. W. S., D. P. V. Braga, E. S. LaHood and P. Keim, 1994generic hybrids, Saccharum officinarum 3 Erianthus arundinaceus,Phylogenetic analysis of chloroplast restriction enzyme site muta-with molecular markers and DNA in situ hybridization. Theor.tions in the Saccharinae Griseb. Subtribe of the AndropogoneaeAppl. Genet. 91: 320–326. Dumort. Tribe. Theor. Appl. Genet. 87: 843–853.Doggett, H., and K. E. Prasada Rao, 1995 Sorghum, pp. 173–180 Sreenivasan, T. V., B. S. Ahloowalia and D. J. Heinz, 1987 Cytoge-in Evolution of Crop Plants, edited by J. Smartt and N. W. Sim- netics, pp. 211–253 in Sugarcane Improvement Through Breeding,
monds. Longman Group, Burnt Mill, England. edited by D. J. Heinz. Elsevier Press, Amsterdam.Dufour, P., M. Deu. L. Grivet, A. D’Hont, F. Paulet et al., 1997 Wendel, J. F., M. M. Goodman and C. W. Stuber, 1985 Mapping
Construction of a composite sorghum genome map and compari- data for 34 isozyme loci currently being studied. Maize Genet.son with sugarcane, a related complex polyploid. Theor. Appl. Coop. News Lett. 59: 90.Genet. 94: 409–418. Whalen, M. D., 1991 Taxonomy of Saccharum (Poaceae). Baileya
Glaszmann, J. C., Y. H. Lu and C. Lanaud, 1990 Variation of nuclear 23: 109–125.ribosomal DNA in sugarcane. J. Genet. Breed. 44: 191–198. Wu, K. K., W. Burnquist, M. E. Sorrells, T. L. Tew, P. H. Moore
Grivet, L., A. D’Hont, D. Roques, P. Feldmann, C. Lanaud et al., et al., 1992 The detection and estimation of linkage in polyploids1996 RFLP mapping in cultivated sugarcane (Saccharum spp.): using single-dose restriction fragments. Theor. Appl. Genet. 83:genome organization in a highly polyploid and aneuploid inter- 294–300.specific hybrid. Genetics 142: 987–1000.
Guimaraes, C. T., G. R. Sills and B. W. S. Sobral, 1997 Compara- Communicating editor: M. A. Asmussen




















