
The effects of dietary phytoestrogens on aromatase activity in human endometrial stromal cells

© The Author 2013. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved. For permissions, please email: [email protected].
Coexposure to Phytoestrogens and Bisphenol A Mimics Estrogenic Effects in an Additive Manner
Anne Katchy,*,1 Caroline Pinto,*,1 Philip Jonsson,* Trang Nguyen-Vu,* Marchela Pandelova,† Anne Riu,*
Karl-Werner Schramm,†,‡ Daniel Samarov,§ Jan-Åke Gustafsson,*,¶ Maria Bondesson,* and Cecilia Williams*,2
*Center for Nuclear Receptors and Cell Signaling, Department of Biology and Biochemistry, University of Houston, Houston, Texas 77204-5056; †Helmholtz Zentrum München (GmbH), Molecular EXposomics (MEX), Ingolstädter Landstr.1, Neuherberg D-85764, Germany; ‡TUM, Wissenschaftszentrum
Weihenstephan für Ernährung und Landnutzung, Department für Biowissenschaften, Weihenstephaner Steig 23, Freising 85350, Germany; §Statistical Engineering Division, Information Technology Laboratory, National Institute of Standards and Technology, Gaithersburg, Maryland 20899; and ¶Department of
Biosciences and Nutrition, Novum, Karolinska Institutet, Stockholm 141 83, Sweden
1These authors contributed equally to this study.2To whom correspondence should be addressed at Center for Nuclear Receptors and Cell Signaling, Department of Biology and Biochemistry, University of
Houston, Houston, TX 77204-5056. Fax: (713) 743-0634. E-mail: [email protected].
Received May 23, 2013; accepted November 6, 2013
Endocrine-disrupting chemicals (EDC) are abundant in our environment. A number of EDCs, including bisphenol A (BPA) can bind to the estrogen receptors (ER), ERα and ERβ, and may contribute to estrogen-linked diseases such as breast cancer. Early exposure is of particular concern; many EDCs cross the placenta and infants have measurable levels of, eg, BPA. In addition, infants are frequently fed soy-based formula (SF) that contains phytoes-trogens. Effects of combined exposure to xeno- and phytoestro-gens are poorly studied. Here, we extensively compared to what extent BPA, genistein, and an extract of infant SF mimic estrogen-induced gene transcription and cell proliferation. We investigated ligand-specific effects on ER activation in HeLa-ERα and ERβ reporter cells; on proliferation, genome-wide gene regulation and non-ER–mediated effects in MCF7 breast cancer cells; and how coexposure influenced these effects. The biological relevance was explored using enrichment analyses of differentially regulated genes and clustering with clinical breast cancer profiles. We dem-onstrate that coexposure to BPA and genistein, or SF, results in increased functional and transcriptional estrogenic effects. Using statistical modeling, we determine that BPA and phytoestrogens act in an additive manner. The proliferative and transcriptional effects of the tested compounds mimic those of 17β-estradiol, and are abolished by cotreatment with an ER antagonist. Gene expres-sion profiles induced by each compound clustered with poor prog-nosis breast cancer, indicating that exposure may adversely affect breast cancer prognosis. This study accentuates that coexposure to BPA and soy-based phytoestrogens results in additive estrogenic effects, and may contribute to estrogen-linked diseases, including breast cancer.
Key Words: bisphenol A; phytoestrogens; estrogen receptor; transcriptional activation; proliferation; concentration addition.
Endocrine-disrupting chemicals (EDCs) include a large group of synthetic agents that are capable of disrupting nor-mal hormonal signaling. Many EDCs, including bisphenol A (BPA), interfere with estrogen signaling. BPA, an industrial monomer found in plastics, can leach into food and drinks con-sumed by humans (Brotons et al., 1995; Singleton et al., 2004). Daily adult intake is between 0.05 and 10 μg/kg body weight per day and virtually all persons have measurable levels in their blood and other bodily fluids (Dong et al., 2011; Ikezuki et al., 2002; Ouchi and Watanabe, 2002; Sasaki et al., 2005). BPA has estrogenic activity, and it is well established that a high lifetime estrogen exposure correlates with an increased incidence of breast cancer (Yager and Davidson, 2006). Epidemiologic data indicate that BPA exposure is related to diseases of the breast in women (vom Saal and Hughes, 2005) and other reproduc-tive abnormalities (Newbold et al., 2007). Different risk assess-ments have led to varying conclusions regarding the safety of BPA, but it is agreed that infants have the highest BPA expo-sure in the general population (Beronius et al., 2010). Although fetuses and infants are normally shielded from endogenous estrogen exposure, BPA crosses the placenta and is excreted into breast milk. Formula-fed babies are further exposed to BPA through food, water, and plastic containers. The impact of such early-life exposure in humans is not known. In animals; how-ever, prenatal and early BPA exposure alters mammary gland development (Markey et al., 2001) in females and testis devel-opment in males (Krementsov et al., 2013), and prepubertal exposure increases the susceptibility to mammary carcinogens
Disclaimer: Certain commercial equipment, instruments, or materials that are identified in this manuscript are to foster understanding. Such identification does not imply recommendation or endorsement by the National Institute of Standards and Technology, nor does it imply that the materials or equipment identified are necessarily the best available for the purpose.
toxicological sciences 138(1), 21–35 2014doi:10.1093/toxsci/kft271Advance Access publication November 27, 2013
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from

KATCHY ET AL.
(Betancourt et al., 2012). We also know that prenatal exposure to another synthetic estrogen, diethylstilbestrol, increases the incidence of breast cancer in human adulthood (Collaborative group on Hormonal factors in Breast Cancer, 2002; Yager and Davidson, 2006). Moreover, current risk assessments of BPA have not taken into consideration the combined effect of expo-sure to multiple estrogenic compounds, such as phytoestrogens present in soy-based infant formula.
The most abundant phytoestrogens found in soybeans include the isoflavones, genistein and daidzein. Although con-sumption of soy-rich diets can be beneficial to adults, contro-versy has developed over the adequacy and safety of soy-based formula (SF) for infant use. The timing of exposure and endog-enous estrogen environment play critical roles in the potential chemopreventive or tumor-stimulating effects of genistein or soy products on mammary cancer in vivo (Betancourt et al., 2012; Helferich et al., 2008). Neonatal exposure to genistein has deleterious effects on the reproductive system in rodents (Cimafranca et al., 2010; Jefferson et al., 2005; Newbold et al., 2001). Soy-based infant formulas account for nearly 25% of the formula market in the United States (Bhatia and Greer, 2008). Infants fed SF are exposed to 6–11 mg/kg body weight of isofla-vones per day, while the intake of phytoestrogens from human milk (calculated to be <0.01 mg/day) is negligible in compari-son (Setchell et al., 1998). Measured circulating concentrations of total genistein and daidzein in 4-month-old infants fed SF are 2.53 ± 1.64 and 1.16 ± 0.23μM, respectively, and these lev-els are maintained constant due to frequent feeding and long half-life of the phytoestrogens (Setchell et al., 1997, 1998). The serum concentration levels in these babies are thereby 13 000–22 000 times higher than the levels of endogenous estradiol in early life (Winter et al., 1976), and may produce significant biological effects in infants (Setchell et al., 1998).
Toxicology testing today is largely performed on one com-pound at a time. However, there is a concern that different EDCs act in synergy and may result in so-called cocktail effects. The goal of this study was to determine the specific molecular and genomic effects that BPA, genistein, and extract of SF mediate in combination and in comparison to 17β-estradiol (E2). These compounds mediate their main activities through binding to the estrogen receptors (ER), ERα and ERβ. The ERs bind to their cognate DNA estrogen-responsive elements (EREs) and regulate gene transcription. It has been suggested that differ-ent ligand-dependent conformational changes allow different sets of cofactors to complex with the ERs, influencing the chro-matin binding and target gene transcription (Gage et al., 2012; Kershaw and Flier, 2004). Thus, different compounds may induce different targets genes via the same ER. Using HeLa cervical cancer reporter cell lines stably transfected with either ERα or ERβ, and ERα-positive MCF7 breast cancer cells, we investigated to what extent these agents mirror estrogen, whether they activate compound-specific target genes and non-ERα–mediated effects, and what the effects of coexposures are. An increased knowledge of the molecular actions of xeno- and
phytoestrogens will help to understand how coexposure to these compounds affects us and assists in risk assessment.
MATERIALS AnD METHoDS
Cell culture and treatments. MCF7 parental cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 5% fetal bovine serum (FBS) and 1% penicillin-streptomycin (Life Technologies, Grand Island, New York). Before treatment, the medium was changed to phenol red-free DMEM medium supplemented with 5% dextran-coated charcoal serum (DCC-FBS) for 24 h and then reduced to 0.5% DCC-FBS for 48 h.
For transactivation studies, we used HeLa cells stably transfected with an ERE-driven luciferase reporter gene (Glob-Luc-SVNeo) and with an expres-sion plasmid of human ERα (pSG5-Puro-hERα) or ERβ (pSG5-Puro-hERβ), referred to as HeLN-ERα and HeLN-ERβ (Escande et al., 2006). The HeLN cells were cultured in phenol red-free DMEM supplemented with 5% DCC, 1% glutamine, 1% penicillin, 1% streptomycin, 1 mg/ml G418, and 0.5 g/ml puro-mycin. All cells were incubated in a 5% CO
2 humidified atmosphere at 37°C.
The cells were treated with various concentrations, as indicated in each experiment, of E2, BPA, genistein, or daidzein, ICI 182,780 (all purchased from Sigma-Aldrich, St Louis, Missouri), SF, and ethanol, dimethyl sulfoxide or acetone/hexane (1:3) (Ac/Hx) as vehicle controls, diluted in fresh DMEM-5% DCC-FBS, for 0–48 h.
Preparation of SF. Extraction of 20 g fresh weight of soy-based infant formula was carried out on Accelerated Solvent Extractor (ASE 200; Dionex GmbH, Idstein, Germany). The procedure was performed using a mixture of n-hexane:acetone (Ac/Hx 75:25, vol/vol) at 120°C and at a pressure of 12 MPa (Cok et al., 2009). Two static cycles of 10 min were applied for a complete extrac-tion. Polychlorinated dibenzo-p-dioxins, dibenzofurans and polychlorinated biphe-nyls levels in human breast milk from different regions of Turkey.
Quantification of genistein and daidzein in SF extract. The levels of genistein and daidzein in the SF extract were measured by high-performance liquid chromatography (HPLC). Analyses were performed on a Binary HPLC pump 1525 (Waters, Milford, Massachusetts) equipped with an autosampler 2707 injector and a photodiode array detector 2998 set at 260 nm (Waters). HPLC profiles of genistein, daidzein, and SF extract were carried out with an HPLC system based on a Nucleodur C
18 column (250 × 4 mm, 5 μm; Macherey
Nagel, Bethlehem, Pennsylvania) in the following conditions (adapted from [Riu et al., 2008]): mobile phases: A: 20mM ammonium acetate pH 3.5/ace-tonitrile (95/5, vol/vol), B: 100% acetonitrile; gradient: 0–5 min, linear gradient from A: 100% to A:B 90:10; 5–25 min, linear gradient from A:B 90:10 to A:B 70:30; 25–35 min, linear gradient from A:B 70:30 to A:B 50:50; 35–36 min, linear gradient from A:B 50:50 to B 100%; 36–41 min, B: 100% isocratic. Under these conditions, daidzein and genistein were eluted with a retention time of 20.4 and 26.4 min, respectively. The isoflavones were quantified in the SF extract by measuring the area under the peak, based on their respective standard curves as previously established. In SF extract, the concentrations of genistein and daidzein were 81.0 and 101.3μM, respectively.
RNA preparation and cDNA synthesis. Treated cells were collected in TRIzol. RNA was extracted using the RNeasy Mini Kit (Qiagen, Valencia, California) with DNase I DNA degradation according to the manufactur-er’s protocol. RNA concentrations were measured using Nanodrop 1000 Spectrophotometer (Thermoscientific, Pittsburgh, Pennsylvania) and RNA integrity was measured using Agilent 2100 Bioanalyzer (Agilent, Santa Clara, California). For complementary DNA (cDNA) synthesis, 1 μg of the extracted total RNA was converted to cDNA using SuperScript III and random hexamer primers as previously described (Katchy et al., 2012).
qPCR analysis. In each quantitative PCR (qPCR) reaction, 10 ng cDNA was amplified using 1 pmol of forward and reverse primers, and 2X SYBR green PCR master mix (Life Technologies) in a 10-μl final reaction volume.
22
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from

COEXPOSURE TO PHYTOESTROGENS AND BISPHENOL A
ARHGDIA was used for normalization of messenger RNA expression. All runs were performed in triplicates using the 7500 Fast Real-Time PCR System (Life Technologies) and relative expressions were calculated using the comparative threshold cycle (ΔΔCt) formula. Melting curve analysis was performed for all reactions, confirming the amplification of one specific fragment. Student’s t test was used for statistical analysis, using 2-tailed distribution and 2-sample unequal variance parameters. A p value <.05 was considered significant.
Luciferase ERE transactivation reporter assay. HeLN-ERα and HeLN-ERβ cell lines were seeded at 4 × 104 cells/well in 96-well white opaque cell culture plates and maintained in DMEM without phenol red, supplemented with 5% DCC-FBS. Cells were incubated for 16 h with 10nM E2 (positive control), 0.00001%–0.1% SF (0.00001%, 0.0001%, 0.001%, 0.01%, and 0.1% of the original concentrated SF extract), 10−8 to 10−5 M BPA (10nM, 30nM, 100nM, 300nM, 1μM, 3μM, and 10μM). For genistein and daidzein, dose-response curves from 10−9 to 10−5 M (1nM, 3nM, 10nM, 30nM, 100nM, 300nM, 1μM, 10μM) were tested in HeLN-ERα cells and 10−10 to 10−6 M (0.1nM, 0.3nM, 1nM, 3nM, 10nM, 30nM, 100nM, 300nM, 1μM) in HeLN-ERβ cells. Media was removed and replaced with fresh media containing 0.3mM luciferin (Escande et al., 2006) and luminescence was measured for 2 s using the Victor X4 Multilabel Plate Reader (Perkin Elmer, Waltham, Massachusetts). Nonlinear regression curve fits were used to plot the dose-response curves using GraphPad Prism version 5 (GraphPad Software, La Jolla, California). Each treatment was performed in quadruplicates, and repeated in 3–5 individual experiments.
Concentration addition prediction. To determine the EC50 and hillslope for genistein, daidzein, BPA, and SF, the activation of luciferase expression was depicted as percentage of E2 response (10nM). Activities higher than 100% E2 response are commonly detected for phytoestrogens at higher doses, because of ER-independent unspecific activation (Escande et al., 2006). Therefore, activi-ties higher than 100% were set to 100%. The technical quadruplicates from each biological experiment were averaged, and the averages (3–5 values for each compound and dose) were used as input and plotted in GraphPad Prism using a 4-parameter logistic regression (4PL) curve fit to generate an EC50 and a hillslope for each compound.
Dose addition–predicted values were calculated using the concentration addi-tion (CA) model (power function written in Microsoft Excel format): R = (1/(1 + (1/(POWER((∑C
i/EC50
i), average hillslope))))) × 100. This formula calculates
a predictive effect of a mixture of chemicals, where R equals the response of the mixture, C
i is the concentration of each chemical in the mixture, EC50
i is the con-
centration of the chemical that causes a 50% response, and “i” refers to individual compounds (Rider and LeBlanc, 2005). EC50s and hillslope values were also placed in the Computational Approach to the Toxicity Assessment of Mixtures model developed by Dr Leblanc (http://www.ncsu.edu/project/toxresearch/model5/linked_files/ php_files /page3.php) to confirm our predicted responses. Observed and predicted data for mixture exposures were plotted in GraphPad Prism using a nonlinear regression (4PL) curve fit. Statistical calculations of 95% prediction intervals (PIs) for the mixtures were calculated using GraphPad Prism, accommodating for the variations between the biological experiments.
Proliferation assay. MCF7 cells were seeded in 96-well plates (5 × 103 cells/well) in DMEM supplemented with 10% FBS. Before treatment, cells were incubated in 5% DCC-FBS for 48 h and serum starved with 0.5% DCC-FBS for 24 h. Cells were treated with vehicle, 10nM E2, 0.01%–0.1% SF, or 0.1nM to 10μM of BPA, genistein, or daidzein, as described above. Proliferation was assayed 48 h after incubation with ligands using CellTiter 96 AQueous One Solution Cell Proliferation Assay [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium] (MTS) (Promega Corporation, Madison, Wisconsin) according to the manufacturer’s protocol. Each treatment was performed in quadruplicates, and repeated in 8 individual experiments. The technical quadruplicates were averaged for each biological experiment and used to calculate fold change in proliferation.
Microarray experiment and analysis. Operon’s long oligonucleotide-spotted array covering the known transcriptome of 35 000 genes and vari-ants, complemented with 133 oligos specifically synthesized for analysis of nuclear receptors, splice variants, and coregulators were used (Microarray Inc.,
Huntsville, Alabama). Using 2-color comparative microarray, each comparison was replicated and dye swapped (Cy5/Cy3) and analyzed essentially as pre-viously described (Williams et al., 2008). In brief, 15–20 μg total RNA was used per cDNA synthesis and labeling reaction, and hybridized to the arrays for 35–40 h at 42°C. The arrays were washed and scanned at 10-μm resolution using the Axon GenePix 4400A microarray scanner (MDS Analytical Technologies, Sunnyvale, California). Obtained TIFF images were analyzed using the GenePix Pro 6.1 software (MDS Analytical Technologies), and subsequent data analysis performed in the R environment as previously described (Richter et al., 2006). Differentially expressed genes were identified using the empirical Bayes mod-erated t test within the Limma package (Smyth, 2004). Cutoff for differential expression was set to p value <.05 and 2 different secondary cutoffs for logFC (>|0.7|, >|1.2|) were investigated. Microarray data is available on NCBI’s GEO under accessions: GSE45557 (E2) and GSE45721 (BPA, genistein, and SF).
Bioinformatics. Gene set enrichment analysis and subnetwork enrichment analyses were performed for differentially expressed genes using the Pathway Studio program (Elsevier Inc., Maryland). Fisher’s exact test was applied to determine enriched Gene Ontology functional groups among the differentially expressed genes, and p values <.05 were considered significant.
Survival cluster analysis. The expression profiles of the differentially expressed genes were used to group 258 primary breast cancer samples from a patient cohort from Uppsala, Sweden, whose expression profiles have been previ-ously defined (Miller et al., 2005), using hierarchical clustering (Eisen Cluster and TreeView). Disease-free survivals of the clusters were graphed using the Kaplan-Meier plot function in the SigmaPlot software, and significant associations with disease parameters (ERα status, progesterone receptor (PR) status, lymph node positivity, grade, recurrence, distant metastasis, death) were determined using Fisher’s exact test.
RESULTS
Comparison of Dose-Dependent Transcriptional ER Activation by BPA, Genistein, Daidzein, and SF
We first set out to determine at what concentrations BPA, genistein, daidzein, and SF induced ERα and ERβ transacti-vation in HeLa cells. We observed that all tested compounds mediated a dose-dependent transcriptional activation of the ERE-luciferase construct in both HeLN-ERα and ERβ cell lines. Genistein induced ERα-mediated transactivation at 10nM, whereas ERβ was activated already at 1nM (Figs. 1A–C). At concentrations higher than 1μM, both genistein and daidzein exceeded maximal E2 response (10nM). These results agree with previously published data (Escande et al., 2006).
BPA reached a maximal transactivation efficacy at approxi-mately 3μM, peaking at approximately 80% activation com-pared with 10nM E2 (defined as 100% activation). At 100nM, BPA activated both ERα and ERβ. Similar to genistein and daidzein, SF showed a higher potency toward ERβ than ERα (Figs. 1B and D) and at approximately 0.01% (ERβ) and 0.1% (ERα), it mediated transcriptional activation similar to 10nM E2. The EC50s for all compounds and SF are shown in Supplementary Table S1.
Next, we analyzed the dose-dependent transcriptional acti-vation of endogenous ERα in MCF7 cells. We and others have previously found that a 24-h exposure to E2 produces signifi-cant changes in ERα-mediated gene transcription in breast can-cer cells (Alvarez-Baron et al., 2011; Williams et al., 2008). We
23
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from
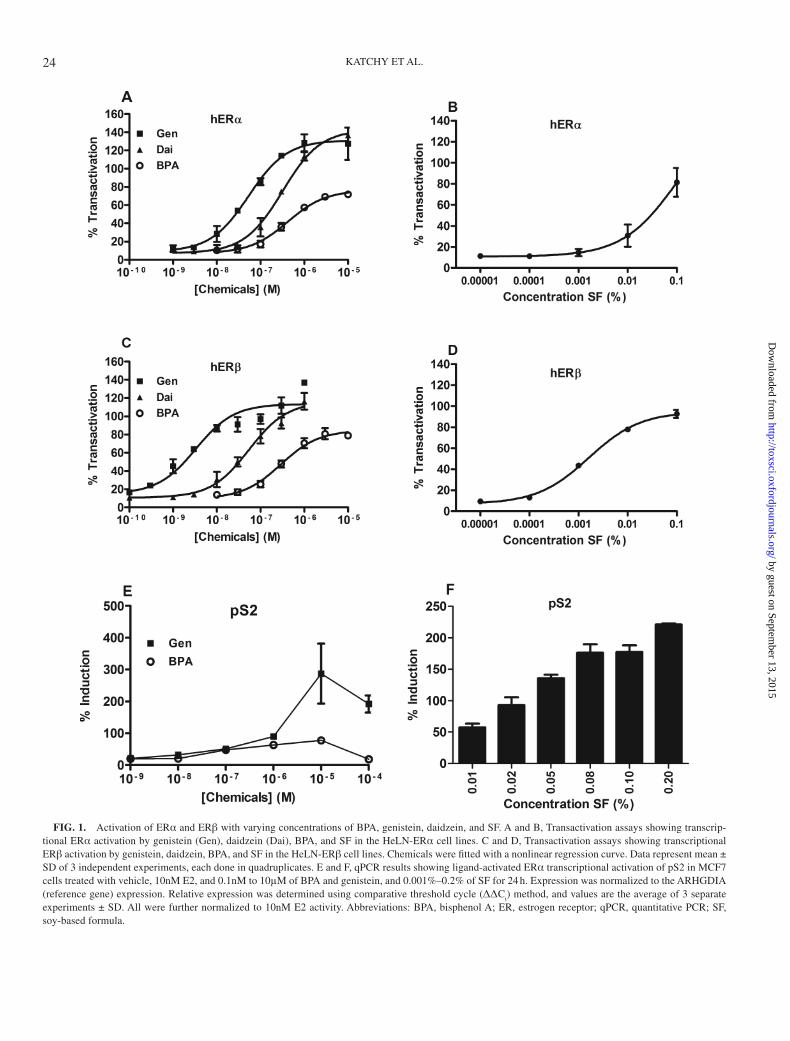
KATCHY ET AL.
FIG. 1. Activation of ERα and ERβ with varying concentrations of BPA, genistein, daidzein, and SF. A and B, Transactivation assays showing transcrip-tional ERα activation by genistein (Gen), daidzein (Dai), BPA, and SF in the HeLN-ERα cell lines. C and D, Transactivation assays showing transcriptional ERβ activation by genistein, daidzein, BPA, and SF in the HeLN-ERβ cell lines. Chemicals were fitted with a nonlinear regression curve. Data represent mean ± SD of 3 independent experiments, each done in quadruplicates. E and F, qPCR results showing ligand-activated ERα transcriptional activation of pS2 in MCF7 cells treated with vehicle, 10nM E2, and 0.1nM to 10μM of BPA and genistein, and 0.001%–0.2% of SF for 24 h. Expression was normalized to the ARHGDIA (reference gene) expression. Relative expression was determined using comparative threshold cycle (ΔΔC
t) method, and values are the average of 3 separate
experiments ± SD. All were further normalized to 10nM E2 activity. Abbreviations: BPA, bisphenol A; ER, estrogen receptor; qPCR, quantitative PCR; SF, soy-based formula.
24
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from
COEXPOSURE TO PHYTOESTROGENS AND BISPHENOL A
assessed the classical ERE target gene pS2 after treatment with various concentrations of each compound. Similar to the HeLN-ERα cells, genistein and BPA mimicked the effect of 10nM E2 at 1μM concentrations. At higher concentrations, genistein induced a stronger response than 10nM E2 (Fig. 1E). SF extract induced endogenous pS2 transcription at 0.01% concentration and its efficacy was comparable to 10nM E2 at 0.02%–0.05% (Fig. 1F).
BPA and Phytoestrogens Have Additive Effects on ER-Mediated Transcriptional Activation
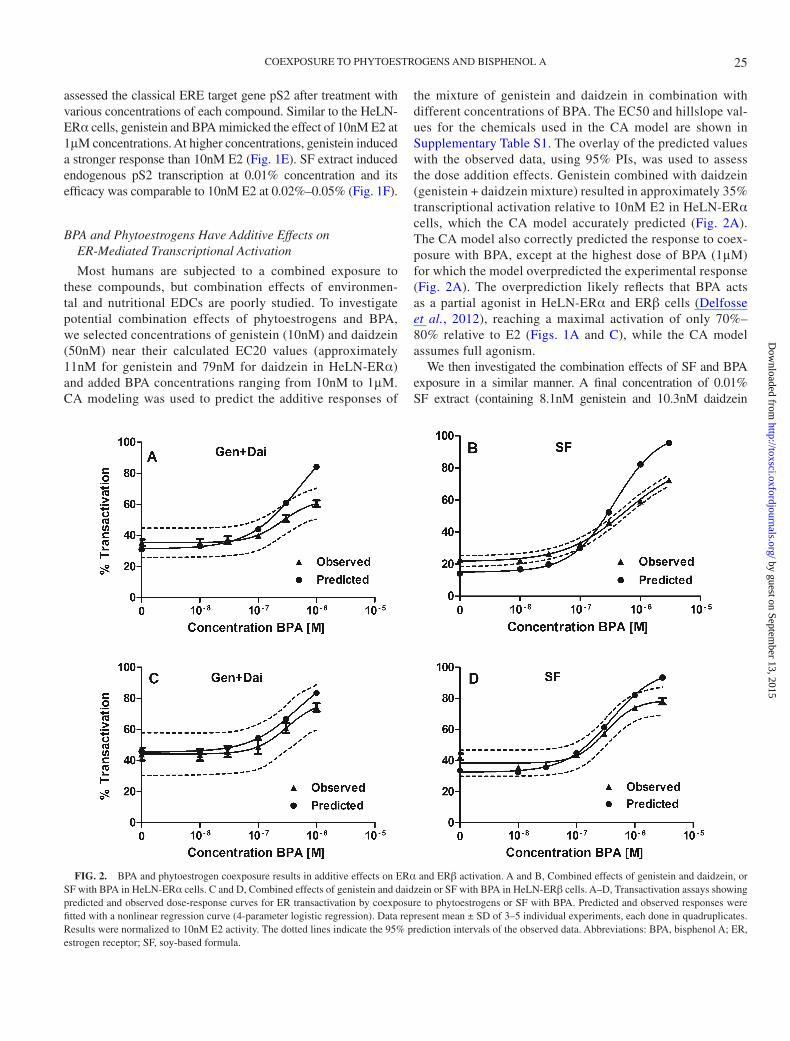
Most humans are subjected to a combined exposure to these compounds, but combination effects of environmen-tal and nutritional EDCs are poorly studied. To investigate potential combination effects of phytoestrogens and BPA, we selected concentrations of genistein (10nM) and daidzein (50nM) near their calculated EC20 values (approximately 11nM for genistein and 79nM for daidzein in HeLN-ERα) and added BPA concentrations ranging from 10nM to 1μM. CA modeling was used to predict the additive responses of
the mixture of genistein and daidzein in combination with different concentrations of BPA. The EC50 and hillslope val-ues for the chemicals used in the CA model are shown in Supplementary Table S1. The overlay of the predicted values with the observed data, using 95% PIs, was used to assess the dose addition effects. Genistein combined with daidzein (genistein + daidzein mixture) resulted in approximately 35% transcriptional activation relative to 10nM E2 in HeLN-ERα cells, which the CA model accurately predicted (Fig. 2A). The CA model also correctly predicted the response to coex-posure with BPA, except at the highest dose of BPA (1μM) for which the model overpredicted the experimental response (Fig. 2A). The overprediction likely reflects that BPA acts as a partial agonist in HeLN-ERα and ERβ cells (Delfosse et al., 2012), reaching a maximal activation of only 70%–80% relative to E2 (Figs. 1A and C), while the CA model assumes full agonism.
We then investigated the combination effects of SF and BPA exposure in a similar manner. A final concentration of 0.01% SF extract (containing 8.1nM genistein and 10.3nM daidzein
FIG. 2. BPA and phytoestrogen coexposure results in additive effects on ERα and ERβ activation. A and B, Combined effects of genistein and daidzein, or SF with BPA in HeLN-ERα cells. C and D, Combined effects of genistein and daidzein or SF with BPA in HeLN-ERβ cells. A–D, Transactivation assays showing predicted and observed dose-response curves for ER transactivation by coexposure to phytoestrogens or SF with BPA. Predicted and observed responses were fitted with a nonlinear regression curve (4-parameter logistic regression). Data represent mean ± SD of 3–5 individual experiments, each done in quadruplicates. Results were normalized to 10nM E2 activity. The dotted lines indicate the 95% prediction intervals of the observed data. Abbreviations: BPA, bisphenol A; ER, estrogen receptor; SF, soy-based formula.
25
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from

KATCHY ET AL.
according to the quantification of the respective isoflavones in the SF extract) was used in combination with concentrations of BPA ranging from 10nM to 3μM. The EC50s and hillslopes of gen-istein and daidzein were used for the CA predictions of the mix-ture effects of the SF with BPA, because most of the estrogenic activity induced by soy-based infant formula has been associated with the presence of these phytoestrogens (Riu et al., 2008). The effects observed with SF in combination with low BPA concentra-tions (10 and 30nM BPA) were slightly higher than the predicted values, and at higher BPA concentrations the model overpredicted the observed response (Fig. 2B), similarly to what was observed with the mixture of genistein, daidzein, and BPA.
Similarly, in order to analyze the effects of coexposure to phytoestrogens and BPA through ERβ, a combination of 1nM genistein and 10nM daidzein (calculated EC20 values were 0.37 and 9.5nM, respectively), or 0.001% SF was selected. The CA model prediction fitted into the 95% PIs for all concentra-tions of genistein and daidzein combined with BPA, whereas the model overpredicted the effect of SF with the highest con-centration of BPA (3μM) (Figs. 2C–D).
Overall, our data shows that low BPA concentrations exert an additive effect when combined with the phytoestrogens pre-sent in soy formula, whereas high BPA concentrations display a subadditive effect likely due to the partial agonism of BPA in the HeLN cell context.
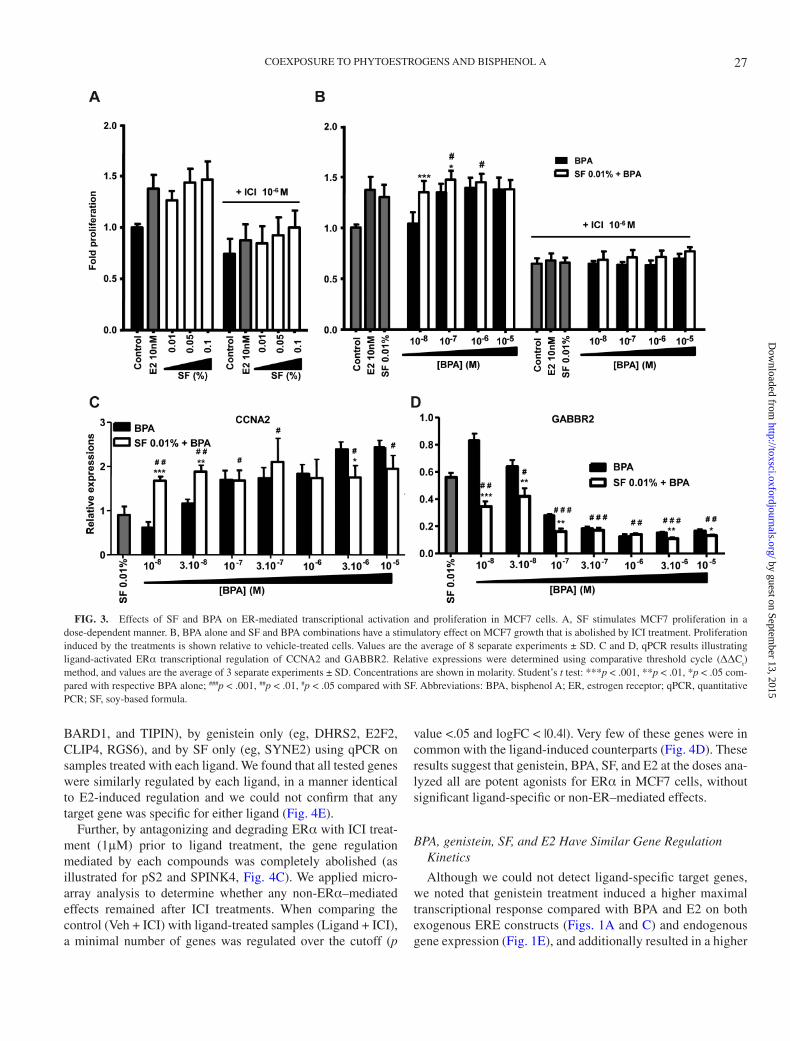
BPA and SF Treatment Mimic E2-Induced Proliferation of Breast Cancer Cells
E2-activated ERα is known to drive proliferation of ERα-positive breast cancer cells. Because both BPA and SF were able to activate transcription through ERα, they may induce prolif-eration of breast cancer cells similarly to E2. We compared pro-liferation of MCF7 cells after the addition of SF and BPA, alone or in mixture, to that of E2-treated cells. E2, at a concentration of 10nM, induced cell growth by 38% compared with vehicle control. SF at 0.01%, 0.05%, and 0.1% significantly increased cell proliferation by 26%, 43%, and 46% relative to control, respectively (Fig. 3A). BPA alone at 10nM had no significant effect on MCF7 proliferation, whereas 100nM, 1μM, and 10μM of BPA stimulated cell growth by 34%, 39%, and 37%, respectively (Fig. 3B). Significant increase on cell proliferation was observed when 100nM BPA was added to 0.01% SF, com-pared with either BPA or SF alone (Fig. 3B). Cotreatment with 1μM (7α,17β)-7-{9-[(4,4,5,5,5-pentafluoropentyl)sulfinyl]nonyl}estra-1,3,5(10)-triene-3,17-diol (ICI) abolished the pro-liferative effect induced by SF, BPA, or a combination thereof, demonstrating that the proliferative effect was only mediated through ER (Figs. 3A and B). Since we know that these MCF7 cells do not express significant levels of ERβ (Jonsson et al., unpublished data) (Holbeck et al., 2010), the measured activity of BPA and SF on proliferation was through ERα.
To explore whether these compounds had combination effects also on endogenous ERα-mediated transcriptional activation,
we performed a parallel analysis in MCF7 cells (Figs. 3C and D). We investigated the combination of SF (0.01%) and BPA (10nM to 10μM) on one known ERα-induced cell cycle gene, cyclin A2 (CCNA2), and one ERα-repressed gene (GABBR2). Compared with the response to 0.01% SF extract, we observed a significant increase in the upregulation of CCNA2 (Fig. 3C) and the repression of GABBR2 (Fig. 3D) when low concentra-tions of BPA (10 and 30nM, and also 100nM for GABBR2 only) were combined with the 0.01% SF extract. These results show that a coexposure of either genistein/daidzein or SF together with low concentrations of BPA activates the ERs to a higher degree than each compound or extract individually does.
BPA, Genistein, SF, and E2 Induce Identical Transcriptional Responses Through ERα
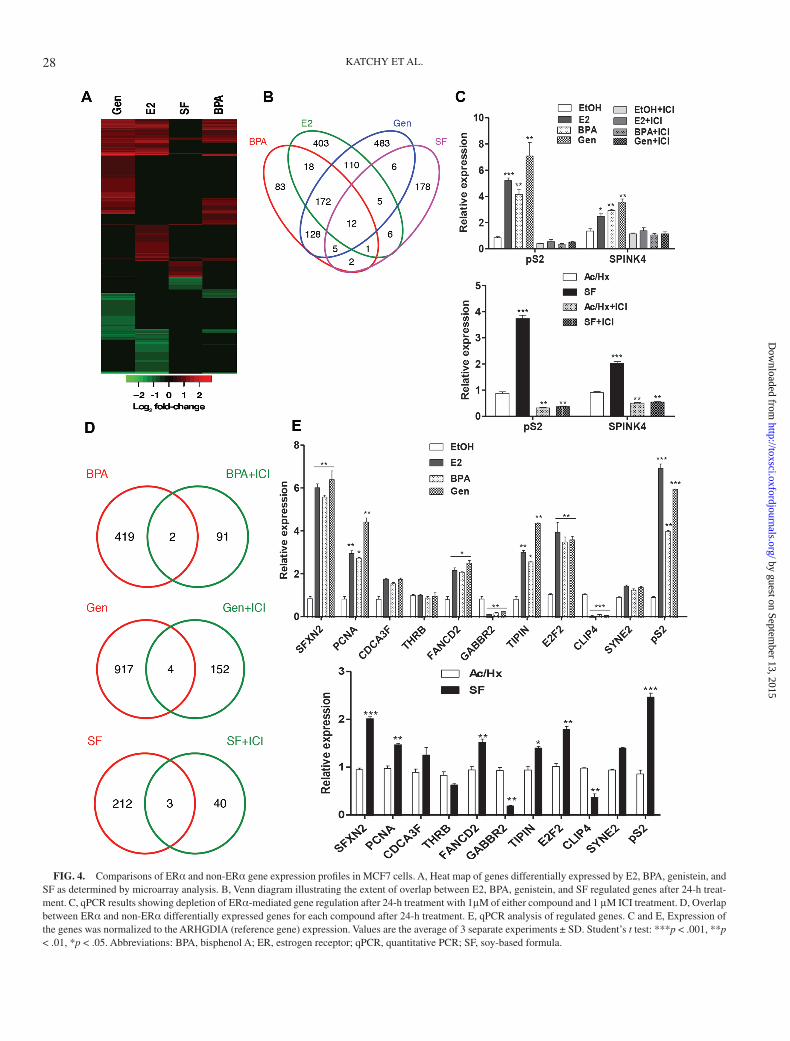
It has been suggested that different ligands induce differ-ent conformational changes of ERα, resulting in ligand-spe-cific gene regulation (Singleton et al., 2006). BPA has, eg, been reported to induce a different set of genes compared with E2 (Singleton et al., 2006). In addition, genistein has ER-independent activities that may result in regulation of a distinct subset of genes (Lucki and Sewer, 2011; Maggiolini et al., 2004). In order to define ligand-specific target genes in MCF7 cells for these compounds, we employed microarray gene expression profiling, followed by qPCR confirmations. At selected cutoff (p value <.05, logFC > |0.7|), BPA significantly regulated 421 genes, E2 727 genes, genistein 921 genes, and SF 215 genes (Figs. 4A and B). A comparison of target genes showed that whereas BPA affected the least number of genes and genistein the highest, many genes were similarly regu-lated by all tested ligands. In addition, the microarray analy-sis identified significant ligand-specific gene sets. This is in line with previous observations also using microarray analysis (Satih et al., 2010; Singleton et al., 2006). To classify our array gene list and explore if the different ligands induced different biological effects, we searched for biological processes that were overrepresented upon treatment by the different ligands. However, we found that most processes were similarly affected by all ligands (Supplementary Tables S2–S4), and no ligand induced processes that were distinctly different from all ligands (Table 1). Each ligand, including E2, affected regulation of the cell cycle, cell migration, DNA repair, mitogen-activated pro-tein kinase activity/cell signaling, lipid metabolism, and cho-lesterol homeostasis.
Although the microarray technique is useful for genome-wide screening purposes, the technique is optimized to detect genes that are changed, not to define genes that are not changed. Therefore, genes that the array detected as regulated by one ligand but not by another one need to be confirmed by qPCR. Thus, we investigated the regulation of genes that the array results indicated were regulated by all compounds (eg, SFXN2 and PCNA), regulated by BPA only (eg, CDCA3F and THRB), by BPA and genistein but not E2 (eg, FANCD2, GABBR2,
26
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from
COEXPOSURE TO PHYTOESTROGENS AND BISPHENOL A
BARD1, and TIPIN), by genistein only (eg, DHRS2, E2F2, CLIP4, RGS6), and by SF only (eg, SYNE2) using qPCR on samples treated with each ligand. We found that all tested genes were similarly regulated by each ligand, in a manner identical to E2-induced regulation and we could not confirm that any target gene was specific for either ligand (Fig. 4E).
Further, by antagonizing and degrading ERα with ICI treat-ment (1μM) prior to ligand treatment, the gene regulation mediated by each compounds was completely abolished (as illustrated for pS2 and SPINK4, Fig. 4C). We applied micro-array analysis to determine whether any non-ERα–mediated effects remained after ICI treatments. When comparing the control (Veh + ICI) with ligand-treated samples (Ligand + ICI), a minimal number of genes was regulated over the cutoff (p
value <.05 and logFC < |0.4|). Very few of these genes were in common with the ligand-induced counterparts (Fig. 4D). These results suggest that genistein, BPA, SF, and E2 at the doses ana-lyzed all are potent agonists for ERα in MCF7 cells, without significant ligand-specific or non-ER–mediated effects.
BPA, genistein, SF, and E2 Have Similar Gene Regulation Kinetics
Although we could not detect ligand-specific target genes, we noted that genistein treatment induced a higher maximal transcriptional response compared with BPA and E2 on both exogenous ERE constructs (Figs. 1A and C) and endogenous gene expression (Fig. 1E), and additionally resulted in a higher
FIG. 3. Effects of SF and BPA on ER-mediated transcriptional activation and proliferation in MCF7 cells. A, SF stimulates MCF7 proliferation in a dose-dependent manner. B, BPA alone and SF and BPA combinations have a stimulatory effect on MCF7 growth that is abolished by ICI treatment. Proliferation induced by the treatments is shown relative to vehicle-treated cells. Values are the average of 8 separate experiments ± SD. C and D, qPCR results illustrating ligand-activated ERα transcriptional regulation of CCNA2 and GABBR2. Relative expressions were determined using comparative threshold cycle (ΔΔC
t)
method, and values are the average of 3 separate experiments ± SD. Concentrations are shown in molarity. Student’s t test: ***p < .001, **p < .01, *p < .05 com-pared with respective BPA alone; ###p < .001, ##p < .01, #p < .05 compared with SF. Abbreviations: BPA, bisphenol A; ER, estrogen receptor; qPCR, quantitative PCR; SF, soy-based formula.
27
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from
KATCHY ET AL.
FIG. 4. Comparisons of ERα and non-ERα gene expression profiles in MCF7 cells. A, Heat map of genes differentially expressed by E2, BPA, genistein, and SF as determined by microarray analysis. B, Venn diagram illustrating the extent of overlap between E2, BPA, genistein, and SF regulated genes after 24-h treat-ment. C, qPCR results showing depletion of ERα-mediated gene regulation after 24-h treatment with 1μM of either compound and 1 μM ICI treatment. D, Overlap between ERα and non-ERα differentially expressed genes for each compound after 24-h treatment. E, qPCR analysis of regulated genes. C and E, Expression of the genes was normalized to the ARHGDIA (reference gene) expression. Values are the average of 3 separate experiments ± SD. Student’s t test: ***p < .001, **p < .01, *p < .05. Abbreviations: BPA, bisphenol A; ER, estrogen receptor; qPCR, quantitative PCR; SF, soy-based formula.
28
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from

COEXPOSURE TO PHYTOESTROGENS AND BISPHENOL A
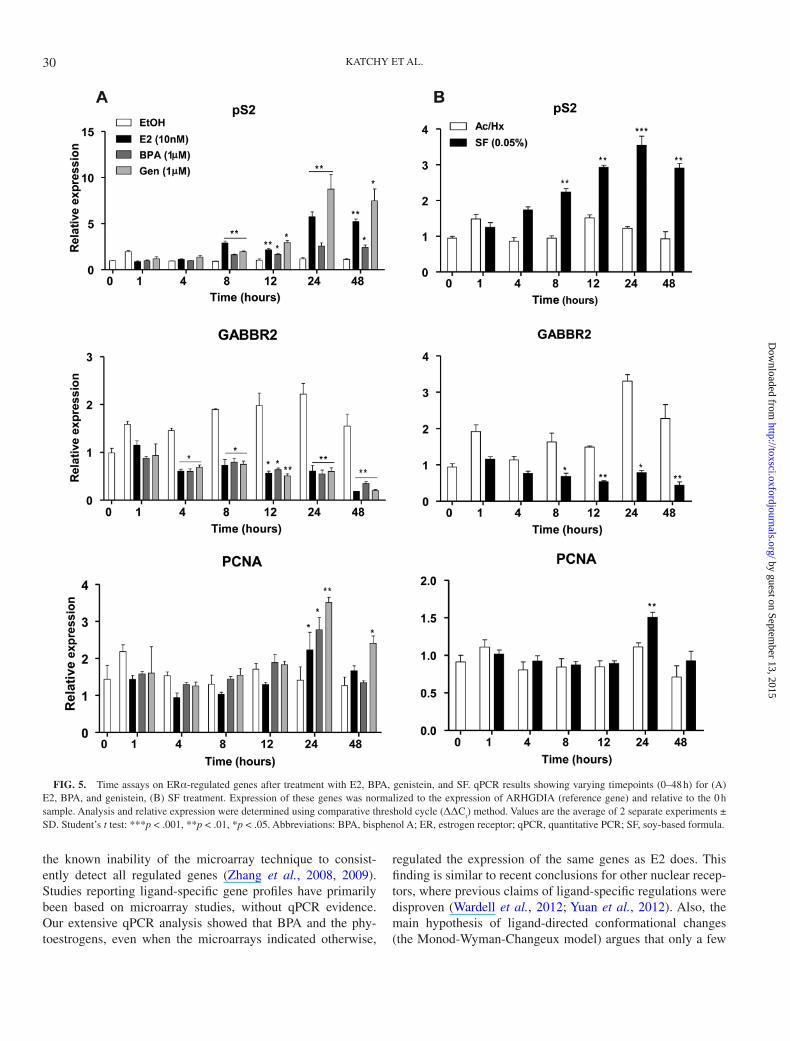
detection frequency by the microarray analysis (Fig. 4B). Upon investigation using qPCR, we noted that genistein (1μM) induced the expression of certain, but not all, endog-enous transcription targets to a higher extent than 10nM E2 (Fig. 4E). We speculated that there might be a ligand-specific kinetics of transcription. Thus, a time-dependent assay was conducted, comparing the kinetics of E2 (10nM), BPA (1μM), genistein (1μM), and SF (0.05%) in mediating gene regula-tion of pS2, GABBR2, and proliferating cell nuclear antigen (PCNA) in MCF7 cells. For pS2, all compounds induced sig-nificant upregulation of transcription at 8 h, reaching the maxi-mal response at 24 h (Figs. 5A and B). PCNA had a weaker and later response and was induced by all compounds at 24 h. We noted that for the upregulated genes, genistein mediated an enhanced response compared with E2 at later timepoints (24–48 h). Expression of the GABBR2 gene was quickly repressed after treatment by each compound, was maximally repressed at 48 h, and no difference in the kinetics between genistein and E2 treatment was noted. This indicated that although the kinet-ics of ERα-mediated regulation may be gene specific, they were not ligand specific, with the exception that genistein has a higher efficacy at later timepoints (24 and 48 h) for the induced genes compared with BPA (1μM) or E2 (10nM).
BPA- and Phytoestrogen-Induced Gene Profile Define Poor Prognosis of Patients
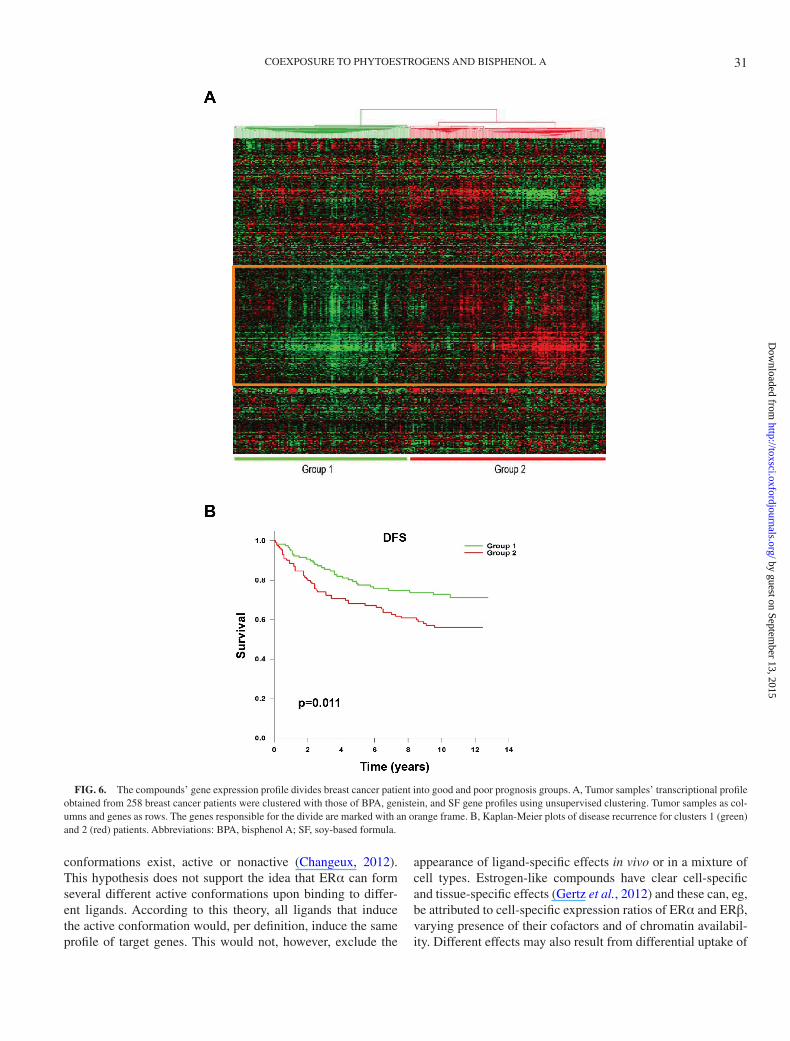
To explore the impact that exposure to these compounds may have on breast cancer, we clustered the gene expression profile of 258 primary breast tumors with the transcript profile that these compounds induced. The transcriptional profiles of the breast tumors and corresponding clinicopathological variables have been described previously (Miller et al., 2005). After establishing that all compounds induced the same target genes in breast cancer cells, based on results of the above microar-ray and qPCR confirmation, we generated their combined
transcription profile composed by genes detected as regulated by treatment with at least 2 different ligands. We then explored whether this profile would divide the clinical breast tumors into groups with different clinicopathological parameters and/or variable survival parameters. Unsupervised clustering using this profile did separate the patients into 2 groups (Figs. 6A and B), with significant differences in terms of tumor grade, ER/PR positivity, lymph node status, and outcome (Table 2). We observed similar results when each compound’s transcription profile was analyzed separately (data not shown). This suggests that genes regulated by these compounds in MCF7 cells have a physiologic relevance to breast cancer and gene expression in vivo. We further evaluated the genes responsible for the cluster division (orange box in Fig. 6A) for how they were regulated by each compound. We found that genes upregulated in the poor prognosis group were consistently upregulated by the different compounds in the MCF7 cells (Supplementary Table S5).
Combined, we show that BPA and phytoestrogens mimic E2-induced gene expression changes in ERα-positive breast cancer cells, and these gene regulations correlate to poor prog-nosis in breast cancer patients.
DISCUSSIon
The constant human exposure to environmental and nutri-tional compounds with endocrine abilities, such as BPA and phytoestrogens, accentuates the urgency of understanding the effects of these compounds on human health. Since estrogenic activity is linked to development of breast cancers, a risk assess-ment in this respect is critical. It is also important to increase the knowledge of how EDCs affect the normal hormonal sign-aling at a molecular level. Thus, we have here performed both functional and gene expression–profiling analysis of selected compounds in order to decipher their molecular mechanism.
We investigated different xeno- and phytoestrogenic com-pounds and complex mixtures and compared them to E2 in terms of transcriptional activation of exogenous ERE constructs, endogenous genome-wide regulation, and proliferative potential for breast cancer cells. We also explored the extent of coexposure effects using CAs and statistical modeling prediction. We found that all ligands mimicked E2, and we did not find any evidence for ligand-specific target genes. We showed that, at the doses and in the models analyzed here, the responses of cells to phytoestrogens and BPA were equivalent to those of pure estrogen. We found that low doses of phytoestrogens (genistein, daidzein, and SF) together with BPA resulted in additive effects on transcriptional activation, and we found that coexposure also enhanced endog-enous gene regulation and functional effects on proliferation.
Regulated Target Genes and Kinetics of Their Expression
Although our analyses taken together do not support previ-ous findings of ligand-specific target genes for these ligands in MCF7 cells (Satih et al., 2010), our microarray analysis alone seemed to support this notion. We suggest that this reflects
TABLE 1Biological Functions overrepresented Among the Genes Detected as Commonly Regulated by BPA, Genistein, SF, and E2 in MCF7
breast cancer cells
Biological Process p value
Nucleosome assembly 1.08E-13Cell cycle 2.94E-05DNA replication 4.56E-04DNA repair 1.18E-03Telomere maintenance 2.86E-03Cell division 9.85E-03DNA recombination 1.12E-02Regulation of phosphorylation 2.14E-02Actin cytoskeleton organization 3.04E-02Nucleotide-excision repair 3.26E-02Glycolysis 3.82E-02
Abbreviations: BPA, bisphenol A; SF, soy-based formula.
29
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from
KATCHY ET AL.
the known inability of the microarray technique to consist-ently detect all regulated genes (Zhang et al., 2008, 2009). Studies reporting ligand-specific gene profiles have primarily been based on microarray studies, without qPCR evidence. Our extensive qPCR analysis showed that BPA and the phy-toestrogens, even when the microarrays indicated otherwise,
regulated the expression of the same genes as E2 does. This finding is similar to recent conclusions for other nuclear recep-tors, where previous claims of ligand-specific regulations were disproven (Wardell et al., 2012; Yuan et al., 2012). Also, the main hypothesis of ligand-directed conformational changes (the Monod-Wyman-Changeux model) argues that only a few
FIG. 5. Time assays on ERα-regulated genes after treatment with E2, BPA, genistein, and SF. qPCR results showing varying timepoints (0–48 h) for (A) E2, BPA, and genistein, (Β) SF treatment. Expression of these genes was normalized to the expression of ARHGDIA (reference gene) and relative to the 0 h sample. Analysis and relative expression were determined using comparative threshold cycle (ΔΔC
t) method. Values are the average of 2 separate experiments ±
SD. Student’s t test: ***p < .001, **p < .01, *p < .05. Abbreviations: BPA, bisphenol A; ER, estrogen receptor; qPCR, quantitative PCR; SF, soy-based formula.
30
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from
COEXPOSURE TO PHYTOESTROGENS AND BISPHENOL A
conformations exist, active or nonactive (Changeux, 2012). This hypothesis does not support the idea that ERα can form several different active conformations upon binding to differ-ent ligands. According to this theory, all ligands that induce the active conformation would, per definition, induce the same profile of target genes. This would not, however, exclude the
appearance of ligand-specific effects in vivo or in a mixture of cell types. Estrogen-like compounds have clear cell-specific and tissue-specific effects (Gertz et al., 2012) and these can, eg, be attributed to cell-specific expression ratios of ERα and ERβ, varying presence of their cofactors and of chromatin availabil-ity. Different effects may also result from differential uptake of
FIG. 6. The compounds’ gene expression profile divides breast cancer patient into good and poor prognosis groups. A, Tumor samples’ transcriptional profile obtained from 258 breast cancer patients were clustered with those of BPA, genistein, and SF gene profiles using unsupervised clustering. Tumor samples as col-umns and genes as rows. The genes responsible for the divide are marked with an orange frame. Β, Kaplan-Meier plots of disease recurrence for clusters 1 (green) and 2 (red) patients. Abbreviations: BPA, bisphenol A; SF, soy-based formula.
31
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from

KATCHY ET AL.
ligands by the different tissues or cells, and by different rates of metabolism, stability, and activation potential. Thus, differ-ent cell contexts can result in ligand-specific effects, albeit this may not necessarily be through ligand-specific target genes.
All proliferative and genome-wide transcriptional effects of BPA, genistein, and SF in our study could be inhibited by ICI treatment, showing that the effects are mediated exclusively through the ERs. Since the MCF7 cells used here express ERα and no functionally significant levels of ERβ (Jonsson et al., unpublished data), the effects of the 3 compounds in these cells were mediated solely through ERα. For unknown reasons, breast cancer cell lines often lose expression of ERβ (Holbeck et al., 2010). In other cells though, the effects would also depend on the levels of ERβ. In fact, many phytoestrogens are somewhat selective for ERβ (Kuiper et al., 1998; Turner et al., 2007). The gene regulatory pattern mediated by BPA, phytoestrogens, or E2 via ERβ was not explored in this study.
We did not detect any non-ER–mediated effects of the above compounds in this study. Other reports that have indicated that phytoestrogens and BPA may induce non-ER–mediated path-ways, eg, through activation of the G-protein–coupled recep-tor 30 (GPR30), Erk1/2, and c-fos transcription (Dong et al., 2011; Stratton and Rahman, 2008). These studies, however, used higher concentration (10μM) than we used in our genome-wide analysis (1μM). Other studies support that BPA lacks non-genomic activity at low concentrations (Bulayeva and Watson, 2004). Isoflavones, especially genistein, have been reported to have agonistic effects at low doses in breast cancer cell, but antagonistic effects at high doses (Wu et al., 2008). We observed significant changes in relative gene expression for each com-pound also at low doses. Our dose-dependent proliferation analysis indicated that both low (100nM) and high BPA concen-trations (up to 10μM) as well as SF concentrations mimicked
the proliferative effect of E2, and were completely abolished by ICI (Fig. 3B). We therefore conclude that these compounds can mimic the effect of E2 at both low and high doses.
We noted higher efficacy in transcriptional responses to gen-istein for many target genes at later timepoints (24 and 48 h, Figs. 4E and 5). We speculate that a reason for the higher efficacy might be related to ER stability and that different ligands, due to their structural shape, may affect the degrada-tion pattern of the ERα protein differently. Another reason for ligand-specific efficacy could be that the binding affinity or the metabolism of the compounds varies and therefore the extent of receptor activation over time may differ.
Additive Effects of EDCs and Implications for Breast Cancer Development
The fact that humans are continually exposed to a mixture of endocrine-disrupting compounds raises concerns with regard to health risk. Although the levels of BPA that humans are exposed to are low, there is increasing evidence of low-dose effects, eg, on the induction of chemoresistance to multiple anticancer drugs (Lapensee et al., 2009; Singh and Li, 2012; Weng et al., 2010; Wolstenholme et al., 2012). We demonstrated that com-bined exposure to BPA and genistein or SF results in additive activation of ER-mediated transcriptional transactivation and also enhanced their proliferative effects. We noted that all com-pounds could induce proliferation of breast cancer cells similarly to estrogen, at both lower (100nM) and higher doses (10μM). The compounds also regulated the same genes and had similar kinetics. Given that the EDCs in our study all acted similarly and in an additive manner, it is possible that the dietary intake of phytoestrogens will add to the effect of BPA exposure and enhance estrogenic exposure. When we related the compound-regulated genes to breast cancer gene expression, we found that there was a significantly lower disease-free survival for patients whose tumors had a higher expression of the genes that BPA, genistein, and SF upregulated. This suggests that such exposure may have adverse effects in terms of breast cancer. The expo-sure to BPA in the environment has also been reported to induce tumor aggressiveness and poor patient outcome (Dairkee et al., 2008). On the other hand, there may be other positive effects of consuming phytoestrogens, possibly through activation of ERβ and/or effects in other tissues (reviewed in [Messina and Wood, 2008; Zhang et al., 2010]). Based on our studies, we conclude that BPA and phytoestrogens can have additive effects, which are similar to E2-mediated effects through ERα, and these effects may be negative from a breast cancer perspective.
Infant Exposure to EDCs and Phytoestrogens
Infants who are fed SF have high levels of circulating phy-toestrogens that may produce significant biological effects (as referenced in introduction). For example, a correlation between soy formula intake during infancy and uterine leiomyoma in adult women has been noted (D’Aloisio et al., 2010). We
TABLE 2Clinicopathological Parameters and Disease outcome for the 2 Patient Groups That Were Separated by Their Correlation
to the BPA/Phytoestrogen Gene Expression Profile, as Illustrated in Figure 6
Cluster 1 (n = 118) Cluster 2 (n = 140) p value
LN+ 26 58 5.82E-04LN− 89 76ER+ 110 108 4.43E-05ER− 5 31PgR+ 103 89 1.36E-05PgR− 15 51G1+G2 115 83 0G3 1 54Recurrence (DFS) 32 59 0.011Metastasis (DMFS) 23 46 0.025Death (DSS) 17 38 0.007
Abbreviations: BPA, bisphenol A; DFS, disease-free survival; DMFS, dis-tant metastasis-free survival; DSS, disease-specific survival; ER, estrogen receptor; G1–G3, tumor grade; LN, lymph node; PgR, progesterone receptor.
32
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from

COEXPOSURE TO PHYTOESTROGENS AND BISPHENOL A
observed that SF was able to fully activate both estrogen recep-tors and could stimulate the proliferation of MCF7 breast can-cer cells with the same efficacy as 10nM E2 (Figs. 1 and 3). As infants also have high levels of BPA in their blood, up to 11 times higher than in adults or in the 1–19.4nM range (Edginton and Ritter, 2009; Vandenberg et al., 2007), the effects of com-bined exposure of BPA and phytoestrogens are important to assess. Our results demonstrate that MCF7 cell proliferation can be induced by 100nM BPA, a concentration that could pos-sibly be reached in the plasma of newborns according to this toxicokinetic model. Further, coexposure to SF and BPA at concentrations relevant to human early-life exposure resulted in additive effect on transcription activation of E2 target genes. These findings raise questions on the potential adverse effects induced by SF feeding in early life, in particular when taking into account a possible coexposure to EDCs, such as BPA.
In conclusion, EDCs present in our immediate environment and diet are potential health risks, because they modulate hor-monal signaling. This study contributes to an increased under-standing of the mechanisms of EDC action in that we show that phyto- and xenoestrogens activate the same ERα target genes as E2 does, and increase cell proliferation of MCF7 breast cancer cells. We further show that the combined exposure to BPA and phytoestrogens increases transcription in an additive manner. Considering that humans are exposed to thousands of chemicals, both through environmental pollution and through the diet, a thorough analysis of additive effects of mixtures and their physiological effects in humans need to be undertaken to create a more reliable risk assessment of EDCs.
SUPPLEMEnTARy DATA
Supplementary data are available online at http://toxsci.oxfordjournals.org/.
FUnDInG
Environmental Protection Agency (R834289 to J.-Å.G., M.B.); the Texas Emerging Technology Fund (agreement 300-9-1958); the Robert A Welch Foundation (E-0004 to J.-Å.G.); National Cancer Institute at the National Institutes of Health (R01CA172437 to C.W.); National Institute of Environmental Health Sciences (1R21ES020036 to J.-Å.G., M.B.); faculty start-up funding from the University of Houston (to C.W.).
ACknoWLEDGMEnTS
We thank Dr Patrick Balaguer (INSERM, IRCM Montpellier, France) for the kind gift of the HeLN-ERα and HeLN-ERβ cell lines and for discussions on the results, Dr Xiaolian Gao (University of Houston) for generously providing the microar-ray scanner for our use, and Dr Paul Webb (Houston Methodist Research Institute) for valuable comments on the manuscript.
The views expressed in this article reflect the views of the authors and not necessarily of the funders.
REFEREnCES
Alvarez-Baron, C. P., Jonsson, P., Thomas, C., Dryer, S. E., and Williams, C. (2011). The two-pore domain potassium channel KCNK5: Induction by estrogen receptor alpha and role in proliferation of breast cancer cells. Mol. Endocrinol. 25, 1326–1336.
Beronius, A., Rudén, C., Håkansson, H., and Hanberg, A. (2010). Risk to all or none? A comparative analysis of controversies in the health risk assessment of Bisphenol A. Reprod. Toxicol. 29, 132–146.
Betancourt, A. M., Wang, J., Jenkins, S., Mobley, J., Russo, J., and C. A. Lamartiniere (2012). Altered carcinogenesis and proteome in mammary glands of rats after prepubertal exposures to the hormonally active chemicals bisphenol a and genistein. J. Nutr. 142, 1382S–1388S.
Bhatia, J., and Greer, F.; American Academy of Pediatrics Committee on Nutrition. (2008). Use of soy protein-based formulas in infant feeding. Pediatrics 121, 1062–1068.
Brotons, J. A., Olea-Serrano, M. F., Villalobos, M., Pedraza, V., and Olea, N. (1995). Xenoestrogens released from lacquer coatings in food cans. Environ. Health Perspect. 103, 608–612.
Bulayeva, N. N., and Watson, C. S. (2004). Xenoestrogen-induced ERK-1 and ERK-2 activation via multiple membrane-initiated signaling pathways. Environ. Health Perspect. 112, 1481–1487.
Changeux, J. P. (2012). Allostery and the Monod-Wyman-Changeux model after 50 years. Annu. Rev. Biophys. 41, 103–133.
Cimafranca, M. A., Davila, J., Ekman, G. C., Andrews, R. N., Neese, S. L., Peretz, J., Woodling, K. A., Helferich, W. G., Sarkar, J., Flaws, J. A., et al. (2010). Acute and chronic effects of oral genistein administration in neonatal mice. Biol. Reprod. 83, 114–121.
Collaborative group on Hormonal factors in Breast Cancer (2002). Breast can-cer and breastfeeding: Collaborative reanalysis of individual data from 47 epidemiological studies in 30 countries, including 50302 women with breast cancer and 96973 women without the disease. Lancet 360, 187–195.
Cok, I., Donmez, M. K., Uner, M., Demirkaya, E., Henkelmann, B., Shen, H., Kotalik, J., and Schramm, K. W. (2009). Polychlorinated dibenzo-p-dioxins, dibenzofurans and polychlorinated biphenyls levels in human breast milk from different regions of Turkey. Chemosphere 76, 1563–1571.
D’Aloisio, A. A., Baird, D. D., DeRoo, L. A., and Sandler, D. P. (2010). Association of intrauterine and early-life exposures with diagnosis of uter-ine leiomyomata by 35 years of age in the Sister Study. Environ. Health Perspect. 118, 375–381.
Dairkee, S. H., Seok, J., Champion, S., Sayeed, A., Mindrinos, M., Xiao, W., Davis, R. W., and Goodson, W. H. (2008). Bisphenol A induces a profile of tumor aggressiveness in high-risk cells from breast cancer patients. Cancer Res. 68, 2076–2080.
Delfosse, V., Grimaldi, M., Pons, J. L., Boulahtouf, A., le Maire, A., Cavailles, V., Labesse, G., Bourguet, W., and Balaguer, P. (2012). Structural and mech-anistic insights into bisphenols action provide guidelines for risk assessment and discovery of bisphenol A substitutes. Proc. Natl. Acad. Sci. U.S.A. 109, 14930–14935.
Dong, S., Terasaka, S., and Kiyama, R. (2011). Bisphenol A induces a rapid activation of Erk1/2 through GPR30 in human breast cancer cells. Environ. Pollut. 159, 212–218.
Edginton, A. N., and Ritter, L. (2009). Predicting plasma concentrations of bis-phenol A in children younger than 2 years of age after typical feeding sched-ules, using a physiologically based toxicokinetic model. Environ. Health Perspect. 117, 645–652.
Escande, A., Pillon, A., Servant, N., Cravedi, J. P., Larrea, F., Muhn, P., Nicolas, J. C., Cavaillès, V., and Balaguer, P. (2006). Evaluation of ligand selectivity
33
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from

KATCHY ET AL.
using reporter cell lines stably expressing estrogen receptor alpha or beta. Biochem. Pharmacol. 71, 1459–1469.
Gage, M., Wattendorf, D., and Henry, L. R. (2012). Translational advances regarding hereditary breast cancer syndromes. J. Surg. Oncol. 105, 444–451.
Gertz, J., Reddy, T. E., Varley, K. E., Garabedian, M. J., and Myers, R. M. (2012). Genistein and bisphenol A exposure cause estrogen receptor 1 to bind thousands of sites in a cell type-specific manner. Genome Res. 22, 2153–2162.
Helferich, W. G., Andrade, J. E., and Hoagland, M. S. (2008). Phytoestrogens and breast cancer: A complex story. Inflammopharmacology 16, 219–226.
Holbeck, S., Chang, J., Best, A. M., Bookout, A. L., Mangelsdorf, D. J., and Martinez, E. D. (2010). Expression profiling of nuclear receptors in the NCI60 cancer cell panel reveals receptor-drug and receptor-gene interac-tions. Mol. Endocrinol. 24, 1287–1296.
Ikezuki, Y., Tsutsumi, O., Takai, Y., Kamei, Y., and Taketani, Y. (2002). Determination of bisphenol A concentrations in human biological fluids reveals significant early prenatal exposure. Hum. Reprod. 17, 2839–2841.
Jefferson, W. N., Padilla-Banks, E., and Newbold, R. R. (2005). Adverse effects on female development and reproduction in CD-1 mice following neonatal exposure to the phytoestrogen genistein at environmentally relevant doses. Biol. Reprod. 73, 798–806.
Katchy, A., Edvardsson, K., Aydogdu, E., and Williams, C. (2012). Estradiol-activated estrogen receptor α does not regulate mature microRNAs in T47D breast cancer cells. J. Steroid Biochem. Mol. Biol. 128, 145–153.
Kershaw, E. E., and Flier, J. S. (2004). Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 89, 2548–2556.
Krementsov, D. N., Katchy, A., Case, L. K., Carr, F. E., Davis, B., Williams, C., and Teuscher, C. (2013). Studies in experimental autoimmune encepha-lomyelitis do not support developmental bisphenol a exposure as an environ-mental factor in increasing multiple sclerosis risk. Toxicol. Sci. 135, 91–102.
Kuiper, G. G., Lemmen, J. G., Carlsson, B., Corton, J. C., Safe, S. H., van der Saag, P. T., van der Burg, B., and Gustafsson, J. A. (1998). Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology 139, 4252–4263.
Lapensee, E. W., Tuttle, T. R., Fox, S. R., and Ben-Jonathan, N. (2009). Bisphenol A at low nanomolar doses confers chemoresistance in estrogen receptor-alpha-positive and -negative breast cancer cells. Environ. Health Perspect. 117, 175–180.
Lucki, N. C., and Sewer, M. B. (2011). Genistein stimulates MCF-7 breast cancer cell growth by inducing acid ceramidase (ASAH1) gene expression. J. Biol. Chem. 286, 19399–19409.
Maggiolini, M., Vivacqua, A., Fasanella, G., Recchia, A. G., Sisci, D., Pezzi, V., Montanaro, D., Musti, A. M., Picard, D., and Andò, S. (2004). The G protein-coupled receptor GPR30 mediates c-fos up-regulation by 17beta-estradiol and phytoestrogens in breast cancer cells. J. Biol. Chem. 279, 27008–27016.
Markey, C. M., Luque, E. H., Munoz De Toro, M., Sonnenschein, C., and Soto, A. M. (2001). In utero exposure to bisphenol A alters the develop-ment and tissue organization of the mouse mammary gland. Biol. Reprod. 65, 1215–1223.
Messina, M. J., and Wood, C. E. (2008). Soy isoflavones, estrogen therapy, and breast cancer risk: analysis and commentary. Nutr. J. 7, 17.
Miller, L. D., Smeds, J., George, J., Vega, V. B., Vergara, L., Ploner, A., Pawitan, Y., Hall, P., Klaar, S., Liu, E. T., et al. (2005). An expression signature for p53 status in human breast cancer predicts mutation status, transcriptional effects, and patient survival. Proc. Natl. Acad. Sci. U.S.A. 102, 13550–13555.
Newbold, R. R., Banks, E. P., Bullock, B., and Jefferson, W. N. (2001). Uterine adenocarcinoma in mice treated neonatally with genistein. Cancer Res. 61, 4325–4328.
Newbold, R. R., Jefferson, W. N., and Padilla-Banks, E. (2007). Long-term adverse effects of neonatal exposure to bisphenol A on the murine female reproductive tract. Reprod. Toxicol. 24, 253–258.
Ouchi, K., and Watanabe, S. (2002). Measurement of bisphenol A in human urine using liquid chromatography with multi-channel coulometric elec-trochemical detection. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 780, 365–370.
Richter, K., Wirta, V., Dahl, L., Bruce, S., Lundeberg, J., Carlsson, L., and Williams, C. (2006). Global gene expression analyses of hematopoietic stem cell-like cell lines with inducible Lhx2 expression. BMC Genomics 7, 75.
Rider, C. V., and LeBlanc, G. A. (2005). An integrated addition and interaction model for assessing toxicity of chemical mixtures. Toxicol. Sci. 87, 520–528.
Riu, A., Balaguer, P., Perdu, E., Pandelova, M., Piccinelli, R., Gustafsson, J. A., Leclercq, C., Schramm, K. W., Dagnino, S., Debrauwer, L., et al. (2008). Characterisation of bioactive compounds in infant formulas using immo-bilised recombinant estrogen receptor-alpha affinity columns. Food Chem. Toxicol. 46, 3268–3278.
Sasaki, N., Okuda, K., Kato, T., Kakishima, H., Okuma, H., Abe, K., Tachino, H., Tuchida, K., and Kubono, K. (2005). Salivary bisphenol-A levels detected by ELISA after restoration with composite resin. J. Mater. Sci. Mater. Med. 16, 297–300.
Satih, S., Chalabi, N., Rabiau, N., Bosviel, R., Fontana, L., Bignon, Y. J., and Bernard-Gallon, D. J. (2010). Gene expression profiling of breast cancer cell lines in response to soy isoflavones using a pangenomic microarray approach. OMICS 14, 231–238.
Setchell, K. D., Zimmer-Nechemias, L., Cai, J., and Heubi, J. E. (1997). Exposure of infants to phyto-oestrogens from soy-based infant formula. Lancet 350, 23–27.
Setchell, K. D., Zimmer-Nechemias, L., Cai, J., and Heubi, J. E. (1998). Isoflavone content of infant formulas and the metabolic fate of these phy-toestrogens in early life. Am. J. Clin. Nutr. 68, 1453S–1461S.
Singh, S., and Li, S. S. (2012). Epigenetic effects of environmental chemicals bisphenol a and phthalates. Int. J. Mol. Sci. 13, 10143–10153.
Singleton, D. W., Feng, Y., Chen, Y., Busch, S. J., Lee, A. V., Puga, A., and Khan, S. A. (2004). Bisphenol-A and estradiol exert novel gene regulation in human MCF-7 derived breast cancer cells. Mol. Cell. Endocrinol. 221, 47–55.
Singleton, D. W., Feng, Y., Yang, J., Puga, A., Lee, A. V., and Khan, S. A. (2006). Gene expression profiling reveals novel regulation by bisphenol-A in estrogen receptor-alpha-positive human cells. Environ. Res. 100, 86–92.
Smyth, G. K. (2004). Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 3, Article3.
Stratton, M. R., and Rahman, N. (2008). The emerging landscape of breast cancer susceptibility. Nat. Genet. 40, 17–22.
Turner, J. V., Agatonovic-Kustrin, S., and Glass, B. D. (2007). Molecular aspects of phytoestrogen selective binding at estrogen receptors. J. Pharm. Sci. 96, 1879–1885.
Vandenberg, L. N., Hauser, R., Marcus, M., Olea, N., and Welshons, W. V. (2007). Human exposure to bisphenol A (BPA). Reprod. Toxicol. 24, 139–177.
vom Saal, F. S., and Hughes, C. (2005). An extensive new literature concerning low-dose effects of bisphenol A shows the need for a new risk assessment. Environ. Health Perspect. 113, 926–933.
Wardell, S. E., Kazmin, D., and McDonnell, D. P. (2012). Research resource: Transcriptional profiling in a cellular model of breast cancer reveals func-tional and mechanistic differences between clinically relevant SERM and between SERM/estrogen complexes. Mol. Endocrinol. 26, 1235–1248.
Weng, Y. I., Hsu, P. Y., Liyanarachchi, S., Liu, J., Deatherage, D. E., Huang, Y. W., Zuo, T., Rodriguez, B., Lin, C. H., Cheng, A. L., et al. (2010). Epigenetic influences of low-dose bisphenol A in primary human breast epithelial cells. Toxicol. Appl. Pharmacol. 248, 111–121.
Williams, C., Edvardsson, K., Lewandowski, S. A., Ström, A., and Gustafsson, J. A. (2008). A genome-wide study of the repressive effects of estrogen
34
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from

COEXPOSURE TO PHYTOESTROGENS AND BISPHENOL A
receptor beta on estrogen receptor alpha signaling in breast cancer cells. Oncogene 27, 1019–1032.
Winter, J. S., Hughes, I. A., Reyes, F. I., and Faiman, C. (1976). Pituitary-gonadal relations in infancy: 2. Patterns of serum gonadal steroid concentra-tions in man from birth to two years of age. J. Clin. Endocrinol. Metab. 42, 679–686.
Wolstenholme, J. T., Edwards, M., Shetty, S. R., Gatewood, J. D., Taylor, J. A., Rissman, E. F., and Connelly, J. J. (2012). Gestational exposure to bisphe-nol a produces transgenerational changes in behaviors and gene expression. Endocrinology 153, 3828–3838.
Wu, A. H., Yu, M. C., Tseng, C. C., and Pike, M. C. (2008). Epidemiology of soy exposures and breast cancer risk. Br. J. Cancer 98, 9–14.
Yager, J. D., and Davidson, N. E. (2006). Estrogen carcinogenesis in breast cancer. N. Engl. J. Med. 354, 270–282.
Yuan, C., Lin, J. Z., Sieglaff, D. H., Ayers, S. D., Denoto-Reynolds, F., Baxter, J. D., and Webb, P. (2012). Identical gene regulation patterns of T3 and selective thyroid hormone receptor modulator GC-1. Endocrinology 153, 501–511.
Zhang, C., Ho, S. C., Lin, F., Cheng, S., Fu, J., and Chen, Y. (2010). Soy prod-uct and isoflavone intake and breast cancer risk defined by hormone receptor status. Cancer Sci. 101, 501–507.
Zhang, M., Yao, C., Guo, Z., Zou, J., Zhang, L., Xiao, H., Wang, D., Yang, D., Gong, X., Zhu, J., et al. (2008). Apparently low reproducibility of true differential expression discoveries in microarray studies. Bioinformatics 24, 2057–2063.
Zhang, M., Zhang, L., Zou, J., Yao, C., Xiao, H., Liu, Q., Wang, J., Wang, D., Wang, C., and Guo, Z. (2009). Evaluating reproducibility of differen-tial expression discoveries in microarray studies by considering correlated molecular changes. Bioinformatics 25, 1662–1668.
35
by guest on September 13, 2015
http://toxsci.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















