
Characterization of benign and malignant prostate epithelial Hoechst 33342 side populations
13
The Prostate Characterization of Benign and Malignant Prostate Epithelial Hoechst 33342 Side Populations Mick D. Brown, 1 * Paul E. Gilmore, 1,2 Claire A. Hart, 1 Joanne D. Samuel, 1,2 Vijay A.C. Ramani, 3,4 Nicholas J. George, 3 and Noel W. Clarke 1,2,4 1 ProMPTGenito-Urinary Cancer Research Group, Paterson Institute for Cancer Research, University of Manchester, Manchester,UK 2 Department of Urology, Salford Royal Hospital NHS Trust, Salford,UK 3 Department of Urology, South Manchester University Hospital NHS Trust, Manchester,UK 4 Department of Urology,Christie Hospital NHS Trust,West Didsbury, Manchester,UK BACKGROUND. The prostate epithelial stem cell has been proposed as the primary origin of neoplastic change in prostate cancer. However, the isolation and characterization of unexpanded prostate epithelial stem cells have proven problematic. METHODS. A prostate epithelial side population (SP) has been isolated utilizing a modified Hoechst 33342 dye efflux assay from both benign and malignant prostate tissue. CD45 ve , integrin a2 þve Hoechst 33342 SP and NSP cells were isolated by FACS, immunophenotyped and functionally characterized in 3D culture. RESULTS. FACS analysis revealed a verapamil sensitive SP accounting for 0.93 0.12% and 0.57 0.11% of the total epithelial population from both benign and malignant prostates. The benign SP phenotype revealed a heterogeneous cell population consisting predominantly of small basal cells containing minimal cytoplasm. Conversely, the malignant SP was of undetermined acinar origin and with a complete loss of expression of the CDK2 inhibitor p21 WAF1/Cip1 . In vitro androgen-enhanced 3D culture of the benign and malignant SP cells led to the production of spheroids which had acinus like morphology and expressed primitive and basal cell markers. Incorporation of the CD133 marker isolated a further SP sub-fraction accounting for 0.037 0.01% of epithelial cells. CONCLUSIONS. Our observations are consistent with the Hoechst 33342 dye efflux assay isolating a stem cell enriched population which can be further sub-fractionated by CD133 selection. Moreover, the loss of the CDK inhibitor in malignancy is consistent with the hypothesis that neoplastic change originates in the stem cell compartment. Prostate # 2007 Wiley-Liss, Inc. KEY WORDS: prostate cancer; stem cells; SP; CD133 INTRODUCTION Benign prostatic hyperplasia (BPH) and prostate cancer (CaP) are major problems in males. BPH is present in the majority of men over 60 years and prostate cancer is one of the commonest male malig- nancies, resulting in 10,200 deaths in the UK each year [1]. Both abnormalities are characterized by deranged cellular proliferation and behavior. Stem cells are clonogenic, are capable of undergoing self-renewal and differentiation, and there is increasing evidence demonstrating that the root of aberrant cellular proliferation begins within stem cell popula- tions. Hemopoietic stem cells (HSCs) which have Grant sponsor: British Urological Foundation; Grant sponsor: The Fellows Fellowship of the Royal College of Surgeons of England. *Correspondence to: Mick D. Brown, ProMPT Genito-Urinary Cancer Research Group, Paterson Institute for Cancer Research, University of Manchester, Wilmslow Road, Manchester M20 4BX, UK. E-mail: [email protected] Received 15 January 2007; Accepted 11 May 2007 DOI 10.1002/pros.20620 Published online in Wiley InterScience (www.interscience.wiley.com). ȣ 2007 Wiley-Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Characterization of benign and malignant prostate epithelial Hoechst 33342 side populations

The Prostate
Characterizationof Benign andMalignant ProstateEpithelialHoechst 33342 Side Populations
Mick D. Brown,1* Paul E. Gilmore,1,2 Claire A. Hart,1 Joanne D. Samuel,1,2
Vijay A.C. Ramani,3,4 Nicholas J. George,3 and Noel W. Clarke1,2,4
1ProMPTGenito-UrinaryCancer ResearchGroup,Paterson Institute for Cancer Research,UniversityofManchester,Manchester,UK
2DepartmentofUrology, Salford RoyalHospitalNHSTrust, Salford,UK3DepartmentofUrology, SouthManchesterUniversityHospitalNHSTrust,Manchester,UK
4DepartmentofUrology,ChristieHospitalNHSTrust,WestDidsbury,Manchester,UK
BACKGROUND. The prostate epithelial stem cell has been proposed as the primary origin ofneoplastic change in prostate cancer. However, the isolation and characterization ofunexpanded prostate epithelial stem cells have proven problematic.METHODS. A prostate epithelial side population (SP) has been isolated utilizing amodified Hoechst 33342 dye efflux assay from both benign and malignant prostate tissue.CD45�ve, integrin a2þve Hoechst 33342 SP and NSP cells were isolated by FACS,immunophenotyped and functionally characterized in 3D culture.RESULTS. FACS analysis revealed a verapamil sensitive SP accounting for 0.93� 0.12% and0.57� 0.11% of the total epithelial population from both benign and malignant prostates. Thebenign SP phenotype revealed a heterogeneous cell population consisting predominantly ofsmall basal cells containing minimal cytoplasm. Conversely, the malignant SP was ofundetermined acinar origin and with a complete loss of expression of the CDK2 inhibitorp21WAF1/Cip1. In vitro androgen-enhanced 3Dculture of the benign andmalignant SP cells led tothe production of spheroids which had acinus like morphology and expressed primitive andbasal cell markers. Incorporation of the CD133 marker isolated a further SP sub-fractionaccounting for 0.037� 0.01% of epithelial cells.CONCLUSIONS. Our observations are consistent with the Hoechst 33342 dye efflux assayisolating a stem cell enriched population which can be further sub-fractionated by CD133selection. Moreover, the loss of the CDK inhibitor in malignancy is consistent withthe hypothesis that neoplastic change originates in the stem cell compartment. Prostate# 2007 Wiley-Liss, Inc.
KEY WORDS: prostate cancer; stem cells; SP; CD133
INTRODUCTION
Benign prostatic hyperplasia (BPH) and prostatecancer (CaP) are major problems in males. BPH ispresent in the majority of men over 60 years andprostate cancer is one of the commonest male malig-nancies, resulting in 10,200 deaths in the UKeach year [1]. Both abnormalities are characterizedby deranged cellular proliferation and behavior.Stem cells are clonogenic, are capable of undergoingself-renewal anddifferentiation, and there is increasingevidence demonstrating that the root of aberrant
cellular proliferation begins within stem cell popula-tions. Hemopoietic stem cells (HSCs) which have
Grant sponsor: British Urological Foundation; Grant sponsor: TheFellows Fellowship of the Royal College of Surgeons of England.
*Correspondence to: Mick D. Brown, ProMPT Genito-UrinaryCancer Research Group, Paterson Institute for Cancer Research,University of Manchester, Wilmslow Road, Manchester M20 4BX,UK. E-mail: [email protected] 15 January 2007; Accepted 11 May 2007DOI 10.1002/pros.20620Published online in Wiley InterScience(www.interscience.wiley.com).
� 2007Wiley-Liss, Inc.

accumulated a series of mutations have been shown tobe the fundamental cause of both human acutemyeloidand lymphoid leukemia [2–4]. There is also increasingevidence for the involvement of stem cells in solidcancers. Observations that solid tumors are typicallyheterogeneous and contain only a small number ofclonogenic cells [5,6] has recently been supported bythe isolation of putative cancer stem cells from breastcancer [7], glioblastoma [8], and melanoma [9].
Prostatic stem cells have been implicated in theetiology of prostate cancer and are believed to reside inthe basal layer of the prostate glandular acini [10].In vitro studies have substantiated this by demonstrat-ing that basal cells have enhanced powers of prolife-ration and survival. Isolation of an a2 integrin basal cellpopulation resulted in an enriched cell populationwitha capacity for prolonged clonal expansion and differ-entiation in vitro [11,12]. Physical isolation of a pureprostatic stem cell population has proven elusive. Anumber of markers have been proposed, for example,Prostate Stem Cell Antigen [13], Prostate MembraneAntigen [14], pp32 [15], and the basal marker integrina2 [11,16]: none have been proved to be tissue orlocation specific [17,18]. Richardson et al. [19] recentlyisolated a prostate epithelial cell (PEC) population,based on the co-expression of CD133 and a2b1, highlyenriched for cells exhibiting enhanced proliferativepotential and colony forming efficiency compared tounrefinedbasal cells andable to formprostate neo-aciniin culture.
Stemcell behavior rather thanmarker expressionhasshown promise, with populations of HSCs beingenriched amongst populations of cells which activelyefflux Hoechst 33342 [20–24]. These lineage�ve
CD34þve ‘‘side population’’ (SP) cells have demon-strated properties of stem cells enabling FACS isolationof normal blood cells from leukemic patients, allowingautologous transplantation [25–27]. Tissue homeostasisrequires the stem cell to differentiate and proliferatein response to changes in its environment or niche[28]. The proposed HSC expressing CD34, Thy-1, andSca-1 have shown a propensity for the generation andregeneration of both bone marrow and immunologicalsystems, although there is variable specificity ofproposed markers [29,30]. This technique has beentranslated for SP isolation from solid tissues such asbreast, kidney, lung, liver, heart, skin, brain, andskeletal muscle [31–35].
Previously, we have isolated a valid prostateepithelial SP from both benign and malignant tissue[36]. However, the cellular labeling techniquesrequired for this isolation method preclude compre-hensive immunohistochemical analysis. Here, wepresent a refined Hoechst 33342 technique enablingisolation of an enriched viable and unlabelled stem
cell population from both benign and malignantprostatic tissue and characterization of both benignand malignant prostate epithelial SP cells.
MATERIALSANDMETHODS
Materials
Tissue culture media and reagents were purchasedfrom Invitrogen (Paisley, UK) with the exception ofMetrizamide, Sigma Fast 2,3 diaminobenzidine perox-idase substrate kit, 30% hydrogen peroxidase, citricacid, Tween-20, and verapamil hydrochloride (V106)from Sigma-Aldrich (Poole, Dorset, UK); collagenasetype I and trypsin from Worthington, Lorne Laborato-ries Ltd (Twyford, UK); fetal calf serum (FCS) fromBioWhittaker Europe (Verviers, Belgium), cell cultureinserts andMatrigelTM fromBectonDickinson (NJ) andbovine serumalbumen fromPAALaboratories (Yeovil,UK). Vectastain ABC kit and VectorShield weresupplied by Vector Laboratories (CA); Hoechst 33342trihydrochloride and SYTO 61 by Molecular ProbesEurope (BV, Leiden, The Netherlands); and PervexMounting Medium by CellPath (Powys, Wales, UK).
Antibodies
Antibodies against pan-cytokeratin (clone C-11)cytokeratin 18 (clone CY90), cytokeratin 8 (cloneM20), cytokeratin 14 (Clone CKB1), p63, streptavidinPE-Cy5 (Quantum RedTM) and streptavidin-Cy3 werefrom Sigma-Aldrich. Anti cytokeratin 5, CD49b (a2integrin), CD45-FITC, and g1-IgG1a-FITC antibodieswere from BD Biosciences (San Jose, CA). Anti-epithelial Ber-EP4 (pure and FITC conjugate), prostatespecific antigen (ER-PR8), p21WAF1/Cip1, p27kip1 andbiotinylated secondary antibodies were supplied byDako Ltd (Cambridge, UK); mouse IgG1A and 2B, (pureand FITC conjugate) by R&D Systems (Abingdon, UK);Cleaved Notch, b-catenin and non-murine IgG byCell Signaling Technologies (Hitchin, Herts, UK);androgen receptor (AR) from Novocastra LaboratoriesLtd (Newcastle-upon-Tyne, UK), CD133-biotin fromMiltenyl Biotec Ltd (Bisley, Surrey, UK) and Notch1 (M-20) from Santa Cruz Biotechnology, Inc. (SantaCruz, CA). Musashi-1 (Msi-1) antibody courtesy of Dr.R Clarke, Breast Biology Research Group, Manchester,UK [37].
Cell Lines
The STO cell line was obtained from the ECACC(Porton Down, UK) and cultured in DMEM supple-mented with 10% FCS 2 mM L-glutamine ina humidified atmosphere at 378C and 5% CO2 [38].
The Prostate DOI 10.1002/pros
2 Brown et al.

Tissue Collection and Isolation of Epithelial Cells
All tissue was collected from consenting patientsundergoing trans-urethral resection of the prostate(TURP) for benign or malignant disease under localResearch Ethics Committee approval. TURP chippingswere bisected for independent histopathological anal-ysis and for tissue culture. Epithelial cells were isolatedas previously described by Lang et al. [39]. BrieflyTURP chips were diced and agitated overnight at 378Cin collagenase type I at 200U/ml inRPMI 1640mediumwith 2% serum.Thedigestwas agitated in 0.1% trypsin,1% BSA/1 mM EDTA in PBS for 15 min. The cellsuspension was washed in PBS 1% BSA/1 mM EDTAand resuspended in RPMI 1640 10% FCS. Fibroblastswere removed by differential centrifugation (360g,1 min without braking) prior to PEC isolation bycentrifugation over ametrizamide cushion (1.079 g/ml,800g, 15 min, 48C).
Hoechst 33342 Stainingof Prostate Epithelial Cells
Hoechst 33342 staining was modified from themethod described by Bhatt et al. [36]. Briefly, 1� 106
PEC in 1 ml Hoechst buffer (Hank’s buffered saltsolution, 10% FCS, 20 mM HEPES, and 1% D-glucose)were stained with 5 mM Hoechst 33342 for 90 min at378C with continuous agitation (5 Hz). Cells werewashed and resuspended in ice-cold Hoechst bufferprior to FACS analysis. Hoechst 33342 dye efflux SPwas defined by 50 mM verapamil hydrochloride block-adeofHoechst 33342dye efflux.Hoechst 33342 stainingwas detected using a Becton Dickinson FACS VantageSE flow cytometer (FACS), exciting at 357 nm anddetecting Hoechst Blue with a 424/44 broad pass (BP)filter and Hoechst Red with a 675/20 BP filter (OmegaOptical, Brattleboro VT).
PhenotypicAnalysis
Cells were immobilized on poly-L-lysine coatedslides and fixed with 4% paraformaldehyde prior topermeabilization with methanol at �208C followedby acetone at �208C. Cells were blocked with 10%serum and incubated overnight at 48C with primaryantibodies. Primary antibodies were then labeledwith biotinylated secondary antibodies and tertiarystreptavidin PE-Cy3 label. Samples were then co-labeled with Ber-EP4-FITC and stained with SYTO61(1 ml/ml of PBS) before mounting with VectaShieldwith DAPI.
Cells were viewed by confocal microscopy using aZeiss LSM510 based around an AxioVert 100M.SYTO61 was excited at 633 nm and streptavidin PE-Cy3 at 543 nm using a 1 mW helium neon laser, andFITC at 488 nm by a 25 mW argon laser. Z series datawere collected at 1 mm intervals.
Prostate Epithelial Spheroid Co-Culture
Prostate epithelial spheroids were cultured usingthe protocol defined by Lang et al. [40]. Briefly, viablecells (as determined by trypan blue exclusion) wereresuspended in keratinocyte serum free medium(KSFM) with supplied 50 mg/ml bovine pituitaryextract (BPE) and 5 ng/ml epithelial growth factor(EGF) supplements, 2 mM L-glutamine, 2% FCS,dihydrotestosterone (2� 10�7 M), and estradiol(10 ng/ml) at 2� 103 cells/ml and mixed on ice withMatrigel at a 1:1 ratio. Aliquots (0.25 ml) were platedout in 0.4 mmpore culture inserts and theMatrigel set at378C for 30 min. The inserts were co-culturedsuspended above a confluent layer of irradiatedSTO cells (50 gray) at 378C, 5% CO2 and media wasrefreshedevery 48hr. Brightfield spheroid imagesweretaken with an inverted Olympus BX51 microscope bySoft Imaging Systems (SIS) GmbH (Munster, Germany)and analyzed with Adobe Photoshop version 6.1(Adobe, San Jose, CA). Spheroids were fixed overnightat 48C in 4% formalin in PBS before embedding inparaffin.
Immunohistochemistry
Paraffin embedded sections were de-waxed inxylene for 5 min and rehydrated through gradedalcohols and water prior to citrate antigen retrieval.Sections were blocked with 10% serum and 0.3% H2O2
and stained with primary antibodies. Primary anti-bodies were labeled with biotinylated secondary anti-body and avidin DH/biotinylated horseradishperoxidase complex (VectastainEliteABCkit). Sectionswere developed by the addition of DAB substrate for5 min and counterstained with hematoxylin. Sectionswere viewed on an Olympus BX51 upright microscopeand processing of image data was carried out usingAdobe Photoshop 6.0.
RESULTS
Hoechst Prof|le of CD45�ve ProstateCells
We have previously shown that a basal epithelialcell Hoechst 33342 SP could be isolated from solidprostate tissue [36]. However, the isolation protocolused generated an SP population pre-labeled with Ber-EP4 and integrin a2 preventing further phenotyping.Comparison of the forward (FSC) and side scatter (SSC)plots (Fig. 1A) showed that the majority of cellsremoved by this protocol displayed similar FSC versusSSC plots to CD45þve lymphocytes. Primary prostatecell suspensions were labeled with anti CD45-FITCand the CD45þve cells gated out from the sort(Fig. 1B). Hoechst Red/Blue analysis of the CD45
The Prostate DOI 10.1002/pros
Prostate StemCell Isolation 3

depleted cell population generated similar verapamilsensitive SP profiles from both benign and malignanttissue with the SP accounting for 0.93� 0.12% (n¼ 18)and 0.57� 0.11% (n¼ 7) of the PEC, respectively. Boththe unlabelled SP and Non-SP (NSP) CD45�ve cellswere collected for phenotypic analysis (Fig. 1C).
Phenotype of SP andNSPPopulations
SP and NSP cell populations were stained forthe stem cell markers p21WAF1/Cip1, p27Kip1, b-catenin,Msi-1, Notch, Cleaved Notch and the differentiationmarkers cytokeratin k5, k8, k14, and k18, AR, PSA, andp63 (Fig. 2Bi). All cells were visualized by stainingwithSYTO61 and epithelial phenotype was confirmed bylabeling with Ber-EP4-FITC. Captured x, y, and z datawere imported into Imaris (Bitplane AG) where anisosurface was created, utilizing 3D thresholding,and applied to each cell (Fig. 2Bii). Conversion of theisosurface pixels to voxels provided volumetric datafor each cell. Figure 2A shows that the NSP populationcontained a significantly higher proportion of largecells, mean 1708.84 mm3, compared to the smaller cellsfound in the SP, mean 648.68 mm3; (P¼ 0.0000036).
Using this protocol the phenotype of the cells in boththe SP and NSP populations were determined for bothmalignant and benign cells (Fig. 2C). All cells fromboththe SP and NSP populations were of Ber-EP4þve
epithelial origin. Significant difference in the pheno-type between benign SP and NSP populations wasobserved. The benign SP population was enriched forbasal cells labeled with k5 and k14 compared to thecorresponding NSP (59.5� 7.1% and 76� 9.1% vs.32.8� 6.3% and 29� 3.8% (P¼ 0.014 and 0.0003)respectively). In contrast, the benign NSP populationwas enriched for cells of a luminal phenotype,expressing k8 and k18 (55.4� 7.1% and 53.7� 4.7% ofNSP vs. 32.2� 10.56% and 29� 10.7% of SP (P¼ 0.089and 0.054) respectively). This enriched SP basalphenotype is reflected in the cells expression ofp63 (52.7� 4.3% SP vs. 32.1� 3.8% NSP; P¼ 0.024),PSA (21.95� 5.2% SP vs. 52.8� 5.8% NSP; P¼0.00319). However, there was no significant differencein the number of cells within each population express-ing AR (15.5� 4.9% SP vs. 28� 3.3% NSP; P¼ 0.081).Significant difference between SP and NSP in thenumbers of cells expressingp21WAF1/Cip1 (10.5� 2%vs.2.1� 1%; P¼ 0.0016), p27kip1 (27.7� 3.5 vs. 9.5� 3%;
The Prostate DOI 10.1002/pros
Fig. 1. Isolationof anunlabelledprostate epithelialcell SP.A:Histogramsdepictingprostateepithelialenrichedpopulations labeledwith theepithelialmarkerBer-EP4, integrina2,CD45or isotype control (shadedarea).Markershighlightpositivelylabeledcellpopulations.B:FSCandSSC plots of total cell and integrin a2 Ber-EP4 positive prostate epithelial populations defined regions for Hoechst 33342 Red/Blueanalysis.Hoechst 33342Red/Blueprofiles for the integrin a2Ber-EP4 positiveprostate epithelialpopulationswith the SPpopulations definedby verapamilhydrochloride sensitivityonequivalentnumberof events.The final collectionHoechst 33342Red/Blue sort is shown.C:FSC andSSCplots showingboth theCD45þve andCD45�vepopulations andHoechst33342Red/Blueprofiles for theCD45�veprostate epithelialpop-ulationwith the SP populations defined by verapamil hydrochloride sensitivity on equivalent number of events.The final collection Hoechst33342Red/Blue sortis shown.
4 Brown et al.
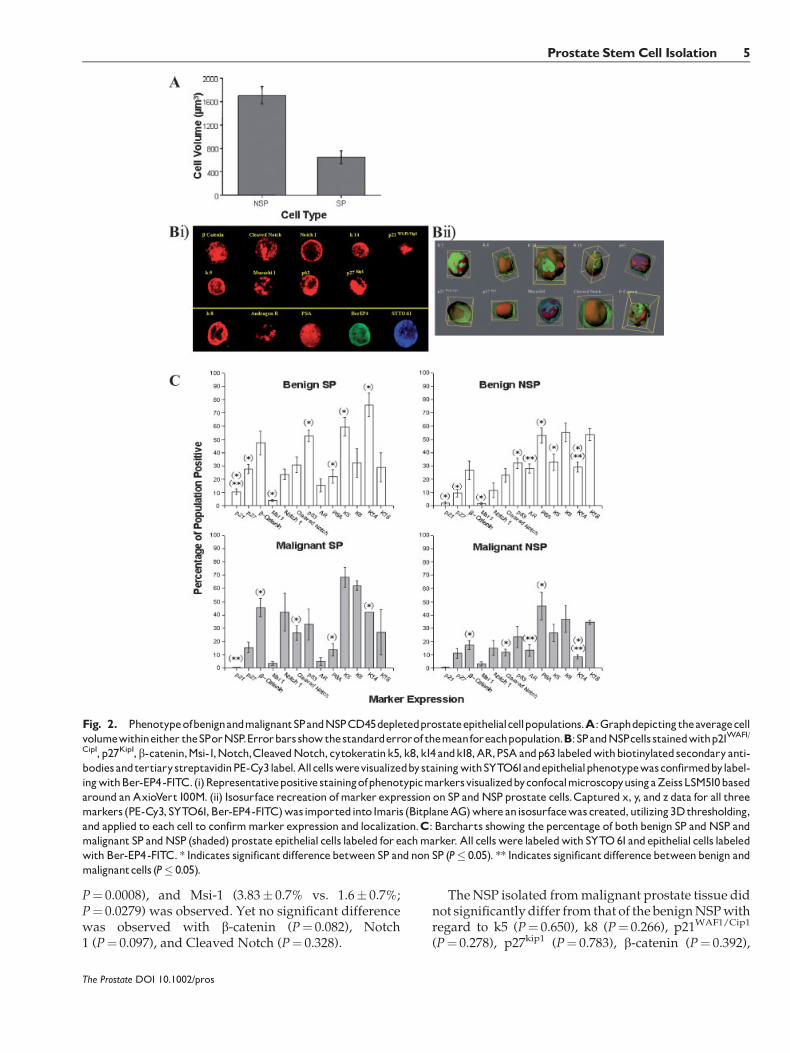
P¼ 0.0008), and Msi-1 (3.83� 0.7% vs. 1.6� 0.7%;P¼ 0.0279) was observed. Yet no significant differencewas observed with b-catenin (P¼ 0.082), Notch1 (P¼ 0.097), and Cleaved Notch (P¼ 0.328).
TheNSP isolated frommalignant prostate tissue didnot significantly differ from that of the benignNSPwithregard to k5 (P¼ 0.650), k8 (P¼ 0.266), p21WAF1/Cip1
(P¼ 0.278), p27kip1 (P¼ 0.783), b-catenin (P¼ 0.392),
The Prostate DOI 10.1002/pros
Fig. 2. PhenotypeofbenignandmalignantSPandNSPCD45depletedprostateepithelialcellpopulations.A:Graphdepictingtheaveragecellvolumewithineither theSPorNSP.Errorbarsshowthestandarderrorof themeanforeachpopulation.B:SPandNSPcells stainedwithp21WAF1/
Cip1, p27Kip1,b-catenin,Msi-1,Notch,CleavedNotch, cytokeratink5,k8,k14andk18,AR,PSAandp63 labeledwithbiotinylatedsecondaryanti-bodiesandtertiarystreptavidinPE-Cy3label.AllcellswerevisualizedbystainingwithSYTO61andepithelialphenotypewasconfirmedbylabel-ingwithBer-EP4-FITC.(i)RepresentativepositivestainingofphenotypicmarkersvisualizedbyconfocalmicroscopyusingaZeissLSM510basedaround anAxioVert100M. (ii) Isosurface recreation ofmarker expression on SP andNSP prostate cells.Captured x, y, and z data for all threemarkers (PE-Cy3,SYTO61,Ber-EP4-FITC)wasimportedinto Imaris (BitplaneAG)whereanisosurfacewascreated,utilizing3Dthresholding,and applied to each cell to confirmmarker expression and localization.C: Barcharts showing the percentage of both benign SP andNSP andmalignant SP andNSP (shaded) prostate epithelial cells labeled for eachmarker. All cellswere labeledwith SYTO61andepithelial cells labeledwith Ber-EP4-FITC. * Indicates significantdifferencebetween SP andnon SP (P� 0.05). ** Indicates significantdifferencebetweenbenign andmalignantcells (P� 0.05).
Prostate StemCell Isolation 5
Msi-1 (P¼ 0.346), Notch 1 (P¼ 0.755), Cleaved Notch1 (P¼ 0.172), and PSA (P¼ 0.591). Greater differenceswere observed with k14 and k18 (8.5� 1.5% vs.29� 3.8%; P¼ 0.035 and 34.5� 1.5% vs. 53.7� 4.7%;P¼ 0.089, respectively). Interestingly, fewer cellswithin the malignant NSP expressed AR as comparedwith benignNSP (13.3� 4.4%vs. 28� 3.3%;P¼ 0.0421).
There were considerable differences in the pheno-type of the malignant SP cells. Unlike the benign SP,whichwas enriched for basal cells, itwasnot possible todetermine the basal/luminal nature of the cells on thebasis of cytokeratin staining. The k5, k8, k14, andk18 status of the malignant SP was 68.5� 7.5%,62.25� 3.75%, 42� 0%, and 27� 17%, respectively,suggesting the presence of cells expressing bothcytokeratin k5 and k8. It was also noted that only0.3� 0.3% (n¼ 6) of cells within the malignant SPexpressed p21WAF1/Cip1.
Tissue PrimaryMarker Localization
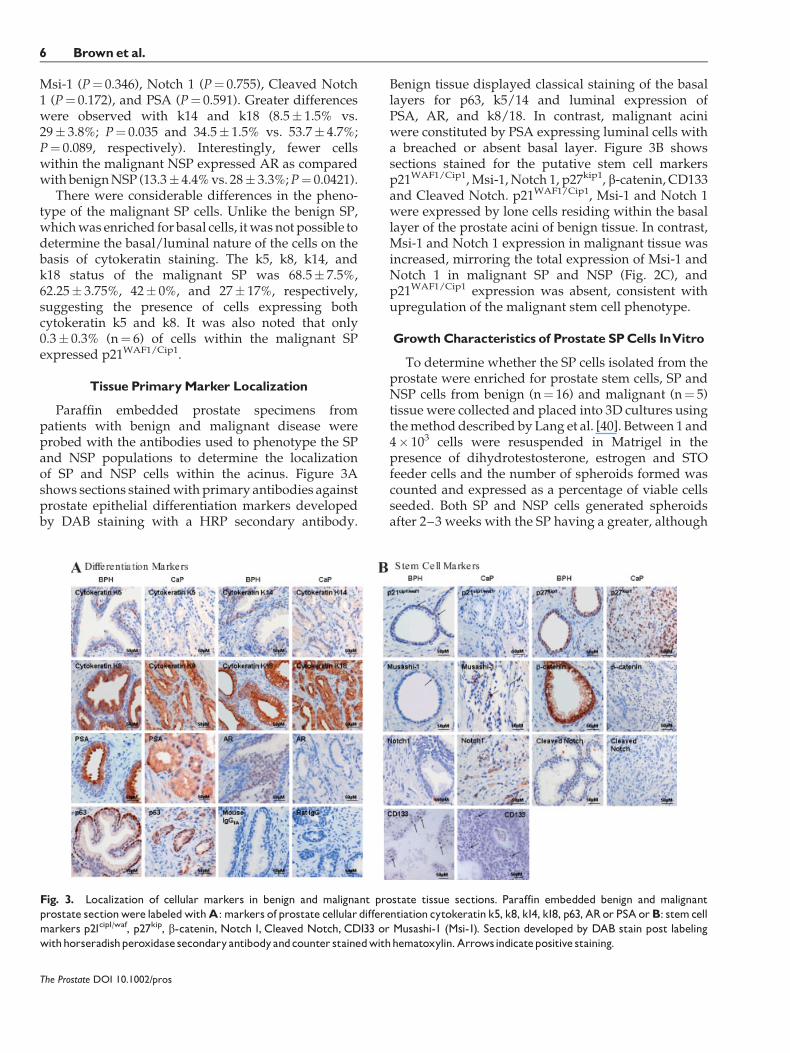
Paraffin embedded prostate specimens frompatients with benign and malignant disease wereprobed with the antibodies used to phenotype the SPand NSP populations to determine the localizationof SP and NSP cells within the acinus. Figure 3Ashows sections stainedwith primary antibodies againstprostate epithelial differentiation markers developedby DAB staining with a HRP secondary antibody.
Benign tissue displayed classical staining of the basallayers for p63, k5/14 and luminal expression ofPSA, AR, and k8/18. In contrast, malignant aciniwere constituted by PSA expressing luminal cells witha breached or absent basal layer. Figure 3B showssections stained for the putative stem cell markersp21WAF1/Cip1,Msi-1, Notch 1, p27kip1, b-catenin, CD133and Cleaved Notch. p21WAF1/Cip1, Msi-1 and Notch 1were expressed by lone cells residing within the basallayer of the prostate acini of benign tissue. In contrast,Msi-1 and Notch 1 expression in malignant tissue wasincreased, mirroring the total expression of Msi-1 andNotch 1 in malignant SP and NSP (Fig. 2C), andp21WAF1/Cip1 expression was absent, consistent withupregulation of the malignant stem cell phenotype.
GrowthCharacteristics of Prostate SPCells InVitro
To determine whether the SP cells isolated from theprostate were enriched for prostate stem cells, SP andNSP cells from benign (n¼ 16) and malignant (n¼ 5)tissue were collected and placed into 3D cultures usingthemethod described by Lang et al. [40]. Between 1 and4� 103 cells were resuspended in Matrigel in thepresence of dihydrotestosterone, estrogen and STOfeeder cells and the number of spheroids formed wascounted and expressed as a percentage of viable cellsseeded. Both SP and NSP cells generated spheroidsafter 2–3 weeks with the SP having a greater, although
The Prostate DOI 10.1002/pros
Fig. 3. Localization of cellular markers in benign and malignant prostate tissue sections. Paraffin embedded benign and malignantprostate sectionwere labeledwithA: markers of prostate cellular differentiation cytokeratin k5, k8, k14, k18, p63, ARor PSAorB: stemcellmarkers p21cip1/waf, p27kip, b-catenin, Notch 1, Cleaved Notch, CD133 or Musashi-1 (Msi-1). Section developed by DAB stain post labelingwithhorseradishperoxidasesecondaryantibodyandcounter stainedwithhematoxylin.Arrowsindicatepositive staining.
6 Brown et al.
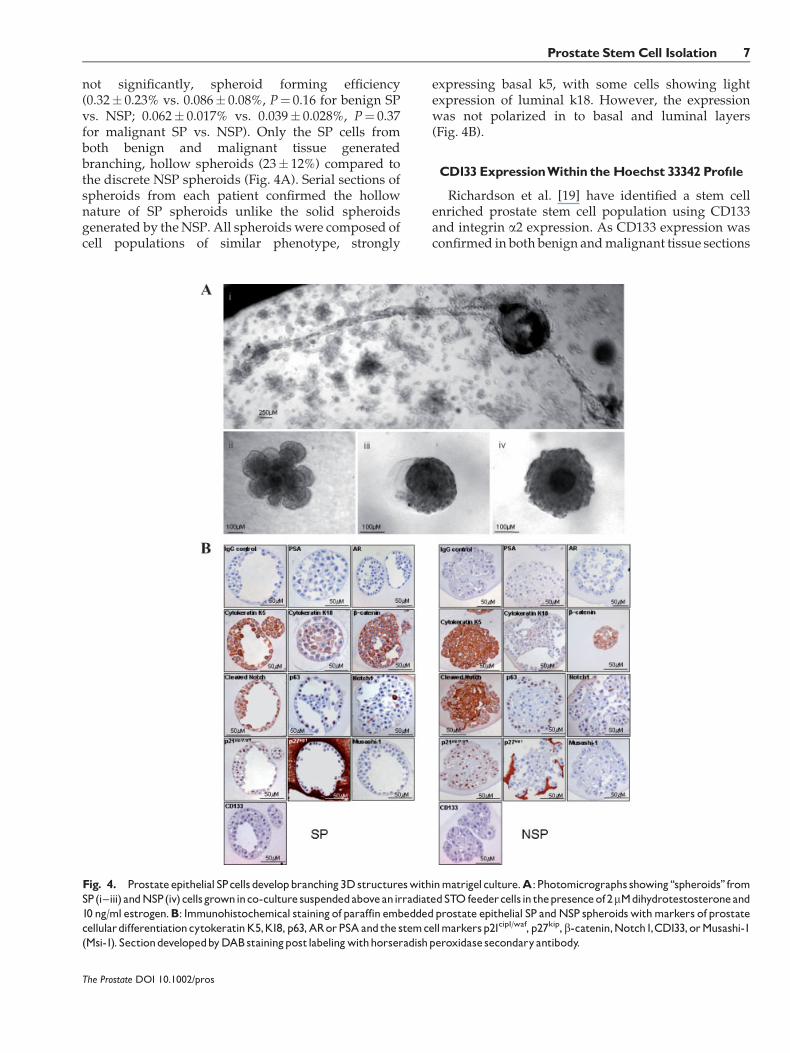
not significantly, spheroid forming efficiency(0.32� 0.23% vs. 0.086� 0.08%, P¼ 0.16 for benign SPvs. NSP; 0.062� 0.017% vs. 0.039� 0.028%, P¼ 0.37for malignant SP vs. NSP). Only the SP cells fromboth benign and malignant tissue generatedbranching, hollow spheroids (23� 12%) compared tothe discrete NSP spheroids (Fig. 4A). Serial sections ofspheroids from each patient confirmed the hollownature of SP spheroids unlike the solid spheroidsgenerated by theNSP. All spheroids were composed ofcell populations of similar phenotype, strongly
expressing basal k5, with some cells showing lightexpression of luminal k18. However, the expressionwas not polarized in to basal and luminal layers(Fig. 4B).
CD133 ExpressionWithin theHoechst 33342 Prof|le
Richardson et al. [19] have identified a stem cellenriched prostate stem cell population using CD133and integrin a2 expression. As CD133 expression wasconfirmed in both benign andmalignant tissue sections
The Prostate DOI 10.1002/pros
Fig. 4. Prostateepithelial SPcellsdevelopbranching3Dstructureswithinmatrigelculture.A:Photomicrographs showing‘‘spheroids’’ fromSP(i^iii) andNSP(iv)cellsgrowninco-culture suspendedaboveanirradiatedSTO feedercellsin thepresenceof2mMdihydrotestosteroneand10 ng/ml estrogen.B: Immunohistochemical stainingofparaffin embeddedprostate epithelial SP andNSP spheroidswithmarkers of prostatecellulardifferentiationcytokeratinK5,K18,p63,ARorPSAandthe stemcellmarkersp21cip1/waf, p27kip,b-catenin,Notch1,CD133,orMusashi-1(Msi-1). SectiondevelopedbyDABstainingpostlabelingwithhorseradishperoxidasesecondaryantibody.
Prostate StemCell Isolation 7
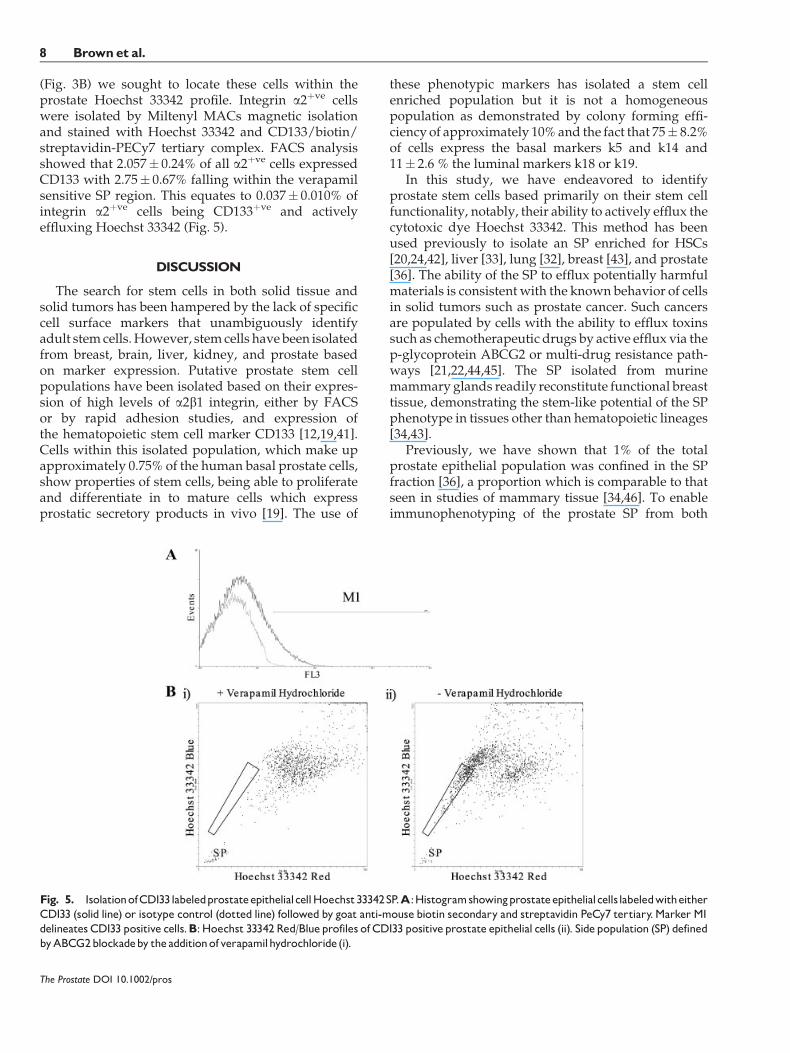
(Fig. 3B) we sought to locate these cells within theprostate Hoechst 33342 profile. Integrin a2þve cellswere isolated by Miltenyl MACs magnetic isolationand stained with Hoechst 33342 and CD133/biotin/streptavidin-PECy7 tertiary complex. FACS analysisshowed that 2.057� 0.24% of all a2þve cells expressedCD133 with 2.75� 0.67% falling within the verapamilsensitive SP region. This equates to 0.037� 0.010% ofintegrin a2þve cells being CD133þve and activelyeffluxing Hoechst 33342 (Fig. 5).
DISCUSSION
The search for stem cells in both solid tissue andsolid tumors has been hampered by the lack of specificcell surface markers that unambiguously identifyadult stemcells.However, stemcells havebeen isolatedfrom breast, brain, liver, kidney, and prostate basedon marker expression. Putative prostate stem cellpopulations have been isolated based on their expres-sion of high levels of a2b1 integrin, either by FACSor by rapid adhesion studies, and expression ofthe hematopoietic stem cell marker CD133 [12,19,41].Cells within this isolated population, which make upapproximately 0.75% of the human basal prostate cells,show properties of stem cells, being able to proliferateand differentiate in to mature cells which expressprostatic secretory products in vivo [19]. The use of
these phenotypic markers has isolated a stem cellenriched population but it is not a homogeneouspopulation as demonstrated by colony forming effi-ciency of approximately 10%and the fact that 75� 8.2%of cells express the basal markers k5 and k14 and11� 2.6 % the luminal markers k18 or k19.
In this study, we have endeavored to identifyprostate stem cells based primarily on their stem cellfunctionality, notably, their ability to actively efflux thecytotoxic dye Hoechst 33342. This method has beenused previously to isolate an SP enriched for HSCs[20,24,42], liver [33], lung [32], breast [43], and prostate[36]. The ability of the SP to efflux potentially harmfulmaterials is consistent with the known behavior of cellsin solid tumors such as prostate cancer. Such cancersare populated by cells with the ability to efflux toxinssuch as chemotherapeutic drugs by active efflux via thep-glycoprotein ABCG2 or multi-drug resistance path-ways [21,22,44,45]. The SP isolated from murinemammary glands readily reconstitute functional breasttissue, demonstrating the stem-like potential of the SPphenotype in tissues other than hematopoietic lineages[34,43].
Previously, we have shown that 1% of the totalprostate epithelial population was confined in the SPfraction [36], a proportion which is comparable to thatseen in studies of mammary tissue [34,46]. To enableimmunophenotyping of the prostate SP from both
The Prostate DOI 10.1002/pros
Fig. 5. IsolationofCD133labeledprostateepithelialcellHoechst33342SP.A:HistogramshowingprostateepithelialcellslabeledwitheitherCD133 (solid line) or isotype control (dotted line) followed by goat anti-mouse biotin secondary and streptavidin PeCy7 tertiary.Marker M1delineates CD133 positive cells.B:Hoechst 33342Red/Blueprofiles of CD133 positiveprostate epithelial cells (ii). Sidepopulation (SP) definedbyABCG2blockadeby theadditionofverapamilhydrochloride(i).
8 Brown et al.

benign and malignant PECs the Hoechst 33342 dyeefflux described by Bhatt et al. [36] was adapted toisolate an unlabelled SP and NSP cell populations.This modified approach isolated a similar benign SPof 0.93� 0.12% of epithelial cells. Interestingly, thepercentage of SP cells isolated frommalignant prostatewas significantly lower at 0.57� 0.11%. This was alsoobserved byCollins et al. [47]who noted that the a2b1hi
CD133 population decreased from 1% in benign tissueto 0.1–0.3% in malignant prostate tissue.
Characterization of the cells within the prostate SPshows that the cells share similar properties topreviously describedHSC. Prostate SP cells, like limbalepithelial SP cells [48], are smaller (648 mm3) than cellswithin the NSP (1708.84 mm3), having little cytoplasm.This feature was first noted in our previous study,which first described a Hoechst 33342 SP in PECs [36].Cell cycle analysis of the prostate SP showed that thecells were in either G0 or G1. Back gating these cell cyclestages on to FSC and SSC plots revealed two distinctpopulations based on cell size and granularity, with thequiescent G0 displaying low FSC and SSC profilesindicative of small cells with little cytoplasm [36]. Thequiescent nature of the putative prostate stem cells wasalso observed by Richardson et al. [19] who found thatthe CD133þve a2b1hi cells did not express the cellularproliferation marker Ki-67.
Utilization of a modified Hoechst 33342 dye effluxassay has enabled the isolation of CD45þve depletedprostate epithelial SP and NSP from both benignand malignant prostate for phenotypic analysis. Itis hypothesized that the prostate stem cell niche lieswithin the basal layer of the prostate acinus and stemcells would have a basal phenotype [49–51]. Initialphenotypic analysis of benign PECs reveals that theSP is a heterogeneous population, containing cellsof both a luminal (k8/k18) and basal (k5/k14)phenotype. However, unlike the NSP phenotype,which shows a luminal to basal ratio of 3:1, reflectingthe benign hyperplasic origin of the TURP chips, thebenign SP is enriched for cells expressing k5 and k14.The SP is also enriched for cells expressing p63,which has also been shown to be expressed byproliferative myoepithelial cells of mammary tissuerequired for the normal development of the glandulartissue [50–55]. Tissue localization of the p63 markerwithin the basal cell layer of the prostate acinussupports the hypothesis that the proliferative stem cellniche is within the basal layer.
The organized luminal andbasal pattern is lost in themalignant SP. This change in overt basal phenotype inmalignancy is not a new finding; diminished basalcytokeratin expression inbothSPandNSPcomplementthe global decrease in basal layer integrity in prostatecancer [56]. The malignant SP phenotype cannot be
truly determined as either basal or luminal cells on thebasis of their cytokeratin expression as there is aproportion of cells that are k5þve and k8þve. The k8expression within the malignant SP may represent thediminished representative basal cell component of theneoplastic prostate acini although the emergence ofan intermediate cell, the transit amplifying cell, hasbeen hypothesized, with co-localization of both basaland luminal protein expression representing a shift ofthe stem cells into their immediate progenitors [57–60].The co-expression of both luminal and basalmarkers inthemalignant SPmay also suggest that the cancer stemcells have de-differentiated, acquiring a fetal pheno-type. Wang et al. [49] found that the majority of theembryonic urogenital sinus epitheliumanddevelopingprostatic epithelium expressed both luminal and basalcytokeratin markers and proposed that these cellscould give rise to both differentiated basal and luminalcells. Thus, the increased numbers of cells of indeter-minate origin in the malignant SP as compared to thebenign SPmaybedue to an increase in stem cell/transitamplifying cell numbers [61] or de-differentiation ofcells to a more fetal phenotype.
Phenotypic analysis of both the SP and NSP withputative stem cell markers suggested an enrichment ofstemcellswithin the SP. The SP is enriched for theMsi-1marker, an RNA binding protein which is required forasymmetrical division of stem cells [37,62]. The role ofMsi-1 as a stem cell marker is supported by recent datawhich shows thatMsi-1 is required for themaintenanceof the stem cell phenotype in germ-line stem cells indrosophila and murine testes [63] and is a marker forboth neural and colonic stem cells [64,65]. The benignSP is also enriched for cells expressing the cyclin-dependent kinase inhibitor (CDKI) p21cip1/waf1. Anincreased number of cells expressing both Msi-1 andp21cip1/waf1 within the SP populations suggest that theSP contains a greater population of cells with stem cellcharacteristics. We found few cells in tissue sectionsexpressing p21cip1/waf1 and Msi-1, but all of these cellswere found in the basal cell layer and constituted lessthan 1% of the cells in any one acinus section. Whencomparing the benign to the malignant SP there was asignificant reduction of p21cip1/waf1, mirroring thereport by Cheng et al. [66] that the loss of p21 is amarker of poor prognosis in prostate cancer. The loss ofthe control of entry in to cell cycle at theG0/G1 gate andthe associated diminished control of cell cycle inneoplastic stem cells supports the observation thatcancers have the ability to produce proliferating cellswith a capacity for differentiation potential and theability for self-renewal. The decrease in p21cip/waf1 andthe changes in Msi-1 expression correlates with the‘‘cancer stem cell’’ theory that these highly plasticcells are able to cycle, differentiate, proliferate, and
The Prostate DOI 10.1002/pros
Prostate StemCell Isolation 9

self-renew outside the confines of normal tissuearchitecture and that they have the ability to behavemore like amulti-potent stem,with decreased cell cyclearrest at the G0/G1 gate [67,68].
The stem cell marker p27kip1 acts as a barrier toprogression towards S phase, potentially signaling exitfrom the cell cycle and quiescence [69,70]. P27kip1 haspreviously been shown to be restricted to the luminalcells in the normal prostate. Expression in the basalcompartment is variable with some acini not express-ing p27kip1 whilst others may show some or evencomplete staining for p27kip1 [71].Here,wehave shownthat there is increased expression of p27kip1 in thebenign SP, confirming the increase in quiescent cellswithin the SPdemonstrated by Bhatt et al. [36]. Stainingof benign acini shows that the majority of p27kip1þve
cells are luminal, but there are occasional small basalcells also expressing p27kip1. Malignant prostate tissuep27kip1 expression differs from that in benign tissue.There is a reduction in the number of p27kip1 cells inboth the SP and NSP in malignant tissue as comparedwith benign tissue and there is no difference betweenthe SP and NSP in malignant tissue. Loss of p27kip1 hasbeen previously shown to be a poor prognostic markerin prostate as well as other solid organ cancers[66,72,73].
Stem cells classically have the characteristicsof being pluripotent, clonogenic, self-renewing pro-genitor cells and it is proposed that adult prostate stemcells are capable of regenerating all the cell types thatconstitute the prostate organ [49,57,74], although in thisstudy we have not sort to determine their ability todifferentiate in to the prostate neuroendocrine cells.The classical proof of stem activity is the formation ofglandular structures/tumors in murine xenografts.Richardson et al. [19] demonstrated that 104�5� 104
a2b1hi CD133þve PECs were required to form aglandular acinus upon murine xenograft. Due to thevery low numbers of cells isolated from TURP chips bythe Hoechst 33342 dye efflux protocol, often less than103, we were unable to conduct in vivo murinexenograft experiments to confirm these characteristics.In vitro expansion and long term proliferation of adultstem cells continues to be problematic due theirasymmetric cell kinetics (reviewed in Pare and Sherley[75]) leading to the generation of a stem cell, by selfrenewal, and a terminally differentiating daughter cell.Therefore, in order to isolate larger numbers of SP cellswithout in vitro expansion we are currently adaptingour protocols to isolate cells from prostates isolated byradical prostatectomy.
However, we have used in vitro 3D cell cultures toassess both the colony forming efficiency and the abilityof cells to differentiate in to prostate acinar likestructures and give rise to both basal and luminal
epithelial cells. Previously, the co-culture of ratmammary epithelial cells with extra-cellular matrix incombination with stromal cells has given an insightinto this behavior and has provided an opportunityto assess how these microorgans grow in response togrowth factors and specific hormones in their micro-environment [76]. This methodology was translatedfor use with primary prostate CD44�ve basal cells todevelop a co-culture which in the presence of the STO(mouse fibroblast) feeder layer, allowing single pros-tate cells with stem cell features to produce 3Dstructures or ‘‘spheroids’’ exhibiting the propensityfor differentiation [40]. From very small numbers of SPcells we were able to grow spheroids consistently.These showed not only the ability to form prostateacinus-like spheres but also the capacity to differentiateto produce defined tubular structures. This property ofSP cells differed from the NSP cultures. Although thelatter formed spheroids (to a much lesser extent), theydid not show functional behavior of the type exhibitedby the SP cells. It should be noted that although all cellswere viable at time of plating, the reduced number ofspheroids formed by the NSP may be a result of theirinability to efflux the cytotoxic Hoechst 33342dye. However, this pattern of growth by NSP cells hasalso been described in mammary tissue. In this study,there were no signs of functional differentiation intobranching structures from NSP breast epithelial cells,unlike breast epithelial SP cells which were able togive rise to cells of both luminal and myoepitheliallineages [43].
The data presented here have shown that we haveisolated an SP population from both benign andmalignant prostate epithelial tissue which is enrichedfor cells with a stem cell phenotype. However, it isclear that this is an enriched population, nota homogenous population. Recently, studies using theputative hematopoietic stem cell marker CD133[41] have been used to define a group of a2b1hiCD133hi
prostate basal cells which exhibit the primitive cellculture characteristics of high proliferative potentialin vitro and the ability to reconstitute prostatic-likeacini in vivo [19]. Again this population is enrichedfor cells with a stem like phenotype but it isnot homogeneous. We therefore sought to combineCD133 marker selection with our modified Hoechst33342 dye efflux assay to determine the presence of aCD133þve prostate epithelial SP. Our studies haveconfirmed the presence of a2b1þve CD133þve cellswhich constitute 2.75% of all a2b1 PECs. Thisis higher than the reported 0.75% of basal cells reportedby Richardson et al. but this was due to selection of alla2b1 cells rather than selecting cells with high levels ofexpression. However, incorporation of the Hoechst33342 dye efflux assay isolated a small subpopulation
The Prostate DOI 10.1002/pros
10 Brown et al.

of basal cells which represents 0.04% of all prostatebasal epithelial cells. This is comparable with thereported figures that 0.75% of the basal cells expressCD133, of which approximately 10% are able to formcolonies [19]. We are currently characterizing thispopulation for stem cell activity.
In summary, the prostate Hoechst 33342 SP has beenshown to consist of small basal epithelial cells with anenriched population of cells with the stem cell featuresof differentiation and proliferation. Furthermore,the comparative analysis of this stem cell enrichedpopulation with a control population of NSPcells has shown the latter to consist mainly of cells,which display a more differentiated phenotype. Whencultured, these display the characteristics expectedfrom a progenitor population. The loss of expression ofthe important cell cycle controller p21cip1/waf1 in themalignant SP and the increased expression of theproliferationmarker b-catenin is consistent with the SPbeing stem cell enriched and is consistent with thenotion that these cells are important in malignantchange. Further research with promising new markerssuch as CD133 has the potential to provide additionalpurification of the Hoechst 33342 dye efflux stem cellmodel.
ACKNOWLEDGMENTS
We thank the urological surgeons of theChristie Hospital NHS Trust, Manchester, UK, SalfordRoyal Hospitals NHS Trust, Salford, UK, and SouthManchester University Hospitals NHS Trust, Man-chester, UK for prostate tissue collection, Mr.M Hughes and Mr. J Barry Cancer Research UK FlowCytometry Facility, Dr. S Bagley Cancer Research UKAdvanced Imaging Facility for providing technicalassistance and Dr. Hideyuki Okano for supplying theMsi-1 antibody. We would also like to thank Dr. RBClarke and the Breast Biology Group, Cancer ResearchUKDepartment of Medical Oncology,Manchester, UKtheir support and the British Urological Foundation,and The Fellows Fellowship of the Royal College ofSurgeons of England for their financial support.
REFERENCES
1. Statistics: National Office of Statistics UK 2004.
2. Bonnet D, Dick JE. Human acute myeloid leukemia is organizedas a hierarchy that originates from a primitive haematopoieticcell. Nat Med 1997;3(7):730–737.
3. George AA, Franklin J, Kerkof K, Shah AJ, Price M, Tsark E,Bockstoce D, Yao D, Hart N, Carcich S, Parkman R, Crooks GM,WeinbergK. Detection of leukemic cells in the CD34(þ)CD38(�)bone marrow progenitor population in children with acutelymphoblastic leukemia. Blood 2001;97(12):3925–3930.
4. Miyamoto T, Weissman IL, Akashi K. AML1/ETO-expressingnonleukemic stem cells in acute myelogenous leukemia with
8;21 chromosomal translocation. Proc Natl Acad Sci USA2000;97(13):7521–7526.
5. Heppner GH. Tumor heterogeneity. Cancer Res 1984;44(6):2259–2265.
6. Fidler IJ, Hart IR. Biological diversity in metastatic neoplasms:Origins and implications. Science 1982;217(4564):998–1003.
7. Al-Hajj M,WichaMS, Benito-Hernandez A,Morrison SJ, ClarkeMF. Prospective identification of tumorigenic breast cancer cells.Proc Natl Acad Sci USA 2003;100(7):3983–3988.
8. Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire J,Dirks PB. Identification of a cancer stem cell in human braintumors. Cancer Res 2003;63(18):5821–5828.
9. Grichnik JM, Burch JA, Schulteis RD, Shan S, Liu J, Darrow TL,Vervaert CE, Seigler HF. Melanoma, a tumor based on a mutantstem cell? J Invest Dermatol 2006;126(1):142–153.
10. Bonkhoff H. Role of the basal cells in premalignant changes ofthe human prostate: A stem cell concept for the development ofprostate cancer. Eur Urol 1996;30(2):201–205.
11. Collins AT, Habib FK, Maitland NJ, Neal DE. Identification andisolation of human prostate epithelial stem cells based onalpha(2)beta(1)-integrin expression. J Cell Sci 2001;114(Pt 21):3865–3872.
12. Hudson DL, O’Hare M, Watt FM, Masters JR.Proliferative heterogeneity in the human prostate: Evidence forepithelial stem cells. Lab Invest 2000;80(8):1243–1250.
13. Reiter RE, Gu Z, Watabe T, Thomas G, Szigeti K, Davis E, WahlM, Nisitani S, Yamashiro J, Le Beau MM, Loda M, Witte ON.Prostate stem cell antigen: A cell surface marker overexpressedin prostate cancer. Proc Natl Acad Sci USA 1998;95(4):1735–1740.
14. Israeli RS, Powell CT, Corr JG, FairWR, HestonWD. Expressionof the prostate-specific membrane antigen. Cancer Res 1994;54(7):1807–1811.
15. Kadkol SS, Brody JR, Epstein JI, Kuhajda FP, Pasternack GR.Novel nuclear phosphoprotein pp32 is highly expressed inintermediate- and high-grade prostate cancer. Prostate1998;34(3):231–237.
16. Lang SH, Clarke NW, George NJ, Testa NG. Primary prostaticepithelial cell binding to human bone marrow stroma and therole of alpha2beta1 integrin. Clin ExpMetastasis 1997;15(3):218–227.
17. LiuH,Moy P, Kim S, Xia Y, RajasekaranA,Navarro V, KnudsenB, Bander NH. Monoclonal antibodies to the extracellulardomain of prostate-specific membrane antigen also react withtumorvascular endothelium.CancerRes 1997;57(17):3629–3634.
18. Chang SS, Reuter VE, Heston WD, Bander NH, Grauer LS,Gaudin PB. Five different anti-prostate-specific membraneantigen (PSMA) antibodies confirm PSMA expression intumor-associated neovasculature. Cancer Res 1999;59(13):3192–3198.
19. Richardson GD, Robson CN, Lang SH, Neal DE, Maitland NJ,CollinsAT.CD133, anovelmarker for humanprostatic epithelialstem cells. J Cell Sci 2004;117(Pt 16):3539–3545.
20. Goodell MA, Brose K, Paradis G, Conner AS, Mulligan RC.Isolation and functional properties of murine haematopoieticstem cells that are replicating in vivo. J Exp Med 1996;183(4):1797–1806.
21. Zhou S, Schuetz JD, Bunting KD, Colapietro AM, Sampath J,Morris JJ, Lagutina I, Grosveld GC, Osawa M, Nakauchi H,Sorrentino BP. The ABC transporter Bcrp1/ABCG2 is expressedin a wide variety of stem cells and is a molecular determinant ofthe side-population phenotype. Nat Med 2001;7(9):1028–1034.
The Prostate DOI 10.1002/pros
Prostate StemCell Isolation 11

22. Zhou S, Morris JJ, Barnes Y, Lan L, Schuetz JD, Sorrentino BP.Bcrp1 gene expression is required for normal numbers of sidepopulation stem cells in mice, and confers relative protection tomitoxantrone in haematopoietic cells in vivo. ProcNatl Acad SciUSA 2002;99(19):12339–12344.
23. KimM, TurnquistH, Jackson J, SgagiasM, YanY, GongM,DeanM, Sharp JG, Cowan K. The multidrug resistance transporterABCG2 (breast cancer resistance protein 1) effluxes Hoechst33342 and is overexpressed in haematopoietic stem cells. ClinCancer Res 2002;8(1):22–28.
24. Scharenberg CW, Harkey MA, Torok-Storb B. The ABCG2transporter is an efficient Hoechst 33342 efflux pump and ispreferentially expressed by immature human haematopoieticprogenitors. Blood 2002;99(2):507–512.
25. Storms RW, Goodell MA, Fisher A, Mulligan RC, Smith C.Hoechst dye efflux reveals a novel CD7(þ)CD34(-) lymphoidprogenitor in human umbilical cord blood. Blood 2000;96(6):2125–2133.
26. Guo Y, Lubbert M, Engelhardt M. CD34(-) haematopoietic stemcells: Current concepts and controversies. Stem Cells 2003;21(1):15–20.
27. Feuring-Buske M, Hogge DE. Hoechst 33342 efflux identifies asubpopulation of cytogenetically normal CD34(þ)CD38(-) pro-genitor cells from patients with acute myeloid leukemia. Blood2001;97(12):3882–3889.
28. Foster CS, Dodson A, Karavana V, Smith PH, Ke Y. Prostaticstem cells. J Pathol 2002;197(4):551–565.
29. Spangrude GJ, Heimfeld S, Weissman IL. Purification andcharacterization of mouse haematopoietic stem cells. Science1988;241(4861):58–62.
30. Baum CM, Weissman IL, Tsukamoto AS, Buckle AM, Peault B.Isolation of a candidate human haematopoietic stem-cell population. Proc Natl Acad Sci U S A 1992;89(7):2804–2808.
31. Asakura A, Rudnicki MA. Side population cells from diverseadult tissues are capable of in vitro haematopoietic differ-entiation. Exp Hematol 2002;30(11):1339–1345.
32. SummerR,KottonDN, SunX,MaB, FitzsimmonsK, FineA. Sidepopulation cells and Bcrp1 expression in lung. Am J PhysiolLung Cell Mol Physiol 2003;285(1):L97–L104.
33. Uchida N, Leung FY, Eaves CJ. Liver and marrow of adult mdr-1a/1b(-/-) mice show normal generation, function, and multi-tissue trafficking of primitive haematopoietic cells. ExpHematol2002;30(8):862–869.
34. Alvi AJ, ClaytonH, Joshi C, Enver T, AshworthA, VivancoMM,Dale TC, SmalleyMJ. Functional andmolecular characterisationof mammary side population cells. Breast Cancer Res 2003;5(1):R1–R8.
35. Challen GA, Little MH. A side order of stem cells: The SPphenotype. Stem Cells 2006;24(1):3–12.
36. Bhatt RI, Brown MD, Hart CA, Gilmore P, Ramani VA, GeorgeNJ, Clarke NW. Novel method for the isolation and character-isation of the putative prostatic stem cell. Cytometry 2003;54(2):89–99.
37. Okano H, Kawahara H, Toriya M, Nakao K, Shibata S, Imai T.Function of RNA-binding protein musashi-1 in stem cells. ExpCell Res 2005;306:349–356.
38. Martin GR, Evans MJ. Differentiation of clonal lines ofteratocarcinoma cells: Formation of embryoid bodies in vitro.Proc Natl Acad Sci USA 1975;72(4):1441–1445.
39. Lang SH, Clarke NW, George NJ, Allen TD, Testa NG.Interactionofprostate epithelial cells frombenignandmalignant
tumor tissue with bone-marrow stroma. Prostate 1998;34(3):203–213.
40. Lang SH, Stark M, Collins A, Paul AB, Stower MJ, Maitland NJ.Experimental prostate epithelial morphogenesis in response tostroma and three-dimensional matrigel culture. Cell GrowthDiffer 2001;12(12):631–640.
41. Yin AH, Miraglia S, Zanjani ED, Almeida-Porada G, Ogawa M,LearyAG,Olweus J,Kearney J, BuckDW.AC133, anovelmarkerfor human haematopoietic stem and progenitor cells. Blood1997;90(12):5002–5012.
42. Goodell MA, Rosenzweig M, Kim H, Marks DF, DeMaria M,Paradis G, Grupp SA, Sieff CA, Mulligan RC, Johnson RP. Dyeefflux studies suggest that haematopoietic stem cells expressinglow or undetectable levels of CD34 antigen exist in multiplespecies. Nat Med 1997;3(12):1337–1345.
43. Clarke RB, Spence K, Anderson E, Howell A, Okano H, PottenCS. Aputative human breast stem cell population is enriched forsteroid receptor-positive cells. Dev Biol 2005;277(2):443–456.
44. Siegsmund MJ, Kreukler C, Steidler A, Nebe T, Kohrmann KU,Alken P. Multidrug resistance in androgen-independent grow-ing rat prostate carcinoma cells is mediated by P-glycoprotein.Urol Res 1997;25(1):35–41.
45. Van Brussel JP, Jan Van Steenbrugge G, Van Krimpen C,Bogdanowicz JF,VanDerKwastTH,Schroder FH,MickischGH.Expression of multidrug resistance related proteins and pro-liferative activity is increased in advanced clinical prostatecancer. J Urol 2001;165(1):130–135.
46. WelmBE,Tepera SB,VeneziaT,GraubertTA,Rosen JM,GoodellMA. Sca-1(pos) cells in the mouse mammary gland represent anenriched progenitor cell population. Dev Biol 2002;245(1):42–56.
47. Collins AT, Berry PA, Hyde C, Stower MJ, Maitland NJ.Prospective identification of tumorigenic prostate cancer stemcells. Cancer Res 2005;65(23):10946–10951.
48. Umemoto T, Yamato M, Nishida K, Yang J, Tano Y, Okano T.Limbal epithelial side-population cells have stem cell-likeproperties, including quiescent state. Stem Cells 2006;24(1):86–94.
49. Wang Y, Hayward S, Cao M, Thayer K, Cunha G. Celldifferentiation lineage in the prostate. Differentiation 2001;68(4–5):270–279.
50. Signoretti S, Waltregny D, Dilks J, Isaac B, Lin D, Garraway L,Yang A, Montironi R, McKeon F, LodaM. p63 is a prostate basalcell marker and is required for prostate development. Am JPathol 2000;157(6):1769–1775.
51. Parsons JK, Gage WR, Nelson WG, De Marzo AM. p63 proteinexpression is rare in prostate adenocarcinoma: Implications forcancer diagnosis and carcinogenesis. Urology 2001;58(4):619–624.
52. Nylander K, Vojtesek B, Nenutil R, Lindgren B, Roos G,Zhanxiang W, Sjostrom B, Dahlqvist A, Coates PJ. Differentialexpressionof p63 isoforms in normal tissues andneoplastic cells.J Pathol 2002;198(4):417–427.
53. Brunner HG, Hamel BC, Bokhoven Hv H. P63 gene mutationsand human developmental syndromes. Am J Med Genet 2002;112(3):284–290.
54. Taylor-Papadimitriou J, Stampfer M, Bartek J, Lewis A, BoshellM, Lane EB, Leigh IM. Keratin expression in human mammaryepithelial cells cultured from normal and malignant tissue:Relation to in vivo phenotypes and influence of medium. J CellSci 1989;94(Pt 3):403–413.
55. Barbareschi M, Pecciarini L, Cangi MG, Macri E, Rizzo A, VialeG, Doglioni C. p63, a p53 homologue, is a selective nuclear
The Prostate DOI 10.1002/pros
12 Brown et al.

marker of myoepithelial cells of the human breast. Am J SurgPathol 2001;25(8):1054–1060.
56. Oliai BR, Kahane H, Epstein JI. Can basal cells be seen inadenocarcinoma of the prostate?: An immunohistochemicalstudy using high molecular weight cytokeratin (clone34betaE12) antibody. Am J Surg Pathol 2002;26(9):1151–1160.
57. Bonkhoff H, Stein U, Remberger K. Multidirectional differ-entiation in the normal, hyperplastic, and neoplastic humanprostate: Simultaneous demonstration of cell-specific epithelialmarkers. Hum Pathol 1994;25(1):42–46.
58. Verhagen AP, Ramaekers FC, Aalders TW, Schaafsma HE,Debruyne FM, Schalken JA. Colocalization of basal and luminalcell-type cytokeratins in human prostate cancer. Cancer Res1992;52(22):6182–6187.
59. Robinson EJ, Neal DE, Collins AT. Basal cells are progenitors ofluminal cells in primary cultures of differentiating humanprostatic epithelium. Prostate 1998;37(3):149–160.
60. Xue Y, Smedts F, Debruyne FM, de la Rosette JJ, Schalken JA.Identification of intermediate cell types by keratin expression inthe developing human prostate. Prostate 1998;34(4):292–301.
61. vanLeendersG,DijkmanH,Hulsbergen-vandeKaaC,RuiterD,Schalken J. Demonstration of intermediate cells during humanprostate epithelial differentiation in situ and invitro using triple-staining confocal scanning microscopy. Lab Invest 2000;80(8):1251–1258.
62. OkanoH, Imai T,OkabeM.Musashi:A translational regulator ofcell fate. J Cell Sci 2002;115(Pt 7):1355–1359.
63. Siddall NA,McLaughlin EA,MarrinerNL, HimeGR. The RNA-binding protein Musashi is required intrinsically to maintainstem cell identity. Proc Natl Acad Sci USA 2006;103(22):8402–8407.
64. Toda M, Iizuka Y, Yu W, Imai T, Ikeda E, Yoshida K, Kawase T,Kawakami Y, Okano H, Uyemura K. Expression of the neuralRNA-binding protein Musashi1 in human gliomas. Glia 2001;34(1):1–7.
65. Potten CS, Booth C, Tudor GL, Booth D, Brady G, Hurley P,Ashton G, Clarke R, Sakakibara S, Okano H. Identification of a
putative intestinal stemcell andearly lineagemarker;musashi-1.Differentiation 2003;71(1):28–41.
66. Cheng L, Lloyd RV, Weaver AL, Pisansky TM, ChevilleJC, Ramnani DM, Leibovich BC, Blute ML, Zincke H, BostwickDG. The cell cycle inhibitors p21WAF1 and p27KIP1 areassociated with survival in patients treated by salvage prosta-tectomy after radiation therapy. ClinCancer Res 2000;6(5):1896–1899.
67. ReyaT,Morrison SJ, ClarkeMF,Weissman IL. Stem cells, cancer,and cancer stem cells. Nature 2001;414(6859):105–111.
68. Masters JR, Foley CL, Bisson I, AhmedA. Cancer stem cells. BJUInt 2003;92(7):661–662.
69. Coats S, Flanagan WM, Nourse J, Roberts JM. Requirement ofp27Kip1 for restriction point control of the fibroblast cell cycle.Science 1996;272(5263):877–880.
70. Sherr CJ. Cancer cell cycles. Science 1996;274(5293):1672–1677.
71. De Marzo AM, Meeker AK, Epstein JI, Coffey DS. Prostate stemcell compartments: Expression of the cell cycle inhibitor p27Kip1in normal, hyperplastic, and neoplastic cells. Am J Pathol1998;153(3):911–919.
72. Lloyd RV, Erickson LA, Jin L, Kulig E, Qian X, Cheville JC,Scheithauer BW. p27kip1: A multifunctional cyclin-dependentkinase inhibitor with prognostic significance in human cancers.Am J Pathol 1999;154(2):313–323.
73. Tsihlias J, Kapusta L, Slingerland J. The prognostic significanceof altered cyclin-dependent kinase inhibitors in human cancer.Annu Rev Med 1999;50:401–423.
74. Morrison SJ, ShahNM,AndersonDJ. Regulatorymechanisms instem cell biology. Cell 1997;88(3):287–298.
75. Pare JF, Sherley JL. Biological principles for ex vivo adult stemcell expansion. Curr Top Dev Biol 2006;73:141–171.
76. Darcy KM, Zangani D, Shea-Eaton W, Shoemaker SF, Lee PP,Mead LH,Mudipalli A, Megan R, IpMM.Mammary fibroblastsstimulate growth, alveolar morphogenesis, and functionaldifferentiation of normal rat mammary epithelial cells. In VitroCell Dev Biol Anim 2000;36(9):578–592.
The Prostate DOI 10.1002/pros
Prostate StemCell Isolation 13




















