
Creating Biomimetic Surfaces through Covalent and Oriented Binding of Proteins

Original Article
Collagen-Binding Peptidoglycans: A BiomimeticApproach to Modulate Collagen Fibrillogenesis
for Tissue Engineering Applications
John E. Paderi, Ph.D., Rizaldi Sistiabudi, Ph.D., Albena Ivanisevic, Ph.D., and Alyssa Panitch, Ph.D.
The small leucine-rich proteoglycans (SLRPs), prevalent in collagenous tissues, regulate collagen fibrillogenesisand provide a host of biochemical cues critical to tissue function and homeostasis. Incorporating SLRPs mayenhance tissue engineering designs that mimic the native extracellular matrix, although SLRPs purified fromanimal sources bear low yields and lack design control. Consequently, we have designed synthetic peptido-glycans, inspired by the native SLRP decorin, that contain a collagen-binding peptide attached to a glycos-aminoglycan (GAG) chain. These peptidoglycans modulate collagen fibrillogenesis and decrease fibril diameterin vitro, similarly to decorin, while maintaining the characteristic D-banded fibrils. Application for tissue engi-neering is demonstrated as these peptidoglycans are incorporated into collagen gels seeded with smooth musclecells. Gels formed with peptidoglycans and decorin show a faster rate of gel compaction, and one peptidoglycanuniquely increases elastin production. The peptidoglycan design can be tailored with respect to the peptidesequence and GAG identity and is expected to have versatile application in tissue engineering.
Introduction
Collagen, the most abundant protein in mammals, isthe principle component of numerous tissues in the
body. Its abundance, together with its ability to self-assemblein vitro to form insoluble fibrils, make collagen a useful andversatile material for tissue-engineered scaffolds. Based uponaims in tissue engineering to design scaffolds that mimic themechanical and biochemical signals of the native extracel-lular matrix (ECM), researchers have employed collagen fornumerous applications.1–4 The ability to control fibril orien-tation and diameter size is important for engineering themechanical characteristics of collagen-based scaffolds; how-ever, although methods for controlling fibril alignmenthave been developed, methods for controlling fibril diameterwhile maintaining physiological conditions have yet to bedeveloped.5–8
The small leucine-rich proteoglycans (SLRPs), prevalent incollagenous tissues, have gained interest for their role inregulating collagen fibrillogenesis and providing a host ofbiochemical cues critical to tissue function and homeosta-sis.9–18 Several SLRPs have been identified and are known tocontain a protein core with an attached glycosaminoglycan(GAG) chain.19,20 Decorin is a well-studied SLRP and isknown to delay fibrillogenesis in vitro and modulate fibrildiameter in vitro and in vivo.17,21–23 The core protein of dec-orin (decoron)24 contains many functional domains, includ-
ing two regions known to bind collagen.25,26 Decorin bindingto collagen inhibits lateral aggregation of collagen duringfibrillogenesis, thus modulating the fibril diameter.17,27
Simplifying decorin to its collagen-binding function withattached GAG, we have previously designed a peptido-glycan, which contains a collagen-binding peptide and acovalently attached dermatan sulfate (DS) GAG.28 Thispeptidoglycan modulates collagen fibrillogenesis while in-corporating a biochemically active GAG chain. It has alsobeen shown to increase the stiffness of collagen gels, simi-larly to decorin. For collagen-based tissue engineering, pep-tidoglycans may provide unique application; they mimic inpart, native SLRPs, by binding collagen and incorporating aGAG chain, but unlike SLRPs purified from animal tissues,peptidoglycans have the benefit of design control and ease ofsynthesis.
Collagen-based tissue-engineered scaffolds have been usedfor vascular tissue engineering with promising results.29 Onedrawback to collagen-based scaffolds is that smooth musclecells (SMCs), which maintain the vessel ECM, only minimallydegrade collagen and synthesize little new matrix.2,30 In-corporating GAGs, SLRPs, or peptidoglycans may improvethe design of collagen-based scaffolds by modulating collagenfibril structure and incorporating the biochemically active in-gredients present in the native tissue.2,31,32
The present work broadens the scope of our previouslydescribed peptidoglycan, by demonstrating tailorability of
Weldon School of Biomedical Engineering, Purdue University, West Lafayette, Indiana.
TISSUE ENGINEERING: Part AVolume 15, Number 10, 2009ª Mary Ann Liebert, Inc.DOI: 10.1089=ten.tea.2009.0014
2991

the peptidoglycan molecule, analyzing the morphologicalcharacteristics of collagen fibrils with incorporated peptido-glycans, and demonstrating tissue engineering application.We hypothesized that these molecules would behave simi-larly to native SLRPs in modulating collagen fibrillogenesisand would influence cellular remodeling of collagen matri-ces. The designed peptidoglycans offer a tailorable biomi-metic approach to influence collagen fibril morphology whileincorporating biochemical cues that influence cell function;as such, these peptidoglycans are expected to have versatileapplication in tissue engineering.
Materials and Methods
Materials
Pepsin-treated Nutragen Type I Collagen purified from bo-vine hide was purchased from Inamed Biomaterials (Freemont,CA) at a stock concentration of 6.4 mg=mL in 10 mM of hy-drochloric acid (HCl). DS (41 kDa, 6.85% sulfur) and its oxi-dized form, containing an average of 1.1 aldehydes per polymerchain, were purchased from Celsus Laboratories (Cincinnati,OH). Decorin from bovine tendon (100 kDa, 50 kDa of which isDS) was purchased from Sigma-Aldrich (St. Louis, MO). Aminoacids were purchased from Anaspec (San Jose, CA), Knorr resinwas purchased from SynBioSci Corporation (Livermore, CA),and all other supplies were purchased from VWR or Sigma-Aldrich, West Chester, PA unless otherwise noted.
Synthesis/characterization DS-Dc13
The peptide sequences RRANAALKAGELYKSILYGC(SILY), SYIRIADTNITGC (Dc13), and ZSYIRIADTNITGC(ZDc13), where Z designates dansyl glycine, were synthesizedon a solid-phase Symphony Peptide Synthesizer (ProteinTechnologies, Tucson, AZ) with Knorr resin. Peptides werecleaved from the resin using 92.5% trifluoroacetic acid, 2.5%MilliQ water, 2.5% triisopropylsilane, and 2.5% ethane dithiol.Peptides were purified using reverse-phase chromatographyon an AKTA Explorer FPLC (GE Healthcare, Piscataway, NY)using a C-18 column (Grace-Vydac, Deerfield, IL) with aceto-nitrile=water gradient, and purity was confirmed using massspectroscopy (Applied Biosystems 4700, Foster City, CA).
Conjugation of peptides to dermatan sulfate
Peptides were conjugated to oxidized DS as described previ-ously using the heterobifunctional cross-linker 3-(2-pyridyldithio)propionyl hydrazide (PDPH; Pierce, Rockford, IL), reactive toaldehyde and thiol groups.28 Briefly, PDPH was reacted incoupling buffer (0.1 M of sodium phosphate, 0.25 M of sodiumchloride, pH 7.2) at 10-fold molar excess with oxidized DScontaining 1.1 aldehyde groups per DS polymer chain. ExcessPDPH was removed using gel filtration by a column packedwith Sephadex G-25 medium (GE Healthcare). Purified DS-PDPH was then reacted in coupling buffer with 5-fold molarexcess of peptide. The reaction was monitored according to theproduction of pyridine-2-thione as described. Excess peptidewas removed using gel filtration, and pure DS-peptide waslyophilized and stored at �208C until further testing.
Binding of DS-Dc13 to collagen
Collagen and bovine serum albumin was immobilized ona high-bind 384-well black=clear-bottom plate by incubating
each at 2 mg=mL for 1 h at 378C in 10 mM of HCl and 1�phosphate buffered saline (PBS), respectively. Wells werethen washed three times with 1�PBS. Untreated wells wereused as a positive control. A mix of 60% DS-PDPH cappedwith L-cysteine and 40% fluorescently labeled DS-ZDc13 wasdissolved in 1�PBS at varying concentrations from 100mM to1 nM and incubated in the treated or control wells for 30 minat 378C. Unbound DS-ZDc13 was removed by washing threetimes with 1�PBS, and fluorescence (ex: 335 nm em: 490 nm)was measured on a SpectraMax M-5 spectrophotometer(Molecular Devices, Sunnyvale, CA). All measurements wereperformed in triplicate.
Collagen gel preparation and fibrillogenesis assays
Samples compared in this study include collagen alone andcollagen mixed with DS, DS-SILY, DS-Dc13, intact decorin, orfree peptides SILY or Dc13. Unless noted, treatment concen-trations were added at a 10:1 molar ratio of collagen:additive.Molar ratios were calculated by an estimate of molecularweights: collagen, 285 kDa, DS 41, kDa; DS-SILY, 43.2 kDa; anddecorin, 100 kDa. Gels were formed by mixing stock collagensolution with 10�PBS, 1 M of sodium hydroxide, and 1xPBS onice to a final concentration of 4 mg=mL, pH 7.4, and ionicstrength of 164 mM. Treatments were added as a part of thefinal 1�PBS addition and incubated on ice for 30 min beforeinitializing fibrillogenesis by warming to 378C. Fibrillogenesiswas monitored turbidimetrically by absorbance at 313 nm asdescribed previously.8,20,22 Samples of 50mL were added at 48Cto each well of a 384-well plate, and measurements were takenat 1-min intervals for up to 6 h in a SpectraMax M-5 spectro-photometer set at 378C.
Cryo-scanning electron microscopyand fibril diameters
Gels for cryo-scanning electron microscopy (SEM) wereformed directly on the scanning electron microscope stage at378C overnight as described previously.33 The stages were thensecured in a cryo-holder and plunged into liquid nitrogenslush. Samples were then transferred to a Gatan Alto, Plea-santon, CA, 2500 pre-chamber cooled to �1708C under vac-uum. A free-break surface was created using a cooled scalpel,and the sample was sublimated for 20 min, followed by plati-num sputter coating for 120 s. Samples were transferred to thecryo-stage at �1308C, where regions with similar orientationwere imaged for comparison across treatments. Fibril diame-ters were measured using ImageJ software (National Institutesof Health (NIH), Bethesda, MD), in which regions orthogonalto the fibril were measured. Three independent observersblinded to the treatment recorded at least 45 measurements foreach treatment and were instructed to measure diameters onlyof fibrils that could be resolved to individual fibrils, rather thanthose appearing fused, overlapping, or obscured by otherartifacts.
Atomic force microscopy
Gels were formed on cut glass cover slides and incubatedovernight at 378C. Samples were dehydrated and washedwith MilliQ water, and the glass slides were fixed to theatomic force microscope (AFM) metal specimen disc (#16208)purchased from Ted Pella (Redding, CA). Tapping mode
2992 PADERI ET AL.

AFM was performed using multimode scanning probe micros-copy from Veeco Instruments (Santa Barbara, CA). A single-beam silicon tip (#OTESPA) was purchased from Veeco Probes(Camarillo, CA) and had spring constant of 42 N=m. AFM im-aging was performed at room temperature under ambient hu-midity. Images were collected at a 1 Hz scan rate.
Cell culture and gel compaction
Human coronary artery (HCA) SMCs (Cascade Biologics,Portland, OR) were cultured in growth medium (Medium231 supplemented with smooth muscle growth factor). Cellsfrom passage 3 were used for all experiments. Differentiationmedium (Medium 231 supplemented with 1% fetal bovineserum and 1� penicillin=streptomycin) was used for all ex-periments unless otherwise noted. This medium differs frommanufacturer protocol in that it does not contain heparin.
Collagen gels were prepared with each additive, as de-scribed, with the exception that the 1�PBS addition wasomitted to accommodate the addition of cells in media. Col-lagen pregel was incubated on ice for 30 min, followed byaddition of HCA SMCs in differentiation medium to a finalconcentration of 1�106 cells=mL. Gels were formed in qua-druplicate in 48-well non-tissue culture treated plates (Costar,Corning, NY) for 6 h before adding 500mL per well of differ-entiation medium. Gels were freed from the well edges after24 h to allow free gel compaction. After 24 h, growth mediumwas collected and then replaced every 2 to 3 days; images forcompaction were taken at the same time points using a GelDoc System (Bio-Rad, Hercules, CA). The cross-sectional areaof circular gels correlating to degree of compaction was de-termined using ImageJ software (NIH). Gels containing nocells were used as a negative control, and cells in collagen gelsabsent additive were used as a positive control.
Collagen degradation
Collagen degradation was measured by determining thehydroxyproline content released from gels into the cell me-dium, following a protocol adopted from Reddy and En-wemeka, assuming that 12% of the weight of collagen ishydroxyproline.28,34 Medium (200 mL) was collected every 2 to3 days and was hydrolyzed in 6 M of HCl at 1108C overnight.After HCl removal by evaporation, Chloramine T reagent wasfreshly made, and 450mL was added to each sample and al-lowed to incubate at room temperature for 25 min. Ehrlich’saldehyde reagent was freshly made and 500mL mixed witheach sample and then incubated at 658C for 20 min. Absor-bance was read at 550 nm, and hydroxyproline concentrationwas determined from a collagen standard curve.
Elastin production
Collagen gels seeded with HCA SMCs were prepared asdescribed. Differentiation medium was changed every threedays and gels were cultured for 10 days. Collagen gelscontaining no cells were used as a control. At day 10, gelswere rinsed in 1� PBS overnight, and gels were tested forelastin content using the Biocolor, United Kingdom, Fastinelastin assay, per the manufacturer’s protocol (Biocolor,County Atrim, United Kingdom). Briefly, gels were solubi-lized in 0.25 M of oxalic acid by incubating at 1008C for 1 h.Elastin was precipitated using Fastin Precipitation Solution,
and samples were then centrifuged at 11,000�g for 10 min.The solubilized collagen supernatant was removed, and theelastin pellet was stained using Fastin Dye Reagent for90 min at room temperature. Samples were centrifuged at11,000�g for 10 min, and unbound dye in the supernatantwas removed. Dye from the elastin pellets was released bythe Fastin Dye Dissociation Reagent, and 100-mL sampleswere transferred to a 96-well plate (Costar). Absorbance wasmeasured at 513 nm, and elastin content was calculated froman a-elastin standard curve.
Cytotoxicity
HCA SMCs were seeded at 4.8�104 cells=mL in growthmedium onto a 96-well tissue-culture black=clear-bottom plate(Costar) and allowed to adhere for 4 h. Growth medium wasaspirated, and 600mL of differentiation medium containingeach additive at a concentration equivalent to the concentra-tion within collagen gels (1.4�10�6 M) was added to each well.Cells were incubated for 48 h and then tested for cytotoxicityusing Live-Dead and CyQuant (Invitrogen, Carlsbad, CA)assays, per the manufacturer’s protocol. Cells in differentia-tion medium containing no additive were used as control.
Statistics
All experiments were performed in at least triplicate andare presented as averages� standard errors. Significance wasdetermined using Design Expert (StatEase, Minneapolis, MN)software, with a¼ 0.05.
Results and Discussion
We have previously described the design of a syntheticpeptidoglycan based on the SLRPs such as decorin in which theGAG DS is conjugated to a collagen-binding peptide. Here wehave expanded the design, demonstrating tailoribility usingtwo different collagen-binding peptide sequences, one derivedfrom decorin SYIRIADTNITGC (Dc13) and one from theplatelet receptor to type I collagen RRANAALKAGELYKSI-LYGC (SILY).35,36 Although the exact binding location alongthe collagen fibril was not determined in this study, it is as-sumed that these sequences, derived from different sources andwith different primary structure, bind collagen in different lo-cations, and they were thus studied for comparison. In addi-tion, each sequence not containing an internal cysteine could bemodified using a terminal glycine (spacer) and cysteine toemploy specific sulfhydryl chemistry. We have compared theeffects of these peptidoglycans on collagen fibrillogenesis andmorphology of collagen gels, showing a level of control overthe gel biophysical properties, and demonstrated tissue engi-neering application by culturing HCA SMCs in collagen gelswith incorporated peptidoglycans.
Synthesis and characterizationof DS-Dc13 and DS-SILY
As shown in Figure 1, approximately 1 Dc13 peptide wascoupled per DS molecule, resulting in a peptidoglycan that issimilar to DS-SILY and is designed to mimic in part nativeSLRPs such as decorin, which contains one collagen-bindingdecoron per GAG chain.28,37 To ensure that conjugation ofDS to Dc13 did not inhibit the ability of Dc13 to bind tocollagen, fluorescently labeled DS-ZDc13, where Z denotes
COLLAGEN-BINDING PEPTIDOGLYCANS 2993

dansyl glycine, was conjugated to DS and in a microplateassay, bound specifically to collagen in a dose-dependentmanner shown in Fig. 2. Because saturation binding was notachieved at the maximum concentration of 100mM, thebinding affinity could not be calculated. DS is known to in-teract electrostatically with collagen and can self-associate;thus increasing the concentration to reach saturation bindingof the peptidoglycan was not possible, although specificbinding to collagen was demonstrated, making DS-Dc13 asuitable collagen-binding peptidoglycan.38
Collagen fibrillogenesis and morphology
The effects on fibrillogenesis and fibril morphology wereassessed according to turbidity. DS-SILY and DS-Dc13 delayfibrillogenesis in a dose-dependent manner, as shown inFigure 3, in which, at high concentrations (1:1 molar ratio),marked inhibition is observed, following the same trends asobserved for native SLRPs.22,39 Free peptide controls did nothave an effect on fibrillogenesis even at the high concentra-tions shown in Figure 3 and previously.28
A proposed mechanism for the interaction of collagen-binding peptidoglycans, depicted in Figure 4, is the same asthat proposed for SLRPs, in which the binding of thesemolecules to collagen monomer or onto forming fibrils in-hibits lateral fibril growth during fibrillogenesis.15,17,22,37,40,41
In vitro, collagen self-associates into aggregates in the form offibrils, which are specifically arranged, quarter-staggered col-lagen molecules.42 Delayed fibrillogenesis is dose dependent,as illustrated at the 1:1 molar ratio of collagen:peptidoglycan,
FIG. 2. Microplate fluorescence binding of dermatan sul-fate (DS)-ZDc13 to collagen. DS-ZDc13 bound specifically tothe collagen surface in a dose-dependent manner, althoughsaturation was not achieved at the maximum concentration.(101�62 mm) (300�300 DPI). NT, no treatment; BSA, bovineserum albumin.
FIG. 3. Collagen fibrillogenesis according to turbidity mea-surements. (A) Dermatan sulfate (DS)-Dc13 delayed fibrillo-genesis and decreased optical density in a dose-dependentmanner. Free Dc13 peptide in contrast had little difference ineffect on fibrillogenesis from collagen alone. (B) DS-SILY andDS-Dc13 at 10:1 and 1:1 molar ratios delay fibrillogenesis anddecrease optical density by similar amounts (101�109 mm)(300�300 DPI).
FIG. 1. Conjugation of Dc13 to dermatan sulfate (DS).Production of pyridine-2-thione, measured according to anincrease in absorbance at 343 nm, indicates 0.99 Dc13 pep-tides per DS polymer chain (101�65 mm) (300�300 DPI).
FIG. 4. Peptidoglycan–collagen interaction mechanism.Proposed inhibition of lateral aggregation by binding ofpeptidoglycan or small leucine-rich proteoglycans (SLRPs)along a collagen molecule. The required quarter-stagger ar-rangement of collagen in the lateral direction is prevented bybound peptidoglycan or SLRPs, limiting growth to the axialdirection. The exact binding location of peptidoglycan is notproposed here but a schematic of the mechanism of lateralinhibition (101�47 mm) (300�300 DPI).
2994 PADERI ET AL.

indicating that, as more peptidoglycans are bound to collagen,they inhibit the required quarter-stagger arrangement of col-lagen, thus preventing or delaying fibril formation. Unlikepeptidoglycans, DS alone increases the rate of fibrillogenesis.Because DS interacts electrostatically with collagen, it is likelythat collagen fibrils are brought together, nucleating fibrilsmore quickly and speeding fibrillogenesis. Because the inter-action between collagen and DS is weaker than that betweencollagen and peptide, DS does not remain bound where itwould inhibit lateral aggregation of collagen.43
Fibril morphology was assessed using cryo-SEM tech-niques and is depicted in Figure 5. Cryo-SEM does not re-quire dehydration steps, which can collapse the fibrillarstructure, making fibril diameter measurements less accu-rate, and is thus a preferred method for assessing collagengel structure.33 Inhibited lateral aggregation would result insmaller diameter fibrils, which is indicated by decreased
optical density observed from turbidity studies and confirmedby direct measurement of fibril diameters (Fig. 4). Histo-grams illustrating fibril size distribution show non-normaldistribution for some treatments, so the Kolmogorov-Smirnovtest, which does not assume normal distribution, was used forstatistical analysis. Gels formed in the presence of decorin,DS-Dc13, and DS-SILY resulted in significantly smaller averagefibril diameter (10%, 15%, and 27%, respectively) than collagen.The ability to modulate collagen fibril diameter under physi-ological conditions is significant for collagen-based tissue en-gineering applications because fibril size plays an importantrole in cellular response, as well as the mechanical properties ofcollagen scaffolds. Native tissues contain fibrils ranging in di-ameter from 10s to 100s of nanometers, depending on the tis-sue, which contributes to the unique mechanical and cellularenvironments.44 Synthetic peptidoglycans also incorporate thebiochemically active DS GAG chain, so fibril diameter cannot
FIG. 5. Cryo-canning electron microscopy images and fibril diameter distribution. The addition of decorin, dermatan sulfate(DS)-SILY, and DS-Dc13 resulted in significantly smaller fibril diameter than collagen alone or collagen with DS. Free peptideSILY also reduced fibril diameter, whereas Dc13 had no effect. *Significance vs collagen or collagenþDS, **DS-SILY sig-nificant vs decorin (254�190 mm) (300�300 DPI).
COLLAGEN-BINDING PEPTIDOGLYCANS 2995
be isolated as a distinct variable in cellular response or me-chanical properties.
The free peptide SILY also decreased fibril diameter,whereas Dc13 did not. The size of these peptide sequences, inwhich SILY containing 20 amino acid residues could steri-cally hinder collagen–collagen interactions, whereas Dc13containing only 13 amino acid residues may not may in partexplain this. Likewise, when the 40-kDa DS molecule is at-tached, both peptidoglycans are large enough to inhibit lat-eral aggregation of collagen. Because free SILY peptide doesnot incorporate DS, its may be a useful peptide for studyingthe effect of fibril diameter independently.
In addition to fibril diameter, cryo-SEM images also pro-vided information on the branching structure of the fibrillarnetwork of collagen gels, revealing a highly branched net-work, especially in the presence of DS-SILY. The specificbinding location of the peptidoglycan on growing collagenfibrils may influence fibril branching, which is currently be-ing studied.
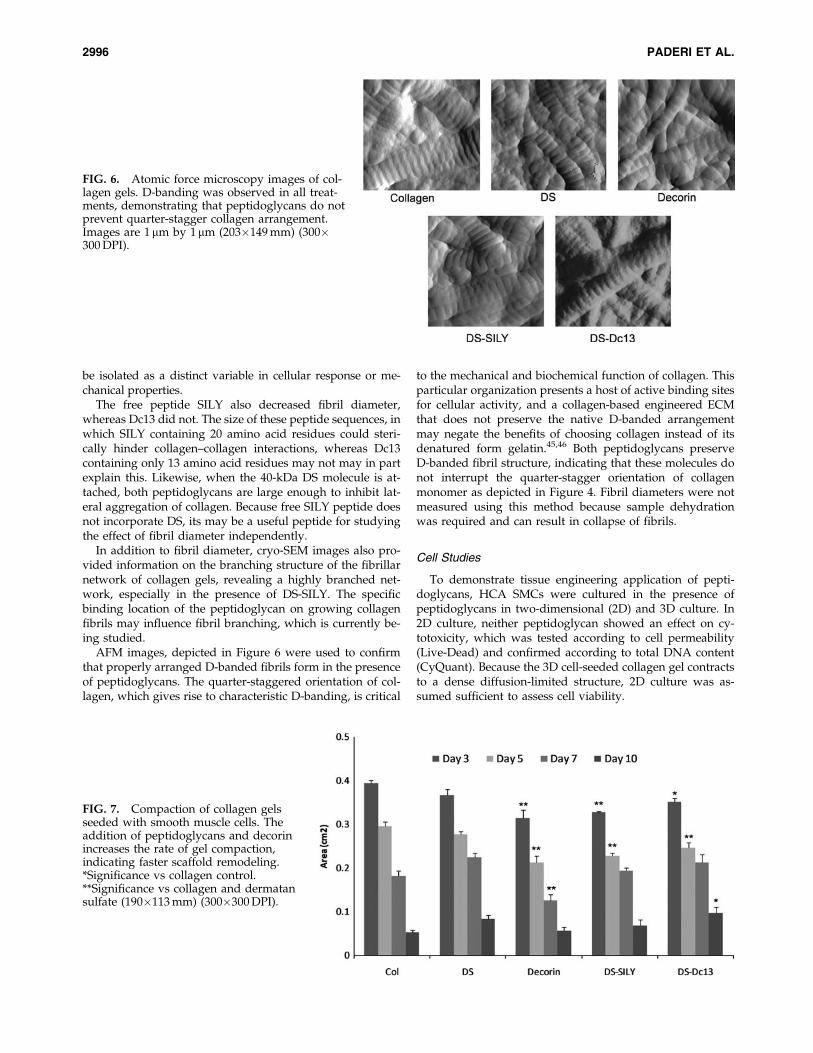
AFM images, depicted in Figure 6 were used to confirmthat properly arranged D-banded fibrils form in the presenceof peptidoglycans. The quarter-staggered orientation of col-lagen, which gives rise to characteristic D-banding, is critical
to the mechanical and biochemical function of collagen. Thisparticular organization presents a host of active binding sitesfor cellular activity, and a collagen-based engineered ECMthat does not preserve the native D-banded arrangementmay negate the benefits of choosing collagen instead of itsdenatured form gelatin.45,46 Both peptidoglycans preserveD-banded fibril structure, indicating that these molecules donot interrupt the quarter-stagger orientation of collagenmonomer as depicted in Figure 4. Fibril diameters were notmeasured using this method because sample dehydrationwas required and can result in collapse of fibrils.
Cell Studies
To demonstrate tissue engineering application of pepti-doglycans, HCA SMCs were cultured in the presence ofpeptidoglycans in two-dimensional (2D) and 3D culture. In2D culture, neither peptidoglycan showed an effect on cy-totoxicity, which was tested according to cell permeability(Live-Dead) and confirmed according to total DNA content(CyQuant). Because the 3D cell-seeded collagen gel contractsto a dense diffusion-limited structure, 2D culture was as-sumed sufficient to assess cell viability.
FIG. 6. Atomic force microscopy images of col-lagen gels. D-banding was observed in all treat-ments, demonstrating that peptidoglycans do notprevent quarter-stagger collagen arrangement.Images are 1 mm by 1 mm (203�149 mm) (300�300 DPI).
FIG. 7. Compaction of collagen gelsseeded with smooth muscle cells. Theaddition of peptidoglycans and decorinincreases the rate of gel compaction,indicating faster scaffold remodeling.*Significance vs collagen control.**Significance vs collagen and dermatansulfate (190�113 mm) (300�300 DPI).
2996 PADERI ET AL.

HCA SMCs seeded into 3D collagen gels with incorpo-rated peptidoglycans characteristically contracted the gel tovarying degrees, which correlated to fibril diameter at earlytime points, as shown in Figure 7. Gels with incorporatedpeptidoglycans or decorin, containing fibrils of smaller di-ameters, contracted faster, which a greater surface area onwhich cells can exert traction could explain. By day 10, thecompaction differences were largely lost, because the gels allhad substantially compacted from their original size, andsubtle differences in compaction were undetectable. Freepeptide controls were compared with collagen alone andshowed that, at day 5, SILY resulted in greater compactionthan collagen, but by day 10, both peptides resulted in less-compact gels than collagen alone (data not shown). Theimplications of a greater rate of compaction indicate a fasterrate of remodeling, which is necessary for restoring thefunction of tissue-engineered constructs.3
Degradation of collagen matrices by incorporation of pep-tidoglycans or decorin did not increase over the 2-week cul-ture time, indicating a lack of remodeling by breaking downof the existing collagen matrix (data not shown). No variationsin trends were found at each time point, so hydroxyprolinetotals for the 2-week culture time were used for analysis.
For tissue-engineered vessels, the synthesis of elastin,which is responsible for the elasticity of arteries, is critical.Incorporation of DS-SILY into HCA SMC–seeded collagengels increased elastin production more than all other treat-ments, as shown in Figure 8, indicating great promise for thisparticular peptidoglycan in collagen-based vascular con-structs. Although the increase over collagen with no additivewas significant, but modest, DS itself resulted in a decrease
in elastin, and DS-SILY–incorporated gels resulted in a moresubstantial increase than DS-incorporated gels. This suggeststhat not only chemical identity of the GAG, but also pre-sentation of the GAG in the form of a peptidoglycan is im-portant not only for biophysical function, but also forbiochemical function.
Conclusions
We have described here the synthesis of collagen-bindingpeptidoglycans that can be tailored with respect to the peptidesequence as well as the attached GAG chain. These peptido-glycans modulate collagen fibrillogenesis and decrease fibrildiameter in collagen gels similarly to the SLRP decorin. Char-acteristic D-banding is observed in collagen fibrils formed withand without peptidoglycans or decorin, and turbidity resultstogether with smaller fibril diameters suggest inhibited lateralaggregation of collagen molecules by the binding of peptido-glycans. We are further investigating this mechanism and thepossible role of binding location along the collagen molecule onmorphology of collagen fibrils.
The potential use of peptidoglycans for tissue engineeringapplications was demonstrated by culturing HCA SMCs withincollagen gel matrices formed with added peptidoglycans. Freepeptides and peptidoglycans did not exhibit cytotoxicity in 2Dculture studies. Cell-seeded collagen gels formed with addedpeptidoglycans resulted in a faster rate of compaction, and in-corporation of DS-SILY increased elastin synthesis. Furtherstudies for vascular tissue engineering, as well as other appli-cations for collagen-based tissue-engineered constructs withincorporated peptidoglycans, are being studied. Additionally,variant forms of peptidoglycans designed with different GAGidentities and peptide sequences are being explored as a bio-mimetic method of modulating collagen-based constructs.
Acknowledgments
This work was funded in part by the NIH (K25HL074968)and the National Science Foundation (CBET-0651643). Theauthors thank Debby Sherman at Purdue Life Sciences Mi-croscopy Facility for help with cryo-SEM images.
Disclosure Statement
No competing financial interests exist.
References
1. Cen, L., Liu, W., Cui, L., Zhang, W.J., Cao, Y.L. Collagentissue engineering: development of novel biomaterials andapplications. Pediatr Res 63, 492, 2008.
2. Couet, F., Rajan, N., Mantovani, D. Macromolecular bio-materials for scaffold-based vascular tissue engineering.Macromolec Biosci 7, 701, 2007.
3. Freed, L.E., Guilak, F., Guo, X.E., Gray, M.L., Tranquillo, R.,Holmes, J.W., Radisic, M., Sefton, M.V., Kaplan, D., Vunjak-Novakovic, G. Advanced tools for tissue engineering: Scafflds,bioreactors, and signaling. Tissue Eng 12, 3285, 2006.
4. Stegemann, J.P., Kaszuba, S.N., Rowe, S.L. Review: Ad-vances in vascular tissue engineering using protein-basedbiomaterials. Tissue Eng 13, 2601, 2007.
5. Barocas, V.H., Girton, T.S., Tranquillo, R.T. Engineeredalignment in media equivalents: Magntic prealignment andMandrel compaction. J Biomech Eng 120, 660, 1998.
FIG. 8. Elastin production according to Fastin assay. (A)The addition of peptidoglycan dermatan sulfate (DS)-SILYresulted in significantly greater elastin production than allother treatments. DS and DS-Dc13 resulted in significantlyless elastin production than collagen. (B) The addition of freepeptides did not significantly affect elastin production(101�114 mm) (300�300 DPI).
COLLAGEN-BINDING PEPTIDOGLYCANS 2997

6. Cheng, X., Gurkan, U.A., Dehen, C.J., Tate, M.P., Hillhouse,H.W., Simpson, G.J., Akkus, O. An electrochemical fabrica-tion process for the assembly of anisotropically orientedcollagen bundles. Biomaterials 29, 3278, 2008.
7. Pins, G.D., Christiansen, D.L., Patel, R., Silver, F.H. Self-assembly of collagen fibers. Influence of fibrillar alignmentand decorin on mechanical properties. Biophys J 73, 2164,1997.
8. Williams, B.R., Gelman, R.A., Poppke, D.C., Piez, K.A.Collagen fibril formation - optimal in vitro conditions andpreliminary kinetic results. J Biol Chem 253, 6578, 1978.
9. Birk, D.E., Nurminskaya, M.V., Zycband, E.I. Collagen fi-brillogenesis in-situ—fibril segments undergo postdeposi-tional modifications resulting in linear and lateral growthduring matrix development. Dev Dyn 202, 229, 1995.
10. Chakravarti, S., Magnuson, T., Lass, J.H., Jepsen, K.J., La-Mantia, C., Carroll, H. Lumican regulates collagen fibril as-sembly: skin fragility and corneal opacity in the absence oflumican. J Cell Biol 141, 1277, 1998.
11. Danielson, K.G., Baribault, H., Holmes, D.F., Graham, H.,Kadler, K.E., Iozzo, R.V. Targeted disruption of decorinleads to abnormal collagen fibril morphology and skin fra-gility. J Cell Biol 136, 729, 1997.
12. Graham, H.K., Holmes, D.F., Watson, R.B., Kadler, K.E.Identification of collagen fibril fusion during vertebratetendon morphogenesis. The process relies on unipolar fibrilsand is regulated by collagen-proteoglycan interaction. J MolBiol 295, 891, 2000.
13. Jepsen, K.J., Wu, F., Peragallo, J.H., Paul, J., Roberts, L.,Ezura, Y., Oldberg, A., Birk, D.E., Chakravarti, S. A syndromeof joint laxity and impaired tendon integrity in lumican- andfibromodulin-deficient mice. J Biol Chem 277, 35532, 2002.
14. Scott, J.E., Haigh, M., Identification of specific binding-sitesfor keratan sulfate proteoglycans and chondroitin dermatansulfate proteoglycans on collagen fibrils in cornea by the useof cupromeronic blue in critical-electrolyte-concentrationtechniques. Biochem J 253, 607, 1988.
15. Scott, J.E., Orford, C.R. Dermatan sulfate-rich proteoglycanassociates with rat tail tendon collagen at the D-band in thegap region. Biochem J 197, 213, 1981.
16. Svensson, L., Aszodi, A., Reinholt, F.P., Fassler, R., Heine-gard, D., Oldberg, A. Fibromodulin-null mice have abnor-mal collagen fibrils, tissue organization, and altered lumicandeposition in tendon. J Biol Chem 274, 9636, 1999.
17. Vogel, K.G., Trotter, J.A. The effect of proteoglycans on themorphology of collagen fibrils formed in vitro. Coll Relat Res7, 105, 1987.
18. Yung, S., Chan, T.M. Peritoneal proteoglycans: much morethan ground substance. Perit Dial Int 27, 375, 2007.
19. Iozzo, R.V. Matrix proteoglycans: from molecular design tocellular function. Annu Rev Biochem 67, 609, 1998.
20. Milan, A.M., Sugars, R.V., Embery, G., Waddington, R.J.Modulation of collagen fibrillogenesis by dentinal proteo-glycans. Calcif Tissue Int 76, 127, 2005.
21. Bierbaum, S., Douglas, T., Hanke, T., Scharnweber, D.,Tippelt, S., Monsees, T.K., Funk, R.H. W., Worch, H. Col-lageneous matrix coatings on titanium implants modifiedwith decorin and chondroitin sulfate: characterization andinfluence on osteoblastic cells. J Biomed Mater Res A 77, 551,2006.
22. Douglas, T., Heinemann, S., Bierbaum, S., Scharnweber, D.,Worch, H. Fibrillogenesis of collagen types I, II, and III withsmall leucine-rich proteoglycans decorin and biglycan. Bio-macromolecules 7, 2388, 2006.
23. Reed, C.C., Iozzo, R.V. The role of decorin in collagen fi-brillogenesis and skin homeostasis. Glycoconj J 19, 249, 2003.
24. Alberty, R.A. Recommendations for nomenclature and tablesin biochemical thermodynamics. Eur J Biochem 240, 1, 1996.
25. Scott, J.E. Proteodermatan and proteokeratan sulfate (dec-orin, lumican=fibromodulin) proteins are horseshoe shaped.Implications for their interactions with collagen. Biochem-istry 35, 8795, 1996.
26. Weber, I.T., Harrison, R.W., Iozzo, R.V. Model structure ofdecorin and implications for collagen fibrillogenesis. J BiolChem 271, 31767, 1996.
27. Nareyeck, G., Seidler, D.G., Troyer, D., Rauterberg, J.,Kresse, H., Schonherr, E. Differential interactions of decorinand decorin mutants with type I and type VI collagens. Eur JBiochem 271, 3389, 2004.
28. Paderi, J.E., Panitch, A. Design of a synthetic collagen-binding peptidoglycan that modulates collagen fibrillogen-esis. Biomacromolecules 9, 2562, 2008.
29. Ratcliffe, A. Tissue engineering of vascular grafts. MatrixBiol 19, 353, 2000.
30. Swartz, D.D., Russell, J.A., Andreadis, S.T. Engineering offibrin-based functional and implantable small-diameterblood vessels. Am J Physiol Heart Circ Physiol 288, H1451,2005.
31. Chupa, J.M., Foster, A.M., Sumner, S.R., Madihally, S.V.,Matthew, H.W. T. Vascular cell responses to polysaccharidematerials: in vitro and in vivo evaluations. Biomaterials 21,
2315, 2000.32. Pieper, J.S., Hafmans, T., Veerkamp, J.H., van Kuppevelt,
T.H. Development of tailor-made collagen-glycosaminoglycanmatrices: EDC=NHS crosslinking, and ultrastructural aspects.Biomaterials 21, 581, 2000.
33. Stuart, K., Panitch, A. Influence of chondroitin sulfate oncollagen gel structure and mechanical properties at physio-logically relevant levels. Biopolymers 89, 841, 2008.
34. Long, J.L., Tranquillo, R.T. Elastic fiber production in car-diovascular tissue-equivalents. Matrix Biol 22, 339, 2003.
35. Chiang, T.M., Kang, A.H. A synthetic peptide derived fromthe sequence of a type I collagen receptor inhibits typecollagen-mediated platelet aggregation. J Clin Invest 100,
2079, 1997.36. Kalamajski, S., Aspberg, A., Oldberg, A. The decorin se-
quence SYIRIADTNIT binds collagen type I. J Biol Chem282, 16062, 2007.
37. Scott, J.E, Supramolecular organization of extracellular-matrix glycosaminoglycans, in vitro and in the tissues. FASEBJ 6, 2639, 1992.
38. Obrink, B. Study of interactions between monomeric tro-pocollagen and glycosaminoglycans. Eur J Biochem 33, 387,1973.
39. Vogel, K.G., Paulsson, M., Heinegard, D. Specific-inhibitionof type-i and type-ii collagen fibrillogenesis by the smallproteoglycan of tendon. Biochem J 223, 587, 1984.
40. Neame, P.J., Kay, C.J., McQuillan, D.J., Beales, M.P., Hassell,J.R. Independent modulation of collagen fibrillogenesis bydecorin and lumican. Cell Mol Life Sci 57, 859, 2000.
41. Ruehland, C., Schonherr, E., Robenek, H., Hansen, U., Iozz,R.V., Bruckner, P., Seidler, D.G. The glycosaminoglycanchain of decorin plays an important role in collagen fibrilformation at the early stages of fibrillogenesis. FEBS J 274,
4246, 2007.42. Chapman, J.A. Staining pattern of collagen fibrils. 1. Ana-
lysis of electron micrographs. Connect Tissue Res 2, 137,1974.
2998 PADERI ET AL.

43. Obrink, B. Influence of glycosaminoglycans on formation offibers from monomeric tropocollagen in-vitro. Eur J Biochem34, 129, 1973.
44. Parry, D.A. D., Barnes, G.R. G., Craig, A.S. Comparison ofsize distribution of collagen fibrils in connective tissues as afunction of age and a possible relation between fibril sizedistribution and mechanical-properties. Proc R Soc LondonB Biol Sci 203, 305, 1978.
45. Di Lullo, G.A., Sweeney, S.M., Korkko, J., Ala-Kokko, L.,San Antonio, J.D. Mapping the ligand-binding sites anddisease-associated mutations on the most abundant pro-tein in the human, type I collagen. J Biol Chem 277, 4223,2002.
46. Zeugolis, D.I., Khew, S.T., Yew, E.S. Y., Ekaputra,A.K., Tong, Y.W., Yung, L.Y. L., Hutmacher, D.W., Shep-
pard, C., Raghunath, M. Electro-spinning of pure collagennano-fibres—just an expensive way to make gelatin? Bio-materials 29, 2293, 2008.
Address correspondence to:Alyssa Panitch, Ph.D.
Purdue University206 S. Martin Jischke Dr.West Lafayette, IN 47907
E-mail: [email protected]
Received: January 7, 2009Accepted: March 26, 2009
Online Publication Date: May 5, 2009
COLLAGEN-BINDING PEPTIDOGLYCANS 2999

Copyright © 2022 FDOKUMEN




















