
Development of a Marker of Estrogenic Exposure in Human Serum
8
CLIN. CHEM. 41/12, 1888-1895 (1995) #{149} European Beckman Conference 1888 CLINICAL CHEMISTRY, Vol. 41, No. 12, 1995 Development of a Marker of Estrogenic Exposure in Human Serum Carlos Sonnenschein,”2 Ana M. Soto,”2’5 Mariana F. Fernandez,’ Nicolas Olea,3 M. Fatima Olea-Serrano,4 and M. Dolores Ruiz-Lopez4 The deleterious, disruptive effects of estrogen mimics on the endocrine system were discovered after the com- pounds were released into the environment. Their chem- ical structure does not obviously resemble that of steroid hormones; hence, their estrogenic effects were totally unexpected. In addition to occupational exposures, en- vironmental estrogens may have played a role in de- creasing the quantity and quality of human semen during the last 50 years and in increasing the incidences of testicular cancer and cryptorchidism in men and breast cancer in women and men in industrialized countries. Testing the environmental estrogen hypothesis will re- quire developing appropriate biomarkers of exposure and measuring these biomarkers at developmental points where exposure is critical. We report the ongoing development of a method to extractand separate xe- noestrogens from ovarian estrogens with human serum as a source, followed by determination of xenoestrogen concentration by a bioassay. We also critically assess bioassays currently available to measure the cumulative effect of xenoestrogens, e.g., (a) the E-SCREEN assay, which measures the proliferative effect of estrogens on their target cells, and (b) the induction by estrogens of specific gene products, such as progesterone receptor and pS2. Indexing Terms: xenoestrogens/endocrine disruptors/cancer/en- vironmental estrogens For the last 40 years, substantial evidence has sur- faced on the hormone-like effects of environmental chemicals in fish, wildlife, and humans (1). These unintended effects were discovered long after these chemicals were released into the environment. Estro- gen mimics are a class of endocrine disruptors. The chemical structure of xenoestrogens does not obviously resemble that of steroid hormones; hence, their estro- genic effects were unexpected. In humans, occupa- tional exposures revealed that certain pesticides were endocrine disruptors. In 1949, Singer (2) reported that aviation crop dusters handling dichlorodiphenyltri- ‘Department of Anatomy and Cellular Biology, Tufts Univer- sity School of Medicine, Boston, MA 02111. 2 These authors contributed equally to this work. Department of Radiology, School of Medicine, University of Granada, Spain. Department of Bromatology, School of Pharmacy, University of Granada, Spain. Author for correspondence. Fax 617-636-6536; e-mail [email protected]. Received June 23, 1995; accepted August 24, 1995. chloroethane (DDT) had reduced sperm counts.6 Two decades later,workers at a plantproducing the insec- ticide Kepone were reported to have lost their libido, become impotent, and had low sperm counts (3). These effects were similar to those observed after administra- tion of natural estrogens to men and were attributed to estrogen-mediated suppression of gonadotropins, which in turn yielded decreased production of testos- terone and reduced spermatogenesis. In occupational exposure cases, a single agent is usuallyidentifiedastheculprit,whereaswildlifeshow- ing signs of reproductive damage are typically exposed to a combination ofendocrine disruptors.In reviewing wildlife studies, the abnormal development of the re- productive system in gull embryos exposed in ovo to DDT and other pesticides suggested that these chemi- cals were the causal agents; this was verified in labo- ratory experiments with various bird species (4). Sim- ilarly, the decreased reproductive success of alligators and turtles in Lake Apopka, FL, could be linked to a spill of Kelthane, a pesticide formulation containing dichlorodiphenyldichloroethylene (DDE) (5). More of- ten, decreased reproductive success and signs of femi- nization and demasculinization in wildlife could not be ascribed to a single agent; in these cases, chemical analysis of the specimens detected multiple com- pounds, some of them known to have hormone-like activity (1). In the last 5 years, several environmental chemicals have been added to the list of xenoestrogens, including the pesticides toxaphene, dieldrin, and endosulfan; anti- oxidants such as dibutylhydroxyamsole and alkylphe- nols; and plasticizers such as benzylbutylphthalate and bisphenol A (6). The relevance of these newly discovered estrogens to wildlife and human health is now starting to emerge. Feminized male fish were found near sewage outlets in several rivers in the UK a mixture of chemicals including alkyl phenols, resulting from degradation of detergents during sewage treatment, seem to have caused this endocrine disruption (7).Brotons et al.(8) reported that bisphenol A, released from tin cans lined with lacquers such as polycarbonate, contaminates the contents of canned foods. It has been hypothesized that environmental estro- gens may be responsible for lowering the quantity and 6 Nonstandard abbreviations: DDT, dichlorodiphenyltrichloro- ethane; DDE, dichlorodiphenyldichloroethylene; PCBs, polychlo- mated biphenyls; PgR, progesterone receptor; E2, estradiol-1713; CD, charcoal-dextran; HCB, hexachlorobiphenyl; TCB, tetra- chlorobiphenyl; DCB, dichlorobiphenyl; FBS, fetal bovine serum; and FE, proliferative effect.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Development of a Marker of Estrogenic Exposure in Human Serum

CLIN. CHEM. 41/12, 1888-1895 (1995) #{149}European Beckman Conference
1888 CLINICAL CHEMISTRY, Vol. 41, No. 12, 1995
Development of a Marker of Estrogenic Exposure in Human Serum
Carlos Sonnenschein,”2 Ana M. Soto,”2’5 Mariana F. Fernandez,’ Nicolas Olea,3 M. Fatima Olea-Serrano,4 andM. Dolores Ruiz-Lopez4
The deleterious, disruptive effects of estrogen mimics onthe endocrine system were discovered after the com-pounds were released into the environment. Their chem-ical structure does not obviously resemble that of steroidhormones; hence, their estrogenic effects were totallyunexpected. In addition to occupational exposures, en-vironmental estrogens may have played a role in de-creasing the quantity and quality of human semen duringthe last 50 years and in increasing the incidences oftesticular cancer and cryptorchidism in men and breastcancer in women and men in industrialized countries.Testing the environmental estrogen hypothesis will re-quire developing appropriate biomarkers of exposureand measuring these biomarkers at developmentalpoints where exposure is critical. We report the ongoingdevelopment of a method to extractand separate xe-noestrogens from ovarian estrogens with human serumas a source, followed by determination of xenoestrogenconcentration by a bioassay. We also critically assessbioassays currently available to measure the cumulativeeffect of xenoestrogens, e.g., (a) the E-SCREEN assay,which measures the proliferative effect of estrogens ontheir target cells, and (b) the induction by estrogens ofspecific gene products, such as progesterone receptorand pS2.
Indexing Terms: xenoestrogens/endocrine disruptors/cancer/en-vironmental estrogens
For the last 40 years, substantial evidence has sur-faced on the hormone-like effects of environmentalchemicals in fish, wildlife, and humans (1). Theseunintended effects were discovered long after thesechemicals were released into the environment. Estro-gen mimics are a class of endocrine disruptors. Thechemical structure of xenoestrogens does not obviouslyresemble that of steroid hormones; hence, their estro-genic effects were unexpected. In humans, occupa-tional exposures revealed that certain pesticides wereendocrine disruptors. In 1949, Singer (2) reported thataviation crop dusters handling dichlorodiphenyltri-
‘Department of Anatomy and Cellular Biology, Tufts Univer-sity School of Medicine, Boston, MA 02111.
2 These authors contributed equally to this work.
Department of Radiology, School of Medicine, University ofGranada, Spain.
Department of Bromatology, School of Pharmacy, Universityof Granada, Spain.
Author for correspondence. Fax 617-636-6536; [email protected].
Received June 23, 1995; accepted August 24, 1995.
chloroethane (DDT) had reduced sperm counts.6 Twodecades later,workers at a plant producing the insec-ticide Kepone were reported to have lost their libido,become impotent, and had low sperm counts (3). Theseeffects were similar to those observed after administra-tion of natural estrogens to men and were attributedto estrogen-mediated suppression of gonadotropins,which in turn yielded decreased production of testos-terone and reduced spermatogenesis.
In occupational exposure cases, a single agent isusuallyidentifiedas the culprit,whereas wildlifeshow-ing signs of reproductive damage are typically exposedto a combination ofendocrine disruptors.In reviewingwildlife studies, the abnormal development of the re-productive system in gull embryos exposed in ovo toDDT and other pesticides suggested that these chemi-cals were the causal agents; this was verified in labo-ratory experiments with various bird species (4). Sim-ilarly, the decreased reproductive success of alligatorsand turtles in Lake Apopka, FL, could be linked to aspill of Kelthane, a pesticide formulation containingdichlorodiphenyldichloroethylene (DDE) (5). More of-ten, decreased reproductive success and signs of femi-nization and demasculinization in wildlife could not beascribed to a single agent; in these cases, chemicalanalysis of the specimens detected multiple com-pounds, some of them known to have hormone-likeactivity (1).
In the last 5 years, several environmental chemicalshave been added to the list of xenoestrogens, includingthe pesticides toxaphene, dieldrin, and endosulfan; anti-oxidants such as dibutylhydroxyamsole and alkylphe-nols; and plasticizers such as benzylbutylphthalate andbisphenol A (6). The relevance of these newly discoveredestrogens to wildlife and human health is now starting toemerge. Feminized male fish were found near sewageoutlets in several rivers in the UK a mixture of chemicalsincluding alkyl phenols, resulting from degradation ofdetergents during sewage treatment, seem to havecaused this endocrine disruption (7).Brotons et al.(8)reported that bisphenol A, released from tin cans linedwith lacquers such as polycarbonate, contaminates thecontents of canned foods.
It has been hypothesized that environmental estro-gens may be responsible for lowering the quantity and
6 Nonstandard abbreviations: DDT, dichlorodiphenyltrichloro-
ethane; DDE, dichlorodiphenyldichloroethylene; PCBs, polychlo-mated biphenyls; PgR, progesterone receptor; E2, estradiol-1713;CD, charcoal-dextran; HCB, hexachlorobiphenyl; TCB, tetra-chlorobiphenyl; DCB, dichlorobiphenyl; FBS, fetal bovine serum;and FE, proliferative effect.

CLINICAL CHEMISTRY, Vol. 41, No. 12, 1995 1889
quality of human semen during the last 50 years andincreasing the incidences of testicular cancer and crypt-orchidism in men and breast cancer in women and menin industrialized countries (9-11). The main risk factorfor developing breast cancer is lifelong exposure toestrogens (12). Studies in animal models point to theexistence of critical periods during development inwhich mammary tissue is most susceptible to chemicalcarcinogenesis (13). Epidemiological studies suggestthat increased plasma concentrations of “bioavailable”ovarian estrogens (not bound to sex-hormone-bindingglobulin) in postmenopausal women correlate posi-tively with breast cancer (14). Among the estrogenicxenobiotics, polychlorinated biphenyls [PCBs; e.g., di-chiorobiphenyls (DCBs), tetrachlorobiphenyls (TCB),and hexachlorobiphenyls (HCBs)] and DDT were con-sidered suitable markers of exposure for breast cancerbecause they were released massively into the environ-ment -50 years ago and are persistent; their presencein serum may represent cumulative lifelong exposure.In an epidemiological study, Wolff et al. (15) reported apositive correlation between plasma concentrations ofDDE, a DDT metabolite, and breast cancer incidence.Krieger et al. (16), however, did not find such a corre-lation. To clarify this issue, larger prospective epidemi-ological studies are being conducted. However, it wouldbe impractical to test only for DDT metabolites andPCBs, because not all the PCB congeners are estro-genic; many other environmental estrogens such as theones already mentioned may also play a role.Thenewly identified estrogens may be less persistent thanPCBs and DDT metabolites, but are widely used; onemay infer that exposure occurs steadily because thoseestrogens are present in foods (17). Analytical chemicaltesting for each of these chemicals in biological samplesis costly, requires large specimens, and may be incom-plete, given that not all xenoestrogens may have beenidentified yet. Alternatively, evidence that xenoestro-gens act cumulatively to induce cell proliferation couldbe used to develop a marker of exposure (17).
We introduce here a procedure to extract and sepa-rate xenoestrogens from ovarian estrogens in humanserum, followed by determination of xenoestrogen con-centration with a bioassay. We discuss the E-SCREEN,a bioassay suitable for measuring the cumulative effectof xenoestrogens; this assay measures the proliferativeeffect of estrogens on their target cells, a featureconsidered the hallmark of estrogen action (18). Fi-nally, we assess the use of other endpoints, namely, theinduction by estrogens of specific gene products, suchas progesterone receptor (PgR) and p52 (19).
Materials and Methods
Chemicals
Estradiol-1713 (E2) was from Calbiochem (Richmond,CA). Radiolabeled E2 was purchased from New En-gland Nuclear Research Products (Boston,MA). Other
steroids were purchased from Steraloids (Wilton, NH).Toxaphene, endosulfan, nonylphenol, and octylphenol
were from Chem Services (West Chester, PA). o,p’-DDT, p,p’-DDT, p,p’-DDE, PCB congeners, hydroxy-lated biphenyls, methoxychlor, dieldrin, phthalates,and antioxidants were from Ultra Scientific (NorthKingstown, RI). Bisphenol A dimethacrylate and bis-phenol A were from Aldrich Chemical, Milwaukee, WI.All these chemicals were dissolved in ethanol or di-methyl sulfoxide before being tested in the variousassay systems. The final solvent concentration in cul-ture medium did not exceed 1 mLIL and did not affectcell yields. All organic solvents were HPLC grade andwere purchased from Fisher Scientific (Pittsburgh,PA), except for ethanol and ethyl ether (from SigmaChemical Co., St. Louis, MO), and petroleum ether(from Aldrich). All other chemicals were of the highestquality available from commercial sources.
Plasma-Derived and Blood-Derived Human Serum
Plasma-derived human serum was prepared fromoutdated plasma supplied by the New England MedicalCenter Blood Bank. Calcium chloride was added to afinal concentration of 30 mmolJL to facilitate clot for-mation. Blood was drawn from healthy adult volun-teers according to a protocol approved by the HumanInvestigation Review Committee at Tufts-New En-gland Medical Center and was allowed to clot in glasscentrifuge tubes for 2-4 h to produce serum. Plasma-and blood-derived sera were clarified by centrifugation(2000g for 10 mm), stripped with charcoal-dextran(CD), and stored in glass tubes at -20 #{176}Cuntil use.
Removal of Sex Steroids by CD Treatment of Serum
Charcoal (Norit A, acid-washed; Sigma) was washedtwice with cold sterile water immediately before use. A50 gIL charcoal-5 gIL dextran T70 (Pharmacia-LKB,Uppsala, Sweden) suspension was prepared. CD sus-pension aliquots of a volume similar to that of serumaliquots to be processed were centrifuged at lOOg for 10mm. Supernates were aspirated and serum aliquotswere mixed with the charcoal pellets. This charcoal-serum mixture was maintained in suspension by roll-ing at 4 cycles/mm at 37 #{176}Cfor 1 h. This suspensionwas centrifuged at 2000g for 20 mm. The supernatewas then filtered through a 0.45-I.Lm (pore size) Nal-gene filter. More than 99% of serum sex steroids wasremoved by this treatment, as measured by removal of[3H1E2 (20); E2 concentrations after CD treatment were<0.01 ng/L as measured by RIA. CD-treated sera werestoredat -20 #{176}Cuntilneeded. Samples kept for1 yearin the freezer remained inhibitory to the proliferationof human estrogen-sensitive breast tumor MCF7 cells;plasma- and blood-derivedsera were equally effective.
ExtractionofXenoestrogensfrom Plasma-DerivedSerum
and Cleanup
PCBs and pesticides were extracted as described byBurse et al. (21), with modifications to extract Xe-noestrogens with polarities intermediate betweenthose of PCBs and ovarian estrogens. To assess therecovery of each xenoestrogen from serum, we added

1890 CLINICAL CHEMISTRY, Vol. 41, No. 12, 1995
known amounts of standard xenoestrogens to 4-mLaliquots of CD-treated serum, equilibrated the solutionfor 2 h at room temperature, and extracted the Xe-noestrogens as described below. CD-treated serum wasused as a blank because extracts from this preparationdo not absorb at 280 nm and therefore do not interferewith the quantification of added xenoestrogens.
Sample extraction. Methanol (2 mL) was added to 4mL of serum; after vortex-mixing, 5 mL of hexane:ethylether (inequal volumes) was added, and the mixturewas first agitated on a rotary mixer for 15 mm and thencentrifuged at 2000g for 5 mm. The organic phase wascollected, and the aqueous phase was extracted twicemore. The organic phases were pooled, concentrated to1 mL under a nitrogen stream, and then pipetted intoa normal-phase silica Sep-Pak cartridge that had beenwashed with 4 mL of petroleum ether. The adsorbedxenoestrogens were eluted serially by manual perfu-sion by means of a glass syringe, with 8 mL of 60 mLILethyl ether in petroleum ether (Fl), 8 mL of 150 milLethyl ether in petroleum ether (F2), and 8 mL of 1000mIlL ethanol (F3). Fl was concentrated to 1 mL, andF2 and F3 were dried down under a gentle stream ofnitrogen.
Acid cleanup of organic phase before HPLC. Concen-trated H2S04 (0.5 mL) was added to Fl (concentratedto 1 mL). The sample was vortex-mixed briefly andcentrifuged for 5 mm at 2000g. The organic phase wascollected, and the aqueous phase was extracted twicemore with 1 mL of hexane. The organic phases werepooled and concentrated to 1 mL under nitrogen.
Monitoring extraction of[3H]E2. [3H]E2 was purifiedimmediately before use by Sephadex LH-20 chromatog-raphy. [3H]E2 (100 000 cpm) was added to 4 mL ofCD-treated serum; after equilibration for 2 h at roomtemperature, the supplemented serum was extractedas described above, and each fraction was evaporatedand counted in a liquid scintillation counter.
HPLC analysis. Separation was performed by themethod of Medina and Sherman (22), modified asdescribed below, in a Kratos chromatograph (solventdelivery system: Spectroflow 400; gradient controllerand absorbance detector:Spectroflow 783; injector!mixer: Spectroflow 491; all from ABI Analytical, SantaClara, CA). Extracts were dried down and resuspendedin hexane, and 500-.tL aliquots were injected into a 4 X
220 mm Spheri 5 silica column (Brownlee Labs, ABI)equilibrated with 1000 mLfL n-hexane. Xenoestrogenswere eluted by a gradient of two mobile phases: n-hexane (phase A), and n-hexane:methanol:2-propanol(40:45:15, by vol) (phase B) at a flow rate of 1.5 mLlminand a pressure of 5 x 106 Pa. The gradient wasdeveloped as follows: after injection, 1000 mL/L phaseA flowed for 2 mm; then the concentration of phase Bincreased from 0 to 400 milL in 15 mm and from 400to 1000 milL in the next 10 mm. The elution profilewas monitored at 280 nm with a Kratos Spectroflow783 detector.
Quantification. Calibration curves produced by in-jecting 0-500 nmol of analytes at 5-7 doses (in dupli-
cates) were obtained for nonylphenol; o,p’-DDT; p,p’-DDT; methoxychlor; 2,2’ ,3,3 ‘ ,6,6’ -HCB; 2,3,4,5-TCB;4-hydroxy-2’ ,5 ‘-DCB; bisphenol A methacrylate; andbenzylbutylphthalate. The correlation coefficient foreach curve was 0.98-0.99.
Bioassays for Measuring Estrogenicity
E-SCREEN test. Theoretical underpinnings of theE-SCREEN assay have been published elsewhere(20,23). Cloned MCF7 cells were trypsinized andplated into 12-well plates (Costar, Cambridge, MA) atinitial concentrations of 20 000 cells per well (24, 25).
Cells were allowed to attach for 24 h; then the seedingmedium [50 mLIL fetal bovine serum (FBS) in DMEM]was replaced with the experimental medium (50 milLCD-treated serum supplemented to phenol red-freeDMEM). A range of concentrations of the test com-pounds was added to this medium. The bioassay wasterminated on the 6th day (late exponential phase) byremoving the media from the wells, adding a cell-lysingsolution [100 mLIL ethylhexadecyldimethylammoniumbromide (Eastman Kodak, Rochester, NY) in 5 milLTriton X-l00, 2 mmol/L MgC12, 15 mmol/L NaCl, and 5mmol/L phosphate buffer,pH 7.4],and counting thenuclei in a Model ZM Coulter Counter (Coulter Elec-tronics, Hialeah, FL).
The best estimate of the proliferative behavior of acell population is tD or doubling time (the time intervalduring which an exponentially growing culture doublesits cell number). Determining tD requires measuringcell yields at several time intervals during the expo-nential proliferation phase. A less cumbersome alter-native is to compare the cell yield achieved by similarcell inocula harvested simultaneously during the lateexponential phase of proliferation. The proliferativeeffect (PE) is measured as the ratio of the highest cellyield obtained with the test chemical to the hormone-free control.Under these conditions,cellyield repre-sents a reliable estimate of the relative rate of prolif-eration achieved by similar inocula exposed to differentproliferation regulators. In our experimental design,MCF7 cell yields were measured 6 days after t0; how-ever,significantdifferencesbetween controland estro-gen-treated cultures are apparent after 4 days (26).
PgR assay. MCF7 cells were seeded in 25-cm2 flasksin 50 mIlL FBS-supplemented DMEM. After 24 h, themedium was changed to 50 milL CD-treated humanserum, and the chemicals to be tested were added.Control cultures were treated with vehicle. After 72 hof exposure to the test xenoestrogens, the medium wasaspirated, the cell layer was rinsed with phosphate-buffered saline, and cells were frozen in liquid N2. Forextracting receptor molecules, cells were incubatedwith 1 mL of extraction buffer (0.5 mol!L KC1, 10mmol/L potassium phosphate, 1.5 mmolfL EDTA, and1 mmoIIL monothioglycerol, pH 7.4) at 4 #{176}Cfor 30 mm(27). After centrifugation to pellet the cell debris,receptor concentrations were measured in l00-p.L ex-tract aliquots by enzyme immunoassay with the Abbott
Retention time, mm
2.07
2.22
2.40
3.18
3.40
3.70
3.89
3.95
4.88
5.34
6.20
8.85
8.89
9.05
12.62
13.64
14.00
14.61
16.38
16.73
17.89
CLINICAL CHEMISTRY, Vol. 41, No. 12, 1995 1891
PgR kit (Abbott Diagnostics,Chicago, IL) according tothe manufacturer’s instructions.
pS2 assay. Culture media were harvested after 144h of exposure to the test chemicals and centrifuged toeliminate floating and detached cells; samples werekept frozen at -80 #{176}Cuntil immunoradiometricallyassayed according to the manufacturer’s protocol(ELSA-PS2; CIS Bio International, Gif-sur-Yvette,France).
Results
Extraction from Serum and Cleanup
The objective of this fractionation protocol was toaccomplish quantitative extraction from serum whileseparating xenoestrogens from ovarian estrogens.[3H]E2-supplemented CD-treated serum was extractedand chromatographed through a Sep-Pak normal-phase cartridge; the binding to and recovery from thematrix was affected by the perfusion rate. Recoverywas optimized at a mean flow rate of 7.5 mL/min (Table1); under these conditions, most of the [3H]E2 waseluted in the F3 phase.
As expected from published data, PCBs and DDTwere eluted preferentially in Fl and F2 (21). Nonylphe-nol and benzylbutylphthalate were extracted from thesupplemented CD-treated serum and were mainlyeluted in Fl and F2 (Table 2). These results indicatedthat the first step, elution with 60 mLIL ethyl ether inpetroleum ether (Fl), could be skipped; that is, xe-noestrogens may be eluted quantitatively from theSep-Pak cartridge in a single step with 150 milL ethylether in petroleum ether, the solvent originally used toelute F2 (Fl + 2) (Table 2). The recovery of xenoestro-gens from CD-treated serum was equally efficient fromsamples supplemented with either a single xenoestro-gen or a mixture of four (Table 3). Most estradiol elutedin F3, and the bulk of xenoestrogens eluted in Fl +2.
Table 1.Recovery (%) of 3H counts from CD plasma-derived serum supplemented with (3H]E2.
AqueousMean flow rate Fl F2 F3 phase15 mL/min 0.45-0.50 1.90-2.20 63.0-65.0 7.0-8.0
7.5 mL/min 0.25-0.31 0.32-0.36 80.2-88.4 5.3-7.1
CD serum aliquots (4 mL) were supplemented with 100 000 cpm of (3H]E2;aliquots were extracted as described in Materials and Methods after 2 h ofequilibration at room temperature. Data represent the range of duplicateexperiments.
Table 2.Recovery (%) of supplemented calibratorsfrom CD serum.
Chemical Fl P2 Fl + 2
2,2’,3,3’,6,6’-HCB 65 25 85
o,p’-DDT 62 27 84
Nonylphenol 61 17 78
4-Hydroxy-2’,5’-DCB 70 24 92
CD serum aliquots (4 mL) were supplemented with 125 nmol of eachchemical. Recovery of the chemicals through the original two-step elutionmethod (Fl and F2) is compared with that obtained in a single step elution (Fl+ 2).
Table 3.Comparison of the recovery of calibratorsfrom CD serum supplemented with a singlexenoestrogen or a combination of four.
Amount % recoveryadded,
Chemical A B
Nonylphenol 26.875 88.5 ± 2.1 89.2 ± 12
4-Hydroxy-2’,5’-DCB 29.9 69.5 ± 2.0 80.7 ± 5.3
Bisphenol A dimethacrylate 45.5 87.8 ± 14 95.4 ± 3.1
Benzylbutylphthalate 195.25 79.8 ± 5.3 90.8 ± 2.9
A denotes the recovery of each of the indicated chemicals when serum wassupplemented with a single chemical; B denotes the recovery of eachchemical when serum was supplemented with a mixture of the four chemi-cals. Each result represents the mean ± SD of at least four experiments.
This suggests that an almost complete separation couldbe accomplished in a single step.
Separation of Xenoestrogens, Phytoestrogens, andOvarian Estrogens by HPLC
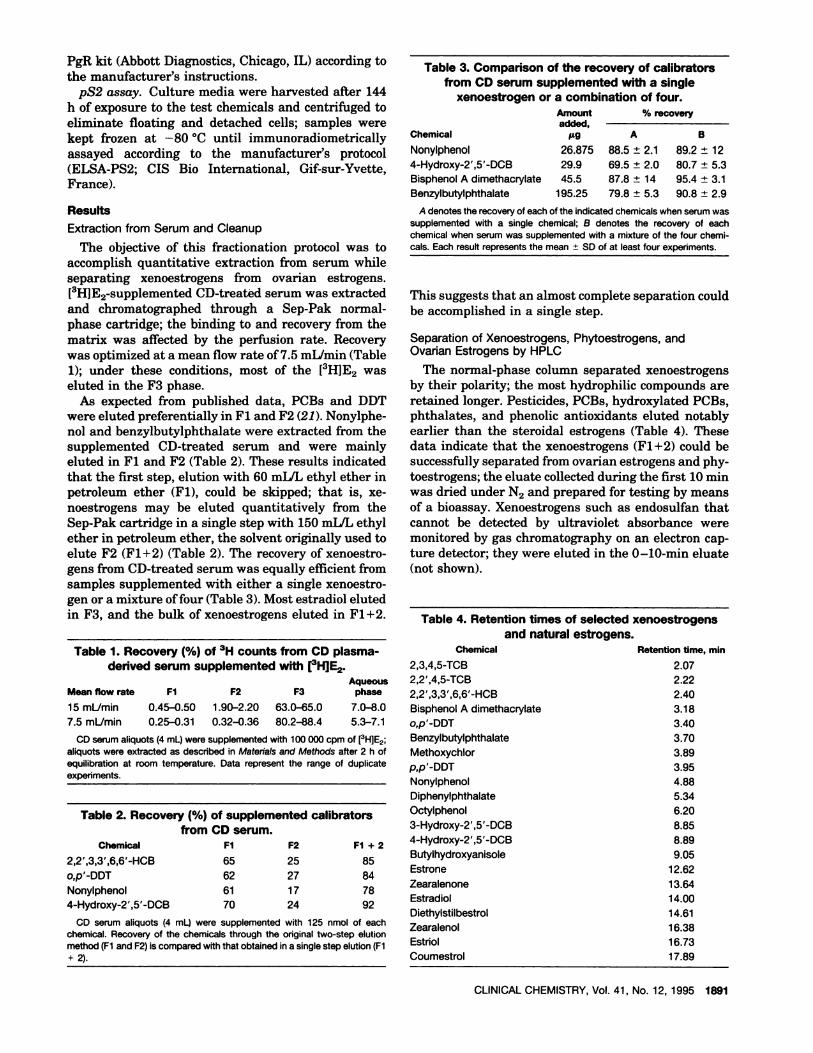
The normal-phase column separated xenoestrogensby their polarity; the most hydrophilic compounds areretained longer. Pesticides, PCBs, hydroxylated PCBs,phthalates, and phenolic antioxidants eluted notablyearlierthan the steroidalestrogens (Table 4).Thesedata indicate that the xenoestrogens (Fl+2) could besuccessfully separated from ovarian estrogens and phy-toestrogens; the eluate collected during the first 10 mmwas dried under N2 and prepared for testing by meansof a bioassay. Xenoestrogens such as endosulfan thatcannot be detected by ultraviolet absorbance weremonitored by gas chromatography on an electron cap-ture detector; they were eluted in the 0-10-mm eluate(not shown).
Table 4.Retentiontimes of selectedxenoestrogensand naturalestrogens.
Chemical
2,3,4,5-TCB
2,2’,4,5-TCB
2,2’,3,3’,6,6’-HCB
Bisphenol A dimethacrylate
o,p’-DDTBenzylbutylphthalateMethoxychiorp,p’-DDTNonylphenolDiphenylphthalateOctylphenol
3-Hydroxy-2’,5’-DCB4-Hydroxy-2’,S’-DCBButylhydroxyanisole
Estrone
ZearalenoneEstradiol
DiethylstilbestrolZearalenol
Estriol
Coumestrol
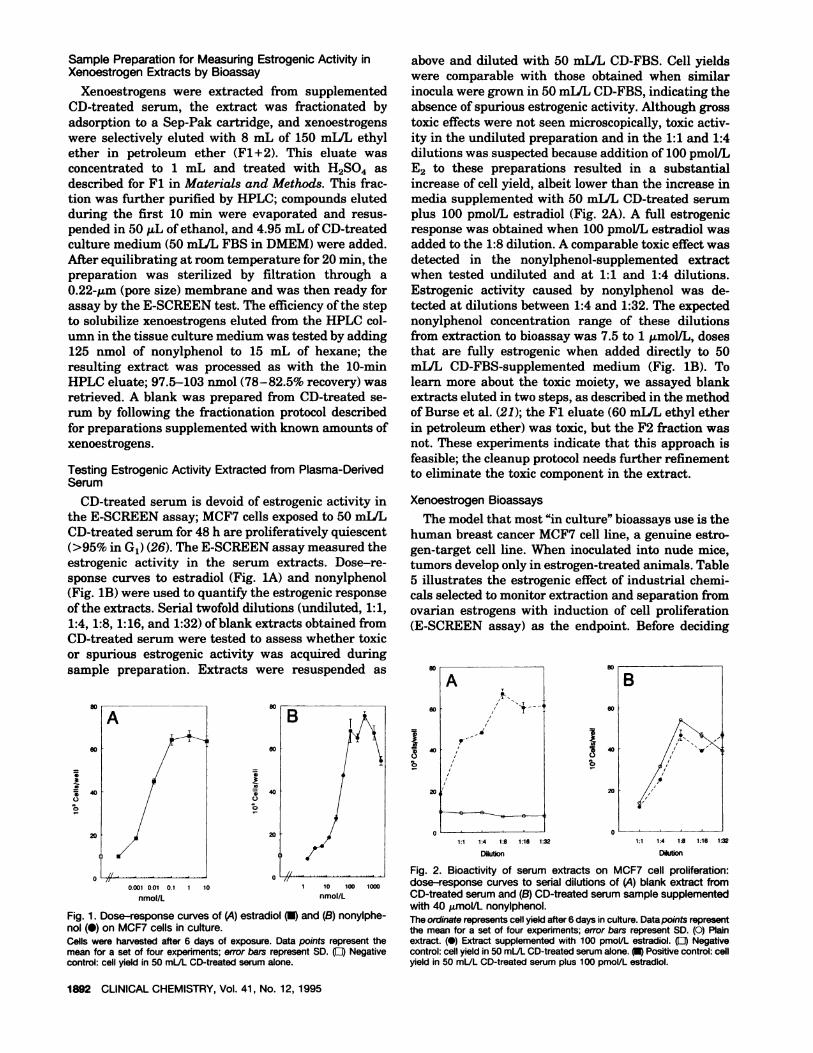
A80
60
40
20
A
U
B
/p
60
40
80I
60
/ -5 400/
40 /
20 0 0II 1:4 1:8 116 1:32 1,1 1.4 1:8 1:16 1:32
Dilution Dilution
Fig. 2. Bioactivity of serum extracts on MCF7 cell proliferation:0 L . -. .J 0
0001 001 01 1 10 10 100 dose-response curves to serial dilutions of (A) blank extract fromnmol/L nmol/L CD-treated serum and (B) CD-treated serum sample supplemented
with 40 imol/L nonyiphenol.Fig. 1. Dose-response curves of (A) estradiol and (B) nonylphe- The ordinate representscell yield after 6 days in culture. Data points representnol (#{149})on MCF7 cells in culture. the mean for a set of four experiments; error bars represent SD. (0) PlainCells were harvested after 6 days of exposure. Data points represent the extract. (#{149})Extract supplemented with 100 pmol/L estradiol. (LI) Negativemean for a set of four experiments; error bars represent SD. (LI) Negative control: cell yield in 50 mLJLCD-treatedserumalone. Positive control: cellcontrol: cell yield in 50 mL/L CD-treated serum alone, yield in 50 mLJL CD-treated serum plus 100 pmoi/L estradiol.
20/
1892 CLINICALCHEMISTRY, Vol. 41, No. 12, 1995
Sample Preparation for Measuring Estrogenic Activity in
Xenoestrogen Extracts by Bioassay
Xenoestrogens were extracted from supplementedCD-treated serum, the extract was fractionated byadsorption to a Sep-Pak cartridge, and xenoestrogenswere selectively eluted with 8 mL of 150 mLIL ethylether in petroleum ether (Fl+2). This eluate wasconcentrated to 1 mL and treated with H2S04 asdescribed for Fl in Materials and Methods. This frac-tion was further purified by HPLC; compounds elutedduring the first 10 mm were evaporated and resus-pended in 50 j.t.L of ethanol, and 4.95 mL of CD-treatedculture medium (50 mLIL FBS in DMEM) were added.After equilibrating at room temperature for 20 mm, thepreparation was sterilized by filtration through a0.22-.tm (pore size) membrane and was then ready forassay by the E-SCREEN test. The efficiency of the stepto solubilize xenoestrogens eluted from the HPLC col-umn in the tissue culture medium was tested by adding125 nmol of nonylphenol to 15 mL of hexane; theresulting extract was processed as with the 10-mmHPLC eluate; 97.5-103 nmol (78-82.5% recovery) wasretrieved. A blank was prepared from CD-treated se-rum by following the fractionation protocol describedfor preparations supplemented with known amounts ofxenoestrogens.
Testing Estrogenic Activity Extracted from Plasma-Derived
Serum
CD-treated serum is devoid of estrogenic activity inthe E-SCREEN assay; MCF7 cells exposed to 50 mIlL
CD-treated serum for 48 h are proliferatively quiescent(>95% in G1) (26). The E-SCREEN assay measured theestrogenic activity in the serum extracts. Dose-re-sponse curves to estradiol (Fig. lA) and nonylphenol(Fig. lB) were used to quantify the estrogenic responseof the extracts. Serial twofold dilutions (undiluted, 1:1,1:4, 1:8, 1:16, and 1:32) of blank extracts obtained fromCD-treated serum were tested to assess whether toxicor spurious estrogenic activity was acquired duringsample preparation. Extracts were resuspended as
above and diluted with 50 milL CD-FBS. Cell yieldswere comparable with those obtained when similarinocula were grown in 50 milL CD-FBS, indicating theabsence ofspurious estrogenicactivity. Although grosstoxic effects were not seen microscopically, toxic activ-ity in the undiluted preparation and in the 1:1 and 1:4dilutions was suspected because addition of 100 pmol/LE2 to these preparations resulted in a substantialincrease of cell yield, albeit lower than the increase inmedia supplemented with 50 mLIL CD-treated serumplus 100 pmolJL estradiol (Fig. 2A). A full estrogenicresponse was obtained when 100 pmol/L estradiol wasadded to the 1:8 dilution. A comparable toxic effect wasdetected in the nonylphenol-supplemented extractwhen tested undiluted and at 1:1 and 1:4 dilutions.Estrogenic activity caused by nonyiphenol was de-tected at dilutions between 1:4 and 1:32. The expectednonylphenol concentration range of these dilutionsfrom extraction to bioassay was 7.5 to 1 .tmolIL, dosesthat are fully estrogenic when added directly to 50mLIL CD-FBS-supplemented medium (Fig. 1B). Tolearn more about the toxic moiety, we assayed blankextracts eluted in two steps, as described in the methodof Burse et al. (21); the Fl eluate (60 mL/L ethyl etherin petroleum ether) was toxic, but the F2 fraction wasnot. These experiments indicate that this approach isfeasible; the cleanup protocol needs further refinementto eliminate the toxic component in the extract.
Xenoestrogen Bioassays
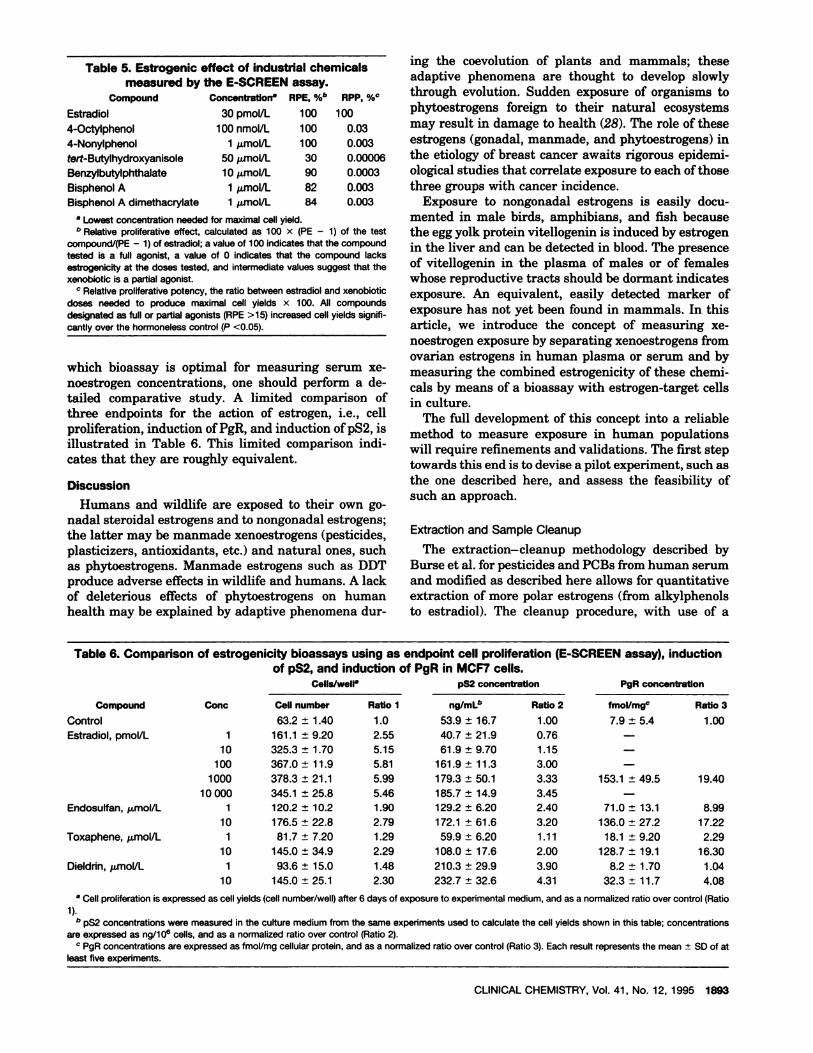
The model that most “in culture” bioassays use is thehuman breast cancer MCF7 cell line, a genuine estro-gen-target cell line. When inoculated into nude mice,tumors develop only in estrogen-treated animals. Table5 illustrates the estrogenic effect of industrial chemi-cals selected to monitor extraction and separation fromovarian estrogens with induction of cell proliferation(E-SCREEN assay) as the endpoint. Before deciding
Compound Conc Cell number Ratio 1 ng/mL”
10
100
1000
10 000
Ratio 2 fmol/mgc
7.9 ± 5.4
Ratio 3
Table 6.Comparison of estrogenicitybioassays using as endpoint cellproliferation(E-SCREEN assay),inductionof pS2, and inductionof PgR inMCF7 cells.
CelIs/well pS2 concentration PgR concentration
Control 63.2 ± 1.40 1.0 53.9 ± 16.7 1.00 1.00
Estradiol, pmol/L 1 161.1 ± 9.20 2.55 40.7 ± 21.9 0.76
325.3 ± 1.70 5.15 61.9 ± 9.70 1.15 -
367.0 ± 11.9 5.81 161.9 ± 11.3 3.00 -
378.3 ± 21.1 5.99 179.3 ± 50.1 3.33 153.1 ± 49.5 19.40
345.1 ± 25.8 5.46 185.7 ± 14.9 3.45 -
Endosulfan, mol/L 1 120.2 ± 10.2 1.90 129.2 ± 6.20 2.40 71.0 ± 13.1 8.99
10 176.5 ± 22.8 2.79 172.1 ± 61.6 3.20 136.0 ± 27.2 17.22
Toxaphene, mol/L 1 81.7 ± 7.20 1.29 59.9 ± 6.20 1.11 18.1 ± 9.20 2.29
10 145.0 ± 34.9 2.29 108.0 ± 17.6 2.00 128.7 ± 19.1 16.30
Dieldrin, mol/L 1 93.6 ± 15.0 1.48 210.3 ± 29.9 3.90 8.2 ± 1.70 1.04
10 145.0 ± 25.1 2.30 232.7 ± 32.6 4.31 32.3 ± 11.7 4.08
a Cell proliferation is expressed as cell yields (cell number/well) after 6 days of exposure to experimental medium, and as a normalized ratio over control (Ratio
1).C pS2 concentrations were measured in the culture medium from the same experiments used to calculate the cell yields shown in this table; concentrations
are expressed as ng/1 08 cells, and as a normalized ratio over control (Ratio 2).C PgR concentrations are expressed as fmoVmg cellular protein, and as a normalized ratio over control (Ratio 3). Each result represents the mean ± SD of at
least five experiments.
CLINICAL CHEMISTRY, Vol. 41, No. 12, 1995 1893
Table 5.Estrogenic effectof industrialchemicalsmeasured by the E-SCREEN assay.
Concentratlon RPE, %b RPP, %#{176}
Estradiol 30 pmol/L 100 100
4-Octylphenol 100 nmol/L 100 0.03
4-Nonylphenol 1 mol/L 100 0.003
tert-Butylhydroxyanisole 50 jmol/L 30 0.00006
Benzylbutylphthalate 10 mol/L 90 0.0003
Bisphenol A 1 mol/L 82 0.003
Bisphenol A dimethacrylate 1 imol/L 84 0.003
a Lowest concentration needed for maximal cell yield.r Relative proliferative effect, calculated as 100 x (PE - 1) of the test
compound/(PE - 1) of estradiol; a value of 100 indicates that the compoundtested is a full agonist, a value of 0 indicates that the compound lacksestrogenicity at the doses tested, and intermediate values suggest that thexenobiotic is a partial agonist.
Relative proliferative potency, the ratio between estradiol and xenobioticdoses needed to produce maximal cell yields x 100. All compoundsdesignated as full or partial agonists (RPE >15) increased cell yields signifi-cantly over the hormoneless control (P <0.05).
which bioassay is optimal for measuring serum xe-noestrogen concentrations, one should perform a de-tailed comparative study. A limited comparison ofthree endpoints for the action of estrogen, i.e., cellproliferation, induction of PgR, and induction of pS2, isillustrated in Table 6. This limited comparison indi-cates that they are roughly equivalent.
Discussion
Humans and wildlife are exposed to their own go-nadal steroidal estrogens and to nongonadal estrogens;the latter may be manmade xenoestrogens (pesticides,plasticizers, antioxidants, etc.) and natural ones, suchas phytoestrogens. Manmade estrogens such as DDTproduce adverse effects in wildlife and humans. A lackof deleterious effects of phytoestrogens on humanhealth may be explained by adaptive phenomena dur-
Compound
ing the coevolution of plants and mammals; theseadaptive phenomena are thought to develop slowlythrough evolution. Sudden exposure of organisms tophytoestrogens foreign to their natural ecosystemsmay result in damage to health (28). The role of theseestrogens (gonadal, manmade, and phytoestrogens) inthe etiology of breast cancer awaits rigorous epidemi-ological studies that correlate exposure to each of thosethree groups with cancer incidence.
Exposure to nongonadal estrogens is easily docu-mented in male birds, amphibians, and fish becausethe egg yolk protein vitellogenin is induced by estrogenin the liver and can be detected in blood. The presenceof vitellogenmn in the plasma of males or of femaleswhose reproductive tracts should be dormant indicatesexposure. An equivalent, easily detected marker ofexposure has not yet been found in mammals. In thisarticle, we introduce the concept of measuring xe-noestrogen exposure by separating xenoestrogens fromovarian estrogens in human plasma or serum and bymeasuring the combined estrogenicity of these chemi-cals by means of a bioassay with estrogen-target cellsin culture.
The full development of this concept into a reliablemethod to measure exposure in human populationswill require refinements and validations. The first steptowards this end is to devise a pilot experiment, such asthe one described here, and assess the feasibility ofsuch an approach.
Extractionand Sample Cleanup
The extraction-cleanup methodology described byBurse et al. for pesticides and PCBs from human serumand modified as describedhere allows for quantitativeextraction of more polar estrogens (from alkylphenolsto estradiol). The cleanup procedure, with use of a

1894 CLINICAL CHEMISTRY, Vol. 41, No. 12, 1995
Sep-Pak cartridge, was also simplified by eluting thexenoestrogens of interest with 150 milL ethyl ether inpetroleum ether practically free of estradiol (Table 1)and with reproducible high yields (Tables 2 and 3).Although this cleanup procedure seemed adequate foranalytical purposes, it resulted in coelution of un-known moieties that were somewhat toxic to MCF7cells. This toxic effect masked the estrogenic activity inthe extract of nonylphenol-supplemented serum; dilu-tion resulted in progressive unmasking of the estro-genic activity.Full recovery occurred at a 1:8dilution(Fig.2B). This toxicactivitywas not evident in blankextracts because cell yields resembled those obtainedin 50 mIlL CD-treated FBS and cells looked healthywhen examined by phase-contrast microscopy; additionof 100 pmoL’L E2 to this extract revealed that fullexpression of the estrogenic effect required a 1:8 dilu-tion of the extract (Fig. 2A). Further cleanup will benecessary to remove toxic moieties from the xenoestro-gen extract.
Note added in proof The toxic effect of serum ex-tracts was removed by deleting the Sep-Pak cleanupstep and by resuspending the HPLC fraction in 100 p.Lofethanol.The ethanol extractwas then transferredtoa clean tube containing 9.9 mL of 50 milL CD-treatedFBS.
Final Separation of Xenoestrogens from Ovarian Estrogens
by HPLC in Silica Column
Estrogenic pesticides, PCBs, hydroxylated PCBs,phenolic antioxidants, and plasticizers eluted from theHPLC column during the first 10 mm; ovarian estro-gens, phytoestrogens, diethylstilbestrol, and mycoes-trogens were retained for >12 mm. We expect that thepopulations to be investigated would not be exposed todiethylstilbestrol and mycoestrogens; these agentshave been used as anabolic agents in cattle but, basedon current regulations, should not exist in meats.Phytoestrogens are full estrogen agonists in animalmodels and in estrogen-target cells in culture; however,some propose that phytoestrogens are protectiveagainst breast cancer (29, 30). This has not been con-firmed. Thus, we propose to measure the combinedeffect of the compounds eluted during the first 10 mmas a marker of exposure to manmade xenoestrogens.
Evaluation of Bioassays That May Be Used to Measure
Estrogenic Activity
The ability of estrogens to induce proliferation oftheir target organs is considered the hallmark of estro-gen action. Therefore, the most reliable bioassay toassess estrogenicity should be one that measures cellproliferation as the endpoint. Uterotropic assays mea-sure increases of uterine wet weight caused by theadditive effect of cell proliferation, hypertrophy, andwater imbibition. Several variations of the uterotropicassay have been used to determine estrogenicity; singleor multiple doses of estrogens were administered dur-ing 24-72 h in immature and ovariectomized mice andrats. This diversity of endpoints implies that there is no
agreed universal “gold standard” of the action of estro-gen among animal bioassays. However, the E-SCREENassay appears to be the best candidate for establishinga quantitative standard of estrogenic activity at thetarget organ level, given that no false positives ornegatives have been observed thus far among theestrogens and nonestrogens tested. In addition, theE-SCREEN assay is the only assay that has beenvalidatedforallthe known estrogens (17).
Estrogenicity may be assessed with other endpoints,such as the induction of PgR and of p52 in MCF7 cells,the same cells used in the E-SCREEN assay. These twoassays are reliable but have yet to be tested for allknown estrogenic compounds. Under the conditionsused here (Table 6), an advantage of the PgR inductionassay is the 20-fold difference between control andmaximal activity by estradiol; in the pS2 assay, differ-
ences between control and experimentals were three-fold. In the E-SCREEN assay, differences were up tosixfold.Cell proliferationassays are more sensitive
than those that induce gene products; several labora-tories (31-33) have reported that the estrogen doseneeded to induce maximal cell proliferation is -50-100times lower than that required to elicit maximal induc-tion of PgR or pS2.
One substantial reservation about gene expressionassays is that estrogen-inducible genes could also beinduced by nonestrogenic substances. For example,prolactin synthesis may be induced by epidermalgrowth factor, thyrotropin-releasing factor, and phor-bol esters (34). Another estrogen-inducible marker,ovalbumin synthesis, is stimulated by other steroidssuch as progesterone and glucocorticoids (35). Induc-tionofreportergenes under controlofestrogen-respon-siveelements has been proposed to assess estrogenic-ity; however, increased basal expression in the absence
of estrogen often occurs and may raise concern aboutthe reliability of these assays. On the other hand, geneexpression assays in general, and particularly thoseinvolving reporter genes, are amenable to use in micro-methods by manipulations that enhance the signal;this potential for increased sensitivity may be exploitedto reduce the amount of biological material required.By contrast, since Allen and Doisy (36) isolated es-trone, the operational definition of estrogen has beenthe ability of these compounds to induce cell prolifera-tion in organs of the female genital tract; therefore,both conceptually and operationally, cell proliferationremains the most reliable endpoint for identifyingestrogenic chemicals.
Our data and analysis attempt to cope with demandsby the public and regulatory agencies to evaluaterigorously the role of xenoestrogens in the increase ofmale genital tract anomalies, testicular cancer, andbreast cancer in human populations.
Note added to proof A report describing the estroge-nicity of plasticizers and antioxidants has been pub-lished (37).

CLINICAL CHEMISTRY, Vol. 41, No. 12, 1995 1895
This work was partially supported by grants from the W. AltonJones Foundation, EPA-CR 820301, NIH-CA-13410, NSF-DCB-9105594, the Silent Spring Institute, and the MassachusettsDepartment of Public Health (DPH 79005-214 Hil). The assis-tance of The Center for Reproductive Research at Tufts Univer-sity (P30 HD 28897) is acknowledged. We are grateful to Maria F.Luizzi for her skillful technical assistance.
References
1. Colborn T, vom Saal FS, Soto AM. Developmental effects ofendocrine-disrupting chemicals in wildlife and humans. EnvironHealth Perspect 1993;1O1:378-84.2. Singer PL. Occupational oligospermia [Letter). JAMA 1949;140:1249.3. Guzelian PS. Comparative toxicology of chlorodecone (kepone)in humans and experimental animals. Annu Rev PharmacolToxicol 1982;22:89-113.4. Fry DM. Sex ratio skew and breeding patterns of gulls:demographic and toxicological considerations. Stud Avian Biol1987;10:26-43.5. Guillette U. Developmental abnormalities of the gonad andabnormal sex hormone concentrations in juvenile alligators fromcontaminated and control lakes in Florida. Environ Health Per-spect 1994;102:680-8.6. Soto AM, Sonnenschein C. Environmental sex hormone ago-nists and antagonists. Comments Toxicol (in press).7. Purdom CE, Hardiman PA, Bye VJ, Eno NC, Tyler CR,Sumpter JP. Estrogenic effects from sewage treatment works.Chem Ecol 1994;8:275-85.8. Brotons JA, Olea-Serrano MF, Villalobos M, Pedraza, V, OleaN. Xenoestrogens released from lacquer coating in food cans.Environ Health Perspect 1995;103:608-13.9. Sharpe RM, Skakkebaek NE. Are oestrogens involved infalling sperm count and disorders of the male reproductive tract?Lancet 1993;341: 1392-5.10. Giwercman A, Carlsen E, Keiding N, Skakkebaek NE. Evi-dence for increasing incidence of abnormalities of the humantestis: a review. Environ Health Perspect 1993;101:65-71.11. Davis D. Medical hypothesis: xenoestrogens as preventablecauses of breast cancer. Environ Health Perspect 1993;101:372-7.12. Sonnenschein C, Soto AM. Control of estrogen-target cellproliferation, environmental estrogens and breast tumorigenesis.Comments Toxicol (in press).13. Russo J, Russo IH. Biological and molecular bases of mam-mary carcinogenesis. Lab Invest 1987;57:112-37.14. Toniolo PG, Levitz M, Zeleniuch-Jacquotte A, Bannerjee 5,Koenig KL, Shore RE, et al. A prospective study of endogenousestrogens and breast cancer in postmenopausal women. J NatlCancer Inst 1995;87:190-7.15. Wolff MS, Paolo G, Toniolo PG, Lee EW, Rivera M, Dubin N.Blood levels of organochiorine residues and risk of breast cancer.J Natl Cancer Inst 1993;85:648-52.16. Krieger N, Wolff MS, Hiatt RA, Rivera M, Vogelman J,Orentreich N. Breast cancer and serum organochlorines: a pro-spective study among white, black, and Asian women. J Nat)Cancer Inst 1994;86:589-99.17. Soto AM, Sonnenschein C, Chung KL, Fernandez MF, OleaN, Olea-Serrano MF. The E-SCREEN assay as a tool to identirestrogens: an update on estrogenic environmental pollutants.Environ Health Perspect 1995 (in press).18. Hertz R. The estrogen problem: retrospect and prospect. In:McLachlan JA, ed. Estrogens in the environment, Vol. 2. NewYork: Elsevier, 1985:1-11.19. Jeltsch JM, Roberts M, Scatz C, Gamier JM, Brown AMC,Chambon P. Structure of the human oestrogen-responsive genepS2. Nucleic Acids Res 1987;15:1401-14.
20. Soto AM, Sonnenschein C. The role of estrogens on theproliferation of human breast tumor cells (MCF-7). J SteroidBiochem 1985;23:87-94.21. Burse VW, Head SL, Korver MP, McClure PC, Donahue JF.Determination of selected organochlorine pesticides and polychlo-rinated biphenyls in human serum. J Anal Toxicol 1990;14:137-42.22. Medina MB, Sherman JT. HPLC separation of anabolicoestrogens and ultraviolet detection of 17-beta oestradiol, zera-no!, diethyistilboestrol or zearalenone in avian muscle tissueextract. Food Addit Contam 1986;3:263-72.23. Soto AM, Sonnenschein C. Mechanism of estrogen action oncellular proliferation: evidence for indirect and negative controlon cloned breast tumor cells. Biochem Biophys Res Commun1984;122:1097-103.
24. Soto AM, Justicia H, Wray JW, Sonnenschein C. p-Nonyl-phenol: an estrogenic xenobiotic released from “modified” poly-styrene. Environ Health Perspect 1991;92: 167-73.25. Soto AM, Lin TM, Justicia H, Silvia RM, Sonnenschein C. An“in culture” bioassay to assess the estrogenicity of xenobiotics. In:Colborn T, Clement C, eds. Chemically induced alterations insexual development: the wildlife/human connection. Princeton,NJ: Princeton Scientific Publishing, 1992:295-309.26. Soto AM, Silvia RM, Sonnenschein C. A plasma-borne spe-cific inhibitor of the proliferation of human estrogen-sensitivebreast tumor cells (estrocolyone-I). J Steroid Biochem Mol Biol1992;43:703-12.27. Maddedu L, Legros N, Devleeschouwer N, Bosman C, PiccartM, Leclercq G. Estrogen receptor status and estradiol sensitivityof MCF-7 cells in exponential growth phase. Eur J Cancer ClinOncol 1988;24:385-90.28. Shutt DA. The effects of plant estrogens on animal reproduc-tion. Endeavour 1976;35:110-3.29. Adlercreutz H. Western diet and Western diseases: somehormonal and biochemical mechanisms and associations. Scand JClin Lab Invest 1990;50(Suppl):3-23.30. Messina MJ, Persky P, Setchell KDR, Barnes S. Soy intakeand cancer risk: a review of the in vitro and in vivo data. NutrCancer 1994;21:113-31.31. Soto AM, Sonnenschein C. Cell proliferation of estrogen-sensitive cells: the case for negative control. Endocr Rev 1987;8:44-52.32. Katzenellenbogen BS, Kendra KL, Norman MJ, Berthois Y.Proliferation, hormonal responsiveness, and estrogen receptorcontent of MCF-7 human breast cancer cells grown in the short-term and long-term absence of estrogens. Cancer Res 1987;47:4355-60.33. Reiner GCA, Katzenellenbogen BS. Characterization of es-trogen and progesterone receptors and the dissociated regulationof growth and progesterone receptor stimulation by estrogen inMDA-MB-134 human breast cancer cells. Cancer Res 1986;46:1124-31.34. Ramsdell JS, Tashjian AH. Thyrotropin releasing hormoneand epidermal growth factor stimulate prolactin synthesis by apathway(s) that differs from that used by phorbol esters: dissoci-ation of actions by calcium dependency additivity. Endocrinology1985;117:2050-60.35. Palmiter RD. Quantitation of parameters that determine therate of ovalbumin synthesis. Cell 1975;4:189-97.36. Allen E, Doisy EA. An ovarian hormone: preliminary reporton its localization, extraction and partial purification, and actionin test animals. JAMA 1923;81:819-21.37. Jobling S, Reynolds T, White R, Parka MG, Sumpter JP. Avariety of environmentally persistent chemicals, including somephthalate plasticizers are weakly estrogenic. Environ HealthPerspect 1995;103:582-7.




















