
CCR7 Mediates Inflammation-Associated Tumor Progression
12
Introduction Inflammatory signals are increasingly linked to the induction and promotion of tumorigenesis. Because chemokine receptors regulate immune cell migration and survival, and are often stimulated by inflammatory sig- nals, their expression on tumors may reflect Abstract Chemokine receptor 7 (CCR7) mediates leukocyte adhesion and chemotaxis from peripheral sites of inflammation through lym- phatic channels to secondary lymphoid organs. Aberrant CCR7 expression has been identified on certain tumor types and been linked to pro-survival and invasive pathways. In metastatic squa- mous cell carcinoma of the head and neck (SCCHN), we have described the selective upregulation of functional CCR7. In this manuscript, we review our understanding of CCR7-mediated sig- naling in metastatic SCCHN and provide evidence for its involve- ment in tumor survival, invasion, and metastasis. Autocrine and paracrine CCR7 activation appears to propagate the response to the CCR7 ligands CCL19 and CCL21, which are expressed by the lym- phatic endothelium, secondary lymphoid tissues, and CCR7-posi- tive tumor cells. Based on our recent findings, the induction of CCR7 expression and the sustenance of the autocrine signaling pathway have been shown to be regulated by NF-κB, similar to sev- eral types of immune cells. While extending these observations to metastatic SCCHN tumor cells, our studies highlight the importance of downstream NF-κB mediated CCR7 signals in the progression of SCCHN malignancy. Key Words Chemokine receptors NF-κB Inflammation Metastasis Head and neck cancer Robert L. Ferris, MD, PhD The Hillman Cancer Center Research Pavilion, 5117 Centre Avenue, Room 2.26b, Pittsburgh, PA 15213 E-mail: [email protected] 61 © 2006 Humana Press Inc. 0257–277X/ (Online)1559-0755/06/ 36/1–3:61–72/$30.00 CCR7 Mediates Inflammation- Associated Tumor Progression Immunologic Research 2006;36/1–3:61–72 Yvonne K. Mburu 1 Jun Wang 2 Michelle A. Wood 3 William H. Walker 3 Robert L. Ferris 1,2 1 Department of Immunology, University of Pittsburgh School of Medicine, Pittsburgh, PA 2 Department of Otolaryngology, University of Pittsburgh Medical Center, Pittsburgh, PA 3 Department of Cell Biology and Physiology, University of Pittsburgh School of Medicine, Pittsburgh, PA
Transcript of CCR7 Mediates Inflammation-Associated Tumor Progression

Introduction
Inflammatory signals are increasinglylinked to the induction and promotion of
tumorigenesis. Because chemokine receptorsregulate immune cell migration and survival,and are often stimulated by inflammatory sig-nals, their expression on tumors may reflect
AbstractChemokine receptor 7 (CCR7) mediates leukocyte adhesion andchemotaxis from peripheral sites of inflammation through lym-phatic channels to secondary lymphoid organs. Aberrant CCR7expression has been identified on certain tumor types and beenlinked to pro-survival and invasive pathways. In metastatic squa-mous cell carcinoma of the head and neck (SCCHN), we havedescribed the selective upregulation of functional CCR7. In thismanuscript, we review our understanding of CCR7-mediated sig-naling in metastatic SCCHN and provide evidence for its involve-ment in tumor survival, invasion, and metastasis. Autocrine andparacrine CCR7 activation appears to propagate the response to theCCR7 ligands CCL19 and CCL21, which are expressed by the lym-phatic endothelium, secondary lymphoid tissues, and CCR7-posi-tive tumor cells. Based on our recent findings, the induction ofCCR7 expression and the sustenance of the autocrine signalingpathway have been shown to be regulated by NF-κB, similar to sev-eral types of immune cells. While extending these observations tometastatic SCCHN tumor cells, our studies highlight the importanceof downstream NF-κB mediated CCR7 signals in the progressionof SCCHN malignancy.
Key WordsChemokine receptorsNF-κBInflammationMetastasisHead and neck cancer
Robert L. Ferris, MD, PhDThe Hillman Cancer Center ResearchPavilion, 5117 Centre Avenue, Room 2.26b,Pittsburgh, PA 15213E-mail: [email protected]
61© 2006Humana Press Inc.0257–277X/(Online)1559-0755/06/36/1–3:61–72/$30.00
CCR7 Mediates Inflammation-Associated Tumor Progression
Immunologic Research 2006;36/1–3:61–72
Yvonne K. Mburu1
Jun Wang2
Michelle A. Wood3
William H. Walker3
Robert L. Ferris1,2
1Department of Immunology,University of Pittsburgh School ofMedicine, Pittsburgh, PA2Department of Otolaryngology,University of Pittsburgh MedicalCenter, Pittsburgh, PA3Department of Cell Biology andPhysiology, University ofPittsburgh School of Medicine,Pittsburgh, PA

aberrant inflammatory signals and lead toenhanced tumor growth, invasion, and metas-tasis. In the clinical setting, despite ongoingadvances in cancer therapy, a large number ofpatients still succumb to metastatic disease.This is, by far, the most devastating aspect ofhuman cancers as it reflects the insidiousnature of malignant cells to escape conven-tional therapies and re-establish tumorgrowth at secondary sites, often in a moreaggressive manner. Thus, understanding thefactors that promote tumor invasion andspread is a critical area in cancer research.
The dual role of the immune system in therecognition and rejection of tumor cells andits potential role in sending procarcinogenicsignals has fascinated immunologists fordecades. Mechanisms of metastasis havereceived scientific investigation for over100 yr since the publication by StephenPaget in 1889 of the “seed and soil” hypoth-esis, which described metastasis as a non-random process (1). Indeed, it has beenobserved clinically that certain tumor types(the seed) have tissue tropism for particularorgans (the soil) where they establish sitesof malignant growth. Squamous cell carci-noma of the head and neck (SCCHN) is ahuman malignancy that metastasizes pre-dictably to the regional lymph nodes (LN).We therefore hypothesized that LN homingreceptors, such as CC chemokine receptor 7(CCR7), could mediate SCCHN tumormetastasis to these secondary lymphoidorgans (2).
Chemokines are small (8–14 kDa)chemotactic cytokines that bind to seven-transmembrane-domain G protein–coupledreceptors. They are classified into four fam-ilies, C, CC, CXC, and CX3C, based on theposition of the first two cysteine residuesadjacent to the N-terminus. Chemokinesand their receptors are either constitutivelyexpressed by some cells or are induced by
inflammatory cytokines, growth factors, andpathogenic stimuli, and mediate the selec-tive recruitment and trafficking of leukocytesubsets (3).
There has been controversy over whetherthe metastatic potential is preprogrammedwithin the primary tumor or that a raremetastatic variant cell selectively and sto-chastically arises within the primary tumorand emanates to regional or distant sites.Immunologists have facilitated our under-standing of lymphocyte migratory and sur-vival molecules, in particular, the findingthat chemokines have important signalingeffects on the invasive and metastatic poten-tial of solid tumors. Chemokine receptorsappear to enable a subset of tumor cells inthe microenvironment to respond to consti-tutive and/or inflammatory chemokine-induced signals (4).
The selective upregulation of CCR7 onmetastatic SCCHN derivatives may explainthe predictable pattern of cervical, nodalmetastasis observed in patients with this dis-ease. The autocrine and/or paracrine secretionof chemokine signals has recently beenobserved, and validates the important role ofCCR7 as a potential oncogene in several tumortypes. The ability of metastatic SCCHNtumors to induce inflammatory nuclear factor-κB (NF-κB) associated CCR7 signals linksinflammatory pathways with CCR7-mediatedinvasion, survival, and metastasis. The role ofNF-κB in propagating an autocrine signalingloop in CCR7-positive SCCHN cells may pro-vide a clinically useful biomarker of diseasestatus and response to therapy. This linkbetween inflammation and carcinogenesisreflects our ongoing understanding of thetumor microenvironment in the progression ofmalignant disease. This review will summa-rize recent findings for CCR7, which appearsto be important in promoting metastaticSCCHN.
Mburu et al.62

From Leukocytes to Tumors: Expression of CC Chemokine Receptor 7
CCR7 is a seven-transmembrane-domain Gprotein–coupled receptor that is important forimmune cell trafficking and homing to thelymph nodes. It is expressed on mature den-dritic cells (DCs), B cells, naïve cells, andsome memory T cells. Its chemokine ligands,CC chemokine ligand 19 (MIP-3β/ELC) andCCL21 (6CKine/SLC), are predominantlyexpressed by stromal cells in lymphoid organsand regulate the recruitment and homing ofCCR7-positive (CCR7+) cells to lymphatictissue. In immune cells, the profile ofchemokine receptor expression is largelydependent on the homeostatic interactionsbetween the cell and its surrounding milieu. Inthe presence of proinflammatory stimuli suchas cytokines, chemokines, and selectins, leuko-cytes are induced to alter their chemokinereceptor expression patterns thereby coordi-nating their maturation and migration out ofperipheral tissues and into the proximal anddistant lymphatics. Chemokine receptors wereinitially described and identified on immunecells; however, it is increasingly apparent thatfunctional chemokine receptors are expressedby some endothelial cells (5) and transformedepithelial cells (4). According to the “chemoat-tractant” theory of tumor invasion, malignantcells that express functional chemokine recep-tors can respond to environmental cuesintended for immune cells, migrate alongchemokine gradients to distant organs, andestablish sites of secondary tumor growth. Thishas been shown experimentally using a murineB16 melanoma model, in which transfectionwith CCR7 allowed tumor cells to migratethrough the lymphatics into the regional lymphnodes (6). Using the same model, other exper-iments have also shown that transfection withCXCR4 leads to enhanced lung metastasis (7)while CCR10 contributes to skin metastases
(8). There is mounting evidence indicatingthat some of these chemokine receptors areselectively expressed in metastatic lesions, fur-ther supporting their role in tumor cell migra-tion and metastasis.
As described above, in SCCHN a pre-dictable pattern of regional cervical lymphnode metastasis is observed in a majority ofpatients at diagnosis. Understanding failure oftherapy against metastatic tumor variantswould be of significant clinical utility. In oursearch for potential predictive biomarkers andtherapeutic targets, we identified anddescribed a distinct pattern of chemokinereceptor expression that is defined by theselective upregulation of CCR7 in metastaticSCCHN cell lines and tumor tissues. Con-versely, autologous primary tumors werefound to express CCR6 and very low (if any)levels of CCR7. Corroboration of these datausing immunohistochemical staining as wellas quantitative reverse-transcription (qRT)PCR on paired tumor biopsies (primary andmetastatic) indicated significantly higherexpression of CCR7 on metastatic tumors ascompared to primary tumor tissue (2). We alsocompared other chemokine receptor expres-sion profiles (CCR1 to CCR10 and CXCR1 toCXCR5) between our primary tumor andmetastatic SCCHN cell lines. Our findings donot exclude the concomitant expression ofother chemokine receptors in addition toCCR7 although none of these has demon-strated a consistent pattern of expression thatdistinguishes the metastatic from primarytumors. Notably, despite reports studying therole of CXCR4 (another nodal homingchemokine receptor) in SCCHN (9,10), wehave not detected any appreciable or consis-tent expression of this receptor on our SCCHNcell lines or tumor tissues. Also, recent evi-dence indicates that CCR7 appears to be astronger mediator of lymph node homing forDCs than CXCR4 (11). This may also hold
NF-κB Mediates Autocrine CCR7Signaling in HNC
63
true in the case of SCCHN tumor cell migra-tion during nodal invasion and metastasis.
A Functional Role for CCR7 in SCCHN Metastasis
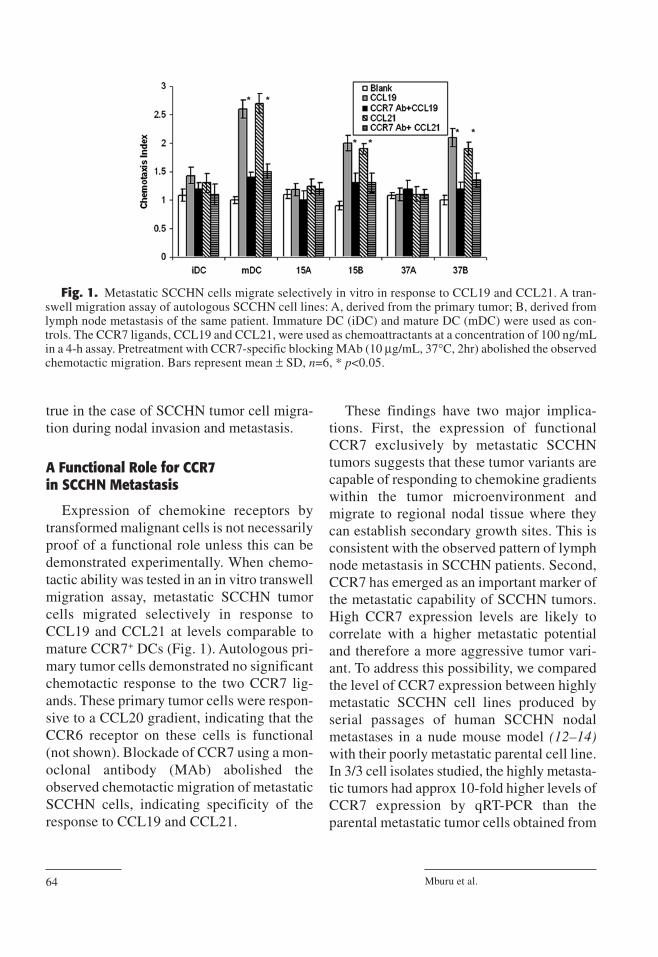
Expression of chemokine receptors bytransformed malignant cells is not necessarilyproof of a functional role unless this can bedemonstrated experimentally. When chemo-tactic ability was tested in an in vitro transwellmigration assay, metastatic SCCHN tumorcells migrated selectively in response toCCL19 and CCL21 at levels comparable tomature CCR7+ DCs (Fig. 1). Autologous pri-mary tumor cells demonstrated no significantchemotactic response to the two CCR7 lig-ands. These primary tumor cells were respon-sive to a CCL20 gradient, indicating that theCCR6 receptor on these cells is functional(not shown). Blockade of CCR7 using a mon-oclonal antibody (MAb) abolished theobserved chemotactic migration of metastaticSCCHN cells, indicating specificity of theresponse to CCL19 and CCL21.
These findings have two major implica-tions. First, the expression of functionalCCR7 exclusively by metastatic SCCHNtumors suggests that these tumor variants arecapable of responding to chemokine gradientswithin the tumor microenvironment andmigrate to regional nodal tissue where theycan establish secondary growth sites. This isconsistent with the observed pattern of lymphnode metastasis in SCCHN patients. Second,CCR7 has emerged as an important marker ofthe metastatic capability of SCCHN tumors.High CCR7 expression levels are likely tocorrelate with a higher metastatic potentialand therefore a more aggressive tumor vari-ant. To address this possibility, we comparedthe level of CCR7 expression between highlymetastatic SCCHN cell lines produced byserial passages of human SCCHN nodalmetastases in a nude mouse model (12–14)with their poorly metastatic parental cell line.In 3/3 cell isolates studied, the highly metasta-tic tumors had approx 10-fold higher levels ofCCR7 expression by qRT-PCR than theparental metastatic tumor cells obtained from
Mburu et al.64
Fig. 1. Metastatic SCCHN cells migrate selectively in vitro in response to CCL19 and CCL21. A tran-swell migration assay of autologous SCCHN cell lines: A, derived from the primary tumor; B, derived fromlymph node metastasis of the same patient. Immature DC (iDC) and mature DC (mDC) were used as con-trols. The CCR7 ligands, CCL19 and CCL21, were used as chemoattractants at a concentration of 100 ng/mLin a 4-h assay. Pretreatment with CCR7-specific blocking MAb (10 µg/mL, 37°C, 2hr) abolished the observedchemotactic migration. Bars represent mean ± SD, n=6, * p<0.05.
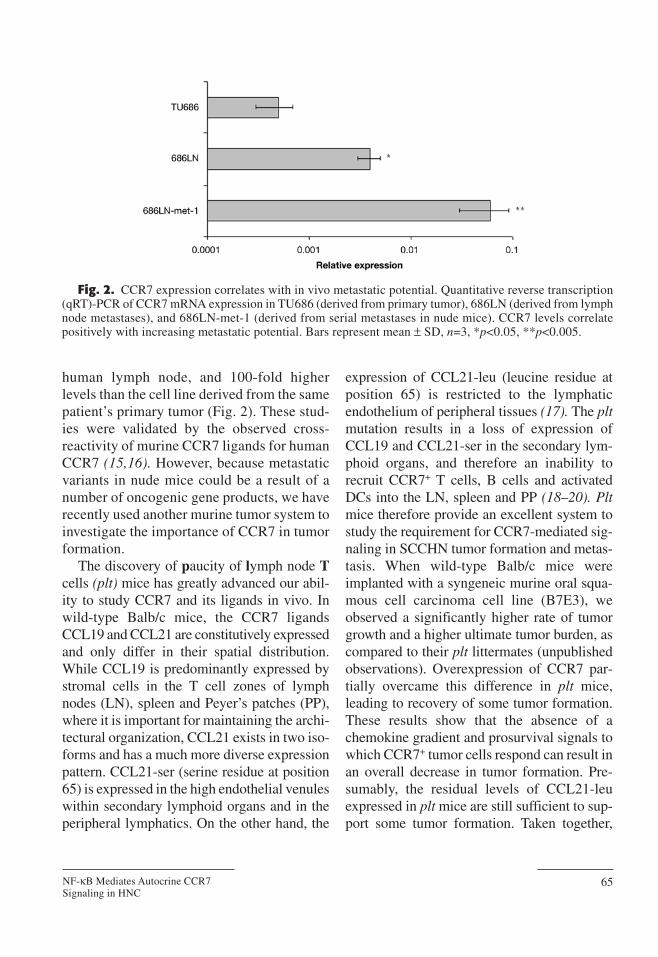
human lymph node, and 100-fold higherlevels than the cell line derived from the samepatient’s primary tumor (Fig. 2). These stud-ies were validated by the observed cross-reactivity of murine CCR7 ligands for humanCCR7 (15,16). However, because metastaticvariants in nude mice could be a result of anumber of oncogenic gene products, we haverecently used another murine tumor system toinvestigate the importance of CCR7 in tumorformation.
The discovery of paucity of lymph node Tcells (plt) mice has greatly advanced our abil-ity to study CCR7 and its ligands in vivo. Inwild-type Balb/c mice, the CCR7 ligandsCCL19 and CCL21 are constitutively expressedand only differ in their spatial distribution.While CCL19 is predominantly expressed bystromal cells in the T cell zones of lymphnodes (LN), spleen and Peyer’s patches (PP),where it is important for maintaining the archi-tectural organization, CCL21 exists in two iso-forms and has a much more diverse expressionpattern. CCL21-ser (serine residue at position65) is expressed in the high endothelial venuleswithin secondary lymphoid organs and in theperipheral lymphatics. On the other hand, the
expression of CCL21-leu (leucine residue atposition 65) is restricted to the lymphaticendothelium of peripheral tissues (17). The pltmutation results in a loss of expression ofCCL19 and CCL21-ser in the secondary lym-phoid organs, and therefore an inability torecruit CCR7+ T cells, B cells and activatedDCs into the LN, spleen and PP (18–20). Pltmice therefore provide an excellent system tostudy the requirement for CCR7-mediated sig-naling in SCCHN tumor formation and metas-tasis. When wild-type Balb/c mice wereimplanted with a syngeneic murine oral squa-mous cell carcinoma cell line (B7E3), weobserved a significantly higher rate of tumorgrowth and a higher ultimate tumor burden, ascompared to their plt littermates (unpublishedobservations). Overexpression of CCR7 par-tially overcame this difference in plt mice,leading to recovery of some tumor formation.These results show that the absence of achemokine gradient and prosurvival signals towhich CCR7+ tumor cells respond can result inan overall decrease in tumor formation. Pre-sumably, the residual levels of CCL21-leuexpressed in plt mice are still sufficient to sup-port some tumor formation. Taken together,
NF-κB Mediates Autocrine CCR7Signaling in HNC
65
Fig. 2. CCR7 expression correlates with in vivo metastatic potential. Quantitative reverse transcription(qRT)-PCR of CCR7 mRNA expression in TU686 (derived from primary tumor), 686LN (derived from lymphnode metastases), and 686LN-met-1 (derived from serial metastases in nude mice). CCR7 levels correlatepositively with increasing metastatic potential. Bars represent mean ± SD, n=3, *p<0.05, **p<0.005.
these results suggest that CCR7-mediated sig-naling may be important not only for thechemotactic migration of invasive CCR7+
tumor cells, but also for the survival of growingtumors. However, because CCR7 expressionhas generally been identified on tumors alreadypresent at metastatic sites, a question stillremains as to the temporal and topologicalimportance of CCR7 expression on metastaticcells emigrating from primary tumors to lymphnodes. In general, elevated CCR7 ligandexpression has been associated with the pres-ence of CCR7+ tumors (21,22).
CCR7 Mediates Tumor Survival and Invasion of Metastatic SCCHN
Recent evidence has reported CCR7 signal-ing to be crucial for providing prosurvival sig-nals to activated DCs and effector CD8+ T cellsthrough the phosphoinositide-3 kinase
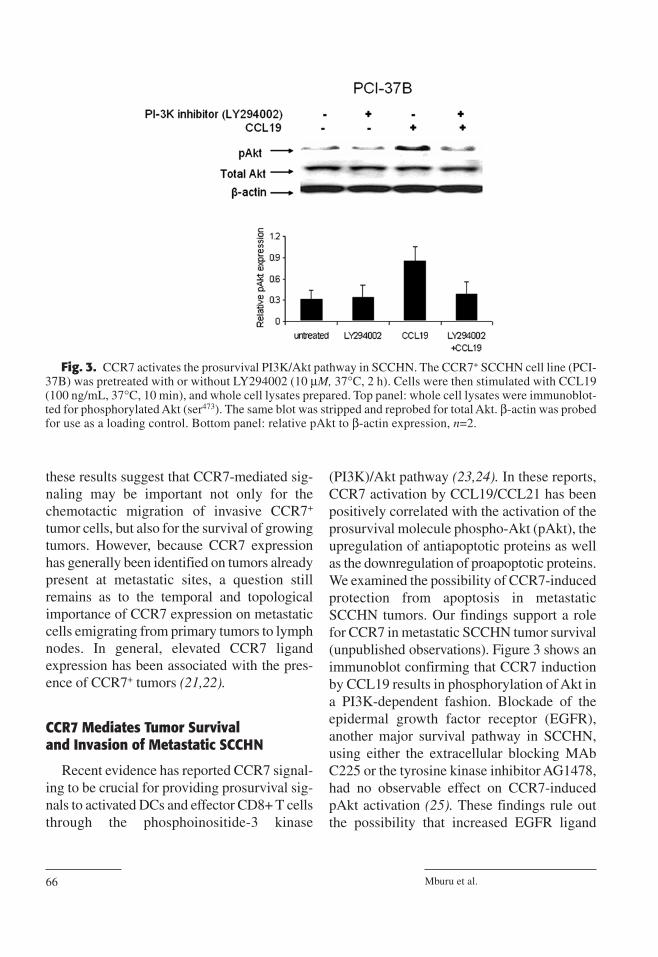
(PI3K)/Akt pathway (23,24). In these reports,CCR7 activation by CCL19/CCL21 has beenpositively correlated with the activation of theprosurvival molecule phospho-Akt (pAkt), theupregulation of antiapoptotic proteins as wellas the downregulation of proapoptotic proteins.We examined the possibility of CCR7-inducedprotection from apoptosis in metastaticSCCHN tumors. Our findings support a rolefor CCR7 in metastatic SCCHN tumor survival(unpublished observations). Figure 3 shows animmunoblot confirming that CCR7 inductionby CCL19 results in phosphorylation of Akt ina PI3K-dependent fashion. Blockade of theepidermal growth factor receptor (EGFR),another major survival pathway in SCCHN,using either the extracellular blocking MAbC225 or the tyrosine kinase inhibitor AG1478,had no observable effect on CCR7-inducedpAkt activation (25). These findings rule outthe possibility that increased EGFR ligand
Mburu et al.66
Fig. 3. CCR7 activates the prosurvival PI3K/Akt pathway in SCCHN. The CCR7+ SCCHN cell line (PCI-37B) was pretreated with or without LY294002 (10 µM, 37°C, 2 h). Cells were then stimulated with CCL19(100 ng/mL, 37°C, 10 min), and whole cell lysates prepared. Top panel: whole cell lysates were immunoblot-ted for phosphorylated Akt (ser473). The same blot was stripped and reprobed for total Akt. β-actin was probedfor use as a loading control. Bottom panel: relative pAkt to β-actin expression, n=2.
secretion or intracellular “cross-talk” betweenEGFR and CCR7 pathways may be responsi-ble for mediating these prosurvival signals.Thus the CCR7-induced prosurvival pathwayappears to be independent of EGFR signaling.
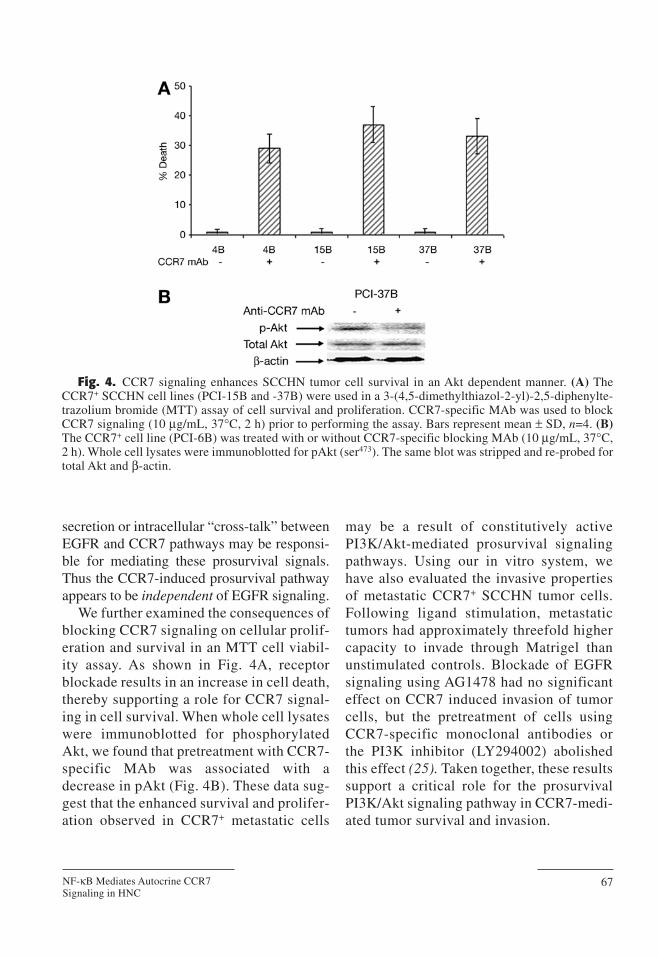
We further examined the consequences ofblocking CCR7 signaling on cellular prolif-eration and survival in an MTT cell viabil-ity assay. As shown in Fig. 4A, receptorblockade results in an increase in cell death,thereby supporting a role for CCR7 signal-ing in cell survival. When whole cell lysateswere immunoblotted for phosphorylatedAkt, we found that pretreatment with CCR7-specific MAb was associated with adecrease in pAkt (Fig. 4B). These data sug-gest that the enhanced survival and prolifer-ation observed in CCR7+ metastatic cells
may be a result of constitutively activePI3K/Akt-mediated prosurvival signalingpathways. Using our in vitro system, wehave also evaluated the invasive propertiesof metastatic CCR7+ SCCHN tumor cells.Following ligand stimulation, metastatictumors had approximately threefold highercapacity to invade through Matrigel thanunstimulated controls. Blockade of EGFRsignaling using AG1478 had no significanteffect on CCR7 induced invasion of tumorcells, but the pretreatment of cells usingCCR7-specific monoclonal antibodies orthe PI3K inhibitor (LY294002) abolishedthis effect (25). Taken together, these resultssupport a critical role for the prosurvivalPI3K/Akt signaling pathway in CCR7-medi-ated tumor survival and invasion.
NF-κB Mediates Autocrine CCR7Signaling in HNC
67
Fig. 4. CCR7 signaling enhances SCCHN tumor cell survival in an Akt dependent manner. (A) TheCCR7+ SCCHN cell lines (PCI-15B and -37B) were used in a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenylte-trazolium bromide (MTT) assay of cell survival and proliferation. CCR7-specific MAb was used to blockCCR7 signaling (10 µg/mL, 37°C, 2 h) prior to performing the assay. Bars represent mean ± SD, n=4. (B)The CCR7+ cell line (PCI-6B) was treated with or without CCR7-specific blocking MAb (10 µg/mL, 37°C,2 h). Whole cell lysates were immunoblotted for pAkt (ser473). The same blot was stripped and re-probed fortotal Akt and β-actin.

CCR7 Downstream Activation of NF-κBCompletes the Autocrine Loop
We noted that untreated cells are able tosustain CCR7-mediated survival even in theabsence of exogenous ligand (Fig. 4A). Fur-thermore, we could detect phosphorylatedAkt in untreated cells, which decreased whencells were pretreated with CCR7-blockingMAb. We reasoned that CCR7+ tumors maybe capable of secreting endogenous ligand,thereby propagating autocrine CCR7 activa-tion. To determine whether there is a basallevel of activation, we assayed cell culturesupernatants by ELISA for the secretion ofchemokine ligands by our SCCHN cell lines.Interestingly, we detected a matching patternof ligand secretion in which primary tumorcell lines expressing CCR6 secrete CCL20,while metastatic cell lines expressing CCR7secrete CCL19. These data have since beenconfirmed and extended by qRT-PCR in bothcell lines and tumor tissues (unpublishedobservations). Furthermore, ligand secretionin metastatic SCCHN cell lines is functionallysignificant, as observed by higher basal levelsof phosphorylated Akt in untreated cells com-pared to cells pretreated with a CCR7-block-ing MAb. This ligand secretion patternsuggests the existence of an autocrine (andparacrine) signaling loop in which chemokinereceptor–expressing tumors are capable ofsecreting and responding to their cognateligand. This autocrine and paracrine produc-tion of chemokines by tumors is similar to thatdescribed in DCs (26), and can contribute tothe malignant transformation of tumor cellsand subsequently their propensity for metas-tasis by providing self-sustaining growth andsurvival signals. Autocrine signaling intumors may also play a role in the selection ofclinically aggressive tumor variants.
The PI3K/Akt signaling pathway has beenreported to lead to the activation of nuclear
factor (NF)-κB, a critical regulator ofinducible gene expression (27,28). Among thegenes regulated by NF-κB are cellular stressresponse genes such as cycloxygenase-2 andinducible nitric oxide synthase; mediators ofimmunity and inflammation such ascytokines, chemokines, proteases, and adhe-sion molecules; and cell cycle regulators suchas c-myc and cyclin D1. These NF-κB–regu-lated factors are important for cell survivaland proliferation. In most cases, NF-κB hasbeen documented to provide cell survival sig-nals, although there have been some reports ofits apoptosis-inducing properties (29,30). Forthese reasons, NF-κB has been the focus ofnumerous recent studies to determine its rolein tumorigenesis and in the malignant trans-formation of cells.
To determine whether CCR7 stimulationby its ligand results in the activation of NF-κBin a metastatic SCCHN cell line, we treatedcells with CCL19 in the presence or absenceof CCR7-blocking antibody and quantifiedNF-κB activation in nuclear extracts using anelectrophoretic mobility shift assay (EMSA)(Fig. 5A). CCL19 stimulation was found toactivate NF-κB to a level comparable to thatinduced by TNF-α. This was specific to theCCR7 receptor as indicated by the ability ofCCR7-specific MAb to block NF-κB activa-tion. The use of an irrelevant ligand (CCL20)did not activate NF-κB. Furthermore, bindingcould be competed away by the addition ofexcess unlabeled but not an irrelevant probe,showing specificity of NF-κB binding.
In the classical pathway of NF-κB activa-tion, IκB keeps NF-κB sequestered and inac-tive in the cytoplasm. Upon phosphorylationby the IκB kinases IKK-1 and IKK-2, IκB istargeted for ubiquitination and proteasomaldegradation, subsequently liberating activeNF-κB, which translocates to the nucleus toregulate target gene expression (31,32). TheIκBβ super-repressor contains Ser to Ala
Mburu et al.68
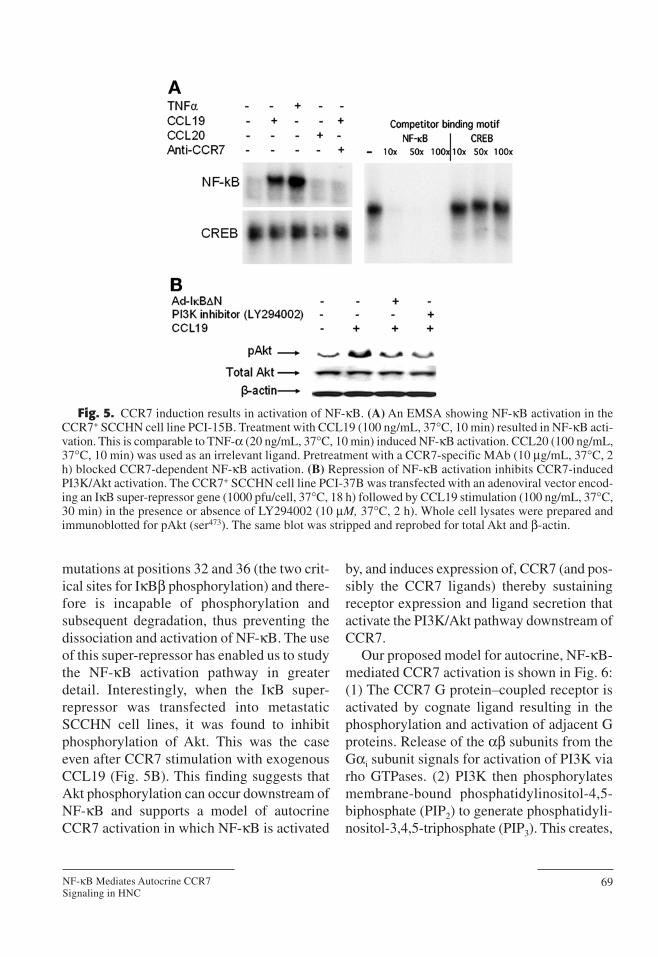
mutations at positions 32 and 36 (the two crit-ical sites for IκBβ phosphorylation) and there-fore is incapable of phosphorylation andsubsequent degradation, thus preventing thedissociation and activation of NF-κB. The useof this super-repressor has enabled us to studythe NF-κB activation pathway in greaterdetail. Interestingly, when the IκB super-repressor was transfected into metastaticSCCHN cell lines, it was found to inhibitphosphorylation of Akt. This was the caseeven after CCR7 stimulation with exogenousCCL19 (Fig. 5B). This finding suggests thatAkt phosphorylation can occur downstream ofNF-κB and supports a model of autocrineCCR7 activation in which NF-κB is activated
by, and induces expression of, CCR7 (and pos-sibly the CCR7 ligands) thereby sustainingreceptor expression and ligand secretion thatactivate the PI3K/Akt pathway downstream ofCCR7.
Our proposed model for autocrine, NF-κB-mediated CCR7 activation is shown in Fig. 6:(1) The CCR7 G protein–coupled receptor isactivated by cognate ligand resulting in thephosphorylation and activation of adjacent Gproteins. Release of the αβ subunits from theGαi subunit signals for activation of PI3K viarho GTPases. (2) PI3K then phosphorylatesmembrane-bound phosphatidylinositol-4,5-biphosphate (PIP2) to generate phosphatidyli-nositol-3,4,5-triphosphate (PIP3). This creates,
NF-κB Mediates Autocrine CCR7Signaling in HNC
69
Fig. 5. CCR7 induction results in activation of NF-κB. (A) An EMSA showing NF-κB activation in theCCR7+ SCCHN cell line PCI-15B. Treatment with CCL19 (100 ng/mL, 37°C, 10 min) resulted in NF-κB acti-vation. This is comparable to TNF-α (20 ng/mL, 37°C, 10 min) induced NF-κB activation. CCL20 (100 ng/mL,37°C, 10 min) was used as an irrelevant ligand. Pretreatment with a CCR7-specific MAb (10 µg/mL, 37°C, 2h) blocked CCR7-dependent NF-κB activation. (B) Repression of NF-κB activation inhibits CCR7-inducedPI3K/Akt activation. The CCR7+ SCCHN cell line PCI-37B was transfected with an adenoviral vector encod-ing an IκB super-repressor gene (1000 pfu/cell, 37°C, 18 h) followed by CCL19 stimulation (100 ng/mL, 37°C,30 min) in the presence or absence of LY294002 (10 µM, 37°C, 2 h). Whole cell lysates were prepared andimmunoblotted for pAkt (ser473). The same blot was stripped and reprobed for total Akt and β-actin.
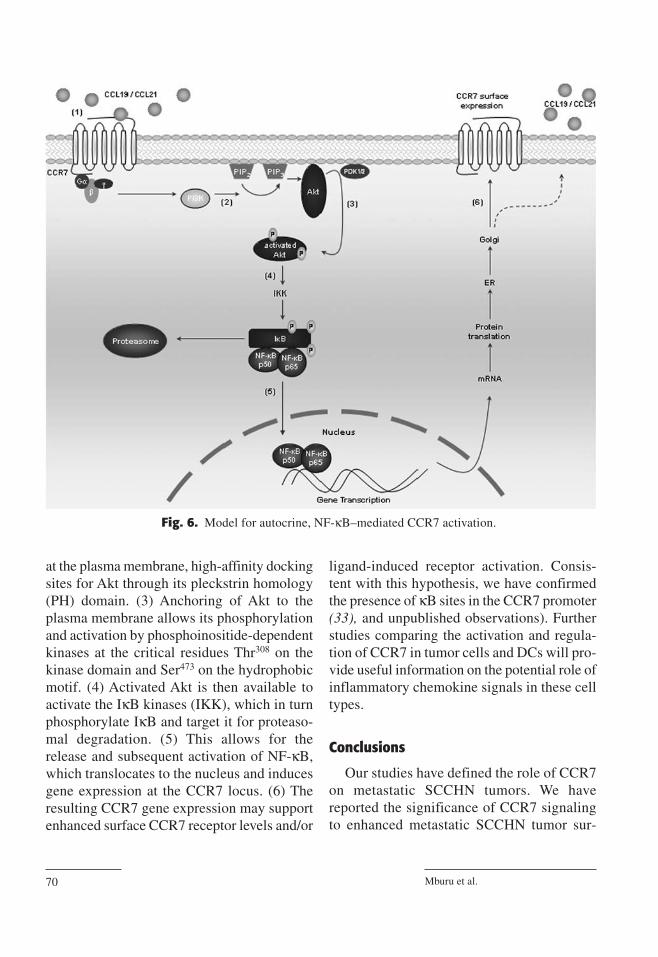
at the plasma membrane, high-affinity dockingsites for Akt through its pleckstrin homology(PH) domain. (3) Anchoring of Akt to theplasma membrane allows its phosphorylationand activation by phosphoinositide-dependentkinases at the critical residues Thr308 on thekinase domain and Ser473 on the hydrophobicmotif. (4) Activated Akt is then available toactivate the IκB kinases (IKK), which in turnphosphorylate IκB and target it for proteaso-mal degradation. (5) This allows for therelease and subsequent activation of NF-κB,which translocates to the nucleus and inducesgene expression at the CCR7 locus. (6) Theresulting CCR7 gene expression may supportenhanced surface CCR7 receptor levels and/or
ligand-induced receptor activation. Consis-tent with this hypothesis, we have confirmedthe presence of κB sites in the CCR7 promoter(33), and unpublished observations). Furtherstudies comparing the activation and regula-tion of CCR7 in tumor cells and DCs will pro-vide useful information on the potential role ofinflammatory chemokine signals in these celltypes.
Conclusions
Our studies have defined the role of CCR7on metastatic SCCHN tumors. We havereported the significance of CCR7 signalingto enhanced metastatic SCCHN tumor sur-
Mburu et al.70
Fig. 6. Model for autocrine, NF-κB–mediated CCR7 activation.

vival, invasion, and migration to regionallymph nodes. Furthermore, our findings sup-port a role for autocrine NF-κB mediated sig-nals in maintaining CCR7 signaling.
CCR7 expression on transformed andmetastatic cells has also been described in anumber of other malignancies includingbreast cancer (4), chronic lymphocyticleukemia (34), gastric carcinoma (35), non-small-cell lung cancer (36), and esophagealsquamous carcinoma (37). In all studiedcases, expression of CCR7 by tumors corre-lates with a high metastatic potential andpoor prognosis. Results from our lab andothers support a role for CCR7 in the lym-phatic migration of tumor cells in a mannerthat is reminiscent of DC immigration intodraining lymph nodes. It is likely that tumorcells expressing functional chemokine recep-tors hijack physiological chemokine-directedmechanisms of leukocyte migration duringmetastasis. This underscores the critical roleof the tumor microenvironment duringimmune cell–mediated inflammation in shap-
ing the repertoire of chemokine expression onthe immune cell infiltrate and possibly on thedeveloping tumor. Recent studies havedescribed a range of mechanisms that pro-gram malignant cells to express functionalchemokine receptors (reviewed in ref. 38). Inparticular, constitutive NF-κB activity hasbeen shown to be important for CCR7 upreg-ulation in classical Hodgkin’s disease (33,39).Our own data in SCCHN are consistent witha critical role for NF-κB in the regulation ofCCR7 expression and downstream signaling(unpublished observations).
In conclusion, we have identified CCR7-mediated signaling as a potential pathway forclinical intervention in the dissemination ofmetastatic SCCHN tumors. Furthermore,there appears to be a role for inflammatoryautocrine signals in CCR7-induced NF-κBactivation. Further work may enable the con-trol of tumor progression and metastasis byextending our understanding CCR7-mediatedtumor growth, survival and invasion.
NF-κB Mediates Autocrine CCR7Signaling in HNC
71
References1. Paget S: The distribution of secondary growths in cancer
of the breast. Lancet 1889;1:571–573.2. Wang J, Xi L, Hunt JL, et al: Expression pattern of
chemokine receptor 6 (CCR6) and CCR7 in squamouscell carcinoma of the head and neck identifies a novelmetastatic phenotype. Cancer Res 2004;64:1861–1866.
3. Zlotnik A, Yoshie O: Chemokines: A new classificationsystem and their role in immunity. Immunity 2000;12:121–127.
4. Muller A, Homey B, Soto H, et al: Involvement ofchemokine receptors in breast cancer metastasis. Nature2001;410:50–56.
5. Gupta SK, Lysko PG, Pillarisetti K, Ohlstein E, StadelJM: Chemokine receptors in human endothelial cells.Functional expression of CXCR4 and its transcriptionalregulation by inflammatory cytokines. J Biol Chem1998;273:4282–4287.
6. Wiley HE, Gonzalez EB, Maki W, Wu M-t, Hwang ST:Expression of CC chemokine receptor-7 and regional
lymph node metastasis of B16 murine melanoma. J NatlCancer Inst 2001;93:1638–1643.
7. Murakami T, Maki W, Cardones AR, et al: Expression ofCXC chemokine receptor-4 enhances the pulmonarymetastatic potential of murine B16 melanoma cells.Cancer Res 2002;62:7328–7334.
8. Murakami T, Cardones AR, Finkelstein SE, et al:Immune evasion by murine melanoma mediated throughCC chemokine receptor-10. J Exp Med 2003;198:1337–1347.
9. Katayama A, Ogino T, Bandoh N, Nonaka S, HarabuchiY: Expression of CXCR4 and its down-regulation byIFN-gamma in head and neck squamous cell carcinoma.Clin Cancer Res 2005;11:2937–2946.
10. Ishikawa T, Nakashiro. K, Hara S, et al: CXCR4 expres-sion is associated with lymph-node metastasis of oralsquamous cell carcinoma. Int J Oncol 2006;28:61–66.
11. Humrich JY, Humrich JH, Averbeck M, et al: Maturemonocyte-derived dendritic cells respond more strongly

to CCL19 than to CXCL12: consequences for directionalmigration. Immunology 2006;117:238–247.
12. Sacks PG: Cell, tissue and organ culture as in vitromodels to study the biology of squamous cell carcino-mas of the head and neck. Cancer Metastasis Rev1996;15:27–51.
13. Zhang X, Liu Y, Gilcrease MZ, et al: A lymph nodemetastatic mouse model reveals alterations of metasta-sis-related gene expression in metastatic human oral car-cinoma sublines selected from a poorly metastaticparental cell line. Cancer (Phila) 2002;95:1663–1672.
14. Chen Z, Zhang K, Zhang X, et al: Comparison of geneexpression between metastatic derivatives and theirpoorly metastatic parental cells implicates crucial tumor-environment interaction in metastasis of head and necksquamous cell carcinoma. Clin Exp Metastasis2003;20:335–342.
15. Campbell JJ, Hedrick J, Zlotnik A, Siani MA, Thomp-son DA, Butcher EC: Chemokines and the arrest of lym-phocytes rolling under flow conditions. Science1998;279:381–384.
16. Dellacasagrande J, Schreurs OJ, Hofgaard PO, et al:Liver metastasis of cancer facilitated by chemokinereceptor CCR6. Scand J Immunol 2003;57:534–544.
17. Luther SA, Tang HL, Hyman PL, Farr AG, Cyster JG:Coexpression of the chemokines ELC and SLC by Tzone stromal cells and deletion of the ELC gene in theplt/plt mouse. Proc Natl Acad Sci USA2000;97:12694–12699.
18. Nakano H, Mori S,Yonekawa H, Nariuchi H, MatsuzawaA, Kakiuchi T: A novel mutant gene involved in T-lym-phocyte-specific homing into peripheral lymphoidorgans on mouse chromosome 4. Blood 1998;91:2886–2895.
19. Vassileva G, Soto H, Zlotnik A, et al: The reducedexpression of 6Ckine in the plt mouse results from thedeletion of one of two 6Ckine genes. J Exp Med1999;190:1183–1188.
20. Gunn MD, Kyuwa S, Tam C, et al: Mice lacking expres-sion of secondary lymphoid organ chemokine havedefects in lymphocyte homing and dendritic cell local-ization. J Exp Med 1999;189:451–460.
21. Takeuchi H, Fujimoto A, Tanaka M,Yamano T, Hsueh E,Hoon DSB: CCL21 chemokine regulates chemokinereceptor CCR7 bearing malignant melanoma cells. ClinCancer Res 2004;10:2351–2358.
22. Wang J, Xi L, Gooding W, Godfrey TE, Ferris RL:Chemokine receptors 6 and 7 identify a metastaticexpression pattern in squamous cell carcinoma of thehead and neck. Adv Otorhinolaryngol 2005;62:121–133.
23. Sanchez-Sanchez N, Riol-Blanco L, de la Rosa G, et al:Chemokine receptor CCR7 induces intracellular signal-ing that inhibits apoptosis of mature dendritic cells.Blood 2004;104:619–625.
24. Kim J-W, Ferris RL, Whiteside TL: Chemokine C recep-tor 7 expression and protection of circulating CD8+ Tlymphocytes from apoptosis. Clin Cancer Res 2005;11:7901–7910.
25. Wang J, Zhang X, Thomas SM, et al: Chemokine recep-tor 7 activates phosphoinositide-3 kinase-mediated inva-sive and prosurvival pathways in head and neck cancercells independent of EGFR. Oncogene 2005;24:5897–5904.
26. Kaiser A, Donnadieu E, Abastado J-P, Trautmann A,Nardin A: CC chemokine ligand 19 secreted by maturedendritic cells increases naive T cell scanning behaviorand their response to rare cognate antigen. J Immunol2005;175:2349–2356.
27. Nidai Ozes O, Mayo LD, Gustin JA, Pfeffer SR, PfefferLM, Donner DB: NF-[kappa]B activation by tumournecrosis factor requires the Akt serine-threonine kinase.Nature 1999;401:82.
28. Kane LP, Shapiro VS, Stokoe D, Weiss A: Induction ofNF-[kappa]B by the Akt/PKB kinase. Curr Biol1999;9:601.
29. Karin M, Ben-Neriah Y: Phosphorylation meets ubiqui-tination: the control of NF-[kappa]B activity. Ann RevImmunol 2000;18:621–663.
30. Bours V, Bentires-Alj M, Hellin A-C, et al: Nuclearfactor-[kappa]B, cancer, and apoptosis. Biochem Phar-macol 2000;60:1085–1089.
31. Karin M: The Beginning of the End: Ikappa B kinase(IKK) and NF-kappa B activation. J Biol Chem1999;274:27339–27342.
32. Karin M, Greten FR: NF-[kappa]B: linking inflamma-tion and immunity to cancer development and progres-sion. Nat Rev Immunol 2005;5:749–759.
33. Hinz M, Lemke P, Anagnostopoulos I, et al: Nuclearfactor {kappa}B-dependent gene expression profiling ofHodgkin’s disease tumor cells, pathogenetic signifi-cance, and link to constitutive signal transducer and acti-vator of transcription 5a activity. J Exp Med 2002;196:605–617.
34. Till KJ, Lin K, Zuzel M, Cawley JC: The chemokinereceptor CCR7 and alpha 4 integrin are important formigration of chronic lymphocytic leukemia cells intolymph nodes. Blood 2002;99:2977–2984.
35. Mashino K, Sadanaga N,Yamaguchi H, et al: Expressionof chemokine receptor CCR7 is associated with lymphnode metastasis of gastric carcinoma. Cancer Res2002;62:2937–2941.
36. Takanami I: Overexpression of CCR7 mRNA in nons-mall cell lung cancer: Correlation with lymph nodemetastasis. International Journal of Cancer 2003;105:186–189.
37. Ding Y, Shimada Y, Maeda M, et al: Association of CCchemokine receptor 7 with lymph node metastasis ofesophageal squamous cell carcinoma. Clin Cancer Res2003;9:3406–3412.
38. Balkwill F: Cancer and the chemokine network. Nat RevCancer 2004;4:540.
39. Hopken UE, Foss H-D, Meyer D, et al: Up-regulation ofthe chemokine receptor CCR7 in classical but not inlymphocyte-predominant Hodgkin disease correlateswith distinct dissemination of neoplastic cells in lym-phoid organs. Blood 2002;99:1109–1116.
Mburu et al.72




















