
Protein Localization in Silica Nanospheres Derived via Biomimetic Mineralization
Upload
khangminh22Category
view
2download
0
BIOMIMETIC MEMBRANES AS NEW MATERIALS FOR APPLICATIONS IN ENVIRONMENTAL ENGINEERING AND BIOLOGY
BY
MANISH KUMAR
DISSERTATION
Submitted in partial fulfillment of the requirements
for the degree of Doctor of Philosophy in Environmental Engineering in Civil Engineering in the Graduate College of the
University of Illinois at Urbana-Champaign, 2010
Urbana, Illinois
Doctoral Committee:
Research Assistant Professor Julie L. Zilles, Chair Professor Mark M. Clark, Director of Dissertation Research Emeritus Professor Vernon L. Snoeyink Professor Wolfgang P. Meier, University of Basel

ii
ABSTRACT
Biological water channel proteins, called aquaporins, provide selective and rapid transport of
water across cell membranes. They utilize an elegant mechanism distinct from and more
efficient than that used in commercial solute separation polymeric membranes such as Reverse
Osmosis (RO) membranes. In this work, the bacterial Aquaporin (AqpZ) was functionally
incorporated into synthetic biomimetic polymer vesicles. Using stopped flow light scattering, the
permeability of such systems was determined to be up to two orders of magnitude higher than
current RO membranes, revealing the potential of this approach. A templating procedure was
then used to make flat membrane films with a high density of AqpZ. The sizes of these films are
small (~500 nm) and more research needs to be performed to scale up this process. However,
this method led to creation of flat 2D or thin 3D crystals of AqpZ in a polymer matrix as
confirmed by electron diffraction. This indicates that the packing efficiency of these polymer-
based systems is extremely high. Additionally, such crystals have the potential to allow for
structural reconstruction of the incorporated aquaporins. This procedure can thus provide
fundamental knowledge regarding the conformation of membrane proteins in block copolymers
and help in design of functional protein-polymer hybrid materials.
This work also led to the serendipitous discovery of AqpZ gating (reversible closure) at low pH
values when incorporated into triblock copolymer vesicles. This gating is also present in bacteria
and has relevance for bacterial survival under acid and osmotic shock. An overall scheme of
osmoregulation and acidic shock survival utilizing coordinated activation and gating of
membrane proteins is proposed.
Several research ideas resulted from this work and are currently being pursued. This includes
determination of insertion efficiency of membrane proteins in block copolymers, the use of block
copolymer membranes for studying gas transport in membrane proteins, block copolymer
vesicles with encapsulated perchlorate degrading enzymes for water treatment, and carbon
capture using active CO2 transporters inserted into block copolymer membranes.
Overall, this work has demonstrated the promise of using hybrid protein-polymer systems for
environmental engineering applications. In particular its applicability to synthetic desalination

iii
membranes is most promising and relevant. The basic approach used here may be applied to any
separation for which a specific transport protein is available or could be engineered. My work
has also contributed to understanding the properties of aquaporins, in particular AqpZ and its
possible role in microbial physiology. Finally, recent successes in immobilizing protein
molecules and in synthesizing 2D crystals of membrane proteins may provide an excellent way
to answer fundamental questions regarding the structure and function of these membrane
proteins in block copolymers.
In broad terms, this work has shown that biology provides excellent paradigms for engineering
materials and processes that are efficient and sustainable and that this “reverse engineering”
approach can enrich our understanding of underlying biological phenomena

iv
To Anu, without whom all this would not be possible.

v
ACKNOWLEDGMENTS
This work needed to be collaborative to achieve whatever modest level of success it has
achieved and I have many people to thank. First, I would like to thank my advisors Mark Clark
and Julie Zilles. Mark, for convincing me to come back to Illinois for my PhD and giving me the
independence required to start my ambitious project. Julie was an excellent advisor and helped
me navigate through my PhD with her steady advice and sympathetic ear. I know that I have
been very demanding of her time and attention and certainly, patience, many times; but she was
always there for me and supported me wholeheartedly. I did follow some crazy ideas and she let
me run through them logically and made sure I did not get distracted from the main goal of my
work. I cannot thank her enough and hope to be able to justify her faith in my abilities through
continued success.
Dr. Wolfgang Meier was a great collaborator, mentor, and committee member. I started
this project inspired by some of his work and he was very welcoming and supportive of me
learning techniques and working in his lab. The months I spent in his lab were probably the
most productive part of my PhD experience.
Dr. Snoeyink was a great committee member and mentor. He has helped me with great
professional advice over the years starting with when I first started graduate studies in 1998. He
was the reason for me to wait “at least 5 years in industry” before returning for a PhD. I think
this was geat advice and I think has served me well.
I would also like to thank my former group members David Ladner and Won-young Ahn
for contributing to my growth as a researcher and as a person. David taught me about careful
planning and independent thinking as well ass opening my eyes to what was possible for a
graduate student to do on his own. I admire his independent thinking and ability to get along
with people of all kinds while getting things done! Won-young helped me learn about the
promise of molecular modeling and how it could be applicable to my work.
My current group members, especially Michelle Marincel and Sean Poust have taught me
a lot about collaborating on a research group. I must have been a constant strain on them with
my incessant stream of advice on how to do things better and how to think independently. I hope
this was somehow helpful enough to them to counterbalance dealing with me on a daily basis.
Michelle has helped me think about how to design experiments and explain my ideas to make
them accessible. Sean has helped me learn how to think my ideas through with his independent

vi
ideas and ability and willingness to raise objections to my harebrained theories. Other members
of the Zilles research group have also taught me many lab techniques and helped me with ideas.
Dr. Meir’s research group has also helped me learn about block copolymers and their
uses. Mariusz Grzelakowski was my closest collaborator and has become a great friend. We can
hopefully collaborate on our slate of “unfinished ideas” in the future. Katarzyna Kita was a great
resource and friend and her frank and insightful comments helped me get work done efficiently
while I was in Basel. Fabian Itel visited our lab in summer 2008 and has been an ongoing
collaborator and friend since. His straight talk, and his ability to make me see his side of the
story while working on problems together has been a great learning experience. Dr. Andreas
Engel and his group in Basel got me started on 2D crystallization of membrane proteins and I am
thankful for their help and advice in this area over the last two years.
The EES faculty at Illinois, both current and past, are outstanding people in addition to
being great researchers and teachers. In particular I would like to mention Drs. Timm
Strathmann, Helen Nguyen, Eberhard Morgenroth, Kevin Finneran and Benito Marinas for their
help and advice over the course of my PhD.
I developed a great working relationships and friendships with EES students over the
years. In particular thanks to Marty Page for helping me out with UV experiments and for being
a great friend. The occupants (past and current) of 4125 deserve my gratitude for expanding my
knowledge and cultural training and for bearing with my habit of discussing ideas for longer than
necessary. These include Kim Milferstedt, Adrienne Menitti, Nick Weihardt, and Xinyu Zhang.
Derek Vardon worked in our lab, was an office mate, a neighbor and a great friend.
Don Cropek and Irene McAllister from CERL, Champaign helped me get the
experience of writing a successful proposal and have been great collaborators. Hopefully, we
can find a way of continuing our collaboration.
Shaoying Qi; our lab manager, has helped by being very supportive with equipment and
lab issues as well as being a great mentor and somebody who I have had great discussions about
what it means to be an engineer and a scientist.
Towards the end of my PhD I visited Dr. Tom Walz and his group and in working with
them I was able to figure some things out in weeks that I though would not be possible during
my PhD. I am thankful to them and hope that my postdoctoral work with them will be
productive.

vii
Steve Daley has been an excellent resource for budget issues and has really helped me
navigate these issues. His ability to find a solution to almost any budget situation is amazing.
Marvis Ozerk, the EES program coordinator helped with the many letters I had to obtain for
fellowships and such and also with other logistical issues. Sue Lowry was very helpful with
purchasing issues and Bobbi Vance helped with reimbursements.
The initial funding for this work was provided through a small but important “loan” from
the CEE department in the first year. A University Dissertation Travel grant provided the next
critical funding step for the summer following the first year for a very productive summer in Dr.
Meier’s group. Funding from the National Water Research Institute came at a critical time to
keep towards the end of this summer. Finally, fellowships from American Water Works
Association and Environmental Protection Agency towards the end of the second year allowed
support for stipends as well as travel. National Science Foundation funding starting at the
beginning of the third year for this project helped us equip our lab completely for making ptotein
polymer membranes and hiring a graduate student. US Arrmy CERL funded an additional
project on perchlorate removal that helped us expand the scope of the work that I had initiated
and helped hire another graduate student and a couple of undergraduate students.
Charlie Werth and my running/social group, the Wandering Elderbarries (especially
Shane, Peter, Barry, Joe, and Jack) were a source of much enjoyment, laughter, as well as sage
advice over the years. They made Saturday mornings a hoot and the trips to the relays we took
part in, memorable!
My parents, Virendra and Sushila Yadav, have instilled in me a love of learning and a
respect for teachers. Thank you for all that you have done for me. My dad was even more
excited than me at the prospect of me becoming a professor someday. Hopefully, I will be able
to justify your enthusiasm by being a good teacher. My brother, Chandra Prakash has always
encouraged me in my studies and I appreciate his advice over the years. My in-laws, especially
my mother-in-law, Annamma Mathew helped with the kids during the last year of my PhD. She
is an amazing woman and never seems to tire. I can only aspire to be as active as her at her age.
Anu, my dear wife, makes me complete; and was a major driving force for me striving to
do my best. She spent many months alone while I was traveling or just spending time in the lab.
Finally, she has given me the greatest gifts of my life, Arun and Sierra, our beautiful kids. I
thank her with all my heart.

viii
TABLE OF CONTENTS 1 INTRODUCTION AND OBJECTIVES................................................................... 1
1.1 WATER AND WASTEWATER TREATMENT MEMBRANES....................... 2
1.2 AQUAPORINS ................................................................................................... 12
1.3 BLOCK COPOLYMERS AS MIMICS OF BIOLOGICAL MEMBRANES.... 17
1.4 OBJECTIVES ..................................................................................................... 22
1.5 ORGANIZATION OF DISSERTATION........................................................... 23
2 HIGHLY PERMEABLE POLYMERIC MEMBRANES BASED ON THE
INCORPORATION OF THE FUNCTIONAL WATER CHANNEL PROTEIN
AQUAPORIN Z ........................................................................................................ 24
2.1 ABSTRACT ........................................................................................................ 24
2.2 INTRODUCTION............................................................................................... 25
2.3 RESULTS............................................................................................................ 27
2.4 DISCUSSION ..................................................................................................... 32
2.5 MATERIALS AND METHODS ........................................................................ 34
3 BIOLOGICAL ROLE FOR BACTERIAL AQUAPORIN GATING
DISCOVERED IN BLOCK COPOLYMER MEMBRANES .............................. 39
3.1 ABSTRACT ........................................................................................................ 39
3.2 INTRODUCTION............................................................................................... 39
3.3 RESULTS............................................................................................................ 42
3.4 DISCUSSION ..................................................................................................... 47
3.5 MATERIALS AND METHODS ........................................................................ 50
4 SELF-ASSEMBLY OF MEMBRANE PROTEINS IN BLOCK
POLYMERS.............................................................................................................. 53
4.1 ABSTRACT ........................................................................................................ 53
4.2 INTRODUCTION............................................................................................... 53
4.3 RESULTS............................................................................................................ 54

ix
4.4 DISCUSSION ..................................................................................................... 58
4.5 MATERIALS AND METHODS ........................................................................ 60
5 SUMMARY, IMPLICATIONS, AND FUTURE RESEARCH............................ 65
5.1 HIGH PERMEABILITY AQPZ RICH MEMBRANE CAN MAKE
IDEAL DESALINATION MEMBRANES AND HIGH SENSITIVITY
SENSORS. .................................................................................................................. 65
5.2 TRIBLOCK COPOLYMERS CAN BE USED AS A PLATFORM FOR
STUDYING MEMBRANE PROTEIN PROPERTIES.............................................. 69
5.3 BLOCK COPOLYMERS CAN BE USED FOR DETERMINING
PROTEIN STRUCTURE. .......................................................................................... 72
5.4 MEMBRANE VESICLES CAN BE USED FOR CONDUCTING
REACTIONS ON SURFACES, ENVIRONMENTAL REMEDIATION AND
CARBON CAPTURE................................................................................................. 76
5.5 CONCLUSION ................................................................................................... 80
6 REFERENCES.......................................................................................................... 81
APPENDIX A.................................................................................................................. 95
APPENDIX B .................................................................................................................. 98
APPENDIX C................................................................................................................ 101
AUTHOR’S BIOGRAPHY.......................................................................................... 109

1
1 INTRODUCTION AND OBJECTIVES
Synthetic membranes are important for environmental applications, particularly for water and
wastewater treatment. They have become more relevant as we are faced with a scarcity of fresh
water sources in many parts of the world. This water scarcity is further complicated by
contamination of existing freshwater supplies and the emergence of new, difficult-to-remove
pollutants (Koplin et al. 2002). Membrane desalination technologies, such as reverse osmosis
(RO), have come to the forefront as excellent technologies for developing new water sources
such as brackish surface and groundwater (Bohdziewicz et al. 1999), seawater (Matsura 2001),
and even recycled wastewater (del Pino and Durham 1999; Goldstein 2006; Judd 2006). RO is
also used for removal of several existing and emerging contaminants (Kimura et al. 2003;
Kimura et al. 2004). However, RO’s environmental footprint is coming under increasing
scrutiny, especially for seawater desalination (Einav et al. 2002). The pressure requirement for
RO is high, resulting in high energy consumption. This is a continuing hurdle to extensive
application of this technology. Additionally, some endocrine disrupting compounds (EDCs) and
pharmaceutically active compounds (PhACs) are not easily treated using RO (Kimura et al.
2004). With increasing concern regarding the impact of climate change on traditional water
supplies (Kundzewicz 2007), the development of alternate sources such as seawater and
reclaimed water is imperative. This has led to a focus on research into enabling technologies
such as RO in recent years.
There has been tremendous development in the efficacy and affordability of RO membranes in
the last three decades. However, limitations still exist on their selectivity, permeability and
energy efficiency. Cell membranes, on the other hand, can be considered ideal solute rejecting
membranes. Studying these membranes may help us improve existing RO membranes. They are
a composite filter consisting of exceptionally selective proteins known as membrane proteins that
transport water, specific ions (K+), or specific macromolecules. Cell membranes conduct
extremely efficient filtration while removing salts and other dissolved solutes. Furthermore, they
transport water at extremely high rates. Current high salt rejection RO membranes have a
permeability of around 3-4 x 10-12 ms-1Pa-1 (Ladner 2009), while membranes mimicking those
present in red blood cells or kidney tubules can reach ~ 4 x 10-11 ms-1Pa-1 (Walz et al. 1994).
This enhanced permeability is due to the presence of water channel proteins, Aquaporins, in

2
these membranes. Aquaporins have exceptional water transport properties, transporting as many
as three billion water molecules per second while excluding solutes (Agre 2004). They have
been found in species from all domains of life and are important for diverse physiological
functions involving water transport.
My work focused on developing a highly permeable membrane for desalination by
inserting Aquaporins into amphiphilic block copolymer membranes. These membranes are
proposed as alternatives to currently used synthetic polymeric RO membranes. As background
for this work, in this chapter I introduce water and wastewater membranes, RO membranes and
their structure, use and limitations during desalination, current efforts to improve RO
membranes, aquaporins, the relevance of important structural motifs and mechanism of transport,
amphiphilic block copolymers, and the concept of aquaporin-based block copolymer biomimetic
membranes.
1.1 WATER AND WASTEWATER TREATMENT MEMBRANES
Synthetic polymeric membranes are widely used for water and wastewater treatment in
environmental engineering. The most porous membranes have controlled pore sizes as large as
several microns. At the opposite end of the spectrum (nonporous membranes) there are no pores
per se, rather transport occurs between overlapping polymer strands. Separation by these
membranes is based primarily on solute size. Figure 1.1 illustrates the size range of membrane
separations, and the contaminants common in water and wastewater treatment.
As seen from Figure 1.1, these membranes are used for separation of dissolved and particulate
matter from water and wastewater. Low pressure membranes (microfiltration and ultrafiltration)
are used for separation of particulate matter while high pressure membranes (RO and
nanofiltration) are used for removal of dissolved materials such as ions and macromolecules.
Membrane treatment technologies pertinent to environmental engineering are summarized in
Table 1.1.

3
Figure 1.1: Membrane processes used in water and wastewater treatment. (Note: All images used in this figure are from the public domain and do not require copyright for use. Cryptosporidium and Giardia image is from www.epa.gov/microbes/cpt_gda.htm ; Viruses: rotovirus top image from http://en.wikipedia.org/wiki/Rotavirus and adenovirus bottom image is from http://en.wikipedia.org/wiki/Adenoviridae ; Humic acid structure is from http://en.wikipedia.org/wiki/Humic_acid)
Of the technologies presented in Table 1.1, high pressure membrane technologies have
the highest power needs due to the mechanism of water transport and the necessity of
overcoming the osmotic pressure created by rejection of soluble material, in particular, salts. I
will refer to high pressure membranes (both Nanofiltration (NF) and Reverse Osmosis (RO))
collectively as RO in this document. Typical operating pressures for low pressure membranes
are either up to ~ 1 bar vacuum (-14.5 psi) or between 0.3 and 2.1 bar pressures (~5 to 30 psi),
while for high pressure membranes operating pressures can range from 7 to 21 bar (~100 -300
psi) (for nanofiltration and brackish water reverse osmosis) to 55-70 bar (~800-1000 psi) for
seawater desalination. Consequently, energy consumption for RO facilities are a large fraction
(up to 60%) of the operating costs (Kumar et al. 2005). The relevance of RO membranes is
growing rapidly in coastal areas as well in inland areas with marginal water quality. Hence the

4
pressure to improve the energy efficiency and thus the sustainability of the RO process has
increased in recent years. New ideas are needed to move this important technology further.
Table 1.1: Applications of membrane treatment technologies
Application Separation Source Membrane Type Technology
Water Treatment Particulate
and microbial
contaminant
removal
Groundwater,
surface water,
seawater
Microfiltration
(MF) and
Ultrafiltration
(UF)
Low Pressure
Membranes
Water Treatment
and Reuse
Dissolved
contaminant
and salt
removal
Tertiary treated
wastewater,
groundwater,
surface water
Nanofiltration
(NF) and Reverse
Osmosis (RO)
High Pressure
Membranes
Wastewater
Treatment
Biomass
separation in
bioreactor
Raw and
primary treated
wastewater
MF/UF Membrane Bioreactors
(MBR)
Desalination Salt and
dissolved ion
removal
Brackish
surface and
groundwater,
seawater
Cation/Anion
Exchange
Membranes
Electrodialysis
(ED)/Electrodialysis
Reversal (EDR)
The following subsections describe the structure of current RO membranes, the current
stage of development and their inherent permeability limitation. This is followed by inspiration
for a high permeability membrane derived from biological membrane protein channels.
1.1.1 Reverse Osmosis Membranes: Structure
Current RO membranes are either asymmetric or thin film composites (Lonsdale 1987;
Ho and Sirkar 1992; Cheryan 1998). Asymmetric RO membranes have a thin selective skin
layer supported on a porous sublayer of the same polymer. Asymmetric membranes are
synthesized using the phase inversion process in which a polymer solution in solvent is
immersed in a non-solvent bath. This leads to precipitation of the polymer into a polymer-rich
solid phase that forms the membrane and a polymer-poor liquid phase that forms the membrane

5
pores or void spaces. Thin film composite (TFC) membranes, on the other hand, consist of a thin
polymer layer formed on one or more porous support layers. TFC membranes are made by
interfacial polymerization in which a porous polymer such as polysulfone is coated with a layer
of polymer or polymer components and then reacted using crosslinking agents. For both the
asymmetric and the TFC membranes, the surface layer determines the flux and separation. In
both cases the surface separation layer is a dense polymer layer about 100 nm thick. This has
implications on the energy efficiency for such membranes as described in the following sections.
1.1.2 Current state of development, limitations, and opportunities for improvement
There have been great improvements in the productivity and decrease in the cost of
synthetic membranes used for water and wastewater applications. Figure 1.2 presents a cost and
productivity index (cost per unit water production) for low pressure membranes and high
pressure membranes (specifically brackish water reverse osmosis (BWRO)) over the last few
decades. This index was calculated based on manufacturer-provided historical and projection
data. An explanation of this data is provided in Appendix A.
Figure 1.2: Cost reduction and productivity improvement in synthetic membranes used in water treatment. The indices present a cost/productivity value indexed to the first year data was available. Fits for UF/MF include two linear regions and for BWRO includes an exponential curve.
0
0.2
0.4
0.6
0.8
1
1.2
1989 1994 1999 2004Year
UF/MF Index
(a)
0
0.2
0.4
0.6
0.8
1
1.2
1980 1990 2000 2010
Year
BWRO Index
(b)

6
As discussed earlier, a critical need at present is to improve the efficiency of high
pressure membranes. Figure 1.2 (b) shows that the decrease in cost per unit productivity has
become relatively flat in recent years. Also researchers have pointed out that the permeability of
RO membranes has almost reached the thermodynamic limit of minimum energy required for
such separations (Zhu et al. 2009). Increases in productivity of such membranes are limited by
the mechanism of water transport through interfacially formed and crosslinked polymers. An
illustration of the mechanism of water transport in such polymer layers is provided in Figure 1.3.
The rate-controlling transport step is through the top interfacial thin film layer (shown in Figure
1.3 (a)). Figure 1.3 (b) shows the distribution of water within pores of the crosslinked structure
of this polymer. These pores are not connected in a static manner; hence RO membranes are
described as being nonporous. Water present in pores of such polymers diffuses through the
polymer layer when pores become interconnected by thermal motion of polymers (Kotelyanskii
et al. 1998; Kotelyanskii et al. 1999). This is called “jump” diffusion of water. Due to the
statistical nature of this phenomenon, it is inherently slow. This is particularly evident when the
scale of transport (~100 nm) is considered compared to the scale of each jump diffusion step (~
3Å) (Kotelyanskii et al. 1998). Also, water transport is related to the amount of crosslinking as
crosslinking reduces the possible connections between adjacent pores. Increasing amounts of
crosslinking also relates to higher salt rejection in polyamide RO membranes such as in high salt
rejection membranes (seawater RO membranes). This, in high rejection membranes the jump
diffusion rate is further decreased, leading to even lower transport rates.
Some natural (Aquaporin) and artificial (carbon nanotube) nanochannels transport water
more rapidly than existing RO membranes due to a fundamental difference in transport
mechanisms. Given the mechanistic constraints of RO water transport, it is instructive to look at
the mechanism of water transport in hydrophobic protein channels, in particular aquaporins and
their synthetic analogue, carbon nanotubes.

7
Figure 1.3. Water transport through RO membranes is limited by the slow “jump diffusion” of water (a) Scanning Electron Micrograph of a Thin Film Composite (TFC) RO membrane. The top separating layer has delaminated from the support in this micrograph. (b) A molecular model of water distribution in a small part of the separating layer and pathway for “jump diffusion of water” as described in (Kotelyanskii et al. 1998) (c) Distribution of pores and water including the crosslinked polymer structure. Note: (b) and (c) are used with permission from (Kalinichev 2007).
Water transport in narrow hydrophobic channels (<100 nm) occurs by frictionless
movement of water molecules in a single file across the channel walls (Figure 1.3 (a) and (b))
across small lengths (10-20 nm). This frictionless single file movement of water molecules
down such channels, under the effects of an osmotic or pressure gradient (Majumder et al. 2005;
Hinds 2007), results in an enhanced transport rate that could be used for high permeability RO
membranes as described in later chapters.

8
Figure 1.4: Water transport in narrow hydrophobic channels (a) and (b) are illustrations of water flow in Aquaporins and Carbon Nanotubes (CNTs) respectively. (c) and (d) are illustrations of the velocity profile in Poiseuille flow (parabolic) and in confined slip flow (blunted parabolic). An additional profile of classical Poiseuille flow is shown in (d) in light blue to indicate comparison to classical flow and show where zero velocity is in relation to the shown velocity profile.
The following discussion provides a simple illustrative description of water flow in smooth
hydrophobic channels such as aquaporins and CNTs. Here, a classical fluid dynamics
description is presented as a simple illustration of the major reason for flux enhancement seen in
such channels.
In confined narrow channels there is a deviation from the classical Poiseuille flow
leading to an apparent velocity, the slip velocity, at the channel wall. In order to account for slip
velocity the Poiseuille equation is modified as follows.
2 2
2
2( ) 1 ,
4sLR r P
u rR R z
where, u (r) is velocity of water at a radial distance r from the center,
R is the radius of the channel,

9
Ls is the slip length at the solid liquid interface,
μ is the viscosity of water, and
P
z
is the pressure drop across the channel length z
The slip length is a characteristic scale that represents the discontinuity of the velocity
profile at the channel wall. In the above equation, the slip length is given by (described in
(Thomas et al. 2009))
( )
,s
r R
u rL
dudr
where, du/dr is the velocity gradient at r.
For CNTs, the slip length has been empirically determined to depend on carbon
nanochannel diameter and is given by (as described in (Thomas et al. 2009))
, 3( ) ,s s
CL d L
d
Where, Ls,∞ = 30 nm, the slip length over a flat grapheme (single layer molecular carbon
sheet) sheet, and C = 352 nm (a fitting parameter reported in (Holt et al. 2006)), and d is
diameter of the channel.
The effect of slip in these narrow hydrophobic channels is that the parabolic shape of the
velocity profile seen in Poiseuille flow is “blunted” (Figure 1.4 (b) and (c)) with decrease in
radius. This is illustrated using numerical calculations of the flow profile in Figure 1.5 (a).
Additionally as the channels become narrower and the slip length increases the velocity increases
dramatically when compared to average velocity in Poiseuille flow as shown in Figure 1.5 (b).
The preceding discussion applies to the case where the size of these channels are beyond
the continuum limit (>1.6 nm) for water flow. This size is much larger than AqpZ channel
“pores” (~0.23 nm at the narrowest part) and slightly larger than current CNT membranes (~1.8
nm) pores. However a simple continuum discussion of transport in smooth hydrophobic
channels provides insight into the fundamental mechanism of transport enhancement. Transport
enhancement becomes more pronounced at non-continuum scale in such membranes. However,
at the smaller non-continuum scale the description is somewhat complicated by the structure of
water in confined geometries as well as by coordinated motion of water molecules and can only
be described using molecular dynamics simulations.

10
Figure 1.5. Comparison of velocity profiles in Poiseuille flow (R>200nm) to that in confined slip flow. (a) Velocity profiles normalized to the maximum velocity for different channel radii showing the change in profile from parabolic to blunt profile. (b) Velocity profiles across the channel where the y axis represents the ratio of the radial velocity, u(r), to the average Poiseuille velocity for different channel radii. Note the large increase in velocity (and hence flow) as the R value goes from >200 nm (2x avg. Poiseville velocity as expected in Poiseville flow) to 0.8 nm. (~580 x avg. Poiseville velocity). The above was calculated using the approach presented elsewhere (Thomas et al. 2009).
The efforts to develop high permeability RO membranes including membranes that utilize the
above approach are summarized in the following section.
1.1.3 Ongoing efforts for developing high permeability RO membranes
There are currently three main approaches being investigated for developing high
permeability RO membranes. These are: (1) thin film nanocomposite (TFN) membranes, (2)
carbon nanotube (CNT) membranes and (3) Aquaporin-based biomimetic membranes.
The thin film nanocomposite (TFN) membrane is a direct improvement on the current RO
membranes and improves water transport efficiency in the thin selective skin of TFC polyamide
RO membranes. This is achieved by adding zeolite nanoparticles during active layer formation.
Adding these nanoparticles is hypothesized to increase local order and structuring in these thin
selective layers (Jeong et al. 2007). In accordance with this hypothesis, TFN membranes have
increased smoothness, hydrophilicity and negative charge. These properties may explain the
approximately 100 to 200 % (up to two times) improvement in permeability (Jeong et al. 2007)

11
seen over conventional TFC RO membranes. Additionally, incorporating nanoparticles of
different compositions and at different amounts provides control over mechanical, transport and
surface properties of such membranes (Hoek et al. 2008; Lind et al. 2009). The membranes are
currently being commercialized for implementation as full scale membrane modules.
Performance data on these modules are currently unavailable.
CNT membranes rely on the use of aligned, single wall carbon nanotubes in an impermeable
matrix. Currently synthesized CNTs have diameters in the range of ~ 1.8 nm or higher. There
are two commonly used approaches to synthesizing aligned carbon nanotubes that may be
suitable for membrane applications (Noy et al. 2007). Holt et al have developed a low pressure
chemical vapor deposition process to encapsulate a vertically aligned array of CNTs in silicon
nitride (Holt et al. 2006). After encapsulation the ends of the CNTs are opened up using an
etching process. Another approach to synthesizing aligned CNTs (Kim et al. 2007) uses a
polymer matrix. CNTs are first functionalized with amine groups and then dispersed in the
solvent - tetrahydrofuran (THF). This suspension is filtered through a hydrophobic Poly Tetra
Fluoro Ethylene (PTFE) filter to align the CNTs. A polymer solution of polysulfone is spin
coated to make the support matrix. Molecular dynamics and initial reports have shown great
promise for desalination using narrow CNT membranes (<0.5 nm). However, currently
synthesized CNT membranes have the major drawback of not being able to remove small ions
such as sodium and chloride. This is due to the fundamental difficulty of making carbon
nanotubes smaller than the hydrated diameter of these ions (Na+ has a diameter of ~0.2 nm).
Attempts to make narrower CNTs and functionalization of the internal surface of these CNTs are
being considered as options to improve ion rejection. However, no current publications report
success in making salt rejecting CNTs.
While the work presented in this thesis resulted in the first publication on AQP-based
membranes, to my knowledge three other research groups are actively pursuing similar ideas.
Most of the other ideas regarding the use of Aquaporin-based membranes prior to 2010 are
presented in patents. A research group at the Technical University of Denmark (as a part of a
European Union project called Membaq and a company Aquaporin AS) has patented an AQP-
based water treatment approach that utilizes lipid membranes (Jensen and Keller 2006; Jensen
2007). The Aquaporin AS group has recently published work on the development of a
microporous support for painting lipid bilayers or PMOXA-PDMS-PMOXA based monolayers

12
where aquaporins insertion was demonstrated (Gonzalez-Perez et al. 2009; Vogel et al. 2009).
This support is manufactured by laser ablation of an Ethylene Tetra Fluoro Ethylene (ETFE)
polymer film. They have shown insertion of several membrane proteins. However functional
insertion of Aquaporins has not been demonstrated at present. Montemagno et al have proposed
using triblock copolymers for protein insertion and using vesicles embedded in a polymer matrix
for water treatment (Motemagno 2006). Montemagno et al have not published this work,
although the group has patented several schemes for use of membrane proteins embedded in
block copolymer membranes (Montemagno et al. 2007). Kaufmann et al have recently published
a method to reliably coat a NF membrane with a lipid bilayer by depositing a tuned mixture of
lipids on the surface (Kaufman et al. 2010). They have demonstrated that the lipid bilayer is
continuous and could be a platform for insertion of Aquaporins. However, insertion of
Aquaporins and permeability of such membranes has still not been demonstrated. The rest of
this dissertation will discuss my results and ongoing efforts to develop Aquaporin rich
membranes for developing desalination membranes, as well as related findings on fundamental
properties and other possible applications of membrane proteins in block copolymers. Currently,
work on AQP-membranes in the other groups is focused on achieving protein insertion and on
development of larger scale membranes in order to directly measure permeability. Additionally,
work is ongoing on developing an appropriate support for such membranes. AQP-based
membranes have exceptional promise for both high permeability and high selectivity. However
this technology is at an early stage.
1.2 AQUAPORINS
Aquaporins are a class of membrane-spanning proteins that primarily transport water
across the cell membrane (Agre 2004). Their existence had been hypothesized for many decades
due to observation of anomalously large fluxes across some biological membranes such as
erythrocyte plasma membranes (summarized in (Finkelstein 1987)). These fluxes could not be
solely explained by diffusion across the cell membrane, so the presence of water channel
proteins was proposed. Agre and coworkers first demonstrated the water channel activity of a 28
KDa erythrocyte membrane protein expressed in Xenopus laevis oocytes (Preston et al. 1992).
This was the human aquaporin subsequently named AQP1. Since then 10 other
human/mammalian aquaporins have been discovered (named AQP0 through AQP10) with

13
critical physiological functions (Agre and Kozono 2003). Additionally, aquaporins have been
found in plants, yeast, bacteria and archaea (Calamita et al. 1995; Maurel 1997; Bonhivers et al.
1998; Kozono et al. 2003). Most aquaporins have functions related to water transport but solute
transport (reviewed in (King et al. 2004)) as well as formation of intercellular junctions (AQP0,
AQP4) by aquaporins have also been reported (Engel et al. 2008).
Aquaporins have a unique structure that allows them to transport water while excluding all
other solutes including protons. This flow is in the direction of the osmotic gradient. They have
an hourglass architecture and “pore openings” of around 3 Å lined with mostly hydrophobic
amino acids. They have excellent solute rejection properties. The mechanisms responsible for
solute rejection are summarized in Figure 1.6 (de Groot and Grubmuller 2001; Kozono et al.
2002). Aquaporins have 6 membrane spanning domains that are composed of alpha helices
(Figure 1.7) connected by 5 loops (A through E). Two of these domains (loop B and E) dip
halfway into the center of the hydrophobic core of the lipids, forming a hour glass structure that
provides a path for water (Jung et al. 1994). Both these loops contain a universally conserved
asparagine-proline-alanine (NPA) motif where the loops fold into the center of the membrane.
These two sets of NPA motifs form a part of the selectivity mechanism of the Aquaporin
depicted in Figure 1.6. The presence of a charged arginine residue close to this region and
interacting with water molecules passing down the lumen (pore) of the aquaporin in a single file
is responsible for inhibiting proton transport across aquaporins. There are several explanations
for proton diffusion in water. For example the Grotthuss mechanism (Agmon 1995) suggests
transfer via hydrogen bonds and transient hydronium ions. Thus a single file of water molecules
would be expected to efficiently transport protons by acting as a proton wire. This is indeed the
case in channels such as Gramicidin A where such an unbroken proton wire is invoked (Pomes
and Roux 2002). However, due to the presence of the two sets of NPA residues and a conserved
Arginine residue (Arg 189 in AqpZ, Arg 185 in AQP1) in a defined configuration, every water
molecule passing through this region has to form a series of hydrogen bonds with the aquaporin
residues in a manner that leads to reorientation of the water molecule, thus breaking the proton
wire (Murata et al. 2000). As first seen from molecular dynamics simulations and eventually
experimentally confirmed by X-ray crystallography, the water molecules face different directions
in the two halves of the protein (de Groot and Grubmuller 2001; Tajkhorshid et al. 2002).

14
Aquaporin exists as a tetramer in its native state; however each monomer is functional on its
own.
Figure 1.6: Mechanisms for rejection of solutes and transport of water through aquaporins illustrated using AQP1 as a model. The amino acid residues important for these rejection mechanisms are highlighted. Histidine 180 (His 180) and Arginine 185 (Arg 185) form the narrowest part of the channel (2.8 Å) and this size restriction responsible for size exclusion of larger solutes. Arg 185 is positively charged and the presence of this residue at the narrowest part of the channel helps reject positively charged solutes by electrostatic repulsion. Water molecules undergo a molecular reorientation by hydrogen bonding with a residue near the pore (Murata et al. 2000; de Groot and Grubmuller 2001; Chakrabarti et al. 2004). This molecular reorientation (water dipole reorientation) is responsible for preventing passage of H+ ions by breaking extensive hydrogen bonding that normally allows rapid H+ transfer in bulk water. The above illustration is based on Kozono at al (Kozono et al. 2002) and Jung et al (Jung et al. 1994) and was recreated using an example simulation (trajectory file from a molecular dynamics simulation of water passage through AQP1 (Grabmueller 2010)).

15
Figure 1.7: Predicted topology and sequence of AqpZ (a) based on (Borgnia et al. 1999). The transmembrane domains are shown as tubes and are numbered 1 through 6 starting from the N-terminus. The loops connecting these domains are shown as ribbons and numbered A through E. The highly conserved NPA residues are shown on loops B and E. It can be seen that this structure is symmetrical around loop C. (b) shows the folding of this polypeptide chain into a channel with the intracellular loops orienting parallel to each other in one of two tilt angles. (c) and (d) are side and top views of AqpZ monomer shown in ribbon representation.
The bacterial Aquaporin, Aquaporin Z (AqpZ) was selected for this study because of the
ease of laboratory expression and purification of this protein in its native host, Escherichia coli.
Additionally, this protein has been reported to be robust under various solution conditions and
active upon reconstitution into lipid vesicles (Borgnia et al. 1999). It also has one of the highest
water permeabilities of aquaporins that have been tested (Hashido et al. 2005). The sequence

16
and predicted topology (structure and layout in the membrane) (based on (Borgnia et al. 1999))
as well as the determined structure of AqpZ are shown in Figure 1.7.
As can been seen from the placement of the six transmembrane domains of Aquaporin
and its location within the membrane, the lipid layer is required to stabilize the structure of the
protein. The hydrophobic interactions between the transmembrane parts of the protein and the
hydrophobic core of the lipid bilayer are important for correct orientation and stability of
aquaporins. This is illustrated in Figure 1.8. All membrane proteins have hydrophobic
transmembrane regions, usually in the form of a band on its outer surface. In the absence of a
hydrophobic “cushion” around the protein, interactions between proteins lead to their
precipitation and inactivation. These cushions most commonly are made from lipids. During
purification of membrane proteins these lipid cushions are replaced by appropriate amphiphilic
detergent molecules. In order to utilize the function of these proteins they have to be placed in a
lipid bilayer-like environment. Such a lipid-like environment is provided in this work by
utilizing block copolymers that self assemble into structures that allow for membrane protein
insertion.
Figure 1.8: Schematic of the lipid bilayer with inserted channel protein (transmembrane protein) (a) shows the insertion of the channel into the lipid bilayer with the pink region representing the hydrophobic surface of the protein that interacts with the hydrophobic core of the membrane bilayer formed from the tails of the lipid molecules illustrated in (b). (c) depicts the hydrophobic residues on the AqpZ monomer (in surface representation) in pink as an illustration of the hydrophobic band in membrane proteins.

17
1.3 BLOCK COPOLYMERS AS MIMICS OF BIOLOGICAL MEMBRANES
Amphiphilic block copolymers can self assemble into structures mimicking lipid bilayers in
aqueous solutions. Such block copolymers are synthesized as diblock co-polymers with one
hydrophilic block and one hydrophobic block linked together by a covalent bond or triblock co-
polymers with two hydrophilic end blocks and a hydrophobic middle block. These block
copolymers assemble into a variety of structures in aqueous solutions, including micelles, rods
and vesicles depending on the amount of water present (Discher and Eisenberg 2002). The
morphology seen in such systems is a result of hydrophobic interactions between the polymer
molecules and the shape of individual polymer molecules; this is dictated by the ratio of the
hydrophilic to hydrophobic block as well as their individual conformations (Zhang and
Eisenberg 1995).
1.3.1 Amphiphilic block copolymers
Block copolymers can be designed to have hydrophobic and hydrophilic blocks. In a solvent
selective for one of these blocks, polar solvents for the hydrophilic block and nonpolar solvent
for the hydrophobic block, one of the blocks interacts favorably with the solvent while the other
block is shielded leading to a phase separation at the scale of the hydrophobic or hydrophilic
blocks. This phase separation occurs while being constrained by the covalent bond between the
blocks and in some cases leads to formation of monolayers or bilayers that mimic the structure of
the lipid bilayer. These amphiphilic molecules aggregate into structures such as vesicles and
form an alternative platform for insertion of membrane proteins. These protein-polymer
aggregates can then be utilized for studying membrane protein properties and for making new
engineering materials. The following paragraphs and subsections provide a brief introduction to
block copolymer properties and summarize available literature on insertion of membrane
proteins in block copolymers.
Amphiphilic block copolymers were used in this study as biomimetic membranes. They
provide several advantages over artificial and synthetic lipids in engineering applications. These
include:
i) High mechanical and chemical stability when compared to lipid membranes. Lipids have
the following disadvantages when used for applications (reviewed in (James C-M. Lee
2001)). Lipid membranes cannot be stretched beyond 5% of their original area without

18
rupture under osmotic or other stresses (Needham and Zhelev 1996). Because of
susceptibility to thermal phase transitions the permeability of lipid membranes is highly
variable, leading to solute leakage (Needham and Zhelev 1996). Block copolymer
membranes, on the other hand, perform much better than lipid membranes in terms of rupture
strength (toughness) (Discher et al. 1999) and thermal stability (James C-M. Lee 2001). The
cohesive energy density or toughness for polyethylethylene-polyethyleneoxide (EE-EO)
block copolymer membranes are 5-50 times higher than lipid membranes (Discher et al.
1999). Liposomes can also be strongly affected by pH, ionic strength, and temperature (Grit
and Crommelin 1993), complicating studies that need to be conducted under these conditions.
In this work, I show that the PMOXA-PDMS-PMOXA block copolymer vesicles are
insensitive to extreme pH values (between 2 and 12) and high ionic strengths (of up to 1 M).
They could also be engineered to be sensitive (Sauer et al. 2001) to pH changes.
ii) Low permeability for water (James C-M. Lee 2001; Kumar et al. 2007) and gases (ongoing
work, see Summary and Conclusions section) while being impermeable to charged species.
The structure and function of membrane transport proteins, including aquaporins, are typically
studied via reconstitution in a lipid matrix. However, lipids have high permeability for water
and gases, making it difficult to study transport of these molecules (Verkman 2002; Hill et al.
2004). The low permeability of block copolymers enhances the difference in transport
behavior between the membranes with and without transport proteins and allows sensitive
measurement of transport rates of the inserted transport proteins.
iii) Customizable properties. They can be engineered to provide the desired properties such as
fluidity or morphology by the choice of blocks and block lengths (Discher and Eisenberg
2002) or their length ratios (Zhang and Eisenberg 1995). Additionally, the choice of blocks
can provide a range of properties to these membranes. A highly relevant example is the use of
PMOXA as the hydrophilic block which provides the possibility of adding antifouling
properties to proposed desalination membranes and other applications (Konradi et al. 2008)
where protein deposition is undesirable.
iii) Ability to perform end group functionalization. Their end groups can be modified (Napoli
2008) by molecules such as biotin (for recognition and immobilization) (Grzelakowski et al.
2009), methacrylate (for stabilization by crosslinking) (Nardin et al. 2000), fluorescent

19
molecules (for imaging) (Grzelakowski et al. 2009) and even drugs (for drug delivery) (Peer
et al. 2007).
The amphiphlic block copolymers used in this work were poly-(2-methyloxazoline)-
poly-(dimethylsiloxane)- poly-(2-methyloxazoline) (PMOXA-PDMS-PMOXA) based
triblock polymers (Nardin et al. 2000). The general structure of this polymer is shown in
Figure 1.9.
Figure 1.9: Structure of the PMOXA-PDMS-PMOXA polymers used in this study
1.3.2 Membrane protein insertion into block copolymer membranes
Self assembled block copolymers in aqueous solutions, in particular the PMOXA-PDMS-
PMOXA system, have been studied over the last decade with a focus on insertion of membrane
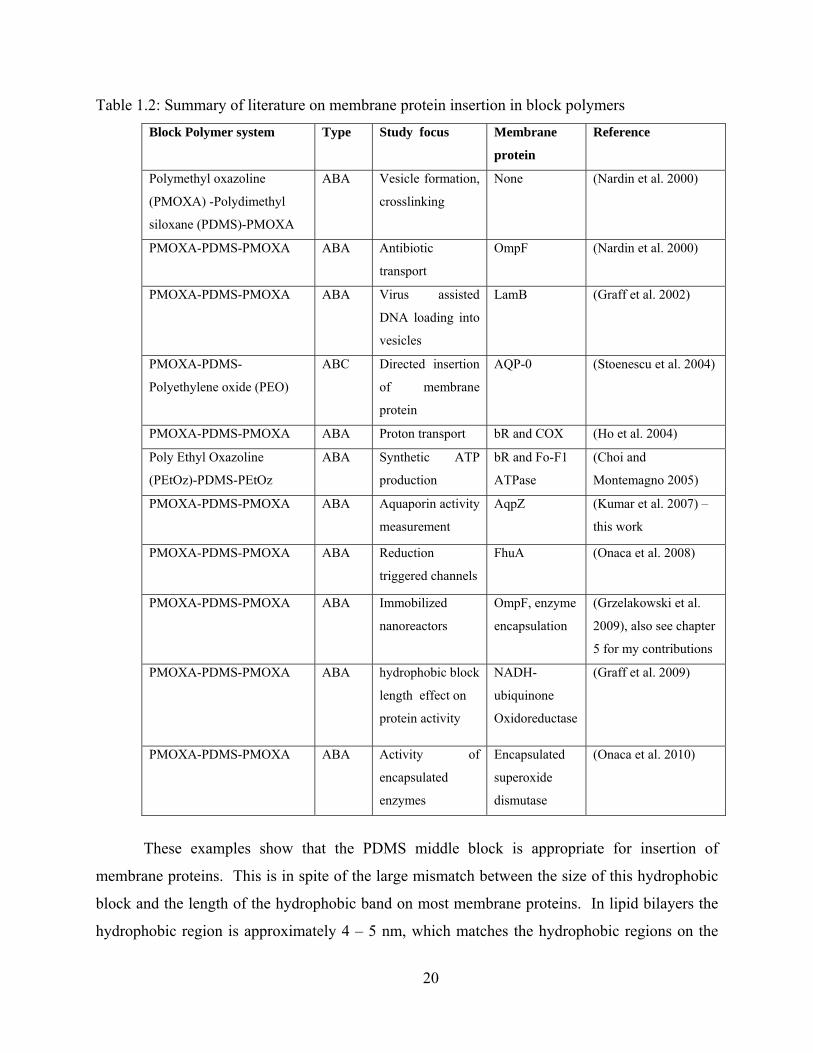
proteins and realization of biochemical reactions within vesicle. Table 1.2 presents a summary
of the studies conducted with block copolymer systems and membrane proteins.
All the reported studies shown in Table 1.2 have used triblock copolymers with the same middle
block – Polydimethylsiloxane (PDMS). All the studies discussed below are referenced in the
above table. The most common hydrophilic block was polymethyl oxazoline (PMOXA).
Polyethyl oxazoline (PEtOz) was used in one study while another study used Polyethylene oxide
(PEO) as one of the hydrophilic blocks. The inserted proteins include the bacterial outer
membrane proteins OmpF, LamB, and FhuA, the bacterial water channel protein AqpZ, the
bovine eye lens aquaporin (AQP0), the purple membrane H+ pump bacteriorhodopsin (bR), the
cytochrome oxidase (COX), the bacterial Fo-F1 ATPase and the NADH-ubiquinone
Oxidoreductase. Of the above, all proteins (except AQP0) were shown to be active in this block
copolymer.
N NSi
O OSi Si
n OOHHO
O m m
Polydimethylsiloxane Poly‐2‐methyl‐2‐oxazoline Poly‐2‐methyl‐2‐oxazoline
Hydrophilic
Hydrophobic
20
Table 1.2: Summary of literature on membrane protein insertion in block polymers
These examples show that the PDMS middle block is appropriate for insertion of
membrane proteins. This is in spite of the large mismatch between the size of this hydrophobic
block and the length of the hydrophobic band on most membrane proteins. In lipid bilayers the
hydrophobic region is approximately 4 – 5 nm, which matches the hydrophobic regions on the
Block Polymer system Type Study focus Membrane
protein
Reference
Polymethyl oxazoline
(PMOXA) -Polydimethyl
siloxane (PDMS)-PMOXA
ABA Vesicle formation,
crosslinking
None (Nardin et al. 2000)
PMOXA-PDMS-PMOXA ABA Antibiotic
transport
OmpF (Nardin et al. 2000)
PMOXA-PDMS-PMOXA ABA Virus assisted
DNA loading into
vesicles
LamB (Graff et al. 2002)
PMOXA-PDMS-
Polyethylene oxide (PEO)
ABC Directed insertion
of membrane
protein
AQP-0 (Stoenescu et al. 2004)
PMOXA-PDMS-PMOXA ABA Proton transport bR and COX (Ho et al. 2004)
Poly Ethyl Oxazoline
(PEtOz)-PDMS-PEtOz
ABA Synthetic ATP
production
bR and Fo-F1
ATPase
(Choi and
Montemagno 2005)
PMOXA-PDMS-PMOXA ABA Aquaporin activity
measurement
AqpZ (Kumar et al. 2007) –
this work
PMOXA-PDMS-PMOXA ABA Reduction
triggered channels
FhuA (Onaca et al. 2008)
PMOXA-PDMS-PMOXA ABA Immobilized
nanoreactors
OmpF, enzyme
encapsulation
(Grzelakowski et al.
2009), also see chapter
5 for my contributions
PMOXA-PDMS-PMOXA ABA hydrophobic block
length effect on
protein activity
NADH-
ubiquinone
Oxidoreductase
(Graff et al. 2009)
PMOXA-PDMS-PMOXA ABA Activity of
encapsulated
enzymes
Encapsulated
superoxide
dismutase
(Onaca et al. 2010)

21
outside of most membrane proteins (Figure 1.8). A large hydrophobic mismatch between these
two regions might lead to an energetic penalty and limit the stability of such self assembled
systems. At first glance, such seems to be the case in the block copolymer membranes, where
the hydrophobic region is around 10 nm. However, Pata and Dan (Pata and Dan 2003) have
suggested that the flexibility of the hydrophobic block as well as its polydispersivity could lead
to regions immediately adjacent to the membrane proteins matching the hydrophobic band of the
membrane protein. This is illustrated in Figure 1.10(a). PDMS is a highly flexible molecule
and may be subject to such a phenomenon. Figure 1.10(b) presents a conceptual model of how
this principle would result in a block copolymer membrane with inserted aquaporins. Regardless
of the mechanism of stable aquaporin insertion in these membranes, the measured activity
indicates that we have obtained functional incorporation.
Figure 1.10: Possible mechanism of insertion of membrane proteins in thick block copolymers. (a) and (b) Flexible middle blocks are usually highly stretched and could shrink up to half their length to accommodate membrane proteins with sizes that are half the length of the hydrophobic block without excessive energetic penalties (Pata and Dan 2003). (c) A conceptual model for a PMOXA-PDMS-PMOXA membrane with inserted membrane proteins based on the above

22
mechanism, from (Mecke et al. 2006) (http://dx.doi.org/10.1039/b605165k), Reproduced by permission of The Royal Society of Chemistry.
As demonstrated in my work, the functional insertion of AqpZ in block copolymers, the
stability of block copolymers, and the high permeabilities and selectivity expected suggest AqpZ
molecules inserted into PMOXA-PDMS-PMOXA block copolymers may be an excellent starting
point for development of high efficiency biomimetic desalination membranes.
1.4 OBJECTIVES
The main hypothesis of this work is that efficient desalination membrane can be developed
by insertion of the bacterial aquaporin, AqpZ into PMOXA-PDMS-PMOXA block copolymers.
The specific objectives and a brief summary of the work conducted under each objective are
provided below.
o Permeability measurements of ABA type PMOXA-PDMS-PMOXA block copolymer
vesicles with and without AqpZ to demonstrate enhanced permeability of membranes with
incorporated AqpZ (Chapter 2). This chapter discusses proof of principle for using AqpZ
inserted in block copolymers as desalination membranes. Here I show that up to 3000
times increase in permeability can be obtained by inserting Aquaporins in block
copolymers while maintaining selectivity. The permeabilities measured for these AqpZ-
ABA membranes were an order of magnitude or more above that current commercial
membranes.
o Determination of pH behavior of such AqpZ-ABA membranes (Chapter 3). In this chapter
the discovery of low pH-induced gating of bacterial aquaporin is reported. In particular,
the use of block copolymer enabled the measurement of permeability of AqpZ over a
wide pH range. This is in contrast to lipid membranes commonly used for such studies,
which are unstable at low pH values. This work demonstrates the presence of aquaporin
gating in bacteria and discusses its physiological significance.
o Development of a flat Aquaporin rich membrane by using an interfacial templating
monolayer (Chapter 4). In this chapter a templating polymer monolayer was used to
arrange histidine-tagged aquaporins. This was further used to assemble AqpZ-rich
triblock copolymer films under this template. I was successful in making small, protein-

23
packed films that are crystalline and may have relevance to protein structure
determination in block copolymers. I also observed what may be 2D crystallization of
the PMOXA-PDMS-PMOXA polymer alone or in a unique manner (distinct from
crystallization in lipids) with AqpZ. This has not been reported before for any polymer
using the detergent removal method employed here. These crystals need to be studied
further but may be microporous materials with unique properties.
1.5 ORGANIZATION OF DISSERTATION
This dissertation is organized into an introduction, followed by papers on three specific areas of
membrane proteins in block copolymers (Chapters 2 through 4 introduced above). The final
section summarizes my dissertation work with focus on a) broader implications of the conducted
work, b) related projects that were inspired by and initiated during the course of this work and to
which I contributed, and c) proposed future work that could be conducted to further develop the
ideas developed in this dissertation and associated projects. The appendix contains additional
information and details on an approach to characterize crosslinking in methacrylated PMOXA-
PDMS-PMOXA polymers to determine their stability.

24
2 HIGHLY PERMEABLE POLYMERIC MEMBRANES BASED ON THE
INCORPORATION OF THE FUNCTIONAL WATER CHANNEL PROTEIN
AQUAPORIN Z
Ackowledgements: I performed all the experiments described in this chapter except for
PMOXA-PDMS-PMOXA polymer synthesis, transformation of the ptrcHis10AqpZ plasmid into
E.coli and cryo-electron microscopy.
The results described in this chapter have been published. The citation is as follows: Kumar, M;
Grzelakowski, M; Zilles, J; Clark, M; Meier, W. 2007. Highly permeable polymeric membranes
based on the incorporation of the functional water channel protein Aquaporin Z, Proceedings of
the National Academy of Sciences. 2007;104(52):20719 – 20724. All figures, tables and text are
reproduced from this published work. Copyright (2007) National Academy of Sciences, U.S.A.
Explicit permission is not required as I am author of this work.
2.1 ABSTRACT
The effects of incorporation of the bacterial water channel protein, Aquaporin Z (AqpZ) on
permeability and solute transport characteristics of highly impermeable amphiphilic triblock
polymer vesicles were investigated. These vesicles were made of a novel polymer with
symmetric poly-(2-methyloxazoline)-poly (dimethylsiloxane)-poly (2-methyloxazoline)
(PMOXA15-PDMS110-PMOXA15) repeat units. Permeability measurements were conducted by
measuring light scattering of the polymer vesicles placed under an outwardly directed osmotic
gradient of salt in a stopped flow apparatus. These polymer vesicles were found to be highly
impermeable. However, a large enhancement in water productivity (permeability per unit
driving force) of up to ~800 times that of pure polymer was observed when AqpZ was
incorporated. The activation energy (Ea) of water transport for the protein-polymer vesicles (3.4
kcal/mol) corresponded to that reported for water-channel-mediated water transport. The solute
reflection coefficients of glucose, glycerol, salt and urea were also calculated. The productivity
values obtained for AqpZ-incorporated polymers were an order of magnitude or larger than for
existing salt-rejecting polymeric membranes. This approach represents a exciting new direction
for developing more efficient water treatment membranes for municipal and medical

25
applications. The distinct levels of permeability obtained by using different concentrations of
AqpZ may provide a key property for drug delivery applications.
2.2 INTRODUCTION
Biological membranes have excellent water transport characteristics, with certain membranes
able to control permeability over a wide range. Membranes such as those present in the proximal
tubules of the human kidney (Knepper et al. 1996) be induced to insert specific water channel
membrane proteins known as Aquaporins (AQPs) to increase permeability. Other biological
membranes, such as those in mammalian optic lenses (Gorin et al. 1984), erythrocytes (Preston
et al. 1992), and many other cell membranes(Nielsen et al. 1993), are constitutively AQP-rich.
The permeabilities observed in AQP-rich membranes are orders of magnitude higher than that
observed for unmodified phospholipid membranes (Borgnia et al. 1999). Additionally, some
members of the AQP family have excellent solute retention capabilities for very small solutes
such as urea, glycerol and glucose even at high water transport rates (Meinild et al. 1998;
Borgnia et al. 1999). These properties result from the unique structure of the water-selective
AQPs. AQPs have six membrane-spanning domains and a unique hourglass structure (Jung et
al. 1994) with conserved charged residues that form a pore that allows the selective transport of
water while rejecting solutes. The AQP used in this study was a bacterial Aquaporin from
Escherichia coli – Aquaporin Z (AqpZ). AqpZ was selected as it has been shown to enhance
the permeability of lipid vesicles by an order of magnitude (Borgnia et al. 1999)while retaining
small uncharged solutes(Borgnia et al. 1999). Additionally, AqpZ can be expressed in relatively
large quantities in E. coli and has been reported to be quite stable under different reducing
conditions (Borgnia et al. 1999) and at temperatures of 4 ° C for extended periods of time-
properties that make it attractive for applications in drug delivery and water treatment.
The AQP’s high permeability and high specificity could be very valuable for a variety of
applications. High permeabilities and excellent solute retention of small solutes are important
for water treatment in critical medical applications such as dialysis (Pastan and Bailey 1998;
Pontoriero et al. 2003) as it could lead to reduced equipment size and more efficient use of
energy. A significant improvement in the permeability of solute rejecting membranes would be
a large step in improving the economics of desalination for municipal applications as well.
Desalination is becoming increasingly important for water production in semi-arid coastal

26
regions as well as for wastewater recycling (Service 2006; Tal 2006). Reverse Osmosis (RO)
membranes are most commonly used for this application and the use of Forward Osmosis (FO)
membranes is creating substantial interest (Patel-Predd 2006; Service 2006). However, the use
of RO requires large consumption of energy while FO applications need large membrane areas.
This is due to the low productivity of currently used commercial RO and FO membranes.
Finally, the ability to produce membranes that can be designed to have low initial permeabilities
is desirable for drug delivery applications (Discher et al. 1999); however, a high level of control
over this permeability could be an additional advantage (Dinsmore et al. 2002).
The effects of AQPs on the permeability of biological and synthetic lipid membranes has been
studied by incorporating these proteins into liposomes (Borgnia et al. 1999), frog oocytes
(Preston et al. 1992), and cellular secretory vesicles (Coury et al. 1996). However, the direct use
of biological membranes or synthesized lipid membranes for water treatment and drug delivery
applications has practical disadvantages. The major limitation is the low stability of lipid
membranes (Duncan 2003). Obtaining and processing large volumes of such membranes would
also present technical challenges.
The use of synthetic polymers, in particular lipid-bilayer-like amphiphilic block copolymers, is
exciting in this regard, as they could provide the necessary chemical and mechanical stability
while still providing a amphiphilic structure that allows incorporation and function of membrane
proteins such as AQPs (Taubert et al. 2004). Some membrane proteins have been functionally
inserted into block copolymer membranes (Meier et al. 2000; Graff et al. 2002; Choi and
Montemagno 2005; Choi et al. 2006). These include the beta barrel OmpF protein, the bacterial
receptor protein LamB and bacteriorhodopsin. Directed insertion of the eye lens AQP-
Aquaporin 0 (Aqp-0) into similar block copolymer membranes has been also been demonstrated
(Stoenescu et al. 2004), although this initial study did not examine function. Additionally,
studies on the permeability of block copolymer membranes are few dd and no previous studies
have reported on the magnitude of improvement in permeability that could be obtained in block
copolymer membranes by insertion of AQPs. Data on rejection of dissolved ionic solutes such
as salt (NaCl) has also not been reported before for AQP-rich membranes.
To investigate the potential of protein polymer membranes for water treatment applications,
solute and water transport properties as well as physical characteristics were analyzed for a
specific protein-polymer membrane. This membrane was composed of a novel triblock

27
copolymer with inserted AqpZ and is referred to throughout the text as an AqpZ-ABA
membrane. The composition of the symmetric triblock copolymer was PMOXA15-PDMS110-
PMOXA15. This new ABA polymer has a large hydrophobic block (110 PDMS groups) that is
expected to result in more mechanical stability and low permeability. The physical
characteristics of the ABA triblock copolymer were studied using microscopy techniques, while
light scattering was used to characterize the permeability.
2.3 RESULTS
2.3.1 Synthesis of AqpZ-ABA polymer membranes
Histidine-tagged AqpZ was overexpressed in Escherichia coli and purified using nickel affinity
chromatography (Sulkowski 1985; Borgnia et al. 1999). A large yield of pure protein (between
2.5 and 15 mg/L of culture) was obtained for three different purification runs, indicating the
potential for large yields using the procedures described. PMOXAm-PDMSn-PMOXAm (ABA)
triblock copolymers were synthesized using a ring opening cationic polymerization procedure.
Polymer vesicles were produced using the film rehydration method described in detail in the
methods section.
2.3.2 Characterization of Polymer Vesicles
To determine the permeability and solute rejection properties of polymer vesicles using stopped
flow spectroscopy a knowledge of the physical dimensions (radius in hydrated state) and
morphology (hollow versus solid sphere structure) is necessary. ABA polymer vesicles were
therefore characterized using static and dynamic light scattering, Transmission Electron
Microscopy (TEM), cryo-TEM, and Atomic Force Microscopy (AFM).
Using dynamic light scattering, a hydrodynamic radius (Rh) of approximately 160 nm was
estimated. This size is consistent with similar PMOXA-PDMS based block copolymers with
shorter hydrophobic blocks (Nardin et al. 2000). The ratio of the radius of gyration (Rg) from
static light scattering experiments to the hydrodynamic radius from dynamic light scattering
was ~1 (Rg/Rh = 1.03) and supports a hollow sphere morphology (Nardin et al. 2000). The
molecular weight of each vesicle was estimated at 52x106 g/mol using static light scattering.

28
Transmission Electron Microscopy (TEM) and cryo TEM were also conducted on these
particles. The TEM images show relatively uniform sizes for the different vesicles imaged.
From cryo TEM, the radius of the vesicles in the hydrated state was 117 nm.
Figure 2.1 shows representative images obtained from electron microscopy. Radii estimated
from cryo TEM were used in further calculations of permeability as this method best preserves
the structural features of vesicular structures (Almgren et al. 2000).
The presence of spherical vesicle like aggregates with a hydrophilic corona was further
supported by AFM measurements. During AFM, when the suspension of dilute vesicles is
spread on a hydrophilic mica surface, a film, punctuated with emerging vesicles, was seen to
form (Figure 2.1).
Figure 2.1: Examination of polymer vesicles using microscopy. The cryo TEM images were used for size determination as regular TEM and AFM influence the structure of the observed vesicles. (a) cryo transmission electron micrograph of a ABA polymer vesicle.(scale bar 200nm) (b) Electron micrograph of a cluster of vesicles (scale bar 50 nm) (c)Atomic force micrograph of vesicles on mica in non tapping mode shows that a film of polymer is formed on the hydrophilic mica surface with vesicles located in the films.
2.3.3 Permeability Measurements
The permeabilities of ABA and AqpZ-ABA vesicles were investigated using stopped flow light
scattering experiments as described in Borgnia et al, 1999. Vesicle suspensions were rapidly
mixed with osmotic solutions (1.7 osmol/L) of salt (NaCl). The shrinkage of these vesicles was
followed by monitoring the increase in light scattering with time. The initial rise in the
experimental data was fitted to an exponential rise equation, and the exponential coefficient (k)
was used in calculating permeability.

29
These results are shown in Figure 2.2 for the ABA and AqpZ-ABA vesicles with a protein to
polymer molar ratio of 1:200. For AqpZ-ABA vesicles the time scale for the exponential rise
was between 5 and 20 milliseconds, while up to 10 seconds were required to capture the
exponential rise in light scattering of ABA vesicles because of its low permeability. The
calculated permeabilities of the ABA and the AqpZ-ABA vesicles were 0.8 m/s and 74 m/s
respectively based on these results. This represents a large permeability increase of ~ 90 times
with protein incorporation. Assuming complete incorporation of proteins into the polymer
vesicles, a maximum of 25 monomers per vesicle is expected to be incorporated based on the
molecular weight of the vesicles estimated from static light scattering. Using the calculated
permeability of 74 m/s, 25 monomers per vesicle and the surface area of each vesicle, the
calculated water permeability for each AqpZ tetramer is 13 x 10-14 cm3/s, which is similar to that
reported for AqpZ reconstituted into liposomes (≥10 x 10-14, (Borgnia et al. 1999)).
Figure 2.2: Stopped flow light scattering experiments (a) Increase in relative light scattering with and without reconstituted AqpZ into the ABA polymer at 5.5 ° C at a molar ratio of 200:1 (polymer:protein). Fits are shown as guides. The initial rise rates were used to calculate the permeability. As seen from (a) a rise cannot be calculated for the pure ABA polymer vesicles (b) shows the rise in scattering between 2 and 10 s for the polymer that was used in calculating permeability at 5.5 ° C.
2.3.4 Calculation of Activation Energies
Activation energies of water transport calculated by conducting experiments over a wide
range of temperatures can be used to characterize the transport across the vesicle membranes as

30
diffusion-driven or channel-mediated. Calculated permeabilities for the AqpZ-ABA vesicles
indicate an increase of 38 to 94 times over ABA vesicles in these experiments. In Figure 2.3, the
exponential constants obtained from these experiments are plotted against the inverse of
temperature to determine the Arrhenius activation energies. The Arrhenius activation energy
calculated for the ABA vesicles was 8.7 kcal/mol, while the AqpZ-ABA vesicles had a value of
3.4 kcal/mol. The higher value obtained for ABA membranes is consistent with the values of
activation energies reported for polymer membranes in the measured temperature range
(Garybobo 1971; Mehdizadeh et al. 1989), indicating transport by diffusion through the polymer.
The low activation energy for the AqpZ-ABA vesicles is strong evidence for channel-mediated
water transport across the vesicle membrane. This value is also consistent with the low
activation energy values obtained with proteoliposomes with incorporated AqpZ (Borgnia et al.
1999) and with oocytes with inserted AQP1 (Preston et al. 1992).
Figure 2.3: Arrhenius plots for calculation of activation energy for osmotic transport of water across polymer membranes with incorporated AqpZ (ABA with AqpZ) and pure polymer membranes (ABA).

31
2.3.5 Reflection Coefficients
The reflection coefficient of solutes in membrane systems determines the extent to which a
particular solute will be excluded from the filtered water. The relative reflection coefficients of
salt, urea, glucose, and glycerol were determined using a procedure described by Meinild et al
(1998). Glucose was taken as the reference solute with an assumed reflection coefficient of 1
because of its relatively large molecular size. The calculated reflection coefficients of salt,
glycerol and urea were >1 indicating higher rejection of salt, urea and glycerol when compared
to glucose (data not shown). These reflection coefficients are higher than the values reported by
Meinild et al (1998) for the same solutes but are within the experimental error for these
calculations.
2.3.6 Effect of Protein-to-Polymer ratio on permeability
Stopped flow experiments were conducted with protein to polymer molar ratios of 1:25, 1:50
1:100, 1:200, 1:500 and 1:1000 at 5.5 ° C. A dramatic increase in the permeability followed by a
sharp decrease was observed with increasing concentration of protein as shown in Figure 2.4.
At a ratio of 1:50 the increase in permeability over pure polymer vesicles is approximately 3000
times.
Figure 2.4: Effect of increasing AqpZ concentrations on the permeability of ABA polymers (a) light scattering results normalized to fit between 0 and 1. Curves for different ratios have been offset for clarity and fits are shown as guides. Data at 0 AQP: ABA ratio shown at a fixed offset. (b) Increase in calculated permeability with increase in protein to polymer ratio.

32
2.4 DISCUSSION
Dramatic improvements in the efficiency of water treatment membranes may result from the
development of biomimetic membranes with high permeability and selectivity. In this work we
present the synthesis of a novel ABA triblock copolymer with incorporation of the bacterial
water channel protein- AqpZ, data on the low permeability of the ABA polymer alone, and the
large increase in permeability upon incorporation of AqpZ. This increased permeability varied
with the AqpZ:ABA ratio and showed an activation energy indicative of channel mediated
transport. Furthermore, the selectivity of the AqpZ-ABA membrane for water over small solutes
such as salt, glucose, urea and glycerol was demonstrated.
The dramatic increase in permeability observed upon incorporation of AqpZ indicates that the
water channel protein is functional in the synthetic context, as has been seen previously for other
membrane proteins in a similar polymer (Graff et al. 2002). The magnitude of increase in
permeability and the excellent solute rejection capabilities demonstrate the potential benefit of
such membranes for water treatment and drug delivery. The ABA triblock copolymer was found
to be relatively impermeable when compared to other membranes, with a productivity of 0.22
(m/s)bar-1 at 20°C as compared to the other reported value for a similar diblock copolymer
(Polyethylene oxide-Polyethylethylene designated OE-7) with normalized permeability of 5.7
(m/s)bar-1 at 20°C (calculated from Discher et al, 1999). The large difference in the size of the
hydrophobic blocks between the ABA polymer (110 units of PDMS) and the diblock copolymer
studied by Discher et al (37 units of Polyethyleneethylene (EE)) could account for this
difference. The permeabilities of lipid membranes are reported to be an order of magnitude
higher than OE-7 copolymer (Discher et al. 1999). The incorporation of AqpZ increases the
productivity of ABA membranes by ~800 times (Figure 2.5). Additionally, the productivitites of
AqpZ-ABA membranes reported here exceed those for any salt rejecting membrane. A
comparison of the measured permeability values to those measured by other researchers for
block copolymer membranes and to commercial reverse osmosis and forward osmosis
membranes is shown in Figure 2.5. Although these values were measured using different
procedures, it can be seen from this comparison that the protein-polymer system investigated
here represents more than an order-of- magnitude improvement over existing commercial
solute-rejecting membranes.

33
Figure 2.5: Comparison of reported permeability values for polymeric membranes to those obtained in the study. FO is commercial forward osmosis membrane with data from (McCutcheon and Elimelech 2006) at 20 ° C. RO is commercial reverse osmosis desalination membrane with data from Matsura (Matsura 2001) at room temperature (assumed 25 ° C) PB-PEO is Polybutadiene Polyethylene diblock polymer with data from Discher et al (1999) at temp of 20 ° C. ABA represents the polymer vesicles used in study with permeability calculated at 20 ° C. ABA+ AQP represents the polymer vesicles with incorporated AqpZ at 1:100 molar ratio used in this study at 20 ° C. Data for ABA and AqpZ were obtained at 5.5 ° C and calculated at 20 ° C using Ea values.
The AqpZ-ABA system studied here may benefit from further optimization of the protein
incorporation conditions. This will allow production of membranes with a greater enhancement
in permeability. Increasing the amount of AqpZ leads to a trend of increase in permeability
followed by a decrease similar to that seen with lipid membranes. In experiments with lipid
membranes a limiting concentration of AqpZ was observed around a weight ratio of 1:200
protein to polymer (Borgnia et al. 1999). We see such a limiting concentration at a ratio of 1:19
protein to polymer (1:50 molar ratio). The higher molecular weight of the polymer compared to
lipids might play a role in this difference. The limiting concentration observed is probably due

34
to the method of incorporation utilized. At the higher concentration of AqpZ a large amount of
detergent present in the stock solution of AqpZ was added to the reconstitution mixture leading
to a reconstitution mixture detergent concentration of ~ 0.4%. This may have resulted in a low
efficiency of reconstitution for AqpZ leading to lower permeabilities. Unfortunately, the number
of AqpZ inserted per vesicle have not been determined experimentally for inserted AqpZs in
previous studies (Borgnia et al. 1999) as well as this study to allow a definitive conclusion.
Additionally, other studies have shown that vesicles with a AqpZ to polymer weight ratio of 1:1
can be synthesized using slow detergent removal methods (Dolder et al. 1996; Scheuring et al.
1999). This indicates that higher permeabilities than that reported here might be possible with
other reconstitution methods. These results also indicate that membranes with specified levels of
water permeability can be synthesized by controlling the concentration of AqpZ – a property that
may be useful for drug delivery applications.
The protein polymer membranes examined in this study show excellent salt rejection and high
permeabilities ideal for water treatment membranes used for desalination. Excellent control
over the permeability of polymers was also demonstrated. Incorporation of AQPs or suitable
molecular mimics into compatible synthetic polymers such as the block copolymer system
investigated in this study is an innovative approach for making solute-rejecting membranes for
medical, industrial and municipal desalting applications.
2.5 MATERIALS AND METHODS
The detergent dodecyl maltoside used in protein purification was obtained from Anatrace
Chemicals (Maumee, OH, USA). Ni-NTA agarose beads were obtained from Qiagen,
Inc.(Valencia, CA, USA). Sepahrose 4B was used for chromatographic separation was obtained
from Sigma Aldrich (USA and Switzerland). Other chemicals were obtained as described in the
following sections.
2.5.1 Expression and Purification of AQP-Z
The AqpZ overexpression vector pTrc10His AqpZ was obtained from Dr. Peter Agre (Duke
University Medical Center, Durham, NC, USA) and transformed into the Escherichia coli strain
JM109 by elecroporation. Growth, isopropyl- D-thiogalactoside induction and Ni-NTA
affinity chromatography purification of AqpZ were performed as described in (Borgnia et al.

35
1999) with two modifications. The induction time was optimized at 7-8 hours and this was used
for all the protein production experiments. The ultracentrifugation setup for recovery of
membrane fractions was replaced by the use of a Biomax filter (MWCO of 30 KDa, Billerica,
MA,USA) in a centrifuge for 45-60 minutes for removal of smaller molecular weight material.
Protein purification was followed by solubilization with cracking buffer (Sambrook et al. 1989)
with 1% dodecyl maltoside and running polyacrylamide gel electrophoresis(Sambrook et al.
1989). The purified protein was quantified using the Lowry method (Lowry et al. 1951).
2.5.2 Synthesis of ABA polymer
The ABA polymer was a symmetric poly- (2-methyloxazoline)-block-poly (dimethylsiloxane)-
blockpoly (2-methyloxazoline) (PMOXA15-PDMS110-PMOXA10). The first step of the synthesis
procedure involved acid catalyzed polycondensation of dimethoxydimethyl silane in presence of
water and end-capper resulting in butylhydroxy terminated bi-functional PDMS. Liquid PDMS
was purified by vacuum stripping at 80°C and precipitation in equal (by weight) water/methanol
mixture. Purified PDMS was reacted with triflicacid anhydride in hexane at -10°C for three
hours resulting in triflate-PDMS bi-functional macroinitiator. The reaction mixture was than
filtered under argon through a G4 filter. Hexane was evaporated under vacuum and dry ethyl
acetate was added as reaction solvent. Addition of dry 2-methyl-2-oxazoline resulted in
symmetric ring-opening cationic polymerization of PMOXA blocks on the macroinitiator. The
reaction was terminated by addition of a methanol solution of potassium hydroxide. The
synthesis of a similar lower molecular weight polymer system is described in further detail in
Nardin et al, 2000.
2.5.3 Preparation of ABA vesicles and AQP incorporation
Block copolymer vesicles (1-2 wt. % of polymer in phosphate buffer saline (PBS) at pH 7.4)
were prepared using the film rehydration method. 60 mg of polymer was first dissolved in
chloroform (5-10 ml) and the chloroform was evaporated slowly in a rotary vacuum evaporator
at 40°C and a vacuum of less than 400 mbar to form a even film on the inside of round bottomed
flasks. This film was then further dried under high vacuum of 0.3 mbar for at least four hours.
10 ml PBS was then added drop wise to the film with alternating vigorous vortexing and periodic
sonication (of durations less than 30s) over several minutes. This mixture was then left stirring

36
for at least eight hours. This suspension was then extruded several times through a 0.4 micron
track etched filter (Isopore, Millipore Corporation, Billerca, MA,USA), followed by a 0.2 micron
track etched filter (Nucleopore, Whatman, Newton, MA, USA) to obtain monodisperse
unilamellar vesicles. For reconstitution experiments 500 L of an AqpZ stock solution (1.6 mg
mL-1 in 1.5% dodecyl maltoside, 66 mM KH2PO4, 133 mM NaCl, 13% glycerol, 3.33 mM
mercaptoethanol (BME), and 33.33 mM Tris was added during the formation of the polymer
vesicles and subsequent steps were completed on ice. The resulting protein-containing vesicles
were purified chromatographically using a column packed with Sepharose 4B to remove non-
incorporated protein and detergent traces.
2.5.4 Characterization of vesicles
Light Scattering: The static and dynamic light scattering experiments were performed according
to the description presented in Nardin et al, 2000 (Nardin et al. 2000) using a commercial
goniometer (ALV Langen) equipped with a frequency-doubled Nd:YAG laser (ADLAS,
wavelength 532 nm) at scattering angles between 30° and 150°.
AFM: AFM measurements were conducted using a Picoscan SPM LE scanning probe
microscope equipped with a Picoscan 2100 SPM controller (Agilent Technologies, Chandler,
AZ, USA). Measurements were conducted in tapping mode using a Si cantilever (NCH)
(Nanosensors, Neuchatel, Switzerland). The length of this cantilever was 125 m and the
nominal force constant was 42 N/m. Samples were prepared by placing a dilute vesicle
suspension on freshly cleaved mica for 1 minute and then carefully washing with double distilled
water.
TEM: Transmission Electron microscopy was conducted on vesicle samples using a Phillips 400
TEM. The samples were prepared by dilution up to 1000 times and then stained with 2% uranyl
acetate on plasma treated copper grids.
cryoTEM: The vesicle suspension was deposited on holey carbon grid and frozen rapidly by
plunging into liquid ethane above its freezing point using a cryo holder. This holder was then
transferred to the TEM and imaged in transmission mode at 200kV at liquid nitrogen
temperature. The equipment used for this measurement was a Zeiss 922 Omega microscope with
a Gatan CT3500 Cryo Holder and a Gatan Ultrascan 1000 camera system.

37
Permeability measurements: The permeability of the two types of vesicles were determined
using a stopped flow procedure (Borgnia et al. 1999). The permeability was measured by
following the light scattering of the preparation in a stopped-flow apparatus (SV.17MV, Applied
Photophysics, Letterhead, UK). The water permeability was measured by rapid mixing of the
vesicles with a solution containing the osmotic agent (salt, glucose, glycerol or urea) causing
water efflux from the vesicles. The changes in light scattering caused by this shrinkage were
recorded at an emission wavelength of 600 nm in the stopped flow apparatus. Under these
conditions a reduction in vesicle volume leads to an increase in the relative light scattering
measurements as described in Borgnia et al, 1999 according to the methods developed by Milon
et al, 1986. These experiments were conducted with pressure in the mixing cell held constant at
7 bar to minimize pressure variations that could complicate the analysis of kinetics. Results were
corrected for the mechanical compression resulting from this pressure by subtracting the baseline
observed in control experiments without osmotic agents. These data were then fitted to an
exponential rise equation to calculate the exponential coefficient and the osmotic water
permeability (Pf) was calculated using the following expression (Milon et al. 1986):
Where, k is the exponential rise rate constant for the initial rise in the light scattering curve, S is
the initial surface area of the vesicles, Vo is the initial volume of the vesicles, Vw is the molar
volume of water (18cm3), and Δosm is the difference in osmolarity driving the shrinkage of the
vesicles.
Several sets of experiments were conducted with similar conditions but different preparations
(prepared in different batches) to ensure repeatability. Additionally, initial experiments with up
to 12 traces per experimental condition showed that five traces were adequate for providing
sufficient data for analysis, so in subsequent detailed experiments a minimum of five were
acquired per experimental condition. Data analysis was conducted by averaging a minimum of
five traces for each scattering experiment and then fitting the initial rise to an exponential
equation using the curve fitting toolbox available through the MATLAB software (Mathworks,
Natick, Massachusetts, USA).
osmWof VVSkP )//(

38
Stopped flow experiments were repeated at different temperatures (5.5, 7.5, 10, 12.5 and 15 °C)
for the ABA and AqpZ-ABA vesicles. The exponential rise rates calculated from these
experiments exponential were plotted against the inverse of temperature to determine the
Arrhenius activation energies.
The reflection coefficients for salt, urea and glycerol were calculated based on the method
presented by Meinild et al, 1998 (Meinild et al. 1998). Glucose was used as the reference solute
and the comparative experiments were all conducted at 15°C.

39
3 BIOLOGICAL ROLE FOR BACTERIAL AQUAPORIN GATING DISCOVERED IN
BLOCK COPOLYMER MEMBRANES
Ackowledgements: I performed all the experiments described in this chapter except for
PMOXA-PDMS-PMOXA polymer synthesis and transformation of the ptrcHis10AqpZ plasmid
in to E.coli.
The results described in this chapter have been submitted for publication.
3.1 ABSTRACT
Block copolymers can mimic lipids in structure and function, while being engineered for
desired chemical and mechanical properties. Use of a pH-insensitive, low-permeability triblock
polymer (Polymethyl oxazoline- Polydimethyl siloxane - Polymethyl oxazoline or PMOXA-
PDMS-PMOXA) as a matrix for reconstitution allowed discovery of pH gating of the bacterial
water channel protein Aquaporin Z (AqpZ). AqpZ was reversibly closed at pH ≤ 4 in polymer
vesicles and in whole cells of Escherichia coli. Furthermore, pH gating of AqpZ was important
for survival of acid and simultaneous acid and hypoosmotic shock, demonstrating the biological
relevance of this phenomenon. This work utilized a hybrid biological-synthetic system to provide
new insight into the physiological role of microbial aquaporins.
3.2 INTRODUCTION
Aquaporins mediate rapid and highly specific water transport across cell membranes
(Preston et al. 1992), and defects in aquaporins are associated with several human diseases (King
et al. 2000). In contrast, the role of aquaporins in microorganisms is uncertain. The large surface
area to volume ratios found in microorganisms should provide sufficient water transport via
passive diffusion without the need for a dedicated channel protein (Hill et al. 2004; Tanghe et al.
2006). Aquaporins are not essential in microorganisms, as evidenced by their incomplete
distribution (Zardoya 2005) and by the subtle effects observed in strains where the aquaporins
have been eliminated via inactivation of the aqp gene (Calamita et al. 1998; Soupene et al. 2002;
Hernandez-Castro et al. 2003; Shapiguzov et al. 2005). Indeed, some laboratory strains of yeast

40
appear to have gained a competitive advantage by inactivating their aquaporins (Bonhivers et al.
1998). Their existence, on the other hand, suggests some benefit. Proposed roles are diverse and
include involvement in osmoregulation, sporulation, cell expansion prior to cell division, and
freeze tolerance (reviewed by Tanghe et al (Tanghe et al. 2006) and Calamita (Calamita 2000)).
In plant and mammalian aquaporins, water transport is often regulated, including closure or
gating in response to changes in pH or osmolarity, and this regulation has provided insight into
the physiological roles of specific aquaporins (reviewed by Hedfalk et al (Hedfalk et al. 2006)).
Previous pH studies generally focused on the range between pH 6 and pH 8, perhaps due to
constraints imposed by pH effects on the liposome or oocyte systems used, and found a wide
range of effects. The plant plasma membrane intrinsic protein 2 (PIP2) subfamily of aquaporins
and the human AQP3 have reduced permeability at pH 6 or below pH 7, respectively (Tournaire-
Roux et al. 2003; Zelenina et al. 2003), while the mammalian AQP0 (Nemeth-Cahalan et al.
2004) and AQP6 (Yasui et al. 1999) have increased permeability at acidic pH, and mammalian
AQP4 & AQP5 are reported to be insensitive to pH (Zelenina et al. 2003). However, the
Escherichia coli aquaporin AqpZ is insensitive to two conditions affecting other aquaporins: 1
mM mercury and changes in pH between 5 and 7 (Borgnia et al. 1999), and to our knowledge no
experimental evidence of gating in a prokaryotic aquaporin has been reported prior to this work.
The structure and function of membrane proteins, including aquaporins, are typically
studied via reconstitution in a lipid matrix, but these investigations are limited by the lipid
properties. In particular, lipids have high permeability for water and gases, making it difficult to
study transport of these molecules (Verkman 2002; Uehlein et al. 2003; Hill et al. 2004).
Liposomes can also be strongly affected by pH, ionic strength, and temperature(Grit and
Crommelin 1993), complicating studies that need to be conducted under these conditions. To
investigate a broader range of membrane protein properties and as a platform for technology
development, we envision the increased use of synthetic block copolymer membranes with
tailorable properties.
Symmetric, ABA-type triblock copolymers composed of poly-(2-methyloxazoline)-poly-
(dimethylsiloxane)- poly-(2-methyloxazoline) (PMOXA-PDMS-PMOXA, referred to as ABA
throughout the rest of this paper) mimic lipid bilayers in their structure (Figure. 3.1) and ability
to form vesicular structures (Nardin et al. 2000). The feasibility of using ABA polymers for the
study of membrane proteins is supported by studies demonstrating insertion and activity of

membrane proteins, including AqpZ (Kumar et al. 2007). Specifically, AqpZ-rich ABA block
copolymer biomimetic membranes may have an important role in addressing world water supply
issues as alternative desalination membranes (Kumar et al. 2007). The use of PMOXA as the
hydrophilic block also provides the possibility of adding antifouling properties in this and other
applications (Konradi et al. 2008) where protein deposition is undesirable. The polymer vesicles
have several advantages over liposomes: i) they can be engineered to provide the desired
properties such as fluidity or morphology by the choice of blocks and block lengths (Discher and
Eisenberg 2002) or their length ratio (Zhang and Eisenberg 1995) ii) their end groups can be
modified (Nardin et al. 2000; Napoli et al. 2008) by molecules such as biotin (for recognition and
immobilization ) (Grzelakowski et al. 2009), methacrylate (for stabilization by crosslinking)
(Nardin et al. 2000), fluorescent molecules (for imaging) (Grzelakowski et al. 2009) and even
drugs (for drug delivery) (Peer et al. 2007) iii) they are highly stable (Discher et al. 1999), and
iv) they can be engineered to be sensitive (Sauer et al. 2001) or insensitive to pH changes (Figure
3.2). In this work, the use of pH-stable, low permeability ABA triblock copolymers permitted the
discovery of pH gating of AqpZ.
Poly‐2‐methyl‐2‐oxazoline Polydimethylsiloxane Poly‐2‐methyl‐2‐oxazoline
N NSi
O OSi Si
n OOHHO
O m m
Hydrophilic
Hydrophobic
Figure 3.1: Structure of PMOXA-PDMS-PMOXA polymer
41

Figure 3.2: AqpZ-ABA vesicles are stable over a wide pH range. To evaluate the pH stability of the AqpZ-ABA vesicles, dynamic light scattering measurements were conducted following overnight incubation at different pH. The results demonstrate the stability and uniform size of the vesicles between pH 2 and pH 12. Each point was calculated from 5 data points at different scattering angles. Error bars represent 95% Confidence Interval (C.I.) values.
3.3 RESULTS
3.3.1 Closure of AqpZ at low pH in polymer vesicles
During a systematic investigation of the permeability of ABA polymer vesicles with
incorporated AqpZ (AqpZ-ABA vesicles), we noticed that permeability decreased by 81%
between pH 6 and pH 4 (Figures 3.3 (a) and (b)). Two subsequent experiments suggested that
this was not due to damage of the polymer. First, sizing experiments suggested that the ABA-
AqpZ vesicles are stable at low pH (Figure 3.2). Second, the activation energies of water
transport for the AqpZ-ABA vesicles at pH values of 4, 7.2, and 12 are in the range of channel-
mediated transport (1.8 to 3.1 kcal/mole (Figure 3.4)) and lower than that through pure polymer
(8.7 kcal/mol (Kumar et al. 2007)), indicating that transport is still mediated by AqpZ throughout
this pH range. The permeability of AqpZ-ABA vesicles at pH 2 and 4 (84 and 107 m/s,
respectively), while lower than at neutral pH, is still higher than pure polymer vesicles (0.8
m/s(Kumar et al. 2007)), indicating that not all AqpZ activity is eliminated under these
conditions.
42

Figure 3.3: AqpZ-ABA vesicles have a lower permeability at acidic pH and this decrease is reversible. (a) Normalized light scattering response from a representative hyperosmotic shock experiment on AqpZ-ABA vesicles prepared at pH 4, 7.2, and 12. (b) A significant decrease in permeability (no overlap of 95% Confidence Interval (C.I.) error bars) is seen between pH 6 and 4. (c) AqpZ closure is reversible. AqpZ-ABA vesicles were tested at pH 4.0 (or 7.2), resuspended at pH 7.2 (or 4.0), and tested again. Error bars are 95% C.I. values from exponential fits of trace data (>8) for each condition.
Figure 3.4: Activation Energy for AqpZ-ABA vesicles at pH 4, pH 7 and pH 12. Based on permeability measurements at different temperatures, the activation energies of water transport at pH values of 4 (squares), 7.2 (circles), and 12 (triangles) are between 1.8 to 3.1 kcal/mole, indicating that transport is channel mediated.
To distinguish between damage and gating of AqpZ at low pH, the reversibility of this pH
effect was examined. The toughness of these vesicles allowed a batch of vesicles to be recovered
43

after stopped flow and reused at a different pH. Vesicles transferred from pH 4.0 to pH 7.2
regained 123% of the permeability of vesicles originally at pH 7.2 (Figure 3.3c). The recovery
following exposure to pH 2 was also complete (Figure 3.5). Thus, the low permeability observed
at acidic pH was fully reversible, indicating gating of AqpZ.
Figure 3.5: pH gating reversibility and control experiments for AqpZ-ABA vesicles. pH gating reversibility and control experiments shown for pH values between 7.2 and 2.0. Permeabilities were calculated from average of 8 or more stopped flow traces and error bars represent 95% C.I. values.
3.3.2 Closure of AqpZ at low pH in E. coli
This pH gating is an intrinsic property of AqpZ rather than an artifact of the synthetic
polymer environment, based on similar experiments with whole cells of E. coli (Figure 3.6a).
Comparing the permeabilities of strains with normal, overexpressed, and no aqpZ at pH 7.4, the
overexpression strain had the expected increase in permeability compared to the wild type strain.
The activation energy of water transport in the overexpression strain was 1.6 kcal/mol for pH 7.4
and 2.6 kcal/mol for pH 4 (Figure 3.7), indicating that the increased transport is still channel
44

mediated. With respect to pH, the wild type aqpZ strains (ME9062 and JM109 pTrc10His1) and
the aqpZ overexpression strain showed 15%, 44%, and 50% lower permeability, respectively, at
pH 4 than pH 7.4, while no decrease was observed in the aqpZ null strain. These results
demonstrate pH gating of AqpZ in E. coli.
Figure 3.6: Whole cells also exhibited an AqpZ-dependent, reversible decrease in permeability at low pH. (a) E. coli cells were less permeable at low pH. This decreased permeability was due to AqpZ, as an overexpression strain (AqpZ+) showed a larger decrease (50%) and no significant decrease was detected in an aqpZ null mutant (AqpZ-). Inset shows the less permeable strains at higher resolution. The results are averaged over 5 or more experimental traces. (b) Reversibility. Permeability of the overexpression strain recovered when the pH was changed from 4 to pH 7.4. Average permeabilities from three independent cultures are shown. Error bars represent 95 % C.I. values.
45

Figure 3.7: Activation Energy for E. coli water transport at pH 4 and pH 7. Permeability measurements were conducted at temperatures between 5 and 25C. The activation energies of water transport at pH values of 4 (circles) and 7.4 (squares) were calculated at 2.6 and 1.6 kcal/mol respectively.
As in the polymer vesicles, the observed decrease in the permeability of E. coli cells at pH
4 is not due to irreversible damage, because the original permeability is recovered upon
resuspension at neutral pH (Figure 2b). When the pH is increased from pH 4 to pH 7.4, the
resulting permeability of the overexpression strain is 93 % (95 % C.I. values, 74%, 115%) of its
original permeability at pH 7.4, indicating that the decrease in permeability at low pH is
reversible.
3.3.3 Effect of AqpZ on survival of acid and osmotic shock
While considering the possible physiological relevance of pH gating of AqpZ, we
investigated the effect of AqpZ on cell survival under separate and simultaneous acid and
osmotic shock (Figure 3.8). To avoid complications resulting from different growth rates in the
induced aqpZ overexpression strain and the other strains considered here, the overexpression
strain was only tested under uninduced conditions. However, even under uninduced conditions,
the permeability of the uninduced aqpZ overexpression strain is high, and at pH 4 its
46

permeability is higher than the wild type and null mutant under any conditions. In agreement
with the permeability data, this strain also has the most pronounced response to acid shock
(Figure 3.8 (b)). For osmotic shock alone, no significant difference in survival was observed
between the three strains, in agreement with previous reports indicating that AqpZ is not
essential for survival of osmotic shock (Calamita et al. 1998; Soupene et al. 2002). The
simultaneous acid and osmotic shock had the most pronounced effect, with all strains showing
lower survival than would be predicted based on separate shock experiments, and again with the
most permeable strain showing the lowest survival.
Figure 3.8. Lower permeability strains survive acid and simultaneous acid/hypoosmotic shock better. The acid shock was from pH 7.4 to 2.5 and the osmotic shock was from 300 to 100 mOsm (hypoosmotic), both for 30 minutes. All error bars represent 95 % C.I. values. At least three independent cultures were examined for each experimental condition.
3.4 DISCUSSION
This work demonstrates the biological relevance of a property discovered in a synthetic
membrane, illustrating the potential of hybrid biological-synthetic systems to contribute new
insight to biological systems. Our results demonstrate the reversible closure of AqpZ at pH 4 in
synthetic polymer vesicles and in whole cells. The low intrinsic permeability of these polymer
vesicles (Kumar et al. 2007) allowed sensitive determination of the change in AqpZ
permeability, and the insensitivity of the polymer to pH changes allowed investigation of a broad
range of pH conditions. The discovery of pH gating in the bacterial aquaporin AqpZ has
identified new directions for investigating the molecular mechanism of aquaporin gating and the
role of aquaporins in bacterial physiology.
47

The neutral pH permeabilities of our strains were consistent with previous reports using
different wild type (Mallo and Ashby 2006), null and overexpression strains (Hubert et al. 2005)
of E. coli. However, one apparent contradiction should be noted. Mallo and Ashby detected
<10% of wild type permeability in an aqpZ null mutant (Mallo and Ashby 2006), while our data
showed 84% of wild type permeability for another aqpZ null mutant. The reason for this
discrepancy has not been determined, but it could reflect differences in the strains or
experimental protocols.
The presence of pH-mediated AqpZ closure in both polymer vesicles and whole cells
indicates that pH has a direct effect on AqpZ. Detailed molecular mechanisms for pH gating
between pH 6 and pH 7 by plant and mammalian aquaporins have been proposed (Tournaire-
Roux et al. 2003; Zelenina et al. 2003; Nemeth-Cahalan et al. 2004; Törnroth-Horsefield et al.
2006). However, the molecular mechanism for the gating observed here is likely to be distinct
from those in plant and mammalian aqauporins, because the residues involved in these
mechanisms are not conserved in AqpZ (Figure 3.9). The pH range suggests the involvement of
a histidine residue, and the experimental assays used in this work provide a basis for
investigating the molecular mechanism.
Figure 3.9: Residues involved in pH gating in plant and mammalian aquaporins are not conserved in bacterial aquaporins. Aquaporin amino acid alignments are shown for regions identified in the literature as important for pH gating. The first segment corresponds to Loop A, the second and third to Loop C, and the fourth segment is from Loop D. Specific residues that affect pH gating are circled in the figure and include: H40 for hAQP0 (Nemeth-Cahalan et al. 2004); H53, Y124, S152 and H154 for hAQP3 (Zelenina et al. 2003); H193 and L197 for SoPIP 2;1 (Zelenina et al. 2003) and H197 and L201 for PIP 2;2 (Tournaire-Roux et al. 2003). Amino acid sequences were retrieved from the Genbank database (http://www.ncbi.nlm.nih.gov/) with the sequence for hAQP3 obtained from Uniprot database (http://www.uniprot.org/). The alignment was conducted using the Clustal W program (http://align.genome.jp/) and results are displayed using Cinema 5 (http://utopia.cs.manchester.ac.uk/cinema).
48

To consider the physiological relevance of pH gating of AqpZ, we developed a model
integrating pH gating of AqpZ with existing information on bacterial responses to osmotic and
acid shock (Figure 3.10). When exposed to hypoosmotic shock, aquaporins may provide a rapid
equilibration mechanism, while the increased pressure (and accompanying membrane stretching)
resulting from hypoosmotic shock activates mechanosensitive channels (Berrier et al. 1992;
Levina et al. 1999) to release small solutes. This prevents cells from bursting when a sudden
downshift in osmotic pressure occurs. At low pH, an increase in activation pressure of the
mechanosensitive channel MscL has been demonstrated (Kloda et al. 2006) and serves to prevent
acidification of the cytoplasm. However, this regulation also reduces the protective action of
MscL. The ability of cells to avoid a rapid influx of water, via pH gating of AqpZ, might
therefore be an important protective step under combined acid and hypoosmotic shock. This
model is supported by the reduced survival of more permeable strains under acid and combined
acid/osmotic shock (Figure 3.8).
Figure 3.10: Model for coordinated activity of aquaporins and mechanosensitive channels under pH and osmotic downshift conditions. Upon hypoosmotic shock aquaporins mediate rapid water transport into the cell, leading to rapid swelling. To prevent cells from bursting, mechanosensitive channels open and allow small solutes to be released. Under acid shock, mechanosensitive channel activation is lowered. The cell is therefore especially vulnerable to combined acid and hypoosmotic shock, unless water permeability of AqpZ is also reduced under acidic conditions.
49

While many questions remain about the physiological role of AqpZ, this report suggests a
new direction for such investigations, namely acid stress. Acid stress response also provides a
potential explanation for the apparent preferential distribution of AQP homologs in Gram
negative bacteria (Zardoya 2005), as the acid response in these organisms relies in part on
periplasmic acidification, while the Gram positive bacteria utilize different protective
mechanisms (Foster 2000). The acid and combined acid and osmotic shock conditions
considered here for whole cells are environmentally relevant, similar to conditions E. coli and
other pathogens might be exposed to upon passage through a host digestive system, and therefore
insights into bacterial survival under these conditions may also have relevance to public health
and food safety.(Diez-Gonzalez et al. 1998)
This work demonstrates that specifically designed biomimetic polymeric materials can
facilitate discovery of biologically relevant fundamental properties in hybrid synthetic-biological
systems. The robust block copolymer membrane platform provides the flexibility to conduct
assays of membrane protein properties under conditions that were previously limited by the
mechanical and chemical properties of lipid membranes.
3.5 MATERIALS AND METHODS
3.5.1 AqpZ-ABA vesicles.
AqpZ, poly (2-methyloxazoline)-block-poly (dimethylsiloxane)-block-poly (2-
methyloxazoline) (PMOXA15-PDMS110-PMOXA15) polymer, and protein-polymer vesicles were
prepared as described previously (Kumar et al. 2007) using a protein to polymer molar ratio of
1:100 and adjusting pH with HCl or NaOH. Vesicles were purified using Sepharose 2B. Size and
stability of AqpZ-ABA vesicles synthesized at different pH values were evaluated before and
after overnight storage using dynamic light scattering as previously described (Kumar et al.
2007).
3.5.2 Vesicle permeability.
Permeabilities were calculated from stopped flow light scattering results(Kumar et al.
2007) with 0.85 M NaCl as the osmotic agent for polymer vesicles. Three sets of experiments
were conducted with similar conditions but independent preparations to ensure repeatability. A
minimum of eight traces were averaged for each experimental condition. Reversibility
50

experiments were conducted by collecting the vesicles after stopped flow, transferring them to
the second pH by repeated washes in a centrifugal filter (20kDa, Amicon Ultra 15, Millipore),
and repeating the stopped flow experiment.
3.5.3 Bacterial strains.
Four strains were used for the experiments reported here, an aqpZ null mutant, an aqpZ
overexpression strain, and their respective parents. The wild type E. coli strain was the parent
strain used for construction of the Keio collection (BW25113) as obtained from the National
BioResource Project (Japan) under strain number ME9062. Its genotype is rrnB lacZ4787
hsdR514 (araBAD)567 (rhaBAD)568 rph-1. The aqpZ null mutant (JW0859) was constructed
in ME9062 as part of the Keio collection and contains a kanamycin resistance gene inserted in
aqpZ (Baba et al. 2006). The AqpZ overexpression vector pTrc10HisAqpZ was obtained from
Dr. Peter Agre (Duke University Medical Center, Durham, NC, USA) and transformed into E.
coli strain JM109 (endA1 recA1 gyrA96 thi hsdR17 (rK-, mK
+) relA1 supE44 Δ(lac-proAB) [F'
traD36 proAB lacIqZΔM15]) by electroporation. To construct an isogenic control for the
overexpression strain, AqpZ was deleted from the pTrc10HisAqpZ plasmid using a EcoRI and
Pst1 double digest, overhangs were filled in with T4 DNA polymerase, and the resulting DNA
(pTrc10His1) was blunt end ligated and introduced into JM109 by electroporation.
3.5.4 Whole Cell Permeability Experiments.
These experiments were conducted using a stopped flow apparatus as described in Mallo and
Ashby (Mallo and Ashby 2006). All strains were grown overnight (15 hours) with shaking at 37
deg C in LB medium. 50 g/ml ampicillin was included as needed to maintain plasmids. The
cultures were then subcultured (1:100) into fresh LB medium. For induction of AqpZ, IPTG was
added 4 hours after subculturing. Eight hours after subculturing, the cells were harvested by
centrifugation at 10,000 g for 5 minutes at 4 deg C. The cell pellet was washed three times with
a solution of 100 mM PBS, with the addition of 50 g/ml kanamycin (or chloramphenicol for the
aqpZ null strain) to suppress protein synthesis during sample preparation and stopped flow
analysis. After washing, the cells were resuspended in a variable volume of PBS – antibiotic
solution at pH 7.4 or 4.0 to obtain a solution with a fixed transmittance (701%) and tested
within an hour.
51

Stopped flow experiments were conducted on the above cell suspensions with 1 M proline (also
adjusted to pH 7.4 or 4.0) as the osmotic agent and time bases between 0.2 and 5 s. The initial
volume (Vo) and surface area (S) of the cells were calculated as described in Mallo and Ashby
(Mallo and Ashby 2006) by assuming a length of 5 m and a diameter of 1 m, average values
for K-12 strains of E. coli under these growth conditions (Van De Merwe et al. 2004). Simple
exponential equations were used to determine the initial rise rate using the curve fitting toolbox
in MATLAB. The fit values were also tested using the curve fitting software provided with the
Applied Photophysics stopped flow for one set of experiments.
The reversibility of permeability change between pH values was tested as follows. An induced
culture of the overexpression strain was resuspended in pH 7.4 (or 4) buffer and part of this
culture was used to measure permeability. The remaining culture was spun down and transferred
to the second pH. Thus different aliquots of the same culture were tested at both pH values.
Five or more traces were obtained for each condition. At least three independent cultures were
tested.
3.5.5 E. coli survival experiments.
The experiments were conducted using a procedure similar to that described by Levina et al
(Levina et al. 1999). Single colonies from fresh overnight plates were grown overnight (12-15
hours) with shaking at 37 deg C in LB medium. They were subcultured 1:50 (for two sets of
experiments with AqpZ- and ME9062 experiments) or 1:100 (all other experiments) into fresh
LB medium supplemented with 0.2 M NaCl and grown for 4 hours. These cultures were diluted
1:10 (for experiments under both acid and osmotic shock conditions) or 1:100 (all other
experiments) into different media preparations to provide the desired shock. For control
experiments, dilution was performed in the same medium, while the acid shocks were performed
in LB at pH 2.5 and the osmotic shocks were in LB without supplemental NaCl. After 30
minutes of incubation at 37 deg C, serial dilutions were spread on LB agar plates and incubated
at 37 deg C overnight. Survival was calculated based on results of the control plates for each
culture.
52

4 SELF-ASSEMBLY OF MEMBRANE PROTEINS IN BLOCK POLYMERS
Ackowledgements: I performed all the experiments described in this chapter except for
PMOXA-PDMS-PMOXA and NTA-polymer synthesis, transformation of the ptrcHis10AqpZ
plasmid in to E.coli, surface pressure measurements (Figure , and Brewster Angle Microscopy
measurements.
4.1 ABSTRACT
Incorporation of membrane proteins at high densities in lipid membranes to produce 2D
crystals is an excellent but challenging technique to study membrane protein structure in the lipid
environment (Jap et al. 1992). Such arrays could also have relevance to applications where a
large concentration of membrane proteins is beneficial. Increased sensitivity in sensor-based
applications and transport in transport-oriented applications are examples of such applications.
In this work we show evidence for crystallization of membrane proteins in a biomimetic triblock
copolymer. A templating diblock copolymer film with a nickel functionalized hydrophilic block
was first assembled at the air water interface. A ternary mixture of triblock copolymer, detergent
and histidine-tagged membrane protein was then added to the subphase. Slow detergent removal
using polymeric adsorbents led to formation of flat protein rich areas up to 1 micron in length.
Electron microscopy imaging and electron diffraction show that some of these areas are highly
ordered (crystalline). In addition to protein-polymer crystals that look similar to previously
described, lipid-based AqpZ 2D crystals and display loose ordering, strongly diffracting
structures were also observed. These structures may be pure block copolymer crystals or novel
2D crystals of block copolymers and AqpZ. The structures observed and the electron diffraction
results for these crystals are different from lipid-based 2D crystals.
4.2 INTRODUCTION
Membrane proteins provide specific and elegant transport mechanisms for water, ions and
nutrients needed by a cell to survive. They are also the cell’s sensor for environmental
conditions that range from presence of specific toxins to change in pH. Thus, understanding the
structure of membrane proteins is fundamentally important. In addition, incorporating these
proteins into cell membrane analogs could provide materials with targeted applications in water
53

purification (Kumar et al. 2007), enzymatic reactions (Astier et al. 2005), and contaminant
recognition, transport, and sensors (Gu et al. 1999; Braha et al. 2000; Nielsen 2009).
Incorporation of membrane proteins at high densities in lipid membranes to produce 2D crystals
is widely used to study membrane protein structure in the lipid environment (Jap et al. 1992).
Block copolymer membranes mimic the architecture of natural lipid cell membranes and allow
incorporation of functional proteins across the membrane (Taubert et al. 2004). However, in
contrast to lipid-based membranes, block copolymers can be tailored to have the stability and
toughness attributable to polymeric materials (Discher et al. 1999) and hence are more suitable to
formation of membrane protein based devices and applications.
Here we provide the first report on the possible formation of 2D membrane protein crystals
using amphiphilic block copolymers. The bacterial water channel protein Aquaporin Z was used
for these crystallization experiments. The procedure used was a monolayer based templating
procedure described previously for protein crystallization (Uzgiris and Kornberg 1983; Lévy et
al. 1999). However, instead of the commonly used nickel chelated lipid monolayer, a highly
detergent-resistant monolayer consisting of a nickel (Ni2+) functionalized Polybutadiene-
Polyethylene Oxide diblock copolymer (Nehring et al. 2009) was used. Additionally the 2D
crystal was assembled in a PMOXA-PDMS-PMOXA based triblock copolymer instead of the
natural or synthetic lipids used previously.
4.3 RESULTS
4.3.1 Monolayer formation
Langmuir trough film compression studies revealed transition between the dehydration of
the hydrophilic blocks of the diblock copolymer to the self assembly of the hydrophobic
polybutadiene blocks at around a mean molecular area of 100 Angstrom2 (Figure 4.1). Brewster
Angle microscopy of this self assembly process using the precursor hydroxyl terminated
hydrophilic block polymer (PB-PEO-OH) reveled that large areas (>100 microns) of stable
monolayer areas could be formed.
54

Figure 4.1: Film compression data for the Ni NTA parent diblock copolymer at the air water interface.
4.3.2 Templating AqpZ particles
Histidine tagged AqpZ molecules could be templated onto the NTA diblock copolyment
monolayer. Figure 4.2 shows a comparison between single particle images of AqpZ (panel a)
and templated AqpZ particles on the NTA polymer at different coverages (panels b and c) when
the monolayer was incubated with 40 g/ml protein. As seen from this figure both regularly
distributed templated proteins (Figure 4.2 (b)) as well as areas with aggregated proteins (Figure
4.2 (c)) are seen. This shows that templating of His-tagged membrane protein is possible using
this interfacial film.
Figure 4.2: Templating of AqpZ tetramers on Ni NTA block copolymer film. a) Single particle AqpZ micrograph showing AqpZ tetramers, negatively stained with 1% uranyl acetate. Scale bar 0.1 micron. b) Templated AqpZ molecules on the Ni NTA block copolymer film (Scale bar
55

0.2 micron) c) Higher density of templated AqpZ molecules showing aggregated areas. (Scale bar 0.2 micron)
4.3.3 Self assembly of protein-lipid films on templating layer
PC/PS lipids formed large micron size folded sheets at a LPR of 2 with a number of
wrinkled vesicles also seen. At lower LPR values vesicles and smaller flat sheets were observed.
Figure 4.3(a) and (b) show folded sheets and vesicles with bright edges indicating the presence
of packed proteins. The use of DOPC/POPC lipids show formation of flat single or
multilayered layered crystals at a LPR of 2 as shown in Figure 4.3(c) and 4.3(d). This result
was expected as POPC/DOPC lipids have been used for 2 D crystallization of AqpZ using other
(non-templating) methods (Ringler et al. 1999)
Figure 4.3: 2 D crystals of AqpZ formed in lipid using templating block copolymers. a) Large folded sheet formed at a LPR of 2 with PC/PS lipids. Scale bar 0.5 micron b) Wrinkled vesicles formed at a LPR of 2 with PC/PS lipids. Scale bar 0.5 microns. c) and d) Flat sheets formed at a LPR of 2 with DOPC/POPC lipids. Scale bar 0.2 microns.
4.3.4 Self assembly of protein-polymer films on templating layer
Three common structures were formed with an ABA polymer to protein ratio of four.
Figure 4.4(a) shows that large vesicles that were also formed under these conditions but were
56

apparently unordered as seen from a lack of diffraction spots in the fourier transform of their
image. The structures shown in Figure 4.4(b) seem to be crystalline. This is seen from the
power spectrum of the raw images (inset in Figure 4.4(b)) which reveals that this structure has a
tetragonal structure as would be expected from a AqpZ 2D crystal. The weak spots seen here
indicate that the crystal is weekly ordered. Figure 4.4(c) shows large flat-plate like structures, but
the power spectrum of the image by itself does not show diffraction spots. Of the above three
types of structures, only the structures shown in Figure 4.4(b) has the behavior expected of lipid-
based membrane protein 2D crystals, with diffraction spots occurring in the image power
spectrum at 40,000-50,000 magnification.
Figure 4.4: Representative structures formed using the monolayer technique. (a) Small ordered areas formed at a polymer to protein ratio of four. Inset shows power spectrum of the image showing spots indicating a tetragonal crystal. Scale bar 50 nm (b) large vesicles. Scale bar 100 nm. (c) Large flat plate like structures. Scale bar 200 nm.
However, as shown in Figure 4.5, direct electron diffraction of the flat plate like
structures commonly seen in such samples diffract electron quite strongly (4th order). This is the
limit of what could be expected to be resolved using a negatively stained membrane protein
crystal. However, these diffraction patterns are unexpected as AqpZ crystals in lipids rarely
show such bright diffraction spots under negative stain conditions. One clue to understanding
the origin of these spots is to determine the lattice spacing indicated calculated from these
diffraction patterns.
57

Figure 4.5: 2 D crystals formed in PMOXA PDMS PMOXA triblock copolymer (a) Large sheet formed at a polymer to protein ratio of four with PMOXA PDMA PMOXA polymers. Scale bar 0.2 micron (b) Close up of a) Scale bar 50 nm. (c) Electron diffraction of crystal shown in a using selected area diffraction.
4.4 DISCUSSION
This work was focused on synthesizing protein-rich polymer films. Such films were
formed at the scale of ~ 1 micron. Specifically, there were two significant findings of this work
that are discussed in the following paragraphs: a) Formation of a stable Ni2+-functionalized,
polymer monolayer at the air water interface which was resistant to detergent (up to 1% DDM),
which represents 180 times the critical micelle concentration, and b) Formation of 2D crystals of
AqpZ in block copolymers.
Stable functionalized diblock co-polymer monolayers such as those used in this study
could have two direct applications. The first application is for immobilizing proteins from
solutions. In particular, the high detergent resistance of this film is important to applications such
as the Ni-chelating lipid-based monolayer techniques proposed by Kelly et al (Kelly et al. 2008)
and an extension of this technique, the so-called “affinity grid”. In the affinity grid technique Ni-
chelating lipids are deposited onto a TEM grid, dried and stored (Kelly et al. 2008a; Kelly et al.
2008b). Both the monolayer and the affinity grid techniques have been suggested for extracting
specifically tagged proteins from crude cell lysate for direct three dimensional cryogenic electron
microscopy-based (cryo-EM) tomography and structure determination (Kelly et al. 2008). These
templating di-block co-polymers could be an excellent replacement for chelating lipids for such
applications, particularly as high concentrations of detergent are sometimes necessary for
solubilization of membrane proteins. The higher molecular weight of this polymer indicates, in
addition to a higher detergent resistance, higher mechanical and chemical stability. For the
“affinity grid” loss of the lipid monolayer to glycerol and/or detergents limits some of the
58

functionality of this method (Kelly et al. 2008a). Additionally, the hydrophilic block of the
diblock copolymer could be functionalized in a variety of ways in order to provide specific
recognition for proteins and other macromolecules. Here, the hydrophilic block was
functionalized with nickel providing recognition for histidine tagged proteins or possibly with
proteins with surface histidines (Pack et al. 1997). Functionalizing the hydrophilic end with
other affinity tags could provide a customized templating procedure. Secondly, the block
copolymer monolayer could be successfully used for 2D crystallization in lipids. 2D
crystallization of AqpZ using a monolayer technique has not been shown before. Here we tested
both a PC/PS based system as well as a previously used POPC/DOPC system. This versatile
diblock polymer system could be used in place of oxidation sensitive lipid based Ni-chelating
monolayers in 2D crystallization studies.
2D crystallization of AqpZ in block copolymers is the most significant finding of this
chapter. The crystals formed had a similar morphology to lipid-based AqpZ 2D crystals. The
quality of the crystals obtained thus far is not adequate for further processing. Additional efforts
should be directed towards making larger crystals, using other methods including dialysis, if
final structure of the protein in block copolymers is to be determined. Alternative methods of
transfer to the TEM grid and better imaging techniques including the use of cryogenic and low-
dose electron microscopy techniques under various tilt conditions should be utilized.
Nevertheless, this is a highly promising initial finding. The ability to form 2D crystals opens the
door to structure determination of membrane proteins in these polymers. Accurate structure
determination of proteins in polymers can provide a molecular basis for anomalous effects seen
after incorporation of membrane proteins into block copolymers and provides a basis for design
of novel functional materials. Thus, attempts should be made to from 2D crystals of such
proteins where a large change in conformation is suspected from activity measurements, and that
are of interest for applications. Moreover, a clear understanding of this phenomenon will
provide us with the tools to manipulate the extent of hydrophobic mismatch to control the level
of protein activity.
The presence of highly diffracting, large flat areas that do not seem to be similar to
previously described lipid-based AqpZ crystals was surprising. To better characterize these
areas, the electron diffraction pattern shown in Figure 4.6(b) was used to calculate the lattice d-
spacing values for the crystal shown in Figure 4.6(a). For lipid-based AqpZ crystals the d
59

spacing should be about 10 nm, as the unit cell size is in the range of a=b=10 nm. However, for
these flat areas the d spacing was quite large (~ 28 (100) and 20 nm (110)) indicating that this is
not a typical protein crystal. The possibility of this being an artifact from the staining process
was eliminated, as similar diffracting structures were found in unstained, cryo-plunged samples.
Another common cause of artifacts in TEM sample, parts of the carbon support, could also be
eliminated as a potential artifact as these are amorphous materials and would not show sharp
diffraction spots. These areas could be either pure block copolymer crystals or a novel form of
block copolymer – AqpZ 2D crystal. If these are pure block copolymer crystals, it would be the
first report of PDMS block copolymer crystallization though self assembly in an aqueous
solution and would be applicable to determination of the polymer structure. If these are novel
AqpZ-ABA block copolymer crystals it could be studied further to confirm the composition and
structure of these unique materials, particularly if AqpZ can be shown to be active.
Figure 4.6: Image (a) and selected area diffraction pattern (b) of a flat crystalline structures seen during crystallization at a polymer to protein ratio of four using a nickel templating monolayer.
4.5 MATERIALS AND METHODS
The lipids used in this study were obtained from Avanti Polar Lipids, Alabaster, AL, USA.
The detergents dodecyl maltoside (DDM) and Octyl glucoside (OG) used in protein purification
were obtained from Anatrace Chemicals (Maumee, OH, USA). Ni-NTA agarose beads were
obtained from Qiagen, Inc. (Valencia, CA, USA). Transmission electron microscopy grids
were obtained from Ted Pella, Redding, CA, USA (Cu grids) or Pacific Grid-Tech, San
60

Francisco, CA, USA (Mo grids). N,N-Bis[(tert-butyloxycarbonyl)methyl]-L-lysine tert-
Butylester (Lys-NTA) was a gift from Prof. Robert Tampé’s group, Institute of Biochemistry,
Biocenter, Goethe Universität, Frankfurt/ Main and was used as received. Other chemicals and
supplies were obtained as described in the following sections.
4.5.1 Nickel functionalized diblock copolymer synthesis
This synthesis has been described in detail in a recent report (Nehring et al. 2009) and
therefore is described only briefly here. The polybutadiene-polyethyleneoxide diblock copolymer
(PB39-PEO36-O-) was synthesized via anionic polymerization. Poly (ethylene oxide)
polymerization was initiated by adding ethylene oxide to the living PB anions. After activation
with dicyclohexylcarbodiimide the polymeric carboxylic acid PB39-PEO36-SA-OH was reacted
with Lys-NTA. After deprotection and further purification, the yield of the functionalized block
copolymer was found to be 35%. To obtain the metal-polymer complex, the block copolymers
were stirred with a Nickel Chloride solution overnight until a green polymer solution was
obtained. This polymer is referred to as NTA polymer throughout this paper.
4.5.2 Triblock copolymer synthesis
The Polymethyloxazoline-Polydimethylsiloxane (PMOXA12PDMS55PMOXA12) block-
copolymer was synthesized by cationic ring-opening polymerization of 2-methyloxazoline using
a bi-triflate activated polydimethylsiloxane macroinitiator as described previously. The synthetic
route followed the one originally described in Nardin et al (Nardin et al. 2000). The molecular
weight of the polymer obtained from 1H NMR was 6325g/mol.
4.5.3 Membrane protein purification
Growth, isopropyl--D-thiogalactoside (IPTG) induction and Ni-NTA affinity
chromatography purification of histidine-tagged AqpZ were performed as described in Borgnia et
al (1999) with modifications described in Kumar et al (2007). The purified protein was
quantified using the Pierce 660-nm assay (Pierce Biochemical, Rockford, IL, USA). The
purified protein samples were dialyzed against elution buffer without imidazole prior to
experiments.
4.5.4 Monolayer formation studies
61

To determine appropriate conditions for formation of a stable diblock copolymer
monolayer at the air-water interface, film compression and Brewster Angle Microscopy (BAM)
techniques were used. Surface pressure-area (π-A) isotherms were obtained by using either a
Langmuir-Blodgett minitrough (surface area 273 cm2) or a BAM trough (surface area 432 cm2).
Both troughs were obtained from KSV Instruments Ltd., Helsinki, Finland. Prior to experiments,
the troughs were thoroughly cleaned with chloroform and ethanol, rinsed with double distilled
water, and filled with the aqueous subphase (either pure water or a phosphate buffer saline, 50
mM Na2HPO4, 10 mM KCl (Fluka AG, Buchs, Switzerland)), pH 7.4. The barriers were cleaned
with ethanol and rinsed with water. The Wilhelmy plate used in surface pressure measurements
was made of chromatography paper (ashless Whatman #1) with a perimeter of 20 mm. It was
equilibrated for at least 20 min. The surface was cleaned repeatedly through compression-
aspiration-expansion cycles and checked for impurities. The NTA polymer was spread dropwise
from chloroform solution (c = 1 mg/mL) on the air/water interface. The solvent was allowed to
evaporate for 10 min, and the monolayers were compressed at the rate of 10 mm/min. After each
measurement, the surface was cleaned and checked for impurities. All experiments were
performed at 20°C in a dust-free room, with the trough housed in a Plexiglas cabinet.
4.5.5 2 D crystallization studies
2D crystallization experiments were conducted in microtroughs built into Teflon block as
described in Lévy et al (Lévy et al. 1999). Each block had eight cylindrical troughs with a
diameter of 4 mm. Each well had a side port for addition of solutions and the overall volume
was approximately 60 microL. Citrate buffer (20 mM) containing 200 mM NaCl, 100 mM
MgCl2, 3 mM NaN3 and 10% glycerol at a pH of 7 was used for all the crystallization
experiments. A schematic of this process is provided in Figure 4.7. 60 L of buffer was first
added to each well to form a surface with a flat meniscus. To this 0.5 L of the templating
polymer dissolved in chloroform was added, targeting more than one but less than 2 monolayers
at a surface pressure equivalent to a mean molecular area of approximately 100 Å2 (determined
from Langmuir isotherm studies). This was equivalent to a concentration of 0.28 mg/ ml of
polymer in chloroform added to the surface of each cylindrical trough. A detailed calculation is
provided in Appendix B. The chloroform was left to evaporate for an hour.
62

A ternary mixture of ABA triblock copolymer, the detergent dodecyl maltoside (DDM),
and AqpZ protein was prepared and equilibrated overnight at 4 deg C. This mixture was then
added to the subphase of this monolayer. This mixture was designed to target a concentration of
100 g/mL of protein in the subphase. Polymer to protein ratios of between 0.2 and 4 were
screened and a 10% detergent (DDM) was necessary to solubilize ABA block copolymers. For
experiments with lipids, lipid to protein ratios of between 0.2 and 2 were screened. A 80%:20%
chicken egg phosphatidylcholine (PC)/ porcine brain phosphatidyl serine (PS) mixture and a
50%:50% 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC)/ 1-palmitoyl-2-oleoyl-sn-glycero-
3-phosphocholine (POPC) mixture was used for lipid experiments at lipid to protein ratios
between 0.2 to 2. A detergent concentration of 2% DDM was used to solubilize the lipids. After
addition of the ternary mixture the system was allowed to rest for 3-4 hours to allow binding of
the ternary micelles to the Ni2+ groups on the monolayer. Following this stabilization period
absorbent polymeric beads (Biobeads SM2) were added to the subphase at a dose of ten times the
detergent amount in the subphase, divided into equal batches of 6-8 additions over a 24-48 hour
period. Detergent removal drives self assembly of the protein polymer mixture.
water
NTA
PB-PEO-Ni NTA copolymer (in trough)
air
PB
PEO
Mixed Detergent, AqpZ, and
ABAmicelles
Slow detergent r
emoval
(Biobeads)
Ni2+Ni2+ Ni2+ Ni2+
airwater
Ni2+
Inject in subphase
Aquaporin Z
His tagHis tag
His tag
Dodecyl Maltoside(Detergent)
+
PMOXA-PDMS-PMOXA (block polymer)
PMOXA
PDMS
PMOXA
(a)
(b)
(c)
(d)
(e)
AqpZ 2D Crystalin block copolymer
NTA
Ni2+
NTA
Ni2+
NTA
Ni2+
water
NTA
PB-PEO-Ni NTA copolymer (in trough)
air
PB
PEO
Mixed Detergent, AqpZ, and
ABAmicelles
Mixed Detergent, AqpZ, and
ABAmicelles
Mixed Detergent, AqpZ, and
ABAmicelles
Slow detergent r
emoval
(Biobeads)
Ni2+Ni2+ Ni2+ Ni2+
airwater
Ni2+Ni2+ Ni2+ Ni2+ Ni2+Ni2+ Ni2+ Ni2+
airwater
Ni2+
Inject in subphase
Aquaporin Z
His tagHis tag
His tag
Aquaporin Z
His tagHis tag
His tag
Aquaporin Z
His tagHis tag
His tag
Aquaporin Z
His tagHis tag
His tag
Dodecyl Maltoside(Detergent)
+
PMOXA-PDMS-PMOXA (block polymer)
PMOXA
PDMS
PMOXA
Dodecyl Maltoside(Detergent)Dodecyl Maltoside(Detergent)
+
PMOXA-PDMS-PMOXA (block polymer)
PMOXA
PDMS
PMOXA
PMOXA-PDMS-PMOXA (block polymer)
PMOXA
PDMS
PMOXA
(a)
(b)
(c)
(d)
(e)
AqpZ 2D Crystalin block copolymer
NTA
Ni2+
NTA
Ni2+
NTA
Ni2+
Figure 4.7: Overall scheme for 2D crystallization of AqpZ in triblock copolymer using templating Ni2+ functionalized diblock copolymers.
63

The 2D crystals attached to the templating film were transferred onto non-glow
discharged Cu or Mo grids with thin carbon support for electron microscopy and negatively
stained with 1% uranyl acetate. Experiments were also performed with 40 g/ml AqpZ only (no
polymer or lipid) added to the monolayer subphase for comparison.
4.5.6 Electron Microscopy and electron diffraction
Electron microscopy was conducted on either a Phillips CM200 microscope at 120 keV
or on a JOEL 2100 microscope at 200 keV, both with LaB6 emitters. A liquid nitrogen cooled
Gatan Instruments Cryo-stage was used for cryo-electron microscopy with a FEI vitrobot used
for preparing hydrated frozen samples.
Single particle and templated AqpZ particles. AqpZ stock solutions, diluted to 25 microg/ml in
protein buffer, were applied to glow discharged copper grids and stained with 1% uranyl acetate
to visualize single AqpZ particles. AqpZ templated on monolayer (subphase concentration of ~
40 g/ml) was transferred onto non-glow discharged Cu grids and negative stained using 1%
uranyl acetate.
Electron microscopy of 2D Crystals and electron diffraction. 2D crystals were transferred onto
grids by depositing grids directly onto the monolayer for 2 minutes. The grids were then washed
twice with double disitilled water and stained with 1% uranyl acetate. Electron diffraction of
selected areas was recorded using Selected Area Diffraction apertures to select appropriate areas
or using the nanobeam diffraction mode for smaller diffracting areas.
Cryo-electron microscopy. In order to obtain images that were free from staining artifacts and
that may be suitable for further image processing 2D crystal solutions were embedded in a 2%
glucose or trehalose solution and loaded into the electron microscope using a liquid nitrogen
cooled cryo-stage. Electron diffraction patterns at low temperatures were recorded using a CCD
camera at appropriate exposure times to resolve the maximum number of spots possible.
64

5 SUMMARY, IMPLICATIONS, AND FUTURE RESEARCH
This work was the first to demonstrate the potential of biomimetic membranes for
desalination, has led to the discovery of pH gating in bacterial aquaporins, and has provided an
exciting first step in the determination of membrane protein structure in biomimetic membranes.
It has also led to extensions that have applications in understanding membrane protein insertion
in block copolymers, membrane protein gas transport studies, perchlorate removal, carbon
capture and. This chapter summarizes these outcomes of my dissertation research, looks at its
broad relevance, introduces some additional projects that resulted from this work, and provides
suggestions on future research.
5.1 HIGH PERMEABILITY AQPZ RICH MEMBRANE CAN MAKE IDEAL
DESALINATION MEMBRANES AND HIGH SENSITIVITY SENSORS.
This work started with the objective of inserting aquaporins into biomimetic block
copolymers to develop an alternative to current, polyamide-based, thin film composite RO
membranes. Aquaporins had been inserted into block copolymers prior to this study but their
activity was not tested (Stoenescu et al. 2004). I have demonstrated that the bacterial aquaporin,
Aquaporin Z, can be functionally inserted into an ABA-type, PMOXA-PDMS-PMOXA
membrane. Upon insertion of AqpZ, the permeability of this membrane increased by
approximately three orders of magnitude (protein to polymer molar ratio of 1:50, Figure 5.1).
Additionally, these membranes were shown to reject salt, urea, glycerol and glucose.
Flat membranes are most common for desalination and sensor applications. However,
most work with PMOXA-PDMS-PMOXA membranes with inserted membrane proteins has
been performed on the vesicle configuration (Nardin et al. 2000; Graff et al. 2002; Ho et al.
2004; Stoenescu et al. 2004; Kumar et al. 2007; Onaca et al. 2008; Graff et al. 2009;
Grzelakowski et al. 2009). A large part of this work was focused on developing a flat
configuration of AqpZ-rich block copolymer membranes. This is described in Chapter 4. The
highly packed polymer films achieved in these studies were small (up to 500 nm) and further
optimization would be needed to develop larger areas that would be practical for applications.
65

Figure 5.1: a) Insertion of AqpZ into block copolymer membranes increases permeability by an order of magnitude. b) AqpZ based membranes have higher permeability than currently used membranes. FO is commercial forward osmosis membrane, while RO is commercial reverse osmosis desalination membrane. PB-PEO is Polybutadiene Polyethylene diblock polymer. ABA represents the polymer vesicles used in study and ABA+ AqpZ represents the polymer vesicles with incorporated AqpZ at 1:50 molar ratio (see Section 2 for details).
5.1.1 Extensions
Determining insertion efficiency of membrane proteins in block copolymers. A major
experimental unknown regarding insertion of membrane proteins in lipid or polymer membranes
is the question of how many membrane proteins are inserted per unit membrane area. Efforts are
currently underway in our research group to covalently label individual aquaporins with suitable
fluorophores and determine the number of aquaporins inserted per vesicle using Fluorescence
Correlation Spectroscopy techniques modified from those presented in Rigler et al (Rigler and
Meier 2006). This work will have relevance to determining the optimum strategy for inserting
membrane proteins in block polymers. More broadly, it would provide clues to understanding
the phenomenon of membrane-membrane protein hydrophobic mismatch, its role in limiting
insertion of membrane proteins, and its possible effect on the activity of inserted membrane
proteins (Andersen and Koeppe 2007; Graff et al. 2009).
66

5.1.2 Implications
This work is proof of principle for using a biomimetic approach to develop vastly
improved desalination membranes. I have shown that incorporating AqpZ molecules into block
copolymers can increase the permeability of these polymers by orders or magnitude. More
importantly, this enhanced permeability is approximately 2-3 orders of magnitude higher than
commercial desalination membranes (Figure 5.1 (b)) while maintaining similar or higher salt
rejection capabilities. Additionally, the polymer used for this part of the study was highly
impermeable when compared to other block copolymers such as the PB-PEO membrane (Figure
5.1 (b)). This ensures that after protein insertion, flow is mostly through aquaporins and not the
membrane matrix, thus decreasing the possibility of solute leakage through the membrane
matrix. All of the above point to the promise of developing block copolymers with inserted
Aquaporins as desalination membranes.
High density membrane protein arrays may have practical applications in sensing. I
have demonstrated that protein packed structures including vesicles and small flat sheets can be
assembled. Such high density arrays have relevance to applications where a high concentration
of membrane proteins would be beneficial. In sensor applications based on membrane proteins
(Cornell et al. 1997; Martin 2007; Nielsen 2009) increased sensitivity could result from the
increased signal to noise ratio that would result from “multiplexing” multiple membrane
proteins. Similarly, in transport applications, a high density of membrane proteins would result
in enhanced transport (Kumar et al. 2007; Nielsen 2009).
5.1.3 Future Work
I have shown proof-of-concept for use of block copolymer membranes for desalination.
Further, by making templated, protein-rich films; I have been able to show that a large amount of
aquaporins can be packed into polymer films. This makes such a system attractive in terms of
the permeabilities that can be expected. However, scale up of these systems and development of
appropriate support systems for molecularly thin membranes will be a major challenge. Some
steps were made in this direction using a templating approach described in Chapter 4. However,
other efforts, including those where immobilized or deposited vesicles could be used to form
membranes, should also be pursued as these may have the ability to cover larger areas (albeit at a
lower AqpZ density). Several filtration experiments were conducted with AqpZ rich vesicles
67

deposited on an alumina filter. No rejection of salts was observed. This may be due to the lack
of sealing between the vesicles and filter pores. Thus methods where proper sealing can be
achieved should be developed.
Another question that needs to be addressed is the stability of the protein-polymer system
developed in this work. It has been reported that the chemical and mechanical stability of block
co-polymers are superior to lipid bilayer membranes. This claim is supported by studies on
bending modulus where block copolymer vesicles were aspirated into pipettes and the
deformation measured (Discher et al. 1999). Other methods should be used to further quantify
the mechanical properties and stability of block copolymer membranes. The stability of block
copolymers can be greatly enhanced by incorporating crosslinkable groups such as methacrylate
and performing crosslinking reactions. In this work (reported in Appendix C) I have tested a
method to determine the rate and extent of UV crosslinking of methacrylated PMOXA-PDMS-
PMOXA polymers using Fourier Transform Infrared (FTIR) Spectroscopy. However, these
experiments were conducted on solid polymer films. Vesicle stability tests should be conducted
before and after crosslinking. An approach to determining mechanical stability of such vesicles
in an aqueous suspension would be to evaluate change in size and/or polydispersivity (indicating
rupture) by light scattering after subjecting the vesicle suspension to varying shear rates in a cone
on plate viscometer. Additionally, osmotic shock of different magnitudes can be used to
determine the “burst pressure” of these vesicles. This experiment however needs a careful
control on the rate of mixing (such as that achieved in a stopped flow apparatus) as both the
magnitude and rate of application of osmotic pressure is relevant to bursting of vesicles. Also
such experiments could be done on vesicles stored under different conditions and over time to
determine stability.
More broadly, this work could be extended to incorporate membrane proteins with specific
transport and sensing mechanisms. Specific applications that I have begun exploring are CO2
capture (described in section 5.2.3) and perchlorate transport and degradation (described in
section 5.4.1). In terms of sensing the most promising membrane protein candidates would be
the ones currently being engineered by the Bayley group for sensing various analytes such as
organic molecules, divalent ions (Gu et al. 1999; Braha et al. 2000; Bayley and Cremer 2001)
and even DNA sequencing (Howorka et al. 2001).
68

5.2 TRIBLOCK COPOLYMERS CAN BE USED AS A PLATFORM FOR STUDYING
MEMBRANE PROTEIN PROPERTIES.
In this work ABA block copolymers were used to study the response of AqpZ to pH.
Permeability of AqpZ-ABA membranes decreased with pH from the neutral range to an acidic
range (Figure 7.2 (a)). This permeability decrease was found to be reversible with subsequent
increase in pH indicating an AqpZ gating mechanism. This gating was demonstrated in E coli
cells, and it is physiologically relevant based on experiments with wild type and mutant E. coli
strains. E. coli cells overexpressing aqpZ exhibited reduced survival under both acid and
simultaneous acid and hypoosmotic shock.
Figure 5.2. (a) pH gating of AqpZ observed in block copolymer. It was seen that the permeability of block copolymer vesicles decrease reversibly between pH 6 and 4. (b) AqpZ – ABA vesicles are stable over a wide pH range as seen from light scattering results on vesicles stored overnight at the indicated pH values (x-axis).
5.2.1 Extensions
Study of gas transport in membrane proteins using block co-polymers. Membrane
protein-mediated water and ion transport is studied by incorporation into lipid bilayers.
However, gas permeabilities of lipids are high, making it difficult to conclusively show gas
transport of incorporated proteins (Hill et al. 2004). In particular, CO2 transport across
aquaporins has been a controversial topic (Verkman 2002). The high gas permeability of lipid
membranes should preclude the need for specialized membrane proteins. Additionally, the use
of AQPs reconstituted in lipids for permeability assays has further contributed to this controversy
69

because the observed differences in permeability between lipids alone and lipids with aquaporins
is small . We are studying CO2 transport in block copolymers with incorporated aquaporins. In
preliminary studies, the CO2 permeabilities of these membranes were orders of magnitude lower
than lipid bilayers (10-6 cm/s vs. 10-3 cm/s) making this a sensitive system. In future studies
tailoring of membrane permeabilities using specific membrane proteins could be investigated.
5.2.2 Implications
Triblock copolymers can provide a stable platform for studying membrane protein
properties. PMOXA-PDMS-PMOXA polymer was used to study the permeability of AqpZ at
pH values from 2 to 12. The vesicles formed at these different pH values were stable (Figure 5.2
(b)) and even survived high pressure (~8 bar) stopped flow experiments followed by recovery
using centrifugation assisted membrane concentration. These results are a definite improvement
over lipid vesicles. In particular, lipids have high permeability for water and gases, making it
difficult to study transport of these molecules (Hill et al. 2004; Wang et al. 2007). Lipid vesicles
can also be strongly affected by pH, ionic strength, and temperature (Grit and Crommelin 1993)
complicating studies that need to be conducted under these conditions. The use of biomimetic
block copolymers to investigate a broader range of membrane protein properties provides an
excellent alternative to the lipid-based system. Further, the use of stable polymers makes it
feasible for such hybrid protein polymer systems to be used in engineering applications.
pH gating of microbial AqpZ is physiologically relevant and may be important for cell
survival in mammalian digestive tracts. Many microorganisms contain aquaporins, and some
microbial aquaporins transport water rapidly and specifically (Calamita et al. 1995; Bonhivers et
al. 1998; Hernandez-Castro et al. 2003; Shapiguzov et al. 2005), just like their eukaryotic
counterparts. However, because microorganisms have large surface area to volume ratios,
allowing fast diffusive transport, the physiological relevance of channel-mediated water transport
in microorganisms is unknown (Hill et al. 2004; Tanghe et al. 2006). Genetic analysis has not
resolved the issue, as elimination of microbial aquaporins typically results in subtle defects
(Calamita et al. 1998; Soupene et al. 2002; Hernandez-Castro et al. 2003; Shapiguzov et al.
2005), and in some cases even improves growth (Bonhivers et al. 1998). Experiments with
AqpZ mutants showed that AqpZ gating may have a role in bacterial survival under conditions of
hypoosmotic and acid shock. Figure 5.3 presents a proposed model for the role of Aquaporins
70

in preventing cell death when exposed to combined acid and osmotic shock. The acid and
combined acid and osmotic shock conditions considered here for whole cells are similar to
conditions E. coli and other pathogens might be exposed to upon passage through a host
digestive system; therefore, insights into bacterial survival under these conditions have relevance
to public health and food safety (Diez-Gonzalez et al. 1998).
Figure 5.3: Model for coordinated activity of aquaporins and mechanosensitive channels under pH and osmotic downshift conditions. Upon hypoosmotic shock aquaporins mediate rapid water transport into the cell, leading to rapid swelling. To prevent cells from bursting, mechanosensitive channels open and allow small solutes to be released. Under acid shock, mechanosensitive channel activation is lowered. The cell is therefore especially vulnerable to combined acid and hypoosmotic shock, unless water permeability of AqpZ is also reduced under acidic conditions.
5.2.3 Future Work
This work has shown that using block copolymers to incorporate membrane properties
allows flexibility in investigating a broader range of properties for these important biomolecules.
Using pH-stable block copolymers led to the discovery of pH gating in AqpZ in block copolymer
membranes and confirmation of its existence in E. coli cells. pH gating has been reported in
71

eukaryotic aquaporins, however the residues involved in gating in these aquaporins are not
conserved in AqpZ (Figure 5.4). Further work on determining the molecular mechanism of
AqpZ should be conducted as it is not clear from its structure and homology with other gated
aquaporins what residues could be involved in this behavior. Single point mutations of
protonated residues with pKa’s in the physiological range, including Histidine residues –His 56,
His 61, His 124, His 150, and His 174 should be conducted and the resulting strains should be
subjected to permeability measurements under different pH conditions to determine if pH gating
can be abolished by change in one or more residues. This will provide an indication of which
residues could be involved in the observed pH gating. Additionally, these mutated proteins
could be purified and reconstituted into block copolymer vesicles to provide further
demonstration of gating and the residues involved in gating.
Similar work could use this proposed system to study relevant properties of other
membrane proteins. An interesting case would be to study mechanosensitive channels in block
copolymers. Since the distribution of membrane stresses in a block copolymer would be
different than in a lipid membrane due to the difference in hydrophobic mismatch magnitudes, it
would be interesting to correlate the relationship between osmotic pressure required for pore
activation with the level of hydrophobic mismatch. This will provide more insight into the
gating of mechanosensitive channels and the mechanisms of osmoprotection provided by these
important membrane proteins.
Another research area would be to find candidate membrane proteins where the catalytic
activity of the protein could be enhanced from stresses induced by hydrophobic mismatch. An
excellent recent example is the NADH Ubiquinone Oxoreductase protein that was found to have
increased activity (~two times) with increasing hydrophobic block thickness (from 20 to 120
units of PDMS) (Graff et al. 2009).
5.3 BLOCK COPOLYMERS CAN BE USED FOR DETERMINING PROTEIN
STRUCTURE.
In addition to providing a platform for incorporation of membrane protein, these polymers
may provide a method for determining protein structure. In this work, a PB-PEO diblock
copolymer with a nickel functionalized hydrophilic block (PEO) (Nehring et al. 2009) was used
to provide a stable interfacial film for immobilizing AqpZ or mixed micelles of AqpZ, detergent
72

and ABA polymer. Immobilized AqpZ could be directly imaged on these films using electron
microscopy (Figure 5.4(a)). Additionally, when detergent was slowly removed, flat AqpZ
packed layers of ABA polymer were formed. These structures diffract electrons and seem to
indicate a tetragonal crystal structure (Figure 5.4(b)). A tetragonal crystal structure may be
indicative of AqpZ crystals as all reported aquaporin 2D crystals have a tetragonal unit cell
(Ringler et al. 1999; Braun et al. 2000; Gonen et al. 2004; Kukulski et al. 2005; Schenk et al.
2005; Viadiu 2007; Viadiu 2007; Ho et al. 2009). 2D crystal formation of AqpZ in block
copolymers, if confirmed, would be a major breakthrough as described in 5.3.1.
Figure 5.4: (a) Templating of AqpZ tetramers on Ni NTA block copolymer film. Inset shows a schematic of the templating system. (b) 2 D crystals of AqpZ formed in PMOXA PDMS triblock copolymer using templating block copolymers. Inset shows the electron diffraction pattern indicating a tetragonal crystal structure.
5.3.1 Implications
In this study membrane proteins could be immobilized on the surface of a chelating
diblock polymer monolayer or within the matrix of a triblock copolymer. Both of these findings
have implications for protein structure determination as described in the paragraphs below.
Membrane proteins selectively immobilized on the surface of a stable polymer
monolayer can be used to determine structure using single-particle cryo-electron microscopy.
73

The Ni-NTA polymer could be used to successfully immobilize AqpZ molecules (Figure 5.4
(b)). This has excellent application for preparing samples for imaging using single particle,
three-dimensional, cryogenic electron microscopy (cryo-EM). In single particle cryo-EM,
samples deposited on a Transmission Electron Microscopy (TEM) grid are plunge-frozen in
liquid ethane to form a thin layer of vitreous ice thus preserving the native hydrated structure of
biological macromolecules. Images of single macromoelcules can then be taken under cryogenic
conditions and averaged to obtain 3D structures of molecules. To this effect, Kelly et al have
developed the concept of an “affinity grid” where Ni –chelating lipids are deposited onto a TEM
grid, dried and stored (Kelly et al. 2008; Kelly et al. 2008a). This grid can then be directly
placed on a small sample of a crude cell lysate with added imidazole to directly recover Histidine
tagged proteins for direct imaging using cryo-EM. The Ni NTA polymer used in this work
forms a stable templating monolayer and is an excellent candidate to be used similarly. The
higher molecular weight of this polymer indicates a higher detergent resistance and overall
higher mechanical and chemical stability. For the “affinity grid” loss of the lipid monolayer to
glycerol and/or detergents limits some of the functionality of this method (Kelly et al. 2008a).
Preliminary work with these grids using polymer monolayers seems to indicate high stability
when used for membrane protein templating.
2D crystallization of membrane protein arrays in block copolymers may be a stable
platform for membrane protein structure determination. 2D crystallization of proteins in block
copolymers is an important finding of this work. 2D protein-crystals, after careful imaging using
low dose cryo-electron microscopy, can be used to determine the 3D structure of membrane
proteins in synthetic membranes (Henderson and Unwin 1975; Hankamer et al. 2007). In single
particle 3D cryo-EM each particle needs to be large enough and present in different orientations
to allow reconstruction. Conversely, 2D crystals present perfectly aligned particles in a grid,
making the mathematical process of structure reconstruction easier and possible for smaller
membrane proteins. Thus, the ability to form 2D crystals opens the door to structure
determination of membrane proteins in these polymers. The structure of AqpZ and many other
membrane proteins is known in lipids. Lipids represent the native state of membrane proteins
and structures obtained are considered the native structures of these important proteins. In this
work and future work, membrane proteins inserted in block copolymers is envisaged as a
platform for developing technologies for environmental and other applications. Further studies
74

need to be conducted to determine the structure of various candidate membrane proteins in these
materials in order to determine the suitability and properties for specific membrane-protein based
devices. Since membrane protein structure directly dictates function a careful structural
comparison of membrane proteins in lipids and the block polymer system could allow us to
explain differences in activity, or more importantly design increases or variations in activity of
these proteins (Andersen and Koeppe 2007; Graff et al. 2009). Moreover, systems could be
designed to provide conformational strain on incorporated proteins to obtain desired function or
level of activity.
5.3.2 Future Work
I have obtained promising results for crystallizing membrane proteins in triblock
copolymers obtained using the templating approach. Electron diffraction of these samples
indicates that there is crystallinity in the samples that may be a result of AqpZ crystallization,
particularly because the diffraction pattern points towards a p4 crystal such as that expected
from an Aquaporin 2D crystal (Ringler et al. 1999). However, as discussed in Chapter 4, while
promising, the images and diffraction patterns are a bit unusual. Further work should include
confirming that the crystals formed are indeed protein-polymer 2D crystals, or just pure polymer
2D crystals. If final structure of the protein in block copolymers is to be determined, additional
efforts should be directed towards making larger crystals, using other methods including dialysis.
Alternative methods of transfer to the TEM grid and better imaging techniques including the use
of cryogenic and low-dose electron microscopy techniques under various tilt conditions should
be utilized to obtain information that could lead to structure determination.
Accurate structure determination of proteins in polymers can provide a molecular explanation for
anomalous effects seen after incorporation of membrane proteins into block copolymers and
provide a basis for design of novel functional materials. Thus, attempts should be made to from
2D crystals of such proteins where a large change in conformation is suspected from activity
measurements, and that are of interest for applications.
Validation of the potential of the affinity grid technique utilizing stable Ni-NTA polymers would
to contribute to fundamental science of single particle 3D cryo-EM. In this regard, the
templating polymer should be further optimized with proteins of known structure and amenable
to 3D cryo-EM.
75

5.4 MEMBRANE VESICLES CAN BE USED FOR CONDUCTING REACTIONS ON
SURFACES, ENVIRONMENTAL REMEDIATION AND CARBON CAPTURE.
Immobilized vesicles can be used for creating surfaces with novel properties that can be
used to engineer systems for applications. In order to illustrate this concept, I participated in
collaborative work on vesicles permeabilized using a membrane protein (OmpF) and containing
the enzyme acid phosphatase. Further, the block copolymer vesicle membrane contained biotin,
and use of biotin-avidin interactions allowed controlled patterning of these vesicles on a surface
(summarized in Figure 5.5). This system can be considered an analogue of biofilm reactors
commonly used in environmental applications, where cells immobilized on surfaces are used for
degrading contaminants. Use of a fluorescent substrate enabled measurement of reaction rates by
monitoring fluorescence change. This work has been published (Grzelakowski et al. 2009) and
demonstrates how a biofilm-reactor mimic can be designed using block copolymer vesicles with
active enzymes.
Proteo-vesicle
Streptavidin
Biotin
Acid phosphatase
Elf 97® substrateBSA
Figure 5.5: Immobilized nanoreactors. Polymer nanoreactors are permeabilized with the channel OmpF and encapsulate the enzyme acid phosphatase. The enzyme substrate (ELF 97) enters the nanoreactor through OmpF and is converted by the acid phosphatase to a fluorescent product which can be monitored to determine reaction rate. The hydrophilic block of the triblock copolymer forming the nanoreactor is functionalized with biotin. A glass surface is functionalized with bovine serum albumin – biotin which binds streptavidin which then forms the link between this surface and the vesicle surface immobilizing it.
76

5.4.1 Extensions
Nanoreactors for environmental remediation of perchlorate. In this work, we have
encapsulated perchlorate degrading enzymes from specialized bacteria into block copolymer
vesicles. We also incorporated a purified outer membrane channel protein (OmpF) to allow
perchlorate to enter these vesicles (Figure 5.6). Work is currently underway to demonstrate that
perchlorate enters these vesicles through OmpF channels and is degraded. Future work includes
purification and insertion of a specific, active perchlorate transporter – the Sodium Iodide
Symporter (NIS). NIS transports perchlorate against its concentration gradient (uphill transport)
by coupling its transport to a downhill sodium gradient. Thus, the use of this transporter will
allow a high level of control of concentration of perchlorate within these nanoreactors based on
the sodium gradient utilized. The proposed system may have several advantages over currently
used biological treatment approaches as discussed in the following subsection. Further, the
ability to immobilize these nanoreactors to create a perchlorate degrading surface would be a
direct application for environmental engineering.
Figure 5.6: Perchlorate reducing nanoreactors
5.4.2 Implications
Environmental remediation using functionalized, immobilized vesicles could be an
excellent alternative to traditional biological treatment. These hybrid biological/synthetic
systems could be used for contaminants for which biological treatment is the only sustainable
option and where maintaining suitable reactor conditions is challenging. Additionally, the
77

system described above provides the flexibility of using genetically engineered proteins without
the risk of accidental release and propagation of genetically modified DNA in the environment.
Use of a CO2 impermeable block-copolymer membrane for inserting membrane
proteins could help in developing technologies for carbon capture. Our ongoing work with
block copolymer gas permeability suggests that these materials might have extremely low
permeabilities for gas. Incorporating an active transporter for CO2 can thus lead to concentration
of CO2 inside a block copolymer based vesicle. Enzymes could then convert this CO2 into
organic compounds and lead to CO2 capture. This idea is developed further in the next
subsection.
The approach proposed in this subsection could be applied in for any compound where a
specific transporter exists. Additionally, the block copolymer vesicle system allows us to create
cells with a combination of functions that do not exist in nature. This is illustrated by the use of
NIS (a mammalian protein) to concentrate perchlorate while using bacterial enzymes to degrade
it in a single synthetic cell.
5.4.3 Future Work
The feasibility of perchlorate reduction is being studied using nanoreactors with
encapsulated cell derived enzymes. Further work should include the purification of NIS, its
insertion into these block copolymers and activity studies. The insertion of membrane proteins
in the ABA block polymer system could be in either direction. Unlike the bidirectional AqpZ
and OmpF proteins, when directional active proteins (such as NIS) are inserted into such a
system, there will be difficulty in maintaining a stable concentration within these nanoreactors as
there will be constant influx as well as efflux of perchlorate over small time periods depending
on the concentration gradients set up. However, asymmetric ABC-type polymers can be used for
directional insertion and would be valuable in such cases. For the perchlorate system, the
enzymes used also need to be in a reduced state. Thus approaches to provide an external electron
donor should be investigated.
The proposed approach should be extended to other micropollutants or mixtures of
pollutants in order to understand the broad applicability of this technology. Additionally, on the
fundamental side, the effect of confinement of enzymes in a small volume should be further
studied to determine the effect of molecular crowding on enzyme activity. Further, use of
78

reactive surfaces made with such nanoreactors should be further investigated with environmental
applications; scale-up of such systems should be studied.
Ongoing work on gas transport in aquaporins incorporated into block copolymer could be
extended to develop a system for CO2 capture and conversion. This work would mimic the CO2
concentrating mechanisms (CCM) in plant cells and cyanobacteria. The central feature of this
CCM is the organelle carboxysome which is enclosed in a highly impermeable membrane.
Concentration within organelles is achieved with the use of active carbon dioxide transporters
such as the Na+/HCO3- transporter (Romero et al. 1997). This could be mimicked by inserting
the Na+/HCO3- transporter into block copolymer vesicle membranes to create a carboxysome
mimic. Incorporating the enzyme Rubisco within these vesicles could then convert CO2 into
long chain carbohydrates similar to the functioning of a carboxysome. This proposed scheme is
summarized in Figure 5.7.
Rubisco
HCO3‐
CO2
RubP
PGA+
Rubisco
HCO3‐
CO2
RubP
PGA+
Figure 5.7. Carboxysome- mimic vesicles. Carbonic anhydrase (CA) first converts dissolved CO2 into bicarbonate which could be transported into the centre of the organelle using active bicarbonate transporter channel proteins. The CO2 accumulated in this impermeable “artificial organelle” can then be combined with short chain sugars such as Ribulose Phosphate (RubP) and converted to long chain sugars such as phophphoglycolate (PGA) by the encapsulated enzyme Rubisco.
79

5.5 CONCLUSION
This work has demonstrated the promise of using hybrid membrane protein-polymer
systems for environmental engineering applications. In particular, the applicability to synthetic
desalination membranes is most promising and relevant. The basic approach used here may be
applied to any separation for which a specific transport protein is available or could be
engineered. It has also contributed to understanding the properties of aquaporins, in particular
AqpZ and its possible role in microbial physiology. Recent successes in immobilizing protein
molecules and in synthesizing 2D crystals of membrane proteins may provide an excellent path
to answer fundamental questions regarding the structure and function of these membrane
proteins in block copolymers.
80

6 REFERENCES
Agmon, N. (1995). The grotthuss mechanism. Chemical Physics Letters. 244: 456-462.
Agre, P. (2004). Nobel Lecture. Aquaporin water channels. Bioscince Rep 24(3): 127-63.
Agre, P. and D. Kozono (2003). Aquaporin water channels: molecular mechanisms for human
diseases1. FEBS letters. 555: 72-78.
Almgren, M., K. Edwards and G. Karlsson (2000). Cryo transmission electron microscopy of
liposomes and related structures. Colloids and Surfaces a-Physicochemical and
Engineering Aspects 174(1-2): 3-21.
Andersen, O. S. and R. E. Koeppe (2007). Bilayer thickness and membrane protein function: an
energetic perspective. Annual Review of Biophysics and Biomolecular Structure 36: 107-
130.
Astier, Y., H. Bayley and S. Howorka (2005). Protein components for nanodevices. Current
opinion in chemical biology 9(6): 576-584.
Baba, T., T. Ara, M. Hasegawa, Y. Takai, Y. Okumura, M. Baba, K. Datsenko, M. Tomita, B.
Wanner and H. Mori (2006). Construction of Escherichia coli K-12 in-frame, single-gene
knockout mutants: the Keio collection. Molecular systems biology 2(1).
Bayley, H. and P. S. Cremer (2001). Stochastic sensors inspired by biology. Nature. 413: 226-
230.
Berrier, C., A. Coulombe, I. Szabo, M. Zoratti and A. Ghazi (1992). Gadolinium ion inhibits loss
of metabolites induced by osmotic shock and large stretch-activated channels in bacteria.
European Journal of Biochemistry 206(2): 559-565.
Bohdziewicz, J., M. Bodzek and E. Wasik (1999). The application of reverse osmosis and
nanofiltration to the removal of nitrates from groundwater. Desalination 121(2): 139-147.
Bonhivers, M., J. Carbrey, S. Gould and P. Agre (1998). "Aquaporins in Saccharomyces:
Genetic and Functional Distinctions between laboratory and wild-type strains." Journal of
Biological Chemistry 273(42): 27565-27572.
Borgnia, M., D. Kozono, G. Calamita, P. Maloney and P. Agre (1999). Functional reconstitution
and characterization of AqpZ, the E. coli water channel protein. Journal of Molecular
biology 291(5): 1169-1179.
81

Braha, O., L. Gu, L. Zhou, X. Lu, S. Cheley and H. Bayley (2000). Simultaneous stochastic
sensing of divalent metal ions. Nature Biotechnology 18(9): 1005-1007.
Braun, T., A. Philippsen, S. Wirtz, M. J. Borgnia, P. Agre, W. Kuhlbrandt, A. Engel and H.
Stahlberg (2000). The 3.7 Angstrom projection map of the glycerol facilitator GlpF: a
variant of the aquaporin tetramer. European Molecular Biology Organization Reports
1(2): 183.
Calamita, G. (2000). The Escherichia coli aquaporin-Z water channel. Molecular Microbiology
37(2): 254-262.
Calamita, G., W. R. Bishai, G. M. Preston, W. B. Guggino and P. Agre (1995). Molecular
cloning and characterization of AqpZ, a water channel from Escherichia coli. Journal of
Biological Chemistry. 270: 29063 -29066.
Calamita, G., B. Kempf, M. Bonhivers, W. R. Bishai, E. Bremer and P. Agre (1998). Regulation
of the Escherichia coli water channel gene aqpZ. Proceeding of the National Academy of
Sciences of the USA 95(7): 3627-31.
Chakrabarti, N., E. Tajkhorshid, B. Roux and R. Pomes (2004). Molecular basis of proton
blockage in aquaporins. Structure 12(1): 65-74.
Cheryan, M. (1998). Ultrafiltration and microfiltration handbook, CRC Press.
Choi, H. J., J. Germain and C. D. Montemagno (2006). Effects of different reconstitution
procedures on membrane protein activities in proteopolymersomes. Nanotechnology
17(8): 1825-1830.
Choi, H. J. and C. D. Montemagno (2005). Artificial organelle: ATP synthesis from cellular
mimetic polymersomes. Nano Letters. 5: 2538-2542.
Cornell, B. A., V. L. B. Braach-Maksvytis, L. G. King, P. D. J. Osman, B. Raguse, L. Wieczorek
and R. J. Pace (1997). A biosensor that uses ion-channel switches. NATURE: 580-582.
Coury, L. A., J. C. Mathai, J. L. Brodsky, P. Agre and M. L. Zeidel (1996). Use of yeast
secretory (sec) vesicles to express and characterize aquaporin (AQP) 1 and 2 water
channels. Journal of the American Society of Nephrology 7(9): A0088-A0088.
de Groot, B. L. and H. Grubmuller (2001). Water permeation across biological membranes:
mechanism and dynamics of aquaporin-1 and GlpF. Science. 294: 2353-2357.
del Pino, M. and B. Durham (1999). Wastewater reuse through dual-membrane processes:
opportunities for sustainable water resources. Desalination 124(1): 271-277.
82

Diez-Gonzalez, F., T. R. Callaway, M. G. Kizoulis and J. B. Russell (1998). Grain feeding and
the dissemination of acid-resistant Escherichia coli from cattle. Science 281(5383): 1666-
8.
Dinsmore, A. D., M. F. Hsu, M. G. Nikolaides, M. Marquez, A. R. Bausch and D. A. Weitz
(2002). Colloidosomes: selectively permeable capsules composed of colloidal particles.
Science 298(5595): 1006-1009.
Discher, B., Y. Won, D. Ege, J. Lee, F. Bates, D. Discher and D. Hammer (1999).
Polymersomes: tough vesicles made from diblock copolymers. Science 284(5417): 1143.
Discher, D. E. and A. Eisenberg (2002). Polymer vesicles. Science 297(5583): 967-73.
Dolder, M., A. Engel and M. Zulauf (1996). "The micelle to vesicle transition of lipids and
detergents in the presence of a membrane protein: Towards a rationale for 2D
crystallization." FEBS Letters 382(1-2): 203-208.
Duncan, R. (2003). "The dawning era of polymer therapeutics. Nature Reviews:Drug Discovery
2(5): 347-360.
Einav, R., K. Harussi and P. D. (2002). The footprint of desalination processes on the
environment. Desalination 152: 141-154.
Engel, A., Y. Fujiyoshi, T. Gonen and T. Walz (2008). Junction-forming aquaporins. Current
opinion in structural biology. 18: 229-235.
Finkelstein, A. (1987). Water movement through lipid bilayers, pores and plasma membranes.
Theory and reality, John Wiley and Sons.
Foster, J. (2000). Microbial responses to acid stress. Bacterial Stress Responses. G. Storz,
Hengge-Aronis, R. Washington, DC, ASM Press: 99–115.
Garybobo, C. M. (1971). Effect of Geometrical and Chemical Constraints on Water Flux across
Artificial Membranes. Journal of General Physiology 57(5): 610-622.
Goldstein, J. (2006). Sustainable water supplies with wastewater recycling. Biocycle 47(1): 24-
25.
Gonen, T., P. Sliz, J. Kistler, Y. Cheng and T. Walz (2004). Aquaporin-0 membrane junctions
reveal the structure of a closed water pore. Nature 429(6988): 193-197.
Gonzalez-Perez, A., K. B. Stibius, T. Vissing, C. H. Nielsen and O. G. Mouritsen (2009).
Biomimetic Triblock Copolymer Membrane Arrays: A Stable Template for Functional
Membrane Proteins. Langmuir. 25: 10447-10450.
83

Gorin, M. B., S. B. Yancey, J. Cline, J.-P. Revel and J. Horwitz (1984). The major intrinsic
protein (MIP) of the bovine lens fiber membrane: Characterization and structure based on
cDNA cloning. Cell 39(1): 49-59.
Grabmueller, H. (2010).
http://www.mpibpc.mpg.de/home/grubmueller/teaching/past_Lectures/WS2007_2008/pr
actical_05/index.html. accesed on May 9th, 2010.
Graff, A., C. Fraysse-Ailhas, C. G. Palivan, M. Grzelakowski, T. Friedrich, C. Vebert, G.
Gescheidt and W. Meier (2009). Amphiphilic Copolymer Membranes Promote NADH:
Ubiquinone Oxidoreductase Activity: Towards an Electron-Transfer Nanodevice.
Macromolecular Chemistry and Physics. 211: 229-238.
Graff, A., M. Sauer, P. Van Gelder and W. Meier (2002). Virus-assisted loading of polymer
nanocontainer. Proceedings of the National Academy of Sciences of the United States of
America. 99: 5064-.
Grit, M. and D. J. A. Crommelin (1993). Chemical stability of liposomes: implications for their
physical stability. Chemistry and Physics of Lipids 64(1-3): 3-18.
Grzelakowski, M., O. Onaca, P. Rigler, M. Kumar and W. Meier (2009). Immobilized Protein-
Polymer Nanoreactors. Small 5(22): 2545 -2548.
Gu, L., O. Braha, S. Conlan, S. Cheley and H. Bayley (1999). Stochastic sensing of organic
analytes by a pore-forming protein containing a molecular adapter. Nature 398(6729):
686-690.
Hankamer, B., R. Glaeser and H. Stahlberg (2007). Electron crystallography of membrane
proteins. Journal of structural biology 160(3): 263.
Hashido, M., M. Ikeguchi and A. Kidera (2005). Comparative simulations of aquaporin family:
AQP1, AQPZ, AQP0 and GlpF. FEBS letters. 579: 5549-5552.
Hedfalk, K., S. Tornroth-Horsefield, M. Nyblom, U. Johanson, P. Kjellbom and R. Neutze
(2006). Aquaporin gating. Current Opinion on Structural Biology 16(4): 447-56.
Henderson, R. and P. N. T. Unwin (1975). Three-dimensional model of purple membrane
obtained by electron microscopy. Nature. 257: 28-32.
Hernandez-Castro, R., M. C. Rodriguez, A. Seoane and J. M. Garcia Lobo (2003). The aquaporin
gene aqpX of Brucella abortus is induced in hyperosmotic conditions. Microbiology
149(Pt 11): 3185-92.
84

Hill, A., B. Shachar-Hill and Y. Shachar-Hill (2004). What Are Aquaporins For? Journal of
Membrane Biology 197(1): 1-32.
Hinds, B. (2007). Molecular dynamics: A blueprint for a nanoscale pump. Nature
Nanotechnology. 2: 673-674.
Ho, D., B. Chu, H. Lee and C. D. Montemagno (2004). Protein-driven energy transduction
across polymeric biomembranes. Nanotechnology. 15: 1084-1094.
Ho, J. D., R. Yeh, A. Sandstrom, I. Chorny, W. E. C. Harries, R. A. Robbins, L. J. W. Miercke
and R. M. Stroud (2009). Crystal structure of human aquaporin 4 at 1.8 Å and its
mechanism of conductance." Proceedings of the National Academy of Sciences of the
USA 106(18): 7437.
Ho, W. S. W. and K. K. Sirkar (1992). Membrane handbook, Van Nostrand Reinhold New York.
Hoek, V., M. Eric, A. K. Ghosh and J. M. Nygaard (2008). Micro-and nanocomposite support
structures for reverse osmosis thin film membranes, University of California, Los
Angeles.
Holt, J. K., H. G. Park, Y. Wang, M. Stadermann, A. B. Artyukhin, C. P. Grigoropoulos, A. Noy
and O. Bakajin (2006). Fast mass transport through sub-2-nanometer carbon nanotubes.
Science. 312: 1034.
Howorka, S., S. Cheley and H. Bayley (2001). Sequence-specific detection of individual DNA
strands using engineered nanopores. Nature Biotechnology 19(7): 636-639.
Hubert, J., L. Duchesne, C. Delamarche, A. Vaysse, H. Gueuné and C. Raguénès-Nicol (2005).
Pore selectivity analysis of an aquaglyceroporin by stopped-flow spectrophotometry on
bacterial cell suspensions. Biology of the Cell 97: 675-686.
James C-M. Lee, H. B., Bohdana M. Discher, Maureen A. Sheehan, You-Yeon Won, Frank S.
Bates, Dennis E. Discher, (2001). Preparation, stability, and in vitro performance of
vesicles made with diblock copolymers. Biotechnology and Bioengineering 73(2): 135-
145.
Jap, B., M. Zulauf, T. Scheybani, A. Hefti, W. Baumeister, U. Aebi and A. Engel (1992). 2D
crystallization: from art to science. Ultramicroscopy 46(1-4): 45.
Jensen, P. and D. Keller (2006). Membrane for Filtering of Water, WO Patent
WO/2006/122,566.
85

Jensen, P. H. (2007). Biomimetic Water Membrane comprising Aquaporins used in the
production of Salinity Power, WO Patent WO/2007/033,675.
Jeong, B. H., E. Hoek, Y. Yan, A. Subramani, X. Huang, G. Hurwitz, A. K. Ghosh and A. Jawor
(2007). Interfacial polymerization of thin film nanocomposites: A new concept for
reverse osmosis membranes. Journal of Membrane Science. 294: 1-7.
Judd, S. (2006). The MBR Book: Principles and Applications of Membrane Bioreactors for
Water and Wastewater Treatment, Elsevier.
Jung, J. S., G. M. Preston, B. L. Smith, W. B. Guggino and P. Agre (1994). Molecular structure
of the water channel through aquaporin CHIP. The hourglass model. Journal of
Biological Chemistry. 269: 14648.
Kalinichev, A. (2007). Lecture 12, Slide 22, from University of Illinois course GEOL593 MM:
Molecular Modeling of Aqueous Solutions and Substrate-Solution Interfaces, Spring
2007.
Kaufman, Y., A. Berman and V. Freger (2010). Supported Lipid Bilayer Membranes for Water
Purification by Reverse Osmosis. Langmuir.
Kelly, D., D. Dukovski and T. Walz (2008). Monolayer purification: A rapid method for
isolating protein complexes for single-particle electron microscopy. Proceedings of the
National Academy of Sciences of the USA 105(12): 4703.
Kelly, D. F., P. D. Abeyrathne, D. Dukovski and T. Walz (2008a). The Affinity Grid: A Pre-
fabricated EM Grid for Monolayer Purification. Journal of molecular biology. 382: 423-
433.
Kim, S., J. R. Jinschek, H. Chen, D. S. Sholl and E. Marand (2007). Scalable fabrication of
carbon nanotube/polymer nanocomposite membranes for high flux gas transport. Nano
Lett 7(9): 2806-2811.
Kimura, K., G. Amy, J. Drewes, T. Heberer, T. Kim and Y. Watanabe (2003). Rejection of
organic micropollutants (disinfection by-products, endocrine disrupting compounds, and
pharmaceutically active compounds) by NF/RO membranes. Journal of Membrane
Science 227(1-2): 113-121.
Kimura, K., S. Toshima, G. Amy and Y. Watanabe (2004). Rejection of neutral endocrine
disrupting compounds (EDCs) and pharmaceutical active compounds (PhACs) by RO
membranes. Journal of Membrane Science 245(1-2): 71-78.
86

King, L. S., D. Kozono and P. Agre (2004). From structure to disease: the evolving tale of
aquaporin biology. Nature Reviews Molecular Cell Biology. 5: 687-698.
King, L. S., M. Yasui and P. Agre (2000). Aquaporins in health and disease. Molecular Medicine
Today 6(2): 60-65.
Kloda, A., A. Ghazi and B. Martinac (2006). C-Terminal Charged Cluster of MscL, RKKEE,
Functions as a pH Sensor. Biophysical Journal 90(6): 1992-1998.
Knepper, M. A., J. B. Wade, J. Terris, C. A. Ecelbarger, D. Marples, B. Mandon, C. L. Chou, B.
K. Kishore and S. Nielsen (1996). Renal aquaporins. Kidney Int 49(6): 1712-7.
Konradi, R., B. Pidhatika, A. Muhlebach and M. Textor (2008). Poly-2-methyl-2-oxazoline: A
peptide-like polymer for protein-repellent surfaces. Langmuir. 24: 613-616.
Koplin, D., E. Furlong, M. Meyer, E. Thurman, S. Zaugg, L. Barber and H. Buxton (2002).
Pharmaceuticals, Hormones, and Other Organic Wastewater Contaminants in US
Streams: A National Reconnaissance,(1999-2000). Environmental Science and
Technology 36(6).
Kotelyanskii, M. J., N. J. Wagner and M. E. Paulaitis (1998). Atomistic simulation of water and
salt transport in the reverse osmosis membrane FT-30. Journal of Membrane Science.
139: 1-16.
Kotelyanskii, M. J., N. J. Wagner and M. E. Paulaitis (1999). Molecular dynamics simulation
study of the mechanisms of water diffusion in a hydrated, amorphous polyamide.
Computational and Theoretical Polymer Science 9(3-4): 301-306.
Kozono, D., X. Ding, I. Iwasaki, X. Meng, Y. Kamagata, P. Agre and Y. Kitagawa (2003).
Functional expression and characterization of an archaeal aquaporin. Journal of
Biological Chemistry. 278: 10649.
Kozono, D., M. Yasui, L. S. King and P. Agre (2002). Aquaporin water channels: atomic
structure molecular dynamics meet clinical medicine. Journal of Clinical Investigation
109(11): 1395-9.
Kukulski, W., A. D. Schenk, U. Johanson, T. Braun, B. L. De Groot, D. Fotiadis, P. Kjellbom
and A. Engel (2005). The 5 Å structure of heterologously expressed plant aquaporin
SoPIP2; 1. Journal of molecular biology 350(4): 611-616.
Kumar, M., S. S. Adham and W. Pearce (2005). Model Developed for Brackish and Reclaimed
Water. Desalination and Water Reuse 15(3): 38-46.
87

Kumar, M., M. Grzelakowski, J. Zilles, M. Clark and W. Meier (2007). Highly permeable
polymeric membranes based on the incorporation of the functional water channel protein
Aquaporin Z. Proceedings of the National Academy of Sciences of the USA 104(52):
20719.
Kundzewicz, Z. W., L.J. Mata, N.W. Arnell, P. Döll, P. Kabat, B. Jiménez, K.A. Miller, T. Oki,
Z. Sen and I.A. Shiklomanov (2007). Freshwater resources and their management.
Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working
Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate
Change,. O. F. C. M.L. Parry, J.P. Palutikof, P.J. van der Linden and C.E. Hanson, Eds.
Cambridge, UK, 173-210, Cambridge University Press,.
Ladner, D. A. (2009). Effects of Bloom-Forming Algae on Fouling of Integrated Membrane
Systems in Seawater Desalination. Department of Civil and Environmental Engineering.
Urbana, University of Illinois.
Levina, N., S. Tötemeyer, N. Stokes, P. Louis, M. Jones and I. Booth (1999). Protection of
Escherichia coli cells against extreme turgor by activation of MscS and MscL
mechanosensitive channels: identification of genes required for MscS activity. The
EMBO Journal 18: 1730-1737.
Lévy, D., G. Mosser, O. Lambert, G. Moeck, D. Bald and J. Rigaud (1999). Two-dimensional
crystallization on lipid layer: A successful approach for membrane proteins. Journal of
Structural Biology 127(1): 44-52.
Lind, M. L., A. K. Ghosh, A. Jawor, X. Huang, W. Hou, Y. Yang and E. M. Hoek (2009).
Influence of Zeolite Crystal Size on Zeolite-Polyamide Thin Film Nanocomposite
Membranes. Langmuir: the ACS journal of surfaces and colloids.
Liu, D. Z., W. Y. Chen, L. M. Tasi and S. P. Yang (2000). Microcalorimetric and shear studies
on the effects of cholesterol on the physical stability of lipid vesicles. Colloids and
Surfaces A: Physicochemical and Engineering Aspects. 172: 57-67.
Lonsdale, H. K. (1987). The evolution of ultrathin synthetic membranes. Journal of Membrane
Science. 33: 121-136.
Lowry, O. H., N. J. Rosebrough, A. L. Farr and R. J. Randall (1951). Protein measurement with
the Folin phenol reagent. Journal of Biological Chemistry 193(1): 265-75.
88

Majumder, M., N. Chopra, R. Andrews and B. J. Hinds (2005). Nanoscale
hydrodynamicsEnhanced flow in carbon nanotubes. Nature 438(7064): 44.
Mallo, R. and M. Ashby (2006). AqpZ-Mediated Water Permeability in Escherichia coli
Measured by Stopped-Flow Spectroscopy. Journal of Bacteriology 188(2): 820-822.
Martin, D. K. (2007). Nanobiotechnology of biomimetic membranes, Springer Verlag.
Matsura, T. (2001). Progress in membrane science and technology for desalination - a review.
Desalination 134: 47-54.
Maurel, C. (1997). Aquaporins and water permeability of plant membranes. Annual Review of
Plant Biology. 48: 399-429.
McCutcheon, J. and M. Elimelech (2006). Influence of concentrative and dilutive internal
concentration polarization on flux behavior in forward osmosis. Journal of Membrane
Science 284(1-2): 237-247.
Mecke, A., C. Dittrich and W. Meier (2006). Biomimetic membranes designed from amphiphilic
block copolymers. Soft Matter. 2: 751-759.
Mehdizadeh, H., J. M. Dickson and P. K. Eriksson (1989). Temperature Effects on the
Performance of Thin-Film Composite, Aromatic Polyamide Membranes. Industrial &
Engineering Chemistry Research 28(6): 814-824.
Meier, W., C. Nardin and M. Winterhalter (2000). Reconstitution of channel proteins in
(polymerized) ABA triblock copolymer membranes. Angewandte Chemie-International
Edition 39(24): 4599.
Meinild, A. K., D. A. Klaerke and T. Zeuthen (1998). Bidirectional water fluxes and specificity
for small hydrophilic molecules in aquaporins 0-5. Journal of Biological Chemistry
273(49): 32446-32451.
Milon, A., T. Lazrak, A. M. Albrecht, G. Wolff, G. Weill, G. Ourisson and Y. Nakatani (1986).
Osmotic Swelling of Unilamellar Vesicles by the Stopped-Flow Light-Scattering Method
- Influence of Vesicle Size, Solute, Temperature, Cholesterol and 3 Alpha,Omega-
Dihydroxycarotenoids. Biochimica Et Biophysica Acta 859(1): 1-9.
Montemagno, C., J. Schmidt and S. Tozzi (2007). Biomimetic membranes.
Motemagno, C. D. (2006). http://www.cnsi.ucla.edu/arr/paper?paper_id=195905.
89

Murata, K., K. Mitsuoka, T. Hirai, T. Walz, P. Agre, J. B. Heymann, A. Engel and Y. Fujiyoshi
(2000). Structural determinants of water permeation through aquaporin-1. Nature. 407:
599-605.
Napoli, A., Diana Sebök, Alex Senti, Wolfgang Meier, (2008). Block Copolymer Vesicles.
Block Copolymers in Nanoscience. Editors: L. G. Massimo L, Lecommandoux S: 39-71.
Nardin, C., T. Hirt, J. Leukel and W. Meier (2000). Polymerized ABA triblock copolymer
vesicles. Langmuir. 16: 1035-1041.
Nardin, C., S. Thoeni, J. Widmer, M. Winterhalter and W. Meier (2000). "Nanoreactors based on
(polymerized) ABA-triblock copolymer vesicles." Chemical Communications 2000(15):
1433-1434.
Needham, D. and D. Zhelev (1996). The Mechanochemistry of Lipid Vesicles Examined by
Micropipet Manipulation Techniques. Surfactant science series 62: 373-444.
Nehring, R., C. G. Palivan, O. Casse, P. Tanner, J. Tuxen and W. Meier (2009). Amphiphilic
diblock copolymers for molecular recognition: metal-nitrilotriacetic acid functionalized
vesicles. Langmuir: the ACS journal of surfaces and colloids. 25: 1122.
Nemeth-Cahalan, K., K. Kalman and J. Hall (2004). Molecular basis of pH and Ca2+ regulation
of aquaporin water permeability. Journal of General Physiology 123(5): 573.
Nielsen, C. (2009). Biomimetic membranes for sensor and separation applications. Analytical
and Bioanalytical Chemistry 395(3): 697-718.
Nielsen, S., B. L. Smith, E. I. Christensen and P. Agre (1993). Distribution of the aquaporin
CHIP in secretory and resorptive epithelia and capillary endothelia. Proceedings of the
National Academy of Sciences of the USA. 90(15): 7275-9.
Noy, A., H. G. Park, F. Fornasiero, J. K. Holt, C. P. Grigoropoulos and O. Bakajin (2007).
Nanofluidics in carbon nanotubes. Nano Today. 2: 22-29.
Onaca, O., D. W. Hughes, V. Balasubramanian, M. Grzelakowski, W. Meier and C. G. Palivan
(2010). SOD Antioxidant Nanoreactors: Influence of Block Copolymer Composition on
the Nanoreactor Efficiency. Macromolecular Bioscience 10(5): 531-538.
Onaca, O., P. Sarkar, D. Roccatano, T. Friedrich, B. Hauer, M. Grzelakowski, A. Gauven, M.
Fioroni and U. Schwaneberg (2008). Functionalized nanocompartments (Synthosomes)
with a reduction-triggered release system. Angewandte Chemie International Edition
120(37): 7137-7139.
90

Pack, D., G. Chen, K. Maloney, C. Chen and F. Arnold (1997). A metal-chelating lipid for 2D
protein crystallization via coordination of surface histidines. Journal of the American
Chemical Society 119(10): 2479-2487.
Pastan, S. and J. Bailey (1998). Dialysis therapy. N Engl J Med 338(20): 1428-37.
Pata, V. and N. Dan (2003). The effect of chain length on protein solubilization in polymer-
based vesicles (polymersomes). Biophysical journal. 85: 2111-2118.
Patel-Predd, P. (2006). Water desalination takes a step forward. Environmental Science &
Technology 40(11): 3454-3455.
Peer, D., J. M. Karp, S. Hong, O. C. Farokhzad, R. Margalit and R. Langer (2007). Nanocarriers
as an emerging platform for cancer therapy. Nature nanotechnology. 2: 751-760.
Pomes, R. and B. Roux (2002). Molecular mechanism of H+ conduction in the single-file water
chain of the gramicidin channel. Biophysical journal. 82: 2304-2316.
Pontoriero, G., P. Pozzoni, S. Andrulli and F. Locatelli (2003). The quality of dialysis
water.Nephrology Dialysis and Transplantation 18 Suppl 7: vii21-5; discussion vii56.
Preston, G. M., T. P. Carroll, W. B. Guggino and P. Agre (1992). Appearance of Water Channels
in Xenopus Oocytes Expressing Red-Cell Chip28 Protein. Science 256(5055): 385-387.
Rigler, P. and W. Meier (2006). Encapsulation of fluorescent molecules by functionalized
polymeric nanocontainers: investigation by confocal fluorescence imaging and
fluorescence correlation spectroscopy. Journal of the American Chemical Society. 128:
367-373.
Ringler, P., M. Borgnia, H. Stahlberg, P. Maloney, P. Agre and A. Engel (1999). Structure of the
water channel AqpZ from Escherichia coli revealed by electron crystallography. Journal
of molecular biology 291(5): 1181-1190.
Romero, M. F., M. A. Hediger, E. L. Boulpaep and W. F. Boron (1997). Expression cloning and
characterization of a renal electrogenic Na+/HCO3- cotransporter. Nature, 387, 409 - 413
Sambrook, J., E. F. Fritsch and T. Maniatis (1989). Molecular Cloning: A Lanoratory Manual.
N.Y., Cold Spring Harbor Laboratory.
Sauer, M., D. Streich and W. Meier (2001). pH-sensitive nanocontainers. Advanced Materials.
13: 1649.
91

Schenk, A. D., P. J. L. Werten, S. Scheuring, B. L. de Groot, S. A. Muller, H. Stahlberg, A.
Philippsen and A. Engel (2005). The 4.5 Å structure of human AQP2. Journal of
Molecular Biology 350: 278-289.
Scheuring, S., P. Ringler, M. Borgnia, H. Stahlberg, D. J. Muller, P. Agre and A. Engel (1999).
High resolution AFM topographs of the Escherichia coli water channel aquaporin Z.
European Molecular Biology Journal. 18(18): 4981-7.
Service, R. F. (2006). Desalination freshens up. Science 313(5790): 1088-90.
Shapiguzov, A., A. A. Lyukevich, S. I. Allakhverdiev, T. V. Sergeyenko, I. Suzuki, N. Murata
and D. A. Los (2005). Osmotic shrinkage of cells of Synechocystis sp. PCC 6803 by
water efflux via aquaporins regulates osmostress-inducible gene expression.
Microbiology 151(Pt 2): 447-55.
Soupene, E., N. King, H. Lee and S. Kustu (2002). Aquaporin Z of Escherichia coli:
reassessment of its regulation and physiological role. Journal of Bacteriology 184(15):
4304-7.
Stoenescu, R., A. Graff and W. Meier (2004). Asymmetric ABC-triblock copolymer membranes
induce a directed insertion of membrane proteins. Macromolecular bioscience. 4: 930-
935.
Sulkowski, E. (1985). Purification of proteins by IMAC. Trends in Biotechnology 3(1): 1-7.
Tajkhorshid, E., P. Nollert, M. O. Jensen, L. J. W. Miercke, J. O'Connell, R. M. Stroud and K.
Schulten (2002). Control of the selectivity of the aquaporin water channel family by
global orientational tuning. Science. 296: 525.
Tal, A. (2006). Seeking sustainability: Israel's evolving water management strategy. Science
313(5790): 1081-4.
Tanghe, A., P. Van Dijck and J. Thevelein (2006). "Why do microorganisms have aquaporins?"
Trends in Microbiology 14(2): 78-85.
Taubert, A., A. Napoli and W. Meier (2004). Self-assembly of reactive amphiphilic block
copolymers as mimetics for biological membranes. Current opinion in chemical biology
8(6): 598-603.
.
92

Thomas, J. A., A. J. H. McGaughey and O. Kuter-Arnebeck (2009). Pressure-driven water flow
through carbon nanotubes: Insights from molecular dynamics simulation. International
Journal of Thermal Sciences.
Törnroth-Horsefield, S., Y. Wang, K. Hedfalk, U. Johanson, M. Karlsson, E. Tajkhorshid, R.
Neutze and P. Kjellbom (2006). Structural mechanism of plant aquaporin gating. Nature
439: 688-694.
Tournaire-Roux, C., M. Sutka, H. Javot, E. Gout, P. Gerbeau, D. Luu, R. Bligny and C. Maurel
(2003). Cytosolic pH regulates root water transport during anoxic stress through gating of
aquaporins. Nature 425(6956): 393-397.
Uehlein, N., C. Lovisolo, F. Siefritz and R. Kaldenhoff (2003). The tobacco aquaporin NtAQP1
is a membrane CO2 pore with physiological functions. Nature. 425: 734-737.
Uzgiris, E. and R. Kornberg (1983). Two-dimensional crystallization technique for imaging
macromolecules, with application to antigen–antibody–complement complexes. Nature
301, 125 – 129.
Van De Merwe, W., J. Czégé, M. Milham and B. Bronk (2004). Rapid optically based
measurements of diameter and length for spherical or rod-shaped bacteria in vivo.
Applied optics 43(28): 5295-5302.
Verkman, A. S. (2002). Does aquaporin-1 pass gas? An opposing view. The Journal of
Physiology. 542: 31.
Viadiu, H. (2007). Projection map of aquaporin-9 at 7 Angstrom resolution. Journal of molecular
biology 367(1): 80.
Vogel, J., M. Perry, J. S. Hansen, P. Y. Bolinger, C. H. Nielsen and O. Geschke (2009). A
support structure for biomimetic applications. Journal of Micromechanics and
Microengineering 19(2): 25026-25031.
Walz, T., B. L. Smith, M. L. Zeidel, A. Engel and P. Agre (1994). Biologically-Active 2-
Dimensional Crystals of Aquaporin CHIP. Journal of Biological Chemistry 269(3): 1583-
1586.
Wang, Y., J. Cohen, W. Boron, K. Schulten and E. Tajkhorshid (2007). Exploring gas
permeability of cellular membranes and membrane channels with molecular dynamics.
Journal of Structural Biology 157(3): 534-544.
93

Yasui, M., A. Hazama, T.-H. Kwon, S. Nielsen, W. B. Guggino and P. Agre (1999). Rapid
gating and anion permeability of an intracellular aquaporin. Nature 402(6758): 184-187.
Zardoya, R. (2005). Phylogeny and evolution of the major intrinsic protein family. Biology of
the Cell 97: 397-414.
Zelenina, M., A. Bondar, S. Zelenin and A. Aperia (2003). Nickel and Extracellular
Acidification Inhibit the Water Permeability of Human Aquaporin-3 in Lung Epithelial
Cells. Journal of Biological Chemistry 278(32): 30037-30043.
Zhang, L. and A. Eisenberg (1995). Aggregates of Polystyrene-b-poly (acrylic acid) Block
Copolymers. Science. 268: 23.
Zhu, A., P. D. Christofides and Y. Cohen (2009). On RO membrane and energy costs and
associated incentives for future enhancements of membrane permeability. Journal of
Membrane Science. 344: 1-5.
94

APPENDIX A
Calculation of productivity index for BWRO and MF/UF membranes
95

MF/UF Index
The data for this index was obtained from Warren Johnson, US Filter Corporation, and is
shown in Table A.1. The raw data shown in columns 1 and 2 include the year and the cost per
unit production rate of membranes sold (price per kilo liters per day) by US Filter, respectively.
For the year 1999, 2001 and 2002 a similar value was calculated for the contracted value of
membranes at the Carmicheal WaterDistrict, CA
(http://www.carmichaelwd.org/watertreatmentplant.html), Otay Water Treatment Plant (not
built), Chula Vista, CA; and Columbia Heights Filtration Plant, Minnesota, MN, respectively.
The index was obtained by simply indexing the costs to the first year this information was
available (1989).
Table A-1: MF/UF Cost Index Data
Year cost/production rate Time UF/MF Index
$/KL/day years
1989 2000 0 1
1990 1500 1 0.75
1991 900 2 0.45
1992 700 3 0.35
1993 280 4 0.14
1994 270 5 0.135
1995 150 6 0.075
1996 120 7 0.06
1997 80 8 0.04
1998 50 9 0.025
1999 65 10 0.0325
2001 60 12 0.03
2002 65 13 0.0325
BWRO index
The data for this index was obtained from David Furukawa, Separation Consultants Inc, Poway,
CA and is shown in Table A.2. The raw data shown in columns 1, 2 and 4 include the year, the
cost per eight inch brackish water RO (BWRO) module, and the productivity (flux per unit
applied pressure), respectively. This module cost is indexed to the year 1980 (column 3). Using
96

this indexed membrane module cost and diving it by productivity of the module provides a
cost/productivity number. This number is then indexed again to 1980 to obtain the BWRO
Improvement index shown in Figure 1.2 (b).
Table A-2: Brackish Water RO Cost Index Data
Year OEM cost
Indexed
membrane
Cost
Membrane
Productivity Years Cost/Productivity
Improvement
index
$/module indexed cost gfd/psi Indexed cost/productivity
1980 800 1.0 0.06 0 16.07 1
1985 650 0.8 0.07 5 11.42 0.711
1990 550 0.7 0.08 10 8.59 0.535
1995 450 0.6 0.10 15 5.75 0.358
2000 350 0.4 0.13 20 3.40 0.212
2010 300 0.375 0.17 30 2.14 0.133
97

APPENDIX B
Calculations for addition of NTA-polymer to 4 mm wells to obtain 100 Å2/molecule
98

Surface area for spreading of the NTA polymer in 4 mm diameter wells
= = 1.257 x10-5 m2
(4 / 4 3 210 )
The calculation of excess amount of polymer added is based on a lipid based Ni chelating
layer from Levy et al ( Journal of Structural Biology, 127, 44-52 (1999).
They target a surface area per molecule of 60 Angstrom2.
Calculating the number of molecules of Ni2+ chelating lipids (Ni2+-NTA-DOGS) needed
=1.257x10-5 m2/ (60x10-20) m2 = 2.094 x 1013 molecules
Or, 3.477 x 10-11 moles of chelating lipids needed to form one layer.
Now they suggest adding more than necessary for one layer but less than two layers to
make up for loss to solution due to presence of detergent in subphase
They actually add 0.5 microL of 0.1 mg/ml lipids (MW 1057.02) which in terms of moles
is
3110.1 / 0.5 10
4.73 101000 / 1057.02 /
mg ml mlmoles
mg g g mol
Thus the excess amount they added was = 4.73/3.48 = approx 1.4 times the theoretical
amount
For our case, The NTA polymer has a MW of 4754 g/mol and the target Mean Molecular
Area from Rainer’s data seems to be 100 Angstrom2.
Number of molecules of Ni2+ chelating polymer needed for one layer:
99

=1.257x10-5 m2/ (100x10-20) m2 = 1.257 x 1013 molecules
Or, 2.086 x 10-11 moles of chelating lipids needed to form one layer.
x 1.4 excess = 2.92 x10-11 moles
To add this much polymer on the surface in a volume of 0.5 microL,
we will need 0.5 microL of x mg/ml NTA polymer
3110.5 10
2.92 104754000 /
0.28
mg mlx moles
ml mg mol
mgx
ml
100

APPENDIX C
Crosslinking of methacrylated PMOXA PDMS triblock co-polymers using FTIR and the
effect of crosslinking on chemical and mechanical stability
101

Crosslinking of methacrylated PMOXA PDMS triblock co-polymers using FTIR and the
effect of crosslinking on chemical and mechanical stability
Summary: A series of experiments were conducted to systematically investigate UV-induced
crosslinking behavior of ABA type methacrylated PMOXA-PDMS triblock copolymer. A
knowledge of the kinetics and dose requirements of this process is important for application of
polymerized triblock copolymer to situations where the mechanical and chemical stability of the
polymer resulting from crosslinking would be important. Two polymers were used,
methacrylated ABA3 (PMOXA15-PDMS110-PMOXA15) and methacrylated ABA 4 (PMOXA12-
PDMS55-PMOXA12). These polymers were dissolved in chloroform and coated onto a glass
slide to make a thick film, from which chloroform was either evaporated at room temperature
(ABA3) or by using a refrigerated rotary vacuum evaporator (ABA4). Attenuated Total
Reflection – Fourier Transform Infra Red (ATR-FTIR) spectroscopy was performed on this
polymer to obtain a spectrum for the pure polymer. The polymer was then subjected to UV
irradiation from a solar simulator with UV intensity of 17mW/cm2 in the 340 - 390 nm range and
an overall intensity of 407 W/cm2 over different lengths of time. The polymer sample was
analyzed after each exposure with FTIR to determine the kinetics of crosslinking between the
methacrylated groups. Chemical stability of the crosslinked polymer was evaluated by
conducting dissolution tests in 100% ethanol. Mechanical stability tests have not been
performed, but could be done by subjecting vesicles to controlled shear conditions to determine
the maximum shear that such vesicles can withstand. It is known that lipid vesicles can
withstand a shear rate of only 500 s-1 in water, equivalent to an average shear stress of 5.01 x10-1
N/m2. It is anticipated that the even the unpolymerized vesicles will be able to withstand much
larger shear stresses than lipid vesicles and this will be further enhanced (maybe by a large
amount) by crosslinking of these vesicles by UV irradiation.
For both the polymers the FTIR spectrum shows distinct peaks for the PDMS and the PMOXA
parts of the polymer, indicating that this method is an excellent method to characterize this class
of polymers. The methacrylated ABA3 polymer seems to show a strong FTIR signal, and the
amount of crosslinking could be followed using a distinct peak. Also, after crosslinking the
polymer was extremely insoluble in 100% ethanol. Exposure to ambient light at room
102

temperature also caused crosslinking and led to insolubililty of the polymer in ethanol. The
crosslinking of ABA4 polymer, on the other hand, is difficult to analyze using FTIR. There are
no distinct peaks that evolve as a result of UV curing. This polymer also had no resistance to
100% ethanol. The level of methacrylation for this polymer seems to be low; increasing
methacrylation might lead to better UV curing and solvent resistance properties.
Materials and Methods
Sample prep
A 30-50 mg sample of polymer was dissolved in 5 ml CHCl3 and coated on a glass slide. In the
case of ABA3 it was left overnight to dry and form a thick film. For ABA 4 the glass slide was
put in a vacuum evaporator to dry off the CHCl3.
Fourier Transfer Infrared (FTIR) spectroscopy.
Attenuated total reflectance Fourier transform infrared (ATR-FTIR) spectrometry was used to
assess changes in characteristic functional groups of the methacrylated triblock co-polymer
during UV crosslinking. Analyses were performed with a NEXUS 670 FTIR spectrometer
(Thermo Nicolet Corporation, Madison, WI) equipped with a smart golden gate accessory,
DTGS-KBr detector, KBr beam-splitter and diamond crystal. An IR source at 45◦ incident angle
was employed. Samples of polymer were put in direct contact with the diamond crystal and 32-
64 scans were conducted in the range of 600 to 4000 cm-1 for each sample. The information
from these scans was averaged by the instrument to provide IR spectra which were then
analyzed.
UV exposure
The samples were exposed to UV light in a collimated beam apparatus to induce crosslinking of
the methacrylated polymers. The samples were exposed for different lengths of time (0.5 min to
3.5 hours) to provide different doses of UV irradiation for crosslinking. After each exposure the
samples were analyzed using FTIR to monitor changes in functional groups. The UV lamp was
a solar simulator with an overall intensity of 407 mW/cm2 and an intensity of 17 mW/cm2 in the
324 - 400 nm wavelength range expected to cause curing of polymers.
Data Analysis Methodology (Selection of appropriate peaks)
103

To investigate polymer crosslinking kinetics, FTIR spectra were examined to determine if
crosslinking of methacrylate groups causes an appreciable change in any part of the spectrum.
For the ABA3 polymer, UV exposure causes a large change in the intensity of the peak at 750
cm-1, so this was used to determine the kinetics of crosslinking. Usually ~810 cm-1 is used to
monitor crosslinking in acrylates (-CH our of plane bending) but this was shifted to 750 cm-1 in
our case. This is likely to be due to the large PDMA, PMOXA groups that the acrylates are
attached to. The change in area under the 750 cm-1 peak provided values for the change in
crosslinking with time.
For the ABA 4 polymer it was much more difficult to find an independent peak that would
confirm methacrylate crosslinking. No peak was seen at 750 cm-1 for this polymer. However,
the ~790 cm-1 peak increases as a result of UV exposure. We propose using this peak for
monitoring increase in crosslinking. It represents the formation of methylene bonds (-CH2)
bonds, which is usually observed at 814 cm-1. The use of this peak for the ABA polymer,
however, is not ideal for monitoring crosslinking as we do not have a way of determining
independent increase in crosslinking; but in the absence of independent peaks, it is a good
choice??.
Results and Discussion
The following presents a detailed description of the FTIR spectra of the two methacrylated
PMOXA-PDMS ABA polymers. It also describes the UV crosslinking of these two polymers by
monitoring change in the FTIR spectrum at different UV exposure time. Additionally the
chemical stability tests conducted with 100% ethanol are described, and a set of experiments are
proposed to characterize the mechanical (shear) stability of these polymers.
FTIR Spectrum of uncrosslinked polymers.
Figure C-1 presents the FTIR spectrum of the two methcrylated PDMS-PMOXA polymers. The
peaks corresponding to pure PDMS are evident in the spectrum. These peaks include the 798
cm-1 representing the Si-C stretching and CH3 rocking vibrations. The twin peaks at 1018 and
1092 cm-1 represent the Si-O-Si stretching vibrations. The peak at 1260 cm-1 represents the CH3
symmetric deformation of Si-CH3 bonds. The peak at 1632 cm-1 may represent the amide C=O
104

bond from the PMOXA. These peaks are common to ABA 3 and ABA 4 polymers. Several
differences are also seen between the two polymers. In particular the ABA3 polymer shows
several extra peaks including those at 696 cm-1, 756 cm-1, 1453 cm-1, 1493 cm-1 as well as at
1608 cm-1.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
600 800 1000 1200 1400 1600 1800 2000
wavelength (cm-1)
Ab
sorb
ance
(ar
b u
nit
s)
ABA3
ABA41
2
3 4
5
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
600 800 1000 1200 1400 1600 1800 2000
wavelength (cm-1)
Ab
sorb
ance
(ar
b u
nit
s)
ABA3
ABA41
2
3 4
5
Figure C-1: FTIR spectrum of ABA 3 and ABA 4 polymers. Peaks represent the PDMS and
PMOXA components of the block copolymer. These are identified as 1. (~798 cm-1) Si-C
stretching and CH3 rocking (from PDMS). 2. & 3. (~1018 and 1092 cm-1) Si-O-Si stretching
vibrations (from PDMS). 4. (~1260cm-1) CH3 symmetric deformation of Si-CH3 (from PDMS).
5. (~1632 cm-1) amide C=O bond (from PMOXA)
Crosslinking of ABA3
The exposure of the ABA 3 polymer to UV radiation caused a large decrease in the peak around
750 cm-1. This may represent the crosslinking of the methacrylate groups by representing a
decrease in the C-H bond out of plane bending. This peak is usually seen at ~814 cm-1 but the
presence of the large PDMS group attached to the methcrylate group may have resulted in a
shifting of this peak. The spectra of ABA3 polymer after exposure to different UV doses is
shown in Figure C-2(a).
105

The degree of crosslinking was calculated by determining the area under this peak, while its
evolution over time represents the kinetic of crosslinking for this polymer. This is shown in
Figure C-2(b).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
600 800 1000 1200 1400 1600 1800 2000
Wavelength (cm-1
)
No
rmal
ized
Ab
sorb
ance
0 min
12.5 min
72.5 min
192.5 min
C-H out of plane bond (shifted)
0%
5%
10%
15%
20%
25%
30%
35%
0 50 100 150 200 250
UV Dose 324 -400 nm (J/cm 2)
cro
sslin
kin
g (
%)
(b)
(a)
Figure C-2: (a) FTIR Spectra of ABA 3 polymer exposed to increasing UV dose (time) (b)
Kinetics of crosslinking of ABA 3 polymer
Crosslinking of ABA 4
As seen in Figure C-3, it was much more difficult to find an independent peak that would
confirm the assumption of methacrylate crosslinking in the ABA 4 polymer. No peak was seen
106

at 750 cm-1 for this polymer. However, the ~790 cm-1 peak increased as a result of UV exposure.
We propose using this peak for monitoring increase in crosslinking. It represents the formation
of methylene bonds (-CH2-) bonds, which is usually observed at 814 cm-1.
The kinetics of crosslinking seem to show a lower level of crosslinking for this polymer as seen
from Figure C-3(b).
Figure C-3: (a) FTIR Spectra of ABA 4 polymer exposed to increasing UV dose (time) (b)
Kinetics of crosslinking of ABA 4 polymer
Chemical Stability
(b)
0%
1%
2%
3%
4%
5%
6%
7%
8%
9%
10%
0 50 100 150 200 250 300UV Dose324-400 nm (J/cm2)
cro
ss
link
ing
(%
)
0
0.1
0.2
0.3
0.4
0.5
0.6
600 700 800 900 1000 1100 1200
Wavelength (1/cm)
No
rma
lize
d A
bs
orb
anc
e
0 min
2.5 min
32.5 min
122.5 min
302.5 min
(a)
(b)
107

The ABA3 polymer completely dissolved in 100% ethanol without crosslinking. However, after
192.5 minutes of exposure to UV, less than 10% of mass was lost from 9 mg of the polymer after
two days of exposure to 100 % ethanol. Both the untreated and treated ABA 4 polymer dissolved
easily in 100% ethanol. This is consistent with the low level of crosslinking observed in ABA 4
polymer. The level of methacrylation for this polymer may have been quite low, resulting in
lower possibility for crosslinking.
Mechanical Stability: Proposed experiments.
We will subject both crosslinked and uncrosslinked polymer vesicles to increasing shear stresses
in a cone on plate viscometer and determine the highest shear rate that these vesicles can
withstand before rupture. A light scattering approach will be used to determine the point where a
large amount of the vesicles are ruptured. Observed rupture stresses can then be compared to
those reported in literature for Phosphatidyl choline (PC) lipid vesicles (~ 500 – 1000 mPa) (Liu
et al. 2000).
References
Liu, D. Z., W. Y. Chen, et al. (2000). Microcalorimetric and shear studies on the effects of
cholesterol on the physical stability of lipid vesicles. Colloids and Surfaces A: Physicochemical
and Engineering Aspects. 172: 57-67.
108

AUTHOR’S BIOGRAPHY
See attached Cirriculum Vitae
109

MANISH KUMAR 205 N. Mathews Ave, 4125
Urbana, IL 61801 (619) 917‐7392
EDUCATIONAL HISTORY Doctor of Philosophy in Environmental Engineering, Department of Civil and Environmental Engineering, University of Illinois, Urbana, IL, 08/2006 – 05/2010 (expected). Advisors: Drs. Mark M. Clark and Julie Zilles; Project: Biomimetic High Efficiency Desalination Membranes Master of Science in Environmental Engineering, Department of Civil and Environmental Engineering University of Illinois, Urbana, IL, 08/1998 – 01/2000. Advisor: Dr. Mark M. Clark; Project: Fouling of Ultrafiltration Membranes by Natural Organic Matter (NOM) Bachelor of Technology in Chemical Engineering, Department of Chemical Engineering National Institute of Technology, Trichy, India, 08/1994 – 05/ 1998 EXPERIENCE
EPA Science to Achieve Results (STAR) Fellow, University of Illinois, Urbana, IL, 2008 – current, Project: Development of highly‐efficient Aquaporin‐based water treatment membranes for desalination and contaminant removal Visiting Researcher, Wolfgang Meier Group and Andreas Engel Group. Department of Chemistry, University of Basel, Switzerland, Summer 2007 and Fall 2008
University of Illinois Fellow and Research Assistant, University of Illinois, Urbana, IL, 2007 ‐ 2008,
Project: Characterization of US Seawaters and Development of Standardized Protocols for Evaluation of Foulants in Seawater Reverse Osmosis Desalination (project in addition to dissertation research).
Senior Engineer, Applied Research Department, MWH, Pasadena, CA, 2001 – 2006, Expertise and Focus: Membrane water and wastewater treatment
Staff Engineer, NCS, Phoenix, AZ, 2000 – 2001, Expertise and Focus: Arsenic treatment, Membrane water and wastewater treatment
Research Assistant, University of Illinois, Urbana, IL, 1998 – 2000 Research Intern, Bhabha Atomic Research Center, Mumbai, India, Summer 1997
Research Intern, Central Electrochemical Research Institute, Karaikudi, India. 1996 – 1997
LICENSES Professional Engineer – Civil, California, P.E. License #69423, expiration July 2010
110

AWARDS
Richard and Mary Engelbrecht Fellowship, Department of Civil and Environmental Engineering, UIUC, 2009
Mavis Memorial Fund Scholarship, College of Engineering, UIUC, 2009
European Union Marie Curie Training Course Fellowship to attend 7th NCCR Practical Course on 2D Membrane Protein Crystallization and Observation , October 2008, Switzerland
US Environmental Protection Agency STAR Fellowship, 2008 – 2011, USEPA
Abel Wolman Fellowship, 2008‐2009, American Water Works Association
Henry Ford II Scholar, 2008, College of Engineering, UIUC
National Water Research Institute Fellowship for Advanced Water and Wastewater Treatment Technologies, 2007 – 2008, NWRI
Included in the “Incomplete list of Instructors ranked as excellent by their students, Fall 2006, UIUC” Courses CEE 347 and CEE 442 and in the "List of Instructors ranked as excellent by their students, Spring 2008, UIUC" CEE 437
Dissertation Travel Grant, Summer 2007, UIUC
University of Illinois Fellowship, 2006 – 2007, UIUC
MWH President’s Award for most Innovative Project, 2006
Oil and Natural Gas Corporation (Government of India) Scholarship, 1994 – 1998 INVENTION DISCLOSURES AND PATENT APPLICATIONS
1. Kumar, M; Meier, W.; Zilles, J.; Clark, M.; Grzelakowski, M.; Nehring, R. Highly permeable polymer
membranes, PCT patent application, filed Dec 4th, 2008, Assignee: University of Illinois, Corresponding Inventor: Manish Kumar
2. Kumar, M; Poust, S; Meier, W; Clark, M; Zilles, J; Cropek, D; McAllister, I. Environmental remediation with functionalized vesicles, disclosed to the University of Illinois, Dec 14th, 2009, Corresponding Inventor: Manish Kumar
PUBLICATIONS
1. Kumar, M; Marincel, M; Poust, S; Zilles, J. 2010. Aquaporin rich polymer membranes. In Biomimetic
membranes, Ed. Claus H. Nielsen, Springer (Invited Contribution, in preparation) 2. Kumar, M; Grzelakowski, M; Clark, M; Meier, W; Zilles, J. 2010. Biological role for bacterial
aquaporin gating discovered in block copolymer membranes (submitted) 3. Ladner, DA; Subramani, A; Kumar, M; Adham, S; and Clark, MM. 2009. Bench‐scale evaluation of
seawater desalination by reverse osmosis (in press) Desalination. 4. Grzelakowski, M., Onaca, O., Rigler, P., Kumar, M., & Meier, W. 2009 Immobilized Protein‐Polymer
Nanoreactors. Small, published online August 13, 2009. 5. Badruzzman M; Oppenheimer, J; Adham, S; and Kumar, M. 2009. Innovative Beneficial Reuses of
Reverse Osmosis Concentrate Using Bipolar Membrane Electrodialysis and Electrochlorination Processes, Journal of Membrane Science. 326 (2009) 392 – 399
6. Kumar, M; Grzelakowski, M; Zilles, J; Clark, M; Meier, W. 2007. Highly permeable polymeric membranes based on the incorporation of the functional water channel protein Aquaporin Z, Proceedings of the National Academy of Sciences. 2007;104(52):20719 – 20724. (featured on cover and in a commentary article in PNAS issue, also on Sciencedaily.com and several other sites)
111

7. Kumar, M; Adham, S; Pearce, W. 2007. Comparative seawater reverse osmosis pretreatment evaluation using bench and pilot scale testing, Journal of the American Water Works Association. 99:9: 168 – 178
8. Kumar, M; Adham, S; and DeCarolis, J. 2007. Reverse Osmosis Integrity Monitoring, Desalination, 214: 138 – 149
9. Kumar, M; Badruzzaman, M; Adham, S; Oppenheimer, J. 2007. Beneficial phosphate recovery from reverse osmosis (RO) concentrate of an integrated membrane system using polymeric ligand exchanger (PLE). Water Research 41, (10), 2211 – 2219
10. Kumar, M; Adham, S; & Pearce, W. 2007. An Integrated Approach to Pretreatment Evaluation for Seawater Reverse Osmosis using Bench and Pilot Scale Testing, in Membrane Treatment for Drinking Water and reuse Applications: A Compendium of Peer‐Reviewed Papers, Ed. Kerry Howe, AWWA, Denver Co.
11. Kumar, M; Adham, S, and Pearce, W. 2006. Development of a protocol for evaluation of new generation RO membranes, Journal of the American Water Works Association, 98, 4, 22 – 132
12. Kumar, M; Adham, S, and Pearce, W. 2006. Investigation of Seawater Reverse Osmosis Fouling And Its Relationship To Pretreatment Type, Environmental Science and Technology, 40, 2037 – 2044. (featured in a news story in ES&T)
13. Kumar, M; Adham, S, and Pearce, W. 2006. Evaluation of New Generation RO membranes for Brackish Groundwater Desalination, In Desalination of Seawater and Brackish Water, William Lauer (ed), Edition: 2006, Hardback, ISBN 1583213287
14. DeCarolis, J, Kumar, M, Adham, S, Pearce, W, Wasserman, L. 2005 Double Filtered for Freshness: An Integrated membrane System could expand water reuse options without excessive maintenance, Water Environment and Technology, March 2005, pg.41 – 47
15. Adham S; Kumar, M; and Pearce, W. 2005. Model Developed for Brackish and Reclaimed Water, Desalination and Water Reuse, 15(3)38 – 46
CO–AUTHORED & FUNDED RESEARCH PROJECTS*
1. Development of highly efficient aquaporin‐based membranes for aqueous separations. Mark Clark (PI) and Julie Zilles (Co‐PI). Funded by the National Science Foundation, Chemical, Bioengineering, Environmental, and Transport Systems, Chemical and Biological Separations, $400,000. Project # CBET 08‐28512, 2008 – 2011
2. Synthetic cells imbued with functional biomolecular machinery. Irene McAllister (PI), Mark Clark (Co‐PI) and Julie Zilles (Co‐PI). Funded by the U.S. Army Engineer Research and Development Center, 6.1 Basic Research , $750,000, 2008 – 2012
3. Development of highly‐efficient Aquaporin‐based water treatment membranes for desalination and contaminant removal, Manish Kumar (PI). Funded by the EPA Science to Achieve Results (STAR) Program (Fellowship), $111,000, 2008 – 2011
4. Aquaporin based membranes for water treatment, Mark Clark (PI) and Manish Kumar (Co‐PI). Funded by National Water Research Institute (Fellowship), $20,000, 2007 – 2009
5. Participated in 7 other winning proposals to Water Research Foundation (WRF). Water Reuse Foundation, and National Water Research Institute during industrial experience
* The University of Illinois does not allow graduate students to be listed as PI’s, however, I contributed substantially to idea and
collaboration development and writing the proposals listed.
112

TEACHING AND MENTORSHIP ACTIVITIES
Volunteered to teach a short block of one or more Environmental Engineering courses in every semester of PhD study. Complete responsibility for the course materials and lectures (3 – 4 lecture sessions), creating, and grading homework assignment and exam question(s). Fall 2006, CEE 398 SEU: Sustainable Urban Engineering Spring 2007, Spring 2008 and Spring 2009, CEE 437: Water Quality Engineering Fall 2007 and Fall 2008, CEE 442: Physical Principles of Environmental Engineering Spring 2009, CEE 498WTS: Water Treatment Case Studies
Teaching Assistant for 3 courses in the department. I was responsible for all assignments, exams and grading as well as teaching several lectures. Fall 2006, CEE 442: Physical Principles of Environmental Engineering Fall 2006 and Fall 2007, CEE 437: Water Quality Engineering
Invited lecture in an advanced Physics course (with a Biophysics focus). Spring 2008, PHYS 498 PNM: Physics of Nanomachines
Committed and active member in recruiting and job mentoring activities for Illinois EES program
Recruited, mentored and hosted a Masters student from University of Basel, Department of Chemistry who conducted his Masters project research at Illinois
PROFESSIONAL MEMBERSHIPS, SERVICE AND ACTIVITIES
Board Member, The Spout Institute – A non profit focused on water quality education and for development of sustainable water treatment technologies for developing countries, 2009 – present
Co‐Chair, Gordon Research Conference, Graduate Research Symposium, Membranes‐Materials and Processes, 2010
Member, Association of Environmental Engineering and Science Professors, since 2008
Member, American Society for Microbiology, 2008 – present
Member, American Water Works Association, 1998 – present
Reviewer: Journal of Membrane Science, Environmental Science and Technology, Water Research, Journal AWWA, Desalination, ASCE ‐ Journal of Environmental Engineering, AQUA, Water Science and Technology
Member, Young Professionals Group, MWH, Pasadena, CA 2005 – 2006
Founding Board Member, Network of Indian Professionals, San Diego, 2001 – 2003
Vice Chair, Chemical Engineering Association, Trichy, India 1997 – 1998 INVITED PRESENTATIONS
1. “Biomimetic Membranes for Desalination Water Treatment”, invited presentation at the Swisstech
Conference, Nov 2008, Basel, Switzerland 2. “Testing of RO Concentrate Minimization and Beneficial Reuse Alternatives”, Southern California
Salinity Coalition Symposium organized by the US Bureau of Reclamation, May 2, 2006, Metropolitan Water District of Southern California, Los Angeles, CA
3. “Current Status of Worldwide MF/UF Full‐scale Plants”, MF IV symposium, National Water Research Institute, Mar 23, 2006, Orange, CA
4. “Current State of Desalination in California”, Feb 7, 2006, Southern Water, Brighton, UK 5. “Pretreatment Strategies for Seawater Desalination”, Orange County Water District, Sep 2005,
Fountain Valley, CA
113

6. “Pretreatment Strategies for Seawater Reverse Osmosis”, Metropolitan Water District of Southern California, Sep 2005 Los Angeles, CA
CONFERENCE PRESENTATIONS
1. Kumar, M., Grzelakowski, M., Meier, W., Clark ,M., and Zilles, J. Bioinspired Membranes for
Environmental Applications, poster presented at Association for Environmental Engineering and Science Professors meeting, Iowa City, July 2009
2. Kumar, M., Grzelakowski, M., Clark ,M., Meier, W. and Zilles, J. pH Gating of the Escherichia coli Water Channel Protein, Aquaporin Z, poster presented at the American Society for Microbiology meeting, Philadephia, May 2009
3. Kumar, M., Clark M and Zilles, J., Biomimetic desalination membranes, poster presented at the Gordon Research Conference (Graduate Research Symposium) – Membranes and Materials, New London, New Hampshire, August 2008
4. Ladner, D. A.; Vardon, D. R.; Kumar, M., Clark, M. M. Fouling by algogenic organic matter in seawater reverse osmosis desalination. AWWA Annual Conference and Exibition, Atlanta, GA. June 2008
5. Kumar, M., Grzelakowski, M., Meier, W. Hochselektive Wasserfilter auf der Basis von Blockcopolymer Membranen, Macromolecular Colloquium, University of Freiburg, Institute for Macromolecular Chemistry, Freiburg, Germany, 2008 (presented by Dr. Meier)
6. Adham, S. , Oppenheimer, J., Kumar, M., Badruzzaman, M., Liu, L. 2007. Innovative Approaches to RO Concentrate Management : Beneficial Reuse and Concentrate Minimization, Proceedings of the International Desalting Association Conference, Gran Canaria, 2007
7. Kumar, M., Oppenheimer, J., Adham, S., and Kottenstette, R. Innovative Technologies for Beneficial Reuse of RO Concentrate, presented at the AWWA Annual Conference and Exhibition, 2006, San Antonio, Texas, 2006
8. Kumar, M.; Adham, S., Decarolis, J., Burbano, A. and Pearce, W. Pretreatment Evaluation For Seawater Reverse Osmosis Using Bench And Pilot Scale Testing, Presented at the AWWA Annual Conference, San Francisco, CA, 2005
9. Oppenheimer, J.; Chiu, K; DeCarolis, J.; Kumar, M.; Adham, S.; Snyder, S.; and Pearce, W.; Evaluating the Effect of UV Peroxide for Control of NDMA on Endocrine Disrupters, Pharmaceuticals, and Personal Care Products, presented at the AWWA Annual Conference 2006, San Antonio, Texas, 2006
10. Oppenheimer, J.; Kumar, M.; Badruzzaman, M.; Webb, L.; Kottenstette, R.; Adham, S. Using RO Concentrate as a Resource by Application of Innovative Technologies, presented at the AWWA Water Quality Technology Conference November, Denver, Colorado 2006
11. Adham, S.; Oppenheimer, J.; Kumar, M.; Webb, L.; Kottenstette, R. Innovative Approaches to RO Concentrate Management: Beneficial Reuse and Concentrate Minimization, presented at WEFTEC, Dallas, Texas, 2006
12. Kumar, M., Oppenheimer, J., Adham, S., and Kottenstette, R. Innovative Technologies for Beneficial Reuse of RO Concentrate, presented at the AWWA Annual Conference and Exhibition, 2006, San Antonio, Texas, 2006
13. Ladner, D.; Clark, M., M; Kumar, M.; and Adham, S. Bench‐Scale Studies to Characterize Organic Foulants in Seawater Reverse Osmosis Desalination, presented at the North American Membrane Society meeting, Chicago, Illinois May 12 – 17, 2006
14. Kumar, M.; Adham, S.; & Pearce, W. An Integrated Approach to Pretreatment Evaluation for Seawater Reverse Osmosis using Bench and Pilot Scale Testing, Presented at the AWWA Membrane Technology Conference, Phoenix, AZ, 2005
114

15. Kumar, M.; Adham, S., Decarolis, J., Burbano, A. and Pearce, W. Pretreatment Evaluation For Seawater Reverse Osmosis Using Bench And Pilot Scale Testing, Presented at the AWWA Annual Conference, San Francisco, CA, 2005
16. Kumar, M.; Decarolis, J.; Adham, S.; Wasserman, L. Desalting of MBR treated wastewater: Integrated Membrane Systems for wastewater Reclamation, Proceedings of the California Nevada Section of AWWA, Sacramento, CA, October 2004
17. Kumar, M.; Adham, S.; & Pearce, W. Development of a Cost Model for Brackish and Reclaimed Water Desalination, Paper presented at the AWWA Annual Conference, Orlando, FL, 2004
18. Kumar, M.; DeCarolis, J.; Adham, S.; Pearce, W; Wasserman, L. Integrated Membrane Systems for Wastewater Reuse, Paper presented at the Water Environment Federation Technology (WEFTEC) conference, New Orleans, LA, 2004
19. Pearce, W.; Kumar, M. Evaluation of New Generation Low Pressure Brackish Water Reverse Osmosis Membranes, Proceedings of the California Nevada AWWA 2004 Spring Conference, April 13, 2004, Las Vegas, NV, 2004
20. Kumar, M.; Adham, S.; & Pearce, W., Development of a Spreadsheet Tool for Brackish and Reclaimed Water Membrane Desalination Costs, Paper presented at the AWWA Annual Conference, Anaheim, CA, June 2003
21. Pearce, W.; Kumar, M.; & Adham, S., Evaluation of New Generation Low Pressure Brackish Water Reverse Osmosis Membranes, Paper presented at the AWWA Annual Conference, Anaheim, CA, June 2003
22. Kumar, M.; Adham, S.; & Pearce, W., Spreadsheet Tool for Brackish and Reclaimed Water Membrane Desalination Costs, Paper presented at the AWWA Membrane Conference, Atlanta, GA, March 2003
23. Oppenheimer, J., DeCarolis, J., Kumar, M., and Gagliardo, P., Adham, S., Environmental Technology Verification of Low‐ and Medium Pressure Ultraviolet Systems, Presented at the American Water Works Association Water Quality Technology Conference. Seattle, WA, November 2002
24. Kumar, M.; Hsu, Y.; Devkota, L.; Welch, M.; and Smith, B. , Optimizing Arsenic Removal in Phoenix Area Water Treatment Plants, Paper presented at the Arizona Water Pollution Control Association (AWPCA) Annual Conference, Chandler, AZ, May 2001
25. Wang, Y.; Clark, M.M.; & Kumar, M., Characterization and fouling of Sulfonated Polyether Sulfone/Polyethersulfone – Polysulfone (SPEES/PES‐PS) UF membranes, Paper presented at the North American Membrane Society (NAMS) Conference, Denver, CO, May 2000
115
Copyright © 2022 FDOKUMEN




















