
Examining the Progression of Ecological Theory Through ...
82
Examining the Progression of Ecological Theory Through Arachnid Recovery of Mount St. Helens: Why Panarchy is Next in the Successional Line Katherine Fiedler ENVS Program Lewis & Clark College 2011 Abstract: Ecological theories attempt to make sense of the noise of ecological systems. The way we consider the recovery of ecosystems following a disturbance has changed over time, progressing from Clementsian succession to alternative stable states and now to panarchy. In this evolution of thought, the notions of recovery, stability, and resilience have also changed. Panarchy acts as a paradigm presenting new considerations regarding these concepts, and in turn powerful implications for conservation and management. In order to fully grasp the implications of these theories, we must consider them in the context of a living ecological system. When applied to a quantitative analysis of arachnid recovery of Mount St. Helens following the 1980 eruption, we can begin to see how each theory considers recovering systems differently. The same data elicit different understandings depending on the language and framework that are applied. Through this analysis, I have illuminated how panarchy can further our thought regarding ecological recovery.
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of Examining the Progression of Ecological Theory Through ...

Examining the Progression of Ecological Theory Through
Arachnid Recovery of Mount St. Helens: Why Panarchy is Next
in the Successional Line
Katherine Fiedler ENVS Program
Lewis & Clark College 2011
Abstract:
Ecological theories attempt to make sense of the noise of ecological systems. The way we consider the recovery of ecosystems following a disturbance has changed over time, progressing from Clementsian succession to alternative stable states and now to panarchy. In this evolution of thought, the notions of recovery, stability, and resilience have also changed. Panarchy acts as a paradigm presenting new considerations regarding these concepts, and in turn powerful implications for conservation and management. In order to fully grasp the implications of these theories, we must consider them in the context of a living ecological system. When applied to a quantitative analysis of arachnid recovery of Mount St. Helens following the 1980 eruption, we can begin to see how each theory considers recovering systems differently. The same data elicit different understandings depending on the language and framework that are applied. Through this analysis, I have illuminated how panarchy can further our thought regarding ecological recovery.

Fiedler 2
Table of Contents 1. Introduction 4
1.1. Mount St. Helens as a Model System 5 1.2. Ecological Theories of System Recovery 7 1.3. Argument 9
2. Music from the Noise: Reviews of Ecological Theories 10 2.1. Review of Clementsian Successional Theory 10
2.1.1. Successional Theory in the Field of Ecology 13 2.1.2. Critiques of Clementsian Succession 13
2.2. Review of Alternative Stable State Theory 14 2.3. Review of Panarchy 19 2.4. Comparing the Theories 26
2.4.1. How These Theories Will Be Tested 27 3. Listening to the Landscape: A Case Study 29
3.1. Arachnid Recovery of Mount St. Helens 31 3.1.1. Arachnid Sampling Methodology 32
3.1.1.1. Limitations of Sampling Methodology 37 3.1.2. Arachnid Dispersal Methods 39 3.1.3. Spider Feeding Guilds 42 3.1.4. Spider Habitat Preferences 43 3.1.5. Results 44
3.1.5.1. Total number of taxa 45 3.1.5.2. Ballooners 46 3.1.5.3. Feeding Guilds 48 3.1.5.4. Habitat Preferences 50 3.1.5.5. Representative Patterns 53
3.1.5.5.1. Higher Abundance in Less Disturbed Sites 54 3.1.5.5.2. Higher Abundance in More Disturbed Sites 55 3.1.5.5.3. Similar Distribution Across All Research Sites 56 3.1.5.5.4. Successful invaders 57
3.1.6. First on the Scene 58 3.1.7. Comparison to Beetle Data 60
4. When Theories and Data Collide 60 4.1. Deconstructing the Theories in Context 61
4.1.1. The Clementsian Succession Perspective 61 4.1.2. The Alternative Stable States Perspective 63

Fiedler 3
4.1.3. The Panarchy Perspective 65 4.1.4. Summary of Perspectives 66
4.2. Spotlight on Panarchy 67 5. Putting Theory into Practice: Conservation Implications 68
5.1. Collapsing Panarchy 69 5.2. Conservation Decisions 70 5.3. Panarchy in Action 72 5.4. Panarchy as Part of Continuing Paradigm Shift 73 5.5. Conclusion 74
6. References 76 Appendices: Appendix 1. Summary Table of Individuals 79 Appendix 2. Family Distribution of 1981-‐1985 Pumice Plain Spider Samples 81 Appendix 3. Distribution of Individuals per Family: 1990, 2000, and 2010 82

Fiedler 4
1. Introduction
We all use theories and models to understand variation and change, to make
predictions, and to better inform our actions. Ecologists use them to make sense of systems
lacking borders in time or place, in which relevant variables seem infinite and much is due to
randomness or endless cascades of response. Perhaps ecologists are envious of those sciences
that have the luxury of laws rather than highly contested theories; however Daniel Simberloff
notes that, “What physicists view as noise is music to the ecologist.”1 While they may embrace
this complexity, ecologists do try to create some order from the noise, although even the
theories themselves cannot be described as static.
Understanding community response to disturbance is one such example. Ecologists have
attempted to quantify recovery through complex mathematical models or to simply visualize
such change with ball-‐in-‐cup diagrams. The last century witnessed the progression of ecological
thought in this field, from Clementsian succession to alternative stable state theory, and now,
to panarchy. This progression reflects perhaps a successional theory of theories, in which each
builds upon the last, and thus is not wholly independent. Thomas Kuhn details this notion in his
discussion of scientific revolutions as paradigm shifts. He wrote, “…that in sciences fact and
1 Daniel Simberloff, A Succession of Paradigms in Ecology: Essentialism to Materialism and Probabilism, p.
63-‐99, In: Esa Saarinen ed., 1982, Conceptual Issues in Ecology, Boston, MA: D. Reidel Publishing Company, 85.

Fiedler 5
theory, discovery and invention, are not categorically and permanently distinct, we can
anticipate overlap between this section and the last.”2 It is only the most surprising scientific
discoveries that do not overlap with those that came before.3 The shift between Clementsian
succession, alternative stable states, and panarchy can also be described as a paradigm shift.
These theories have become increasingly relevant in our desire to understand how to treat
systems that we ourselves have disturbed. However, these theories, along with any other for
that matter, are useless unless they can properly handle the complexities of real living systems.
In order to fully understand these theories and their applications we must see if they can be
applied to a real ecological disturbance, for “Landscapes tell stories, if we know how to listen.”4
The 1980 eruption of Mount St. Helens can tell us such stories.
1.1. Mount St. Helens as a Model System
Mount St. Helens is an active stratovolcano in the Cascade Mountain range of western
Washington.5 On May 18, 1980, Mount St. Helens erupted and left behind a distinct
2 Thomas Kuhn, 1970, The Structure of Scientific Revolutions, Chicago, IL: The University of Chicago Press,
66. 3 Ibid., 66. 4 Charles Goodrich, Kathleen Dean Moore, and Frederick J. Swanson, eds., 2008, In the blast zone:
catastrophe and renewal on Mount St. Helens, Corvallis, OR: Oregon State University Press, ix. 5 Rodney L. Crawford, Patrick M. Sugg, and John S. Edwards, 1995, “Spider arrival and primary
establishment on terrain depopulated by volcanic eruption at Mount St. Helens, Washington,” American Midland
Naturalist 133 (1): 60-‐75, 61.

Fiedler 6
disturbance gradient amidst an unrecognizable landscape. The range of volcanic forces,
landslides, pyroclastic flows (hot gas and rock), tephra (volcanic ash and rock) fall, and the blast
itself, created a patchwork of environments altered to different degrees. The pyroclastic flow
completely scoured and sterilized the immediate landscape. The force of the blast leveled trees
and scorched the vegetation as far as seventeen miles away. Even further away, tephra rained
down onto forests, covering the ground and low-‐lying vegetation.6 After the eruption, much of
the landscape appeared to be lifeless. However, the barren landscape was soon overcome as
surviving pockets of life and the recolonization of many populations began to transform the
land.
Mount St. Helens has become a working field laboratory for researchers in almost every
biological and geophysical field.7 The ecological recovery following the eruption has provided
information regarding the recovery of populations and entire ecosystems following
disturbances. Congress passed the National Volcanic Monument Act in 1982, creating an
110,000-‐acre national monument, within which the recovery of these systems could occur
without human interference or exploitation. An ideal natural experiment of ecosystem recovery
had begun. In just one day, Mount St. Helens became a model system for the observation of
6 Goodrich et al. eds. 2008, x.
7 Virginia H. Dale, Frederick J. Swanson, and Charles M. Crisafulli, eds., 2005, Ecological Responses to the
1980 Eruption of Mount St. Helens, New York, NY: Springer.

Fiedler 7
countless environmental processes, including the recovery of ecosystems following a range of
disturbances.
1.2. Ecological Theories of System Recovery
Ecological theories have long aimed to describe complex living systems that have
continually evaded the confines of strict patterns and equations. Many ecological theories
attempt to describe ecosystem recovery, or the trajectory of ecosystem response toward its
previous state or some newly organized state, following a disturbance. Over time, the theories
themselves have evolved to acknowledge and incorporate more complexity and new
interpretations of stability, while still aiming to be broadly applicable.
The following is only a brief introduction to the three theories I will be considering, as I
will explain each in detail shortly thereafter. Ecological succession, in short, was the first of such
theories regarding community change over time, describing a linear and predictable pattern of
recovery toward a single and repeatable climax state.8 However, ecologists have moved away
from this notion of a predictable trajectory toward a monoclimax in favor of alternative stable
states theory. The theory of alternative stable states focuses on the potential for systems to
settle in different states of stability, triggered by an internal or external disturbance. This theory
8 Frederic E. Clements, 1916, Plant Succession, an Analysis of the Development of Vegetation, p. 140-‐143,
In: Edward J. Kormondy, ed., 1965, Readings in Ecology, Englewood Cliffs, NJ: Prentice-‐Hall, Inc., 140.

Fiedler 8
still focuses on these stable states themselves.9 Panarchy branches from alternative stable
states theory describing systems as being constantly in flux. Community change within systems
can be viewed as series of dynamic cycles of different temporal and spatial scales. Disturbances
act as a trigger to prompt these cycles to either continue on their same adaptive cycles or to
allow for novelty.10 This theory has yet to take hold in ecological discourse; however it may
present us with new ways of discussing the responses of living systems to ecological
disturbances.
We can better understand these theories when we consider them in the context of real
living systems. It is only then that we can see exactly what they focus on and what they largely
ignore. The disturbance zones of Mount St. Helens provide a context through which we can
consider the theory of panarchy and its place in ecological theory. Within these communities, it
is helpful to center our focus on specific taxonomic groups in order to properly manage the
complexity of ecosystems as a whole. Arthropods play important roles in the structuring of
communities. They can provide information regarding the vegetation and other organisms
9 B. E. Beisner, D. T. Haydon, and K. Cuddington, 2003, “Alternative stable states in ecology,” Frontiers in
Ecology and the Environment 1 (7), 376. 10 C.S. Holling, 2001, “Understanding the Complexity of Economic, Ecological, and Social Systems,”
Ecosystems, 4:390-‐405, 390.

Fiedler 9
present in a system by considering their prey items, predators, and preferred habitats.11
Ecological theories of recovery are traditionally considered through investigations of plant
communities, looking at the base of food chains, which act as a necessary predecessor to
consumers. Consumers can prove to be equally valuable in these analyses, as they reflect both
the presence of vegetation and predators of themselves. Consumers might show slow recovery
initially, followed by a rapid influx of taxa following the establishment of plant communities.
However, the diversity and complexity of consumer species interactions might result in a
recovery quite similar to that of plants, in terms of niche diversity. For the purpose of my
analysis, I consider ecosystem recovery with a focus on arachnids.
1.3. Argument
The different theories of ecosystem recovery are not correct or incorrect, but rather
they may or may not be sufficient to explain the recovery of ecological systems. They may also
prove to illuminate certain aspects of the process of recovery while ignoring others, altering our
perceptions of stability and conservation. In this paper, I focus on the language and modeling of
panarchy as a paradigm describing the dynamism and recovery of ecosystems, as compared to
11 Robert R. Parmenter et al., 2005, Posteruption Arthropod Succession on the Mount St. Helens Volcano:
The Ground-‐Dwelling Beetle Fauna (Coleoptera), p. 139-‐150, In: Virginia H. Dale, Frederick J. Swanson, and Charles
M. Crisafulli eds., 2005, Ecological Responses to the 1980 Eruption of Mount St. Helens, New York, NY: Springer,
139.

Fiedler 10
Clementsian succession and alternative stable states. I will apply these theories to the recovery
of arachnids in the disturbance zones of Mount St. Helens following the 1980 eruption. The
theory of panarchy focuses on the dynamism of ecological systems and challenges the idea of
stability, serving as the next step in this ecological paradigm shift. Furthermore, due to its new
and relevant perspectives on conservation, panarchy should be considered in environmental
discourse and its subsequent application.
2. Music from the Noise: Reviews of Ecological Theories
In order to apply these theories to empirical data, we must first understand their
history, framework, and language. I will present these theories independently of each other and
of the Mount St. Helens arachnid case study in order to first focus on the applicability of these
theories as they were conceived. Even though each theory describes distinct ecological
processes, while often not addressing them directly, I will avoid augmenting these theories with
other ecological discourse. This will display each theory as it is presented in its founding
discourse and subsequent evolution.
2.1. Review of Clementsian Successional Theory
Succession, in its simplest form, means the occurrence of events in a particular temporal
or spatial sequence. The term was first used in an ecological context by a French biologist,
Adolphe Dureau de la Malle in 1825. However, the idea of ecological succession is said to be

Fiedler 11
much older, perhaps dating as far back as the scientific wonderings of William King in 1685.12 It
was not until Frederic Clements published Plant Succession: An Analysis of the Development of
Vegetation in 1916 that succession became widely accepted among ecologists.13 Clements
described succession as a series of plant communities, or seres, that culminate in a stable
monoclimax. Each sere is progressively more complex and constructed of life forms of higher
trophic levels that might also require more intricate habitats. The process is an explicitly
deterministic response to some disturbance, as each sere is a step in the development of this
climax formation.14 In Clements’ own words:
As an organism the formation arises, grows, matures, and dies…Furthermore, each climax formation is able to reproduce itself, repeating with essential fidelity the stages of its development. The life-‐history of a formation is a complex but definite process, comparable in its chief feature with the life-‐history of an individual plant.15
Clements believed that, like the development of an organism, an ecosystem experiences
distinct life stages that can be replicated. Each time a certain system recovers from a
disturbance, it will follow the same path to its climax state.
12 Frank B. Golley ed., 1977, Ecological succession, Stroudsburg, Pa: Dowden, Hutchinson & Ross, 1. 13 William H. Drury and Ian C.T. Nisbet, 1973, Succession, p. 287-‐368. In: Frank B. Golley, ed., 1977,
Ecological Succession, Stroudsburg, PA: Dowden, Hutchinson & Ross, 289. 14 Clements, 1916, 140-‐141. 15 Clements 1916, 140.

Fiedler 12
A system at the climax state resembles an organism of the highest order and
complexity.16 The system functions in the most energy efficient manner, in terms of biomass
and organismal interactions.17 According to Clements, the path to reach this state is predictable
and replicable, each step dependent on the last. Furthermore, Clements claimed that climate
determines the specific characteristics of these climax states through the large and constant
control that it has over the system. For each unique climatic region, there must only exist one
climax state. If any other state appears within that region, it must be that the system has not
yet reached this most mature and complex stage.18 Yet inevitably, the system will reach its
climax state. As Worster described, “…eventually nature will find a way to get back on track.”19
Once the climax state is reached, assuming external conditions remain constant, it can remain
there indefinitely,20 or as Clements, himself, wrote, “Such a climax is permanent because of its
entire harmony with a stable habitat.”21 He even goes so far as to say that the term stabilization
is synonymous with succession.22
16 Donald Worster, 1994, Nature's economy: a history of ecological ideas, Cambridge: Cambridge
University Press, 210. 17 Eugene P. Odum, 1969, The Strategy of Ecosystem Development, p. 278-‐286, In: Frank B. Golley ed.,
1997, Ecological Succession, Stroudsburg, PA: Dowden, Hutchinson & Ross, 278-‐279. 18 Worster 1994, 210. 19 Ibid., 210. 20 Ibid., 210. 21 Clements 1916, 143. 22 Ibid., 142.

Fiedler 13
2.1.1. Successional Theory in the Field of Ecology
Clementsian succession integrated the ideas behind many ecological processes.
Clements explains the progression from the initial disturbance to the final monoclimax as, “…(1)
nudation, (2) migration, (3) ecesis [or establishment of local populations], (4) competition, (5)
reaction, (6) stabilization.”23 Community ecologists have since detailed out these processes,
with the ideas of primary succession, in which the disturbed site holds no memory of the
previous community, and secondary succession, in which ecosystems still contain pieces of the
old community structure. Furthermore, Clementsian thought echoes the concept of
dominance-‐controlled communities, or the process of competition, where only a few dominant
species out of the colonizers successfully remain in an area.24
2.1.2. Critiques of Clementsian Succession
While much of Clementsian succession still dominates ecological thought, the idea of a
monoclimax was met with much resistance. Ecologists contested that due to the inherent
variation of environmental factors—such as soil, microclimate, geography, and topography—it
is unlikely that only one climax state can be attributed to a climatic region. Instead, we might
23 Ibid., 141. 24 Michael Begon, Colin R. Townsend, and John L. Harper, 2006, Ecology: from individuals to ecosystems,
Malden, MA: Blackwell Pub, 489.

Fiedler 14
observe a continuum of climax states.25 H.A. Gleason, one of Clementsian succession’s
strongest critics, stated that, “…succession is an extraordinarily mobile phenomenon whose
processes are not to be stated as fixed laws, but only as general principles of exceedingly broad
nature, and whose results need not, and frequently do not, ensue in any definitely predictable
way.”26 Other critics noted the role of chance in community development. An organism must
not only disperse into an area, but the proper resources must be available at that time for it to
survive and establish. Thus, chance also strongly challenges this notion of a monoclimax.27
Modern definitions of succession still strongly reflect the thoughts of Clements and the
progression toward a stable ecosystem. However, ecologists largely treat succession as a way of
understanding how ecosystems reassemble after a disturbance and have reconsidered the
nature of the climax state, as can be seen in alternative stable state theory.
2.2. Review of Alternative Stable State Theory
Alternative stable state theory provides an alternative to the monoclimax. The roots of
this theory are traced back to Richard C. Lewontin’s 1969 paper, The Meaning of Stability, in
which Lewontin uses mathematical ecological modeling to justify the possibility of multiple
25 Ibid., 488. 26 Edward Goldsmith, “Ecological succession rehabilitated,” Edward Goldsmith,
http://www.edwardgoldsmith.org/page119.html (16 November 2010), 2. 27 R.H. Whittaker, 1953, A Consideration of Climax Theory: The Climax as a Population and Pattern, p. 240-‐
277, In: Frank B. Golley, ed., 1977, Ecological Succession, Stroudsburg, PA: Dowden, Hutchinson & Ross, 254.

Fiedler 15
stable states for a given system.28 C.S. Holling’s 1973 paper, Resilience and Stability of
Ecological Systems,29 and John P. Sutherland’s 1974 paper, Multiple Stable Points in Natural
Communities,30 fleshed out the theory of alternative stable states, which has now taken hold in
modern ecological thought.
If the state variables of a system can exist in multiple arrangements, each at an
equilibrium that is stable at the local scale, then that system has alternative stable states. These
state variables include abundances of species or feeding guilds, population demographics,
spatial distributions, or abiotic factors. This idea can be considered in two ways. First, as the
community perspective describes, a certain community may exist in a number of different
states, even under the same external environmental conditions (Figure 1, left). Different stable
states are capable of being achieved under the same conditions. In order for the system to
move from one stable state to another, a large disturbance to the state variables must occur.
These variables describe the internal characteristics of the community. For example, drastic
changes in population demographics or densities might cause this shift. On the other hand, the
ecosystem perspective attributes the existence of alternative stable states to changes in the
28 Richard C. Lewontin,1969, "The meaning of stability," Brookhaven Symposia in Biology, 22: 13-‐24, 13. 29 C. S. Holling, 1973, "Resilience and Stability of Ecological Systems," Annual Review of Ecology and
Systematics, 4: 1-‐23. 30 John P. Sutherland, 1974, "Multiple Stable Points in Natural Communities," American Naturalist, 108
(964): 859-‐873.

Fiedler 16
parameters that determine the behavior of or interactions between populations, such as birth
and death rates or predation (Figure 1, right). Environmental changes might shift these
parameters, causing the stable states to shift. The new stable state need not have been
possible under the previous parameters.31 These two perspectives differ from one another in
whether the shift originates in the community itself (variables) or in the ecosystem containing
that community (parameters).
Figure 1: Ball-‐in-‐cup diagram depicting the community perspective (left) and the ecosystem perspective (right) of alternative stable state theory.32
Shifts between stable states can occur at varying degrees and rates. A smooth response
occurs when a small change to variables or parameters results in a small change in the state of
the system. A threshold response occurs when most changes in conditions result in a small shift
31 Beisner et al. 2003, 376. 32 Ibid., 377.

Fiedler 17
in the system state, but when small changes in conditions, around a threshold value, result in a
drastic shift, or a critical transition.
For a system to return to a previous stable state it is often more difficult than simply
shifting the conditions back to where they were before. Hysteresis occurs when a system must
shift back to conditions far beyond those at the point of the critical transition in order to return
to a previous stable state (Figure 2).33 However, hysteresis is not a necessary condition for
alternative stable states. Minor changes can be absorbed by a system, yet large disturbances or
small changes around a critical point might result in a transition to a different stable state.
Figure 2. A model of a system exhibiting hysteresis. Small changes in conditions near point F1 or F2 result in a critical transition to the alternative stable state. Forward shift and backward shift show that the system must return to conditions far beyond points F1 and F2 in order to return to the original stable state.34
33 Marten Scheffer, Alternative Stable States and Regime Shifts in Ecosystems, p. 395-‐406, In: Simon A.
Levin ed., 2009, The Princeton Guide to Ecology, Princeton, NJ: Princeton University Press, 396. 34 Ibid., 397.

Fiedler 18
This theory requires the distinction between a system that can reach multiple stable
states and one that entirely lacks stability. Alternative stable states theory allows for a shift in
the location of the stable point of a given system, yet still maintains the notion that there do
exist several stable states that the system shifts between. Lewontin explains:
...it is a stable point, since a small perturbation of the system will result in the system returning to that point. It is necessary to specify that the perturbation is small because there may be several such stable points in the space and each will have its own basin of attraction. If the perturbation is sufficiently large to carry the system out of one basin of attraction into another, the original point is still a stable point even though the system did not return to it.35
Thus, a system is at a stable point if the state variables and parameters remain constant for a
given time and place, despite outside changes that threaten to affect them. Of course, the
spatial and temporal scale in which the system is considered must be appropriate in order to
understand its stability.36
Alternative stable state theory has received much attention in the field of ecology,
particularly due to increasing evidence of alternative stable states prompted by human
activities.37 It has modified successional theory to allow for systems to have multiple stable
states, while transforming our views of disturbed systems, and in turn conservation. However,
35 Lewontin 1969, 15. Original emphasis. 36 Sutherland 1974, 860. 37 Michael L. Cain, William D. Bowman, and Sally D. Hacker, 2008, Ecology, Sunderland, MA: Sinauer
Associates, Inc., 358.

Fiedler 19
the notion of “stable states” in itself can be contested in ecological thought. Panarchy has
recently begun to challenge this idea, and once again to transform our perspectives on stability
and conservation.
2.3. Review of Panarchy
The ideas behind panarchy as a whole was first presented in Lance Gunderson and C.S.
Holling’s 2001 synthesis, Panarchy: Understanding Transformation in Human and Natural
Systems.38 Panarchy provides a way to understand natural systems as a series of nested
adaptive cycles, continually in flux between growth, accumulation, restructuring, and renewal
throughout their life cycles.39 The paradigm of panarchy does not allow for any state of
stability, as did succession and alternative stable states. An adaptive cycle can describe any
dynamic system under consideration, which is influenced by other adaptive cycles of larger,
slower scale in conjunction with those of smaller, faster scale. These cycles are represented by
Gunderson and Holling as a figure-‐eight shape (Figure 3). Adaptive cycles progress through
scales of both potential and connectedness. Potential, or wealth, describes the possibilities for
change of a system. A higher potential means that the system has more future alternative
states. Connectedness, or controllability, describes the control a system has over its own
38 Lance H. Gunderson and C. S. Holling eds., 2002, Panarchy: understanding transformations in human
and natural systems, Washington, DC: Island Press, 21. 39 Holling 2001, 392.
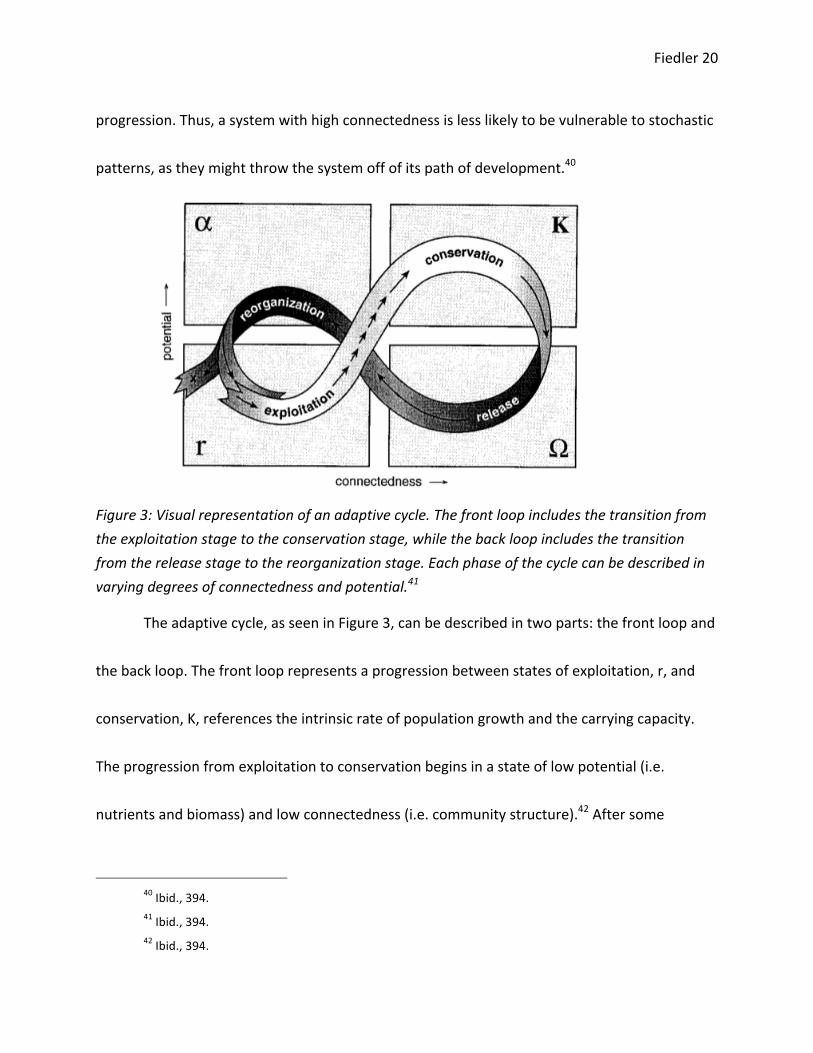
Fiedler 20
progression. Thus, a system with high connectedness is less likely to be vulnerable to stochastic
patterns, as they might throw the system off of its path of development.40
Figure 3: Visual representation of an adaptive cycle. The front loop includes the transition from the exploitation stage to the conservation stage, while the back loop includes the transition from the release stage to the reorganization stage. Each phase of the cycle can be described in varying degrees of connectedness and potential.41
The adaptive cycle, as seen in Figure 3, can be described in two parts: the front loop and
the back loop. The front loop represents a progression between states of exploitation, r, and
conservation, K, references the intrinsic rate of population growth and the carrying capacity.
The progression from exploitation to conservation begins in a state of low potential (i.e.
nutrients and biomass) and low connectedness (i.e. community structure).42 After some
40 Ibid., 394. 41 Ibid., 394. 42 Ibid., 394.

Fiedler 21
disturbance, the system begins with “biotic legacies,” or those species that survived, and other
abiotic factors that remain unchanged. Vegetation that can withstand this disturbed
environment will quickly gain dominance in the r phase, taking advantage of the low
connectedness. As new species immigrate into the system, “A period of contest competition
among entrepreneurial pioneers and surviving species from previous cycles ensues.”43 As new
and surviving species compete and populations become established, a more structured
community is developed, with increasing nutrients, biomass, and species interactions, and thus
increasing connectedness, quite similar to the progression described in succession.44 At the
height of this period of intense competition, we might find a peak in species diversity, as
thereafter species will begin to die out if they are unfit competitors, even if they were able to
take advantage of the disturbed environment. Again, reflecting successional thought, the
transition from the r-‐phase to the K-‐phase is a period of increasing control and less variability.
As the system approaches K, or the conservation phase of the cycle, the system is less likely to
experience novelty, or some alternative ecosystem structure.45 In ecological terms, the final
trajectory of the system can be either dominance-‐ or founder-‐controlled. The composition of
dominance-‐controlled communities, as described under successional theory, is dictated by
43 Gunderson and Holling eds. 2002, 43. 44 Ibid., 43-‐44. 45 Ibid., 44.

Fiedler 22
those species that are more competitively dominant than others.46 The composition of founder-‐
controlled communities, however, is dictated by a competitive lottery: no species is
competitively dominant, thus the composition is either determined by random population
fluctuations or the first colonizers of the area.47
However, the adaptive cycle of panarchy does not describe this increasing control and
connectedness as synonymous with stability, like succession and alternative stable states
theory suggest. Instead, panarchy views this increasing connectedness as rigidity. As the system
becomes more structured, it actually becomes more vulnerable to disturbances. Each species
has settled in a unique niche, thus any slight perturbation puts the entire system at risk.48 Once
a disturbance occurs, “a gale of creative destruction can be released in the resulting Ω
phase.”49 Potential drops as the disturbance destroys many of the resources that had
accumulated.
The disturbance moves the system into the back loop of the adaptive cycle. The back
loop is defined by the release (Ω) phase and the reorganization (α) phase (Figure 3). The length
of the back loop is defined by the nature of the disturbance, and whether it acts slowly or
46 Begon et al. 2006, 489. 47 Ibid., 493. 48 Holling 2001, 394. 49 Gunderson and Holling eds. 2002, 45.

Fiedler 23
suddenly. As the disturbance is acting, connectedness is destroyed in the system, thus allowing
for novelty in reorganization to follow. This process is unpredictable, rather than
deterministic.50 New relationships can be formed between variables, or immigrants can
dominate a system, where they otherwise would be outcompeted. The entire system can follow
a new trajectory due to the events during the reorganization phase.51 The adaptive cycle
continues into the exploitation phase and connectedness increases once again. The very nature
of this connectedness may be vastly different than it was before because of the events of the
back loop.
The phases of panarchical adaptive cycles can be better understood when we consider
them as nested cycles of different temporal and spatial scales (Figure 4). Forces of different
scales will influence any system considered. Larger and slower forces will cause the system to
“remember,” as these forces are less likely to fluctuate and maintain constant control. The
larger and slower cycle might be climatic control, source populations of immigrants, or the
remnants of potential in soil nutrients or stored seeds from the system prior to the disturbance.
The smaller and faster cycle that acts upon the system being considered might cause it to
50 Holling 395. 51 Gunderson and Holling eds. 2002, 46.

Fiedler 24
“revolt.” Its processes are quick and fluctuating and might spark novelty.52
Figure 4. Visual representation of nested adapted cycles.53
This force could be rooted in an invasive entering a disturbed system and establishing a strong
population, thus altering the trajectory of recovery, for example. The release of a system may
lead to revolt, but does not require this to be so. As C.S. Holling describes the nature of
panarchical cycles, “The conservative nature of established panarchies certainly slows change,
while at the same time accumulating potential that can be released periodically if the decks are
cleared of constraining influences by large, extreme events.”54 Whether a system revolts or
remembers is triggered not only by the influences of adaptive cycles of different scales, but also
52 Holling 2001, 398. 53 Ibid., 398. 54 Ibid., 399.

Fiedler 25
the scale of the disturbance and the variables that either remain or were eliminated. The
theory of panarchy cannot predict whether a system will revolt or remember after a
disturbance, yet it can describe conditions that influence each option.
Notions of resilience underlie the theory of panarchy, and balance its discussion of
rigidity and revolt. The concept of rigidity is contrary to that of resilience, or, “…the magnitude
of disturbance that can be absorbed before the system changes its structure by changing the
variables and processes that control behavior.”55 That is to say that not every disturbance will
project the system into the back loop, and certainly not every disturbance will occur in the
conservation phase. The resilience of a system is not a constant quality, but changes depending
on where the system is in the adaptive cycle. For example, as the system progresses through
the back loop, at levels of high potential and low connectedness, resilience is high. Thus, if the
system undergoes a disturbance at this point in the cycle, it will very easily return back to its
previous state. Resilience is low during the conservation phase of the front loop, where rigidity
is high. When a disturbance occurs during this phase, it is less likely that it will be able to return
to that state.56 This idea echoes that of hysteresis of alternative stable state theory.
55 Gunderson and Holling eds. 2002, 28. 56 Holling 2001, 395.

Fiedler 26
Panarchy adapts ideas from successional and alternative stable state theory, while
progressing ecological theory of recovery to account for non-‐linearity in adaptive cycles. It is
difficult to classify panarchy as a theory (however, I am comparing it to theories here), as it is
not testable or falsifiable, rather it is a paradigm that can illuminate ecological realities that we
observe. Panarchy has not taken hold of ecological discourse, perhaps due to its goals of acting
as a way of explaining human and natural systems in conjunction. It aims to apply to economic,
political, and ecological systems, and their intersection. However, despite the ambitious nature
of this paradigm, it is capable of thoroughly explaining living ecological systems. The application
of panarchy will illuminate its new perspective.
2.3. Comparing the theories
These three theories or paradigms are not independent from one another. Alternative
stable state theory was developed largely in response to succession, while panarchy is now
continuing the development of ecological theory or paradigms of response to disturbance. In
many ways, the later theories echo that which came before them. However, each theory also
uses a unique language and new perspectives in developing thought regarding ecosystem
recovery from disturbances. While much of the distinction between these theories can only be
illuminated when they are applied to living ecological systems, we can still begin to see how
Fiedler 27
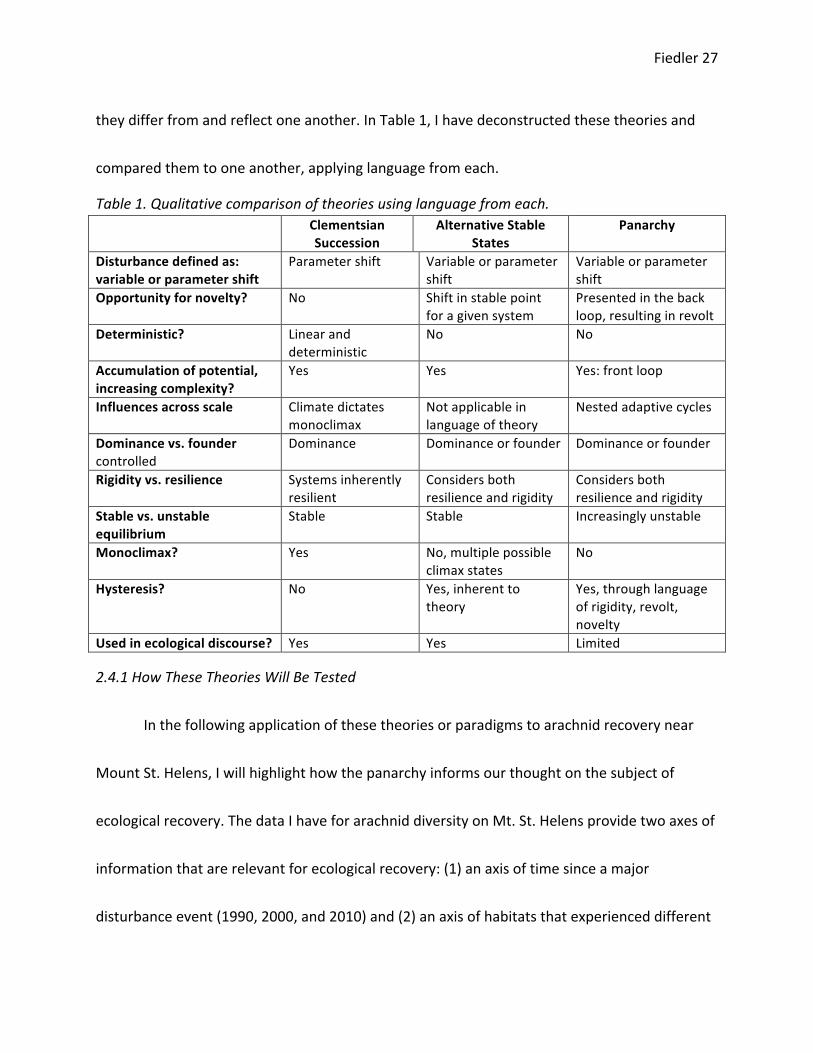
they differ from and reflect one another. In Table 1, I have deconstructed these theories and
compared them to one another, applying language from each.
Table 1. Qualitative comparison of theories using language from each. Clementsian
Succession Alternative Stable
States Panarchy
Disturbance defined as: variable or parameter shift
Parameter shift Variable or parameter shift
Variable or parameter shift
Opportunity for novelty? No Shift in stable point for a given system
Presented in the back loop, resulting in revolt
Deterministic? Linear and deterministic
No No
Accumulation of potential, increasing complexity?
Yes Yes Yes: front loop
Influences across scale Climate dictates monoclimax
Not applicable in language of theory
Nested adaptive cycles
Dominance vs. founder controlled
Dominance Dominance or founder Dominance or founder
Rigidity vs. resilience Systems inherently resilient
Considers both resilience and rigidity
Considers both resilience and rigidity
Stable vs. unstable equilibrium
Stable Stable Increasingly unstable
Monoclimax? Yes No, multiple possible climax states
No
Hysteresis? No Yes, inherent to theory
Yes, through language of rigidity, revolt, novelty
Used in ecological discourse? Yes Yes Limited
2.4.1 How These Theories Will Be Tested
In the following application of these theories or paradigms to arachnid recovery near
Mount St. Helens, I will highlight how the panarchy informs our thought on the subject of
ecological recovery. The data I have for arachnid diversity on Mt. St. Helens provide two axes of
information that are relevant for ecological recovery: (1) an axis of time since a major
disturbance event (1990, 2000, and 2010) and (2) an axis of habitats that experienced different

Fiedler 28
levels of disturbance in that event (pumice plain, blowdown forest, tephra forest, and reference
forest). To consider the application of theories of ecological recovery for this system, I will
apply my data in several ways. First, analyses of the total number of taxa in each research site
across time will provide an example of how one aspect (species diversity) of ecosystem
complexity has changed throughout recovery. Second, an analysis of the ballooning and
cursorial spider species present in each site over time will show how the species pool of
potential inhabitants and, subsequently, successful colonizers are dependent on what species
are capable of reaching the site. I expect to see more ballooning species than cursorial species
in disturbed sites, yet over time the number of each should increase, as would be consistent
with each of the three theories.
Through an analysis of feeding guilds and habitat preferences, I will consider ecosystem
complexity and how it differs between research sites and over time. Each theory predicts an
increase in ecosystem complexity as a part of recovery. However, the character of that
complexity is allowed to diverge from the preexisting state, according to alternative stable state
theory and panarchy only. Finally, I will isolate patterns of individual families or species that are
relevant to the course of recovery. Patterns that exhibit higher abundance of a particular taxon
in less or more disturbed sites might suggest different trajectories of recovery, while similar
abundance of a taxon across all sites would suggest the potential for the same trajectory of

Fiedler 29
recovery. Successful invaders in disturbed sites might indicate the occurrence of novelty and
creativity within the course of recovery, as panarchy and alternative stable states theory
describe. While any fluctuations in the reference forest samples will indicate that the site has
not reached its climax or stable state in successional and alternative stable state theories,
respectively, it could also simply indicate that the system is in the conservation phase of
panarchy and increasing in rigidity. As each theory builds off of that which came before it, many
of the predictions each theory makes might reflect those found in another theory, yet with
different language. Ultimately, however, panarchy emphasizes the dynamism of systems, while
succession and alternative stable states do not.
3. Listening to the Landscape: A Case Study
The 1980 eruption of Mount St. Helens captured the interest of natural scientists in
virtually every discipline. Scientists were able to examine ecological response and recovery
immediately after the eruption, rather than decades or centuries after, as is often the case. This
research has contributed not only to a better understanding of ecosystem recovery at Mount
St. Helens, but recovery following any volcanic disturbance, or even after disturbances in

Fiedler 30
general. Today, more than half of all studies published globally on the ecosystem responses to
volcanic eruptions were done on Mount St. Helens.57
One such study, Parmenter et al. (2005), done at Mount St. Helens examined the
succession of ground-‐dwelling beetles following the eruption. This study used the same
research sites and sampling dates that were used in the collection of my samples, as will be
explained in the following sections. The study also used pitfall trap sampling, also used for my
study, which is a common method of collecting and preserving ground dwelling arthropods.
This sampling method is biased towards those taxa who are most active on the ground.58
Parmenter et al. (2005) found a total of 27,074 beetles of 279 species and 39 families in four
research sites and four sampling years (1987, 1990, 1995, 2000).59 The authors found that
beetle species sequentially replaced other species throughout time in the disturbed sites as
conditions, such as vegetation and prey availability, changed. As would be expected, the most
disturbed site, the pumice plain, least resembled the undisturbed reference forest. Even in the
year 2000, 20 years after the eruption, the beetle species composition of each disturbed
57 Virginia H. Dale, Frederick J. Swanson, and Charles M. Crisafulli, 2005, Disturbance, Survival, and
Succession: Understanding Ecological Responses to the 1980 Eruption of Mount St. Helens, p. 3-‐11, In: Virginia H.
Dale, Frederick J. Swanson, and Charles M. Crisafulli eds., 2005, Ecological Responses to the 1980 Eruption of
Mount St. Helens, New York, NY: Springer, 3-‐4. 58 Parmenter et al. 2005. 59 Ibid., 142.

Fiedler 31
research site still did not resemble that of the reference forest. Thus, the authors hypothesized
that we will continue to see changes in community structure in the future.60 This study can
serve as an example of how we can understand data obtained from this sampling methodology,
and a comparison of another arthropod taxon’s recovery.
3.1. Arachnid Recovery of Mount St. Helens
Arachnids have been able to colonize virtually every terrestrial habitat. Of course, not
every species of arachnid can itself successfully live in different habitats. Each species is limited
by physical factors of the microhabitat, including humidity and temperature. Biological factors,
such as prey type and vegetation, also limit the distribution of species. Thus, many species have
distinctly defined ecological niches, which can be described by both habitat preference and
feeding guild.61 Where they live, arachnids serve a variety of ecological roles. Spiders (Order
Araneae) are entirely predatory, while Opiliones (harvestmen) and Acari (mites and ticks) can
be predators, herbivores, or detritivores. Even among predatory spiders, taxa have evolved
specific feeding biologies that take place in different parts of the ecosystem. Some may wander
or build foraging webs off of the ground, while others are only found on the ground or in leaf
litter.62
60 Ibid., 148-‐149. 61 Jan Beccaloni, 2009, Arachnids, Berkeley: University of California Press, 17. 62 Ibid.

Fiedler 32
From the identification of an arachnid, we can deduce information about the ecosystem
by using our knowledge of the individual’s habitat requirements. In my analysis, I will use
habitat preference and feeding guild to describe the ecological niche of arachnids. By focusing
on arachnid recovery, we can explore the potential to understand ecosystem recovery as a
whole. Furthermore, spiders have a history of being some of the first colonizers in the most
devastated areas following volcanic eruptions. Within just a year of the 1883 eruption of
Krakatoa, which left the island completely devoid of life, a linyphiid became the first to
successfully colonize. Within 50 years, there were over 90 species of spiders on the island.63
3.1.1. Arachnid Sampling Methodology
Samples were collected by a National Forest Service crew, working under Charlie
Crisafulli, from four study sites at Mount St. Helens: reference forest, tephra forest, blowdown
forest, and pumice plain (Figure 5). All four sites are located at similar elevations, from 1040 m
to 1175 m. Each research site was a standing old-‐growth forest, which had never been logged,
at the time of the 1980 eruption, yet each was disturbed differently and to varying degrees.
Each of these four sites was affected differently by the eruption, and thus may present different
courses of recovery. The reference forest is located 40km northeast of Mount St. Helens on
Lonetree Mountain. Parmenter et al. (2005) claims that the reference forest was unaffected by
63 Rainer F. Foelix, 1996, Biology of spiders, New York: Oxford University Press, 235.

Fiedler 33
the 1980 eruption and treats the site as a reference for the populations and communities that
were present before the eruption in the three disturbance sites.64 However, due to the scale of
the eruption it is possible that this site was still subject to at least ash fall, which in turn could
affect small arthropods.
Figure 5. Locations of research sites. Sites labeled as “beetle sites” are those from which arachnids were collected.65
64 Parmenter et al. 2005, 140. 65 Ibid., 141.

Fiedler 34
The reference forest consists of trees with an understory that is predominantly leaf litter
and bryophytes cover (Table 2). The tephra forest site is located directly east of the blowdown
forest, in the Hemlock Forest area. In this site, the eruption buried leaf litter and much of the
understory vegetation with tephra fall. Thirty years later, new leaf litter has covered much of
the tephra, and seedlings and shrubs have reemerged.66 The blowdown forest site is located
near Norway Pass, northeast of the pumice plain where the old-‐growth forest was leveled by
the strength of the blast, leaving no standing trees and minimal surviving vegetation. The
majority of ground cover for this site is downed wood and herbaceous vegetation. The pumice
plain sample site is located within the pyroclastic flow, south of Spirit Lake. This site is
dominated by bare ground and rock as it was entirely scoured and covered by pumice from the
eruption. Only a few pockets of vegetation survived these pyroclastic flows. Small amounts of
herbaceous vegetation and bryophyte vegetation have recovered at this site.
These four research sites represent a disturbance gradient in which different aspects of
the communities were disturbed and at varying degrees. This gradient is arranged spatially,
where the most disturbed sites are closest to the crater of the volcano, and the least disturbed
sites are located farthest away. The undisturbed site (the reference forest) and other
corresponding undisturbed localities which represent intact source populations are also
66 Ibid., 140-‐141.

Fiedler 35
farthest away from the crater and, in turn, the pumice plain. As the level of disturbance
decreases, the distance from source populations also decreases, potentially impacting the rate
of immigration and recovery. It is important to note this distance gradient from source
populations when considering the data, as each of these sites does not act independently. Each
disturbance area acts as a corridor for the movement of species. Overtime, the intermediately
disturbed sites will become source populations for the most disturbed site, which is furthest
from the reference forest. Thus, each site is interconnected temporally and spatially in terms of
the movement of taxa.
Table 2. Mean cover values (%) for four research sites, as measured in 1995. 67 Disturbance Zone Habitat Variable
Bryophyte Herb Shrub Tree Litter Wood Rock/ground Reference Forest 23.3 1.5 12.8 34.1 76.3 14.1 0.0 Tephra Forest 1.5 0.0 12.6 15.7 91.1 11.7 4.5 Blowdown Forest 11.5 26.9 1.9 0.0 12.7 28.6 12.4 Pumice Plain 5.9 7.9 0.0 0.0 3.1 0.0 83.2
Arachnids and other ground-‐dwelling arthropods were collected in pitfall traps at the
four research sites each year since the eruption. The pitfall traps were comprised of six inch
deep plastic cups placed into the ground, with the tops of the cups flush with the ground
surface. The cups were filled with propylene glycol to act as a preservative for the samples
67 Charles M. Crisafulli, James A. MacMahon, and Robert R. Parmenter, 2005, Small Mammal Survival and
Colonization on the Mount St. Helens Volcano: 1980-‐2002, p. 199-‐218, In: Virginia H. Dale, Frederick J. Swanson,
and Charles M. Crisafulli eds., 2005, Ecological Responses to the 1980 Eruption of Mount St. Helens, New York, NY:
Springer, 203.

Fiedler 36
collected. Ten pitfall traps were placed 10m apart along a transect within each study site. Traps
were set from late May through October and were emptied every three weeks. Once removed,
samples were stored in 70% ethanol.68
For my investigation, I used samples collected in 1990, 2000, and 2010, to provide a
summary of the changes in arachnid populations since the eruption. Though the eruption
occurred in 1980, I chose the 1990 trap data because it was the oldest complete and
methodologically consistent data set and could therefore be compared to those from 2000 and
2010. I used samples from three sampling dates per year, one each in July, August, and
September. I summed the data within each year, thus including the three sampling dates with
ten pitfall traps each, in order to account for any seasonal variation in the activity of certain
taxa and the variation within each site.
I separated the arachnids from the other ground-‐dwelling arthropods in the pitfall trap
samples. I identified the spiders to the finest taxonomic level possible, using Vincent D. Roth et
al.’s Spiders of North America: an Identification Manual69, a dichotomous key, and personal
communications with Rod Crawford.70 I identified the opilionids to the genus or species level
68 Parmenter et al. 2005, 140-‐141. 69 Vincent D. Roth, Darrell Ubick, and N. Dupérré, 2005, Spiders of North America: an identification
manual, Poughkeepsie, NY: American Arachnological Society. 70 Rod Crawford, personal communication, March 2, 2011.

Fiedler 37
under the guidance of Shahan Derkarabetian.71 Mites were separated and counted, but were
not identified for this study. Identifications were performed in a random order, thus avoiding
any sampling bias due to improving identification skills.
3.1.1.1. Limitations of Sampling Methodology
While pitfall trap sampling can provide a general understanding of the species
composition of the research sites, it does present some biases that must be taken into account
when analyzing the data. This trapping methodology will preferentially trap wandering ground
species. Sit-‐and-‐wait or arboreal species will be highly underrepresented or not trapped at all.
Thus, the data from this sampling technique does not represent absolute density, but rather
the relative activity of the species present.72 Furthermore, pitfall trap sampling might better
represent certain research sites, as opposed to others, due to habitat complexity. I predict that
the data collected using this sampling method will best represent the species diversity of the
pumice plain research site, as the limited vegetation will result in mostly ground-‐dwelling taxa.
The species diversity reference forest will most likely be the most underrepresented, as its
habitat complexity implies that a large number of taxa live in vegetation or trees far above the
ground.
71 Shahan Derkarabetian, personal communication, February 11, 2011. 72 Crawford et al. 1995, 67.

Fiedler 38
I am only considering pitfall trap samples from three sampling years. While this will
provide an overview of the changes in community composition over time, it may present
problems in understanding the underlying processes that dictate the changes that I observe.
However, similar analyses have been done on the recovery of beetles following the Mount St.
Helens eruption with only four sampling years, summing together the beetles found in all the
pitfall traps of a single sampling site and year in their analyses.73 My latest sampling date is a
mere 30 years after the eruption, thus my samples do not represent ecosystem recovery to its
completion. Moreover, I am not able to assess the degree of change in the first ten years after
the disturbance. Crawford et al. (1995), however, does provide some initial observations in the
years immediately following the eruption that can only be loosely compared with my data set
due to differing sampling methodology.74 Thus, the trajectory of ecosystem recovery can be
hypothesized and estimated from the data I have obtained. Finally, due to time and resource
constraints, not all samples were identified to species or even family level. Again, this must be
taken into account, as taxon numbers and certain ecological classifications might be
misrepresented. This limitation, however, applies uniformly to all research sites and sampling
years.
73 Parmenter et al. 2005. 74 Crawford et al. 1995.

Fiedler 39
As discussed in Section 2.4.1., I will present a summary of my data in a way that will
facilitate a better understanding of the three ecological theories. Table 2 summarizes what each
category of data provides for my assessment of these theories. While each data categorization
will describe certain aspects of recovery, they are interpreted differently by each theory.
Table 2. Summary of how each data category will be applied to theory analysis. Data What Data Provide Analysis
Total Number of Taxa Species diversity reflecting ecosystem complexity Ballooners vs. Cursorial Dispersers Species pool, limitations of individuals to disperse and
reach disturbed research sites Feeding Guilds Ecosystem complexity, compare to reference forest Habitat Preferences Ecosystem complexity, compare to reference forest Higher Abundance in Less Disturbed Sites
Potential for different trajectories of recovery, revolt
Higher Abundance in More Disturbed Sites
Potential for different trajectories of recovery
Similar Distribution Across All Research Sites
Potential for similar trajectories of recovery, remember
Successful Invaders Potential for novelty, revolt 3.1.2. Arachnid Dispersal Methods
Understanding the dispersal methods of arachnids is essential in applying these data to
each ecological theory. Dispersal ability is a likely influence on which species are able to
successfully colonize a disturbed area and the development of the community thereafter. From
those species that are able to reach an area, only those that can meet their specific habitat
needs upon arrival have the potential to establish populations. Arachnids can disperse by
ballooning, walking, rafting on physical objects, or phoresy (using other organisms for

Fiedler 40
transport).75 Ballooning and walking are the dispersal methods most relevant to the
recolonization of the disturbed environments of Mount St. Helens. Juvenile or small spiders, as
well as some mites, can employ ballooning as an active mode of dispersal. Large spiders, due to
physical constraints, and those arachnids without the ability to produce silk threads, are unable
to balloon. In order to balloon, spiders stand in an exposed area on their tiptoes, extend a
strand of thread, and face the wind. This strand is caught by the wind, carrying the spider up
into the air. These spiders can travel vast distances, and have even been spotted at altitudes of
several thousand meters in the air.76 While aloft, they can, to some extent, control where they
land by pulling on their threads or by re-‐ballooning until they have reached an appropriate
destination. However, the direction of travel is dominated by the wind currents and can result
in spiders dispersing to less than preferable environments.77
The act of ballooning is a high-‐risk method of dispersal due to the low probability that
the spider will land in a habitat more, or even just equally, suitable to the one from which it
travelled. In habitats that are patchily distributed across a landscape, much like the landscape
that resulted from the Mount St. Helens eruption, ballooning is even more of a risk. If another
75 Beccaloni 2009, 17. 76 Foelix 1996. 77 Rodney L. Crawford and John S. Edwards, 1986, "Ballooning Spiders as a Component of Arthropod
Fallout on Snowfields of Mount Rainier, Washington, U.S.A.," Arctic and Alpine Research, 18 (4): 429-‐437, 429.

Fiedler 41
suitable patch is potentially far away, there is a small likelihood of a spider reaching the site
with low wind speeds.78
Ballooning and other modes of dispersal are likely promoted by natural selection.
Habitat variability, competition with relatives, and the avoidance of inbreeding will all select for
dispersal methods, while a temporally stable environment, diverse habitat, and niche
specialization will all select against dispersal.79 Spiders that are highly specialized are far less
likely to balloon than their generalist counterparts. Environmental pressures will promote
dispersal into new habitats, as long as the potential benefits of doing so outweigh the risks of
landing in an unsuitable habitat.80
It is important to note that quantifying dispersal is difficult, as resident individuals
cannot be differentiated from incoming dispersers. However, in heavily disturbed environments
where no resident populations are present, newly arriving individuals can be readily
identified.81 It is also important to differentiate between newly arriving dispersers and
78 Dries Bonte, Nele Vandenbroecke, Luc Lens, and Jean-‐Pierre Maelfait, 2003, "Low propensity for aerial
dispersal in specialist spiders from fragmented landscapes," The Royal Society, 270 (1524): 1601-‐7, 1601. 79 Dries Bonte, Jeroen Vanden Borre, Luc Lens, and Jean-‐Pierre Maelfait, 2006, "Geographical variation in
wolf spider dispersal behaviour is related to landscape structure," Animal Behaviour, 72 (3): 655-‐662, 655. 80 Bonte 2003, 1601. 81 Crawford et al. 1995.

Fiedler 42
successful colonizers. Many incoming individuals are not able to successfully establish
populations.
3.1.3. Spider Feeding Guilds
Spiders are some of the most common predators in terrestrial ecosystems, filling this
role in two hugely different ways. Feeding guilds can describe foraging behavior, prey type, and
how the habitat is used. Spider predators can either be web builders or wanderers. Web
builders utilize one of four types of webs to capture prey: orbs, tangles, sheets, and funnel
webs.82 Meanwhile, wandering spiders do not use webs to capture prey. Instead, some spiders
actively hunt prey, while others wait in one place to surprise their prey. For example, jumping
spiders (Salticidae) actively hunt and stalk their prey, while crab spiders (Thomisidae) wait on
flowers, leavers, or bark to attack.83 Both wandering and sit-‐and-‐wait spiders can be found on
the ground or vegetation.84
Insects and other arthropods are the most common prey types for spiders. In general,
very few spiders consume vertebrates, yet some are capable of eating fish (such as the spider
Dolomedes) or geckos (such as the spider Leucorchestris). Insects that are found in significantly
82 David H. Wise, 1993, Spiders in ecological webs, Cambridge: Cambridge University Press, 17. 83 Ibid., 17. 84 Crawford 2011.

Fiedler 43
large numbers, like flies and Collembola, are naturally quite important in the diet of spiders.85
However, the feeding guild will often determine the type of prey a spider will consume. Web-‐
builders will only consume those prey that become ensnared, thus insects who are capable of
avoiding webs or who do not inhabit the areas where webs are built will be avoided.
Meanwhile, spiders that wait on vegetation before attacking their prey will also only encounter
certain prey types. However, despite the selective nature of these feeding types, most spiders
are generalists. As an extreme example, one spider, Linyphia triangularis, consumed 150 out of
153 prey types in an experimental setting.86
3.1.4. Spider Habitat Preferences
Spiders tend to be found in highly specific habitats, dependent on abiotic factors such as
temperature, humidity, wind, and light, and biotic factors, such as vegetation, prey availability,
competitors, and predators. Habitats can be delineated vertically in an ecosystem, as physical
conditions, or microclimate, often vary accordingly. Thus, some spiders are most commonly
found on the ground, while others are found on low vegetation, shrubs, tree trunks, or
treetops.87 The diverse range of habitats in which spiders exist may be determined by their
85 Foelix 1996, 240-‐241. 86 Ibid., 241. 87 Ibid., 236.

Fiedler 44
feeding guild and prey type; however, it may also be a result of the avoidance of interspecific
competition or other environmental requirements.88
3.1.5. Results
Samples from 1990, 2000, and 2010, with three sample collections per year, contained
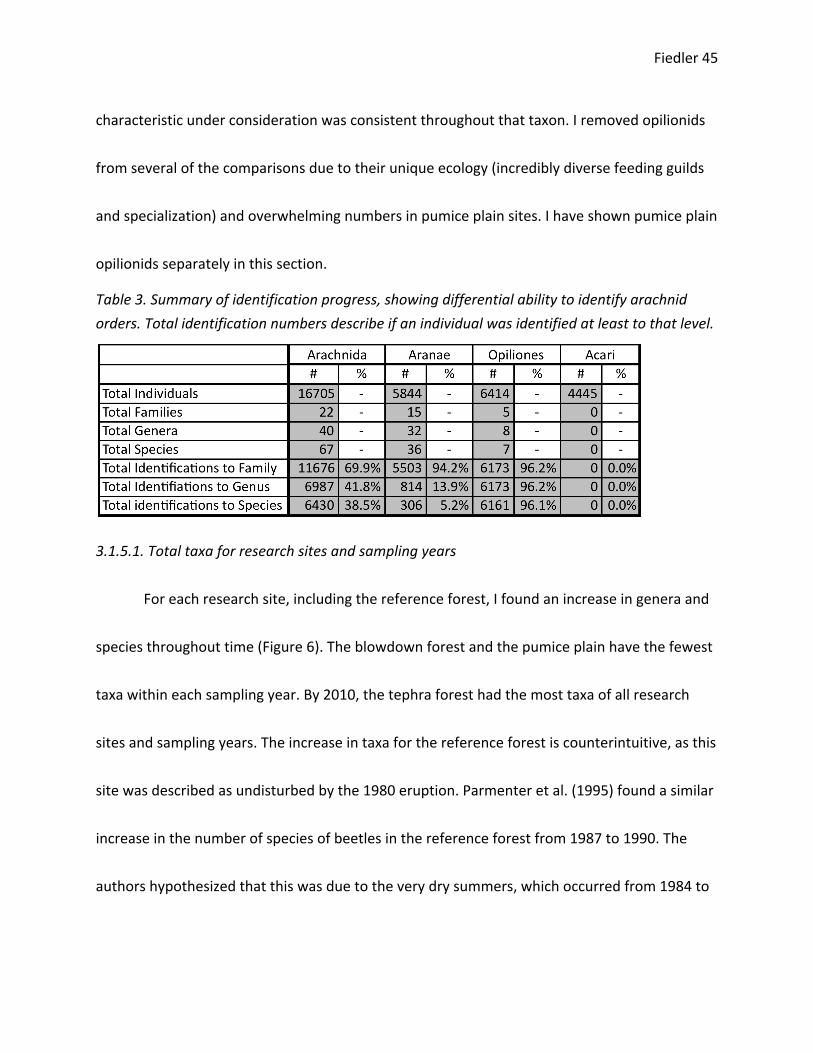
16,705 arachnids. Of these individuals, 70% were identified at least to the family level, 42% at
least to genus, and 39% to species. These identifications included 22 different families, 40
genera, and 67 species (summarized in Table 3). I was differentially able to identify arachnid
orders to generic and species levels with greater success for Opiliones, less so for spiders
(Araneae), and minimally for Acari. Considering spiders alone, I was able to identify 94, 14, and
5% to family, genus, and species respectively. A complete list of identifications divided by
research site and year is included in Appendix 1.
The following is a summary of results and patterns observed. Information regarding
arachnid dispersal methods, feeding guilds, and habitat preferences was provided by Rod
Crawford through personal communication.89 Results assessed by taxa were done using the
finest resolution of identification possible. Thus, the total number of taxa includes families,
genera, and species. However, families and genera were only considered when the
88 Ibid., 238. 89 Crawford 2011.
Fiedler 45
characteristic under consideration was consistent throughout that taxon. I removed opilionids
from several of the comparisons due to their unique ecology (incredibly diverse feeding guilds
and specialization) and overwhelming numbers in pumice plain sites. I have shown pumice plain
opilionids separately in this section.
Table 3. Summary of identification progress, showing differential ability to identify arachnid orders. Total identification numbers describe if an individual was identified at least to that level.
3.1.5.1. Total taxa for research sites and sampling years
For each research site, including the reference forest, I found an increase in genera and
species throughout time (Figure 6). The blowdown forest and the pumice plain have the fewest
taxa within each sampling year. By 2010, the tephra forest had the most taxa of all research
sites and sampling years. The increase in taxa for the reference forest is counterintuitive, as this
site was described as undisturbed by the 1980 eruption. Parmenter et al. (1995) found a similar
increase in the number of species of beetles in the reference forest from 1987 to 1990. The
authors hypothesized that this was due to the very dry summers, which occurred from 1984 to
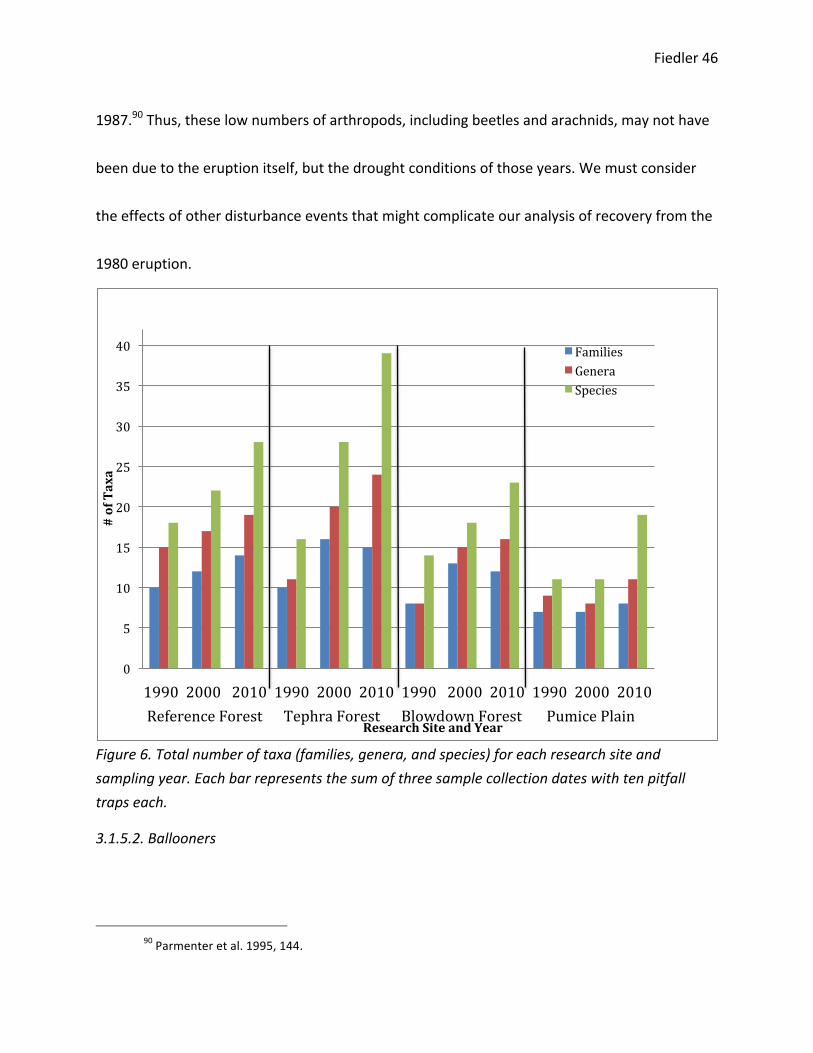
Fiedler 46
1987.90 Thus, these low numbers of arthropods, including beetles and arachnids, may not have
been due to the eruption itself, but the drought conditions of those years. We must consider
the effects of other disturbance events that might complicate our analysis of recovery from the
1980 eruption.
Figure 6. Total number of taxa (families, genera, and species) for each research site and sampling year. Each bar represents the sum of three sample collection dates with ten pitfall traps each.
3.1.5.2. Ballooners
90 Parmenter et al. 1995, 144.
0
5
10
15
20
25
30
35
40
# of Taxa
Research Site and Year
Families Genera Species
1990 2000 2010 1990 2000 2010 1990 2000 2010 1990 2000 2010 Reference Forest Tephra Forest Blowdown Forest Pumice Plain PR

Fiedler 47
Each research site had a distinct temporal pattern in the number of ballooning and
cursorial dispersing taxa (Figure 7). The reference forest shows relatively similar numbers of
ballooning and cursorial taxa, with numbers of both increasing overall. The tephra forest shows
a consistent increase in both ballooning and cursorial taxa throughout time. This site has many
more ballooning taxa than cursorial taxa, and it has the highest numbers of ballooning taxa for
each year of any site. The blowdown forest shows no pattern throughout time. In 1990 and
2010, there are many more ballooning taxa than cursorial taxa; however 2000 does not follow
this pattern. The number of ballooning taxa in the pumice plain increased consistently
throughout time, while I saw only one cursorial taxon each year.
Figure 7. Total number of ballooning and cursorial taxa for each research site and sampling year. Each bar represents the sum of three sample collection dates with ten pitfall traps each.
0
5
10
15
20
25
# of Taxa
Research Site and Year
Ballooner
Cursorial
1990 2000 2010 Reference Forest
1990 2000 2010 Tephra Forest
1990 2000 2010 Blowdown Forest
1990 2000 2010 Pumice Plain
Intact Source Pop. Closest to Source -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐> Farthest from Source

Fiedler 48
3.1.5.3. Feeding Guilds
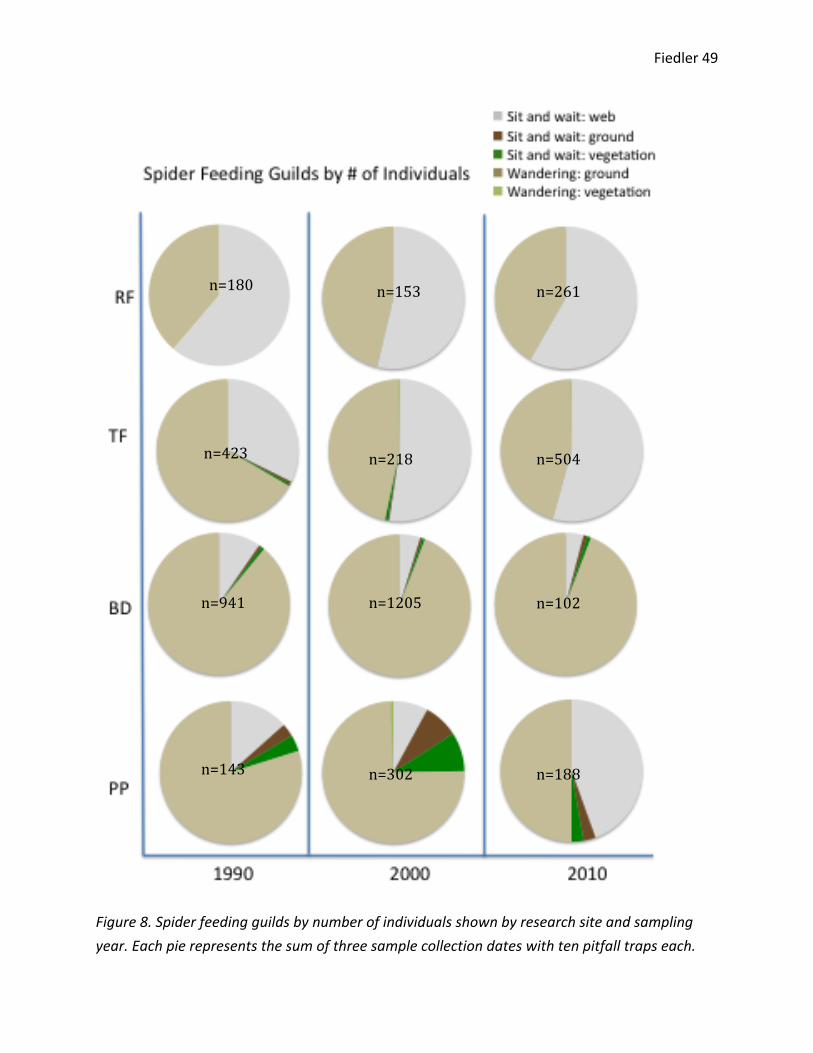
I compared the number of spider individuals in each feeding guild (Figure 8). Only
spiders were considered as they are all predators, while other arachnids fill very different
ecological roles and thus complicate comparisons. I classified the guilds as: sit-‐and-‐wait on
vegetation, ground, or web, and wandering on ground or on vegetation.91 These classifications
describe the dominant feeding guild for each taxon.
The reference forest showed consistent patterns throughout time. For each sampling
year, slightly fewer than ½ of trapped spider individuals were sit-‐and-‐wait web feeders. The
reference forest also showed the fewest feeding guilds, perhaps reflecting a sampling bias that
can be attributed to the complexity of the habitat. In the tephra forest’s 1990 samples, slightly
fewer than ¾ of the individuals were wandering ground feeders, and around ¼ were sit-‐and-‐
wait web feeders. There were also small numbers of sit-‐and-‐wait vegetation and sit-‐and-‐wait
ground feeders. In the 2000 and 2010 samples, the tephra forest feeding guild trends reflected
those of the reference forest. The blowdown forest shows a consistent trend in feeding guilds
throughout time. In all sampling years, around 4/5 of spider individuals were wandering ground
feeders; the remaining 1/5 of individuals were either sit-‐and-‐wait vegetation or ground feeders.
91 Ibid.
Fiedler 49
Figure 8. Spider feeding guilds by number of individuals shown by research site and sampling year. Each pie represents the sum of three sample collection dates with ten pitfall traps each.
n=180
n=153 n=261
n=423
n=218
n=504
n=941
n=1205
n=1022
n=143
n=302 n=188

Fiedler 50
In the pumice plain, there were major shifts in feeding guilds throughout time. In the
1990 samples, around ¾ of spider individuals were wandering ground feeders, 1/6 were sit-‐and-‐
wait wandering feeders, and the remaining individuals were wither sit-‐and-‐wait vegetation or
sit-‐and-‐wait ground feeders. In the 2000 samples, around ¾ of the individuals were wandering
ground feeders, and the remaining 25% of individuals was evenly divided between sit-‐and-‐wait
web, sit-‐and-‐wait ground, and sit-‐and-‐wait vegetation feeders. There were also a small number
of wandering vegetation feeders. In the 2010 sample, around ½ of spider individuals were
wandering ground feeders, less than ½ were sit-‐and-‐wait web feeders, and the remaining
individuals were either sit-‐and-‐wait ground or sit-‐and-‐wait vegetation feeders.
3.1.5.4. Habitat Preferences
I considered patterns of change in primary habitat preferences of the spiders collected
(Figure 9). Habitat preferences were categorized in the following categories: tree
canopy/trunk/understory foliage, deciduous and coniferous litter, moss, dead wood, and
exposed environments.92 Spiders were categorized into the habitat that they would are most
commonly found in, regardless of the habitat of the site in which they were collected. The
reference forest, tephra forest, and blowdown forest were all relatively consistent over time in
92 Ibid.

Fiedler 51
the distribution of habitat preferences of spider individuals, while the pumice plain was more
variable.
In 1990, slightly less than ½ of individuals collected in the reference forest are known to
inhabit dead wood habitats, while ¼ live in exposed environments, ¼ in deciduous and
coniferous litter, and very few in moss habitats. In the 2000 samples, around ½ of the collected
individuals are known to live in exposed environments, ½ in dead wood, and the rest either in
deciduous and coniferous litter, moss, or tree habitats. In 2010, around ½ of the individuals
collected are known to live in exposed environments, ½ in dead wood habitats, and around 1/6
in deciduous and coniferous litter. The tephra forest was even more consistent throughout time
than the reference forest. In 1990, around ½ of the individuals collected are known to live in
exposed environments, slightly less than ½ in dead wood, and the remaining individuals in leaf
litter. In the 2000 and 2010 samples, the distribution of habitat preferences remained similar to
that in 1990, with also a few individuals who are known to live in tree and moss habitats.

Fiedler 52
Figure 9. Spider habitat preferences by number of individuals shown by research site and sampling years. Each pie represents the sum of three sample collection dates with ten pitfall traps each.
n=180
n=153 n=261
n=423
n=218
n=504
n=941
n=1205
n=1022
n=143
n=302 n=188

Fiedler 53
The blowdown forest is again even more consistent throughout time. In all three
sampling years, around ½ of the individuals collected are known to live in exposed
environments and ½ in dead wood. In the 1990 samples, there were also a few individuals who
inhabit leaf litter. The pumice plain was highly variable throughout time. In the 1990 samples, ½
of the spider individuals collected are known to live in exposed environments, slightly less than
½ in dead wood, and some individuals in leaf litter and tree habitats. In 2000, ½ of the spider
individuals collected are known to live in exposed environments, slightly less than ½ in dead
wood habitats, and some individuals in leaf litter and moss. In 2010, 1/3 are known to live in
exposed environments, 1/3 in dead wood habitats, and 1/3 in leaf litter.
3.1.5.5. Representative Patterns
By considering patterns of individual taxa between research sites and across time, I
observed several patterns that represented larger trends in the community development of
these communities. Some taxa are found primarily in the least disturbed sites, others are found
mostly in disturbed sites, and some are found in high numbers in all four research sites. Some
invasive taxa have established strong numbers in the disturbed sites, yet they still remain few in
number in the less disturbed sites. Each of the following examples represents one of these
patterns that are observed across several taxa in my data set.

Fiedler 54
3.1.5.5.1. Higher Abundance in Less Disturbed Sites
Cybaeids are non-‐ballooning spiders that prefer dead wood or exposed environments.93
They are primarily sit-‐and-‐wait web or wandering ground feeders. I found individuals in the
family Cybaeidae primarily in the least disturbed sites: the reference forest and tephra forest
(Figure 10). Many other taxa in my samples follow this distributional trend, such as mites or
individuals of the genus Antrodiaetus.
Figure 10: Number of individuals of the family Cybaeidae in research sites over time. Each bar represents the sum of three sample collection dates with ten pitfall traps each.
93 Ibid.
106 122
187
131 107
304
9
54
21 1 0 0
0
50
100
150
200
250
300
# of Individuals
Research Site and Year
1990 2000 2010 Reference Forest
1990 2000 2010 Tephra Forest
1990 2000 2010 Blowdown Forest
1990 2000 2010 Pumice Plain
Less disturbed -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐> More disturbed
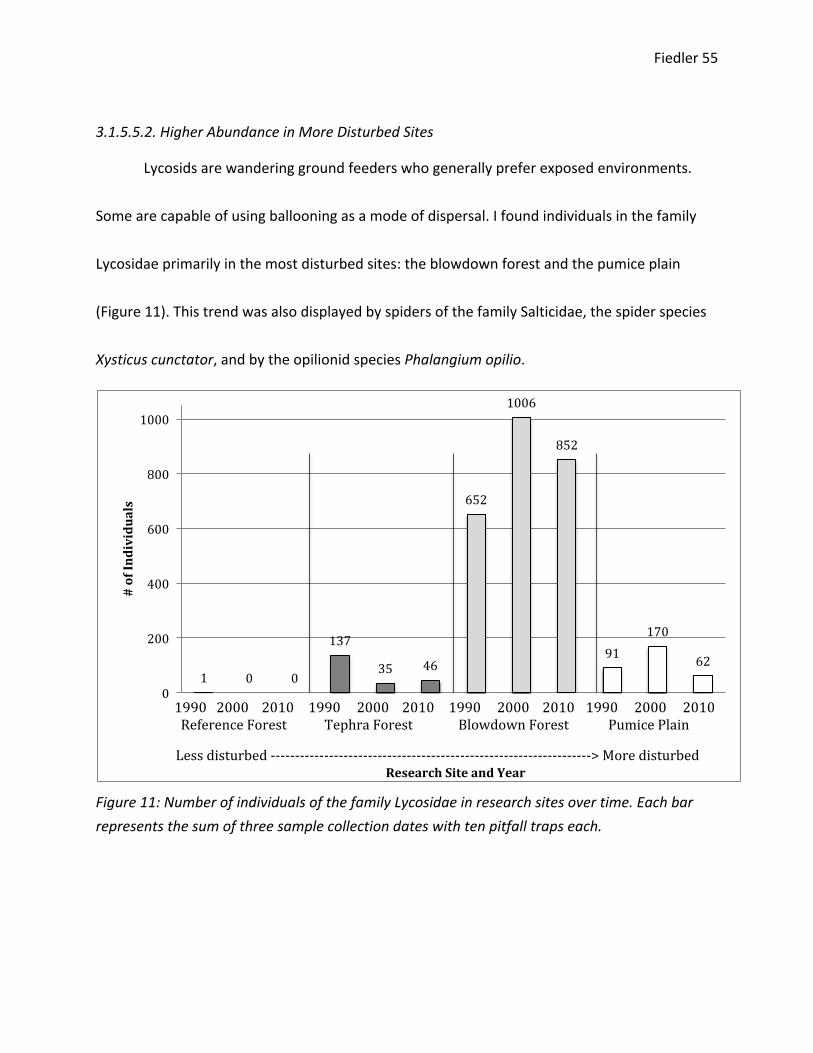
Fiedler 55
3.1.5.5.2. Higher Abundance in More Disturbed Sites
Lycosids are wandering ground feeders who generally prefer exposed environments.
Some are capable of using ballooning as a mode of dispersal. I found individuals in the family
Lycosidae primarily in the most disturbed sites: the blowdown forest and the pumice plain
(Figure 11). This trend was also displayed by spiders of the family Salticidae, the spider species
Xysticus cunctator, and by the opilionid species Phalangium opilio.
Figure 11: Number of individuals of the family Lycosidae in research sites over time. Each bar represents the sum of three sample collection dates with ten pitfall traps each.
1 0 0
137
35 46
652
1006
852
91 170
62
0
200
400
600
800
1000
# of Individuals
Research Site and Year
1990 2000 2010 Reference Forest
1990 2000 2010 Tephra Forest
1990 2000 2010 Blowdown Forest
1990 2000 2010 Pumice Plain
Less disturbed -‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐-‐> More disturbed

Fiedler 56
3.1.5.5.3. Similar distribution across all research sites
Linyphiids are sit-‐and-‐wait web feeders that prefer deciduous and coniferous leaf litter
or dead wood in forested environments.94 Linyphiids are also prolific ballooners, known for
their mass dispersal during autumn and winter months of juveniles and adults alike.95
I found spider individuals of this family in relatively high numbers in each research site and
sampling year (Figure 12).
Figure 12. Number of individuals of the Family Linyphiidae in research sites over time. Each bar represents the sum of three sample collection dates with ten pitfall traps each.
94 Ibid. 95 Eric Duffey, 1956, "Aerial Dispersal in a Known Spider Population," The Journal of Animal Ecology 25 (1):
85-‐111, 86.
64
16
61
48 48
116
83
4
20 17 25
79
0
20
40
60
80
100
120
# of Individuals
Research Site and Year
1990 2000 2010 Reference Forest
1990 2000 2010 Tephra Forest
1990 2000 2010 Blowdown Forest
1990 2000 2010 Pumice Plain
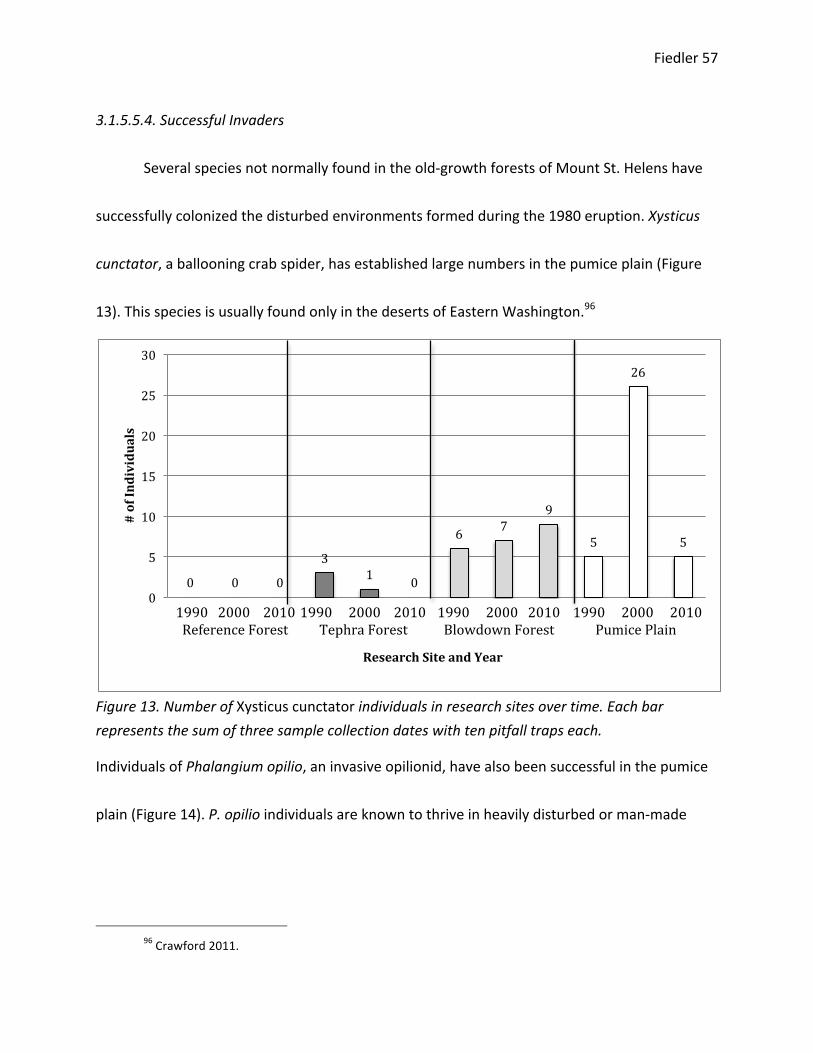
Fiedler 57
3.1.5.5.4. Successful Invaders
Several species not normally found in the old-‐growth forests of Mount St. Helens have
successfully colonized the disturbed environments formed during the 1980 eruption. Xysticus
cunctator, a ballooning crab spider, has established large numbers in the pumice plain (Figure
13). This species is usually found only in the deserts of Eastern Washington.96
Figure 13. Number of Xysticus cunctator individuals in research sites over time. Each bar represents the sum of three sample collection dates with ten pitfall traps each.
Individuals of Phalangium opilio, an invasive opilionid, have also been successful in the pumice
plain (Figure 14). P. opilio individuals are known to thrive in heavily disturbed or man-‐made
96 Crawford 2011.
0 0 0
3 1 0
6 7 9
5
26
5
0
5
10
15
20
25
30
# of Individuals
Research Site and Year
1990 2000 2010 Reference Forest
1990 2000 2010 Tephra Forest
1990 2000 2010 Blowdown Forest
1990 2000 2010 Pumice Plain

Fiedler 58
environments.97 However, each of these species shows a drop in numbers found in my samples
after the year 2000, coinciding with an increase in denser vegetation.
Figure 14. Number of Phalangium opilio individuals in the pumice plain over time. Each bar represents the sum of three sample collection dates with ten pitfall traps each.
3.1.6. First on the Scene
Crawford et al. (1995) showed that some of the first colonizers in the most disturbed
sites of Mount St. Helens, the pumice plain and the blowdown forest were ballooning spiders.
While the methods used for this sampling are not directly comparable to my sampling
methodology, the data collected by Crawford et al. (1995) can still be used to understand the
first steps of arachnid recovery. Of the 1983 samples, 23.3% of the individual arthropods
collected were spiders,98 all of which ballooned into these sites, as there were no survivors of
97Ibid. 98 Crawford et al. 1995, 64.
0
4418
1484
0 500 1000 1500 2000 2500 3000 3500 4000 4500
# of Individuals
Pumice Plain 1990 2000 2010

Fiedler 59
the initial blast in the pumice plain.99 From 1981 to 1986, 14,324 spider individuals were
collected in the pumice plain, representing 125 species (see Appendix 2 for Crawford et al. 1995
figures and Appendix 3 for corresponding figures from my sampling data).
The family Lycosidae contributed the largest number of individuals during this sampling
period, with 49% of all individuals. Linyphiids represented 34% of all individuals, and 50% of all
species.100 Crawford et al. noted that most of the spiders arriving on the pumice plain
immediately after the eruption did not become permanent residents or establish local
populations. Distant populations simply continually supplemented their numbers. However, by
1986, Crawford et al. found two lycosid species and four linyphiid species to be the first
successful colonists, as defined by evidence of successful local reproduction. By 1986, some
vegetation had also successfully colonized, thus provided protected habitat and a greater
source of arthropod prey.101 Lycosids and linyphiids still hold strong numbers on the pumice
plain, despite their differing habitat preferences and feeding guilds, suggesting some habitat
complexity at this site.
99 Ibid., 61. 100 Ibid., 64. 101 Ibid., 72.

Fiedler 60
3.1.7. Comparison to Beetle Data
Despite the later time frame of my sampling, I observed several patterns similar to those
in Parmenter et al. (2005). There appear to be several arachnid taxa that have replaced other
over time, as is seen in the fluctuations in feeding guilds and habitat preferences. Furthermore,
even 30 years after the eruption, the pumice plain arachnid populations appear to be quite
different than those in the reference forest. While the trajectory of the recovery of these
communities is unknown, we can assume that their composition is still rapidly changing.
4. When Theories and Data Collide
The communities impacted by the 1980 Mount St. Helens eruption are still undergoing
recovery. Only 30 years have passed since the eruption occurred, thus we cannot quantitatively
depict the full course of recovery for each of the disturbed sites. These preliminary data can,
however, be used to predict the trajectories of recovery. Each theory interprets these data
differently in terms of ecosystem recovery, and provides different predictions for the future of
each of these systems. I will compare how the data may be interpreted through the lens of each
theory in order to better understand the corresponding evolution of thought regarding
ecosystem recovery. I will follow the framework established in Table 1 to systematically assess
these theories. In this analysis, we must consider the limitations of my methodology, and how
these will impact the subsequent analysis. As my data only represent three points in time, any

Fiedler 61
fluctuations that are observed could be misinterpreted. These fluctuations can be signs of a
long-‐term trend or simply stochastic variation about a stable long-‐term trend. This confusion
cannot be avoided given my data and will be addressed accordingly in my analysis.
4.1. Deconstructing the Theories in Context
The 1980 eruption of Mount St. Helens acted as a disturbance impacting old-‐growth
forests to varying degrees as a function of proximity and orientation relative to the blast. Thus,
this disturbance can be classified as a parameter shift, as external forces altered the
accumulated potential and variables of the system. Each theory accounts for external
disturbances with Clementsian succession calling this disturbance ‘nudation,’ alternative stable
states using the term ‘parameter shift,’ and panarchy classifying it as ‘release.’ However,
because the eruption occurred on such a brief temporal scale, panarchy might predict that this
release will lead to revolt. Each theory describes the subsequent process differently, providing
its own language and predictions.
4.1.1. The Clementsian Succession Perspective
Each disturbed site’s responses to the eruption can be interpreted differently according
to the lens being used. Clementsian succession predicts a linear, deterministic pattern of
recovery, not allowing for novelty, and is the only theory of these three with the ability to
provide any such predictions. As the reference forest was unaffected by the blast, we can

Fiedler 62
assess the stages of recovery for each site through a comparison with the reference forest. The
data regarding habitat preference and feeding guilds show the tephra forest most closely
reflecting the reference forest, or the state from which it was disturbed. The blowdown forest
and pumice plain, however, show much more fluctuation in these patterns over the ten-‐year
periods, and more deviation from those patterns observed in the reference forest. Clementsian
succession suggests that the tephra forest is farther along in its progression towards returning
to its monoclimax. The fluctuations in the more disturbed sites do not suggest a different
trajectory, but rather the stages of migration, ecesis, competition, and reaction. Disturbed sites
with different species compositions can be interpreted as simply exhibiting different
fluctuations within these stages. Thus, these sites, according to succession, are still in the
beginning stages of recovery. As this theory follows the idea of dominance-‐controlled
community development, these disturbed sites will experience a decrease in species richness as
the most successful competitors take control.
The stages of succession, as the theory explains, result in a community of increasing
complexity and efficiency, yet as dominant competitors increase in number, less successful
species are eliminated from the community. Thus, a successional analysis of the number of taxa
for each disturbed site suggests that none of them have reached the climax state. Each
disturbed site appears to be increasing in the number of taxa throughout time, and should

Fiedler 63
exhibit a drop as the system proceeds into its climax state. Clementsian succession insists on
the existence of a monoclimax, so it is to be predicted that each of these sites, given enough
time without a disturbance, will all reach the same climax state. Again, due to the fluctuating
nature of the reference forest, successional theory would suggest that this undisturbed site has
yet to reach its climax state as well. Thus, despite being an old-‐growth forest, the reference
sites are still undergoing periodic disturbances of varying scales that have at least impacted
arachnids, if not other organisms as well. However, the reference forest is farther along in its
successional progress, as it was unaffected by the 1980 eruption, a disturbance that could
affect the entire ecosystem, rather than just certain taxa. According to succession, each
research site is moving along the same trajectory. However, each is experiencing a different
stage due to the variation of the data between sites.
4.1.2. The Alternative Stable States Perspective
Alternative stable state theory provides a much different interpretation of the
responses of these disturbed research sites. This theory suggests a relatively linear process of
recovery, but allows for multiple trajectories towards different stable states. With these
alternatives, this theory is hard to test and falsify. A shift in parameters, or in this case the 1980
eruption, created the opportunity for the variables of the system to achieve a different stable
state that was not available before, as the ecosystem perspective describes, or perhaps pushed

Fiedler 64
the site past a threshold point into a state of hysteresis. The eruption, as it disturbed the
communities that had existed in these sites, created opportunities for different arrangements
of the previously existing and newly arriving variables. Thus, the success of certain species in
the pumice plain, such as Phalangium opilio or Xysticus cunctator, could be indicative of the
system’s progression towards a different stable state. As the tephra forest is already seen to
reflect many of the patterns found in the reference forest, alternative stable state theory might
expect this research site to be returning to its previous state. This theory could allow for either
dominance or founder controlled stability, yet since these systems are still progressing, it is too
soon to determine which will dictate their recovery.
As Clementsian succession also explains, the system will increase in complexity and
efficiency throughout its recovery before reaching a final stable state. This process is reflected
in the increasing numbers of taxa in each of the research sites. As this pattern includes the
reference forest, alternative stable states theory would also suggest that this site has also not
reached its stable state. However, this theory allows these sites to be much farther along in
recovery, unlike succession, as each site is not required to be on the same trajectory. If these
sites are given enough time without a disturbance, each will reach a stable state, either similar
to the trajectory that they were previously on or a different assemblage of variables.

Fiedler 65
4.1.3. The Panarchy Perspective
Panarchy treats the 1980 eruption as a release that results in a rapid dissolution of
connectedness of the disturbed system. Following the disturbance, the system enters into the
back loop where there exists the possibility of novelty as potential increases. This process of
reorganization is reflected in rapid shifts in feeding guilds and habitat preferences in the pumice
plain. The lack of connectedness of the system, according to panarchy, has allowed for novelty
in the forms of new colonizers who otherwise would be unsuccessful in the previously intact
system, such as Phalangium opilio and Xysticus cunctator. These species have taken advantage
of this new environment and its limited competition. Their success could dictate what occurs
during the system’s progression into the front loop, or the progression from exploitation to
conservation. On the other hand, these two populations could be overwhelmed as local
competitors return or as increasing habitat complexity begins to support local forest species.
As each research site undergoes recovery, panarchy predicts that forces of opposing
scale will influence their course. Adaptive cycles of smaller and faster scale will prompt the
system in question to revolt. Thus, the invasion of ballooning spiders into disturbed sites might
cause the system to not recover in the direction of how it was before the disturbance. Adaptive
cycles of larger and slower scale, such as climate and location, will cause the system to

Fiedler 66
remember, or limit the system’s ability to adopt novel forms. For instance, while desert
dwelling Xysticus cunctator has successfully established a population in the pumice plain, it may
not be able to persist if forests do reestablish, eliminating the current desert-‐like habitat.
The progression of a system through the front loop of the adaptive cycle in a panarchy
resembles the steps outlined in Clementsian succession. Competition and immigration shifts
the species composition of the ecosystem, while the potential and connectedness of the system
increase. However, as the system progresses it becomes increasingly rigid, and thus vulnerable
to disturbances. Panarchy, therefore, suggests that the reference forest is in the conservation
stage of the adaptive cycle. Due to the rapid increase in taxa in the disturbed sites, the tephra
forest, blowdown forest, and pumice plain all appear to be in the reorganization phase of the
adaptive cycle. The pumice plain, as it is undergoing the most rapid shifts in community
composition, is the furthest away from the front loop, and the most vulnerable to novelty, if the
larger adaptive cycles allow.
4.1.4. Summary of Perspectives
Each theory interprets the same data in different ways. Using their own language and
perspective, each theory allows us to make predictions regarding the trajectory of the research
sites. These systems are rapidly changing, and only time will tell what direction they are
heading. Table 4 describes how each theory assesses the current state of each research site.

Fiedler 67
While no theory is correct or incorrect, each can drastically alter our perceptions of recovery
and ecosystem development.
Table 4. Comparison of each theory’s assessment of the state of research sites. Clementsian Succession Alternative Stable
States Panarchy
Reference Forest Reaction phase, prior to stabilization
Still transitioning to stable state
Conservation phase, building rigidity
Tephra Forest Reaction phase, prior to stabilization
Signs of resilience, transitioning to stable state along previous trajectory
Exploitation phase, remember
Blowdown Forest Migration, ecesis, competition, and reaction
Initial stages of recovery, potentially leading towards alternative stable state
Reorganization phase, likely to revolt
Pumice Plain Migration, ecesis, competition, and reaction
Initial stages of recovery, potentially leading towards alternative stable state
Reorganization phase, farthest from front loop, likely to revolt
4.2. Spotlight on Panarchy
Each of these three ecological theories interprets the same data in radically different
ways. Panarchy acts as a paradigm, integrating the Clementsian succession and alternative
stable state theories, in understanding the progression from immigration, establishment,
competition, and reorganization, and also the opportunity for alternative trajectories of
recovery. However, panarchy focuses on the transitions of a system from one stage to another,
and that it is in the constant flux that an ecosystem is most resilient. Panarchy treats
equilibrium as a transient state, as rigidity builds alongside connectedness. Panarchy might also
serve as a theoretical model to understanding the compound effects of disturbances; for

Fiedler 68
instance, the effects of a volcanic eruption, invasive species, and climate change, and how each
of these might dictate ecosystem development alone and together.
5. Putting Theory Into Practice: Conservation Implications
Even though the 1980 eruption of Mount St. Helens was a natural disturbance, we can
still use the subsequent recovery as a case study to begin to understand how Clementsian
succession, alternative stable states theory, and panarchy can be applicable to conservation
and management decisions for human disturbances. Our perspectives on recovery, stability,
rigidity, and resilience define how we consider conservation. If we consider stability to mean
“…efficiency, control, constancy, and predictability,”102 as succession and alternative stable
states would define the term, then conservation goals would reflect the desire to maintain a
system within the confines of a single state and level of efficiency. Any fluctuations in the
conditions of that system call for a response. On the contrary, if we consider stability to mean
“…persistence, adaptiveness, variability, and unpredictability,”103 as the theory of panarchy
invokes, then our goals of conservation will shift. Conservation will acknowledge the constant
flux of systems, and emphasize the need to maintain function rather than the efficiency of
specific conditions.104 Thus, we accept the multiplicity of states that an ecosystem can achieve,
102 Gunderson and Holling eds. 2002, 27. 103 Ibid., 27. 104 Ibid., 28.

Fiedler 69
as long as it continues to flow through the adaptive cycle. In this case, novelty can only be
deemed negative under panarchy if the system looses its function; otherwise, no judgment is
made by the theory. Another distinction of language that must be made is between panarchy
and resilience, as these terms are often interchanged in discourse. Panarchies describe the
ecological realities of dynamic systems, yet the term resilience reflects the goal of conservation
that follows from the theory of panarchy. Through our understanding of the dynamism of
adaptive cycles of ecological systems, we aim to maintain the resilience, and in turn the
function, of systems through conservation and preservation work.
5.1. Collapsing Panarchy
While panarchy embraces the lack of predictability and learning of systems, the cycle
can become detrimental to the health of the system. ‘Poverty traps’ occur when the adaptive
cycle collapses due to the elimination of all potential and connectedness, resulting in low
resilience. This can be prompted by a strong disturbance that continues to break down the
potential and connectedness of a system, keeping the system out of the reorganization phase
and in a constant state of release. If a system falls into a poverty trap, the effects can cascade
down to adaptive cycles of different scales. Systems can also collapse into ‘rigidity traps,’ with
high connectedness, potential, and resilience. This system would essentially preserve any
disturbances, creating maladaptive feedback. Eventually, this system would suffer from a

Fiedler 70
disturbance to a degree much greater than would be expected.105 These two traps must be
avoided by restricting perpetual disturbance or the complete lack thereof.
In order to consider conservation in the context of panarchy, it is important to know
what stage of the adaptive cycle the system is in, and what stage is most appropriate to do
some conservation action. Often times, the state of release in the back loop provides the
greatest opportunity for conservation efforts to be applied, allowing the system to reorganize
with these efforts in place.106 Panarchy also emphasizes the importance of some level of
disturbance, in order to promote learning and the resilience of a system, running counter to
successional thought.
5.2. Conservation Decisions
An example of how these theories treat conservation and restoration, and how they
result in vastly different agendas and priorities, lies in current forest management techniques in
the Pacific Northwest. Forest managers fight to balance efficient resource extraction, fire
management, and overall ecosystem health. We have found ourselves in a sort of rigidity trap
as fires have long been suppressed in many of the Northwest forests, yet at the same time
some of these forests are kept within a poverty trap, as ecosystems are not allowed enough
105 Holling 2001, 400. 106 Ibid., 402.

Fiedler 71
time to fully recover after a timber harvest. Douglas County Commissioner Joe Laurance has
testified in support of returning forests to their conditions from the year 1800. This date marks
the period before European Americans were present in the region and, according to Laurance, a
time when these forests were not dangerously packed with fuels (and thus before the
panarchic collapse).107
This restoration approach strongly reflects Clementsian successional theory, in that it is
seeking to return these forests to a former state. Alternative stable state theory would insist
that these forests are trapped in a state of hysteresis, in which they cannot be easily returned
to any previous state at all. Furthermore, these forests are actually able to reach a similar state
of productivity as in 1800, yet under vastly different conditions, or variables. Thus, we might
need to reconsider our perception of what these forests should look like. Panarchy would
declare an extreme sense of urgency in changing our current practices of maintaining what
seems to be both a poverty and rigidity trap. However, panarchical thinking would note that
any goal of a static system is unreasonable and harmful to that system’s resilience. Panarchy
might suggest that a more dynamic approach to restoration and conservation is necessary in
protecting our forests, while maintaining their function. While some environmental problems,
107 SOS Forests: Western Institute for Study of the Environment Commentary, “Douglas Co. Commissioner
Joe Laurance July 15 Testimony,” Last modified July 15, 2010, http://westinstenv.org/sosf/2010/07/15/douglas-‐co-‐
commissioner-‐joe-‐laurance-‐july-‐15-‐testimony/.

Fiedler 72
with linear progressions and clear solutions, can be most efficiently resolved with traditional
“command and control” solutions, others might be considered “wicked problems”, in their
dynamism and complexity (reflective of panarchy) and must be resolved otherwise.108
5.3. Panarchy in Action
The ideas behind panarchy are relatively new in ecological discourse, and thus there are
yet to be explicit conservation instructions that it delineates. Benson and Garmestani (2011)
begin to examine how panarchy can be applied to conservation actions, and specifically to the
National Environmental Policy Act (NEPA).109 The authors propose that panarchical thought
must be coupled with active adaptive management. Active adaptive management essentially
field-‐tests multiple hypotheses of appropriate actions, allowing for challenging and learning
from policies.110 Policies need to be developed that can account for uncertainty and accept the
dynamism of systems. However, changes would need to be made not only in the proposals and
policies that our new understanding of systems would create, but also the institutions that
implement them. According to Benson and Garmestani (2011), our current governing bodies
would not be able to support such conservation actions. The authors detail the needs of NEPA
108 M.H. Benson and A.S. Garmestani, 2011, "Embracing panarchy, building resilience and integrating
adaptive management through a rebirth of the National Environmental Policy Act," Journal of Environmental
Management, 92 (5): 1420-‐7, 1420. 109 Ibid. 110 Ibid. 2011, 1422.

Fiedler 73
to integrate iterative conservation methods, thorough and continual monitoring, and a
reaffirmed responsibility for mitigation. In doing so, conservation actions will better respect the
dynamism suggested by panarchy and tackle those “wicked problems.”111
5.4. Panarchy as Part of a Continuing Paradigm Shift
Panarchy has yet to find its place in the field of ecology, but its discourse is continually
developing. Today, much of the development of the theory can be attributed to the work of the
Resilience Alliance, an organization focused on research on the “resilience, adaptability, and
transformability” of social-‐ecological systems.112 The organization aims to further the
theoretical basis for resilience thinking and to test the ideas of adaptive cycles and applied
adaptive management.113 As the development of all ecological theories aim to do, the
Resilience Alliance promotes this research so we can better understand the complex realities of
the environment, and understand how our actions can reflect these realities the best. The
development of the theory is happening right now, as is seen in Resilience 2011 conference
held in March 2011.114 It is in the actions of the Resilience Alliance and other ecological thinkers
111 Ibid., 1425-‐1426. 112 Resilience Alliance, “About RA,” Last modified January 8, 2004,
http://www.resalliance.org/index.php/about_ra. 113 Ibid. 114 Ibid.

Fiedler 74
and scientists that we will see the development of the theory of panarchy, and, in turn, see it
reflected in future conservation actions.
5.5. Conclusion
There is no perfect ecological theory that can accurately explain and predict any change
within an ecosystem. While ecological realities remain the same, different theories will assess
and understand these realities in vastly different ways. By considering these theories in the
context of numbers and tangible patterns, we can better understand what each theory
illuminates in our understanding of ecological recovery. Panarchy progresses our thought
regarding ecological recovery and should be considered in ecological discourse and
conservation considerations. It embraces change and learning, thus acknowledging the need to
allow systems to exist in their naturally fluctuating states, or to allow for dynamism through our
conservation actions. Our frequent desire to maintain stability and eliminate any sort of change
in living systems is unrealistic and detrimental to overall ecosystem health and resilience.
Panarchy addresses these considerations, while also building from ecological thought of past
theories. However, panarchy acts as more of a paradigm, in teasing out the complexities of
dynamic systems, while deepening our understanding of potential trajectories and past
changes.

Fiedler 75
The arachnid recovery of Mount St. Helens has allowed us to deeply consider the
potential for panarchy to be applied to living ecological systems, and how it compares to
Clementsian succession and alternative stable states theory. While we have only been able to
look at the first steps of recovery of this system, comparing the predictions that each theory
makes to the recovery we may observe in the future will be even more telling of each theory’s
applicability and potential. On the other hand, conservation decisions are rarely made with
ideal data sets, but rather must be made with a limited understanding of the system in question
and its trajectory of recovery. Thus, the limitations of my data set might actually reflect realities
of the choices we make in doing conservation. Again, no theory will be deemed correct or
incorrect in its predictions, but panarchy does prove to be the best in illuminating the
dynamism of natural systems, and thus should be used in any conservation or preservation
considerations. Future analyses for better understanding how these theories describe the
recovery of ecological systems would span a longer time frame and consider the numbers more
frequently in order to observe fluctuations in community composition. However, this analysis of
arachnid recovery has already exemplified how panarchy presents a new perspective regarding
ecosystem recovery, which subsequently changes how we might view conservation and
preservation. Panarchy should be the next in the successional line of ecological thought.

Fiedler 76
6. References
Beccaloni, Jan. 2009. Arachnids. Berkeley: University of California Press. Begon, Michael, Colin R. Townsend, and John L. Harper. 2006. Ecology: from individuals to ecosystems. Malden, MA: Blackwell Pub. Beisner, B. E., D. T. Haydon, and K. Cuddington. 2003. “Alternative stable states in ecology.” Frontiers in Ecology and the Environment 1 (7). Benson, M.H., and A.S. Garmestani. 2011. "Embracing panarchy, building resilience and integrating adaptive management through a rebirth of the National Environmental Policy Act." Journal of Environmental Management. 92 (5): 1420-‐7. Bonte, Dries, Jeroen Vanden Borre, Luc Lens, and Jean-‐Pierre Maelfait. 2006. "Geographical variation in wolf spider dispersal behaviour is related to landscape structure." Animal Behaviour. 72 (3): 655. Bonte, Dries, Nele Vandenbroecke, Luc Lens, and Jean-‐Pierre Maelfait. 2003. "Low propensity for aerial dispersal in specialist spiders from fragmented landscapes." The Royal Society. 270 (1524): 1601-‐7. Cain, Michael L., William D. Bowman, and Sally D. Hacker. 2008. Ecology. Sunderland, MA: Sinauer Associates, Inc. Clements, Frederic E. 1916. Plant Succession, an Analysis of the Development of Vegetation, p. 140-‐143. In: Edward J. Kormondy, ed. 1965. Readings in Ecology. Englewood Cliffs, NJ: Prentice-‐Hall, Inc. Crawford, Rodney L., and John S. Edwards. 1986. "Ballooning Spiders as a Component of Arthropod Fallout on Snowfields of Mount Rainier, Washington, U.S.A." Arctic and Alpine Research. 18 (4): 429-‐437. Crawford, Rodney L., Patrick M. Sugg, and John S. Edwards. 1995. “Spider arrival and primary establishment on terrain depopulated by volcanic eruption at mount St. Helens, Washington.” American Midland Naturalist 133 (1): 60-‐75. Crawford, Rod. Personal communication. March 2, 2011. Crisafulli, Charles M., James A. MacMahon, and Robert R. Parmenter. 2005. Small-‐Mammal Survival and Colonization on the Mount St. Helens Volcano: 1980-‐2002, p. 199-‐218. In: Virginia H. Dale, Frederick J. Swanson, and Charles M. Crisafulli eds. 2005. Ecological Responses to the 1980 Eruption of Mount St. Helens. New York, NY: Springer. Dale, Virginia H., Frederick J. Swanson, and Charles M. Crisafulli. 2005. Disturbance, Survival, and Succession: Understanding Ecological Responses to the 1980 Eruption of Mount St. Helens, p. 3-‐11. In: Virginia H. Dale, Frederick J. Swanson, and Charles M. Crisafulli eds.

Fiedler 77
2005. Ecological Responses to the 1980 Eruption of Mount St. Helens. New York, NY: Springer. Dale, Virginia H., Frederick J. Swanson, and Charles M. Crisafulli, eds. 2005. Ecological Responses to the 1980 Eruption of Mount St. Helens. New York, NY: Springer. Derkarabetian, Shahan. Personal communication. February 11, 2011. Drury, William H. and Ian C.T. Nisbet. 1973. Succession, p. 287-‐368. In: Frank B. Golley, ed. 1977. Ecological Succession. Stroudsburg, PA: Dowden, Hutchinson & Ross. Duffey, Eric. 1956. "Aerial Dispersal in a Known Spider Population." The Journal of Animal Ecology. 25 (1): 85-‐111. Foelix, Rainer F. 1996. Biology of spiders. New York: Oxford University Press. Goldsmith, Edward. “Ecological succession rehabilitated.” Edward Goldsmith, http://www.edwardgoldsmith.org/page119.html (16 November 2010). Golley, Frank B. 1977. Ecological succession. Stroudsburg, Pa: Dowden, Hutchinson & Ross. Goodrich, Charles, Kathleen Dean Moore, and Frederick J. Swanson, eds. 2008. In the blast zone: catastrophe and renewal on Mount St. Helens. Corvallis, OR: Oregon State University Press. Gunderson, Lance H., and C. S. Holling. 2002. Panarchy: understanding transformations in human and natural systems. Washington, DC: Island Press. Holling, C. S. 1973. "Resilience and Stability of Ecological Systems." Annual Review of Ecology and Systematics. 4: 1-‐23. Holling, C.S. 2001. Understanding the Complexity of Economic, Ecological, and Social Systems. Ecosystems, 4:390-‐405. Kuhn, Thomas S. 1970. The Structure of Scientific Revolutions. Chicago, IL: The University of Chicago Press. Lewontin, Richard C. 1969. "The meaning of stability." Brookhaven Symposia in Biology. 22: 13-‐24. Odum, Eugene P. 1969. The Strategy of Ecosystem Development, p. 278-‐286. In: Frank B. Golley,
ed. 1997. Ecological Succession. Stroudsburg, PA: Dowden, Hutchinson & Ross. Parmenter, Robert R. et al. 2005. Posteruption Arthropod Succession on the Mount St. Helens Volcano: The Ground-‐Dwelling Beetle Fauna (Coleoptera), p. 139-‐150. In: Virginia H. Dale, Frederick J. Swanson, and Charles M. Crisafulli eds. 2005. Ecological Responses to the 1980 Eruption of Mount St. Helens. New York, NY: Springer. Resilience Alliance. “About RA.” Last modified January 8, 2004. http://www.resalliance.org/index.php/about_ra. Roth, Vincent D., Darrell Ubick, and N. Dupérré. 2005. Spiders of North America: an identification manual. Poughkeepsie, NY: American Arachnological Society.

Fiedler 78
Scheffer, Marten. Alternative Stable States and Regime Shifts in Ecosystems, p. 395-‐406. In: Simon A. Levin ed. 2009. The Princeton Guide to Ecology. Princeton, NJ: Princeton University Press. Simberloff, Daniel. A Succession of Paradigms in Ecology: Essentialism to Materialism and Probabilism, p. 63-‐99. In: Esa Saarinen ed. 1982. Conceptual Issues in Ecology. Boston, MA: D. Reidel Publishing Company. SOS Forests: Western Institute for Study of the Environment Commentary. “Douglas Co. Commissioner Joe Laurance July 15 Testimony.” Last modified July 15, 2010. http://westinstenv.org/sosf/2010/07/15/douglas-‐co-‐commissioner-‐joe-‐laurance-‐july-‐ 15-‐testimony/. Sutherland, John P. 1974. "Multiple Stable Points in Natural Communities." American Naturalist. 108 (964): 859-‐873. Whittaker, R.H. 1953. A Consideration of Climax Theory: The Climax as a Population and Pattern, p. 240-‐277. In: Frank B. Golley, ed. 1977. Ecological Succession. Stroudsburg, PA: Dowden, Hutchinson & Ross. Wise, David H. 1993. Spiders in ecological webs. Cambridge: Cambridge University Press. Worster, Donald. 1994. Nature's economy: a history of ecological ideas. Cambridge: Cambridge University Press.
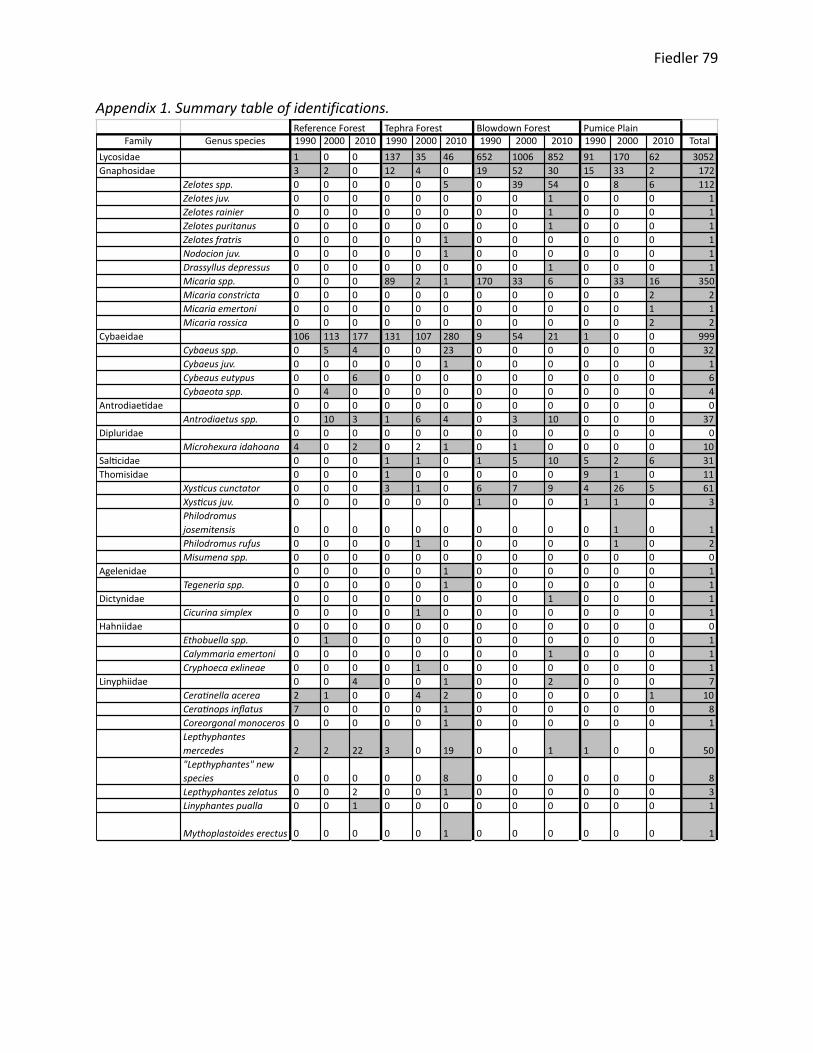
Fiedler 79
Appendix 1. Summary table of identifications.
!"#$%& '()*+,+-(.$(+ /001 2111 21/1 /001 2111 21/1 /001 2111 21/1 /001 2111 21/1 345"%
6&.4+$7"( / 1 1 /89 8: ;< <:2 /11< =:2 0/ /91 <2 81:2
')"->4+$7"( 8 2 1 /2 ; 1 /0 :2 81 /: 88 2 /92
!"#$%"&'&(() 1 1 1 1 1 : 1 80 :; 1 = < //2
!"#$%"&'*+,) 1 1 1 1 1 1 1 1 / 1 1 1 /
!"#$%"&'-./0/"- 1 1 1 1 1 1 1 1 / 1 1 1 /
!"#$%"&'(+-/%.0+& 1 1 1 1 1 1 1 1 / 1 1 1 /
!"#$%"&'1-.%-/& 1 1 1 1 1 / 1 1 1 1 1 1 /
2$3$4/$0'*+,) 1 1 1 1 1 / 1 1 1 1 1 1 /
5-.&&6##+&'3"(-"&&+& 1 1 1 1 1 1 1 1 / 1 1 1 /
7/4.-/.'&(() 1 1 1 =0 2 / /91 88 < 1 88 /< 8:1
7/4.-/.'4$0&%-/4%. 1 1 1 1 1 1 1 1 1 1 1 2 2
7/4.-/.'"8"-%$0/ 1 1 1 1 1 1 1 1 1 1 1 / /
7/4.-/.'-$&&/4. 1 1 1 1 1 1 1 1 1 1 1 2 2
?&@"($7"( /1< //8 /99 /8/ /19 2=1 0 :; 2/ / 1 1 000
96:."+&'&(() 1 : ; 1 1 28 1 1 1 1 1 1 82
96:."+&'*+,) 1 1 1 1 1 / 1 1 1 1 1 1 /
96:".+&'"+%6(+& 1 1 < 1 1 1 1 1 1 1 1 1 <
96:."$%.'&(() 1 ; 1 1 1 1 1 1 1 1 1 1 ;
A)5B47$"(C7"( 1 1 1 1 1 1 1 1 1 1 1 1 1
;0%-$3/."%+&'&(() 1 /1 8 / < ; 1 8 /1 1 1 1 89
D$-%*B$7"( 1 1 1 1 1 1 1 1 1 1 1 1 1
7/4-$<"=+-.'/3.<$.0. ; 1 2 1 2 / 1 / 1 1 1 1 /1
E"%C.$7"( 1 1 1 / / 1 / : /1 : 2 < 8/
3>4#$+$7"( 1 1 1 / 1 1 1 1 1 0 / 1 //
>6&?4+&'4+04%.%$- 1 1 1 8 / 1 < 9 0 ; 2< : </
>6&?4+&'*+,) 1 1 1 1 1 1 / 1 1 / / 1 8
@</#$3-$8+&'
*$&"8/%"0&/& 1 1 1 1 1 1 1 1 1 1 / 1 /
@</#$3-$8+&'-+1+& 1 1 1 1 / 1 1 1 1 1 / 1 2
7/&+8"0.'&(() 1 1 1 1 1 1 1 1 1 1 1 1 1
AF(%()$7"( 1 1 1 1 1 / 1 1 1 1 1 1 /
A"B"0"-/.'&(() 1 1 1 1 1 / 1 1 1 1 1 1 /
D$.5&)$7"( 1 1 1 1 1 1 1 1 / 1 1 1 /
9/4+-/0.'&/8(#"= 1 1 1 1 / 1 1 1 1 1 1 1 /
G">)$$7"( 1 1 1 1 1 1 1 1 1 1 1 1 1
C%<$:+"##.'&(() 1 / 1 1 1 1 1 1 1 1 1 1 /
9.#688.-/.'"8"-%$0/ 1 1 1 1 1 1 1 1 / 1 1 1 /
9-6(<$"4.'"=#/0"." 1 1 1 1 / 1 1 1 1 1 1 1 /
6$)&->$$7"( 1 1 ; 1 1 / 1 1 2 1 1 1 9
9"-.?0"##.'.4"-". 2 / 1 1 ; 2 1 1 1 1 1 / /1
9"-.?0$(&'/0D.%+& 9 1 1 1 1 / 1 1 1 1 1 1 =
9$-"$-B$0.#'8$0$4"-$& 1 1 1 1 1 / 1 1 1 1 1 1 /
E"(%<6(<.0%"&'
8"-4"3"& 2 2 22 8 1 /0 1 1 / / 1 1 :1
FE"(%<6(<.0%"&F'0"G'
&("4/"& 1 1 1 1 1 = 1 1 1 1 1 1 =
E"(%<6(<.0%"&'H"#.%+& 1 1 2 1 1 / 1 1 1 1 1 1 8
E/06(<.0%"&'(+.##. 1 1 / 1 1 1 1 1 1 1 1 1 /
76%<$(#.&%$/3"&'"-"4%+& 1 1 1 1 1 / 1 1 1 1 1 1 /
H(I(B().(,!4B(+5 3(->B",!4B(+5 J%4K74K),!4B(+5 L*#$.(,L%"$)

Fiedler 80
!"#$%#&'#()*+,-./0( ! ! " ! ! ! ! ! ! ! ! ! "
1,/2$3243.%/#().+&-#4( ! ! ! ! ! " ! ! ! ! ! ! "
1,/2$3243.%/#()
"05"$6.(-,./0( ! " ! ! ! ! ! ! " " ! ! #
7-,"$%,()(,8. "$ " % # & "" ' ! " " ! " ()
9.4,%$-25.)',#/",-3, ! ! ! ! ! ! ! ! ! ! ! & &
:.+-;#%.#",.)-$+085,. ! & & ! " ) ! ! ! ! ! ! "(
:.+-;#%.#",.)-$"%0#++. ! ! " ! " ( ! ! ! ! ! ! %
:.+-;#%.#",.)4#++.< ! ! ! ! ! ! ! ! ! ! ! " "
*+,-." % ( / & " ( "& " " # ! ! ("
*+,-.& #& $ ' #( #" &' $% # "& "" &( #% &'!
*+,-./ ! ! ! ( ! % $ ! ! ! ! ! "$
*+,-."! ! ! / ! & % ! ! ! ! ! $ &!
*+,-."" ! ! ! ! & ! ! ! " ! " #" #$
*+,-."$ ! ! ! ! ! / ! ! ! ! ! ! /
*+,-."% ! ! ! ! ! ! ! ! " ! ! & #
*+,-."/ ! ! ! & ( % & ! ! ! ! ! "(
*0-1232234- ! ! ! ! ! ! ! ! ! ! ! ! !
7/#./$'.).+5$8.-0+./. ! ! ! ! " ! ! ! ! ! " & (
=$5#"/0()>,?#"#%( ! ! " ! ! " ! ! ! ! ! ! &
56471892234- ! ! ! ! ! ! ! ! ! ! ! ! !
@.++,$4+0()A@25.#$4(,(B)
(4CDE ! ! " ! / & ! " ! ! ! ! ""
@25.#$4(,()F0>C ! ! ! ! ! " ! ! ! ! ! ! "
:;7928<234- ! ! ! ! ! ! ! ! ! ! ! ! !
@+05,$%.)F0>C ! ! ! ! ! " ! ! ! ! ! ! "
*-;-6234- ! ! ! ! ! ! ! ! ! ! ! ! !
G($*+.)4.-,*-. ! ! " ! ! ! ! ! ! ! ! ! "
*+,-.' & & & ! " % ! ! ! ! ! ! "#
*+,-.) ! ! & ! ! ! ! ! " ! ! $ '
*+,-."& ! ! " ! ! # ! ! ! ! ! ! (
*+,-."# ! ! ! ! ! ' ! ! # ! ! ! ""
*+,-."( ! ! ! ! ! " ! ! ! ! ! ! "
!"#$%&"'&()*&+ ! " " ! ! ! ! ! ! ! ! ! &
,*-$" &!% #! &%! ""' ('/ &%"& "% (") &(# (( ! "! ((($
.)*/*&+$" &/ # ( &) "!" &/ (' & ! ! ! ! &("
=04;4<>2234- 13.+.%?,08)$4,+,$ ! " $ ! / $$ ! ! ! ! (("' "('( $)/!
?494@8<234- 9.".-0()(44C " ! $ ! " ! ! # & ! ! ! "&
*124-<8<+@0234- 1.".%$%2-30()5"0%%#0( ! ! ! ! " " ! ! ! ! ! ! &
:-14A8;4B64C34-
H#(4#"$%#8.(/$8.)
8$'#(/. " & " ! ! ! ! ! ! ! ! ! (
D-64BA864C34- I#%'"$+.(8.)8,".5,+# $ " ( $ & % ! ) ! ! ! ! #&
*124-<8<+@0234-
7-+#"$50%0()
%$%',8$"43,$0( & ! " ! ! ! ! & & ! ! ! /
*124-<8<+@0234- J#/.%$%2-30()(#/0+0( ! " ! ! ! ! ! " ! ! ! ! &
?494@8<234- 7.5.-$%)$--,'#%/.+,( &% "/ && & / %" ' " ! ! ! ! "((
E<23-<CF-3 &! #% #) () #% #& "/ "( #" / "$ ' #!(
*8A4; G<32H2374;B (%' &($ %!# %&% '%! #&)' "!#! "%$% "#!! ")( (/#$ "%)!
I462;2-B "! "& "( "! "% "$ ' "# "& / / '
J-<-14 "$ "/ ") "" &! &( ' "$ "% ) ' ""
?,-@2-B "' && &' "% &' #) "( "' &# "" "" ")

Fiedler 81
Appendix 2. Distribution of 1981-‐1985 pumice plain spider samples. 2a shown as number of individual specimens per family. 2b shown as number of species per family.115
115 Crawford et al. 1995, 65.
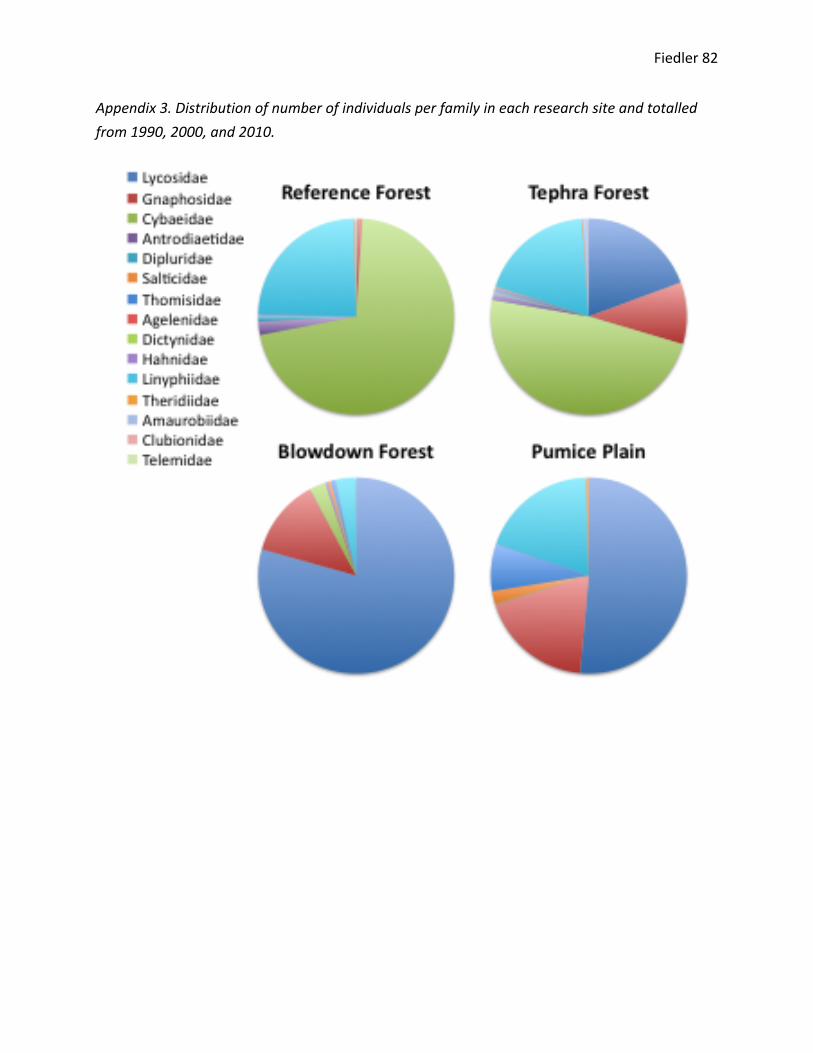
Fiedler 82
Appendix 3. Distribution of number of individuals per family in each research site and totalled from 1990, 2000, and 2010.



![ACCOUNTANCY EXAMINING BOARD[193A]](https://static.fdokumen.com/doc/165x107/6323acc9be5419ea700eb5e1/accountancy-examining-board193a.jpg)
















