
Opposite Fate of Endocytosed CCR7 and Its Ligands: Recycling versus Degradation1
11
of June 15, 2014. This information is current as Ligands: Recycling versus Degradation Opposite Fate of Endocytosed CCR7 and Its Carolina Otero, Marcus Groettrup and Daniel F. Legler http://www.jimmunol.org/content/177/4/2314 doi: 10.4049/jimmunol.177.4.2314 2006; 177:2314-2323; ; J Immunol References http://www.jimmunol.org/content/177/4/2314.full#ref-list-1 , 41 of which you can access for free at: cites 62 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2006 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on June 15, 2014 http://www.jimmunol.org/ Downloaded from by guest on June 15, 2014 http://www.jimmunol.org/ Downloaded from
Transcript of Opposite Fate of Endocytosed CCR7 and Its Ligands: Recycling versus Degradation1

of June 15, 2014.This information is current as
Ligands: Recycling versus DegradationOpposite Fate of Endocytosed CCR7 and Its
Carolina Otero, Marcus Groettrup and Daniel F. Legler
http://www.jimmunol.org/content/177/4/2314doi: 10.4049/jimmunol.177.4.2314
2006; 177:2314-2323; ;J Immunol
Referenceshttp://www.jimmunol.org/content/177/4/2314.full#ref-list-1
, 41 of which you can access for free at: cites 62 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2006 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 15, 2014
http://ww
w.jim
munol.org/
Dow
nloaded from

Opposite Fate of Endocytosed CCR7 and Its Ligands:Recycling versus Degradation1
Carolina Otero, Marcus Groettrup, and Daniel F. Legler2
The chemokine receptor CCR7 and its ligands CCL19 and CCL21 play a crucial role for the homing of lymphocytes and dendriticcells to secondary lymphoid tissues. Nevertheless, how CCR7 senses the gradient of chemokines and how migration is terminatedare poorly understood. In this study, we demonstrate that CCR7(-GFP) is endocytosed into early endosomes containing transferrinreceptor upon CCL19 binding, but less upon CCL21 triggering. Internalization of CCR7 was independent of lipid rafts but reliedon dynamin and Eps15 and was inhibited by hypertonic sucrose, suggesting clathrin-dependent endocytosis. After chemokineremoval, internalized CCR7 recycled back to the plasma membrane and was able to mediate migration again. In contrast,internalized CCL19 was sorted to lysosomes for degradation, showing opposite fate for endocytosed CCR7 and its ligand. TheJournal of Immunology, 2006, 177: 2314–2323.
L eukocyte traffic is fundamental for immune regulationand, hence, is highly coordinated. Tissue- and microen-vironment-selective leukocyte homing is the basis for this
organization. It is now well established that cell migration is or-chestrated mainly by chemokines which, together with adhesionmolecules, deliver the key signal that allows leukocytes to trans-migrate from the bloodstream into the tissue (1–3). All chemokinesact through signaling via seven-transmembrane domain G protein-coupled cell surface receptors, but migration properties varygreatly among different cell types. The central role of the chemo-kine receptor CCR7 and its ligands CCL19 (ELC, Exodus-3, MIP-3�, and CK�11) and CCL21 (SLC, Exodus-2, 6Ckine, andTCA-4) in the homing to secondary lymphoid organs is undis-puted. CCR7 is highly expressed on naive T cells and to a lowerlevel on B cells. A transient increase in CCR7 expression is foundupon T cell activation (4), whereas T cell differentiation towardeffector cells is accompanied by the down-regulation of the recep-tor on the cell surface (5). However, in dendritic cells, CCR7 ex-pression is induced upon maturation (6–8). Mice lacking CCR7show delayed kinetics in Ab responses, delayed-type hypersensi-tivity reactions, and morphological abnormalities in secondarylymphoid organs as a consequence of an impaired homing of ma-ture dendritic cells and lymphocytes (9, 10). The fact that CCR7ligands are mandatory for the homing to secondary lymphoid or-gans has been demonstrated in plt/plt mice lacking CCL19 andCCL21 (11–14).
Although CCR7 and its ligands are essential for eliciting a po-tent cellular immune response, CCR7 signaling and its regulation
is still sparsely investigated (15–20). In particular, information onhow CCR7-mediated migration is stopped after a cell has arrivedat its final destination within the lymph node has remained unclear.CCL19 and CCL21 are both produced by stroma cells within theT cell zone (12). Remarkably, CCL21 is transcytosed to high en-dothelial venules (HEV)3 (21) and mediates LFA-1-mediated ar-rest of the recruited T lymphocytes (22–24). Thus, T lymphocytesand dendritic cells that home to the T zone of lymph nodes seemfirst to be recruited to HEV by CCL21, but then the CCL21 signalmust be overcome by an attraction signal provided by CCL19/CCL21 derived from the T zone. B cells within the lymph nodethat have seen an Ag migrate directionally toward the B zone–Tzone boundary along a gradient of CCL21 (and eventually CCL19)to encounter T cells (25).
One way of rendering a cell unresponsive to chemokines is re-ceptor internalization. Chemokine receptor endocytosis is best de-scribed for the HIV coreceptors CCR5 and CXCR4 but followsdistinct mechanisms (26). Remarkably, binding of CXCL12 toCXCR4 leads to the ubiquitylation of the receptor followed by itsdegradation in lysosomes (27, 28). In contrast, endocytosed CCR5is recycled back to the plasma membrane (29–31). Strikingly,CCR7 internalization was observed by CCL19 triggering, but notby stimulation with CCL21 (32), although binding affinities andG-protein activation are comparable (4, 18). Of note, CCR7 de-sensitization through receptor phosphorylation and �-arrestinbinding was enhanced by CCL19 stimulation, compared withCCL21 (18), whereas T cell polarization mediated by the chemo-kines was indistinguishable (17). However, up to now, the mech-anism of CCR7 signaling and trafficking remains largely unclear,and there is currently no information on the fate of CCL19 afterCCR7 endocytosis.
Cell surface receptors can be internalized by two segregatedpathways: clathrin-dependent and clathrin-independent, lipid raft/caveolae-dependent endocytosis (33, 34). The classical clathrin-dependent pathway is well characterized. Clathrin-coated pits atthe plasma membrane bud and pinch off in a dynamin- and adaptorprotein (such as Eps15)-dependent manner to form clathrin-coatedvesicles. After endocytosis, clathrin-coated vesicles are uncoated
Biotechnology Institute Thurgau, University of Konstanz, Tagerwilen, Switzerland;and Department of Biology, Division of Immunology, University of Konstanz, Kon-stanz, Germany
Received for publication November 10, 2005. Accepted for publication May24, 2006.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the German Research Foundation (DFG, TR-SFB 11),the Thurgauische Stiftung fur Wissenschaft und Forschung, and the State Secretariatfor Education and Research (Section 16 law of research). D.F.L. is a recipient of acareer development award from the Dr. Max Cloetta Foundation, Zurich, Switzerland.2 Address correspondence and reprint requests to Dr. Daniel F. Legler, BiotechnologyInstitute Thurgau, University of Konstanz, Konstanzerstrasse 19, CH-8274 Tager-wilen, Switzerland. E-mail address: [email protected]
3 Abbreviations used in this paper: HEV, high endothelial venule; EGFP, enhancedGFP; HA, hemagluttinin; MCD, methyl-�-cyclodextrin; VSV, vesicular stomatitisvirus; wt, wild type.
The Journal of Immunology
Copyright © 2006 by The American Association of Immunologists, Inc. 0022-1767/06/$02.00
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

and fuse with the early endosomes, the central control organellesfor sorting receptors. Either receptors recycle back to the plasmamembrane via recycling endosomes or are directed to late endo-somes and lysosomes for degradation. Alternatively, receptors canbe endocytosed in a lipid raft/caveolae-dependent manner. Thispathway is ill defined but largely depends on cellular cholesterol(33–35).
In this study, we investigated the route of internalization and thetrafficking of CCR7 by monitoring a newly generated GFP-taggedCCR7. In addition, we tracked CCL19 after receptor binding by achemokine-Fc chimera. Analysis of CCR7 endocytosis and inves-tigations on the routes of CCL19 and CCL21 after receptor trig-gering is critical for a better understanding of how immune cells,such as dendritic cells and lymphocytes, sense a chemokine gra-dient originating in secondary lymphoid organs.
Materials and MethodsAbs and reagents
Abs were obtained from the following sources: PE-Cy5-labeled rat anti-human CCR7 (clone 3D12; BD Pharmingen), goat anti-human IgG1HRP
(Santa Cruz Biotechnology); streptavidin-Cy3 and streptoavidin-FITC(Jackson ImmunoResearch Laboratories); mouse anti-phospho ERK-1/2and mouse anti-total ERK-1/2 (Cell Signaling Technology); mouse anti-hemagluttinin (HA), (Sigma-Aldrich); HRP-conjugated goat anti-mouseIgG (DakoCytomation); and mouse anti-proteasome subunit C7 (�1; pro-vided by Dr. K. Scherrer, Paris, France). Alexa Fluor 546-labeled trans-ferrin and lysotracker Red DND-99 were from Molecular Probes. Humanchemokines CCL19 and CCL21 were purchased from PromoCell. Mono-biotinylated human CCL19 was from RMF Dictagene. Streptavidin-per-oxidase, filipin III, methyl-�-cyclodextrin (MCD), sucrose, cycloheximide,chloroquine, and protein A-Sepharose were obtained from Sigma-Aldrich.Fluo-3/AM was purchased from Calbiochem.
Cells and transfection
The human embryonic kidney cell line, HEK293, was grown in DMEM(Invitrogen Life Technologies) with 10% (v/v) FBS. HEK293 cells werestably transfected in 10-cm dishes by the calcium phosphate procedure.Cell clones were established by limiting dilution in the presence of 0.8mg/ml G-418 (Invitrogen Life Technologies). The pre B cell line 300-19(4, 36) was grown in RPMI 1640 (Invitrogen Life Technologies) with 10%(v/v) FBS, 10% 2-ME, and 2 mM nonessential amino acids. Stable trans-fection was performed by electroporation, and clones were achieved bylimiting dilution as described (4, 36). The human T cell line CEM wascultured in RPMI 1640 supplemented with 10% (v/v) FBS.
Human PBL were isolated from healthy donors by separation on Ficoll-Paque (Pharmacia Biotech), followed by depletion of monocytes usinganti-CD14-conjugated magnetic microbeads (Miltenyi Biotec).
Cells (2 � 106/ml) were cultured in RPMI 1640 supplemented with 10%(v/v) FBS, 10% 2-ME, and 2 mM nonessential amino acids. PBL wereactivated for 4–6 days with 200 U/ml IL-2 and 1 �g/ml PHA-L(Sigma-Aldrich).
Construction of expression plasmids
The entire open reading frame of human CCR7 was amplified by PCR fromSR�puro-CCR7 (4) using the primers CCR7se2 (5�-ATA GAA TTC CGTCAT GGA CCT GGG GAA AC, restriction site underlined) and CCR7as(5�-TAT GCG GCC GCT GGG GAG AAG GTG GTG) and subclonedinto the EcoRI/NotI sites of pcDNA3 (Invitrogen Life Technologies). En-hanced GFP (EGFP) was fused to the N terminus of CCR7 by PCR am-plifying GFP from pEGFP-N1 (Clontech) and subcloning into the XhoI/XbaI sites of pcDNA3-CCR7 using the primers EGFPse (5�-AAA CTCGAG CAG TGA GCA AGG GCG AGG) and EGFPas (5�-AAA TCTAGACTA CTT GTA CAG CTC GTC). A VSV-tagged CCR7 was clonedby PCR amplification of CCR7 using the primers CCR7se3 (5�-TAT GAATTC GAC CTG GGG AAA CCA ATG AAA AGC) and CCR7as3 (5�-TAA TCT AGA CTA TGG GGA GAA GGT GGT G) and subcloning intothe EcoRI/XbaI sites of pCR3-VSV (MT044; provided by M. Thome, Uni-versity of Lausanne, Epalinges, Switzerland). The CCR7-HA construct wasmade by replacing the GFP (XhoI/XbaI) with the annealed oligonucleotidesCCR7-HAse (5�-TCG AGC ATA CCC ATA CGA CGT CCC AGA CTACGC TTA GT) and CCR7-HAas (5�-CTA GAC TAA GCG TAG TCTGGG ACG TCG TAT GGG TAT GC) coding for the HA tag. 300-19 cellsexpressing wild-type (wt), VSV- or HA-tagged CCR7 migrated similarly in
response to CCL21, indicating that the tags do not affect CCR7 functions(data not shown).
Human CCL19 and CCL21 were amplified by PCR using the followingprimers: SLCse (5�-ATA ATA GGA TCC ACA GAC ATG GCT CAGTCA C), SLCas (5�-TAT TAA GAA TTC TGG CCC TTT AGG GGTCTG), ELCse (5�-ATAT AAG CTTCCC TCC ATG GCC CTG) andELCas (5�-TTAT GAA TTC ACT GCT GCG GCG CTT C) and pCR2-SLC and pCR2-ELC (provided by O. Yoshie, Kinki University, Osaka,Japan) as template. Amplified DNAs were subcloned into the HindIII/EcoRI sites of pCR3-Fc (PS521; provided by P. Schneider (University ofLausanne, Eplinges, Switzerland).
GFP-tagged dynamin II (wt, K44A) constructs (37) and GFP-tagged Eps15 constructs (38) were obtained from U. Greber (University of Zurich,Zurich, Switzerland).
Flow cytometry
Cells were washed twice with FACS buffer (PBS containing 2% FBS and5 mM EDTA) and, where required, incubated with the respective Abs for30 min at 4°C. Cells were washed twice and fluorescence was acquired bya FACScan II using CellQuest software (BD Biosciences). Data were an-alyzed with the FlowJo software (Tree Star).
Chemotaxis
Chemotaxis of 300-19 cells was measured by migration through a poly-carbonate filter of 5-�m pore size in 24-well Transwell chambers (CorningCostar). Cell culture medium (600 �l) containing indicated doses of che-mokine, or medium alone as a control for spontaneous migration, wasadded to the lower chamber; a total of 1 � 105 cells in 100 �l was addedto the upper chamber. After 3 h of incubation at 37°C, a 500-�l aliquot ofthe cells that migrated to the bottom chamber was counted by flow cytom-etry acquiring events for a fixed time period of 60 s using CellQuest soft-ware. The number of migrated cells was expressed as percentage of inputcells.
Chemokine-mediated changes in intracellular free calciumconcentrations
Cells were washed twice with Ca2� buffer (145 mM NaCl, 5 mM KCl, 1mM Na2HPO4, 1 mM MgCl2, 5 mM glucose, 1 mM CaCl2, and 10 mMHEPES (pH 7.5)) and resuspended at 1 � 106 cells/ml. Cells were loadedwith 1.5 �l/ml Fluo-3/AM (4 mM in DMSO) for 30 min at 37°C. Cellswere washed, and chemokine-induced calcium mobilization-related fluo-rescence changes of Fluo-3 were measured by flow cytometry.
Confocal laser scanning microscopy
Transfected HEK293 cells were grown overnight on coverslips. 300-19cells were incubated for 1 h on coverslips coated with poly-L-lysin (Sigma-Aldrich). If not stated otherwise, cells were treated with 2 �g/ml CCL19 orCCL21, 3 �g/ml biotinylated CCL19, 10 �g/ml CCL19-Fc or CCL21-Fc,50 �g/ml transferrin, or 50 nM lysotracker. Cells were washed twice withPBS and fixed for 10 min with 4% paraformaldehyde followed by threewashing steps with PBS. For intracellular stainings, cells were permeabil-ized with 0.2% Triton X-100 (Fluka) for 10 min and washed with 0.2%gelatin in PBS. Ab staining of cells was preformed at room temperature for40 min, and cells were washed five times and mounted on glass slides usingFluoromount-G (Southern Biotechnology Associates). Immunofluores-cence was analyzed by a confocal microscope (LSM 510; Zeiss) with a�63 Plan-Apochromat objective (aperture � 1.4). Images were acquiredusing LSM 510 software (Zeiss).
Western blotting
Cells were lysed with 1% Triton X-100 in 150 mM NaCl, 50 mM HEPES,0.1 M EGTA, 2 mM MgCl2, 10% glycerol containing leupeptin, aprotinin,and pepstatin (1 �g/ml each; Roche). Proteins from total cell lysates wereresolved by SDS-PAGE and transferred to Protran nitrocellulose mem-brane (Schleicher & Schuell Microscience). Membranes were blocked withPBS containing 5% of low-fat dry milk and incubated with the respectiveAbs overnight at 4°C or for 1 h at room temperature on a rocking plate.After washing, HRP-conjugated secondary Abs were bound and detectedusing ECL (Pierce/Socochim).
ResultsDifferential endocytosis of CCR7 by CCL19 and CCL21
The homing of lymphocytes and dendritic cells largely depends onthe attraction of the cells by the chemokines CCL19 and CCL21.
2315The Journal of Immunology
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

At its final destination, the migratory signal needs to be shut off,which normally occurs by chemokine receptor down-modulationor receptor desensitization. To unravel the mechanism how CCR7is silenced, we investigated the internalization of CCR7 in IL-2-and PHA-activated human PBL. CCR7 endocytosis by CCL19 wasreadily observed in a concentration- and time-dependant manner(Fig. 1). Cell surface expression of CCR7 on T cells was alreadyreduced by 15% at a CCL19 concentration of 30 ng/ml. More than60% of CCR7 was internalized in the presence of 3 �g/ml CCL19(Fig. 1A). Endocytosis was rapid, because after 2 min of chemo-kine addition, 30% of CCR7 disappeared from the plasma mem-brane. Maximal internalization was reached after 30 min of incu-bation (Fig. 1B). Interestingly, CCR7 endocytosis by CCL21 wasobserved only at high chemokine concentrations (�300 ng/ml) andreached a maximum of �25% (Fig. 1A). Our data on CCL19-mediated endocytosis of CCR7 are largely in agreement with aprevious study by Bardi et al. (32), although they did not find anyinternalization by CCL21 in T cells at all.
Generation of a fluorescent fully functional CCR7
To further investigate CCR7 localization and trafficking, we fusedthe enhanced GFP to the C terminus of human CCR7. We stablyexpressed CCR7-GFP in the murine pre-B cell line 300-19, a cellline that does not respond to CCL19 and CCL21 (4, 36). CCR7-GFP transfected cells readily migrated in response to CCL21, sim-ilar to 300-19 cells expressing wt CCR7, whereas CCR7-GFP-positive cells did not migrate in a Transwell chemotaxis assay inthe absence of chemokines (Fig. 2A). The phosphorylation of theERK-1 and ERK-2 is an early and transient event after chemokinetriggering (16). To test whether CCR7-GFP is fully functional, westimulated 300-19-transfected cells for various time points withCCL21 and analyzed the phosphorylation of ERK-1/2 by Westernblot analysis. Identical kinetics and potency of ERK-1/2 activationupon CCL21 triggering was observed for cells expressing CCR7-GFP and wt CCR7 (Fig. 2B). 300-19 cells are ideal for testingchemotaxis but not for morphological and trafficking studies.Therefore, we stably transfected the human embryonic kidney cellline HEK293 with CCR7-GFP. CCL21 stimulation of HEK293cells expressing CCR7-GFP resulted in the phosphorylation ofERK-1/2 (Fig. 2C), comparable to 300-19 transfectants. As ex-pected, CCR7-GFP mainly localized to the plasma membrane ofboth transfected cell lines as assessed by confocal microscopy(Fig. 2D). These data provide clear evidence that CCR7-GFP isfully functional.
CCR7-GFP colocalizes with early and recycling endosomes butnot with lysosomes
To investigate the intracellular trafficking of CCR7, we stimulatedHEK293 cells expressing CCR7-GFP with CCL19 and CCL21.After 5 min of CCL19 stimulation, CCR7-GFP was endocytosedand appeared as punctuated structures within the cell (Fig. 3A).Some CCR7-GFP was still present at the plasma membrane, con-firming our data from primary T cells. Upon CCL21 stimulation,CCR7-GFP remained mainly at the plasma membrane, althoughintracellular CCR7-GFP spots were reproducibly observed (datanot shown), confirming that CCL19 triggers CCR7 internalizationmore efficiently than does CCL21. Internalized G protein-coupledreceptors are generally degraded in lysosomes or recycled back tothe plasma membrane via early and recycling endosomes. To in-vestigate these two possibilities for CCR7, we incubated HEK293-CCR7-GFP cells with CCL19 together with Alexa Fluor 546-la-beled transferrin. The trafficking of the iron transport protein
FIGURE 1. Internalization of CCR7 in human PBL. A, Human PBLwere cultured for 4–6 days in the presence of IL-2 and PHA and incubatedwith graded concentrations of CCL19 (circles) or CCL21 (triangles) for 30min at 37°C. Cell surface expression of CCR7 was determined by flowcytometry using the mAb 3D12 against CCR7 which can still recognize theepitope in the presence of bound ligand. B, The same T cells were incu-bated for various time points with 3 �g/ml chemokines, and cell surfaceexpression of CCR7 was determined by flow cytometry. Mean values ofthree independent experiments are shown. Error bars are �1%.
FIGURE 2. Characterization of CCR7-GFP. A, Migration of the murinepre-B cell line 300-19 stably transfected with either human CCR7 wt orCCR7-GFP in response to CCL21 (1 �g/ml) was assessed in a Transwellchemotaxis assay. After 3 h of incubation at 37°C, cells in the lower cham-ber were collected and counted by flow cytometry. Mean values and SD ofthree independent experiments are depicted as percentage of migrated cells.B, 300-19 cells expressing either CCR7 wt or CCR7-GPF were incubatedwith 2 �g/ml CCL21 for indicated time points and lyzed, and the activationof ERK-1/2 was determined by Western blot analysis using an Ab recog-nizing the phosphorylated forms of ERK-1 and ERK-2 (pERK-1/2). An Abagainst total ERK-2 (tERK-2) was used to ensure equal protein loading. C,The activation of ERK-1/2 was confirmed in HEK293 cells expressingCCR7-GFP. D, Cell surface expression of transfected CCR7-GFP in300-19 and HEK293 cells was determined by confocal microscopy. Scalebars,10 �m.
2316 OPPOSITE FATE OF CCR7 AND CCL19 TRAFFICKING
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
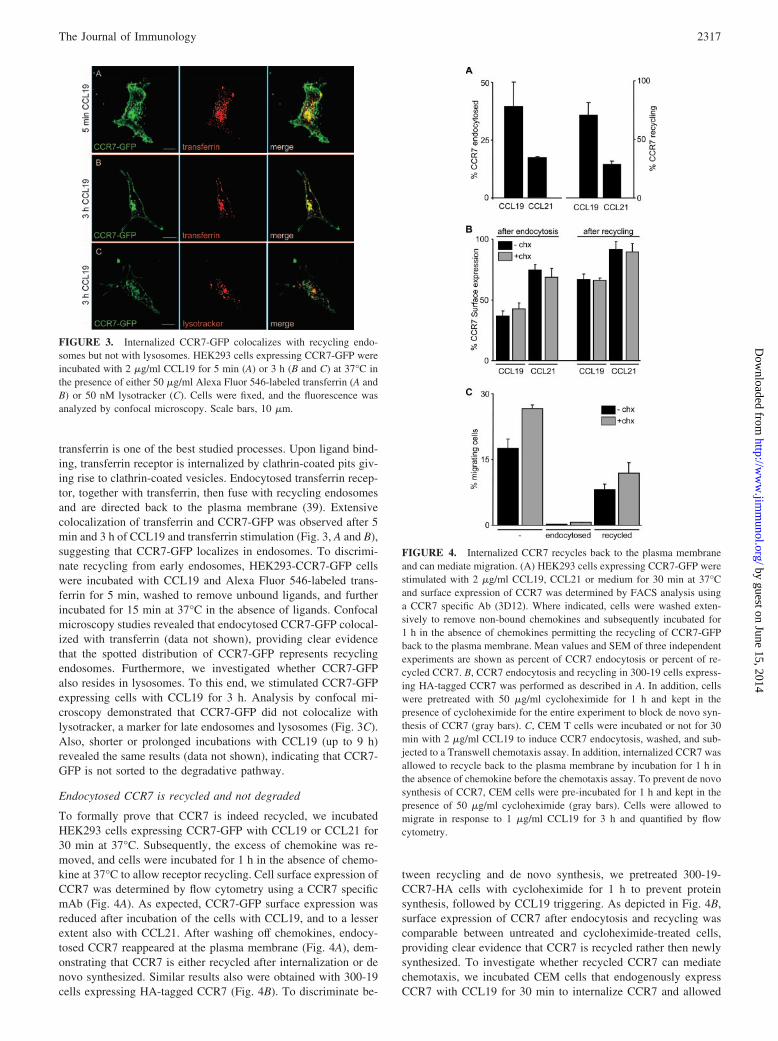
transferrin is one of the best studied processes. Upon ligand bind-ing, transferrin receptor is internalized by clathrin-coated pits giv-ing rise to clathrin-coated vesicles. Endocytosed transferrin recep-tor, together with transferrin, then fuse with recycling endosomesand are directed back to the plasma membrane (39). Extensivecolocalization of transferrin and CCR7-GFP was observed after 5min and 3 h of CCL19 and transferrin stimulation (Fig. 3, A and B),suggesting that CCR7-GFP localizes in endosomes. To discrimi-nate recycling from early endosomes, HEK293-CCR7-GFP cellswere incubated with CCL19 and Alexa Fluor 546-labeled trans-ferrin for 5 min, washed to remove unbound ligands, and furtherincubated for 15 min at 37°C in the absence of ligands. Confocalmicroscopy studies revealed that endocytosed CCR7-GFP colocal-ized with transferrin (data not shown), providing clear evidencethat the spotted distribution of CCR7-GFP represents recyclingendosomes. Furthermore, we investigated whether CCR7-GFPalso resides in lysosomes. To this end, we stimulated CCR7-GFPexpressing cells with CCL19 for 3 h. Analysis by confocal mi-croscopy demonstrated that CCR7-GFP did not colocalize withlysotracker, a marker for late endosomes and lysosomes (Fig. 3C).Also, shorter or prolonged incubations with CCL19 (up to 9 h)revealed the same results (data not shown), indicating that CCR7-GFP is not sorted to the degradative pathway.
Endocytosed CCR7 is recycled and not degraded
To formally prove that CCR7 is indeed recycled, we incubatedHEK293 cells expressing CCR7-GFP with CCL19 or CCL21 for30 min at 37°C. Subsequently, the excess of chemokine was re-moved, and cells were incubated for 1 h in the absence of chemo-kine at 37°C to allow receptor recycling. Cell surface expression ofCCR7 was determined by flow cytometry using a CCR7 specificmAb (Fig. 4A). As expected, CCR7-GFP surface expression wasreduced after incubation of the cells with CCL19, and to a lesserextent also with CCL21. After washing off chemokines, endocy-tosed CCR7 reappeared at the plasma membrane (Fig. 4A), dem-onstrating that CCR7 is either recycled after internalization or denovo synthesized. Similar results also were obtained with 300-19cells expressing HA-tagged CCR7 (Fig. 4B). To discriminate be-
tween recycling and de novo synthesis, we pretreated 300-19-CCR7-HA cells with cycloheximide for 1 h to prevent proteinsynthesis, followed by CCL19 triggering. As depicted in Fig. 4B,surface expression of CCR7 after endocytosis and recycling wascomparable between untreated and cycloheximide-treated cells,providing clear evidence that CCR7 is recycled rather then newlysynthesized. To investigate whether recycled CCR7 can mediatechemotaxis, we incubated CEM cells that endogenously expressCCR7 with CCL19 for 30 min to internalize CCR7 and allowed
FIGURE 3. Internalized CCR7-GFP colocalizes with recycling endo-somes but not with lysosomes. HEK293 cells expressing CCR7-GFP wereincubated with 2 �g/ml CCL19 for 5 min (A) or 3 h (B and C) at 37°C inthe presence of either 50 �g/ml Alexa Fluor 546-labeled transferrin (A andB) or 50 nM lysotracker (C). Cells were fixed, and the fluorescence wasanalyzed by confocal microscopy. Scale bars, 10 �m.
FIGURE 4. Internalized CCR7 recycles back to the plasma membraneand can mediate migration. (A) HEK293 cells expressing CCR7-GFP werestimulated with 2 �g/ml CCL19, CCL21 or medium for 30 min at 37°Cand surface expression of CCR7 was determined by FACS analysis usinga CCR7 specific Ab (3D12). Where indicated, cells were washed exten-sively to remove non-bound chemokines and subsequently incubated for1 h in the absence of chemokines permitting the recycling of CCR7-GFPback to the plasma membrane. Mean values and SEM of three independentexperiments are shown as percent of CCR7 endocytosis or percent of re-cycled CCR7. B, CCR7 endocytosis and recycling in 300-19 cells express-ing HA-tagged CCR7 was performed as described in A. In addition, cellswere pretreated with 50 �g/ml cycloheximide for 1 h and kept in thepresence of cycloheximide for the entire experiment to block de novo syn-thesis of CCR7 (gray bars). C, CEM T cells were incubated or not for 30min with 2 �g/ml CCL19 to induce CCR7 endocytosis, washed, and sub-jected to a Transwell chemotaxis assay. In addition, internalized CCR7 wasallowed to recycle back to the plasma membrane by incubation for 1 h inthe absence of chemokine before the chemotaxis assay. To prevent de novosynthesis of CCR7, CEM cells were pre-incubated for 1 h and kept in thepresence of 50 �g/ml cycloheximide (gray bars). Cells were allowed tomigrate in response to 1 �g/ml CCL19 for 3 h and quantified by flowcytometry.
2317The Journal of Immunology
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

the receptor to recycle back to the plasma membrane for 1 h. Asexpected, cells with internalized CCR7 did not migrate in responseto CCL19, whereas cells with reexpressed CCR7 migrated towardCCL19 (Fig. 4C). Similar results were obtained with 300-19 cellsexpressing either CCR7-GFP or CCR7-HA (data not shown). Re-cycled rather than newly synthesized CCR7 was responsible forchemotaxis, because pretreatment of CEM cells with cyclohexi-mide did not hamper migration (Fig. 4C). Furthermore, recycledCCR7 elicited the mobilization of cytosolic free calcium uponCCL19 stimulation (data not shown), providing clear evidence thatrecycled CCR7 is biologically functional.
To examine the overall rate of CCR7 degradation, potentially bya nonlysosomal pathway, we performed a degradation assay in thepresence of both chemokines. HEK293-CCR7-GFP cells were in-cubated for up to 9 h with CCL19 or CCL21 in the presence orabsence of cycloheximide, to monitor on the one hand the steady-state level of the protein and, in contrast, the impact of de novosynthesis. Using flow cytometry, we found no evidence for CCR7degradation, because the fluorescence derived from CCR7-GFPwas not reduced upon chemokine triggering (Fig. 5A). To corrob-orate these data, we also investigated the degradation of CCR7 byWestern blotting. HEK293 cells expressing CCR7-HA were incu-bated with CCL19 or CCL21 for up to 6 h at 37°C, and the amountof CCR7-HA was determined from total cell lysates using an anti-HA Ab (Fig. 5B). Again, we found no evidence for CCR7 degra-dation, suggesting that the half-life of the receptor is very long.
Generation of functional recombinant CCL19-Fc and CCL21-Fcchemokine fusion proteins
To monitor the fate of CCL19 and CCL21 once they bound toCCR7, we generated expression constructs encoding for chemo-kines fused to the Fc part of human IgG1 as there are no good Absagainst the chemokines available. We expressed human CCL19-Fcand human CCL21-Fc in HEK293 cells and purified the recombi-nant fusion proteins from the supernatants over protein-A columns.Both proteins were purified with a relative mass of �40 kDa in a
monomeric state after reduction and of �80 kDa in a nonreduceddimeric form as judged by SDS-PAGE followed by Coomassiebrilliant blue staining (data not shown). The biological activity ofthe chemokine Fc fusion proteins was tested by the ability to mo-bilize intracellular free calcium and to induce chemotaxis. 300-19cells expressing CCR7 were loaded with Fluo-3/AM and subse-quently exposed to the chemokines, and the calcium-dependentchange in fluorescence was measured over time. Challenging 300-19-CCR7 cells with CCL19, CCL21, or the corresponding chemo-kine-Fc-fusion proteins elucidated comparable transient rises in[Ca2�]i (Fig. 6A), indicating that both CCL19-Fc and CCL21-Fcare functional. No mobilization of [Ca2�]i was observed in paren-tal 300-19 cells lacking CCR7, indicating that the rise in [Ca2�]i
was specific. Additionally, the chemotactic activity of CCL19-Fcand CCL21-Fc were tested in a Transwell chemotaxis assay. Asshown in Fig. 6B, 300-19 cells expressing CCR7 migrated nor-mally in response to CCL19-Fc, CCL19, and CCL21. For an un-known reason, only marginal migration toward CCL21-Fc was
FIGURE 5. Internalized CCR7 is not degraded. A, HEK293-CCR7-GFP cells were incubated for different time periods with 2 �g/ml CCL19(circles) or CCL21 (triangles) in the presence (open symbols) or absence(closed symbols) of cycloheximide. Total GFP-derived fluorescence wasmeasured by flow cytometry. B, HEK293 cells stably expressingCCR7-HA were incubated with 2 �g/ml CCL19 or CCL21 for up to 6 h.Cells were lyzed, and the total amount of CCR7-HA was determined byWestern blotting using an anti-HA Ab. The �1 proteasome subunit wasused as a loading control.
FIGURE 6. Generation of functional CCL19-Fc and CCL21-Fc. A, Pa-rental 300-19 cells (bottom panel) or cells stably expressing CCR7 wereloaded with Fluo-3/AM and stimulated with 4 �g/ml purified CCL19-Fc orCCL21-Fc and chemokine-mediated changes in intracellular free calciumconcentrations were recorded over time by flow cytometry. For compari-son, [Ca2�]i changes in response to 2 �g/ml untagged CCL19 and CCL21were measured. The arrowheads indicate the time point of chemokine ad-dition (B). The migration of 300-19 cells expressing CCR7 in response toCCL19, CCL21 (1 �g/ml), and chemokine-Fc fusion proteins (10 �g/ml)was measured in Transwell chemotaxis assays. After 3 h of incubation,cells migrated to the lower wells were collected and counted by flow cy-tometry. As a control, cell migration in the absence of chemokine wasdetermined.
2318 OPPOSITE FATE OF CCR7 AND CCL19 TRAFFICKING
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

observed at different concentrations of chemokine (Fig. 6B anddata not shown).
CCL19-Fc is internalized together with CCR7 but then sorted tolysosomes for degradation
The functional recombinant proteins enabled us to study the intra-cellular trafficking and interaction of both chemokines with CCR7.CCL19-Fc induced internalization of CCR7-GFP similar toCCL19; and CCL19-Fc colocalized with CCR7-GFP as shown byconfocal microscopy (Fig. 7A). Consistent with CCL21,CCL21-Fc also induced some internalization of CCR7 (data notshown). Remarkably, after incubation of HEK293 cells expressingCCR7-GFP with CCL19-Fc for 30 min at 37°C, followed by wash-ing off the chemokine and an additional incubation for 6 h in the
absence of the chemokine, most of the CCL19-Fc staining disap-peared and vaguely colocalized with CCR7-GFP, which recycledback to the plasma membrane (Fig. 7B). Similar results were ob-tained with monobiotinylated CCL19 (data not shown). To addressthe trafficking of endocytosed chemokine, we stimulated CCR7-transfected HEK293 cells with CCL19-Fc together with AlexaFluor 546-labeled transferrin. After 30 min of incubation,CCL19-Fc partially colocalized with transferrin (Fig. 7C), indicat-ing that CCL19-Fc localizes in early endosomes like CCR7. How-ever, not all intracellular CCL19-Fc spots colocalized with endo-somes. To address the origin of these additional compartments, westimulated CCR7-expressing cells with CCL19-Fc for 8 h in thepresence of lysotracker. As depicted in Fig. 7D, CCL19-Fc alsopartially colocalized with lysosomes. To prove that CCL19-Fc is
FIGURE 7. CCL19-Fc is internalizedtogether with CCR7 and colocalizes withboth transferrin and lysosomes. A,HEK293 cells expressing CCR7-GFPwere incubated with 10 �g/ml CCL19-Fcfor 30 min at 37°C. Cells were fixed andpermeabilized with Triton X-100.CCL19-Fc was visualized using a biotin-ylated anti-human IgG Ab and streptavi-din-Cy3. The localization of CCR7-GFPand CCL19-Fc was determined by con-focal microscopy. B, HEK293-CCR7-GFP cells were stimulated withCCL19-Fc for 30 min as in (A), washedextensively and cultured in the absenceof chemokines for additional 6 h. C,HEK293 cells stably transfected withVSV-CCR7 were incubated withCCL19-Fc for 30 min at 37°C in thepresence of Alexa Fluor 546-labeledtransferrin. CCL19-Fc was monitoredwith a biotinylated anti-human IgG Aband streptavidin-FITC. D, HEK293 cellsexpressing VSV-CCR7 were incubatedwith CCL19-Fc and lysotracker for 8 h.Representative images are shown. Scalebars, 10 �m. E, HEK293 cells expressingCCR7-HA were incubated with 10 �g/mlCCL19-Fc (upper panel) or 3 �g/mlmonobiotinylated CCL19 (lower panel)for 30 min at 37°C in the presence orabsence of 200 �M chloroquine. Cellswere washed, incubated for indicatedtime periods in the presence or absenceof chloroquine, washed, and lyzed, andproteins were separated by SDS-PAGE.CCL19-Fc was detected by Western blot-ting using an anti-human IgG Ab coupledto HRP. Biotinylated CCL19 was de-tected using streptavidinHRP.
2319The Journal of Immunology
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from
indeed degraded in lysosomes, we stimulated HEK293 cells ex-pressing CCR7-HA with CCL19-Fc for 30 min in the presence orabsence of chloroquine. Chemokines were removed and cells werefurther incubated for 3 and 6 h in the presence or absence of chlo-roquine. CCL19-Fc was degraded after 3 and 6 h (Fig. 7E). Treat-ment of the cells with the lysosomotrophic agent chloroquine andsubsequent incubation with CCL19-Fc significantly inhibited che-mokine degradation, providing clear evidence for CCL19-Fc deg-radation in lysosomes (Fig. 7E). To exclude that the degradationwas due to the Fc part, we repeated the experiments with a chem-ically synthesized monobiotinylated CCL19, where a single aminoacid was biotinylated. In fact, biotinylated CCL19 also was de-graded (Fig. 7E) comparable to CCL19-Fc. Chloroquine treatmentabolished the degradation of the biotinylated chemokine leading toan accumulation of CCL19 in intracellular compartments (data notshown).
CCR7 endocytosis is mediated by clathrin-coated pits
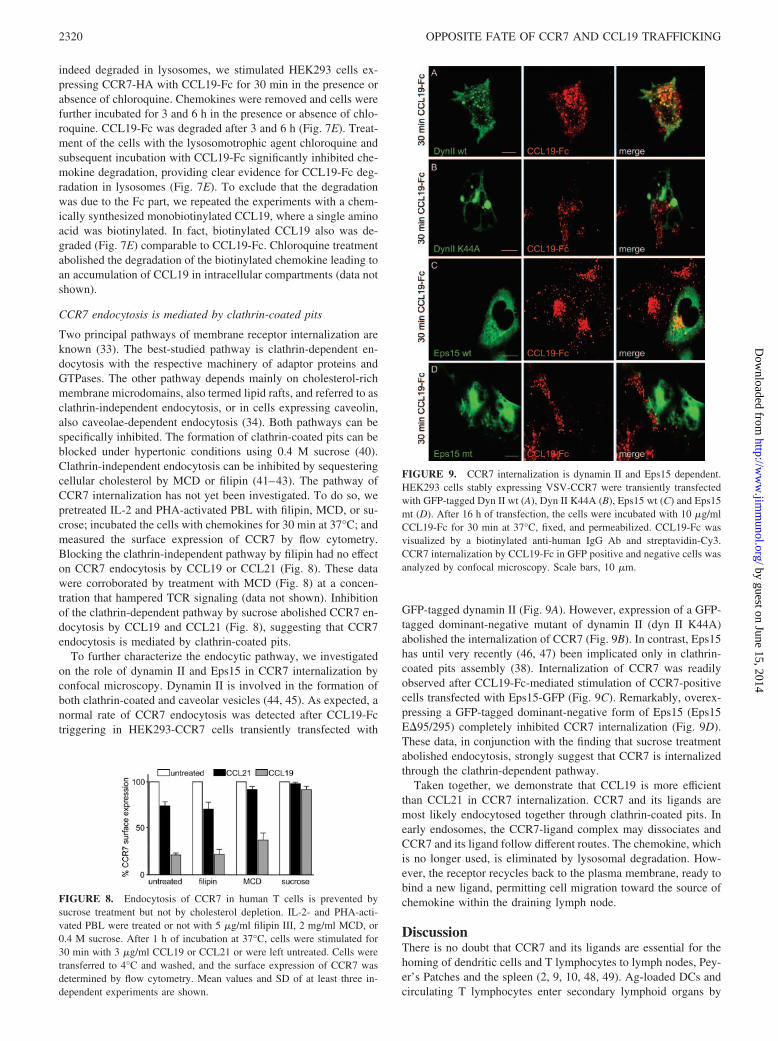
Two principal pathways of membrane receptor internalization areknown (33). The best-studied pathway is clathrin-dependent en-docytosis with the respective machinery of adaptor proteins andGTPases. The other pathway depends mainly on cholesterol-richmembrane microdomains, also termed lipid rafts, and referred to asclathrin-independent endocytosis, or in cells expressing caveolin,also caveolae-dependent endocytosis (34). Both pathways can bespecifically inhibited. The formation of clathrin-coated pits can beblocked under hypertonic conditions using 0.4 M sucrose (40).Clathrin-independent endocytosis can be inhibited by sequesteringcellular cholesterol by MCD or filipin (41–43). The pathway ofCCR7 internalization has not yet been investigated. To do so, wepretreated IL-2 and PHA-activated PBL with filipin, MCD, or su-crose; incubated the cells with chemokines for 30 min at 37°C; andmeasured the surface expression of CCR7 by flow cytometry.Blocking the clathrin-independent pathway by filipin had no effecton CCR7 endocytosis by CCL19 or CCL21 (Fig. 8). These datawere corroborated by treatment with MCD (Fig. 8) at a concen-tration that hampered TCR signaling (data not shown). Inhibitionof the clathrin-dependent pathway by sucrose abolished CCR7 en-docytosis by CCL19 and CCL21 (Fig. 8), suggesting that CCR7endocytosis is mediated by clathrin-coated pits.
To further characterize the endocytic pathway, we investigatedon the role of dynamin II and Eps15 in CCR7 internalization byconfocal microscopy. Dynamin II is involved in the formation ofboth clathrin-coated and caveolar vesicles (44, 45). As expected, anormal rate of CCR7 endocytosis was detected after CCL19-Fctriggering in HEK293-CCR7 cells transiently transfected with
GFP-tagged dynamin II (Fig. 9A). However, expression of a GFP-tagged dominant-negative mutant of dynamin II (dyn II K44A)abolished the internalization of CCR7 (Fig. 9B). In contrast, Eps15has until very recently (46, 47) been implicated only in clathrin-coated pits assembly (38). Internalization of CCR7 was readilyobserved after CCL19-Fc-mediated stimulation of CCR7-positivecells transfected with Eps15-GFP (Fig. 9C). Remarkably, overex-pressing a GFP-tagged dominant-negative form of Eps15 (Eps15E�95/295) completely inhibited CCR7 internalization (Fig. 9D).These data, in conjunction with the finding that sucrose treatmentabolished endocytosis, strongly suggest that CCR7 is internalizedthrough the clathrin-dependent pathway.
Taken together, we demonstrate that CCL19 is more efficientthan CCL21 in CCR7 internalization. CCR7 and its ligands aremost likely endocytosed together through clathrin-coated pits. Inearly endosomes, the CCR7-ligand complex may dissociates andCCR7 and its ligand follow different routes. The chemokine, whichis no longer used, is eliminated by lysosomal degradation. How-ever, the receptor recycles back to the plasma membrane, ready tobind a new ligand, permitting cell migration toward the source ofchemokine within the draining lymph node.
DiscussionThere is no doubt that CCR7 and its ligands are essential for thehoming of dendritic cells and T lymphocytes to lymph nodes, Pey-er’s Patches and the spleen (2, 9, 10, 48, 49). Ag-loaded DCs andcirculating T lymphocytes enter secondary lymphoid organs by
FIGURE 9. CCR7 internalization is dynamin II and Eps15 dependent.HEK293 cells stably expressing VSV-CCR7 were transiently transfectedwith GFP-tagged Dyn II wt (A), Dyn II K44A (B), Eps15 wt (C) and Eps15mt (D). After 16 h of transfection, the cells were incubated with 10 �g/mlCCL19-Fc for 30 min at 37°C, fixed, and permeabilized. CCL19-Fc wasvisualized by a biotinylated anti-human IgG Ab and streptavidin-Cy3.CCR7 internalization by CCL19-Fc in GFP positive and negative cells wasanalyzed by confocal microscopy. Scale bars, 10 �m.
FIGURE 8. Endocytosis of CCR7 in human T cells is prevented bysucrose treatment but not by cholesterol depletion. IL-2- and PHA-acti-vated PBL were treated or not with 5 �g/ml filipin III, 2 mg/ml MCD, or0.4 M sucrose. After 1 h of incubation at 37°C, cells were stimulated for30 min with 3 �g/ml CCL19 or CCL21 or were left untreated. Cells weretransferred to 4°C and washed, and the surface expression of CCR7 wasdetermined by flow cytometry. Mean values and SD of at least three in-dependent experiments are shown.
2320 OPPOSITE FATE OF CCR7 AND CCL19 TRAFFICKING
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

sensing CCL21 presented on HEV. Thereafter, they migrate to theT zone where CCL19, as well as CCL21, are expressed, facilitatingthe contact between naive T lymphocytes and Ag-loaded DCs and,hence, the priming of T cells. For this event, fine-tuning of cellmigration may be critical.
One important way of modulating chemokine receptor respon-siveness is receptor endocytosis after ligand binding. Studies onchemokine receptor endocytosis moved into focus because che-mokine-induced internalization of cell surface receptors was a ma-jor defense mechanism of chemokine-mediated inhibition of HIVinfection. Thus, endocytosis has been studied most intensively onthe HIV coreceptors CCR5 and CXCR4. CXCR4 internalization isinduced by its ligand CXCL12, but also by phorbol esters (50, 51).CXCL12-mediated endocytosis occurs via clathrin-coated pits anddepends on Rab5 and Eps15 (26). After ligand binding, CXCR4 ismonoubiquitylated, endocytosed, and subsequently sorted to lyso-somes for degradation (27). Interestingly, receptor mutants that arenot ubiquitylated internalize normally (27), but CXCR4 ubiquity-lation by AIP4 is required for sorting to lysosomes and its degra-dation (28). However, CXCR4 also was shown to recycle back tothe plasma membrane (26, 50). In contrast, CCR5 internalizationdoes not occur by phorbol esters (52), but only by its ligands (26).However, data on the routes of CCR5 internalization are contro-versial. CCR5 endocytosis was shown to be clathrin dependent(29, 53). Furthermore, intracellular CCR5 colocalized with fluo-rescent-labeled transferrin (54) and �-arrestin (55, 56). In contrast,cholesterol depletion by nystatin and filipin affected CCR5 endo-cytosis, and CCR5 was found to colocalize with caveolin, suggest-ing a role of caveolae/lipid rafts in this process (26, 55). Althoughdifferent routes of CCR5 endocytosis have been described, there isconsensus that CCR5 is recycled back to the cell surface (29, 54, 55).
The mechanism of CCR7 endocytos is poorly investigated. Inthis study, we demonstrate that CCR7 in PBL is rapidly internal-ized after binding of CCL19 and to a lesser extent also by itssecond ligand CCL21 (Fig. 1). This is intriguing because CCR7 isthe only chemokine receptor that is able to turn on different sig-naling pathways depending on different ligands. Our data are es-sentially in agreement with previous findings by Bardi et al. (32),who observed CCL19- but not CCL21-mediated endocytosis ofCCR7 in activated lymphocytes . However, they also found bor-derline internalization of CCR7 by CCL21 in naive T cells, den-dritic cells, and transfectants (32). The differential behavior ofCCL19 and CCL21 is striking because both chemokines have sim-ilar binding affinities and induce G protein activation, calcium mo-bilization, and chemotaxis with equal potency (4, 18). To furthercharacterize the trafficking of CCR7, we generated a CCR7-GFPfusion protein that fulfills all functional properties of wt CCR7(Fig. 2). We provide evidence that CCR7 is internalized by clath-rin-coated pits, because overexpression of dominant-negative mu-tants of dynamin II and Eps15 blocked CCL19-induced endocy-tosis of CCR7 (Fig. 9). For a long time, Eps15 has been describedto be specific for clathrin-dependent endocytosis (33, 38). How-ever, it has been shown recently that Eps15 also may be involvedin clathrin-independent endocytosis (46, 47). As hypertonic su-crose treatment also abolished CCR7 internalization (Fig. 8) is itreasonable to assume that CCR7 uses clathrin-coated pits for en-tering the cells. After endocytosis, clathrin-coted vesicles contain-ing CCR7 fuse with early endosomes, as shown by colocalizationwith transferrin (Fig. 3). Endosomes are the key control organellesfor sorting where the decision is shaped whether receptors aredirected to late endosomes and lysosomes for degradation or arerecycled back to the plasma membrane (33). Internalized CCR7followed the route of recycling back to the plasma membrane (Fig.4 and Ref. 32), and recycled CCR7 was able to mediate chemo-
taxis and flux calcium in response to CCL19. Nevertheless, wealso investigated whether CCR7 is degraded. However, we neitherfound colocalization with lysosomes nor evidence for degradationas measured by the fluorescence intensity of CCR7-GFP over timeand by assessing the protein levels over time by Western blotting(Figs. 3 and 5). Thus, in contrast with CXCR4, which is rapidlydegraded and has a half-life of �3 h (27), CCR7 has a very longhalf-life and recycles (Fig. 5). This finding is important because,for example, dendritic cells that are infected in the periphery witha virus need to migrate over a significant distance after sensingCCR7 ligands for the first time. Viral infection of dendritic cells isoften accompanied by an inhibition of translation (57). Conse-quently, if CCR7 would be degraded after endocytosis-likeCXCR4, infected dendritic cells lacking surface expression ofCCR7 would be insensitive to CCL19/CCL21 and would nevermake it into the lymph node. Under these conditions, an efficientpriming of lymphocytes would not occur because the Ags wouldremain in the periphery.
The fate of chemokines after receptor internalization has re-mained enigmatic. More than a decade ago, even before the firstCC chemokine receptor was cloned, Wang et al. (58) identified thefirst hint of chemokine degradation after endocytosis. Internalized125I-labeled CCL2 was progressively released into the culture su-pernatant of monocytes in a degraded form. CCL2 degradation wasinhibited by ammonium chloride, implicating lysosomal degrada-tion (58). Later, it has been postulated, based on indirect evidence,that CCR5, one of the receptors for CCL2, is recycled back to theplasma membrane together with APO-CCL5, but not with CCL5(29). In contrast, iodinated CCL3L1 was shown to be slowly de-graded in an ammonium chloride-dependent manner after internal-ization via CCR5 (59). Interestingly, the membrane-anchored che-mokine CX3CL1 is expressed at two different locations within thecell, diffuse at the plasma membrane and punctuated in jux-tanuclear compartments, and continuously cycles between the cellsurface and the endomembrane storage compartment in a SNARE-dependent manner (60). The situation for the putative chemokinereceptor D6 that scavenges a large variety of inflammatory CCchemokines is clearer. Internalized 125I-labeled CCL2 andCCL3L1 by D6 were rapidly degraded in an ammonium chloride-dependent manner (59, 61). Noteworthy, CCL19 is not scavengedby D6 (62), and CCL21 scavenging was not addressed; thus, in-formation about internalized CCR7 ligands is missing. Using aCCL19-Fc fusion protein, we could, for the first time, investigatethe trafficking of a CCR7 ligand. CCL19 is internalized togetherwith CCR7 and localized in early endosomes (Fig. 7). There,CCL19 dissociated from the receptor, which was recycled, andwas sorted to lysosomes for degradation as shown by confocalmicroscopy (Fig. 7). Degradation of CCL19 by lysosomes wasfurther supported by the fact that chloroquine treatment abolishedCCL19 degradation as assessed by Western blotting (Fig. 7).
Taken together, we provide strong evidence that CCR7 afterCCL19 triggering is internalized via clathrin-coated pits and istransported to early endosomes followed by its recycling back tothe plasma membrane where it can participate again in chemokinegradient sensing. In contrast, CCL19 dissociates from the endocy-tosed receptor, presumably in early endosomes, and is sorted tolysosomes for degradation. This may be of fundamental impor-tance for inducing an efficient and potent immune response, be-cause CCR7 is critical for the homing of lymphocytes and den-dritic cells to secondary lymphoid organs. The fact that CCR7 isrecycled, rather than degraded, may be essential for virally in-fected dendritic cells to maintain the capacity to sense the chemo-kine gradient until the cells reach their final destination, even if
2321The Journal of Immunology
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

viral infection inhibits translation and, hence, neosynthesisof CCR7.
AcknowledgmentsWe are grateful to Drs. Urs Greber, Osamu Yoshie, Pascal Schneider, andMargot Thome for plasmids, and Matthias Langhorst for assistance in im-age processing.
DisclosuresThe authors have no financial conflict of interest.
References1. Moser, B., M. Wolf, A. Walz, and P. Loetscher. 2004. Chemokines: multiple
levels of leukocyte migration control. Trends Immunol. 25: 75–84.2. Campbell, D. J., C. H. Kim, and E. C. Butcher. 2003. Chemokines in the systemic
organization of immunity. Immunol. Rev. 195: 58–71.3. Muller, G., U. E. Hopken, and M. Lipp. 2003. The impact of CCR7 and CXCR5
on lymphoid organ development and systemic immunity. Immunol. Rev. 195:117–135.
4. Willimann, K., D. F. Legler, M. Loetscher, R. S. Roos, M. B. Delgado,I. Clark-Lewis, M. Baggiolini, and B. Moser. 1998. The chemokine SLC is ex-pressed in T cell areas of lymph nodes and mucosal lymphoid tissues and attractsactivated T cells via CCR7. Eur. J. Immunol. 28: 2025–2034.
5. Sallusto, F., D. Lenig, R. Forster, M. Lipp, and A. Lanzavecchia. 1999. Twosubsets of memory T lymphocytes with distinct homing potentials and effectorfunctions. Nature 401: 708–712.
6. Sallusto, F., P. Schaerli, P. Loetscher, C. Schaniel, D. Lenig, C. R. Mackay,S. Qin, and A. Lanzavecchia. 1998. Rapid and coordinated switch in chemokinereceptor expression during dendritic cell maturation. Eur. J. Immunol. 28:2760–2769.
7. Sozzani, S., P. Allavena, G. D’Amico, W. Luini, G. Bianchi, M. Kataura, T. Imai,O. Yoshie, R. Bonecchi, and A. Mantovani. 1998. Differential regulation of che-mokine receptors during dendritic cell maturation: a model for their traffickingproperties. J. Immunol. 161: 1083–1086.
8. Gunn, M. D. 2003. Chemokine mediated control of dendritic cell migration andfunction. Semin. Immunol. 15: 271–276.
9. Forster, R., A. Schubel, D. Breitfeld, E. Kremmer, I. Renner-Muller, E. Wolf, andM. Lipp. 1999. CCR7 coordinates the primary immune response by establishingfunctional microenvironments in secondary lymphoid organs. Cell 99: 23–33.
10. Ohl, L., M. Mohaupt, N. Czeloth, G. Hintzen, Z. Kiafard, J. Zwirner,T. Blankenstein, G. Henning, and R. Forster. 2004. CCR7 governs skin dendriticcell migration under inflammatory and steady-state conditions. Immunity 21:279–288.
11. Gunn, M. D., S. Kyuwa, C. Tam, T. Kakiuchi, A. Matsuzawa, L. T. Williams, andH. Nakano. 1999. Mice lacking expression of secondary lymphoid organ che-mokine have defects in lymphocyte homing and dendritic cell localization.J. Exp. Med. 189: 451–460.
12. Luther, S. A., H. L. Tang, P. L. Hyman, A. G. Farr, and J. G. Cyster. 2000.Coexpression of the chemokines ELC and SLC by T zone stromal cells anddeletion of the ELC gene in the plt/plt mouse. Proc. Natl. Acad. Sci. USA 97:12694–12699.
13. Vassileva, G., H. Soto, A. Zlotnik, H. Nakano, T. Kakiuchi, J. A. Hedrick, andS. A. Lira. 1999. The reduced expression of 6Ckine in the plt mouse results fromthe deletion of one of two 6Ckine genes. J. Exp. Med. 190: 1183–1188.
14. Nakano, H., and M. D. Gunn. 2001. Gene duplications at the chemokine locus onmouse chromosome 4: multiple strain-specific haplotypes and the deletion ofsecondary lymphoid-organ chemokine and EBI-1 ligand chemokine genes in theplt mutation. J. Immunol. 166: 361–369.
15. Scandella, E., Y. Men, D. F. Legler, S. Gillessen, L. Prikler, B. Ludewig, andM. Groettrup. 2004. CCL19/CCL21-triggered signal transduction and migrationof dendritic cells requires prostaglandin E2. Blood 103: 1595–1601.
16. Tilton, B., L. Ho, E. Oberlin, P. Loetscher, F. Baleux, I. Clark-Lewis, andM. Thelen. 2000. Signal transduction by CXC chemokine receptor 4: stromalcell-derived factor 1 stimulates prolonged protein kinase B and extracellular sig-nal-regulated kinase 2 activation in T lymphocytes. J. Exp. Med. 192: 313–324.
17. Bardi, G., V. Niggli, and P. Loetscher. 2003. Rho kinase is required for CCR7-mediated polarization and chemotaxis of T lymphocytes. FEBS Lett. 542: 79–83.
18. Kohout, T. A., S. L. Nicholas, S. J. Perry, G. Reinhart, S. Junger, andR. S. Struthers. 2004. Differential desensitization, receptor phosphorylation, �-ar-restin recruitment, and ERK1/2 activation by the two endogenous ligands for theCC chemokine receptor 7. J. Biol. Chem. 279: 23214–23222.
19. Riol-Blanco, L., N. Sanchez-Sanchez, A. Torres, A. Tejedor, S. Narumiya,A. L. Corbi, P. Sanchez-Mateos, and J. L. Rodriguez-Fernandez. 2005. The che-mokine receptor CCR7 activates in dendritic cells two signaling modules thatindependently regulate chemotaxis and migratory speed. J. Immunol. 174:4070–4080.
20. Sanchez-Sanchez, N., L. Riol-Blanco, G. De La Rosa, A. Puig-Kroger,J. Garcia-Bordas, D. Martin, N. Longo, A. Cuadrado, C. Cabanas, A. L. Corbi, etal. 2004. Chemokine receptor CCR7 induces intracellular signaling that inhibitsapoptosis of mature dendritic cells. Blood 104: 619–625.
21. Carlsen, H. S., G. Haraldsen, P. Brandtzaeg, and E. S. Baekkevold. 2005. Dis-parate lymphoid chemokine expression in mice and men: no evidence of CCL21synthesis by human high endothelial venules. Blood 106: 444–446.
22. Gunn, M. D., K. Tangemann, C. Tam, J. G. Cyster, S. D. Rosen, andL. T. Williams. 1998. A chemokine expressed in lymphoid high endothelialvenules promotes the adhesion and chemotaxis of naive T lymphocytes. Proc.Natl. Acad. Sci. USA 95: 258–263.
23. Warnock, R. A., J. J. Campbell, M. E. Dorf, A. Matsuzawa, L. M. McEvoy, andE. C. Butcher. 2000. The role of chemokines in the microenvironmental controlof T versus B cell arrest in Peyer’s patch high endothelial venules. J. Exp. Med.191: 77–88.
24. Stein, J. V., A. Rot, Y. Luo, M. Narasimhaswamy, H. Nakano, M. D. Gunn,A. Matsuzawa, E. J. Quackenbush, M. E. Dorf, and U. H. von Andrian. 2000. TheCC chemokine thymus-derived chemotactic agent 4 (TCA-4, secondary lym-phoid tissue chemokine, 6Ckine, exodus-2) triggers lymphocyte function-associ-ated antigen 1-mediated arrest of rolling T lymphocytes in peripheral lymph nodehigh endothelial venules. J. Exp. Med. 191: 61–76.
25. Okada, T., M. J. Miller, I. Parker, M. F. Krummel, M. Neighbors, S. B. Hartley,A. O’Garra, M. D. Cahalan, and J. G. Cyster. 2005. Antigen-engaged B cellsundergo chemotaxis toward the T zone and form motile conjugates with helper Tcells. PLoS Biol. 3:e150.
26. Venkatesan, S., J. J. Rose, R. Lodge, P. M. Murphy, and J. F. Foley. 2003.Distinct mechanisms of agonist-induced endocytosis for human chemokine re-ceptors CCR5 and CXCR4. Mol. Biol. Cell 14: 3305–3324.
27. Marchese, A., and J. L. Benovic. 2001. Agonist-promoted ubiquitination of the Gprotein-coupled receptor CXCR4 mediates lysosomal sorting. J. Biol. Chem. 276:45509–45512.
28. Marchese, A., C. Raiborg, F. Santini, J. H. Keen, H. Stenmark, and J. L. Benovic.2003. The E3 ubiquitin ligase AIP4 mediates ubiquitination and sorting of the Gprotein-coupled receptor CXCR4. Dev. Cell 5: 709–722.
29. Signoret, N., A. Pelchen-Matthews, M. Mack, A. E. Proudfoot, and M. Marsh.2000. Endocytosis and recycling of the HIV coreceptor CCR5. J. Cell Biol. 151:1281–1294.
30. Signoret, N., T. Christophe, M. Oppermann, and M. Marsh. 2004. pH-indepen-dent endocytic cycling of the chemokine receptor CCR5. Traffic 5: 529–543.
31. Mueller, A., and P. G. Strange. 2004. Mechanisms of internalization and recy-cling of the chemokine receptor, CCR5. Eur. J. Biochem. 271: 243–252.
32. Bardi, G., M. Lipp, M. Baggiolini, and P. Loetscher. 2001. The T cell chemokinereceptor CCR7 is internalized on stimulation with ELC, but not with SLC. Eur.J. Immunol. 31: 3291–3297.
33. Le Roy, C., and J. L. Wrana. 2005. Clathrin- and non-clathrin-mediated endocyticregulation of cell signalling. Nat. Rev. Mol. Cell Biol. 6: 112–126.
34. Pfeffer, S. 2003. Membrane domains in the secretory and endocytic pathways.Cell 112: 507–517.
35. Aguilar, R. C., and B. Wendland. 2005. Endocytosis of membrane receptors: twopathways are better than one. Proc. Natl. Acad. Sci. USA 102: 2679–2680.
36. Legler, D. F., M. Loetscher, R. S. Roos, I. Clark-Lewis, M. Baggiolini, andB. Moser. 1998. B cell-attracting chemokine 1, a human CXC chemokine ex-pressed in lymphoid tissues, selectively attracts B lymphocytes via BLR1/CXCR5. J. Exp. Med. 187: 655–660.
37. Cao, H., H. M. Thompson, E. W. Krueger, and M. A. McNiven. 2000. Disruptionof Golgi structure and function in mammalian cells expressing a mutant dynamin.J. Cell Sci. 113 (Pt 11): 1993–2002.
38. Benmerah, A., M. Bayrou, N. Cerf-Bensussan, and A. Dautry-Varsat. 1999. In-hibition of clathrin-coated pit assembly by an Eps15 mutant. J. Cell Sci. 112 (Pt9): 1303–1311.
39. Johnson, A. O., R. N. Ghosh, K. W. Dunn, R. Garippa, J. Park, S. Mayor,F. R. Maxfield, and T. E. McGraw. 1996. Transferrin receptor containing theSDYQRL motif of TGN38 causes a reorganization of the recycling compartmentbut is not targeted to the TGN. J. Cell Biol. 135: 1749–1762.
40. Heuser, J. E., and R. G. Anderson. 1989. Hypertonic media inhibit receptor-mediated endocytosis by blocking clathrin-coated pit formation. J. Cell Biol. 108:389–400.
41. Harder, T., R. Kellner, R. G. Parton, and J. Gruenberg. 1997. Specific release ofmembrane-bound annexin II and cortical cytoskeletal elements by sequestrationof membrane cholesterol. Mol. Biol. Cell 8: 533–545.
42. Keller, P., and K. Simons. 1998. Cholesterol is required for surface transport ofinfluenza virus hemagglutinin. J. Cell Biol. 140: 1357–1367.
43. Orlandi, P. A., and P. H. Fishman. 1998. Filipin-dependent inhibition of choleratoxin: evidence for toxin internalization and activation through caveolae-like do-mains. J. Cell Biol. 141: 905–915.
44. Oh, P., D. P. McIntosh, and J. E. Schnitzer. 1998. Dynamin at the neck of caveo-lae mediates their budding to form transport vesicles by GTP-driven fission fromthe plasma membrane of endothelium. J. Cell Biol. 141: 101–114.
45. De Camilli, P., K. Takei, and P. S. McPherson. 1995. The function of dynaminin endocytosis. Curr. Opin. Neurobiol. 5: 559–565.
46. Chen, H., and P. De Camilli. 2005. The association of epsin with ubiquitinatedcargo along the endocytic pathway is negatively regulated by its interaction withclathrin. Proc. Natl. Acad. Sci. USA 102: 2766–2771.
47. Sigismund, S., T. Woelk, C. Puri, E. Maspero, C. Tacchetti, P. Transidico,P. P. Di Fiore, and S. Polo. 2005. Clathrin-independent endocytosis of ubiquiti-nated cargos. Proc. Natl. Acad. Sci. USA 102: 2760–2765.
48. Moser, B., and P. Loetscher. 2001. Lymphocyte traffic control by chemokines.Nat. Immunol. 2: 123–128.
49. Cyster, J. G. 1999. Chemokines and cell migration in secondary lymphoid organs.Science 286: 2098–2102.
50. Amara, A., S. L. Gall, O. Schwartz, J. Salamero, M. Montes, P. Loetscher,M. Baggiolini, J. L. Virelizier, and F. Arenzana-Seisdedos. 1997. HIV coreceptordownregulation as antiviral principle: SDF-1�-dependent internalization of the
2322 OPPOSITE FATE OF CCR7 AND CCL19 TRAFFICKING
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from

chemokine receptor CXCR4 contributes to inhibition of HIV replication. J. Exp.Med. 186: 139–146.
51. Signoret, N., J. Oldridge, A. Pelchen-Matthews, P. J. Klasse, T. Tran, L. F. Brass,M. M. Rosenkilde, T. W. Schwartz, W. Holmes, W. Dallas, et al. 1997. Phorbolesters and SDF-1 induce rapid endocytosis and down modulation of the chemo-kine receptor CXCR4. J. Cell Biol. 139: 651–664.
52. Signoret, N., M. M. Rosenkilde, P. J. Klasse, T. W. Schwartz, M. H. Malim,J. A. Hoxie, and M. Marsh. 1998. Differential regulation of CXCR4 and CCR5endocytosis. J. Cell Sci. 111 (Pt 18): 2819–2830.
53. Signoret, N., L. Hewlett, S. Wavre, A. Pelchen-Matthews, M. Oppermann, andM. Marsh. 2005. Agonist-induced endocytosis of CC chemokine receptor 5 isclathrin dependent. Mol. Biol. Cell 16: 902–917.
54. Mack, M., B. Luckow, P. J. Nelson, J. Cihak, G. Simmons, P. R. Clapham,N. Signoret, M. Marsh, M. Stangassinger, F. Borlat, et al. 1998. Aminooxypen-tane-RANTES induces CCR5 internalization but inhibits recycling: a novel in-hibitory mechanism of HIV infectivity. J. Exp. Med. 187: 1215–1224.
55. Mueller, A., E. Kelly, and P. G. Strange. 2002. Pathways for internalization andrecycling of the chemokine receptor CCR5. Blood 99: 785–791.
56. Kraft, K., H. Olbrich, I. Majoul, M. Mack, A. Proudfoot, and M. Oppermann.2001. Characterization of sequence determinants within the carboxyl-terminaldomain of chemokine receptor CCR5 that regulate signaling and receptor inter-nalization. J. Biol. Chem. 276: 34408–34418.
57. Freigang, S., H. C. Probst, and M. van den Broek. 2005. DC infection promotesantiviral CTL priming: the “Winkelried” strategy. Trends Immunol. 26: 13–18.
58. Wang, J. M., A. Hishinuma, J. J. Oppenheim, and K. Matsushima. 1993. Studiesof binding and internalization of human recombinant monocyte chemotactic andactivating factor (MCAF) by monocytic cells. Cytokine 5: 264–275.
59. Weber, M., E. Blair, C. V. Simpson, M. O’Hara, P. E. Blackburn, A. Rot,G. J. Graham, and R. J. Nibbs. 2004. The chemokine receptor D6 constitutivelytraffics to and from the cell surface to internalize and degrade chemokines. Mol.Biol. Cell 15: 2492–2508.
60. Liu, G. Y., V. Kulasingam, R. T. Alexander, N. Touret, A. M. Fong, D. D. Patel,and L. A. Robinson. 2005. Recycling of the membrane-anchored chemokine,CX3CL1. J. Biol. Chem. 280: 19858–19866.
61. Fra, A. M., M. Locati, K. Otero, M. Sironi, P. Signorelli, M. L. Massardi,M. Gobbi, A. Vecchi, S. Sozzani, and A. Mantovani. 2003. Cutting edge: scav-enging of inflammatory CC chemokines by the promiscuous putatively silentchemokine receptor D6. J. Immunol. 170: 2279–2282.
62. Bonecchi, R., M. Locati, E. Galliera, M. Vulcano, M. Sironi, A. M. Fra,M. Gobbi, A. Vecchi, S. Sozzani, B. Haribabu, et al. 2004. Differential recogni-tion and scavenging of native and truncated macrophage-derived chemokine(macrophage-derived chemokine/CC chemokine ligand 22) by the D6 decoy re-ceptor. J. Immunol. 172: 4972–4976.
2323The Journal of Immunology
by guest on June 15, 2014http://w
ww
.jimm
unol.org/D
ownloaded from




















