
Prevalence and prognosis of Alzheimer’s disease at the mild cognitive impairment stage
Upload
independentCategory
view
0download
0
ORIGINAL ARTICLE
17b-Oestradiol Stimulation of G-Proteins in Aged and Alzheimer’s HumanBrain: Comparison with PhytoestrogensV. Jefremov,*� A. Rakitin,*� R. Mahlapuu,* K. Zilmer,* N. Bogdanovic,� M. Zilmer* and E. Karelson*
*Department of Biochemistry, Tartu University, Tartu, Estonia.
�Geriatric Department, Neurotec, Karolinska Institute, Huddinge, Sweden.
�Department of Neurology and Neurosurgery, Tartu University Hospital, Tartu, Estonia.
Over the past decade, it has become apparent that the neuropro-
tective action of oestrogens may result from the oestrogen receptor
(ER)-dependent and -independent mechanisms (1, 2). The ER-
dependent mechanisms may be exerted via genomic way (i.e.
through the activation of intracellular ERa and ERb and oestrogen
target genes) (3, 4). In addition, the nongenomic ER-dependent
mechanisms of oestrogens in target cells and tissues have been
described (5, 6). These steroids may interact with the plasma mem-
brane-located ER-like proteins that can activate the G-protein med-
iated down-stream signalling (e.g. cAMP and protein kinase C
Journal ofNeuroendocrinology
Correspondence to:
Ello Karelson, Department of
Biochemistry, University of Tartu,
Ravila 19, 50411 Tartu, Estonia
(e-mail: [email protected]).
The neuroprotective action of oestrogens and oestrogen-like compounds is in the focus of basic
and clinical research. Although such action has been shown to be associated with neuronal
plasma membranes, the implication of G-proteins remains to be elucidated. This study revealed
that micromolar concentrations (lM) of 17b-oestradiol and phytoestrogens, genistein and daidz-
ein, significantly (P < 0.05) stimulate G-proteins ([35S]GTPcS binding) in the post-mortem hippo-
campal membranes of age-matched control women with the respective maximum effects of 28,
20 and 15% at 10 lM. In the frontocortical membranes, the stimulation of G-proteins did not
differ significantly from that in hippocampal membranes. Although in the hippocampus and
frontal cortex of the Alzheimer’s disease (AD) women’s brain, 10 lM 17b-oestradiol produced
significantly (P < 0.05) lower stimulation of G-proteins than in the control regions, stimulation
by phytoestrogens revealed no remarkable decline. 17b-Oestradiol, genistein and daidzein
revealed a selective effect on various G-proteins (Gas, Gao, Gai1 or Ga11 plus Gb1c2) expressed in
Sf9 cells. At a concentration of 10 lM, 17b-oestradiol suppressed the H2O2 and homocysteine
stimulated G-proteins in the frontocortical membranes of control women to a greater extent
than phytoestrogens. In AD, the suppressing effect of each compound was lower than in the
controls. In the cell-free systems, micromolar concentrations of phytoestrogens scavenged OH•
and the 2.2-diphenyl-1-picrylhydrazyl free radical (DPPH•) more than 17b-oestradiol did. In the
frontocortical membranes of control women, the 20 lM 17b-oestradiol stimulated adenylate
cyclase with 20% maximal effect, whereas, in AD, the effect was insignificant. Genistein did not
stimulate enzyme either in control or AD frontocortical membranes. Our data confirm that the
agents stimulate G-proteins in control and AD women’s brains, although 17b-oestradiol and
phytoestrogens have similarities and differences in this respect. We suggest that, besides the
ER-dependent one, the ER-independent antioxidant mechanism is responsible for the oestrogen
stimulation of G-proteins in the brain membranes. Both of these mechanisms could be involved
in the neuroprotective signalling of oestrogens that contributes to their preventive ⁄ therapeutic
action against postmenopausal neurological disorders.
Key words: 17b-oestradiol, phytoestrogens, antioxidativity, G-proteins, aged, Alzheimer’s disease
brain.
doi: 10.1111/j.1365-2826.2008.01696.x
Journal of Neuroendocrinology 20, 587–596
ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd

pathways) (7–10). The available evidence suggests that oestrogens,
like traditional growth factors, recognise the membrane receptors
with intrinsic tyrosine kinase activity (11). These and other mem-
brane-associated mechanisms, such as the alteration of membrane
fluidity (12), activation of K+ channels (13) and stimulation of Ca2+
influx (14), were thought to be involved in the nongenomic neuro-
protection by oestrogens (15).
The ER-independent neuroprotective action of oestrogens
appears to be mainly caused by their antioxidant free-radical scav-
enging properties. The hydroxyl group in the C3 position on the A
ring of the steroid molecule is mainly responsible for these proper-
ties (16–18). In addition, the modulation of neurotransmission and
neuronal excitability may contribute to the ER-independent neuro-
protective action of oestrogens (19, 20). Studies with neuronal cell
lines have shown that nano- to micromolar concentrations of
17b-oestradiol reduce neuronal cell death induced by glutamate,
superoxide anion, H2O2, neurotoxic amyloid-b peptides and other
pro-oxidants (21–23). In vivo administered pharmacological doses
(mg ⁄ kg) of 17b-oestradiol are effective in protecting the animal
brain from the ischaemic injury (24) and deposition of amyloid-bpeptides (25). More interestingly, pharmacological doses of oestro-
gens may exert preventive ⁄ therapeutic effects against ischaemic
stroke and neurodegeneration in postmenopausal women (26, 27)
whereas age-matched men may reveal a weaker response (28).
In recent years, an interest has grown in alternative compounds
that act as selective oestrogen receptor modulators, but have no
unfavourable side-effects of oestrogen. One group of such com-
pounds consists of phytoestrogens, plant-derived (iso)flavones with
oestrogenic activity, which may act as oestrogen receptor agonists
or antagonists in various cells and tissues (29). In the nano- to
micromolar range, these polyphenolic compounds have been shown
to exert multiple protective effects (30), including neuroprotective
ones (31), that are mainly derived from their antioxidant properties
(32). In addition to the antioxidant activities, several other mecha-
nisms appear to be involved in the neuroprotective action of plant
polyphenols; the most important one might be their ability to
improve neuronal signalling and communication (33). However,
despite these findings, the mechanisms responsible for the neuro-
protective action of plant-derived and endogenous oestrogens need
to be studied further, particularly in the human brain.
The present study aimed to compare the effect of micromolar
concentrations of 17b-oestradiol and phytoestrogenic isoflavones
(genistein, daidzein) (Fig. 1) on G-proteins in the membranes of
age-matched control and Alzheimer’s disease (AD) hippocampus
and frontal cortex post-mortem. Both regions revealed morpho-
functional alterations typical of brain ageing and neurodegeneration
(34, 35). The second objective was to test whether the compounds
under investigation can discriminate between the different types of
heterotrimeric G-proteins expressed in Sf9 insect cells. The third
objective was to compare the antioxidant activity of the compounds
as a property that might imply their modulatory action on G-pro-
teins. To that purpose, the influence of 17b-oestradiol and phytoes-
trogens was tested regarding their influence on the oxymodified
membrane G-proteins and the ability to scavenge 2.2-diphenyl-1-
picrylhydrazyl (DPPH•) and OH• radicals in vitro. Finally, the modu-
lation of adenylate cyclase activity as a possible signalling mecha-
nism for the improvement of neuronal function by oestrogens
comprised an interesting problem to be resolved.
Materials and methods
Materials
All reagents used were of highest analytical grade, mainly purchased from
Sigma Chemical Co. (St Louis, MO, USA). [35S]-GTPcS (1250 Ci ⁄ mmol) was
obtained from NEN Life Science Products (Boston, MA, USA).
Brain tissue sampling
Hippocampal and frontal cortical tissues of post-mortem human brain
were obtained from Huddinge Brain Bank, Sweden. The work of the Brain
Bank as well as of the Department of Biochemistry at Tartu University
was approved by the Ethical Committees at Karolinska Institute (Stockholm,
Sweden) and Tartu University (Tartu, Estonia). The present study included
the regions from 12 aged-matched control and eight AD female subjects
(aged 78 � 6 years and 84 � 12 years, respectively). In addition, the
frontocortical tissues from four aged and four AD male persons (aged
72 � 4 years and 78 � 5 years, respectively) were used. The post-mortem
delay for aged and AD persons was 16 � 5 h and 14 � 8 h, respectively.
The AD patients met clinical DSM-IV criteria (36) and neuropathological
CERAD criteria (37, 38) for AD. The control group consisted of aged-
matched persons with no history of neurological or psychiatric disorders.
The respective brain regions were rapidly dissected and kept at )70 �C
prior to the experiment.
Preparation of brain membranes
The hippocampal and frontocortical membranes for [35S]-GTPcS binding
measurement and for the adenylate cyclase assay were mainly prepared
according to protocol of Karelson et al. (39). The membrane pellet was
finally resuspended either in the standard TE-buffer consisting of 10 mM
Tris-HCl and of 0.1 mM ethylenediaminetetraacetic acid (EDTA), pH 7.4
([35S]-GTPcS binding) or in the 4 mM HEPES-Na containing 1.5 mM the-
ophylline, 8.25 mM MgCl2, 0.75 ethylene glycol tetraacetic acid (EGTA),
7.5 mM KCl and 100 mM NaCl, pH 7.4 (adenylate cyclase assay) to obtain
a protein concentration 0.6–1 mg ⁄ ml as determined by the method of
Lowry et al. (40).
O
OHOOH
HOO
OHO
HOGenisteinDaidzein
OH
HO
Fig. 1. Structure of 17b-oestradiol, daidzein and genistein.
588 V. Jefremov et al.
ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 20, 587–596

Preparation of membranes from Sf9 cells
Sf9 cells (the Spodoptera frugiperda insect cell line) were transfected with
baculovirus vectors carrying the genes for the different G-protein alpha-
subunits (Gas, Gao, Gai1 or Ga11) and for Gb1c2. The transfection procedure
was perfomed by Dr Kulliki Saar (Department of Neurochemistry, University
of Stockholm, Sweden) in accordance with a previously described protocol
(41). The transfected Sf9 cells were stored at )70 �C. For the preparation
of membranes, the cells were thawed, resuspended in five volumes of ice-
cold TE-buffer and homogenised. The homogenate was centrifuged at
500 g for 15 min followed by centrifugation of the supernatant at
40 000 g for 40 min. The obtained pellet was washed in ten volumes of
TE-buffer and recentrifuged in the same conditions. The resulting pellet
was resuspended in TE-buffer to obtain the membrane preparation with
2–4 mg ⁄ ml of protein.
[35S]-GTPcS-binding assay
The brain or cell membranes with the final protein concentration of 0.04–
0.05 mg ⁄ ml were incubated in a reaction cocktail containing TE-buffer, GDP
(1 lM), dithiothreitol (1 mM), MgCl2 (5 mM), NaCl (150 mM) and [35S]-GTPcS
(50–70 000 c.p.m. in an aliquot of the reaction cocktail). Incubation of the
membranes was performed at 26 �C in a total volume of 0.1 ml either in
the absence (basal value) or in the presence of various concentrations of
17b-oestradiol or phytoestrogens. Ten minutes of incubation was used to
detect the effect of oestrogens in the human brain membranes. The
membranes of SF9 cells were incubated with oestrogens for 10–30 min
depending on the type of G-protein expressed in the cells. Bound and free
[35S]-GTPcS were separated by vacuum filtration through GF ⁄ B filters
(Whatman International Ltd, Maidstone, UK), which were washed three times
with 5 ml of ice-cold TE-buffer. Radioactivity was quantified using a
Wallac-1409 liquid scintillation counter (Wallac, Turku, Finland). In the data
analysis, the basal (unaffected) [35S]-GTPcS binding was defined as 100%.
Determination of protective effect on the oxymodifiedG-proteins
The protective effect of 17b-oestradiol and phytoestrogens was determined
via the changes that they produced in [35S]-GTPcS binding after the pre-
treatment of membranes with two metabolic pro-oxidants, homocysteine
(Hcy) and H2O2. The 5-min pretreatment of control and AD membranes by
500 lM Hcy or 10 lM H2O2 was carried out at 26 and 30 �C, respectively.
The pre-treatment was terminated by putting the samples into the ice.
Immediately after that, the oestrogen solution and reaction cocktail were
added and [35S]-GTPcS binding was performed. The protective activity was
estimated as a difference between the effects of Hcy or H2O2 on the radioli-
gand binding in the absence and presence of 10 lM 17b-oestradiol or
phytoestrogens.
Measurement of adenylate cyclase activity
The basal adenylate cyclase activity was assayed by determining the cAMP
formed in the reaction media (final volume of 150 ll), which contained a
reaction buffer (30 mM Tris-HCl buffer, 1.5 mM theophylline, 8.25 mM
MgCl2, 0.75 mM EGTA, 7.5 mM KCl and 0.1 M NaCl, 100 lg ⁄ ml bacitracin,
0.05% bovine serum albumin, 10 mM phosphoenol-pyruvate, 45 lg ⁄ ml
pyruvate kinase) and brain membranes with the final protein concentration
of 0.06–0.08 mg ⁄ ml (39). The oestrogenic compounds, dissolved in the 1%
ethanol ⁄ water, were added 2 min before the reaction was initiated by
10 mM ATP ⁄ 10 lM GTP. The reaction at 30 �C was terminated after
15 min by adding 100 mM EDTA and boiling the samples for 3 min. The
cyclic AMP content in the tubes was measured by a competitive protein
saturation assay (42).
Hydroxyl radical scavenging assay
The ability of 17b-oestradiol and phytoestrogens to scavenge OH• was
determined according to (43), using terephthalic acid (THA) as OH• chemical
dosimeter. In brief, 2.0 ml of 10 mM THA solution in 10 mM sodium phos-
phate buffer (pH 7.5) was mixed in a spectrometric cell with 0.1 ml of the
(phyto)estrogen solution. The neutral salt solution consisting of equal
amounts of 0.1 M HCl and 0.1 M NaOH solutions was used as a solvent of
(phyto)estrogens. The mixture of 2.0 ml of THA and 0.1 ml of neutral salt
solutions was used as a control. OH• was generated via the Fenton reaction
initiated by adding the CuSO4 and H2O2 solutions (final concentrations
10 lM and 1 mM, respectively) to the dosimeter solution. The reaction prod-
uct of THA with OH• (THA-OH•) was measured using a Perkin-Elmer LS50B
Luminescence spectrometer (Perkin-Elmer Life Sciences, Inc., Boston, MA,
USA) with excitation at 312 nm and emission at 426 nm. The radical scav-
enging activity of the investigated compounds was expressed as a percent
inhibition of the THA-OH• peaks. The experimental data were analysed using
the one-site binding model (GraphPad Prism, version 3.0; GraphPad Software
Inc., San Diego, CA, USA).
DPPH free radical scavenging assay
The DPPH (2.2-dipenyl-1-picrylhydrazyl) free radical (DPPH•) scavenging
activity of 17b-oestradiol and phytoestrogens was measured according to
the method of Blois (44), as modified by us. In brief, 0.2 ml of the agent
ethanol solution was mixed in a test tube with 0.2 ml of 100 lM DPPH
dissolved in ethanol. The reaction mixture was maintained at room tem-
perature for 100 min. The absorbance of the mixture was recorded at
517 nM by an ultraviolet-visible spectrophotometer Jenway 6405 (Barlo-
world Scientific Ltd, T ⁄ As Jenway, Dunmow, UK). The DPPH• scavenging
activity of 17b-oestradiol and phytoestrogens was calculated by the fol-
lowing equation:
Scavenging ability ð%Þ ¼ ðAo � AcÞ=A0 � 100
where Ao is the absorbance of the DPPH solution (without tested agent),
Ac is the absorbance of the mixture containing both DPPH and the tested
agent.
Preparation of 17b-oestradiol and phytoestrogens solutions
1 mM stock solutions of 17b-oestradiol and phytoestrogens were prepared
in 1% ethanol ⁄ water followed by dilution of stock solutions in the respec-
tive reaction cocktails (for [35S]-GTPcS-binding and adenylate cyclase activity
assays) to get the respective final concentrations 1–30 lM.
Statistical analysis
Each experiment on the compound’s effect on [35S]-GTPcS binding and
adenylate cyclase activity was made independently with the duplicate sam-
ples. The mean � SEM for each statistical group was calculated. The n used
in the statistical analysis was the number of individual brains in the control
or AD group. An unpaired Student’s t-test or one-way ANOVA was used to
identify significant differences (P < 0.05) between the groups. The experi-
ments on the free radical scavenging activities of the compounds were
repeated three to five times and the means � SEM was calculated for each
concentration. The statistical analysis of the experimental data sets was
made using GraphPad Prism software.
17b-oestradiol stimulation of G-proteins in human brain 589
ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 20, 587–596

Results
Effect of 17b-oestradiol, genistein and daidzein on[35S]-GTPcS binding in the hippocampal and frontocorticalmembranes
In the membranes from the hippocampus and frontal cortex of
control and AD females, 17b-oestradiol, genistein and daidzein
showed a capability of stimulation the G-proteins (Figs 2 and 3). In
the control hippocampus, 5–10 lM concentrations of the agents
significantly (P < 0.05) stimulated [35S]-GTPcS binding with the
respective maximum effects of 28%, 20% and 15% over the basal
level (421 � 25 fmol ⁄ mg protein). In the AD hippocampus, 10 lM
17b-oestradiol induced significantly (P < 0.05) weaker stimulation
of the radioligand basal binding (374 � 29 fmol ⁄ mg) than that of
the control, whereas the effect of phytoestrogens revealed no
remarkable decline (Fig. 2). In control frontal cortex, the stimulatory
effect of 17b-oestradiol on [35S]-GTPcS basal binding (391 � 21
fmol ⁄ mg) did not differ significantly from that in the control hip-
pocampus. In the AD frontal cortex, 8 and 10 lM 17b-oestradiol
produced significantly (P < 0.05) less stimulation of basal binding
(358 � 26 fmol ⁄ mg) than in the control region, whereas the effect
of phytoestrogens did not show a significant decrease (Fig. 3). In
both control and AD frontal cortex, the females showed a signifi-
100
105
110
115
120
125
130
135
† †
†
*
†
†
†
† †
†
‡
‡
‡
‡
Co
Genistein Daidzein
Alzheimer's disease
1 3 5 8 10 3 1 5 8 10 1 3 5 8 10 µM
Fig. 2. Effect of 1–10 lM 17b-oestradiol, genistein and daidzein on the [35S]-GTPcS binding in the female age-matched control (Co) and Alzheimer’s disease
(AD) hippocampal membranes (protein 0.05 mg ⁄ ml). 100% corresponds to the basal [35S]-GTPcS binding (means � SEM). In both Co and AD, four to five inde-
pendent experiments with the individual brains were conducted; *P < 0.05 for Co hippocampus versus AD hippocampus; comparison of the stimulated [35S]-
GTPcS binding with the basal level: P < 0.05 in Co (�) and AD (�) hippocampus.
100
105
110
115
120
125
130
†
†
*
*
†
†
††
†
‡‡
‡‡
Co
Genistein Daidzein
Alzheimer's disease
1 3 5 8 10 1 3 5 8 10 1 3 5 8 10 µM
Fig. 3. Effect of 1–10 lM 17b-oestradiol, genistein and daidzein on the [35S]-GTPcS binding in the female control (Co) and Alzheimer’s disease (AD) frontocor-
tical membranes. 100% corresponds to the basal [35S]-GTPcS binding (means � SEM.). In both Co and AD, four to five independent experiments with the indi-
vidual brains were conducted; *P < 0.05 for Co frontal cortex versus AD frontal cortex; comparison of the stimulated [35S]-GTPcS binding with the basal level:
P < 0.05 in Co (�) and AD (�) frontal cortex.
590 V. Jefremov et al.
ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 20, 587–596

cantly (P < 0.05) higher 17b-oestradiol stimulation of G-proteins
than male subjects, whereas the effect of phytoesterogens showed
no gender difference (Fig. 4).
Protective effect of 17b-oestradiol, genistein and daidzeinon the oxymodified G-proteins in the frontocorticalmembranes
Figure 5(A, B) demonstrates suppressing (antioxidative) effects of the
investigated compounds in 10 lM concentration on the stimulation
of G-proteins by the metabolic pro-oxidants H2O2 and Hcy in con-
trol and AD frontal cortex membranes. In the control region, each
of the compounds significantly (P < 0.05) decreased the 25% stim-
ulation of [35S]-GTPcS binding induced by H2O2 and Hcy. 17b-Oes-
tradiol revealed a notably greater antioxidant effect on the H2O2
and Hcy stimulated activity of G-proteins than that of phytoestro-
gens. In AD frontal cortex, the antioxidant effect of 17b-oestradiol
and genistein towards the H2O2 and Hcy stimulation was less pro-
nounced than in the control region, whereas daidzein revealed no
significant protection against the pro-oxidants.
Effect of 17b-oestradiol on the adenylate cyclase activityin the frontocortical membranes
To understand the downstream signalling events induced by oestro-
gens, we investigated the effect of 1–30 lM of 17b-oestradiol on
the adenylate cyclase activity in the membranes of aged control
and AD frontal cortex (Fig. 6). In the control membranes, 17b-oes-
tradiol produced dose-dependent stimulation of adenylate cyclase,
reaching a maximum (plateau) value of 20% at the 20–30 lM con-
centration. In AD, the maximal stimulation of the enzyme was
remarkably lower than in control. Interestingly, the basal adenylate
cyclase activity in AD frontal cortex revealed a significantly
(P < 0.05) lower value (34 � 3 pmol cyclic AMP ⁄ min ⁄ mg protein)
90
100
110
120
130 Gen †*
† ††
†
‡
‡‡
‡
Daidz
Co
[35S]
GTP
S b
ound
, % o
f ba
sal
90
110
120
130
100
Gen
*Daidz
AD
Fig. 4. Effect of 10 lM 17b-oestradiol (17bE), genistein (Gen) and daidzein (Daidz) on the [35S]-GTPcS binding in the female ($) and male (#) frontocortical
membranes from control (Co) and Alzheimer’s disease (AD) brain. 100% corresponds to the basal [35S]-GTPcS binding (means � SEM). In each group, four
independent experiments with the individual brains were conducted; *P < 0.05 for female versus male values; comparison of the stimulated [35S]-GTPcS bind-
ing with the basal level: P < 0.05 in Co (�) and in AD (�).
100
105
110
115
120
125
130
H2O2 + Gen + Daidz 100
105
110
115
120
125
130
+ Daidz + Gen Hcy
[35S]
GTP
S b
indi
ng, %
of
basa
l
[35S]
GTP
S b
indi
ng, %
of
basa
l
H2O2 Co
Hcy Co
Hcy AD
*
*
*
**
**
**
*
H2O2 AD (A) (B)
Fig. 5. Effect of 10 lM 17b-oestradiol (17bE), genistein (Gen) and daidzein (Daidz) on the [35S]-GTPcS binding to control (Co) and Alzheimer’s disease (AD)
frontocortical membranes after the 5-min pretreatment with 10 lM H2O2 at 30 �C (A) or 500 lM homocysteine (Hcy) at 26 �C (B). 100% = basal [35S]-GTPcS
binding (means � SEM). In both, Co and AD, four independent experiments with the individual brains were conducted; *P < 0.05 between the effects of H2O2
or Hcy in the absence and presence of 17bE, Gen or Daidz.
17b-oestradiol stimulation of G-proteins in human brain 591
ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 20, 587–596

than in control (46 � 4). The fact that the nonhydrolyzable G-pro-
tein blocker GDPbS significantly reduced the stimulatory effect of
20 lM 17b-oestradiol on the adenylate cyclase in the control mem-
branes (Fig. 7) indicates that G-proteins are directly implicated in
the stimulation. Unlike 17b-oestradiol, the phytoestrogen genistein
revealed no statistically significant effect on the adenylate cyclase
activity, either in control or AD brain membranes (data not shown).
Effect of 17b-oestradiol, genistein and daidzein on[35S]-GTPcS binding in the membranes of Sf9 cellsexpressing different types of G-proteins
To elucidate the type(s) of G-proteins that might be influenced by
17b-oestradiol and phytoestrogens, the effect of 10 lM agents was
studied in the plasma membranes of Sf9 cells expressing Gas, Gao,
Ga11 or Gai1 together with Gb1c2 (Table 1). In the membranes of
Gasb1c2 expressing cells, 17b-oestradiol increased the basal [35S]-
GTPcS binding by 18% whereas genistein and daidzein slightly
decreased the binding (by 13% and 15%, respectively). In the mem-
branes of Gaob1c2 cells, 17b-oestradiol stimulated the radioligand
binding by 20% whereas the effect of phytoestrogens was negligi-
ble. In the membranes of Ga11b1c2 cells, only phytoestrogens could
significantly increase the radioligand binding. Interestingly, in the
membranes of Gaib1c2i cells, the investigated compounds did not
exert a significant effect on [35S]-GTPcS binding.
OH• and DPPH• scavenging activity of 17b-oestradiol,genistein and daidzein
The investigated compounds concentration-dependently scavenged
the OH• and DPPH• generated in the respective cell-free systems
in vitro. The maximum OH• scavenging activity (Fig. 8) of daidzein
and genistein (84 � 5% and 54 � 3%, respectively) was higher
than that of 17b-oestradiol (13 � 3%). Similarly, the maximum
DPPH• scavenging activity (Fig. 9) of genistein and daidzein
(12 � 0.9 and 8.3 � 0.8%, respectively) was higher than the activ-
ity of 17b-oestradiol (6.1 � 0.5%).
Discussion
The neuroprotective activities of oestrogens and their plant derived
analogs appear to be controlled by a wide range of mechanisms,
most being related to their antioxidant properties (3, 45). Various
models were used to specify the neuroprotective mechanism for
the nano- to micromolar concentrations of oestrogens (16, 32, 46).
However, this study is the first to elucidate whether micromolar
concentrations of 17b-oestradiol and dietary phytoestrogens (geni-
stein, daidzein) affect the G-proteins and adenylate cyclase in the
aged and AD human brain membranes. In parallel, the antioxidant
properties of micromolar concentrations of the agents were tested
in different systems.
We found a stimulatory effect of 5–10 lM 17b-oestradiol and
phytoestrogens on [35S]-GTPcS binding to the post-mortem hip-
pocampal and frontocortical membranes of aged women, which
suggests that G-proteins are involved in the transduction of oes-
trogen signals. Although the effect was not impressive, it might
prove to be an important mechanism for the amplifying neuro-
protective cascade of oestrogens in the living brain. In fact,
10 lM 17b-oestradiol stimulated the Gs- and Go-proteins and
the 10 lM phytoestrogens stimulated the G11-proteins
expressed in Sf9 cells (Table 1). In addition, 17b-oestradiol acti-
vated the Gs- and Gq ⁄ 11-proteins via the membrane ER,
expressed in the recombinant cells (47, 48). These data, together
with the fact that hippocampus and frontal cortex of aged and
AD human brain express Gs-, Go- and Gq ⁄ 11-proteins (49–51),
suggest that 17b-oestradiol may stimulate the three types of
G-proteins, and the pytoestrogens stimulate the G11-proteins
in these regions.
100
105
110
115
120
125
1 5 10 20 30
Co
AD
µM
Ade
nyla
te c
ycla
se a
ctiv
ity,
% o
f ba
sal
**
*
*
Fig. 6. Effect of 1–30 lM 17b-oestradiol on the adenylate cyclase activity in
the female control (Co) and Alzheimer’s disease (AD) frontocortical
membranes. 100% corresponds to the basal adenylate cyclase activity
(means � SEM). Six and five independent experiments with the individual
brains were conducted in Co and AD, repectively; *P < 0.05 for Co versus AD.
Fig. 7. Effect of 20 lM 17b-oestradiol (17bE) on the adenylate cyclase
activity in the female control frontocortical membranes treated with 100 lM
of GDPbS. 100% corresponds to the basal adenylate cyclase activity
(means � SEM). Four independent experiments with the individual brains
were conducted; *P < 0.05 for 17bE versus GDPbS + 17bE.
592 V. Jefremov et al.
ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 20, 587–596
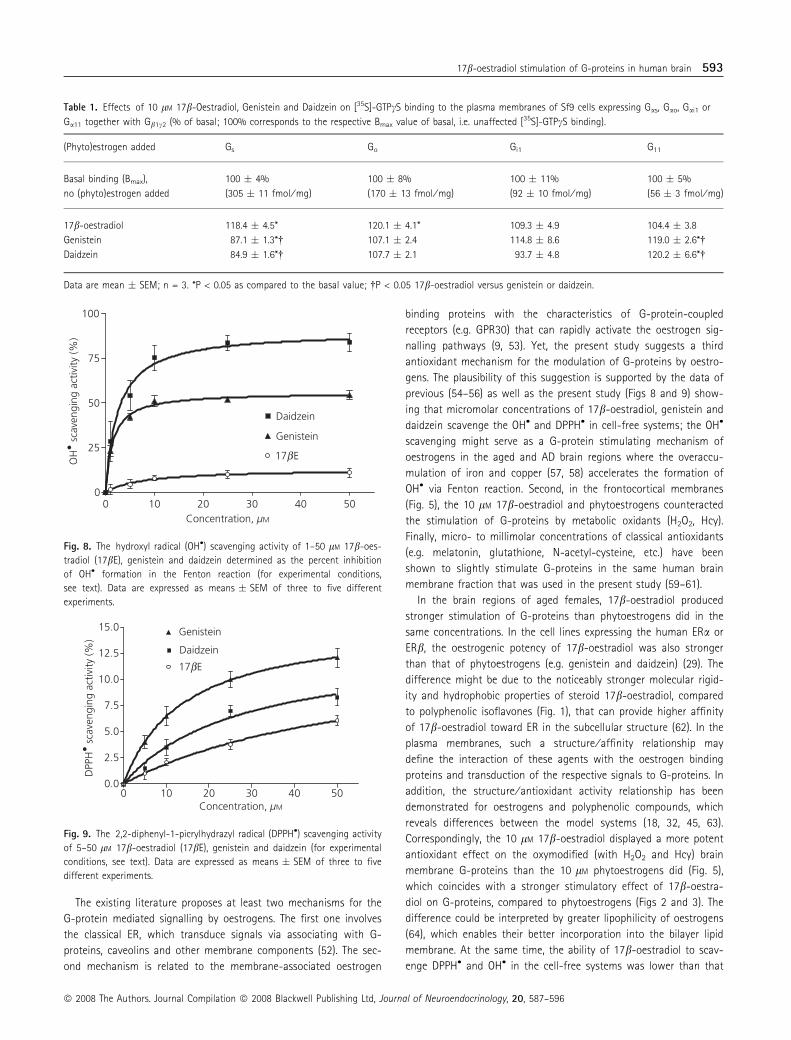
The existing literature proposes at least two mechanisms for the
G-protein mediated signalling by oestrogens. The first one involves
the classical ER, which transduce signals via associating with G-
proteins, caveolins and other membrane components (52). The sec-
ond mechanism is related to the membrane-associated oestrogen
binding proteins with the characteristics of G-protein-coupled
receptors (e.g. GPR30) that can rapidly activate the oestrogen sig-
nalling pathways (9, 53). Yet, the present study suggests a third
antioxidant mechanism for the modulation of G-proteins by oestro-
gens. The plausibility of this suggestion is supported by the data of
previous (54–56) as well as the present study (Figs 8 and 9) show-
ing that micromolar concentrations of 17b-oestradiol, genistein and
daidzein scavenge the OH• and DPPH• in cell-free systems; the OH•
scavenging might serve as a G-protein stimulating mechanism of
oestrogens in the aged and AD brain regions where the overaccu-
mulation of iron and copper (57, 58) accelerates the formation of
OH• via Fenton reaction. Second, in the frontocortical membranes
(Fig. 5), the 10 lM 17b-oestradiol and phytoestrogens counteracted
the stimulation of G-proteins by metabolic oxidants (H2O2, Hcy).
Finally, micro- to millimolar concentrations of classical antioxidants
(e.g. melatonin, glutathione, N-acetyl-cysteine, etc.) have been
shown to slightly stimulate G-proteins in the same human brain
membrane fraction that was used in the present study (59–61).
In the brain regions of aged females, 17b-oestradiol produced
stronger stimulation of G-proteins than phytoestrogens did in the
same concentrations. In the cell lines expressing the human ERa or
ERb, the oestrogenic potency of 17b-oestradiol was also stronger
than that of phytoestrogens (e.g. genistein and daidzein) (29). The
difference might be due to the noticeably stronger molecular rigid-
ity and hydrophobic properties of steroid 17b-oestradiol, compared
to polyphenolic isoflavones (Fig. 1), that can provide higher affinity
of 17b-oestradiol toward ER in the subcellular structure (62). In the
plasma membranes, such a structure ⁄ affinity relationship may
define the interaction of these agents with the oestrogen binding
proteins and transduction of the respective signals to G-proteins. In
addition, the structure ⁄ antioxidant activity relationship has been
demonstrated for oestrogens and polyphenolic compounds, which
reveals differences between the model systems (18, 32, 45, 63).
Correspondingly, the 10 lM 17b-oestradiol displayed a more potent
antioxidant effect on the oxymodified (with H2O2 and Hcy) brain
membrane G-proteins than the 10 lM phytoestrogens did (Fig. 5),
which coincides with a stronger stimulatory effect of 17b-oestra-
diol on G-proteins, compared to phytoestrogens (Figs 2 and 3). The
difference could be interpreted by greater lipophilicity of oestrogens
(64), which enables their better incorporation into the bilayer lipid
membrane. At the same time, the ability of 17b-oestradiol to scav-
enge DPPH• and OH• in the cell-free systems was lower than that
Table 1. Effects of 10 lM 17b-Oestradiol, Genistein and Daidzein on [35S]-GTPcS binding to the plasma membranes of Sf9 cells expressing Gas, Gao, Gai1 or
Ga11 together with Gb1c2 (% of basal; 100% corresponds to the respective Bmax value of basal, i.e. unaffected [35S]-GTPcS binding).
(Phyto)estrogen added Gs Go Gi1 G11
Basal binding (Bmax),
no (phyto)estrogen added
100 � 4%
(305 � 11 fmol ⁄ mg)
100 � 8%
(170 � 13 fmol ⁄ mg)
100 � 11%
(92 � 10 fmol ⁄ mg)
100 � 5%
(56 � 3 fmol ⁄ mg)
17b-oestradiol 118.4 � 4.5* 120.1 � 4.1* 109.3 � 4.9 104.4 � 3.8
Genistein 87.1 � 1.3*� 107.1 � 2.4 114.8 � 8.6 119.0 � 2.6*�
Daidzein 84.9 � 1.6*� 107.7 � 2.1 93.7 � 4.8 120.2 � 6.6*�
Data are mean � SEM; n = 3. *P < 0.05 as compared to the basal value; �P < 0.05 17b-oestradiol versus genistein or daidzein.
0 10 20 30 40 50 0
25
50
75
100
Daidzein
Genistein
17bE
Concentration, µM
OH
• sc
aven
ging
act
ivity
(%)
Fig. 8. The hydroxyl radical (OH•) scavenging activity of 1–50 lM 17b-oes-
tradiol (17bE), genistein and daidzein determined as the percent inhibition
of OH• formation in the Fenton reaction (for experimental conditions,
see text). Data are expressed as means � SEM of three to five different
experiments.
Fig. 9. The 2,2-diphenyl-1-picrylhydrazyl radical (DPPH•) scavenging activity
of 5–50 lM 17b-oestradiol (17bE), genistein and daidzein (for experimental
conditions, see text). Data are expressed as means � SEM of three to five
different experiments.
17b-oestradiol stimulation of G-proteins in human brain 593
ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 20, 587–596

of phytoestrogens (Figs 8 and 9). Such a discrepancy in the ranking
of antioxidant potency of the compounds might be due to the
model system-induced changes in the polarity and phenoxyl radical
stability of 17b-oestradiol and phytoestrogens, as previously dem-
onstrated for phenol and catechol oestrogens (65).
In the brain regions of AD females, the 10 lM 17b-oestradiol
stimulated G-proteins less than it did in the control regions. Simi-
larly, the decrease in the stimulation of G-proteins by glutathione,
melatonin and other antioxidants was shown in AD-injured brain
regions (59–61). These data, together with reports on the excessive
oxidation of proteins in the AD brain (66–68), suggest that one rea-
son for the decrease in 17b-oestradiol stimulation of G-proteins
might be oxymodification of their redox active functional groups.
Next, the decrease in the level of Go- and Gq11 proteins (49, 51)
and functional impairment of Gs-proteins (69) in the AD brain
might reduce the stimulatory response of G-proteins to 17b-oestra-
diol. Finally, local changes in membrane ERa splicing in the AD
brain (70) may diminish the ERa-mediated oestrogen stimulation of
G-proteins.
The gender difference in the 10 lM 17b-oestradiol action on
G-proteins in both, aged and AD frontocortical membranes (Fig. 4),
might be explained by lower production of oestrogens and expres-
sion of oestrogen binding proteins as well as of the respective
co-activators in the male brain membranes compared to those
in the female (71, 72). In addition, the characteristic effects of
oestrogens on the isolated plasma membranes and matrix vesicles
were shown to depend on the gender-specific composition of
membranes and metabolism of their components (particularly of
lipids) (73, 74), which provides another explanation for the gender-
specific 17b-oestradiol stimulation of G-proteins. The fact that
phytoestrogens have lower lipophilicity and incorporate into the
membrane bilayer less potently than classical oestrogens (64)
might help to explain why 10 lM genistein and daidzein, unlike
17b-oestradiol, revealed no gender-specific stimulation of mem-
brane-associated G-proteins.
Several studies have shown the stimulatory effect of nanomolar
concentrations of oestrogens on adenylate cyclase in the non-neuro-
nal cell lines (8, 9, 53). The effect was mediated by the transmem-
brane oestrogen receptor GPR30 and the coupled Gs-protein.
Supportively, 17b-oestradiol significantly potentiated [35S]-GTPcS
binding to the Gs-enriched Sf9 cell membranes (Table 1) and the
G-protein blocker GDPbS remarkably reduced 17b-oestradiol-induced
stimulation of adenylate cyclase in the frontocortical membranes of
aged females (Fig. 7). However, only micromolar concentrations of
17b-oestradiol could stimulate adenylate cyclase in the frontocortical
membranes (Fig. 6). Also, micro- to millimolar concentrations of clas-
sical antioxidants stimulate adenylate cyclase in the brain membranes
(61, 75). These data suggest that oestrogen stimulation of adenylate
cyclase in the brain membranes is mediated at least by two mecha-
nisms: at the nanomolar level, oestrogens stimulate the enzyme via
membrane receptor Gs-protein pathway and, at the micromolar level,
oestrogens may behave as antioxidants that modify the redox active
groups of the enzyme. Based on the recent hypotheses (1–3), both
of these mechanisms might be involved in the downstream neuro-
protective signalling of oestrogens.
In the frontal cortex of AD females, 17b-oestradiol stimulation
of adenylate cyclase was markedly lower than in control group
(Fig. 6). Such a decline in the effect could be explained by reduction
of adenylate cyclase activity and antioxidant sensitivity in the AD
brain (50, 61, 76). In addition, the weakened interaction between
Gs-proteins and adenylate cyclase in AD-injured brain areas (49, 77)
might serve as a causative factor that reduces the adenylate
cyclase stimulatory response to 17b-oestradiol.
The present study is the first to demonstrate that micromolar
concentrations of 17b-oestradiol and dietary phytoestrogens (geni-
stein, daidzein) stimulate G-proteins in the membranes of aged
and AD human brain and that 17b-oestradiol and phytoestrogens
have similarities and differences in this respect. We suggest that,
besides the ER-dependent one, the ER-independent antioxidant
mechanism is responsible for the stimulation of brain membrane
G-proteins by higher concentrations of oestrogens. Both of these
mechanisms could be involved in the G-protein mediated neuro-
protective signalling of oestrogens that contributes to their
preventive ⁄ therapeutic action against postmenopausal neurological
disorders.
Acknowledgements
We sincerely thank Dr Tatsuya Haga (University of Tokyo, Japan) for the
baculovirus tranfer vectors carrying the genes of G-proteins and Dr Kulliki
Saar for her advice and support. Likewise, our thanks are due to Dr Gerald
Finking (Institute of Occupational, Social and Environmental Medicine,
University of Ulm, Germany) for helpful discussions and to Mrs Inga
Volkman (Huddinge Brain Bank) for her excellent technical support. The
project was supported by The Estonian Scientific Foundation (Grants No
5222 and 6574) and the Swedish Foundation of Dementia.
Received: 22 August 2007,
revised 9 January 2008,
accepted 1 February 2008
References
1 Manthey D, Behl C. From structural biochemistry to expression profiling:
neuroprotective activities of estrogen. Neuroscience 2006; 138: 845–
850.
2 Amantea D, Russo R, Bagetta C, Corasaniti MT. From clinical evidence to
molecular mechanisms underlying neuroprotection afforded by estro-
gens. Pharmacol Res 2005; 52: 119–132.
3 Behl C. Oestrogen as a neuroprotective hormone. Nat Rev Neurosci
2002; 3: 433–442.
4 Miller NR, Jover T, Cohen HW, Zukin S, Etgen M. Etrogen can act via
estrogen receptor a and b to protect hipppocampal neurons against glo-
bal ischemia-induced cell death. Endocrinology 2005; 146: 3070–3079.
5 Levin ER. Cellular functions of plasma membrane estrogen receptors.
Steroids 2002; 67: 471–475.
6 Cato AC, Nesti A, Mink S. Rapid actions of steroid receptors in cellular
signalling pathways. Sci STKE 2002; 138: RE9.
7 Rosner W, Hryb DJ, Khan MS, Nakhla AM, Romas NA. Androgen and
estrogen signaling at the cell membrane via G-proteins and cyclic aden-
osine monophospohate. Steroids 1999; 64: 100–106.
8 Filardo EJ, Quinn JA, Frackelton AR Jr, Bland KI. Estrogen action via the
G protein coupled receptor, GPR30: stimulation of adenylyl cyclase and
594 V. Jefremov et al.
ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 20, 587–596

cAMP-mediated attenuation of the epidermal growth factor receptor-to-
MAPK signaling axis. Mol Endocrinol 2002; 16: 70–84.
9 Thomas P, Pang Y, Filardo J, Dong J. Identity of an estrogen membrane
receptor coupled to a G-protein in human breast cancer cells. Endocri-
nology 2005; 146: 624–632.
10 Sylvia VL, Boyan BD, Dean DD, Schwartz Z. The membrane effects of
17b-estradiol on chondrocyte phenotypic expression are mediated by
activation of protein kinase C through phospholipase C and G-proteins.
J Steroid Biochem Mol Biol 2000; 73: 211–224.
11 Toran-Allerand CD, Singh M, Setalo G Jr. Novel mechanisms of estrogen
action in the brain: new players in an old story. Front Neuroendocrinol
1999; 20: 97–121.
12 Whiting KP, Restall CJ, Brain PF. Steroid-hormone induced effects on
membrane fluidity and their potential role in non-genomic mechanisms.
Life Sci 2000; 67: 743–757.
13 Kelly MJ, Qiu J, Rønnekleiv OK. Estrogen modulation of G-protein-cou-
pled receptor activation of potassium channels in the central nervous
system. Ann NY Acad Sci 2003; 1007: 6–16.
14 Zhao L, Chen S, MingWang J, Brinton RD. 17b-estradiol induces Ca2+
influx, dendritic and nuclear Ca2+ rise and subsequent cyclic AMP
response element-binding protein activation in hippocampal neurons: a
potential initiation mechanism for estrogen neurotrophism. Neuroscience
2005; 132: 299–311.
15 Suzuki S, Brown CM, Wise PM. Mechanisms of neuroprotection by estro-
gen. Endocr 2006; 29: 209–215.
16 Behl C, Skutella T, Lezoualc’h F, Post A, Widman M, Newton CJ, Holsboer
F. Neuroprotection against oxidative stress by estrogens: structure-activ-
ity relationship. Mol Pharmacol 1997; 51: 535–541.
17 Green PS, Gordon K, Simpkins JW. Phenolic A ring requirement for neu-
roprotective effects of steroids. J Steroid Biochem Mol Biol 1997; 63:
229–235.
18 Prokai L, Simpkins JW. Structure-nongenomic neuroprotection relation-
ship of estrogens and estrogen-derived compounds. Pharmacol Ther
2007; 114: 1–12.
19 Bicknell RJ. Sex-steroid actions on neurotransmission. Curr Opin Neurol
1998; 11: 667–671.
20 Boulware MI, Mermelstein PG. The influence of estradiol on nervous sys-
tem function. Drug News Perspect 2005; 18: 631–637.
21 Keller JN, Germeyer A, Begley JB, Mattson MP. 17b-estradiol attenuates
oxidative impairment of synaptic Na+ ⁄ K+-ATPase activity, glucose trans-
port, and glutamate transport induced by amyloid b-peptide and iron.
J Neurosci Res 1997; 50: 522–530.
22 Sawada H, Ibi M, Kihara T, Urushitani M, Akaike A, Shimohama S. Estra-
diol protects mesencephalic dopaminergic neurons from oxidative stress-
induced neuronal death. J Neurosci Res 1998; 54: 707–719.
23 Sribnick EA, Ray SK, Nowak MW, Li L, Banik NL. 17b-estradiol
attenuates glutamate-induced apoptosis and preserves electrophysiolog-
ic function in primary cortical neurons. J Neurosci Res 2004; 76: 688–
696.
24 Simpkins JW, Rajakumar G, Zhang YQ, Simpkins CE, Greenwald D, Yu CJ,
Bodor N, Day AL. Estrogens may reduce mortality and ischemic damage
caused by middle cerebral artery occlusion in the female rat. J Neuro-
surg 1997; 87: 724–730.
25 Petanceska SS, Nagy V, Frail D, Gandy S. Ovariectomy and 17b-estradiol
modulate the levels of Alzheimer’s amyloid b peptides in brain. Exp Ger-
ontol 2000; 35: 1317–1325.
26 Dhandapani KM, Brann DW. Protective effects of estrogen and selective
estrogen receptor modulators in the brain. Biol Reprod 2002; 67: 1379–
1385.
27 Simpkins JW, Yang SH, Wen Y, Singh M. Estrogens, progestins, meno-
pause and neurodegeneration: basic and clinical studies. Cell Mol Life
Sci 2005; 62: 271–280.
28 Gooren LJ, Toorians AW. Significance of oestrogens in male (patho)phys-
iology. Ann Endocrinol 2003; 64: 126–135.
29 Kuiper GG, Lemmen JG, Carlsson B, Corton JC, Safe SH, van der Saag PT,
van der Burg B, Gustafsson J-A. Interaction of estrogenic chemicals and
phytoestrogens with estrogen receptor. Endocrinology 1998; 139: 4252–
4263.
30 Duncan AM, Phipps WR, Kurzer MS. Phyto-estrogens. Best Pract Res Clin
Endocrinol Metab 2003; 17: 253–271.
31 Zhao L, Chen Q, Diaz Brinton R. Neuroprotective and neurotrophic effi-
cacy of phytoestrogens in cultured hippocampal neurons. Exp Biol Med
2002; 227: 509–519.
32 Mitchell JH, Gardner PT, McPhail DB, Morrice PC, Collins AR, Duthie GG.
Antioxidant efficacy of phytoestrogens in chemical and biological model
systems. Arch Biochem Biophys 1998; 360: 142–146.
33 Ramassamy C. Emerging role of ployphenolic compounds in the treat-
ment of neurodegenerative diseases: a review of their intracellular tar-
gets. Eur J Pharmacol 2006; 545: 51–64.
34 Simic G, Kostovic I, Winblad B, Bogdanovic N. Volume and number of
neurons of the human hippocampal formation in normal aging and Alz-
heimer’s disease. J Comp Neurol 1997; 379: 482–484.
35 Braak H, Braak E, Bohl J, Bratzke H. Evolution of Alzheimer’s disease
related cortical lesions. J Neural Transm Suppl 1998; 54: 97–106.
36 American Psychiatric Association . Diagnostic and Statistical Manual of
Mental Disorders, 4th edn. Washington, DC: American Psychiatric Associ-
ation, 1994.
37 Bogdanovic N, Morris JC. Diagnostic criteria for Alzheimer’s disease in
multicentre brain banking. In: Cruz-Sanchez FF, Ravid R, Cuziner ML,
eds. Neuropathological Diagnostic Criteria for Brain Banking, Biomedical
and Health Research Series, Vol. 10. Amsterdam: IOS Press, 1985: 20–29.
38 Mirra SS, Heyman A, McKeel D. The Consortium to establish a registry
for Alzheimer’s Disease (CERAD). Part II. Standardization of the neuropa-
thologic assessment of Alzheimer’s disease. Neurology 1991; 41: 479–
486.
39 Karelson E, Laasik J, Sillard R. Regulation of adenylate cyclase by galanin,
neuropeptide Y, secretin and vasoactive intestinal polypeptide in rat
frontal cortex, hippocampus and hypothalamus. Neuropeptides 1995;
28: 21–28.
40 Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement
with the Folin phenol reagent. J Biol Chem 1951; 93: 265–275.
41 Rezaei K, Saar K, Soomets U, Valkna A, Nasman J, Zorko M, Akerman K,
Schroeder T, Bartfai T, Langel U. Role of third intracellular loop of gala-
nin receptor type 1 in signal transduction. Neuropeptides 2000; 34: 25–
31.
42 Brown BL, Ekins RP, Albano JDM. Saturation assay for cyclic AMP using
endogenous binding protein. Adv Cyclic Nucleotide Res 1972; 2: 25–40.
43 Barreto JC, Smith GS, Strobel NH, McQuillin PA, Miller TA. Terephthalic
acid: a dosimeter for the detection of hydroxyl radicals in vitro. Life Sci
1995; 56: PL89–PL96.
44 Blois MS. Antioxidant determinations by the use of stable free radical.
Nature 1958; 181: 1199–2000.
45 Lee YB, Lee HJ, Sohn HS. Soy isoflavones and cognitive function. J Nutr
Biochem 2005; 16: 641–649.
46 Rufer CE, Kulling SE. Antioxidant activity of isoflavones and their major
metabolites using different in vitro assays. J Agric Food Chem 2006; 54:
2926–2931.
47 Razandi M, Pedram A, Greene GL, Levin ER. Cell membrane and nuclear
estrogen receptors (ERs) originate from a single transcript: studies of
ERa and ERb expressed in Chinese hamster ovary cells. Mol Endocrinol
1999; 13: 307–319.
48 Razandi M, Pedram A, Park ST, Levin ER. Proximal events in signalling by
plasma membrane estrogen receptors. J Biol Chem 2003; 278: 2701–
2712.
17b-oestradiol stimulation of G-proteins in human brain 595
ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 20, 587–596

49 O’Neill C, Wiehager B, Fowler CJ, Ravid R, Winblad B, Cowburn RF.
Regionally selective alterations in G-protein subunit levels in the Alzhei-
mer’s disease brain. Brain Res 1994; 636: 193–201.
50 Ross BM, McLaughlin M, Roberts M, Milligan G, McCulloch J, Knowler JT.
Alterations in the activity of adenylate cyclase and high affinity GTPase
in Alzheimer’s disease. Brain Res 1993; 622: 35–42.
51 Kelly JF, Storie K, Skamra C, Bienias J, Beck T, Bennett DA. Relation-
ship between Alzheimer’s disease clinical stage and Gq ⁄ 11 in sub-
cellular fractions of frontal cortex. J Neural Transm 2005; 112:
1049–1056.
52 Hewitt SC, Deroo BJ, Korach KS. A new mediator for an old hormone.
Science 2005; 307: 1572–1573.
53 Thomas P, Dong J. Binding and activation of the seven-transmembrane
estrogen receptor GPR30 by environmental estrogens: a potential novel
mechanism of endocrine disruption. J Steroid Biochem Mol Biol 2006;
102: 175–179.
54 Ayres S, Abplanalp W, Liu JH, Ravi Subbiah MT. Mechanisms involved in
the protective effect of estradiol-17b on lipid peroxidation and DNA
damage. Am J Physiol Endocrinol Metab 1998; 274: 1002–1008.
55 Wenly Y, Yaping Z, Bo S. The radical scavenging activities of radix
puerariae isoflavonoids: a chemiluminescence study. Food Chem 2004;
86: 525–529.
56 Lee CH, Yang L, Xu JZ, Yeung SYV, Huang Y, Chen ZY. Relative antioxi-
dant activity of soybean isoflavones and their glycosides. Food Chem
2005; 90: 735–741.
57 Castellani RJ, Honda K, Zhu X, Cash AD, Nunomura A, Perry G, Smith
MA. Contribution of redox active iron and copper to oxidative damage
in Alzheimer’s disease. Ageing Res Rev 2004; 3: 319–326.
58 Zhu X, Su B, Wang X, Smith MA, Perry G. Causes of oxidative stress in
Alzheimer’s disease. Cell Mol Life Sci 2007; 64: 2202–2210.
59 Karelson E, Fernaeus S, Reis K, Bogdanovic N, Land T. Stimulation of G-
proteins in human control and Alzheimer’s disease brain by FAD mutants
of APP714-723: implication of oxidative mechanisms. J Neurosci Res 2005;
79: 368–374.
60 Reis K, Zharkovsky A, Bogdanovic N, Karelson E, Land T. Critical
role of methionin-722 in the stimulation of human brain G-pro-
teins and neurotoxicity induced by London familial Alzheimer’s
disease (FAD) mutated V717G-APP714-723. Neuroscience 2007; 144:
571–578.
61 Karelson E, Mahlapuu R, Zilmer M, Soomets U, Bogdanovic N, Langel U.
Possible signalling by glutathione and its novel analogue through potent
stimulation of frontocortical G proteins in normal aging and in Alzhei-
mer’s disease. Ann NY Acad Sci 2002; 973: 537–540.
62 Turner JV, Agatanovic-Kustrin S, Glass BD. Molecular aspects of phytoes-
trogen selective binding at estrogen receptors. J Pharm Sci 2007; 96:
1879–1885.
63 Moosmann B, Behl C. The antioxidant neuroprotective effects of estro-
gens and phenolic compounds are independent from their estrogenic
properties. Proc Natl Acad Sci USA 1999; 96: 8867–8872.
64 Cunningham AR, Klopmnan G, Rosenkranz HS. A dichotomy in the lipo-
philicity of natural estrogens, xenoestrogens and phytoestrogens. Envi-
ron Health Perspect 1997; 105: 665–668.
65 Ruiz-Larrea MB, Martın C, Martınez R, Navarro R, Lacort M, Miller NJ.
Antioxidant activities of estrogens against aqueous and lipophilic radi-
cals: differences between phenol and catechol estrogens. Chem Phys
Lipids 2000; 105: 179–188.
66 Butterfield DA, Perluigi M, Sultana R. Oxidative stress in Alzheimer’s dis-
ease brain: new insights from redox proteomics. Eur J Pharmacol 2006;
545: 39–50.
67 Newman SF, Sultana R, Perluigi M, Coccia R, Cai J, Pierce WM, Klein JB,
Turner DM, Butterfield DA. An increase in S-glutathionylated proteins in
the Alzheimer’s disease inferior parietal lobule, a proteomics approach.
J Neurosci Res 2007; 85: 1506–1514.
68 Bogdanovic N, Zilmer M, Zilmer K, Rehema A, Karelson E. The Swedish
APP670 ⁄ 671 Alzheimer’s disease mutation: the first evidence for strik-
ingly increased oxidative injury in the temporal inferior cortex. Dement
Geriatr Cogn Disord 2001; 12: 364–370.
69 Hashimoto E, Ozawa H, Saito T, Gsell W, Takahata N, Riederer P, Frolich
L. Impairment of Gsa function in human brain cortex of Alzheimetr’s dis-
ease: comparison with normal aging. J Neural Transm 2004; 111: 311–
322.
70 Ishunina TA, Swaab DF. Age-dependent ERa MB1 splice variant expres-
sion in discrete areas of the human brain. Neurobiol Aging 2007 [Epub
ahead of print].
71 Tischkau SA, Ramirez VD. A specific membrane binding protein for pro-
gesterone in rat brain: sex differences and induction by estrogen. Proc
Natl Acad Sci USA 1993; 90: 1285–1289.
72 Balthazart J, Baillien M, Charlier TD, Cornil CA, Ball GF. Multiple mecha-
nisms control brain aromatase activity at the genomic and non-genomic
level. J Steroid Biochem Mol Biol 2003; 86: 367–379.
73 Schwartz Z, Gates PA, Nasatzky E, Sylvia VL, Mendez J, Dean DD, Boyan
BD. Effect of 17b-estradiol on chondrocyte membrane fluidity and phos-
pholipid metabolism is membrane-specific, sex-specific, and cell matura-
tion-dependent. Biochim Biophys Acta 1996; 1282: 1–10.
74 Schuessel K, Leutner S, Cairns NJ, Muller WE, Eckert A. Impact of gender
on upregulation of antioxidant defence mechanisms in Alzheimer’s dis-
ease brain. J Neural Transm 2004; 111: 1167–1182.
75 Soomets U, Mahlapuu R, Tehranian R, Jarvet J, Karelson E, Zilmer M,
Iverfeldt K, Zorko M, Graslund A, Langel U. Regulation of GTPase and
adenylate cyclase activity by amyloid b-peptide and its fragments in rat
brain tissue. Brain Res 1999; 850: 179–188.
76 Mahlapuu R, Viht K, Balaspiri L, Bogdanovic N, Saar K, Soomets U, Land
T, Zilmer M, Karelson E, Langel U. Amyloid precursor protein carboxy-
terminal fragments modulate G-proteins and adenylate cyclase activity
in Alzheimer’s disease brain. Brain Res Mol Brain Res 2003; 117: 73–82.
77 Cowburn RF, O’Neill C, Bonkale WL, Ohm TG, Fastbom J. Receptor-G-pro-
tein signalling in Alzheimer’s disease. Biochem Soc Symp 2001; 67:
163–175.
596 V. Jefremov et al.
ª 2008 The Authors. Journal Compilation ª 2008 Blackwell Publishing Ltd, Journal of Neuroendocrinology, 20, 587–596
Copyright © 2022 FDOKUMEN




















