
Structure of the DNA Repair Helicase Hel308 Reveals DNA Binding and Autoinhibitory Domains
Upload
uni-giessenCategory
view
0download
0
1 23
Plant Cell Reports ISSN 0721-7714Volume 34Number 2 Plant Cell Rep (2015) 34:247-263DOI 10.1007/s00299-014-1704-6
Overexpression of EaDREB2 andpyramiding of EaDREB2 with the peaDNA helicase gene (PDH45) enhancedrought and salinity tolerance in sugarcane(Saccharum spp. hybrid)Sruthy Maria Augustine, J. AshwinNarayan, Divya P. Syamaladevi,C. Appunu, M. Chakravarthi,V. Ravichandran, Narendra Tuteja, et al.

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

ORIGINAL PAPER
Overexpression of EaDREB2 and pyramiding of EaDREB2with the pea DNA helicase gene (PDH45) enhance droughtand salinity tolerance in sugarcane (Saccharum spp. hybrid)
Sruthy Maria Augustine • J. Ashwin Narayan •
Divya P. Syamaladevi • C. Appunu • M. Chakravarthi •
V. Ravichandran • Narendra Tuteja • N. Subramonian
Received: 9 July 2014 / Revised: 16 October 2014 / Accepted: 5 November 2014 / Published online: 5 December 2014
� Springer-Verlag Berlin Heidelberg 2014
Abstract
Key message EaDREB2 overexpressed in sugarcane
enhanced tolerance to drought and salinity. When co-
transformed with plant DNA helicase gene, DREB2
showed greater level of salinity tolerance than in single-
gene transgenics.
Abstract Drought is one of the most challenging agri-
cultural issues limiting sustainable sugarcane production
and can potentially cause up to 50 % yield loss. DREB
proteins play a vital regulatory role in abiotic stress toler-
ance in plants. We previously reported that expression of
EaDREB2 is enhanced by drought stress in Erianthus ar-
undinaceus. In this study, we have isolated the DREB2
gene from E. arundinaceus, transformed one of the most
popular sugarcane variety Co 86032 in tropical India with
EaDREB2 through Agrobacterium-mediated transforma-
tion, pyramided the EaDREB2 gene with the gene coding
for PDH45 driven by Port Ubi 2.3 promoter through par-
ticle bombardment and evaluated the V1 transgenics for
soil deficit moisture and salinity stresses. Soil moisture
stress was imposed at the tillering phase by withholding
irrigation. Physiological, molecular and morphological
parameters were used to assess drought tolerance. Salinity
tolerance was assessed through leaf disc senescence and
bud sprout assays under salinity stress. Our results indicate
that overexpression of EaDREB2 in sugarcane enhances
drought and salinity tolerance to a greater extent than the
untransformed control plants. This is the first report of the
co-transformation of EaDREB2 and PDH45 which shows
higher salinity tolerance but lower drought tolerance than
EaDREB2 alone. The present study seems to suggest that,
for combining drought and salinity tolerance together, co-
transformation is a better approach.
Keywords Sugarcane transformation � DREB2 � PDH45 �Erianthus arundinaceus � Drought tolerance � Salinity
tolerance
Introduction
Sugarcane (Saccharum spp. hybrid) is an important com-
mercial crop in India with diverse uses. One of the major
limitations in sugarcane cultivation is the frequent occur-
rence of drought, salinity, extreme temperatures, etc.
Enhanced tolerance to multiple stresses is an important
target for improving the performance of the crop in the
field. DREB genes are the first discovered families of
transcription factors related to abiotic stress gene regula-
tion (Moran et al. 1994). DREB2 is normally induced by
Communicated by Prakash Lakshmanan.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00299-014-1704-6) contains supplementarymaterial, which is available to authorized users.
S. M. Augustine � J. Ashwin Narayan � C. Appunu �M. Chakravarthi � N. Subramonian (&)
Sugarcane Breeding Institute (ICAR), Coimbatore, Tamil Nadu,
India
e-mail: [email protected]
D. P. Syamaladevi
Indian Grass and Fodder Research Institute Regional Station,
Avikanagar, Rajasthan, India
V. Ravichandran
Department of Rice, Tamil Nadu Agricultural University,
Coimbatore, Tamil Nadu, India
N. Tuteja
International Centre for Genetic Engineering and Biotechnology,
New Delhi, India
123
Plant Cell Rep (2015) 34:247–263
DOI 10.1007/s00299-014-1704-6
Author's personal copy

water stress, high salinity and heat shock (Liu et al. 1998;
Kasuga et al. 1999; Nakashima et al. 2000; Sakuma et al.
2006) and its overexpression activates the expression of
genes possessing a CRT/DRE cis-element (Stockinger
et al. 1997; Liu et al. 1998). These genes play a crucial role
in providing tolerance to multiple stresses and display
overlapping responses to different stress conditions.
DREBs control the expression of stress-responsive genes
via the ABA-independent pathway in abiotic stress. In
Arabidopsis, overexpression of DREB2 driven by a con-
stitutive promoter improved tolerance to drought, salinity
and cold without growth retardation (Liu et al. 1998).
Recently, we reported a twofold increase in the relative
expression of DREB2 under water stress in Erianthus ar-
undinaceus compared to the moderately drought-tolerant
sugarcane cultivar Co 86032 (Augustine et al. 2014). Reis
et al. (2014) have recently reported the overexpression of
AtDREB2A in sugarcane imparting drought tolerance, and
the current paper is perhaps the first report on the salinity
tolerance of DREB2 in sugarcane.
Genetic engineering of plants for enhanced drought
tolerance is mostly based on the manipulation of both
transcription and signaling factors or genes that directly
protect the cells against water deficit (Valliyodan et al.
2006). The highly conserved domains in DREB2 proteins
are important for their specific biological functions, and
identifying such critical domains will help in achieving
efficient crop improvement strategies through genetic
engineering. The plants overexpressing DREBs are repor-
ted to impart tolerance to drought, salinity and/or cold in
combination with different promoters (Latha and Prasad
2011). DREB2 is the major transcription factor that binds
to the cis-acting elements of most of the osmotic-stress-
inducible genes responsible for osmotolerance to the plants
under stress conditions (Hussain et al. 2011). Transgenic
tobacco plants overexpressing PgDREB2 (Pennisetum
glaucum) showed enhanced tolerance to both hyperionic
and hyperosmotic stresses (Kasuga et al. 2004). Overex-
pression of PeDREB2 (Populus euphratica) in Arabidopsis
enhanced drought and salinity tolerance (Chen et al. 2009).
Similarly, overexpression of the PgDREB2 and GmDREB2
(Glycine max) separately in tobacco enhanced salt and
osmotic tolerance (Chen et al. 2007; Agarwal et al. 2010).
Rice plants transformed with SbDREB2 (Sorghum bicolor)
improved yield and drought tolerance under water limita-
tion (Bihani et al. 2011).
Helicases are a ubiquitous group of motor proteins
essential for all living organisms. The pea DNA helicase 45
(PDH45), the first plant nuclear DNA helicase, was over-
expressed and purified from a bacterial system (Pham et al.
2000). Studies have shown that PDH45 is upregulated in
pea seedlings under high salt (200 mM of NaCl), and
overexpression of the gene conferred salinity tolerance in
tobacco (Sanan Mishra et al. 2005). The PDH45 gene is
also upregulated by other abiotic stresses including dehy-
dration, wounding and low temperature (Sanan Mishra
et al. 2005; Manjulatha et al. 2014).
To restore cellular functions and induce stress tolerance
in plants, transfer of a single gene encoding a specific stress
protein may not be sufficient to achieve the required tol-
erance levels (Bohnert et al. 1995). This constraint can be
overcome by the two promising approaches, namely
overexpression of a stress inducible transcription factor
that regulates a number of other genes, and pyramiding of
one or more genes (Yamaguchi-Shinozaki and Shinozaki
1994; Chinnusamy et al. 2005). Hence, with a view to
assessing the expression of the abiotic stress tolerance in
the single-gene and double-gene transforments, we have
isolated the gene coding for the transcription factor DREB2
from E. arundinaceus (IK 76-81), transformed the sugar-
cane variety Co 86032 with DREB2 alone and pyramided it
with PDH45 gene and evaluated the transgenics for drought
and salinity tolerance through different morphological,
physiological and molecular parameters.
Materials and methods
Vector construction and generation of transgenics
The DREB2 gene was isolated from E. arundinaceus (IK
76-81) using gene-specific primers designed from Sorghum
bicolor (NCBI accession no. EU500654) and Zea mays
(NCBI accession no. NM 001158997) through polymerase
chain reaction (PCR) (1 cycle at 94 �C for 5 min, followed
by 35 cycles of 94 �C for 45 s, 61.5 �C for 30 s and 72 �C
for 2 min). The pCAMBIA1305.1 vector was restricted
with BamHI and NcoI restriction enzymes to release the
CaMV 35S promoter driving the GUS gene and to insert
Port Ubi 2.3 promoter (Phillip et al. 2013). Port Ubi 2.3
promoter was amplified from pPORT-Ubi-GFP (Phillip
et al. 2013) using two specific primers: forward, 50-GAT
CGGATCCACTATCACCCTCGAGGTG-30 (BamHI site
underlined) and reverse, 50-GATCCCATGGCTGCAAGA
AATAATCACCAA-30 (NcoI site underlined). The newly
constructed plasmid was named pPORT-UBI2.3-GUS.
After sequencing and confirmation, the full-length
sequence of EaDREB2 was amplified with two specific
primers: forward, 50-GATTACTAGTATGGAGCTGGGA
GACGC-30 (SpeI site underlined) and reverse, 50-GAT
TGCTAGCCTAATATGCAAAAAGGCTAAACCCA-30
(NheI site underlined), and cloned into pPORT-UBI2.3-
GUS in place of the GUS gene. The resultant construct
pSBI-DREB2 (Fig. 1a) was transferred to Agrobacterium
tumefaciens LBA4404 by the freeze–thaw method (Sam-
brook and Russel 1989). For Agrobacterium-mediated
248 Plant Cell Rep (2015) 34:247–263
123
Author's personal copy

transformation in sugarcane variety Co 86032, the method
described by Arvinth et al. (2010) was followed, and
transgenic plants were selected on hygromycin (50 mg/l)
selection medium.
The full-length PDH45 gene was amplified from
pRT101 vector (Sanan Mishra et al. 2005) obtained from
Dr. Narendra Tuteja, ICGEB, New Delhi, India, using two
specific primers: forward, 50-GATTCCATGGATGGCGA
CAACTTCTGTGG-30 (NcoI site underlined) and reverse,
50-GGCCCGCTAGCTTATATAAGATCACCAATATTC-
30 (NheI site underlined), and cloned into pPORT-UBI2.3-
GUS in place of GUS and the resultant construct was
named pSBI-PDH45 (Fig. 1b). pSBI-PDH45 was co-
transferred with pSBI-DREB2 to sugarcane variety Co
Fig. 1 Schematic diagram of
the plant expression vectors
a pSBI-DREB2 and b pSBI-
PDH45
Plant Cell Rep (2015) 34:247–263 249
123
Author's personal copy

86032 through particle bombardment using the method
described by Bower and Birch (1992). The transgenic
plants were selected on hygromycin (50 mg/l) selection
medium.
Molecular analyses and selection of transgenic events
Standard PCR technique was used to detect the presence of
the transgene in regenerated putative transgenic sugarcane
plantlets. The promoter-gene fusion primer sequences
50-GTAGCCTAGTTCTTGCTTGGCAT-30 and 50-GTG
TCGGAAACTACTCCTAGCC-30 for EaDREB2 and 50-GCATGTGTGAATGGTGCGATTTG-30 and 50-AGGAG
TTCCAGACACCACGTGAAC-30 for PDH45 were used
for PCR reactions. Plants with the expected size amplicon
were selected for further experiments. Further confirmation
was done with the marker gene (Hpt) using primers 50-GATCTCCAATCTGCGGGATC-30 and 50-ACTCACCG
CGACGTCTGTCG-30 to generate the 800 bp product.
Plant material and stress treatments
Twenty-five independent EaDREB2 transgenic events and
22 co-transformed independent transgenic events (EaD-
REB2 and PDH45) of sugarcane along with the wild-type
(WT-untransformed control sugarcane variety Co 86032)
plants were vegetatively multiplied from V0 plants, thus
taking the events to V1 stage. Five biological replicates for
each event along with the WT plants were planted in 16’’
pots (containing soil, sand and farmyard manure in 1:1:1
ratio) and maintained in a greenhouse for moisture-stress
screening. The pots were arranged in the greenhouse in a
completely randomized design and irrigated daily for
119 days.
Water stress was imposed on the plants at the tillering
phase (120 days after planting) by withholding irrigation
for 10 days and was released on the 11th day, and normal
irrigation was continued. Relative water content and cell
membrane thermostability tests were carried out on the
0th and 10th day of drought induction. Gene expression
and gas-exchange parameters were recorded on the
10th day after drought induction. Chlorophyll content and
photosynthetic efficiency (Fv/Fm) were recorded on the
0th and 10th day after withholding the irrigation and on
15th day after the release of drought (re-irrigation). Soil
samples were collected from five randomly selected pots
at 10, 20 and 30 cm depth using an auger, and moisture
was estimated by gravimetric method using a moisture
analyzer (A&D model Mx-50). Salinity tolerance was
assessed through leaf disc and bud germination assays.
All the observations were recorded from five biological
and three technical replications of both transgenics and
WT.
Visual scoring
The transgenic events along with the WT were visually
observed for wilting of leaves on the 5th and 10th day after
drought induction. Leaf wilting was scored on a scale of
1–4 modified from Engelbrecht et al. (2007) as: 1, no
wilting; 2, slightly wilting; 3, wilting, wherein the plant
showed leaf wilting only during hot hours from which the
leaves recovered; and 4, severe wilting, wherein wilted
leaves did not recover. The mean score was computed from
five plants from each event.
Cell-membrane thermostability
Cell-membrane injury is regarded as an indicatior of the
ability of the plant to tolerate drought. To estimate the cell-
membrane injury percentage, the cell-membrane thermo-
stability test was carried out in both V0 and V1 generations
following the method described by Martineau et al. (1979).
For V0 generation, third fully opened leaves of 120-day-old
plants grown under normal irrigated conditions were used,
and for V1 generation leaves were collected on 0th and
10th day after drought induction. Leaf discs (0.5 cm
diameter) were made up to 200 mg, washed thrice with
distilled water and finally 20 ml of distilled water added to
each control and treatment tubes (2.5 cm 9 15 cm). The
tubes were covered with aluminium foil, incubated at
60 �C in a thermostatically controlled water bath for
20 min and placed at 10 �C for 12 h to allow the diffusion
of electrolytes into the water. After recording the initial
conductance at 30 �C, the tubes were heated at 100 �C for
20 min and final conductance was recorded after cooling.
Membrane Injury percentage was computed using the fol-
lowing formula:
Membrane injury % ¼ 1� 1� T1=T2ð Þ = 1�C1=C2ð Þ½ �� 100
where T and C refer to the values for treatment and control
samples, and the subscripts 1 and 2 denote the initial and
final conductance, respectively.
Leaf water status or relative water content (RWC)
Fully opened third leaves in the whorl from 120-day-old
transgenics and WT plants were subjected to measurement
of leaf relative water content (RWC) on the 0th and
10th day after drought induction. Leaf RWC was calcu-
lated based on fresh (FW), turgid (TW) and dry weights of
0.2 g leaf samples. After recording the fresh weights of
excised leaves, sample leaf discs were soaked in de-ionized
water for 4 h at room temperature in a closed petri dish,
blot-dried and weighed for TW. Samples were then dried at
90 �C for 72 h and the dry weight was recorded. RWC was
250 Plant Cell Rep (2015) 34:247–263
123
Author's personal copy

computed using the formula of Barrs and Weatherley
(1962): RWC ¼ FW�DWð Þ= TW�DWð Þ½ � � 100:
Gas-exchange parameters
Gas-exchange parameters, i.e., photosynthesis rate (A),
stomatal conductance (gs) and transpiration rate (E), were
recorded from the middle part of fully opened third leaf on
the 10th day of drought induction (soil moisture & 8.1 %)
using a portable photosynthesis system (Li-6400, Li-COR
Inc., Lincoln, Nebraska, USA) with a leaf chamber of
2 cm 9 3 cm and an integrated light source (LI-6400-
02B). Gas-exchange parameters were measured at
130 lmol m-2 s-1 [CO2], 40 �C (closed cabinet incubated
to 40 �C) and RH *60–70 %. The external CO2 concen-
tration in air was maintained at 380 lmol mol-1 in the
reference cuvette. All the gas-exchange parameters were
measured at a leaf temperature of 32 ± 2.0 �C at 30 s
intervals over a period of 2 h from 9.00 a.m. to 11.00 a.m.
Chlorophyll measurements
Total chlorophyll content was recorded on the 0th and
10th day after drought induction (soil moisture 25 and
8.1 %, respectively), and 15 days after re-irrigation (25 %
soil moisture) using a chlorophyll meter (SPAD-502, Ko-
nica Minolta, Japan).
Photosynthetic efficiency (Fv/Fm)
The maximum potential photochemical efficiency, defined
as the ratio of variable to maximum fluorescence emitted
by chlorophyll (Fv/Fm), was estimated using a portable
OS-30P chlorophyll fluorimeter (Optisciences, USA).
Photosynthetic efficiency (Fv/Fm) was recorded on the 0th
and 10th day after drought induction and 15 days after the
release of drought. The plants were dark-adapted for
20 min prior to measurement. Maximal fluorescence under
light exposure (Fm0) was obtained by imposing a 1 s sat-
urating flash to the leaf to reduce the entire PS II reaction
centre after attaining steady-state fluorescence (Ft). The
minimal fluorescence immediately after light exposure
(Fo0) was determined by imposing dark while a far red light
was simultaneously switched on to oxidize PS II rapidly by
drawing electrons from PS II to PS I.
Gene expression analysis using quantitative real-time
polymerase chain reaction (qPCR)
To elucidate the role of transgenes, the relative expression
of six abiotic stress (salt, cold and heat) responsive genes,
i.e. RD29 (response to dehydration), LEA (late embryo-
genesis abundant protein), ERF (ethylene responsive
factor), COR15 (cold regulated protein), ERD (early
responsive to dehydration) and BRICK/HSPC300, was
assessed along with the transgenes. Five transgenic events,
each selected from the EaDREB2 and co-transformed
events based on the physiological parameters, were used
for gene expression analysis. Total RNA was isolated from
the third fully opened leaf sample using Trizol reagent
(Sigma Chemicals, USA) following the method of Chom-
czynski and Mackey (1995). First-strand cDNAs were
synthesized from total RNA using the Fermentas first-
strand cDNA synthesis kit and oligo (dT) primers (Fer-
mentas International Inc, Ontario, Canada) following the
manufacturer’s instructions. The actin transcript was used
as an internal control to quantify the relative transcript
levels. To make sure that actin gene did not respond to
drought stress, the expression of the gene was determined
in both irrigated and drought-stressed samples. cDNA
fragments and actin gene were amplified with gene-specific
Table 1 Visual scoring of pSBI-DREB2 and co-transformed trans-
genic events for soil moisture stress
Event no. Score after stress induction
DREB2 transgenics Co-transformed events
5th day 10th day 5th day 10th day
1 1 3 1 3
2 1 2 1 2
3 1 1 1 1
4 1 1 1 2
5 2 3 2 2
6 2 1 1 2
7 1 1 1 3
8 1 1 1 1
9 1 2 1 3
10 1 4 1 2
11 2 2 1 3
12 1 2 1 3
13 1 2 1 3
14 1 1 2 2
15 1 3 1 3
16 1 2 2 2
17 1 1 1 1
18 2 2 2 1
19 2 2 2 1
20 1 3 1 1
21 1 2 1 1
22 1 2 1 1
23 1 3 – –
24 1 1 – –
25 1 4 – –
WT 3 4 3 4
Plant Cell Rep (2015) 34:247–263 251
123
Author's personal copy

primers (Table 1, supplementary data). All PCRs were
performed for 40 cycles (1 cycle at 95 �C for 10 min fol-
lowed by 40 cycles of 95 �C for 15 s and 60 �C for 1 min).
qPCR was performed using MESAGREEN master mix and
StepOne real-time PCR system (Applied Biosystems,
Burlington, ON, Canada). The CT values for both the target
and internal control genes were used for the quantification
of transcripts by comparative CT method normalization.
Later, the products were analysed through a melt-curve
analysis to check the specificity of PCR amplification. Each
reaction was performed three times, and the expression of
target gene was calculated using the formula (Livak and
Schmittgen 2001): 2-DDCt, where DDCt = DCt sample-
DCt actin. DDCt values reflect the relative expression of
the target gene upon exposure to drought stress.
Leaf disc senescence assay for measuring salinity
tolerance
Leaf disc assay was carried out to evaluate the sensitivity
of the transformed and WT plants to sodium chloride
(NaCl) stress as described by Fan et al. (1997). Two grams
of fresh leaf sample (3.0 cm 9 2.0 cm) was excised from
healthy and fully expanded third leaves of 120-day-old
plants. The discs were floated in a 10 ml solution of
250 mM NaCl and experimental control (0 mM NaCl) for
72 h and then used for measuring chlorophyll content
spectrophotometrically after extraction in methanol
(Crafts-Brandner et al. 1984). The damage caused by stress
was reflected in the degree of bleaching observed in the
leaf tissue after 72 h. The treatment was carried out in
continuous white light at 25 ± 2 �C.
Bud germination assay
Ten independent transgenic events were selected for bud
germination assay based on the leaf disc senescence assay.
Single bud setts were planted in cavity trays and grown at
30 �C temperature and 60 % relative humidity. To evaluate
germination under salt stress, single bud setts were irri-
gated daily with 25 ml of 0 mM, 100 mM (5.844 g/l),
200 mM (11.688 g/l) and 300 mM NaCl (17.532 g/l)
solutions for a period of 26 days. The number of buds
germinated and their shoots height were recorded on the
26th day of planting.
Statistical analysis
For statistical analysis of the data, five biological and three
technical replicates from each of the transgenic events were
used. Student’s t test (P B 0.05) was conducted using
XLSTAT 2013.5 to analyse the data under normal and
stressed conditions also to compare the EaDREB2 trans-
genics with co-transformed events.
Results
Isolation of DREB2 gene from E. arundinaceus
and generation of transgenics
The DREB2 gene (1.5 Kb) was isolated from E. arundin-
aceus and the sequence was submitted to Gen Bank
(accession no.KJ670161) with the name EaDREB2. The
isolated DREB2 gene featured a 702-bp-long intron flanked
by a 70-bp-long exon sequence at the 50 end and a 722-bp-
long exon in the 30 end. The isolated gene sequence had
high sequence homology (98–100 %) with the reported
DREB2 gene sequences of Sorghum bicolor and Zea mays.
The translated amino acid sequence showed high homol-
ogy (95–98 %) with other reported DREB2 protein
sequences.
The hygromycin-resistant sugarcane transformants
were confirmed through PCR using primers of promoter-
gene fusion and Hpt marker gene. In promoter-gene
fusion primer screening, the expected amplicon size of
1 kb for DREB2 and 1.2 kb for PDH45 were obtained.
Twenty-five events in EaDREB2 and 22 events in co-
transformed events were positive for the transgene inte-
gration. The morphological and growth characteristics of
transgenic plants were similar to those of the WT plants
(Fig. 2a, c).
Cell-membrane thermostability in V0 transgenics
The standard test for cell-membrane thermostability was
performed in the V0 EaDREB2 and co-transformed trans-
genic events (Fig. 3a). Significantly lower membrane
injury, which indicates higher cell-membrane stability, was
observed in all the EaDREB2 transgenic events
(43–85.03 %) than in WT. As against 95.5 % injury in WT,
56 % of transgenic events (14 out of 25) showed injury
percentage of 50–60 % with the lowest (43.24 %) in the
eighth event and the highest (85.03 %) in the tenth event
(Fig. 3a). In co-transformed events, too, membrane ther-
mostability was significantly higher with a range of
42.37–95.45 %. Sixty-three percent (14 out of 22) of the
events had membrane injury of 50–75 % with the lowest
injury (42.37 %) in the 11th event. The lowest stability was
in the 15th event with 95.45 % membrane injury (Fig. 3b).
There was a significant difference in cell-membrane ther-
mostability between EaDREB2 transgenics and co-trans-
formed events.
252 Plant Cell Rep (2015) 34:247–263
123
Author's personal copy

Morphological and physiological drought stress
characteristics in V1 plants
The wilting symptoms in each of the transgenics on 5th and
10th day (Fig. 2b, d) were recorded (Table 1). In EaD-
REB2 transgenics, 32 % of plants (8 out of 25) and in the
co-transformed events 36 % of plants (8 out of 22) were
almost normal, showing very little wilting which was rated
as 1, whereas in WT all the plants showed the extreme
wilting, which was scored as four.
There was a significant increase in the membrane sta-
bility of V1 EaDREB2 transgenics with decreasing mois-
ture levels (Fig. 3c), though it varied between events. The
membrane injury varied between 43.07 and 82.58 % at
25 % soil moisture (i.e. with normal irrigation). At 8.1 %
soil moisture, the injury percentage ranged between 26.35
Fig. 2 Screening of transgenic
events for soil moisture stress
and bud germination assay for
salinity stress. a EaDREB2 and
WT at 25 % soil moisture,
b EaDREB2 and WT at 8.1 %
soil moisture, c pSBI-PDH45
and WT at 25 % soil moisture,
d pSBI-PDH45 and WT at
8.1 % soil moisture along with
the WT for soil moisture stress.
Bud germination assay for
e WT, f pSBI-DREB2 and
g pSBI-PDH45
Plant Cell Rep (2015) 34:247–263 253
123
Author's personal copy

and 57.14 %. Out of 25 transgenic events screened, 48 %
of the events (12 out of 25) showed lower levels of
membrane injury, i.e., 50–60 % at 25 % soil moisture,
whereas in 68 % of the events (17 out of 25) membrane
injury was between 30 and 50 % at 8.1 % soil moisture,
suggesting enhanced cell-membrane stability under stress
in the transgenic events with the lowest injury percentage
of 26.35 % in the 13th event. A significant increase in the
membrane stability was also observed in co-transformed
events under soil moisture stress. In the co-transformed
events, the membrane injury varied between 39.46 and
95.7 % at 25 % soil moisture (Fig. 3d) and between 24.8
and 84.06 % at 8.1 % soil moisture. Out of the 22 inde-
pendent transgenic events screened, 68 % of the events (15
out of 22) showed reduced levels of membrane injury
(50–80 %) at 25 % soil moisture and 77 % of the events
(17 out of 22) showed 30–50 % injury at 8.1 % soil
moisture. The lowest injury percentage (24.89 %) at 8.1 %
was in the eighth event. At 25 % soil moisture, 5 % of the
transgenic events had the cell-membrane injury close to
that of the WT plants. Cell-membrane injury showed a
decreasing trend with the increasing moisture stress in all
the events studied. However, in WT plants, the membrane
injury increased from 93.2 to 98.71 % when soil moisture
decreased from 25 to 8.1 %. There was no significant
difference in the membrane stability of EaDREB2 trans-
genics and co-transformed events under soil moisture
stress.
Higher relative water content in both EaDREB2 and co-
transformed transgenic events at V1 stage
In EaDREB2 transgenic events, a significantly higher
RWC was observed under moisture stress (Fig. 4a). Four
events exhibited a maximum of 3 % reduction in RWC at
8.1 % soil moisture from the normal irrigated condition.
Fifty-two percent (13 out of 25) of the transgenic events
had maximum of 10 % reduction in RWC, with three
events showing reduced RWC similar to that of the WT,
i.e. 15–30 % reduction from the normal irrigated condition.
In WT, the RWC decreased by 30 % when the soil mois-
ture was reduced from 25 to 8.1 %. In the co-transformed
events, out of 22 independent events screened, 50 % of the
transgenic events (11 out of 22) had a maximum of 10 %
reduction and five events showed a maximum 20 %
reduction in RWC (Fig. 4b). Three events (6, 8, and 9)
exhibited reduced RWC almost equal to that of the WT, i.e.
25–30 % reduction. There was no significant difference in
the RWC between EaDREB2 and co-transformed events
under soil moisture stress condition.
Fig. 3 Cell-membrane thermostability in EaDREB2 transgenics and
WT a V0 stage at 25 % soil moisture, b V1 stage at 8.1 and 25 % soil
moisture stress. All the transgenic events differ significantly from the
WT at P B 0.05, by Student’s t test. Cell-membrane thermostability
in co-transformed events and WT, c V0 stage at 25 % soil moisture
and d V1 stage at 8.1 and 25 % soil moisture stress. Data show a
significant difference in the CMS between transgenic events and WT,
ns- not significant; at P B 0.05, by Student’s t test. Data are presented
as mean ± SD (n = 5) and error bars represent SD
254 Plant Cell Rep (2015) 34:247–263
123
Author's personal copy

Maintaining higher gas-exchange parameters,
chlorophyll content and photosynthetic efficiency
In EaDREB2 transgenic events, the gas-exchange param-
eters, such as stomatal conductance, transpiration rate and
photosynthesis rate, were significantly higher under soil
moisture stress (Fig. 5a–c). The eighth event showed a
10-fold increase in stomatal conductance and transpiration
rate. Sixty-eight percent of the events (17 out of 25) had
1–3 times higher stomatal conductance and transpiration
rate. All the events maintained 2–3 times higher photo-
synthetic rate than WT. In co-transformed events, out of
the 22 events, 18 were screened for stomatal conductance,
transpiration rate and photosynthetic rate at 8.1 % soil
moisture (Fig. 5d–f). Fifty-five percent of the events (10
out of 18) had higher stomatal conductance and transpira-
tion rate, which was two times higher than WT, and all the
other events had similar or lower stomatal conductance and
transpiration rate as that of WT. The highest photosynthetic
rate was in the sixth event, i.e. 11 % more than the WT,
and all the other events except the fifth event had higher
(2–3 times) photosynthetic rate than WT. The stomatal
conductance and transpiration rate showed a significant
difference between EaDREB2 and co-transformed events
but not for photosynthetic rate.
Both EaDREB2 and the co-transformed events had
significantly higher chlorophyll content (Fig. 6a, b). At the
beginning of the induction of stress, the chlorophyll content
(SPAD values) in transgenics varied between 35 and 45
SPAD units, whereas in WT it was around 35 SPAD units,
indicating not much variation under normal irrigated con-
dition. In nine EaDREB2 transgenic events, the maximum
of three SPAD units reduction was observed after the
induction of drought stress, indicating maintenance of a
higher level of chlorophyll content. Fifty-six percent of the
transgenic events (14 out of 25) had maximum of 5 SPAD
units reductions in chlorophyll content under stress, and it
varied between 30 and 40 SPAD units (Fig. 6a). In addi-
tion, an increased level of chlorophyll content (36–42
SPAD units) was observed after re-irrigation. In co-trans-
formed events, 77 % of the transgenics (17 out of 22) had a
maximum of seven SPAD units reduction in chlorophyll
Fig. 4 Relative water content
in transgenics and WT with and
without soil moisture stress
a pSBI-DREB2, b co-
transformed events. Data
followed by an (asterisk) show a
significant difference with
respect to control Co 86032
(P B 0.05; Student’s t test).
Data are presented as
mean ± SD (n = 5) and error
bars represent SD
Plant Cell Rep (2015) 34:247–263 255
123
Author's personal copy

content, and it varied between 26 and 38 SPAD units under
moisture stress, whereas in WT, 10 SPAD units reduction
(25 SPAD units) was observed on the 10th day of stress.
After re-irrigation for 15 days, the SPAD value was around
27 SPAD units in these plants (Fig. 6a, b). The chlorophyll
content (33–41 SPAD units) in all the transgenic events
showed a drastic increase after re-irrigation (Fig. 6b).
There was a significant difference in chlorophyll content
between the EaDREB2 and co-transformed events under
soil moisture stress.
The photosynthetic quantum efficiency was significantly
higher in both DREB2 and co-transformed events com-
pared to the WT (Fig. 6c, d). In EaDREB2 transgenics, the
Fv/Fm values varied between 0.62 and 0.76 under stress
(Fig. 6c). Three events (4, 8 and 14) recorded a maximum
Fv/Fm value of 0.76, and 52 % the events (13 out of 25)
showed a value of 0.60–0.70. After the release of stress, it
varied between 0.73 and 0.77, which was same as the 0th
day value. In co-transformed events, 22 transgenic events
were screened, and the Fv/Fm values varied between 0.59
Fig. 5 Gas exchange parameters in transgenics and WT at 8.1 % soil
moisture. In EaDREB2 transgenics a stomatal conductance, b tran-
spiration rate and c photosynthesis rate. In co-transformed events,
d stomatal conductance, e transpiration rate and f photosynthesis rate.
All the transgenic events differ significantly with respect to control
Co 86032 (P B 0.05; Student’s t test). Data are presented as
mean ± SD (n = 3) and error bars represent SD
256 Plant Cell Rep (2015) 34:247–263
123
Author's personal copy

and 0.70 (Fig. 6d). In 81 % of the events (18 out of 22), it
varied between 0.60 and 0.70, and after the release of stress
the values increased to 0.72–0.76. Under moisture stress,
there was reduction (0.50) in WT, which increased to 0.73
after release of stress. In general, the transgenic events had
better photosynthetic efficiency than WT. There was a
significant difference in the Fv/Fm values between EaD-
REB2 and co-transformed events under soil moisture
stress.
Fig. 6 Chlorophyll content and photosynthetic efficiency in trans-
genics and WT with and without stress. a Chlorophyll content in
EaDREB2 transgenics along with WT, b chlorophyll content in co-
transformed events along with WT, c photosynthetic efficiency in
EaDREB2 transgenics along with WT, d photosynthetic efficiency in
co-transformed events along with WT. Data show a significant
difference in the photosynthetic efficiency between transgenics and
untransformed control, ns not significant at P B 0.05, by Student’s
t test. Data are presented as mean ± SD (n = 5) and error bars
represent standard deviation
Table 2 Bud germination percentage of pSBI-DREB2 transgenic
plants in pots supplied with 0 mM (water), 100 mM, 200 mM and
300 mM NaCl solution for 26 days
Event no. 0 mM (%) 100 mM (%) 200 mM (%) 300 mM (%)
1 100 80 80 60
2 80 100 60 20
4 100 100 60 60
8 100 80 80 40
14 100 100 80 20
15 80 80 0 0
18 100 60 40 0
20 100 60 20 20
22 100 80 80 80
24 100 60 20 20
WT 100 – – –
Table 3 Bud germination percentage of sugarcane co-transformed
with pSBI-DREB2 and pSBI-PDH45 in pots supplied with 0 mM
(water), 100 mM, 200 mM and 300 mM NaCl solution for 26 days
Event no. 0 mM (%) 100 mM (%) 200 mM (%) 300 mM (%)
1 100 40 0 0
2 100 100 80 60
4 100 80 60 0
6 100 80 0 0
9 80 20 20 20
11 100 100 80 60
13 80 40 40 40
15 100 80 80 80
16 100 20 20 20
18 80 40 0 0
WT 100 – – –
Plant Cell Rep (2015) 34:247–263 257
123
Author's personal copy

The pooled data of all the physiological parameters,
together with the visual scoring of both EaDREB2 and co-
transformants along with WT, are given in Tables 2 and 3
(supplementary data).
Expression analysis of stress-inducible genes using
quantitative real-time PCR
In EaDREB2 events, the transgene was 2300-fold upreg-
ulated compared to WT under moisture stress. The relative
expression of HSP70, DNA helicase 45, ERD, RD29, LEA,
ERF, COR15 and BRICK were also upregulated 562-, 57-,
477-, 1075-, 1187-, 117-, 2036- and 1751-fold, respec-
tively, in the transgenic events (Fig. 7a). In the co-trans-
formed events, the EaDREB2 and DNA helicase 45 were
1683- and 526-fold upregulated, respectively, under
moisture stress. The relative expression of HSP70, ERD,
RD29, LEA, ERF, COR15 and BRICK were increased
590-, 226-, 1416-, 1203-, 399-, 325- and 166-fold,
respectively, in the transgenic events (Fig. 7b).
Overexpression of EaDREB2 and EaDREB2-PDH45
events results in tolerance to excess salinity
To understand tolerance level of the transgenic events to
salinity, leaf disc senescence assay was carried out. The
chlorophyll content in the EaDREB2 and co-transformed
events were significantly higher than in WT plants, indi-
cating tolerance to salinity (Fig. 8b, d). Measurement of
the chlorophyll content provided further support for a
positive relationship between the overexpression of the
transgenes and tolerance to excess salinity. In WT, there
was complete bleaching of the excised leaf under stress,
and the chlorophyll reduction was as high as 9 lg.
Salinity-induced chlorosis was less in EaDREB2 over-
expressing events (Fig. 8a). The minimum chlorosis was
observed in the 14th event with 0.03 lg reduction of
chlorophyll, while the maximum chlorosis was in the 16th
event with 6.48 lg reduction of chlorophyll (Fig. 8b). The
chlorophyll loss was significantly lower in co-transformed
DREB2-PDH45 overexpressing events than in WT
(Fig. 8c). The minimum chlorosis was observed in the
fourth event with 0.31 lg reduction of chlorophyll and
the maximum was in the 20th event with 7.75 lg
reduction (Fig. 8d). There was a significant difference in
the chlorophyll content between EaDREB2 and co-trans-
formed events.
Transgenic events had better bud germination
than the WT under salinity stress
Transgenic events and WT were morphologically similar
when grown without NaCl (Fig. 2e–g). Increasing the salt
concentrations significantly affected the germination per-
centage and shoot length of the transgenic events to vary-
ing degrees. Ten EaDREB2 independent events (1, 2, 4, 8,
14, 15, 18, 20, 22 and 24) were selected from the leaf disc
senescence assay and screened along with the WT. The
WT failed to germinate in all the three salt concentrations,
indicating susceptibility of this variety to excess salinity.
Three events (2, 4 and 14) had 100 %, four events (1, 8, 15
and 22) had 80 % and three events (18, 20 and 24) had
60 % germination in 100 mM NaCl. At 200 mM NaCl,
four events (1, 8, 14 and 22) had 80 %, two events (2 and
4) had 60 %, one event (18) had 40 % and two events (20
and 24) had 20 % germination. At 300 mM NaCl, one
event (22) had 80 %, two events (1 and 4) had 60 %, one
event (8) had 40 % and four events (2, 14, 20 and 24) had
20 % germination, suggesting its enhanced tolerance to
excess salinity (Fig. 2f; Table 2). In ten selected co-trans-
formed transgenic events (1, 2, 4, 6,9, 11, 13, 15, 16 and
18), two events (2 and 11) had 100 %, three events (4, 6
and 15) had 80 %, three events (1, 13 and 18) had 40 %
and two events (9 and 16) had 20 % germination in
100 mM NaCl. At 200 mM NaCl, three events (2, 11 and
15) had 80 %, one event (4) had 60 %, one event (13) had
40 % and two events (9 and 16) had 20 % germination, and
three events (1, 6 and 18) failed to germinate. At 300 mm
NaCl, one event (15) had 80 %, two events (2 and 11) had
60 %, one event (13) had 40 % and two events (9 and 16)
Fig. 7 Relative expression of the abiotic stress-responsive genes
a EaDREB2 transgenics and b co-transformed events
258 Plant Cell Rep (2015) 34:247–263
123
Author's personal copy
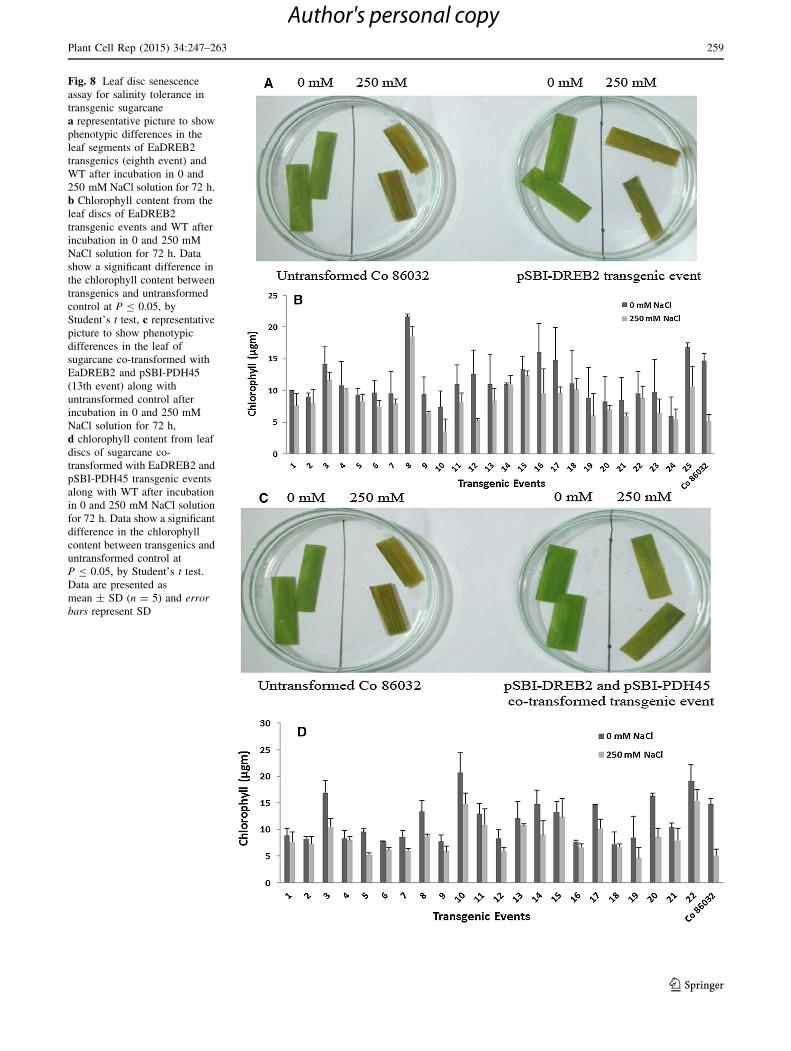
Fig. 8 Leaf disc senescence
assay for salinity tolerance in
transgenic sugarcane
a representative picture to show
phenotypic differences in the
leaf segments of EaDREB2
transgenics (eighth event) and
WT after incubation in 0 and
250 mM NaCl solution for 72 h.
b Chlorophyll content from the
leaf discs of EaDREB2
transgenic events and WT after
incubation in 0 and 250 mM
NaCl solution for 72 h. Data
show a significant difference in
the chlorophyll content between
transgenics and untransformed
control at P B 0.05, by
Student’s t test, c representative
picture to show phenotypic
differences in the leaf of
sugarcane co-transformed with
EaDREB2 and pSBI-PDH45
(13th event) along with
untransformed control after
incubation in 0 and 250 mM
NaCl solution for 72 h,
d chlorophyll content from leaf
discs of sugarcane co-
transformed with EaDREB2 and
pSBI-PDH45 transgenic events
along with WT after incubation
in 0 and 250 mM NaCl solution
for 72 h. Data show a significant
difference in the chlorophyll
content between transgenics and
untransformed control at
P B 0.05, by Student’s t test.
Data are presented as
mean ± SD (n = 5) and error
bars represent SD
Plant Cell Rep (2015) 34:247–263 259
123
Author's personal copy

had 20 % germination and four events (1, 4, 6 and 18)
failed to germinate (Fig. 2g; Table 3).
Shoot height is one of the critical growth parameters,
and was measured as an indicator of salinity tolerance in
transgenic events (Tables 4 and 5: supplementary data). As
there was no germination of the WT in any of the salt
treatments, shoot length measurements could not be
recorded. The shoot length of different transgenics varied
between events under different salt concentrations.
Discussion
Induction of transcription factors contributes to maintain-
ing the integrity of leaf and root cells in soybean during
water stress by triggering the expression of genes associ-
ated with the protection of cell structures against dehy-
dration (Li et al. 2005). DREB2 is the main drought-
responsive transcription factor determining the expression
levels of a number of downstream genes like RD29,
HSP70, etc. (Sakuma et al. 2006; Khan 2011; Mizoi et al.
2011; Qin et al. 2011). The higher transcript level of LEA,
RD29, ERD, ERF, COR15, HSP70 and BRICK genes in
the EaDREB2 overexpressed transgenic sugarcane would
have imparted a higher level of drought and salinity tol-
erance. In the present study the co-expression of the
transgenes resulted in the reduction of relative expression
of the individual transgenes. Reduced expression of the
individual transgenes in co-transformed events may be the
result of perturbation in the entire transcription process of
the plant due to the overexpression of multiple transgenes
under a strong constitutive promoter. Another explanation
for this reduction can be the probable crosstalk between
EaDREB2 and PDH45 pathways. Pyramiding of a tran-
scription factor along with a gene that is not regulated by
the transcription factor per se could not be found in the
literature. In co-transformed events, DREB2, HSP70,
RD29, LEA, ERD, ERF, COR15, BRICK and PDH45 were
significantly upregulated compared to WT but not as high
as in DREB2 transgenics. The screening for drought tol-
erance in these transgenic events has shown that overex-
pression of EaDREB2 led to higher tolerance to drought,
whereas the co-transformed events have shown an
increased tolerance to salinity stress.
It is interesting to note that the transgenics over-
expressing EaDREB2 and the co-transformed events were
found to have increased membrane stability under normal
irrigated condition. However, further increase in the
membrane stability was observed under moisture stress.
The constitutive overexpression of barley group-3 LEA
gene in rice displayed significantly increased tolerance to
water deficit and salinity stresses, which was associated
with higher growth rate, delayed onset of stress damage
symptoms and improved recovery following release of
stress (Xu et al. 1996). Overexpression of LEA in rice also
showed significantly higher RWC, improved turgor, less
reduction in shoot and root growth and improved cell-
membrane stability under prolonged drought conditions. It
was found that LEA did not function as an osmolyte but
contributed to the membrane protection and thus drought
tolerance in rice plants (Chandra Babu et al. 2004). In
EaDREB2 and co-transformed events, the LEA protein was
upregulated 500- and 216-fold, respectively, suggesting
that this might be one of the reasons for its enhanced
membrane stability. Both the EaDREB2 and the co-trans-
formed events showed significantly higher RWC under
moisture stress. This is in accordance with previous reports
on several other transgenic crop plants such as Cenchrus
spp. chickpea, groundnut, wheat, sorghum, and maize
under moisture stress (Nagy et al. 1995; El Hafid et al.
1998; Madhusudan et al. 2002; Bhushan et al. 2007;
Chandra and Dubey 2010).
Drought tolerance associated with stomatal control has
been reported in plants (Pinheiro et al. 2005). The trans-
genic events showed an increased photosynthetic rate than
WT, indicating a substantial protection of photosynthetic
machinery, particularly PS II, during drought stress. It was
found that HSP70 participates in the protection of PS II
against photoinhibition and in the repair of PS II in green
algae (Schroda et al. 1999; Yokthongwattana et al. 2001)
and plays a critical role in the assembly of new PS II core
in tobacco (Tan et al. 2011). This may explain one of the
reasons for the observed higher photosynthetic rate in the
transgenic events. As the main source of energy for plants
is photosynthesis, preservation of the photosynthetic
machinery and its protection contribute significantly to the
plant’s ability to withstand the stress (Mackova et al.
2013). The EaDREB2 and co-transformed events had sig-
nificantly higher chlorophyll content, thus contributing to
the maintenance of higher photosynthetic capacity under
moisture stress. Reduction or no change in chlorophyll
content under drought stress has been observed in different
plant species, and its intensity depends on stress rate and
duration (Rensburg van 1994; Kyparissis et al. 1995; Jag-
tap et al. 1998).
Photochemical quantum efficiency is measured by the
chlorophyll fluorescence parameter Fv/Fm, with maximum
photochemical efficiency of photosystem II in the dark-
adapted state, a trait positively correlated with the organi-
zation and vitality of photosystem II (Aharon et al. 2003).
In healthy leaves, the Fv/Fm value is usually close to 0.8 in
most plant species; therefore, a lower value indicates that a
proportion of PS II reaction centres are damaged or inac-
tivated, a phenomenon termed as photoinhibition and
commonly observed in plants under stress (Baker and
Horton 1987; Baker and Rosenqvist 2004; Zlatev 2009;
260 Plant Cell Rep (2015) 34:247–263
123
Author's personal copy

Vaz and Sharma 2011). Fv/Fm ratios are significantly
higher in EaDREB2 and co-transformed events. The ratio
was not much reduced in transgenic events under moisture
stress, suggesting that the function of reaction centres was
fairly protected in transgenic plants relative to the WT. The
maize transgenic events over expressing PEPC was
reported to show better photosynthetic and water use effi-
ciency with improved dry matter production under soil
moisture stress (Jeanneau et al. 2002).
In WT, the chlorophyll content decreased significantly
compared to the EaDREB2 and co-transformed events
under salinity, suggesting the high salinity tolerance
potential of the transgenic events. Ben Saad et al. (2012)
reported higher stress tolerance in AlSAP (zinc finger
protein) overexpressing rice plants, which maintained the
photosynthetic apparatus integrity by stimulating an
endogenous adaptive potential. Cushman and Bohnert
(2000) suggested that the overexpression or upregulation of
LEA provides salinity tolerance in transgenic plants.
Generally, DREB2 genes are induced by dehydration, high
salinity and heat stress (Liu et al. 1998; Nakashima et al.
2000) and the activation of DREB2 leads to the upregu-
lation of downstream stress-responsive gene expression
and this may be one of the mechanisms for salinity toler-
ance. The mechanism of PDH45-mediated salinity stress
tolerance in transgenics is not clearly understood. How-
ever, based on the properties of PDH45 it was suggested
that there might be two sites of action: (1) it may act at the
translation level to enhance or stabilize protein synthesis,
or (2) it may associate with DNA multisubunit protein
complexes to alter gene expression (Sanan Mishra et al.
2005).
A significant tolerance against salinity stress in trans-
genic events was observed in leaf disc senescence and in
bud germination assays. In numerous crop plants, the
stages of seed germination and early seedling growth are
most susceptible to abiotic stresses (Mito et al. 2010; Se-
khar et al. 2010). However, we could not find any literature
on germination study for salinity tolerance in a vegetatively
propagated plant species. One transgenic event over-
expressing EaDREB2 and one event in the co-transformed
event have shown as high as 80 % germination under high
salt concentration of 300 mM NaCl, while WT failed to
germinate even at lower concentrations. This result agrees
with several previous studies in barley, tomato, Lactuca
sativa, rice and tobacco (Ungar 1978; Bliss et al. 1986;
Bradford 1990; Singla-Pareek et al. 2003; Sanan-Mishra
et al. 2005; Ray and Islam 2008; Dang et al. 2011; Amin
et al. 2012 and Singh et al. 2012), which showed that salt
stress is an important limiting factor for germination in
different crop species. Here, the transgenic plants that
overexpressed EaDREB2 and EaDREB2-PDH45 events
showed increased tolerance to salt stress during bud ger-
mination (Fig. 1e–g).
In conclusion, our results show that transformation of
sugarcane with the EaDREB2 gene under the control of the
Port Ubi 2.3 promoter enhanced the tolerance to water
deficit and salinity stress through improved physiological
adaptation and enhanced stress-related gene expression.
Improved salinity tolerance was observed in the co-trans-
formed events compared to that of EaDREB2. Thus, for
combining drought and salinity tolerance, pyramiding of
these two genes seems to be a better approach. Further
replicated field studies are needed to confirm their toler-
ance to drought and salinity, along with the yield potential
and water use efficiency.
Author contribution statement S. M. Augustine was
responsible for conception and design of experiments,
acquisition of data and analysis, interpretation of data and
drafting of the manuscript; acquisition of data was done by
Ashwin Narayanan, D. P. Syamaladevi, C. Appunu, V.
Ravichandran, and M. Chakravarthi; N. Tuteja provided
the PDH45 construct and N. Subramonian took care of
study conception and design, analysis and interpretation of
data and critical revision.
Acknowledgments This work was supported by the Department of
Biotechnology (DBT) (Grant No. 102/IFD/SAN/325/2013-2014),
Government of India. The authors are grateful to Dr. J. Srikanth,
Principal Scientist, Sugarcane Breeding Institute, Coimbatore, for
critical comments for the improvement of the manuscript.
Conflict of interest The authors declare that they have no conflict
of interest.
References
Agarwal P, Agarwal PK, Joshi AJ, Sopory SK, Reddy MK (2010)
Overexpression of PgDREB2A transcription factor enhances
abiotic stress tolerance and activates downstream stress-respon-
sive genes. Mol Biol Rep 37:1125–1137
Aharon R, Shahak Y, Wininger S, Bendov R, Kapulnik Y, Galili G
(2003) Overexpression of a plasma membrane aquaporin in
transgenic tobacco improves plant vigor under favorable growth
conditions but not under drought or salt stress. Plant Cell
15:439–447
Amin M, Elias SM, Hossain A, Ferdousi A, Rahman MS, Tuteja N,
Seraj ZI (2012) Overexpression of a DEAD box helicase,
PDH45, confers both seedling and reproductive stage salinity
tolerance to rice (Oryza sativa L.). Mol Breed 30:345–354
Arvinth S, Arun S, Selvakesavan RK, Srikanth J, Mukunthan N,
Ananda Kumar P, Premachandran MN, Subramonian N (2010)
Genetic transformation and pyramiding of aprotinin expressing
sugarcane with cry1Ab for shoot borer (Chilo infuscatellus)
resistance. Plant Cell Reports 29:383–395
Augustine SM, Syamaladevi DP, Premachandran MN, Ravichandran
R, Subramonian N (2014) Physiological and molecular insights
Plant Cell Rep (2015) 34:247–263 261
123
Author's personal copy

to drought responsiveness in Erianthus spp. Sugar Tech. doi:10.
1007/s12355-014-0312-7
Baker NR, Horton P (1987) Chlorophyll fluorescence quenching
during photoinhibition. In: Arntzen CJ, Kyle DJ, Osmond CB
(eds) Topics in Photosynthesis, vol 9., photoinhibitionElsevier,
Amsterdam, The Netherlands, pp 145–168
Baker NR, Rosenqvist E (2004) Applications of chlorophyll fluores-
cence can improve crop production strategies: an examination of
future possibilities. J Exp Bot 55:1607–1621
Barrs HD, Weatherley PE (1962) A reexamination of the relative
turgidity technique for estimating water deficits in leaves. Aust J
Biol Sci 15:413–428
Ben Saad R, Walid B, Nabil Z, Jalel A, Delphine M, Emmanuel G,
Radhouane E, Afif H (2012) Marker-free transgenic durum
wheat cv. Karim expressing the AlSAP gene exhibits a high level
of tolerance to salinity and dehydration stresses. Mol Breeding
30:521–533
Bhushan D, Pandey A, Choudhary MK, Datta A, Chakraborty S,
Chakraborty N (2007) Comparative proteomics analysis of
differentially expressed proteins in chickpea extracellular matrix
during dehydration stress. Mol Cell Proteomics 6:1868–1884
Bihani P, Char B, Bhargava S (2011) Transgenic expression of
sorghum DREB2 in rice improves tolerance and yield under
water limitation. J Agric Sci 149:95–101
Bliss RD, Plattaloia KA, Thomson WW (1986) Osmotic sensitivity in
relation to salt sensitivity in germinating barley seeds. Plant Cell
Environ 9:721–725
Bohnert HJ, Nelson DE, Jensen RG (1995) Adaptations to environ-
mental stress. Plant Cell 7:1099–1111
Bradford KJ (1990) A water relation analysis of seed germination
rates. Plant Physiol 94:840–849
Chandra Babu R, Zhang JX, Blum A, Ho DTH, Wu R, Nguyen HT
(2004) HVA1, a LEA gene from barley confers dehydration
tolerance in transgenic rice (Oryza sativa L.) via cell membrane
protection. Plant Sci 166:855–862
Chandra A, Dubey A (2010) Effect of ploidy levels on the activities of
D-pyrroline-5-carboxylate synthetase, superoxide dismutase and
peroxidase in Cenchrus species grown under water stress. Plant
Physiol Biochem 48:27–34
Chen M, Wang QY, Cheng XG, Xu ZS, Li LC, Ye XG, Xia LQ, Ma
YZ (2007) GmDREB2, a soybean DRE-binding transcription
factor, conferred drought and high-salt tolerance in transgenic
plants. Biochem Biophys Res Commun 353:299–305
Chen J, Xia X, Yin W (2009) Expression profiling and functional
characterization of a DREB2-type gene from Populus euphra-
tica. Biochem Biophys Res Commun 378:483–487
Chinnusamy V, Jagendorf A, Zhu JK (2005) Understanding and
improving salt tolerance in plants. Crop Sci 45:437–448
Chomczynski P, Mackey K (1995) Modification of the TRI reagent
DNA/protein isolation procedure for isolation of RNA from
polysaccharide and proteoglycan rich sources. Biotechniques
19:942–945
Crafts-Brandner SJ, Below FE, Harper JE, Hageman RH (1984)
Effects of pod removal on metabolism and senescence of
nodulating and nonnodulating Soybean isolines: II. Enzymes and
chlorophyll. Plant Physiol 75(2):318–322
Cushman JC, Bohnert HJ (2000) Genome approaches to plant stress
tolerance. Curr Opin Plant Biol 3:117–124
Dang HQ, Tran NQ, Gill SS, Tuteja R, Tuteja N (2011) A single
subunit MCM6 from pea promotes salinity stress tolerance
without affecting yield. Plant Mol Biol 76:19–34
El Hafid R, Smith DH, Karrou M, Samir K (1998) Physiological
attributes associated with early-season drought resistance in
spring durum wheat cultivars. Can J Plant Sci 78:227–237
Engelbrecht BMJ, Melvin TT, Thomas AK (2007) Visual assessment
of wilting as a measure of leaf water potential and seedling
drought survival. J Trop Ecol 23:497–500
Fan L, Zheng S, Wang X (1997) Antisense suppression of
phospholipase Da retards abscisic acid and ethylene-promoted
senescence of postharvest Arabidopsis leaves. Plant Cell
9:2183–2196
Hussain SS, Kayani MA, Amjad M (2011) Transcription factors as
tools to engineer enhanced drought tolerance in plants. Am Inst
Chem Eng. doi:10.1002/btpr.514
Jagtap V, Bhargava S, Sterb P, Feierabend J (1998) Comparative
effect of water, heat and light stresses on photosynthetic
reactions in Sorghum bicolor (L.) Moench. J Exp Bot
49:1715–1721
Jeanneau M, Vidal J, Gousset-dupont A, Lebouteiller B, Hodges M,
Gerentes D, Perez P (2002) Manipulating PEPC levels in plants.
J Exp Bot 53:1837–1845
Kasuga M, Liu Q, Miura S, Yamaguchi-Shinozaki K, Shinozaki K
(1999) Improving plant drought, salt, and freezing tolerance by
gene transfer of a single stress inducible transcription factor. Nat
Biotechnol 17:287–291
Kasuga M, Miura S, Shinozaki K, Yamaguchi-Shinozaki K (2004) A
combination of the Arabidopsis DREB1A gene and stress-
inducible rd29A promoter improved drought- and low-temper-
ature stress tolerance in tobacco by gene transfer. Plant Cell
Physiol 45:346–350
Khan MS (2011) The role of DREB transcription factors in abiotic
stress tolerance of plants. Biotechnol Biotechnol
25(3):2433–2442
Kyparissis A, Petropoulou Y, Manetas Y (1995) Summer survival of
leaves in a soft-leaved shrub (Phlomis fruticosa L. Labiatae)
under mediterranean field conditions: avoidance of photoinhib-
itory damage through decreased chlorophyll contents. J Exp Bot
46:1825–1831
Latha C, Prasad Manoj (2011) Role of DREBs in regulation of abiotic
stress responses in plants. J Exp Bot 62(14):4731–4748
Li XP, Tian AG, Luo GZ, Gong ZZ, Zhang JS, Chen SY (2005)
Soybean DRE-binding transcription factors that are responsive to
abiotic stresses. Theor Appl Genet 110:1355–1362
Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki
K, Shinozaki K (1998) Two transcription factors, DREB1 and
DREB2, with an EREBP/AP2 DNA binding domain separate
two cellular signal transduction pathways in drought- and low-
temperature-responsive gene expression, respectively, in Arabi-
dopsis. Plant Cell 10:1391–1406
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression
data using real time quantitative PCR and the 2-DDCT method.
Methods 25:402–408
Mackova H, Marie H, Jana D, Veronika T, Ondrej N, Zuzana L,
Vaclav M, Daniel H, Tomas H, Ilja TP, Alena G, Helena S, Eva
G, Tomas W, Thomas S, Radomıra V (2013) Enhanced drought
and heat stress tolerance of tobacco plants with ectopically
enhanced cytokinin oxidase/dehydrogenase gene expression.
J Exp Bot 64(10):2805–2815
Madhusudan KV, Giridarakumar S, Ranganayakulu GS, Reddy PC,
Sudhakar C (2002) Effect of water stress on some physiolog-
ical responses in two groundnut (Arachis hypogea L.) culti-
vars with contrasting drought tolerance. J Plant Biol 29:
199–202
Manjulatha M, Sreevathsa R, Kumar AM, Sudhakar C, Prasad TG,
Tuteja N, Udayakumar M (2014) Overexpression of a pea DNA
helicase (PDH45) in peanut (Arachis hypogaea L.) confers
improvement of cellular level tolerance and productivity under
drought stress. Mol Biotechnol 56(2):111–125
262 Plant Cell Rep (2015) 34:247–263
123
Author's personal copy

Martineau JR, Specht JE, Williams JH, Sullivan CY (1979) Temper-
ature tolerance in soybeans. I. Evaluation of a technique for
assessing cellular membrane thermostability. Crop Sci 19:75–78
Mito T, Seki M, Shinozaki K, Ohme Takagi M, Matsui K (2010)
Generation of chimeric repressors that confer salt tolerance in
Arabidopsis and rice. Plant Biotechnol J 9:736–746
Mizoi J, Shinozaki K, Yamaguchi-Shinozaki K (2011) AP2/ERF
family transcription factors in plant abiotic stress responses.
Biochim Biophys Acta 1819(2):86–96
Moran JF, Becana M, Iturbe-Ormaetxe I, Frechilla S, Klucas RV,
Aparicio Tejo P (1994) Drought induces oxidative stress in pea
plants. Planta 194:346–352
Nagy Z, Tuba Z, Zsoldos F, Erdei L (1995) CO2 exchange and water
relation responses of sorghum and maize during water stress.
J Plant Physiol 145:539–544
Nakashima K, Shinwari ZK, Sakuma Y, Seki M, Miura S, Shinozaki
K, Yamaguchi-Shinozaki K (2000) Organization and expression
of two Arabidopsis DREB2genes encoding DRE-binding pro-
teins involved in dehydration- and high salinity-responsive gene
expression. Plant Mol Biol 42:657–665
Pham XH, Reddy MK, Ehtesham NZ, Matta B, Tuteja N (2000) A
DNA helicase from Pisum sativum is homologous to translation
initiation factor and stimulates topoisomerase I activity. Plant J
24:219–229
Phillip A, Syamaladevi DP, Chakravarthy M, Gopinath K, Subramo-
nian N (2013) 50 Regulatory region of ubiquitin 2 gene from
Porteresia coarctata makes efficient promoters for transgene
expression in monocots and dicots. Plant Cell Rep 32:1199–1210
Pinheiro HA, Da Matta FM, Chaves ARM, Loureiro ME, Ducatti C
(2005) Drought tolerance is associated with rooting depth and
stomatal control f water use in clones of Coffea canephora. Ann
Bot 96:101–108
Qin H, Qiang G, Junling Z, Sun L, Sundaram K, Yizheng Z, Mark B,
Paxton P, Eduardo B, Hong Z (2011) Regulated expression of an
Isopentenyl transferase gene (IPT) in peanut significantly
improves drought tolerance and increases yield under field
conditions. Plant Cell Physiol 52(11):1904–1914
Ray PKS, Islam MA (2008) Genetic analysis of salinity tolerance in
rice. Bangla J Agric Res 33:519–529
Reis RR, Barbara ADBC, Polyana KM, Maria TBM, Jean CA,
Antonio CJ, Alan CA, Ana PR, Feng Q, Junya M, Yamaguchi-
Shinozaki K, Kazuo N, Josirley FCC, Carlos AFS, Alexandre
LN, Adilson KK, Hugo BCM (2014) Induced over-expression of
AtDREB2A CA improves drought tolerance in sugarcane.
doi:10.1016/j.plantsci.2014.02.003
Sakuma Y, Maruyama K, Osakabe Y, Qin F, Seki M, Shinozaki K,
Yamaguchi-Shinozaki K (2006) Functional analysis of an
Arabidopsis transcription factor, DREB2A, involved in drought
responsive gene expression. Plant Cell 18:1292–1309
Sambrook J, Russel DW (1989) Molecular cloning: a laboratory
manual, III edn. Cold Spring Harbor Laboratory Press, New
York
Sanan Mishra N, Pham XH, Sopory SK, Tuteja N (2005) Pea DNA
helicase 45 overexpression in tobacco confers high salinity
tolerance without affecting yield. Proc Natl Acad Sci USA
102:509–514
Schroda M, Vallon O, Wollman FA, Beck CF (1999) A chloroplast-
targeted heat shock protein 70 (HSP70) contributes to the photo
protection and repair of photosystem II during and after
photoinhibition. Plant Cell 11:1165–1178
Sekhar K, Priyanka B, Reddy VD, Rao KV (2010) Isolation and
characterization of a pigeon pea cyclophilin (CcCYP) gene, and
its over-expression in Arabidopsis confers multiple abiotic stress
tolerance. Plant Cell Environ 33:1324–1338
Singh S, Modi MK, Gill SS, Tuteja N (2012) Rice: genetic
engineering approaches for abiotic stress tolerance retrospects
and prospects. In: Tuteja N, Gill SS, Tuteja R (eds) Improving
crop productivity in sustainable agriculture. Wiley-VCH Verlag
GmbH and Co, KGaA, Germany, pp 203–225
Singla-Pareek SL, Reddy MK, Sopory SK (2003) Genetic engineering
of the glyoxalase pathway in tobacco leads to enhanced salinity
tolerance. Proc Natl Acad Sci USA 100:14672–14677
Stockinger EJ, Gilmour SJ, Thomashow MF (1997) Arabidopsis
thaliana CBF1 encodes an AP2 domain-containing transcrip-
tional activator that binds to the C-repeat/DRE, a cis-acting DNA
regulatory element that stimulates transcription in response to
low temperature and water deficit. Proc the Natl Acad Sci USA
94:1035–1040
Tan W, Meng QW, Brestic M, Olsovska K, Yang XH (2011)
Photosynthesis is improved by exogenous calcium in heat
stressed tobacco plants. J Plant Physiol 168:2063–2071
Ungar IA (1978) Halophyte seed germination. Bot Rev 44:233–264
Valliyodan B, Nguyen HT (2006) Understanding regulatory networks
and engineering for enhanced drought tolerance in plants. Curr
Opin Plant Biol 9:189–195
van Rensburg L, Kruger GHJ (1994) Evaluation of components of
oxidative stress metabolism for use in selection of drought
tolerant cultivars of Nicotiana tabacum L. J Plant Physiol
143:730–737
Vaz J, Sharma PK (2011) Relationship between xanthophyll cycle
and non-photochemical quenching in rice (Oryza sativa L.)
plants in response to light stress. Indian J Exp Bot 49:60–67
Xu D, Duan X, Wang B, Hong B, Ho T, Wu R (1996) Expression of a
late embryogenesis abundant protein gene, HVA1, from barley
confers tolerance to water deficit and salt stress in transgenic
rice. Plant Physiol 110:249–257
Yamaguchi-Shinozaki K, Shinozaki K (1994) A novel cis-acting
element in an Arabidopsis gene is involved in responsiveness to
drought, low-temperature, or high-salt stress. Plant Cell
6:251–264
Yokthongwattana K, Chrost B, Behrman S, Casper-Lindley C, Melis
A (2001) Photosystem II damage and repair cycle in the green
alga Dunaliella salina: involvement of a chloroplast-localized
HSP70. Plant Cell Physiol 42:1389–1397
Zlatev Z (2009) Drought-induced changes in chlorophyll fluorescence
of young wheat plant. Biotechnol Biotechnol Equip 23:437–441
Plant Cell Rep (2015) 34:247–263 263
123
Author's personal copy
Copyright © 2022 FDOKUMEN




















