
Erianthus arundinaceus HSP70 (EaHSP70) overexpression increasesdrought and salinity tolerance in...
12
Plant Science 232 (2015) 23–34 Contents lists available at ScienceDirect Plant Science j ourna l ho me pa ge: www.elsevier.com/locate/plantsci Erianthus arundinaceus HSP70 (EaHSP70) overexpression increases drought and salinity tolerance in sugarcane (Saccharum spp. hybrid) Sruthy Maria Augustine a , J. Ashwin Narayan a , Divya P. Syamaladevi b , C. Appunu a , M. Chakravarthi a , V. Ravichandran c , N. Subramonian a,∗ a Sugarcane Breeding Institute (ICAR), Coimbatore, Tamil Nadu, India b Indian Grass and Fodder Research Institute Regional Station, Avikanagar, Rajasthan, India c Department of Rice, Tamil Nadu Agricultural University, Coimbatore, Tamil Nadu, India a r t i c l e i n f o Article history: Received 1 November 2014 Received in revised form 9 December 2014 Accepted 15 December 2014 Available online 23 December 2014 Keywords: Sugarcane Transformation Drought tolerance Salinity tolerance HSP70 Erianthus arundinaceus a b s t r a c t Heat shock proteins (HSPs) have a major role in stress tolerance mechanisms in plants. Our studies have shown that the expression of HSP70 is enhanced under water stress in Erianthus arundinaceus. In this paper, we evaluate the effects of overexpression of EaHSP70 driven by Port Ubi 2.3 promoter in sugar- cane. The transgenic events exhibit significantly higher gene expression, cell membrane thermostability, relative water content, gas exchange parameters, chlorophyll content and photosynthetic efficiency. The overexpression of EaHSP70 transgenic sugarcane led to the upregulation of stress-related genes. The transformed sugarcane plants had better chlorophyll retention and higher germination ability than con- trol plants under salinity stress. Our results suggest that EaHSP70 plays an important role in sugarcane acclimation to drought and salinity stresses and its potential for genetic engineering of sugarcane for drought and salt tolerance. © 2014 Elsevier Ireland Ltd. All rights reserved. 1. Introduction Erianthus arundinaceus, a wild relative of the genus Saccharum is a broad-leaved species and well known for its drought tolerance [1], high fiber and biomass [2]. It also has resistance to pests and diseases with good ratooning ability. Our previous studies have shown that there is a seven-fold increase in the relative expression of heat-shock protein 70 (HSP70) under water stress in E. arund- inaceus when compared to the commercial sugarcane variety Co 86032. HSPs belong to a class of proteins that are conserved in prokary- otes and eukaryotes and are abundant in plants. Because of the heat shock induction, HSPs are highly expressed in all organisms includ- ing plants [3]. Based on the molecular weight, HSPs are divided into five families: HSP100, HSP90, HSP70, HSP60 and sHSP (small heat shock protein) with a molecular weight ranging from 15 to 42 kDa [4]. The members of the HSP70 family are highly con- served with one HSP70 domain, which have a 44 kDa ATP-binding N-terminal region and a 25–30 kDa substrate-binding C-terminal region. The role of HSP70 family proteins under abiotic stresses has ∗ Corresponding author. E-mail address: monian [email protected] (N. Subramonian). been reported earlier [5]. HSP70 plays different roles during stress and normal conditions. HSP70 stabilizes the proteins by binding to its protein substrates and prevents denaturation or aggregation. In addition, it helps in certain proteins subcellular transport, folding of newly synthesized proteins, the unwanted proteins degradation and protein complexes formation and disassociation. Thus, both during normal and stress conditions, HSP has a major role in protein homeostasis by protein quality controlling and turnover [6]. Previous studies have demonstrated that NtHSP70-1 overex- pression in Nicotiana tabacum plants conferred tolerance to drought stress [7]. The overexpression of HSP70/DnaK gives enhanced heat tolerance as well as early growth and suppression of programmed cell death in transgenic tobacco and rice [8–11]. The HSP70 over- expression in Arabidopsis enhances the tolerance to heat stress and leads to increased transcript levels of two stress-related genes SOS1 and APX1 associated with salinity tolerance [12]. The overexpress- ion of Chrysanthemum HSP70 in Arabidopsis conferred tolerance to abiotic stresses [13]. We have isolated the gene coding for HSP70 from E. arundinaceus and incorporated it into the sugarcane genome. Our results demonstrate that overexpression of EaHSP70 enhances tolerance to drought and salt stresses in sugarcane (Sac- charum spp. hybrid). This is the first report on the overexpression of HSP70 imparting salinity tolerance in food/bioenergy crop plants in general and drought tolerance in sugarcane in particular. http://dx.doi.org/10.1016/j.plantsci.2014.12.012 0168-9452/© 2014 Elsevier Ireland Ltd. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Erianthus arundinaceus HSP70 (EaHSP70) overexpression increasesdrought and salinity tolerance in...

Ed
SMa
b
c
a
ARRAA
KSTDSHE
1
i[dsoi8
osiih4sNr
h0
Plant Science 232 (2015) 23–34
Contents lists available at ScienceDirect
Plant Science
j ourna l ho me pa ge: www.elsev ier .com/ locate /p lantsc i
rianthus arundinaceus HSP70 (EaHSP70) overexpression increasesrought and salinity tolerance in sugarcane (Saccharum spp. hybrid)
ruthy Maria Augustinea, J. Ashwin Narayana, Divya P. Syamaladevib, C. Appunua,. Chakravarthia, V. Ravichandranc, N. Subramoniana,∗
Sugarcane Breeding Institute (ICAR), Coimbatore, Tamil Nadu, IndiaIndian Grass and Fodder Research Institute Regional Station, Avikanagar, Rajasthan, IndiaDepartment of Rice, Tamil Nadu Agricultural University, Coimbatore, Tamil Nadu, India
r t i c l e i n f o
rticle history:eceived 1 November 2014eceived in revised form 9 December 2014ccepted 15 December 2014vailable online 23 December 2014
a b s t r a c t
Heat shock proteins (HSPs) have a major role in stress tolerance mechanisms in plants. Our studies haveshown that the expression of HSP70 is enhanced under water stress in Erianthus arundinaceus. In thispaper, we evaluate the effects of overexpression of EaHSP70 driven by Port Ubi 2.3 promoter in sugar-cane. The transgenic events exhibit significantly higher gene expression, cell membrane thermostability,relative water content, gas exchange parameters, chlorophyll content and photosynthetic efficiency. Theoverexpression of EaHSP70 transgenic sugarcane led to the upregulation of stress-related genes. The
eywords:ugarcaneransformationrought tolerancealinity toleranceSP70rianthus arundinaceus
transformed sugarcane plants had better chlorophyll retention and higher germination ability than con-trol plants under salinity stress. Our results suggest that EaHSP70 plays an important role in sugarcaneacclimation to drought and salinity stresses and its potential for genetic engineering of sugarcane fordrought and salt tolerance.
© 2014 Elsevier Ireland Ltd. All rights reserved.
. Introduction
Erianthus arundinaceus, a wild relative of the genus Saccharums a broad-leaved species and well known for its drought tolerance1], high fiber and biomass [2]. It also has resistance to pests andiseases with good ratooning ability. Our previous studies havehown that there is a seven-fold increase in the relative expressionf heat-shock protein 70 (HSP70) under water stress in E. arund-
naceus when compared to the commercial sugarcane variety Co6032.
HSPs belong to a class of proteins that are conserved in prokary-tes and eukaryotes and are abundant in plants. Because of the heathock induction, HSPs are highly expressed in all organisms includ-ng plants [3]. Based on the molecular weight, HSPs are dividednto five families: HSP100, HSP90, HSP70, HSP60 and sHSP (smalleat shock protein) with a molecular weight ranging from 15 to2 kDa [4]. The members of the HSP70 family are highly con-
erved with one HSP70 domain, which have a 44 kDa ATP-binding-terminal region and a 25–30 kDa substrate-binding C-terminalegion. The role of HSP70 family proteins under abiotic stresses has
∗ Corresponding author.E-mail address: monian [email protected] (N. Subramonian).
ttp://dx.doi.org/10.1016/j.plantsci.2014.12.012168-9452/© 2014 Elsevier Ireland Ltd. All rights reserved.
been reported earlier [5]. HSP70 plays different roles during stressand normal conditions. HSP70 stabilizes the proteins by binding toits protein substrates and prevents denaturation or aggregation. Inaddition, it helps in certain proteins subcellular transport, foldingof newly synthesized proteins, the unwanted proteins degradationand protein complexes formation and disassociation. Thus, bothduring normal and stress conditions, HSP has a major role in proteinhomeostasis by protein quality controlling and turnover [6].
Previous studies have demonstrated that NtHSP70-1 overex-pression in Nicotiana tabacum plants conferred tolerance to droughtstress [7]. The overexpression of HSP70/DnaK gives enhanced heattolerance as well as early growth and suppression of programmedcell death in transgenic tobacco and rice [8–11]. The HSP70 over-expression in Arabidopsis enhances the tolerance to heat stress andleads to increased transcript levels of two stress-related genes SOS1and APX1 associated with salinity tolerance [12]. The overexpress-ion of Chrysanthemum HSP70 in Arabidopsis conferred toleranceto abiotic stresses [13]. We have isolated the gene coding forHSP70 from E. arundinaceus and incorporated it into the sugarcanegenome. Our results demonstrate that overexpression of EaHSP70
enhances tolerance to drought and salt stresses in sugarcane (Sac-charum spp. hybrid). This is the first report on the overexpression ofHSP70 imparting salinity tolerance in food/bioenergy crop plantsin general and drought tolerance in sugarcane in particular.
2 ant Sc
2
2
naga093f5l(bcsEGrst(fmw8fw(At(iha
2
l(wgf5ttefigCb
2
csefpet
4 S.M. Augustine et al. / Pl
. Materials and methods
.1. Vector construction and plant transformation
The HSP70 gene was isolated from E. arundinaceus (accessiono. IK 76-81) from the germplasm collection being maintainedt the Sugarcane Breeding Institute, Coimbatore, India, usingene-specific primers designed from Sorghum bicolor (NCBIccession no. U41653) and Zea mays (NCBI accession no. NM01196236) through polymerase chain reaction (PCR) (1 cycle at4 ◦C for 5 min, followed by 35 cycles of 94 ◦C for 45 s, 62 ◦C for0 s and 72 ◦C for 2 min). Port Ubi 2.3 promoter was amplifiedrom pPORT-UBI-GFP [14] using two specific primers: forward′-GATCGGATCCACTATCACCCTCGAGGTG-3′ (BamHI site under-
ined) and reverse, 5′-ATCCCATGGCTGCAAGAAATAATCACCAA-3′
NcoI site underlined), and cloned into pCAMBIA-3301etween the multiple cloning site SacI and SbfI. The newlyonstructed plasmid was named pPORT-UBI2.3-3301. Afterequencing and confirmation, the full-length sequence ofaHSP70 was amplified using two specific primers: forward 5′-ATTCCTGCAGGATGGCCAAGGGCGAG-3′ (SbfI site underlined) and
everse 5′-GCTCAAGCTTTTAGTCGACCTCCTCGATCTTGG-3′ (HindIIIite underlined), then cloned into pPORT-UBI2.3-3301 betweenhe SbfI and HindIII and the resultant construct pSBI-HSP70Fig. 1) was transferred to Agrobacterium tumefaciens (LBA4404) byreeze–thaw method [15]. For Agrobacterium-mediated transfor-
ation in sugarcane, the method described by Arvinth et al. [16]as followed. Meristematic leaf whorls from the shoot tips of 5- to
-month-old field-grown plants were co-cultivated with A. tume-aciens (LBA4404) harboring pSBI-HSP70, and the transformed cells
ere selected with repeated sub-culturing in a callusing mediumsupplementary Table 3) containing phosphinothricin (3 mg/l).fter six rounds of selection when the calli have attained a size,
he selected calli were transferred to the regeneration mediumsupplementary Table 3) and the regenerated plants were rootedn the rooting medium (supplementary Table 3) and subsequentlyardened and planted in pots containing a mixture of sand, soilnd farmyard manure (1:1:1 ratio).
.2. Molecular analyses and selection of transgenic events
PCR analysis was carried out using the genomic DNA iso-ated from phosphinothricin-tolerant plants. DNA from wild typeWT) plants was used as negative control and plasmid DNAas used as positive control. For PCR, plasmid DNA (10 ng) and
enomic DNA (50 ng) were used as templates. The promoter-geneusion primer sequences 5′-GCATGTGTGAATGGTGCGATTTG-3′ and′-ATGACACCGGCATCCTTGGT-3′ were used for the PCR reac-ion to distinguish EaHSP70 from host plant HSP70. Plants withhe expected size amplicon (1.2 kb) were selected for furtherxperiments. Further, these transgenic events were again con-rmed by amplifying the 300-bp amplicon marker gene (barene) by using primers 5′-AACCACTACATCGAGACAA-3′ and 5′-AACCACGTCTTGAAGCCC-3′ (1 cycle at 94 ◦C for 5 min, followedy 35 cycles of 94 ◦C for 45 s, 51 ◦C for 30 s and 72 ◦C for 30 s).
.3. Plant material and stress treatments
Twenty-five independent transgenic events (V0) of the sugar-ane variety Co 86032 were further propagated through plantingingle bud cuttings from the V0 plants, thus taking the transgenicvents to V1 stage (V1 – vegetative generation). Morphological dif-
erences were not observed between the transgenic events and WTlants (untransformed Co 86032). Five biological replications fromach transgenic event and WT plants were planted in 16′′ pots con-aining soil, sand and farmyard manure in 1:1:1 ratio for moistureience 232 (2015) 23–34
stress screening. Plants were grown with normal irrigation andrecommended fertilizer doses for 120 days. Plants were subjectedto water stress at the tillering phase (120 days after planting) bywithholding irrigation for 10 days. Plants received regular water-ing from the 11th day of water stress, and subsequently regularwatering was continued as required. Relative water content andcell membrane thermostability tests were carried out in the trans-genics and WT plants on 0th and 10th day of drought induction.EaHSP70 transgene expression was analyzed under irrigated con-dition, and endogenous HSP70 in WT sugarcane was assessed indifferent soil moisture stresses, i.e. 0th day (25% soil moisture), 10thday after stress (8.1% soil moisture) and 5 days after the release ofstress (25% soil moisture). Stress-related gene expression and gasexchange parameters were measured in the transgenics and WTplants on the 10th day after inducing drought stress. Chlorophyllcontent and photosynthetic efficiency (Fv/Fm) were recorded onthe 0th day, 10th day after withholding the irrigation and 15 daysafter releasing the drought. All the physiological experiments wereconducted for the 25 independent transgenic events with five bio-logical and three technical replications. The molecular experiment,i.e. gene expression analysis, was conducted in five independenttransgenic events with five biological and three technical replica-tions. Soil moisture percentage was measured through gravimetricmethod using a moisture analyser (A&D model Mx-50) in samplescollected from three different layers (10, 20 and 30 cm). To eval-uate the extent of salinity tolerance of the transgenic events, leafdisk and bud germination assays were carried out. Leaf disk assaywas carried out in the leaves collected from the transgenics andWT plants grown under normal irrigated conditions. The assay wasconducted for 25 independent transgenic events with five biologi-cal and three technical replications. The bud germination assay wascarried out for 10 independent transgenic events selected from leafdisk assay with five biological and three technical replications.
2.4. Morphological analysis (visual scoring) of the transgenicevents
The transgenic events along with the WT were visually observedfor wilting of leaves on the 5th and 10th day after drought induction.Leaf wilting was scored on a scale of 1–4 modified from [17] asfollows: 1 = no wilting; 2 = slightly wilting; 3 = wilting, wherein theplant showed leaf wilting only during hot hours from which theleaves recovered; and 4 = severe wilting, wherein wilted leaves didnot recover. The mean score was computed from five plants fromeach event.
2.5. Cell membrane thermostability analysis
The cell membrane thermostability test was carried out for esti-mating the cell membrane injury percentage, following the methoddescribed by Martineau et al. [18]. The cell membrane injury per-centage is considered as one of the indications of the ability ofthe plant to tolerate drought [19]. This parameter was studied inboth V0 and V1 transgenic generations and compared with WTplants. The third fully opened leaf was collected from transgenicsand WT plants on 0th and 10th day after inducing drought con-dition, as mentioned above. Leaf discs (0.5 cm diameter) weighing200 mg were washed with 20 ml distilled water three times, eachwash extending to 2 min. Immediately after washing, 20 ml of dis-tilled water was added to each of the control and treatment tubes(2.5 cm × 15 cm), the tube mouths were covered with aluminumfoil, the tubes were incubated at 60 ◦C in a thermostatically con-
trolled water bath for 20 min and thereafter both treatment andcontrol sets were placed at 10 ◦C for 12 h to allow the diffusionof electrolytes into the water. After recording the initial conduc-tance at 30 ◦C, the tubes were heated at 100 ◦C for 20 min, and
S.M. Augustine et al. / Plant Science 232 (2015) 23–34 25
of th
fi%ud
2
ddfsasbiiSro
R
2
ctdWIat1ceat
Fig. 1. Schematic diagram
nal conductance was recorded after cooling. Membrane injury = 1 − [(1 − T1/T2)/(1 − C1/C2)] × 100, where T and C refer to the val-es for treatment and control samples, and the subscripts 1 and 2enote the initial and final conductance, respectively.
.6. Plant water status
The relative water content (RWC) of the leaf was evaluated toetermine the leaf hydration status. The excised third leaves of 120-ay-old transgenics and WT plants were subjected to measurementor leaf RWC on the 0th and 10th day after the induction of droughttress. Leaf RWC was calculated based on fresh (FW), turgid (TW)nd dry weights of 200 mg leaf samples. Fresh weights of the leafamples were determined on a mass balance immediately aftereing excised from the plants. TWs were determined after soak-
ng the leaf discs in de-ionized water for 4 h at room temperaturen a closed Petri dish and weighed immediately after blot drying.amples were then dried at 90 ◦C for 72 h and the dry weight wasecorded. Then, the RWC was determined according to the methodf Barrs and Weatherley [20] based on the following calculation:
WC = FW − DWTW − DW
× 100
.7. Gas exchange parameters
Gas exchange parameters, photosynthesis rate (A), stomatalonductance (gs) and transpiration rate (E), were recorded fromhe middle part of the fully opened third leaf on the 10th day ofrought induction (soil moisture 8.1%) from the transgenics andT plants using a portable photosynthesis system (Li-6400, Li-COR
nc., Lincoln, Nebraska, USA) with a leaf chamber of 2 cm × 3 cm andn integrated light source (LI-6400-02B). Gas exchange parame-ers were measured at 40 ◦C (closed cabinet incubated to 40 ◦C),30 �mol m−2 s−1 [CO2] and RH ∼60–70%. The external CO2
oncentration in air was maintained at 380-�mol mol−1 in the ref-rence cuvette. All the gas exchange parameters were measured atleaf temperature of 32 ± 2.0 ◦C, and photon flux density was con-rolled at 1500 �mol m−2 s−1, which gave the light-saturated rates
e plant expression vector.
of photosynthesis (Asat). Measurements were recorded every 30 sover a period of 2 h. Data were recorded for 25 independent eventswith five biological replications and three technical replications perevent.
2.8. Leaf chlorophyll content measurements
Total chlorophyll content was recorded on the 0th and 10thday after the induction of the drought (soil moisture 25% and 8.1%,respectively) and 15 days after the release of drought (re-irrigation,25% soil moisture) from the transgenics and WT plants using achlorophyll meter (SPAD-502; Konica Minolta, Japan).
2.9. Photosynthetic efficiency (Fv/Fm) of the HSP70 transgenicsugarcane
The maximum potential photochemical efficiency, definedas the ratio of variable to maximum fluorescence emitted bychlorophyll (Fv/Fm), was recorded using a portable chlorophyllfluorimeter (OS-30P; Optisciences, USA). Photosynthetic efficiency(Fv/Fm) was recorded on the 0th and 10th day after the induc-tion of drought (soil moisture 8.1%) and 15 days after the releaseof drought (re-irrigation, soil moisture 25%) from the transgenicsand WT plants. The plants were dark-adapted for 20 min beforemeasurement. The maximum fluorescence under light exposure(Fm′) was obtained by imposing a 1-s saturating flash to the leafin order to reduce the entire PS II reaction center after attainingsteady-state fluorescence (Ft). The minimum fluorescence imme-diately after light exposure (Fo′) was determined by imposing darkwhile simultaneously switching on a far red light to oxidize PS IIrapidly by drawing electrons from PS II to PS I.
2.10. Gene expression analysis using quantitative real time PCR(qPCR)
Total RNA was isolated using the method described by Chom-czynski and Mackey [21]. RNA was isolated by using Trizol reagent(Sigma Chemicals, USA) and DNAse (Fermentas International Inc.,Ontario, Canada) treatments were carried out. First-strand cDNAs

26 S.M. Augustine et al. / Plant Sc
Table 1Visual scoring of pSBI-HSP70 transgenic events for soil moisture stress.
Event no. Score after stress induction
5th Day 10th Day
1 2 22 1 23 1 14 1 15 1 36 1 17 1 18 1 19 1 1
10 1 111 1 212 2 213 1 214 1 215 1 316 1 117 1 118 1 219 1 320 1 321 1 222 1 123 1 124 1 1
wsIiasHcasgsrti�rmtifiPtggsccatpMp(btAo
25 1 2WT 3 4
ere synthesized from total RNA using the Fermentas first-trand cDNA synthesis kit using oligo (dT) primers (Fermentasnternational Inc., Ontario, Canada) following the manufacture’snstruction. The relative expression of the transgene EaHSP70 wasssessed in transgenics and WT under normal irrigated and droughttress conditions. In addition, the expression of the endogenousSP70 was also analyzed in WT sugarcane at different soil moistureonditions, i.e. 25% (irrigated condition), 8.1% (drought stressed)nd 25% (5 days after the release of stress). The relative expres-ions of six important abiotic stress (salt, cold and heat) responsiveenes, i.e. RD29 (response to dehydration), LEA (late embryogene-is abundant protein), COR15 (cold regulated protein), ERD (earlyesponsive to dehydration)] [22], ERF (ethylene responsive fac-or) [23] and BRICK/HSPC300 (unpublished data), were assessedn transgenics and WT plants under drought stress condition. The-actin transcript was used as an internal control to quantify the
elative transcript levels. The expression of �-actin was deter-ined in both irrigated and drought-stressed samples to make sure
hat this gene did not respond to drought stress, to use it as annternal control. cDNA fragments and the actin gene were ampli-ed with gene-specific primers. The primers were designed usingrimer Express, version 3 software (Applied Biosystems, Burling-on, ON, Canada) (supplementary data Table 1). The stress-relatedenes used for primer synthesis and their accession numbers areiven in Table 1 (supplementary data Table 1). For each primeret, the primer efficiency was analyzed by constructing standardurves with correlation coefficients of 0.99 or higher, and PCR effi-iencies (calculated by efficiency = [10(−1/slope)] − 1) between 97%nd 101% were accepted. Prior to the qPCR, primer concentra-ions were optimized through melt-curve analysis. All PCRs wereerformed for 40 cycles. qPCR was performed using MESAGREENaster Mix (12.5 �l), Taq DNA polymerase (1 U), gene-specific
rimers (10 pmol each) and the StepOne real-time PCR systemApplied Biosystems, Burlington, ON, Canada). The CT values for
oth the target and internal control genes were used for quan-ification of transcripts by comparative CT method normalization.ll reactions were performed in triplicates, and the expressionf target gene was calculated using the formula: 2−��Ct, whereience 232 (2015) 23–34
��Ct = �Ct sample − �Ct actin [24]. The ��Ct values reflect therelative expression of the target gene upon exposure to droughtstress.
2.11. Leaf disk assay for assessing salinity tolerance
Leaf disk assay was carried out to evaluate the sensitivity ofthe transformed and WT plants to sodium chloride (NaCl) stressas described by Fan et al. [25]. Two grams of fresh leaf sample (each3.0 cm × 2.0 cm size) was excised from healthy and fully expandedthird leaves of the 25 independent transgenic events and WT plants(120 days old). The discs were floated in a 10 ml solution of 250 mMNaCl and experimental control (0 mM NaCl), kept in water for 72 h,and then used for measuring chlorophyll content spectrophotomet-rically after extraction in methanol [26]. The treatment was carriedout in continuous white light at 25 ± 2 ◦C.
2.12. Bud germination assay
Ten independent transgenic events were selected for bud germi-nation assay based on the leaf disk senescence assay. Five biologicalreplications of transgenic events and WT plants single bud settswere planted in cavity trays and grown under identical controlledconditions in a green house. To evaluate the germination under saltstress conditions, single bud setts were irrigated daily with 0, 100,200 and 300 mM NaCl solution for a period of 26 days. The numberof buds germinated and their shoot heights were recorded on the26th day of planting.
2.13. Statistical analysis
For statistical analysis of the data, five biological and three tech-nical replicates from each of the transgenic events were used. Meanvalue, standard deviation and Student’s t-test were evaluated usingthe XLSTAT 2013.5 program to analyze all the data to compare thetransgenic events and WT plants under normal and stress condi-tions. A P-value of (P ≤ 0.05) was considered statistically significant.
3. Results
3.1. Isolation of HSP70 gene from E. arundinaceus anddevelopment of transgenic sugarcane overexpressing EaHSP70gene
The HSP70 gene (1.9 kb) was isolated from E. arundinaceus andthe sequence was submitted to Genbank (accession no. KJ670162).The primers were designed from Z. mays and S. bicolor, so theamplified HSP70 may be possibly a chimera. The gene was namedEaHSP70 and did not have any intron sequence. The isolated genesequence had high sequence homology (98–100%) with the otherreported HSP70 gene sequences of Z. mays and S. bicolor. Thenucleotide sequences translated to amino acid sequence showedhigh sequence homology (95–98%) with other reported HSP70 pro-tein sequences.
To establish the functional significance of the EaHSP70 gene,the complete gene sequence was overexpressed in sugarcane byusing Agrobacterium-mediated transformation. The preliminaryscreening of phosphinothricin-resistant sugarcane transformantswas confirmed through PCR using promoter-gene fusion primers(supplementary Fig. 1A) to distinguish the EaHSP70 from host plantHSP70 and marker gene (supplementary Fig. 1B) at V0 and V1 stages.
Thirty-five independent events were obtained, and 25 events wereselected based on the cell membrane thermostability for subse-quent analysis. The morphological and growth characteristics ofthe transgenic plants were similar to those of WT (Fig. 2A).
S.M. Augustine et al. / Plant Science 232 (2015) 23–34 27
F stressa y toler
3t
fst
Fsm
ig. 2. Screening of EaHSP70 transgenic events along with the WT for soil moisturet 25% soil moisture and (B) at 8.1% soil moisture. Bud germination assay for salinit
.2. Cell membrane thermostability is higher in EaHSP70ransgenic sugarcane events at V0 stage
A standard test of cellular membrane thermostability was per-ormed at the V0 stage to study drought tolerance in transgenicugarcane containing EaHSP70 (Fig. 3A). Though there was varia-ion between the transgenic events, high membrane stability was
ig. 3. Cell membrane thermostability in EaHSP70 transgenics along with WT (A) V0 staghows a significant difference in the CMS between transgenic events and untransformedean ± SD (n = 5) and error bars represent SD.
and bud germination assay for salinity stress. Screening for soil moisture stress (A)ance (C) WT and (D) EaHSP70 transgenic events.
observed in HSP70 transgenics under normal irrigated condition.A high membrane injury indicates low cell membrane stability.Sixty percent of the events had 60–70% membrane injury, whereas
in WT plants the membrane injury was 95.5%. The highest sta-bility was observed in the second event, with 50.8% injury. Thelowest stability was in the seventh event, with 95.11% membraneinjury.e at 25% soil moisture and (B) V1 stage with and without soil moisture stress. Data control, ns: not significant; at P ≤ 0.05, by Student’s t-test. Data are presented as
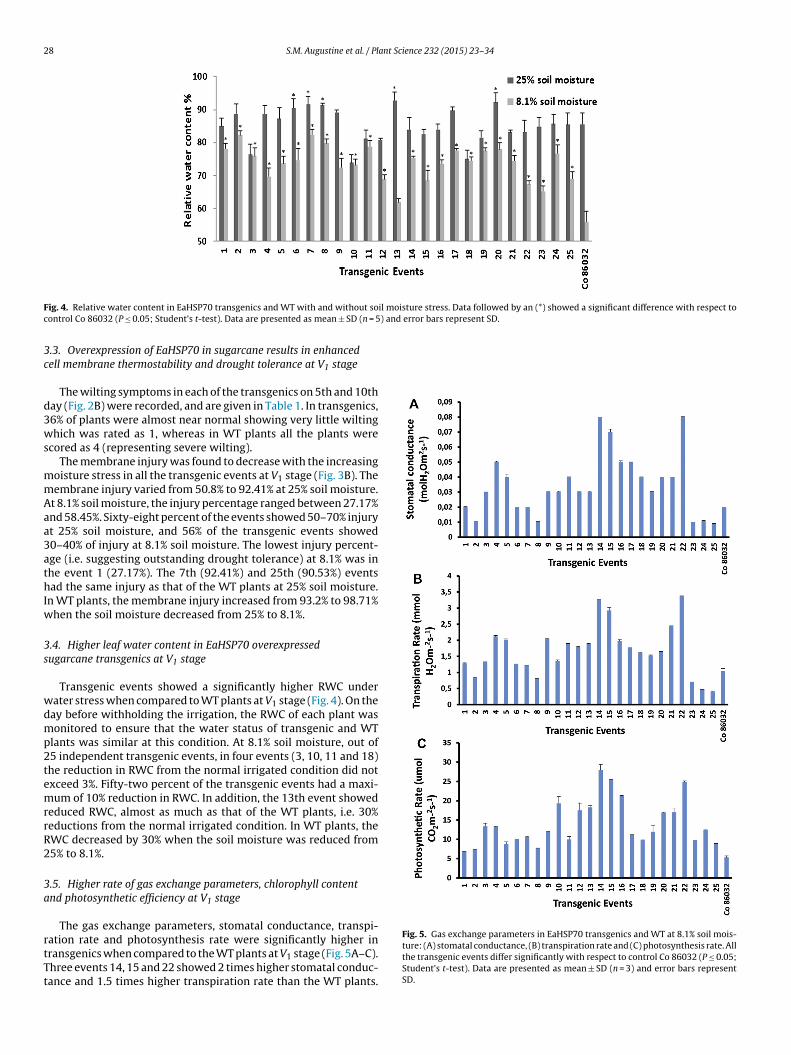
28 S.M. Augustine et al. / Plant Science 232 (2015) 23–34
F il moisture stress. Data followed by an (*) showed a significant difference with respect toc ) and error bars represent SD.
3c
d3ws
mmAaa3athIw
3s
wdmp2temrrR2
3a
rtTt
ig. 4. Relative water content in EaHSP70 transgenics and WT with and without soontrol Co 86032 (P ≤ 0.05; Student’s t-test). Data are presented as mean ± SD (n = 5
.3. Overexpression of EaHSP70 in sugarcane results in enhancedell membrane thermostability and drought tolerance at V1 stage
The wilting symptoms in each of the transgenics on 5th and 10thay (Fig. 2B) were recorded, and are given in Table 1. In transgenics,6% of plants were almost near normal showing very little wiltinghich was rated as 1, whereas in WT plants all the plants were
cored as 4 (representing severe wilting).The membrane injury was found to decrease with the increasing
oisture stress in all the transgenic events at V1 stage (Fig. 3B). Theembrane injury varied from 50.8% to 92.41% at 25% soil moisture.t 8.1% soil moisture, the injury percentage ranged between 27.17%nd 58.45%. Sixty-eight percent of the events showed 50–70% injuryt 25% soil moisture, and 56% of the transgenic events showed0–40% of injury at 8.1% soil moisture. The lowest injury percent-ge (i.e. suggesting outstanding drought tolerance) at 8.1% was inhe event 1 (27.17%). The 7th (92.41%) and 25th (90.53%) eventsad the same injury as that of the WT plants at 25% soil moisture.
n WT plants, the membrane injury increased from 93.2% to 98.71%hen the soil moisture decreased from 25% to 8.1%.
.4. Higher leaf water content in EaHSP70 overexpressedugarcane transgenics at V1 stage
Transgenic events showed a significantly higher RWC underater stress when compared to WT plants at V1 stage (Fig. 4). On theay before withholding the irrigation, the RWC of each plant wasonitored to ensure that the water status of transgenic and WT
lants was similar at this condition. At 8.1% soil moisture, out of5 independent transgenic events, in four events (3, 10, 11 and 18)he reduction in RWC from the normal irrigated condition did notxceed 3%. Fifty-two percent of the transgenic events had a maxi-um of 10% reduction in RWC. In addition, the 13th event showed
educed RWC, almost as much as that of the WT plants, i.e. 30%eductions from the normal irrigated condition. In WT plants, theWC decreased by 30% when the soil moisture was reduced from5% to 8.1%.
.5. Higher rate of gas exchange parameters, chlorophyll contentnd photosynthetic efficiency at V1 stage
The gas exchange parameters, stomatal conductance, transpi-
ation rate and photosynthesis rate were significantly higher inransgenics when compared to the WT plants at V1 stage (Fig. 5A–C).hree events 14, 15 and 22 showed 2 times higher stomatal conduc-ance and 1.5 times higher transpiration rate than the WT plants.Fig. 5. Gas exchange parameters in EaHSP70 transgenics and WT at 8.1% soil mois-ture: (A) stomatal conductance, (B) transpiration rate and (C) photosynthesis rate. Allthe transgenic events differ significantly with respect to control Co 86032 (P ≤ 0.05;Student’s t-test). Data are presented as mean ± SD (n = 3) and error bars representSD.

S.M. Augustine et al. / Plant Science 232 (2015) 23–34 29
Fig. 6. (A) Chlorophyll content in EaHSP70 transgenic events and WT with and without stress. All the transgenic events differ significantly with respect to control Co86032 (P ≤ 0.05; Student’s t-test). (B) Photosynthetic efficiency of EaHSP70 transgenic events and WT with and without stress. Data shows a significant difference in thephotosynthetic efficiency between transgenics and untransformed control, ns: not significant at P ≤ 0.05, by Student’s t-test. Data are presented as mean ± SD (n = 5) anderror bars represent SD.
Scht
nap4ictiti(aoS2
becTpi
the visual scoring for transformants with EaHSP70 are given inTable 2 (supplementary data). Based on the physiological parame-ters, five independent events showing a higher level of tolerance todrought were selected for further gene expression (qPCR) analysis.
Table 2Bud germination (%) of EaHSP70 transgenic plants in pots supplied with 0 mM(water), 100 mM NaCl, 200 mM NaCl and 300 mM NaCl solution for 26 days. Thedata for WT plants could not be obtained in 100 mM NaCl, 200 mM NaCl and 300 mMNaCl solution because these plants failed to sustain growth in the presence of NaCl.
Event no. NaCl concentration
0 mM 100 mM 200 mM 300 mM
5 100% 100% 60% 60%6 80% 60% 40% 40%8 80% 100% 60% 20%
10 100% 100% 60% 60%13 100% 60% 40% 20%19 80% 80% 20% 20%20 100% 60% 40% 20%
ixty-eight percent of the events had 1–2 times higher stomatalonductance and transpiration rate. The 14th event had 5 timesigher photosynthesis rate than WT plants. All the events main-ained significantly higher photosynthesis rate than WT plants.
The chlorophyll content and photosynthetic efficiency were sig-ificantly higher in transgenics than WT plants at V1 stage (Fig. 6And B). On the day before withholding the irrigation, the chloro-hyll content (SPAD values) in transgenics varied between 35 and5 SPAD units, whereas in WT sugarcane it was 35 SPAD units,
ndicating that there was not much variation in the chlorophyllontent at this stage. After the induction of drought stress, out ofhe 25 transgenic events screened, 36% of the transformants exhib-ted only a maximum reduction of 3 SPAD units. Fifty-six percent ofhe transgenic plants showed a maximum reduction of 5 SPAD unitsn chlorophyll content and it varied between 30 and 40 SPAD unitsFig. 6A). However, an increased level of chlorophyll was observedfter re-irrigating for 15 days (36–42 SPAD units). On the 10th dayf stress, there was a reduction of 10 SPAD units in WT plants (25PAD unit), and after re-irrigation for 15 days the value was around7 SPAD units in these plants (Fig. 6A).
The Fv/Fm values were measured at V1 stage, which variedetween 0.55 and 0.75 at 8.1% soil moisture (Fig. 6B). The fourthvent showed a higher value (0.75) at 8.1% soil moisture. Sixty per-
ent of the events were in the range 0.6–0.7 at 8.1% soil moisture.he 13th event showed a reduced value similar to that of the WTlants at 8.1% soil moisture (0.55). After the release of stress (re-rrigation for 15 days), the values varied between 0.71 and 0.73 at
25% soil moisture, which was same as the 0th day value (0.73–0.77).The Fv/Fm for the WT plants at 8.1% soil moisture was 0.5 and afterre-irrigation, it reached 0.71, which was not much different fromthe 0th day value (0.73).
The pooled data of all the physiological parameters along with
22 100% 80% 60% 20%23 100% 80% 80% 60%24 100% 100% 100% 100%WT 100% – – –
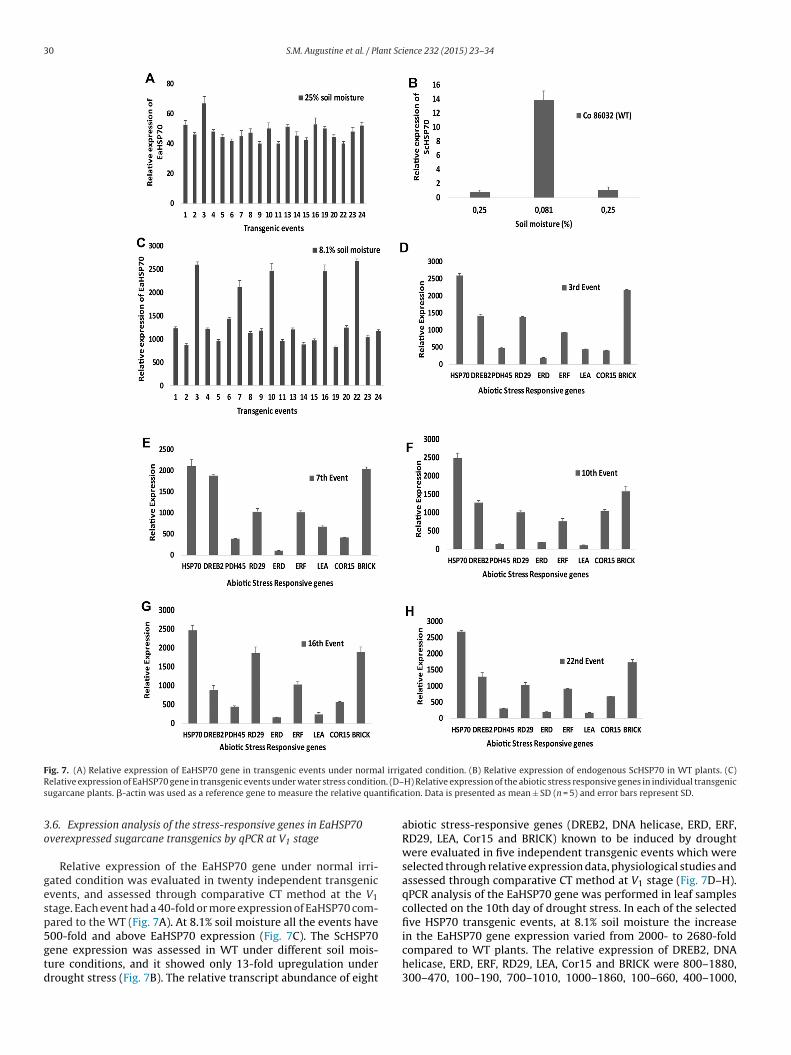
30 S.M. Augustine et al. / Plant Science 232 (2015) 23–34
F l irrigR n. (D–s ntifica
3o
gesp5gtd
ig. 7. (A) Relative expression of EaHSP70 gene in transgenic events under normaelative expression of EaHSP70 gene in transgenic events under water stress conditiougarcane plants. �-actin was used as a reference gene to measure the relative qua
.6. Expression analysis of the stress-responsive genes in EaHSP70verexpressed sugarcane transgenics by qPCR at V1 stage
Relative expression of the EaHSP70 gene under normal irri-ated condition was evaluated in twenty independent transgenicvents, and assessed through comparative CT method at the V1tage. Each event had a 40-fold or more expression of EaHSP70 com-ared to the WT (Fig. 7A). At 8.1% soil moisture all the events have
00-fold and above EaHSP70 expression (Fig. 7C). The ScHSP70ene expression was assessed in WT under different soil mois-ure conditions, and it showed only 13-fold upregulation underrought stress (Fig. 7B). The relative transcript abundance of eightated condition. (B) Relative expression of endogenous ScHSP70 in WT plants. (C)H) Relative expression of the abiotic stress responsive genes in individual transgeniction. Data is presented as mean ± SD (n = 5) and error bars represent SD.
abiotic stress-responsive genes (DREB2, DNA helicase, ERD, ERF,RD29, LEA, Cor15 and BRICK) known to be induced by droughtwere evaluated in five independent transgenic events which wereselected through relative expression data, physiological studies andassessed through comparative CT method at V1 stage (Fig. 7D–H).qPCR analysis of the EaHSP70 gene was performed in leaf samplescollected on the 10th day of drought stress. In each of the selectedfive HSP70 transgenic events, at 8.1% soil moisture the increase
in the EaHSP70 gene expression varied from 2000- to 2680-foldcompared to WT plants. The relative expression of DREB2, DNAhelicase, ERD, ERF, RD29, LEA, Cor15 and BRICK were 800–1880,300–470, 100–190, 700–1010, 1000–1860, 100–660, 400–1000,
S.M. Augustine et al. / Plant Science 232 (2015) 23–34 31
F P70 transgenics (13th event) and WT after incubation in 0 and 250 mM NaCl solution for7 ncubation in 0 and 250 mM NaCl solution for 72 h. Data shows a significant difference int , by Student’s t-test. Data is presented as mean ± SD (n = 5) and error bars represent SD.
ac
3t
td2r7fpWsTiiit1
3g
i
ig. 8. (A) Representative picture to show phenotypic differences in the leaf of EaHS2 h. (B) Chlorophyll content from leaf disks of EaHSP70 transgenics and WT after ihe chlorophyll content between transgenics and untransformed control at P ≤ 0.05
nd 1500–2100 fold upregulated in HSP70 transgenic eventsompared to WT.
.7. Overexpression of EaHSP70 in sugarcane results in toleranceo excess salinity at V1 stage
To understand the level of tolerance of the transgenic eventso salinity, leaf disk assay was carried out at the V1 stage. Leafiscs from V1 transgenics and WT plants were subjected to 0 and50 mM NaCl for 72 h. The damage caused by salinity stress waseflected in the degree of bleaching observed in the leaf tissue after2 h (Fig. 8A). Measurement of the chlorophyll content providedurther support for a positive relationship between the overex-ression of HSP70 and tolerance to excess salinity (Fig. 8B). In theT plants, there was complete bleaching of the excised leaf under
alinity stress, and the chlorophyll reduction was as high as 9 �g.he chlorosis rate was lower because the salt stress was much lowern EaHSP70-overexpressing events compared with WT plants, andt varied between events. The lowest chlorosis rate was observedn the 20th event with 0.21 �g reduction of chlorophyll (i.e. higherolerance to salinity), and the highest rate of chlorosis was in the6th event with 11.54 �g reduction of chlorophyll.
.8. EaHSP70-overexpressed sugarcane has better bud
ermination ability under excess salinity at V1 stageV1 transgenic events and WT plants were morphologically sim-lar when grown without NaCl (Fig. 9A and B). Increasing the
Fig. 9. Screening of EaHSP70 transgenic events along with the WT for bud germi-nation assay for salinity stress. Bud germination assay for salinity tolerance: (A) WTand (B) EaHSP70 transgenic events.
salt concentrations significantly affected the germination percent-age and shoot length of the transgenic events to varying degrees.Ten independent events were selected from the leaf disk assayand screened along with the WT plants. The WT plants failed to

32 S.M. Augustine et al. / Plant Sc
Table 3Shoot height (cm) of EaHSP70 transgenic plants in pots supplied with 0 mM (water),100 mM NaCl, 200 mM NaCl and 300 mM NaCl solution for 26 days.
Event no. NaCl concentration
0 mM 100 mM 200 mM 300 mM
5 4.50 ± 1.17 4.25 ± 0.64 5.0 ± 0.00 4.50 ± 0.006 5.50 ± 1.17 3.60 ± 1.15 4.3 ± 1.15 4.00 ± 0.008 6.87 ± 0.62 5.50 ± 1.73 5.6 ± 0.47 5.50 ± 0.00
10 5.90 ± 2.30 5.30 ± 0.50 4.8 ± 0.20 4.80 ± 1.1513 4.75 ± 1.06 6.00 ± 0.00 5.5 ± 1.40 5.50 ± 2.6019 5.50 ± 0.70 5.00 ± 0.00 4.0 ± 2.12 4.30 ± 1.1520 3.50 ± 0.00 4.00 ± 0.00 2.0 ± 0.00 3.75 ± 0.3522 4.90 ± 0.54 4.40 ± 0.57 4.3 ± 0.76 3.20 ± 0.2623 7.50 ± 1.73 7.20 ± 0.64 7.0 ± 1.40 6.50 ± 2.20
gbe8N82Nog
wgdcpp
4
tchodataetfuts2hsu
igTeicop[
24 6.60 ± 2.17 5.90 ± 0.00 5.5 ± 1.40 5.50 ± 0.86WT 6.26 ± 0.47 – – –
erminate in all the three salt concentrations, indicating suscepti-ility of this variety to excess salinity. In HSP70 transgenics, fourvents (5, 8, 10 and 24) had 100%, three events (19, 22 and 23) had0% and three events (6, 13 and 20) had 60% germination at 100 mMaCl. At 200 mM NaCl, one event (24) had 100%, one event (23) had0%, four events (5, 8, 10 and 22) had 60%, three events (6, 13 and0) had 40% and one event (19) had 20% germination. At 300 mMaCl, one event (24) had 100%, three events (5, 10 and 23) had 60%,ne event (6) had 40% and five events (8, 13, 19, 20 and 22) had 20%ermination, proving their tolerance to excess salinity (Table 2).
The shoot height is one of the critical growth parameters andas measured as an indicator of salinity stress tolerance in V1 trans-
enic events along with the WT plants (Table 3). The height of theifferent transgenics varied between events under different saltoncentrations. It should be noted here that similar data for WTlants grown under salinity could not be obtained because theselants failed to sustain growth in the presence of NaCl.
. Discussion
In the present study, HSP70 was isolated from a highly drought-olerant E. arundinaceus accession with a view to engineering theultivated sugarcane with this gene. Transcriptome engineeringas emerged as a promising avenue for the development of abi-tic stress-tolerant crops. The overexpression of EaHSP70 enhancedrought and salinity tolerance in sugarcane. In transgenic sug-rcane, EaHSP70 overexpressed 2000-fold and above comparedo the WT plants. The physiological and molecular traits associ-ted with drought and salinity tolerance showed a significantlynhanced expression in transgenics compared to WT plants. Inhe present study, a constitutive ubiquitin promoter (Port Ubi 2.3)rom Porteresia coarctata was used for driving the transgene. Maizebi1 promoter has been reported to overexpress under stress inransgenic rice [27,28] and potato Ubi-7 was overexpressed undertress in transgenic potato [29]. It was also reported that Port Ubi.3 has many stress responsive cis-elements [14] and our studiesave shown the induction of the GUS reporter gene under moisturetress (personal observation). This may be the reason for the drasticpregulation of the EaHSP70 gene under moisture stress.
The transgenics overexpressing EaHSP70 were found to havencreased membrane stability than WT plants under normal irri-ated conditions and even higher stability under moisture stress.ransgenic potato plants expressing DcHSP17.7 were shown toxhibit enhanced cellular membrane stability and tuberizationn vitro at elevated temperatures [30]. The overexpression of HSP70
onferred tolerance to heat stress along with improved physi-logical parameters such as water and nutrient use efficiency,hotosynthesis, assimilate partitioning and membrane stability30–32]. Such improvements make plant growth and developmentience 232 (2015) 23–34
possible under drought/heat stress. The transgenic plants main-tained significantly high RWC than WT plants during stress.Generally, osmoregulation is one of the main mechanisms preser-ving turgor pressure in most plant species against water loss [33].In [34], it was reported that better osmotic regulation or lower elas-ticity of the cell walls conferred higher RWC. To reveal the responseto stress intensity leaf RWC is one of the best growth/biochemicalparameter [35]. The plants that have high RWC under droughtstress having higher yield too. The potential of a plant to surviveacute water deficits depends on its ability to restrict water lossthrough the leaf epidermis after the stomata have attained theminimum aperture [36]. Previous studies reported that there is acorrelation between photosynthetic rate and leaf water potential,stomatal conductance and transpiration rate [37].
Photosynthetic capacity of the plant is indicated in its chloro-phyll content [38,39]. In the present study, the transgenic eventswith EaHSP70 showed significantly higher chlorophyll contentunder soil moisture stress. All the transgenic events maintainedhigh, i.e. 30 SPAD units and above, chlorophyll content at 8.1% soilmoisture stress which varied between events. During stress, thechlorophyll content stability in the plant tissues allows the cells tomaintain functional chloroplast and that helps the plant to revivethe photosynthesis after the stress. Hence, the total chlorophyllcontent estimation can help to understand the tolerance of plantsto abiotic stresses. In tomato, genotypes with increased chlorophyllcontent were observed to be tolerant to high temperatures, indicat-ing that increased chlorophyll content is related to thermotolerancein tomato [31].
The Fv/Fm ratio was significantly higher in EaHSP70 transgenicsthan in WT plants under moisture stress. Previous reports statedthat at least in the early maturing varieties there is a genotype-dependent decrease of fluorescence quantum yield (Fv/Fm) inpotato under drought, and small decreases were associated withdrought tolerance [40]. The initial fluorescence (Fo) decline reflectsa regulatory process damage external to P680 (reaction center ofPSII), such as photoprotective processes impairment that facilitatesthe dissipation of excess energy within the leaf [41]. The Fv/Fmreduction indicates either a reversible photo-protective down reg-ulation, or irreversible inactivation of PSII [42,43]. The Fv/Fm ratiowas not much reduced in EaHSP70 transgenic plants under mois-ture stress, suggesting that the function of reaction centers waslittle affected in transgenic plants relative to the control. It indi-cated a substantial protection of the photosystem, especially PSII,during drought stress in EaHSP70 transgenic plants.
In EaHSP70-overexpressed sugarcane, there was a drasticincrease (2000-fold or more) in the upregulation of HSP70 genecompared to the WT plants. In addition, the upregulation of theabiotic stress-responsive genes, i.e. DREB2, DNA helicase 45, LEA,RD29, ERD, ERF, Cor15 and BRICK, was more than 100-fold in eachtransgenic event when compared to WT plants. Possibly, this mightbe one of the reasons for its enhanced drought and salinity toler-ance. So far, there has been no report on the upregulation of DREB2while overexpressing HSP70. DREB2 is one of the main drought-responsive transcription factors determining the expression levelsof a number of downstream genes like RD29, HSP70, etc. [44–46]. Inthe present study, DREB2 was upregulated by 800-fold and morein HSP70-overexpressed transgenics. HSP70 is known to be acti-vated by DREB2, but the mechanism of upregulation of DREB2 inHSP70 transgenics needs to be studied. HSP70 acts as a molecu-lar chaperone by maintaining homeostasis of protein folding andthus helps in maintaining the metabolic and structural integrity ofthe cells [40,47]. The individual HSP70 proteins specific roles can be
determined by their location in different subcellular compartments[48,49], by the differential expression of HSP70s at different stagesof development [50] or by their interaction with specific sets ofHSP70-associated proteins. For example, the cytosolic HSP70 helps
ant Sc
taciolftppict
mCtfoesibr(siApofi(
ppCctao[tIu(s(
cptbfistr
C
A
(I
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
S.M. Augustine et al. / Pl
o prevent protein aggregation, assists folding of de novo proteinsnd organellar precursor proteins maintenance. Mitochondrial andhloroplastic HSP70 proteins are involved in precursor proteinmport and translocation. HSP70 also have a role in regulation ofther stress-associated gene expression [51]. However, the cellu-ar mechanisms of HSP70 function under stress conditions are notully understood. HSP70 also has a role in the regulation of signalransducers such as protein kinase A, protein kinase C and proteinhosphatase [52]. However, HSP70 chaperones are presumed tolay a broad role in regulating many downstream genes expression
n signal transduction pathways, under stress and normal growthonditions. The exact role of HSP70 in the modulation of signalransduction in plants has not been proved.
The upregulation of DREB2 also helps in the upregulation ofany DREB2-responsive genes including LEA, RD29, ERD, ERF and
OR15, which was demonstrated in the present study. LEA pro-eins are quite hydrophilic and are believed to protect plant cellsrom these stresses. These proteins represent another categoryf high molecular weight proteins that are abundant during latembryogenesis and accumulate during seed desiccation, watertress [53] and cold stress [54]. RD29 also encodes proteins sim-lar to LEA [55]. ERD is upregulated by dehydration stress andy natural and dark-induced senescence [56,57]. ERF (ethylene-esponsive element-binding factor) is a subfamily of the AP2/EREBPAPETALA2/ethylene responsive element-binding protein) plant-pecific transcription factor [58]. Enhanced frost tolerance andnduction of cor genes were detected in non-acclimated transgenicrabidopsis [59]. In addition, DRE elements have been found in theromoter of COR15. The BRICK gene helps in the polymerizationf actin [60], and an upregulation of the BRICK gene was identi-ed in the drought-tolerant E. arundinaceus under moisture stressunpublished data).
Overexpression of EaHSP70 in sugarcane also increased thelant’s tolerance against salinity, as reflected by the higher chloro-hyll retention in leaf disk assay and improved bud germination.hlorophyll content measurements revealed that in WT plants thehlorophyll content decreased significantly when compared to theransgenic events under salinity, suggesting the high salinity toler-nce potential of the transgenic events. In plants, the reduction ofxidative damage could provide enhanced resistance to salt stress61]. Overexpression of HSP70 is known to reduce the level of reac-ive oxygen species (ROS), thus conferring tolerance to salinity [62].n general, EaHSP70 transgenics showed higher germination ratender high salt conditions (300 mM NaCl) compared to WT plantsFig. 9A and B). This might be due to the constitutive expression ofeveral salt/osmotic stress-related genes in the transgenic eventsFig. 7C–G).
In conclusion, our results showed that transformation of sugar-ane with the EaHSP70 gene under the control of the Port Ubi 2.3romoter enhanced the tolerance of the sugarcane variety Co 86032o drought and salinity stress to a high extent. As the success of anyiotechnological strategy would ultimately be determined by thenal yield at field conditions, future studies will involve the analy-is of the performance of the sugarcane transgenic events used inhis study at realistic field conditions and under different wateringegimes.
onflicts of interest
The authors have no conflicts of interest to declare.
cknowledgement
This work was supported by the Department of BiotechnologyDBT) (grant no. BT/PR6277/AGII/106/882/2012), Government ofndia.
[
[
ience 232 (2015) 23–34 33
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.plantsci.2014.12.012.
References
[1] S.M. Augustine, D.P. Syamaladevi, M.N. Premachandran, R. Ravichan-dran, N. Subramonian, Physiological and molecular insights to droughtresponsiveness in Erianthus spp., Sugar Tech (2014), http://dx.doi.org/10.1007/s12355-014-0312-7.
[2] V.A. Amalraj, P. Rakkiyappan, D. Neelamathi, S. Chinnaraj, S. Subramanian, Wildcane as a renewable source for fuel and fibre in the paper industry, Curr. Sci.95 (11) (2008) 1599–1602.
[3] E. Vierling, The roles of heat shock proteins in plants, Annu. Rev. Plant Physiol.Plant Mol. Biol. 42 (1991) 579–620.
[4] J.D. Trent, A review of acquired thermotolerance, heat-shock proteins andmolecular chaperones in Archaea, FEMS Microbiol. Rev. 18 (1996) 249–258.
[5] W.R. Boorstein, T. Ziegelhoffer, E.A. Craig, Molecular evolution of the HSP70multigene family, J. Mol. Evol. 38 (1994) 1–17.
[6] C.G. Evans, C. Lyra, E.G. Jason, Heat shock protein 70 (Hsp70) as an emergingdrug target, J. Med. Chem. 53 (12) (2010) 4585–4602.
[7] E.K. Cho, C.B. Hong, Over-expression of tobacco NtHSP 70-1 contributes todrought stress tolerance in plants, Plant Cell Rep. 25 (2006) 349–358.
[8] E.K. Cho, Y.J. Choi, A nuclear-localized HSP70 confers thermo protective activityand drought-stress tolerance on plants, Biotechnol. Lett. 31 (2009) 597–606.
[9] K. Ono, Overexpression of DnaK from a halotolerant cyanobacterium Aphan-othece halophytica enhances the high temperature tolerance of tobacco duringgermination and early growth, Plant Sci. 160 (2001) 455–461.
10] Y. Qi, H. Wang, Y. Zou, C. Liu, Y. Liu, Y. Wang, W. Zhang, Over-expression ofmitochondrial heat shock protein 70 suppresses programmed cell death in rice,FEBS Lett. 585 (2011) 231–239.
11] A. Uchida, A. Hibino, T. Shimada, M. Saigusa, T. Takabe, E. Araki, H. Kajita,T. Takabe, Overexpression of DnaK chaperone from a halotolerant cyanobac-terium Aphanothece halophytica increases seed yield in rice and tobacco, PlantBiotechnol. 25 (2008) 141–150.
12] M. Montero-Barrientos, Transgenic expression of the Trichoderma harzianumhsp70 gene increases Arabidopsis resistance to heat and other abiotic stresses,J. Plant Physiol. 167 (2010) 659–665.
13] A. Song, X. Zhu, F. Chen, H. Gao, J. Jiang, S. Chen, A Chrysanthemum heat shockprotein confers tolerance to abiotic stress, Int. J. Mol. Sci. 15 (2014) 5063–5078.
14] A. Phillip, D.P. Syamaladevi, M. Chakravarthy, K. Gopinath, N. Subramonian, 5′
Regulatory region of ubiquitin 2 gene from Porteresia coarctata makes efficientpromoters for transgene expression in monocots and dicots, Plant Cell Rep. 32(2013) 1199–1210.
15] J. Sambrook, D.W. Russel, Molecular Cloning: A Laboratory Manual, third ed.,Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
16] S. Arvinth, S. Arun, R.K. Selvakesavan, J. Srikanth, N. Mukunthan, P. AnandaKumar, M.N. Premachandran, N. Subramonian, Genetic transformation andpyramiding of aprotinin expressing sugarcane with cry1Ab for shoot borer(Chilo infuscatellus) resistance, Plant Cell Rep. 29 (2010) 383–395.
17] M.J.E. Bettina, T.T. Melvin, A.K. Thomas, Visual assessment of wilting as a mea-sure of leaf water potential and seedling drought survival, J. Trop. Ecol. 23(2007) 497–500.
18] J.R. Martineau, J.E. Specht, J.H. Williams, C.Y. Sullivan, Temperature tolerancein soybeans. I. Evaluation of a technique for assessing cellular membrane ther-mostability, Crop Sci. 19 (1979) 75–78.
19] P. Molaei, A.E.A. Namvar, T.K. Bejandi, Water relation, solute accumulation andcell membrane injury in sesame (Sesamum indicum L.) cultivars subjected towater stress, Ann. Biol. Res. 3 (4) (2012) 1833–1838.
20] H.D. Barrs, P.E. Weatherley, A reexamination of the relative turgidity techniquefor estimating water deficits in leaves, Aust. J. Biol. Sci. 15 (1962) 413–428.
21] P. Chomczynski, K. Mackey, Modification of the TRI reagent DNA/protein isola-tion procedure for isolation of RNA from polysaccharide and proteoglycan richsources, Biotechniques 19 (1995) 942–945.
22] S. Basu, A. Roychoudhury, Expression profiling of abiotic stress inducible genesin response to multiple stresses in rice (Oryza sativa L.) varieties with con-trasting level of stress tolerance, Biol. Med. Res. Int. 2014 (2014), ID-706890.
23] M. Cheng, P. Liao, T. Lin, The Arabidopsis ethylene responsive factor 1 regu-lates abiotic stress responsive gene expression by binding to different cis-actingelements in response to different stress signals, Plant Physiol. 162 (3) (2013)1566–1582.
24] K.J. Livak, T.D. Schmittgen, Analysis of relative gene expression data using realtime quantitative PCR and the 2−�CT method, Methods 25 (2001) 402–408.
25] L. Fan, S. Zheng, X. Wang, Antisense suppression of phospholipase Da retardsabscisic acid and ethylene-promoted senescence of postharvest Arabidopsisleaves, Plant Cell 9 (1997) 2183–2196.
26] S.J. Crafts-Brandner, F.E. Below, J.E. Harper, R.H. Hageman, Effects of podremoval on metabolism and senescence of nodulating and nonnodulating Soy-bean isolines: II. Enzymes and chlorophyll, Plant Physiol. 75 (2) (1984) 318–322.
27] M.J. Cornejo, D. Luth, K.M. Blankenship, O.D. Anderson, A.E. Blechl, Activity of amaize ubiquitin promoter in transgenic rice, Plant Mol. Biol. 13 (1993) 567–581.

3 ant Sc
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
4 S.M. Augustine et al. / Pl
28] I. Takimoto, A.H. Christensen, P.H. Quail, H. Uchimiya, S. Toki, Non-systemicexpression of a stress-responsive maize polyubiquitin gene (Ubi-1) in trans-genic rice plants, Plant Mol. Biol. 26 (1994) 1007–1012.
29] J.E. Garbarino, T. Oosumi, W.R. Belknap, Isolation of a polyubiquitin promoterand its expression in transgenic potato plants, Plant Physiol. 109 (4) (1995)1371–1378.
30] Y.J. Ahn, J.L. Zimmerman, Introduction of the carrot HSP17.7 into potato(Solanum tuberosum L.) enhances cellular membrane stability and tuberizationin vitro, Plant Cell Environ. 29 (2006) 95–104.
31] D. Camejo, P. Rodrıguez, M.A. Morales, J.M. Dellamico, A. Torrecillas, J.J. Alarcon,High temperature effects on photosynthetic activity of two tomato cultivarswith different heat susceptibility, J. Plant Physiol. 162 (2005) 281–289.
32] I. Momcilovic, Z. Ristic, Expression of chloroplast protein synthesis elongationfactor, EF-Tu, in two lines of maize with contrasting tolerance to heat stressduring early stages of plant development, J. Plant Physiol. 164 (2007) 90–99.
33] D. Gunasekera, G.A. Berkowitz, Evaluation of contrasting cellular-level accli-mation responses to leaf water deficits in three wheat genotypes, Plant Sci. 86(1992) 1–12.
34] S.W. Ritchie, H.T. Nguyan, A.S. Holaday, Leaf water content and gas-exchangeparameters of two wheat genotypes differing in drought resistance, Crop Sci.30 (1990) 105–111.
35] A. Alizade, Soil, Water and Plants Relationship, third ed., Emam Reza UniversityPress, Mashhad, Iran, 2002, ISBN: 964-6582-21-4 (in Persian).
36] S. El Jaafari, Durum wheat breeding for abiotic stresses resistance: definingphysiological traits and criteria, Options Mediterr. Ser. A 40 (2000) 251–256.
37] P.J. Kramer, J.S. Boyer, Water Relations of Plants and Soils, Academic Press, SanDiego, 1995, pp. 495.
38] G.C. Wright, R. Nageswara, G.D. Farquhar, Water use efficiency and carbonisotope discrimination in peanut under water deficit conditions, Crop Sci. 34(1994) 92–97.
39] R.C. Nageswara Rao, H.S. Talwar, G.C. Wright, Rapid assessment of specific leafarea and leaf nitrogen in peanut (Arachis hypogaea L.) using a chlorophyll meter,J. Agron. Crop Sci. 186 (2001) 175–182.
40] A. Van der Mescht, J.A. de Ronde, F.T. Rossouw, Chlorophyll fluorescence andchlorophyll content as a measure of drought tolerance in potato, S. Afr. J. Sci.95 (1999) 407–412.
41] K. Angelopoulos, B. Dichio, C. Xiloyannis, Inhibition of photosynthesis in olivetrees (Olea europaea L.) during water stress and rewatering, J. Exp. Bot. 47 (1996)1093–1100.
42] N.R. Baker, J. Bowyer, Photoinhibition of Photosynthesis: From MolecularMechanisms to Field, BOIS Scientific Publishers Ltd., Oxford, UK, 1994.
43] S.P. Long, S. Humphries, P.G. Falkowski, Photoinhibition of photosynthesis innature, Annu. Rev. Plant Physiol. Plant Mol. Biol. 45 (1994) 633–662.
44] M.S. Khan, The role of DREB transcription factors in abiotic stress tolerance ofplants, Biotechnol. Biotechnol. Equip. 25 (3) (2011) 2433–2442.
45] J. Mizoi, K. Shinozaki, K. Yamaguchi-Shinozaki, AP2/ERF family transcriptionfactors in plant abiotic stress responses, Biochim. Biophys. Acta 1819 (2) (2011)86–96.
46] H. Qin, G. Qiang, Z. Junling, K. Li Sun Sundaram, Z. Yizheng, B. Mark, P. Paxton,B. Eduardo, Z. Hong, Regulated expression of an isopentenyl transferase
[
ience 232 (2015) 23–34
gene (IPT) in peanut significantly improves drought tolerance andincreases yield under field conditions, Plant Cell Physiol. 52 (11) (2011)1904–1914.
47] W. Wang, B. Vinocur, O. Shoseyov, A. Altman, Role of plant heat-shock proteinsand molecular chaperones in the abiotic stress response, Trends Plant Sci. 9(2004) 244–252.
48] J. Frydman, Folding of newly translated proteins in vivo: the role of molecularchaperones, Annu. Rev. Biochem. 70 (2001) 603–647.
49] J.A. Miernyk, The 70 kDa stress-related proteins as molecular chaperones,Trends Plant Sci. 2 (1997) 80–87.
50] D.Y. Sung, E. Vierling, C.L. Guy, Comprehensive expression profile analysis ofthe Arabidopsis hsp70 gene family, Plant Physiol. 126 (2001) 789–800.
51] J.H. Lee, F. Schoffl, An Hsp70 antisense gene affects the expression ofHSP70/HSC70, the regulation of HSF and the acquisition of thermotolerancein transgenic Arabidopsis thaliana, Mol. Gen. Genet. 252 (1996) 11–19.
52] X.Z. Ding, G.C. Tsokos, J.G. Kiang, Overexpression of HSP-70 inhibits the phos-phorylation of HSF1 by activating protein phosphatase and inhibiting proteinkinase C activity, FASEB J. 12 (1998) 451–459.
53] G.A. Galau, D.W. Hughes, Coordinate accumulation of homologous transcriptsof seven cotton LEA gene families during embryogenesis and germination, Dev.Biol. 123 (1987) 213–221.
54] A. Tunnacliffe, M.J. Wise, The continuing conundrum of the LEA proteins, Natur-wissenschaften 94 (2007) 791–812.
55] L. Dure, M. Crouch, J. Harada, T.H.D. Ho, J. Mundy, R. Quatrano, Common aminoacid sequence domains among the LEA proteins of higher plants, Plant Mol.Biol. 12 (1989) 475–486.
56] K. Nakashima, T. Kiyosue, K. Yamaguchi-Shinozaki, K. Shinozaki, A nuclear gene,erd1, encoding a chloroplast-targeted Clp protease regulatory subunit homologis not only induced by water stress but also developmentally upregulated dur-ing senescence in Arabidopsis thaliana, Plant J. 12 (1997) 851–861.
57] K. Shinozaki, K. Yamaguchi-Shinozaki, Molecular responses to dehydration andlow temperature: differences and cross talk between two stress-signaling path-ways, Curr. Opin. Plant Biol. 3 (2000) 217–223.
58] J.L. Riechmann, J. Heard, G. Martin, L. Reuber, C. Jiang, J. Keddie, L. Adam,O. Pineda, O.J. Ratcliffe, R.R. Samaha, R. Creelman, M. Pilgrim, P. Broun, J.Z.Zhang, D. Ghandehari, B.K. Sherman, G. Yu, Arabidopsis transcription factors:genome-wide comparative analysis among eukaryotes, Science 290 (5499)(2000) 2105–2110.
59] K.R. Jaglo-Ottosen, S.J. Gilmour, D.G. Zarka, O. Schabenberger, M.F. Thomashow,Arabidopsis CBF1 overexpression induces COR genes and enhances freezingtolerance, Science 280 (1998) 104–106.
60] M.J. Frank, H.N. Cartwright, L.G. Smith, Three brick genes have distinct functionsin a common pathway promoting polarized cell division and cell morphogen-esis in the maize leaf epidermis, Development 130 (2002) 753–762.
61] J. Amudha, G. Balasubramani, Recent molecular advances to combat abi-
otic stress tolerance in crop plants, Biotechnol. Mol. Biol. Rev. 6 (2) (2010)31–58.62] S. Guo, W. Wharton, P. Moseley, H. Shi, Heat shock protein 70 regulates cellularredox status by modulating glutathione related enzyme activities, Cell StressChaperones 12 (3) (2007) 245–254.




















