
Apoptosis and Molecular Pathways in the Seminiferous Epithelium of Aged and Photoinhibited Syrian...
13
Apoptosis and Molecular Pathways in the Seminiferous Epithelium of Aged and Photoinhibited Syrian Hamsters (Mesocricetus auratus) EVA MORALES,* CONCEPCION FERRER,* ADELINA ZUASTI,* JOSE C. GARCIA-BORRON,{ MANUEL CANTERAS,{ AND LUIS M. PASTOR* From the Departments of *Cell Biology (Aging Institute), ÀBiochemistry and Molecular Biology, and `Statistics, Faculty of Medicine, University of Murcia, Murcia, Spain. ABSTRACT: Aging and short photoperiod exposure induce germ cell apoptosis in the Syrian hamster; however, the specific germ cells affected and the molecular pathways triggered have not been elucidated. We analyzed germ cell apoptosis and the expression of the Fas/Fas-L system, Bcl-2 family, and p53 in aged and photoinhibited hamsters and compared with those young maintained in natural photoperiod. Aging increased apoptosis in spermatogonia and spermatocytes, but in photoinhibited hamster testes only an increase in apoptotic spermatocytes was observed. Apoptosis was higher in aged hamsters in stages I–IV, V–VI and VII–VIII. Aging increased apoptosis of spermatogonia in stages I–IV and V–VI. Apoptotic pachytene spermatocytes were significantly higher in stages I–IV, V–VI, and VII–VIII in aging. Apoptotic preleptotene and pachytene spermatocytes were higher in aging, but no differences were observed in leptotene-zygotene. Fas-L was expressed by Sertoli cells, of young, aged, and photoinhibited hamsters. Bcl-x L was strongly expressed in germ cells on young hamsters and slightly in aging and after short photoperiod exposure. Spermatocytes of photoinhibited hamsters were intensively stained with Fas, Bax, Bcl-xs/L, and p53. In conclusion, aging increases apoptosis in spermatogonia and spermatocytes, depending on the stage of the seminiferous epithelium cycle, whereas after a short photoperiod exposure only an increase in apoptotic spermatocytes is observed. The results suggest that Fas, Bcl-x L , Bax, and p53 participate in germ cell apoptosis induction after short photoperiod exposure, whereas only Bcl-x L is involved in aging. Key words: Aging, Bcl-2, Fas, photoperiod, testis. J Androl 2007;28:123–135 G erm cell death occurs via apoptosis, whether spontaneously during normal spermatogenesis or triggered by different stimuli (Blanco-Rodrı ´guez and Martı ´nez-Garcı ´a, 1996, 1998; Cai et al, 1997; Sinha Hikim et al, 1997; Lee et al, 1999). In the Syrian hamster both the short photoperiod exposure, which is charac- terized by a sharp fall in testosterone (Calvo et al, 1997), and aging, characterized by normal androgen levels (Horn et al, 1996; Calvo et al, 1999), are physiological situations of germ cell loss and atrophy of the seminiferous epithelium (Horn et al, 1996; Morales et al, 2002, 2003, 2004). Apoptosis has been involved in testicular germ cell loss during aging in numerous species, including humans (Brinkworth et al, 1997; Kimura et al, 2003) and rodents, such as the mouse (Barnes et al, 1998), rat (Wang et al, 1999; Barnes et al, 1999) and Syrian hamster (Morales et al, 2003). In aged rats germ cell apoptosis has been related to specific stages of the seminiferous epithelium cycle (Wang et al, 1999), although this aspect has not been studied in other species. Also, short photoperiod exposure induces germ cell apoptosis in white-footed mouse (Young et al, 1999, 2000) and Syrian hamster (Morales et al, 2002). However, the specific germ cell population affected has not sufficiently been documented in both models of seminiferous epithelium atrophy. Apoptosis can be triggered by 3 mechanisms: 1) the binding of Fas-L to Fas receptor expressing cells (Itoh et al, 1991; Oehm et al, 1992; Suda et al, 1993); 2) the activation of Bcl-2 family members (Yang and Kors- meyer, 1996; Green and Reed, 1998); and 3) the endo- plasmic reticulum pathway (Nakagawa et al, 2000; Bitko and Barik, 2001; Yoneda et al, 2001). Fas/Fas-L system seems to play an important role in testicular germ cell apoptosis regulation (Pertika ¨inen et al, 1999). Fas is expressed in spermatogonia, spermatocytes, and sperma- tids, all apoptotic cells (Lee et al, 1997). Fas expression Supported by grant PI-56/00866/FS/01 from the Fundacio ´n Se ´neca, Comunidad Auto ´noma de la Regio ´ n de Murcia. Correspondence to: Prof Dr Luis M. Pastor, Department of Cell Biology, Faculty of Medicine, Campus de Espinardo, University of Murcia, E-30071 Murcia, Spain (e-mail: [email protected]). Received for publication September 13, 2005; accepted for publication August 24, 2006. DOI: 10.2164/jandrol.106.000778 A portion of this report has been communicated in abstract form for the 44th American Society of Cell Biology Annual Meeting. Journal of Andrology, Vol. 28, No. 1, January/February 2007 Copyright E American Society of Andrology 123
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Apoptosis and Molecular Pathways in the Seminiferous Epithelium of Aged and Photoinhibited Syrian...

Apoptosis and Molecular Pathways in the Seminiferous Epitheliumof Aged and Photoinhibited Syrian Hamsters(Mesocricetus auratus)
EVA MORALES,* CONCEPCION FERRER,* ADELINA ZUASTI,* JOSE C. GARCIA-BORRON,{MANUEL CANTERAS,{ AND LUIS M. PASTOR*
From the Departments of *Cell Biology (Aging Institute), �Biochemistry and Molecular Biology, and `Statistics,
Faculty of Medicine, University of Murcia, Murcia, Spain.
ABSTRACT: Aging and short photoperiod exposure induce germ
cell apoptosis in the Syrian hamster; however, the specific germ cells
affected and the molecular pathways triggered have not been
elucidated. We analyzed germ cell apoptosis and the expression of
the Fas/Fas-L system, Bcl-2 family, and p53 in aged and
photoinhibited hamsters and compared with those young maintained
in natural photoperiod. Aging increased apoptosis in spermatogonia
and spermatocytes, but in photoinhibited hamster testes only an
increase in apoptotic spermatocytes was observed. Apoptosis was
higher in aged hamsters in stages I–IV, V–VI and VII–VIII. Aging
increased apoptosis of spermatogonia in stages I–IV and V–VI.
Apoptotic pachytene spermatocytes were significantly higher in
stages I–IV, V–VI, and VII–VIII in aging. Apoptotic preleptotene
and pachytene spermatocytes were higher in aging, but no
differences were observed in leptotene-zygotene. Fas-L was
expressed by Sertoli cells, of young, aged, and photoinhibited
hamsters. Bcl-xL was strongly expressed in germ cells on young
hamsters and slightly in aging and after short photoperiod exposure.
Spermatocytes of photoinhibited hamsters were intensively stained
with Fas, Bax, Bcl-xs/L, and p53. In conclusion, aging increases
apoptosis in spermatogonia and spermatocytes, depending on the
stage of the seminiferous epithelium cycle, whereas after a short
photoperiod exposure only an increase in apoptotic spermatocytes is
observed. The results suggest that Fas, Bcl-xL, Bax, and p53
participate in germ cell apoptosis induction after short photoperiod
exposure, whereas only Bcl-xL is involved in aging.
Key words: Aging, Bcl-2, Fas, photoperiod, testis.
J Androl 2007;28:123–135
G erm cell death occurs via apoptosis, whether
spontaneously during normal spermatogenesis or
triggered by different stimuli (Blanco-Rodrıguez and
Martınez-Garcıa, 1996, 1998; Cai et al, 1997; Sinha
Hikim et al, 1997; Lee et al, 1999). In the Syrian hamster
both the short photoperiod exposure, which is charac-
terized by a sharp fall in testosterone (Calvo et al, 1997),
and aging, characterized by normal androgen levels
(Horn et al, 1996; Calvo et al, 1999), are physiological
situations of germ cell loss and atrophy of the
seminiferous epithelium (Horn et al, 1996; Morales et
al, 2002, 2003, 2004).
Apoptosis has been involved in testicular germ cell
loss during aging in numerous species, including humans
(Brinkworth et al, 1997; Kimura et al, 2003) and
rodents, such as the mouse (Barnes et al, 1998), rat
(Wang et al, 1999; Barnes et al, 1999) and Syrian
hamster (Morales et al, 2003). In aged rats germ cell
apoptosis has been related to specific stages of the
seminiferous epithelium cycle (Wang et al, 1999),
although this aspect has not been studied in other
species. Also, short photoperiod exposure induces germ
cell apoptosis in white-footed mouse (Young et al, 1999,
2000) and Syrian hamster (Morales et al, 2002).
However, the specific germ cell population affected has
not sufficiently been documented in both models of
seminiferous epithelium atrophy.
Apoptosis can be triggered by 3 mechanisms: 1) the
binding of Fas-L to Fas receptor expressing cells (Itoh et
al, 1991; Oehm et al, 1992; Suda et al, 1993); 2) the
activation of Bcl-2 family members (Yang and Kors-
meyer, 1996; Green and Reed, 1998); and 3) the endo-
plasmic reticulum pathway (Nakagawa et al, 2000; Bitko
and Barik, 2001; Yoneda et al, 2001). Fas/Fas-L system
seems to play an important role in testicular germ cell
apoptosis regulation (Pertikainen et al, 1999). Fas is
expressed in spermatogonia, spermatocytes, and sperma-
tids, all apoptotic cells (Lee et al, 1997). Fas expression
Supported by grant PI-56/00866/FS/01 from the Fundacion Seneca,
Comunidad Autonoma de la Region de Murcia.
Correspondence to: Prof Dr Luis M. Pastor, Department of Cell
Biology, Faculty of Medicine, Campus de Espinardo, University of
Murcia, E-30071 Murcia, Spain (e-mail: [email protected]).
Received for publication September 13, 2005; accepted for
publication August 24, 2006.
DOI: 10.2164/jandrol.106.000778
A portion of this report has been communicated in abstract form
for the 44th American Society of Cell Biology Annual Meeting.
Journal of Andrology, Vol. 28, No. 1, January/February 2007Copyright E American Society of Andrology
123

has also been related to germ cell degeneration in arrest of
spermatogenesis (Eguchi et al, 2002; Francavilla et al,
2002). There is some discrepancy as to the preciselocalization of Fas-L in the seminiferous tubules, with
some groups showing Fas-L in the Sertoli cells (Bellgrau
et al, 1995; French et al, 1996; Lee et al, 1997; Koji, 2001),
while others have reported that Fas-L is also expressed in
the germ cells (Woolveridge et al, 1999; Francavilla et al,
2000). Bcl-2 and the long form Bcl-x (Bcl-xL) promote cell
survival by inhibiting apoptosis. However, other mem-
bers of the Bcl-2 family (Bax, Bak, Bcl-xS, and Bad)promote cell death (Oltvai et al, 1993). Members of the
Bcl-2 family also regulate spermatogenesis, inducing
apoptosis in spermatogonia, primary spermatocytes,
and spermatids (Oldereid et al, 2001) and participating
in the differentiation process (Oldereid et al, 2001). Also,
p53 has been involved in the elimination of damaged
germ cells as well as those produced in excess (Stephan et
al, 1996), as well as in the control of proliferation andapoptosis in spermatogonia (Beumer et al, 1998).
Although germ cell apoptosis was implicated in aging
and was seen to occur after short photoperiod exposure in
a previous study in our laboratory (Morales et al, 2002,
2003), the specific cell types that display apoptosis, the
relationship of apoptosis with specific stages of the
seminiferous epithelium cycle, and the molecular path-
ways involved have not been sufficiently studied. Theobjectives of the present study were: 1) to study how the
different populations of germ cells are affected by
apoptosis caused by aging and short photoperiod
exposure; 2) to analyze the induction of germ cell
apoptosis during the different stages of the seminiferous
epithelium cycle in aging Syrian hamster; and 3) to
examine the expression of some of the Bcl-2 family
members, the Fas/Fas-L system, and p53 in the seminif-erous epithelium of Syrian hamsters and to investigate the
roles of these proteins in the induction of testicular germ
cell apoptosis in aging and short photoperiod exposure.
Materials and Methods
Animals and Tissue Preparation
A total of 16 male Syrian hamsters (Mesocricetus auratus) were
used in the present study. They were distributed in 3 groups:
a first group of 5 control animals that were 6 months old;
a second group of 6 hamsters that were 24 months old; and
a third group of five 6-month-old animals that were exposed to
short photoperiod. All the animals were maintained at
a temperature of 20uC and given food and water ad libitum.
The illumination was regulated by a programmer so that the 6-
and 24-month-old groups were maintained in a natural
photoperiod of 14:10 (L:D), whereas the third group was
subjected to a short photoperiod cycle of 8:16 (L:D) for
2 months. Animals were sacrificed by an intraperitoneal
overdose of sodium pentobarbital, and both testes were
removed. Right testes were processed for Western blot
analysis, and left testes were processed for in situ detection
of apoptosis and immunohistochemistry. This study was
performed according to Spanish ethical and legal standards
regarding animal protection.
In Situ Germ Cell Apoptosis Detection and Quantification
Left testes were fixed in methacarn (methanol:chloroform:
acetic acid 6:3:1), and representative samples, which were
chosen randomly, were dehydrated, immersed in toluene, and
embedded in Paraplast Plus (Panreac Quımica SA, Barcelona,
Spain). Germ cell apoptosis was examined following the
protocol of TACS TdT in situ Apoptosis Detection kit
(TUNEL reaction; R&D Systems Inc, Minnesota, Minn).
For this, 5-mm-thick sections were deparaffinized, hydrated,
washed, and incubated with proteinase K (1 mg/mL). The
peroxidase activity was quenched with 10% H2O2. The samples
were immersed in 16 TdT labeling buffer (1 mol TACS Safe-
TdT Buffer, 0.5 mg/mL BSA, 0.6 mmol 2-mercaptoethanesul-
fonic acid) at 18u–24uC for 5 minutes and incubated for 1 hour
at 37uC with TdTdNTP Mix (0.25 mmol biotinylated dNTP),
506 Mn+2 (1 mL), TdT Enzyme (1 mL), and 16 TdT labeling
buffer. The reaction was stopped with TdT stop buffer
(0.1 mol EDTA, pH 8.0). Subsequently, the samples were
washed in PBS and incubated with streptavidin-horseradish
peroxidase for 10 minutes at 18u–24uC. After washing in PBS,
the samples were stained with TACS Blue Label and incubated
with Contrast C solution. Negative control sections, processed
without TdT, did not show positive labeling.
The apoptotic germ cells were identified according to their
position within the seminiferous epithelium, the cell size and
nuclear morphology, and stage of the seminiferous epithelium.
The quantity of apoptosis and the percentage of apoptotic
germ cells were recorded in the populations of spermatogonia
(SG) and spermatocytes (SC). For this purpose, 5-mm-thick
sections were collected every 50 mm of tissue and processed for
in situ germ cell apoptosis detection. Of the sections processed,
4 randomly chosen testis cross-sections were counted per
animal. Within each section, 25 random fields were chosen by
systematically moving the microscope lens across the tissue
section without overlap and selecting every second field and
were analyzed (1 field 5 0.018 mm2). In each field of study the
number of TUNEL-positive and negative germ cells was
scored in the populations of spermatogonia and spermato-
cytes. The following apoptotic indices of each population of
germ cells, expressed as the percentage of TUNEL-positive
cells, were calculated: 1) the total apoptotic index (% of
TUNEL-positive SG+SC); 2) the total apoptotic index in
spermatogonia (% of TUNEL-positive SG); and 3) the total
apoptotic index in primary spermatocytes (% of TUNEL-
positive primary SC).
Also, in 6- and 24-month-old hamsters the apoptotic activity
was examined during the different stages of the seminiferous
epithelium cycle. The identification of stages was based on the
description of Leblond and Clermont (1952) and Tiba et al
(1992). Similarly to previous studies (Wang et al, 1999; Sinha
Hikim et al, 2003), the stages were grouped into 4 groups:
124 Journal of Andrology N January �February 2007

I–IV, V–VI, VII–VIII, and IX–XIII. For quantification of
apoptosis during the seminiferous epithelium cycle, at least 100
randomly selected perpendicular seminiferous tubule cross
sections on 4 sections from each animal of each group were
analyzed. In each tubular section, the number of TUNEL-
positive and negative germ cells was scored in the populations
of spermatogonia, preleptotenes, pachytenes, and leptotene-
zygotene spermatocytes. The apoptotic index of each popula-
tion of germ cells during the different stages of the
seminiferous epithelium cycle was expressed as the percentage
of TUNEL-positive cells.
Immunohistochemistry
Five-mm-thick sections fixed in methacarn and embedded in
paraffin were deparaffinized in xylene, hydrated, and trans-
ferred to PBS for 10 minutes. Endogenous peroxidase was
blocked with 1% H2O2 in PBS for 30 minutes. After washing
in PBS, samples were blocked with 1.5% normal rabbit or goat
serum (Jackson ImmunoResearch, West Grove, Pa), depend-
ing on the origin of the secondary antibody. Subsequently, the
samples were washed in PBS and incubated overnight at 4uC,
with the primary antibody diluted in PBS/BSA. The primary
antibodies were purchased from Santa Cruz Biotechnology
(Santa Cruz, Calif), and the concentrations used were the
following: anti-Fas (1:30) (sc-7886/ FL-335), anti-Fas-L (1:50)
(sc-834/N-20), anti-Bcl-xL (1:100) (sc-8392/H-5), anti-Bcl-xS/L
(1:100) (sc-1041/L19), anti-Bcl-2 (1:100) (sc-492/N-19), anti-
Bax (1:100) (sc-526-P19), and anti-p53 (1:20) (sc-6243/FL-393).
The samples were washed in PBS and then incubated for
45 minutes with the corresponding biotinylated secondary
antibody (CHEMICON International, Temecula, Calif) di-
luted 1:500 in PBS/BSA. Sections were again washed in PBS
and subsequently incubated for 45 minutes with HRP-strepta-
vidin (Dako, Glostrup, Denmark) diluted 1:300 in PBS/BSA.
Samples were washed again in PBS; bound antibody was
visualized after the addition of 0.05% solution of 3,39-di-
aminobenzidine tetrachloride in TBS, to which 0.03% H2O2
was added. The slides were subsequently counterstained with
hematoxylin. Control sections, in which the primary antibody
was replaced by PBS, were similarly processed. For all the anti-
bodies analyzed, no immunostaining was detected in control
sections from which the primary antibody had been omitted.
Western Blot Analysis
The right testis was snap frozen in liquid nitrogen and stored at
270uC until required for protein extraction. Tissue was
homogenized in 1% SDS in PBS and a tablet of protease
inhibitors Complete Mini, EDTA-free (Roche, Mannheim,
Germany) for each 10 mL of homogenization solution
(Pentikainen et al, 1999). One mL of homogenization solution
was used for each 100 mg of fresh tissue. After centrifugation
at 17 000 6 g for 30 minutes at 4uC, the supernatants were
collected and their protein concentrations determined by BCA
method (Pierce, Rockford, Ill). The equivalent amounts of
protein were mixed with sample buffer (0.5 mol Tris-HCl
pH 6.8, 20% glycerol, 10% SDS, 0.1% bromophenol blue, and
0.5% ß-mercaptoethanol) and incubated at 95uC for 5 min-
utes. Proteins (10–15 mg) were loaded in 8%–12% SDS-
polyacrylamide gels, and electrophoresis was performed in
the presence of marker standards with molecular weights
between 200 and 6.5 kd (Bio-Rad Laboratories, Hercules,
Calif) at 15 mA during stacking and 25 mA during the
separation. The proteins were transferred to Immobilon-P
membranes (Millipore, Bedford, Mass) by semidry electro-
phoretic transfer for 1 hour at room temperature in transfer
buffer (50 mmol Tris, 40 mmol glycine, 10% SDS and 20%
methanol, pH 9.2) at 22 V. Subsequently, the membrane was
blocked with 5% nonfat milk in PBS for at least 1 hour at
room temperature. Before blocking, the marker standards
were separated and labeled with Amido Black solution (0.1%
Amido Black, 45% methanol, and 10% acetic acid) to check
the protein molecular weight. After 3 washes of 10 minutes in
PBS containing 0.1% Tween 20 (PBST), the membrane was
incubated overnight at 4uC using the same primary antibodies
used for immunohistochemistry diluted 1:1000 in a solution
containing 5% albumin in PBST. After 3 washes of 10 minutes
in PBST, the membranes were incubated for 1 hour at room
temperature with the same biotinylated secondary antibodies
used for immunohistochemistry diluted 1:10 000 PBST. Sub-
sequently, the membranes were washed with PBST and
incubated with HRP-streptavidin (Dako) diluted 1:5000.
Immunoreactive bands were located with the enhanced
chemiluminescence detection Kit (Amersham Biosciences,
Buckinghamshire, United Kingdom) and Hyperfilm ECL
(Amersham Biosciences). The semiquantitative study was
performed for each antibody with an automatic image
analyzer (MIP version 4.5; Consulting Image Digital, Barce-
lona, Spain). After digitalization and inversion of grey level
image, the bands were delimited out manually, and the area
and medium grey were calculated for each band in a range of
0–255. Labeling density was obtained by multiplying the area
times medium grey and was taken as an index of band
intensity. The densitometrical analysis is present as a bar
diagram with arbitrary unit (the densitometrical minor value
was the unit in each bar diagram).
Statistical Analysis
The mean values obtained in aged and photoinhibited
hamsters were compared with data obtained in young
hamsters (control group) using Student’s t test and, for
measurements lacking equal variance, by a Welch test. The
tests were performed using log-transformed data. Data were
back-transformed following analyses and are presented as
means 6 SEM. Statistical evaluation of apoptotic indices
during the different stages of the seminiferous epithelium was
performed by analysis of variance in conjunction with a least
significant difference (LSD) test. Mean differences were
considered statistically significant when P is less than .05.
Results
In Situ Detection of Apoptosis
In young and aged hamsters the germ cells undergoing
apoptosis were predominantly spermatogonia and sper-
Morales et al N Apoptotic Pathways in Aged and Photoinhibited Testes 125
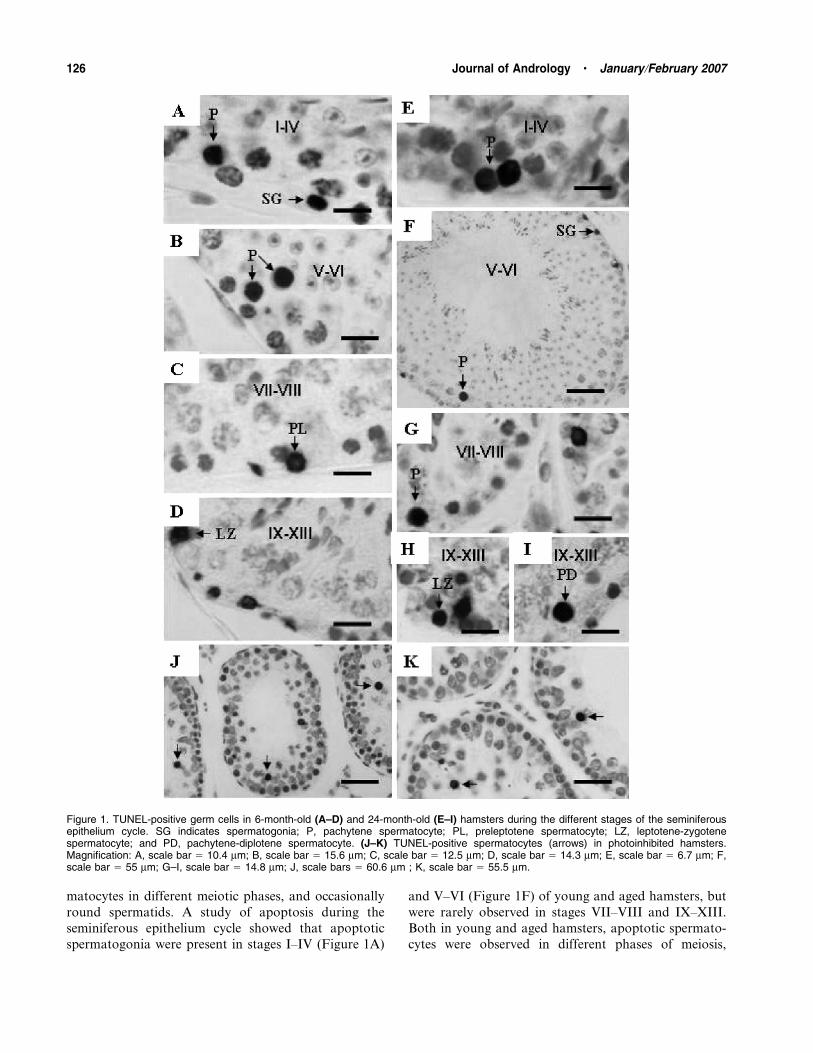
matocytes in different meiotic phases, and occasionally
round spermatids. A study of apoptosis during the
seminiferous epithelium cycle showed that apoptotic
spermatogonia were present in stages I–IV (Figure 1A)
and V–VI (Figure 1F) of young and aged hamsters, but
were rarely observed in stages VII–VIII and IX–XIII.
Both in young and aged hamsters, apoptotic spermato-
cytes were observed in different phases of meiosis,
Figure 1. TUNEL-positive germ cells in 6-month-old (A–D) and 24-month-old (E–I) hamsters during the different stages of the seminiferousepithelium cycle. SG indicates spermatogonia; P, pachytene spermatocyte; PL, preleptotene spermatocyte; LZ, leptotene-zygotenespermatocyte; and PD, pachytene-diplotene spermatocyte. (J–K) TUNEL-positive spermatocytes (arrows) in photoinhibited hamsters.Magnification: A, scale bar 5 10.4 mm; B, scale bar 5 15.6 mm; C, scale bar 5 12.5 mm; D, scale bar 5 14.3 mm; E, scale bar 5 6.7 mm; F,scale bar 5 55 mm; G–I, scale bar 5 14.8 mm; J, scale bars 5 60.6 mm ; K, scale bar 5 55.5 mm.
126 Journal of Andrology N January �February 2007

including preleptotenes in stages VII–VIII of the
seminiferous epithelium cycle (Figure 1C), early pachy-
tenes in stages I–IV (Figure 1A and E), middle
pachytenes in stages V–VI (Figure 1B and F) and VII–
VIII (Figure 1G), late pachytenes-diplotene in stagesIX–XIII (Figure 1I), and also leptotene-zygotenes in
stages IX–XIII (Figure 1D and H).
In photoinhibited hamsters the germ cells undergoing
apoptosis were predominantly early spermatocytes and,
especially, pachytene spermatocytes (Figure 1J and K)
and, on some occasions, spermatogonia and early round
spermatids. Apoptotic spermatocytes were observed in
different meiotic phases: preleptotenes, early, middle,and late pachytenes (Figure 1J and K), and leptotene-
zygotenes.
Aging and Short Photoperiod Exposure IncreaseApoptotic Indexes in the Seminiferous Epithelium ofSyrian Hamsters
Quantitatively, aging induced an increase in the
percentage of both spermatogonia and spermatocytes
in apoptosis. During the cycle of the seminiferous
epithelium, germ cell apoptosis (% of apoptotic SG+SC)
was higher in aging in stages I–IV, V–VI, and VII–VIII
(P , .05) (Table 1). The percentage of apoptoticspermatogonia was significantly increased in stages
I–IV and V–VI (P , .05), with no differences in stages
VII–VIII and IX–XIII (Table 1). The percentage of
spermatocytes in their preleptotene phase and pachytene
in apoptosis was significantly higher in aged than in
young hamsters (P , .05), with no differences in
leptotene-zygotene spermatocytes. The percentage of
pachytene spermatocytes in apoptosis was significantly
increased in stages I–IV, V–VI, and VII–VIII (P , .05),
with no differences in stages IX–XIII.
After 8 weeks of short photoperiod exposure, the
percentage of apoptotic spermatocytes was significantly
increased with respect to animals maintained in a normal
photoperiod (P , .05). But no differences in the
percentage of apoptotic spermatogonia were found
between groups (Table 2).
Immunohistochemistry
Fas/Fas-L System—Immunohistochemistry showed the
presence of Fas-L in cytoplasmic prolongations of
Sertoli cells (Figure 2A) and in Leydig cells of young,
aged (Figure 2B), and photoinhibited hamsters (Fig-
ure 2C). Also, spermatozoon tails were positive for
Fas-L in young (Figure 2A) and aged hamsters. Fas
expression was observed in Leydig cells of young and
aged hamsters (Figure 2D and E), and a slight immu-
noreactivity was also present in the tails of mature
spermatozoa (Figure 2D and E). However, in photo-
inhibited hamsters a strong expression of Fas was
observed in spermatocytes (Figure 2F) and occasionally
in Leydig cells and early round spermatids.
p53—Testicular germ cells of young and aged
hamsters were negative for p53 staining (Figure 2G
and H). However, photoinhibited hamsters showed an
intense immunoreactivity for p53, predominantly in the
spermatocytes (Figure 2I) and occasionally in the
spermatogonia and early round spermatids.
Bcl-2 Family Proteins—Staining for Bax was observed
in round spermatid acrosomes of young and aged
hamsters (Figure 2J and K). In photoinhibited hamsters
an intense immunoreactivity for Bax was observed in
spermatocytes (Figure 2L) and some staining was
observed in Leydig cells. In young and aged hamsters,
the seminiferous tubules were negative for Bcl-2 staining
Table 1. Apoptotic indexes in control (6 months) and agedhamsters (24 months)*
Control Aged hamsters
Total apoptotic index (SG+SC) (%) 0.77 6 0.04 1.51 6 0.233
Stages I–IV 0.23 6 0.08 0.58 6 0.233
Stages V–VI 0.06 6 0.03 0.60 6 0.203
Stages VII–VIII 0.38 6 0.11 0.95 6 0.463
Stages IX–XIII 0.16 6 0.10 0.21 6 0.084
Total apoptotic index in SG (%) 0.24 6 0.03 0.76 6 0.023
Stages I–IV 0.66 6 0.28 1.56 6 0.313
Stages V–VI 0.12 6 0.05 0.61 6 0.053
Stages VII–VIII 0.00 0.00
Stages IX–XIII 0.00 0.00
Total apoptotic index in SC (%) 0.20 6 0.01 0.55 6 0.063
Preleptotenes 0.65 6 0.08 1.53 6 0.343
Leptotenes-zygotenes 0.21 6 0.05 0.18 6 0.054
Pachytenes 0.12 6 0.007 0.45 6 0.063
Stages I–IV 0.18 6 0.06 0.41 6 0.133
Stages V–VI 0.02 6 0.03 0.59 6 0.333
Stages VII–VIII 0.08 6 0.05 0.39 6 0.163
Stages IX–XIII 0.10 6 0.04 0.24 6 0.054
* Values represent mean 6 SEM. SG indicates spermatogonia;
SC, spermatocytes.
3 Significant differences between control group and aged hamsters
(P , .05).
4 No significant differences between groups.
Table 2. Apoptotic indexes in control (6 months) andphotoinhibited hamsters*
Control Photoinhibited
Total apoptotic index (SG+SC) 0.77 6 0.03 2.84 6 0.163
Apoptotic index in SG 0.24 6 0.03 0.76 6 0.174
Apoptotic index in SC 0.19 6 0.01 2.62 6 0.283
* Values represent mean 6 SEM. SG indicates spermatogonia;
SC, spermatocytes.
3 Significant differences between control group and aged hamsters
(P , .05).
4 No significant differences between groups.
Morales et al N Apoptotic Pathways in Aged and Photoinhibited Testes 127
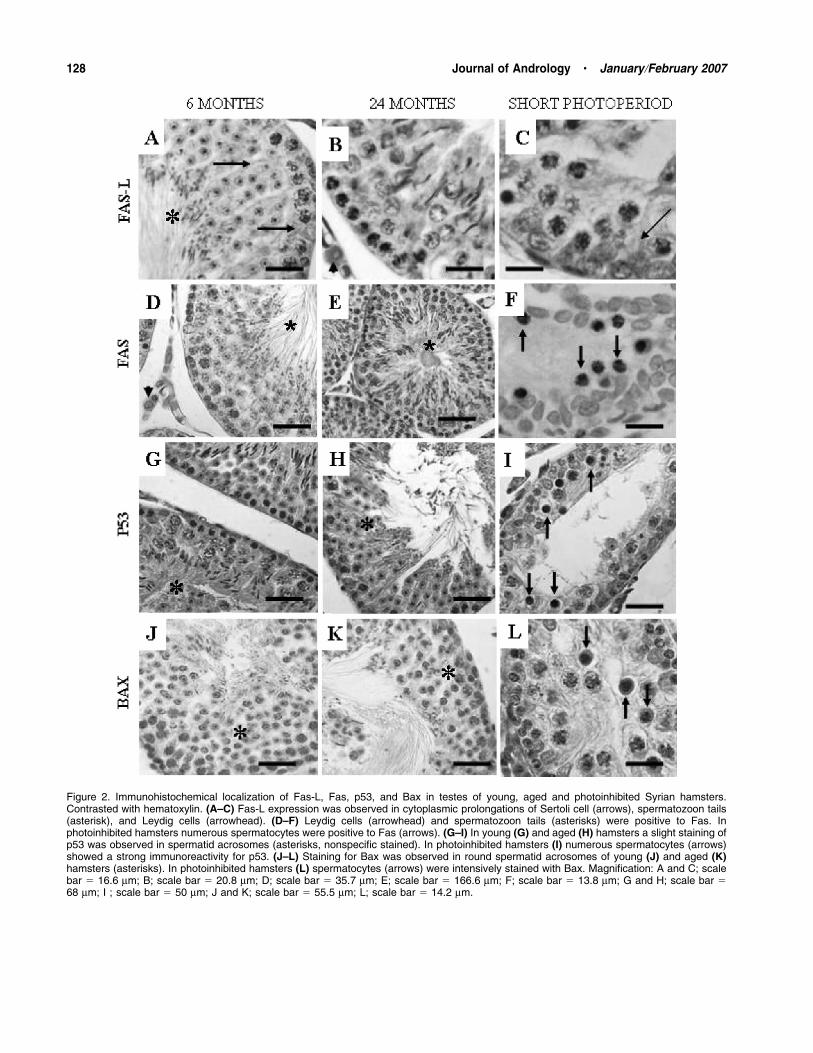
Figure 2. Immunohistochemical localization of Fas-L, Fas, p53, and Bax in testes of young, aged and photoinhibited Syrian hamsters.Contrasted with hematoxylin. (A–C) Fas-L expression was observed in cytoplasmic prolongations of Sertoli cell (arrows), spermatozoon tails(asterisk), and Leydig cells (arrowhead). (D–F) Leydig cells (arrowhead) and spermatozoon tails (asterisks) were positive to Fas. Inphotoinhibited hamsters numerous spermatocytes were positive to Fas (arrows). (G–I) In young (G) and aged (H) hamsters a slight staining ofp53 was observed in spermatid acrosomes (asterisks, nonspecific stained). In photoinhibited hamsters (I) numerous spermatocytes (arrows)showed a strong immunoreactivity for p53. (J–L) Staining for Bax was observed in round spermatid acrosomes of young (J) and aged (K)hamsters (asterisks). In photoinhibited hamsters (L) spermatocytes (arrows) were intensively stained with Bax. Magnification: A and C; scalebar 5 16.6 mm; B; scale bar 5 20.8 mm; D; scale bar 5 35.7 mm; E; scale bar 5 166.6 mm; F; scale bar 5 13.8 mm; G and H; scale bar 568 mm; I ; scale bar 5 50 mm; J and K; scale bar 5 55.5 mm; L; scale bar 5 14.2 mm.
128 Journal of Andrology N January �February 2007

(Figure 3A and B). However, in photoinhibited ham-
sters Bcl-2 was intensively expressed by Sertoli and
Leydig cells (Figure 3C).
Germ cells of young hamsters, including spermato-
gonia, spermatocytes, spermatids, and spermatozoa,
showed an reactivity to Bcl-xL during the different
stages of the seminiferous epithelium (Figure 3D). Also,
some immunoreactivity was observed in Sertoli cells.
However, both in aged and photoinhibited hamsters the
Bcl-xL staining declined in germ cells and Sertoli cells
(Figure 3E and F). Bcl-xS/L was present in germ cells
and Leydig cells of young and aged hamsters (Fig-
ure 3G). Also, in aged hamsters, an intense immunore-
activity for Bcl-xS/L was observed in spermatocytes of
the seminiferous tubules which were in maturation
arrest of spermatogenesis (Figure 3H). In photoinhib-
ited hamsters a strong immunoreactivity for Bcl-xS/L
was observed, especially in spermatocytes (Figure 3I).
Western Blot Analysis
Similar levels of Fas-L (38 kd) were found in the testes
of young, aged, and photoinhibited hamsters (Fig-ure 4A). A slight expression of Fas (48 kd) was observed
in young and aged animals. However, in hamsters
Figure 3. Immunohistochemical localization of Bcl-2, Bcl-xL, and Bcl-xS/L in the testis of young, aged, and photoinhibited Syrian hamsters.Contrasted with hematoxylin. (A–C) No immunostaining for Bcl-2 was observed in testes of young and aged hamsters. Sertoli (asterisks) andLeydig cells of photoinhibited hamsters were strongly positive to Bcl-2. (D–F) A strong immunoreactivity to Bcl-xL was observed in germ cells ofyoung hamsters (D). Bcl-xL staining decreased in aged (E) and photoinhibited hamsters (F). (G–I) Germ cells of young and aged hamsters werepositive to Bcl-xS/L (asterisks). In photoinhibited hamsters spermatocytes (arrows) were intensively stained with Bcl-xS/L. Magnification: A–C;scale bar 5 50 mm; D; scale bar 5 23.5 mm; E and F; scale bar 5 22.4 mm; G; scale bar 5 66.6 mm; H; scale bar 5 100 mm; I; scale bar 535.7 mm.
Morales et al N Apoptotic Pathways in Aged and Photoinhibited Testes 129
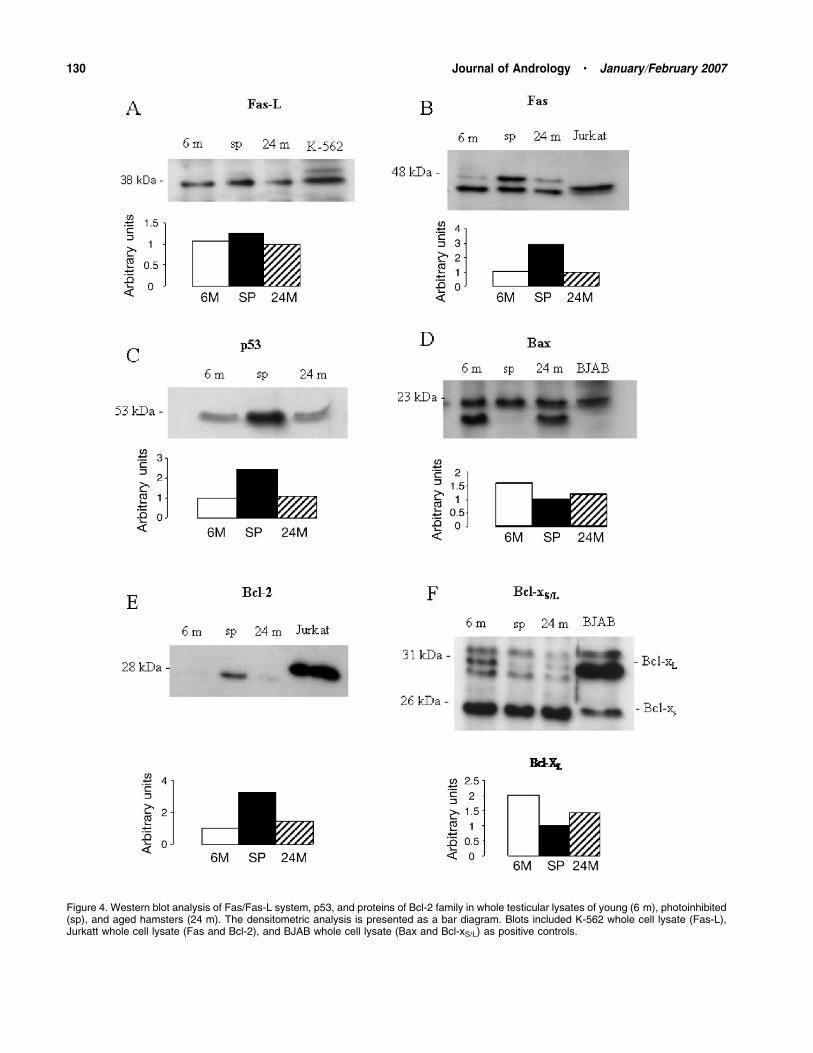
Figure 4. Western blot analysis of Fas/Fas-L system, p53, and proteins of Bcl-2 family in whole testicular lysates of young (6 m), photoinhibited(sp), and aged hamsters (24 m). The densitometric analysis is presented as a bar diagram. Blots included K-562 whole cell lysate (Fas-L),Jurkatt whole cell lysate (Fas and Bcl-2), and BJAB whole cell lysate (Bax and Bcl-xS/L) as positive controls.
130 Journal of Andrology N January �February 2007

exposed to a short photoperiod a strong expression of
Fas was observed (Figure 4B). In hamsters exposed to
a short photoperiod, the levels of p53 were higher than
in young hamsters maintained in a normal photoperiod
(Figure 4C). Aged hamsters showed levels of p53 similar
to young hamsters. Bax (23 kd) levels were similar in
young, aged, and photoinhibited hamsters (Figure 4D).
Western analysis demonstrated similar levels of Bcl-2
(28 kd) in the testes of young and aged hamsters, but
a strong expression was found in photoinhibited
hamsters (Figure: 4E). In aged hamsters and those
exposed to a short photoperiod, the levels of Bcl-xL
(31 kd) showed lower intensity than young hamsters
maintained in normal conditions (Figure 4F).
Discussion
Testicular Germ Cell Apoptosis in Aged Hamsters andAnimals Exposed to a Short Photoperiod
In young hamsters maintained in normal photoperiod
spontaneous apoptosis was observed in spermatogonia
and primary spermatocytes and occasionally in round
spermatids. Apoptotic spermatogonia were found pre-
dominantly in stages I–IV and V–VI of the seminiferous
epithelium cycle. In primary spermatocytes apoptosis
was detected predominantly in preleptotenes, and to
a lesser extent in leptotene-zygotene and pachytene. In
the Syrian hamster, apoptosis constitutes a cellular
mechanism that allows the seminiferous epithelium to
control the number of germ cells supported by Sertoli
cells, eliminating aberrant cells and regulating sperm
production (Hsueh et al, 1996). This control takes place
in stages when spermatogonia are more numerous and
during the beginning of meiosis (preleptotenes), sup-
porting the idea that apoptosis affects spermatogonia in
areas where these are too numerous, and is absent from
areas of low spermatogonia density (De Rooij and
Janssen, 1987; De Rooij and Lok, 1987).
Aging and exposure to a short photoperiod provoked
a significant increase in apoptosis in the seminiferous
epithelium of Syrian hamsters. However, aging induced
increase in apoptotic spermatogonia and spermatocytes,
whereas short photoperiod exposure encouraged an
increase in apoptotic spermatocytes without affecting
the population of spermatogonia.
Aging increased germ cell apoptosis in stages I–IV, V–
VI, and VII–VIII of the seminiferous epithelium cycle of
Syrian hamsters. In previous studies in rats, an increase
in germ cell apoptosis was observed in all the stages of
the seminiferous epithelium cycle, although the increase
was only statistically significant in stages XII–XIV
(Wang et al, 1999). Other experimental situations,
including heat exposure and testosterone deprivation,
have led to differences in the incidence of germ cell
apoptosis during the stages of the seminiferous epithe-lium cycle. After gonadotrophin deprivation and in-
tratesticular testosterone in the rat, apoptosis occurred
in stages VII–VIII, whereas hyperthermia induced germ
cell apoptosis in stages I–IV and XII–XIV (Sinha Hikim
et al, 2003). These studies indicate that apoptosis
depends on the stages of the seminiferous epithelium
cycle and that it differs according to the trigger stimuli
even in the same animal species.In the Syrian hamster, aging increased apoptosis in
the spermatogonia in stages I–IV and V–VI, while
apoptosis was absent in stages VII–VIII and IX–XIII,
similar to the results observed in young animals. As
mentioned above, these results probably reflect the
existence of an optimal and constant regulation of germ
cell density by an increasing apoptosis of spermatogonia
in areas where these cells are too numerous (De Rooijand Janssen, 1987; De Rooij and Lok, 1987). Also, aging
induced an increase in apoptotic pachytene spermato-
cytes over to the low rate found in young hamsters. It is
known that control points are not confined to cells that
divide by mitosis, but also operate during meiosis. Of
particular note is the control checkpoint at the end of the
pachytene phase of the meiotic prophase, when the
meiotic recombination and chromosome synapse areincomplete (Roeder, 1997), preventing the production of
aneuploid gametes (Roeder and Bailis, 2000). Our results
in aging hamsters suggest that this increase in apoptosis
in pachytene spermatocytes provides a control mecha-
nism of meiosis that could eliminate aberrant aged germ
cells. The strong coincidence between spontaneous and
age-induced apoptosis in the same germ cell types and in
the same stages of the seminiferous epithelium cyclesuggests that aging causes an exacerbation of apoptosis at
checkpoints where the germ cells are eliminated sponta-
neously. It is reasonable to think that the high degree of
synchronization required during the spermatogenic pro-
cess imposes a strict control over germ cell apoptosis,
both in normal situations and in situations that imply
a deteriorated spermatogenic capacity of the epithelium,
such as aging.
Germ cell types that suffer apoptosis during testicularregression after exposure to a short photoperiod may
differ between species. In photoinhibited Syrian hamster
the main apoptotic germ cells observed are the
spermatocytes and occasionally spermatogonia and
round spermatid. Similar results to these presented have
been reported in the mouse and birds (Sinha Hikim et al,
1997; Young et al, 1999, 2001). The present study is the
first to demonstrate this phenomenon quantitatively,and to indicate that apoptosis does not increase in the
population of spermatogonia. Testis exposed to short
Morales et al N Apoptotic Pathways in Aged and Photoinhibited Testes 131

photoperiod shows a pattern of cell death similar to
pharmacological deprivation of gonadotrophins (Sinha
Hikim et al, 1997; Young and Nelson, 2001). This
indicates that spermatocytes are more susceptible to
perturbations of the seminiferous epithelium environ-
ment during short photoperiod exposure, which is
characterized by severe testosterone deprivation (Furuta
et al, 1994; Calvo et al, 1997).
Role of Fas/Fas-L System, Bcl-2 Family, and p53 in GermCell Apoptosis in Aging and After ShortPhotoperiod Exposure
In the Syrian hamster Fas-L expression was observed in
Sertoli and Leydig cells as well as in spermatozoon tails
of young and aged animals. Similar results have been
documented in humans (Francavilla et al, 2000) and
rodents, including the mouse (French et al, 1996) and rat
(Lee et al, 1997). Fas-L expression in the normal testis
may be related to the fact that the testis is an
immunologically privileged organ, and Fas-L would be
involved in the elimination of active and infiltrated T
cells that express Fas (Bellgrau et al, 1995). Other
authors have described Fas-L expression in postmeiotic
germ cells, such as mature spermatozoa in the rat and
mouse (D’Alessio et al, 2001) and even in humans
(Francavilla et al, 2002), which is consistent with the
present results obtained in the Syrian hamster. These
results have led us to propose an alternative hypothesis
concerning the role of Fas-L in the reproductive system,
whereby Fas-L may represent an important molecule in
the complex mechanism developed by male gametes to
escape immunological reaction both in the male genital
tract (autoimmune reaction against auto antigens of
sperm cells) and the female genital tract (D’Alessio et al,
2001; Riccioli et al, 2003).
In young and aged Syrian hamsters, a slight expres-
sion of Fas was observed, mainly in Leydig cells, which
agrees with previous results reported in rodents, such as
mouse (Koji et al, 2001) and rat (Lee et al, 1999), as well
as in humans (Francavilla et al, 2000, 2002). These
results suggest that there is no evidence to support
a relationship between the Fas system and germ cell
apoptosis in the testis of young and aged hamsters, as
was found in the mouse (Koji, 2001). Although the Fas
system is essential for germ cell apoptosis in several
pathological situations, it is irrelevant in normal
conditions (Koji, 2001). However, Western blot analysis
revealed a significant increase in Fas expression in
photoinhibited testes. Also, immunohistochemically Fas
was strongly expressed by the spermatocytes of hamsters
exposed to a short photoperiod, which is coincident with
the predominantly TUNEL-positive germ cells observed
in the spermatocytes of these animals. In Fas-mediated
apoptosis, the binding of Fas-L to Fas antigen is
a prerequisite (Koji et al, 2001). So the temporal and
spatial association between Fas and Fas-L expression inhamsters exposed to short photoperiod suggests that the
Fas system is a mediator of germ cell apoptosis
induction after short photoperiod exposure. Although
the Fas system has been correlated with germ cell
degeneration in situations of meiotic and postmeiotic
arrest of spermatogenesis in man (Eguchi et al, 2002;
Francavilla et al, 2002), our immunohistochemical
results in aged hamsters suggest that there is noconnection between TUNEL-positive germ cells and
Fas expression. Also, Western blot analysis revealed no
changes in the expression of Fas and Fas-L in aged
animals, which suggests that the Fas/Fas-L system is not
involved in apoptosis induction of germ cells in the
testes of aged Syrian hamsters.
Another widely known regulatory system of apoptosis
is the Bcl-2 family composed of anti- and proapoptotic
proteins which regulate apoptosis by controlling therelease of cytochrome c and other mitochondrial
changes (Yang and Korsmeyer, 1996; Green and Reed,
1998). In the present study, Western blot analysis and
the immunohistochemical study demonstrated an ex-
pression of the antiapoptotic form of Bcl-xL in germ
cells of young hamsters, whereas aged and photoinhib-
ited hamsters showed a lower degree of expression. On
the other hand, proapoptotic form Bcl-xS was expressedin all the groups (young, aged, and photoinhibited).
With respect to the localization of both proteins, Bcl-xL
was expressed in all the populations of germ cells in
young hamsters. Based on the decrease of Bcl-xL
expression in germ cells of photoinhibited and aged
hamsters, as shown by Western blot analysis and
immunohistochemistry, we can affirm that when Bcl-
xS/L antibody is used, what we see predominantly is theexpression of Bcl-xS in germ cells in these 2 groups of
animals. Accordingly, and based on the immunohisto-
chemical results, in animals exposed to a short photo-
period using Bcl-xS/L antibody we found a strong
expression of Bcl-xS predominately in the spermato-
cytes, which is concordant with TUNEL-positive germ
cells as well as with other proapoptotic proteins studied
in the present report. Also the seminiferous epitheliumof aged hamsters showed germ cells positive for Bcl-xS.
As suggested by other authors, a balance of anti- and
proapoptotic members of the Bcl-2 family is critical for
regulating the survival of testicular germ cells (Oltvai et
al, 1993; Yang and Korsmeyer, 1996; Yamamoto et al,
2001; Sakamaki, 2003). Our results indicate that the
lower expression of the anti-apoptotic form Bcl-xL in
germ cells of aged and photoinhibited hamsters may bedue to a predominance of proapoptotic Bcl-xS forms,
similar to that observed by immunocytochemistry in
132 Journal of Andrology N January �February 2007

aging human testes (Kimura et al, 2003). This imbalance
between members of the Bcl-2 family, with a predomi-
nance of proapoptotic forms in germ cells, seems to beresponsible for the increase in germ cell apoptosis found
in aged hamsters activating the mitochondrial pathway
of apoptosis; the Fas/Fas-L system and p53 also
participate in the increase of germ cell apoptosis in
hamsters exposed to a short photoperiod.
The present results indicate that the levels of Bcl-2 are
low in the testis both of young and aged hamsters. These
results are congruent with previous studies obtained inmouse (Hockenbery et al, 1991; Knudson et al, 1995)
and humans (Beumer et al, 2000; Sakamaki, 2003),
which suggested that Bcl-2 plays no function in the male
gonad in normal conditions, unlike in the ovary, where
this protein is critical for primordial ovarian follicle
formation (Ratts et al, 1995). On the contrary, using
Western blot an overexpression of Bcl-2 by Sertoli and
Leydig cells was observed in hamsters exposed to shortphotoperiod compared with young animals. Studies in
transgenic animals have indicated that the overexpres-
sion of Bcl-2 in somatic cells of the testis produces
alterations in spermatogenesis, including the inhibition
of gamete formation, spermatid malformations, vacuo-
lization of the epithelium, loss of germ cells, and
increased apoptosis, while seminiferous tubules are
characterized by an accumulation of spermatogonia,Sertoli cells, and apoptotic germ cell in the meiotic
prophase (Knudson et al, 1995; Furuchi et al, 1996;
Rodrıguez et al, 1997; Yamamoto et al, 2001). These
spermatogenic defects found in transgenic animals are
comparable with the modifications observed in hamsters
exposed to a short photoperiod, which suggests that
Bcl-2 plays an important role in the atrophy of the
seminiferous epithelium of Syrian hamsters after expo-sure to the short photoperiod.
Previous studies have reported that Bax is expressed
predominantly in primary spermatocytes and sperma-
tids (Oldereid et al, 2001). Our immunohistochemical
study revealed that Bax was expressed by the spermatid
acrosomes of young and aged hamsters, which suggests
that Bax plays a role in the maturation and differenti-
ation process of the acrosome in the hamster, as has
been suggested in other species (Oldereid et al, 2001).Also, Bax has been classified as a proapoptotic member
of the Bcl-2 family and has been related to the induction
of apoptosis of spermatocytes and spermatids in hu-
mans (Oldereid et al, 2001). Based on our immunohis-
tochemical results, Bax does not seen to be involved in
germ cell apoptosis in young or aged hamsters.
However, in hamsters exposed to short photoperiod,
the expression of Bax by the spermatocytes wasconcordant with the expression of Fas and p53 as well
as with TUNEL-positive cells. These results suggest that
Bax is involved in germ cell apoptosis induction after
exposure to a short photoperiod.
Strong immunostaining with p53 was found in
spermatocytes and occasionally in spermatogonia of
photoinhibited hamsters. These results indicate that p53
expression is concordant with Fas and Bax expression as
well as with TUNEL-positive germ cells in photoinhib-
ited hamsters, which suggests that p53 is involved in
germ cell apoptosis induction after short photoperiod
exposure.
In summary, aging of the seminiferous epithelium in
the Syrian hamster is characterized by an increase in
germ cell apoptosis in the populations of spermatogonia
and spermatocytes and is dependent on the stage of the
cycle. After short photoperiod exposure, however, the
increase in apoptosis is only observed in the spermato-
cytes. Different molecular pathways are triggered toinduce germ cell apoptosis in aged animals and those
exposed to a short photoperiod. The results obtained
show both the intrinsic and extrinsic pathways being
activated after short photoperiod exposure, but only the
intrinsic pathway during aging.
ReferencesBarnes CJ, Covington BW IV, Cameron IL, Lee M. Effect of aging on
spontaneous and induced mouse testicular germ cell apoptosis.
Aging (Milano). 1998;10:497–501.
Barnes CJ, Covington BW IV, Lee M. Effect of aging and dietary
restriction on rat testicular germ cell apoptosis. J Gerontol A Biol
Sci Med Sci. 1999;54:B199–204.
Bellgrau D, Gold D, Selawry H, Moore J, Franzusoff A, Duke RC. A
role for CD95 ligand in preventing graft rejection. Nature.
1995;337:630–632.
Beumer TL, Roepers-Gajadien HL, Gademan IS, Lock T, Kal HB, De
Rooij DG. Apoptosis regulation in the testis: involvement of Bcl-2
family members. Mol Reprod Dev. 2000;56:353–359.
Beumer TL, Roepers-Gajadien HL, Gademan IS, Van Buul PP, Gil-
Gomez G, Rutgers DH, De Rooij DG. The role of the tumor
suppressor p53 in spermatogenesis. Cell Death Differ. 1998;5:669–
677.
Bitko V, Barik S. An endoplasmic reticulum-specific stress-activated
caspase (caspase-12) is implicated in the apoptosis of A549
epithelial cells by respiratory syncytial virus. J Cell Biochem.
2001;80:441–454.
Blanco-Rodrıguez J, Martınez-Garcıa C. Apoptosis pattern elicited by
several apoptogenic agents on the seminiferous epithelium of the
adult rat testis. J Androl. 1998;19:487–497.
Blanco-Rodrıguez J, Martınez-Garcıa C. Induction of apoptotic cell
death in the seminiferous tubule of the adult rat testis: assessment
of the germ cell types that exhibit the ability to enter apoptosis after
hormone suppression by oestrodiol treatment. Int J Androl.
1996;19:237–247.
Brinkworth MH, Weinbauer GF, Bergmann M, Nieschlag E.
Apoptosis as a mechanism of germ cell loss in elderly men.
Int J Androl. 1997;20:222–228.
Cai L, Hales BF, Robaire B. Induction of apoptosis in the germ cells of
adult male rats after exposure to cyclophosphamide. Biol Reprod.
1997;56:1490–1497.
Morales et al N Apoptotic Pathways in Aged and Photoinhibited Testes 133

Calvo A, Bustos-Obregon E, Pastor LM. Morphological and
histochemical changes in the epididymis of hamsters (Mesocricetus
auratus) subjected to short photoperiod. J Anat. 1997;191:77–
88.
Calvo A, Pastor LM, Martınez E, Vazquez JM, Roca J. Age-related
changes in the hamster epididymis. Anat Rec. 1999;256:335–346.
D’Alessio A, Riccioli A, Lauretti P, Padula F, Muciaccia B, De Cesaris
P, Filippini A, Nagata S, Ziparo E. Testicular FasL is expressed by
sperm cells. Proc Natl Acad Sci U S A. 2001;98:3316–3321.
De Rooij DG, Janssen JM. Regulation of the density of spermatogo-
nia in the seminiferous epithelium of the Chinese hamster: I.
Undifferentiated spermatogonia. Anat Rec. 1987;217:124–130.
De Rooij DG, Lok D. Regulation of the density of spermatogonia in
the seminiferous epithelium of the Chinese hamster: II. Differen-
tiating spermatogonia. Anat Rec. 1987;217:131–136.
Eguchi J, Koji T, Nomata K, Yoshii A, Shin M, Kanetake H. Fas-Fas
ligand system as a possible mediator of spermatogenic cell
apoptosis in human maturation-arrested testes. Hum Cell. 2002;15:
61–68.
Francavilla S, D’Abrizio P, Cordeschi G, Pelliccione F, Necozione S,
Ulisse S, Properzi G, Francavilla F. Fas expression correlates with
human germ cell degeneration in meiotic and post-meiotic arrest of
spermatogenesis. Mol Hum Reprod. 2002;8:213–220.
Francavilla S, D’Abrizio P, Rucci N, Silvano G, Properzi G, Straface
E, Cordeschi G, Necozione S, Gnessi L, Arizzi M, Ulisse S. Fas
and Fas ligand expression in fetal and adult human testis with
normal or deranged spermatogenesis. J Clin Endocrinol Metab.
2000;85:2692–2700.
French LE, Hahne M, Viard I, Radlgruber G, Zanone R, Becker K,
Muller C, Tschopp J. Fas and Fas ligand in embryos and adult
mice: ligand expression in several immune-privileged tissues and
coexpression in adult tissues characterized by apoptotic cell
turnover. J Cell Biol. 1996;133:335–343.
Furuchi T, Masuko K, Nishimune Y, Obinata M, Matsui Y.
Inhibition of testicular germ cell apoptosis and differentiation
in mice misexpressing Bcl-2 in spermatogonia. Development.
1996;122:1703–1709.
Furuta I, Porkka-Heiskanen T, Scarbrough K, Tapanainen J, Turek
FW, Hsueh AJ. Photoperiod regulates testis cell apoptosis in
Djungarian hamsters. Biol Reprod. 1994;51:1315–1321.
Green DR, Reed JC. Mitochondria and apoptosis. Science. 1998;281:
1309–1312.
Hockenbery DM, Zutter M, Hickey W, Nahm M, Korsmeyer SJ. Bcl-
2 protein is topographically restricted in tissues characterized
by apoptotic cell death. Proc Natl Acad Sci U S A. 1991;88:6961–
6965.
Horn R, Pastor LM, Moreno E, Calvo A, Canteras M, Pallares J.
Morphological and morphometric study of early changes in the
ageing golden hamster testis. J Anat. 1996;188:109–117.
Hsueh AJ, Eisenhauer K, Chun SY, Hsu SY, Billig H. Gonadal cell
apoptosis. Recent Prog Horm Res. 1996;51:433–455.
Itoh N, Yonehara S, Ishii A, Yonehara M, Mizushima S, Sameshima
M, Hase A, Seto Y, Nagata S. The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis.
Cell. 1991;66:233–243.
Kimura M, Itoh N, Takagi S, Sasao T, Takahashi A, Masumori N,
Tsukamoto T. Balance of apoptosis and proliferation of germ
cells related to spermatogenesis in aged men. J Androl. 2003;24:
185–191.
Knudson CM, Tung KS, Tourtellotte WG, Brown GA, Korsmeyer SJ.
Bax-deficient mice with lymphoid hyperplasia and male germ cell
death. Science. 1995;270:96–99.
Koji T. Male germ cell death in mouse testes: possible involvement of
Fas and Fas ligand. Med Electron Microsc. 2001;34:213–222.
Krajewski S, Bodrug S, Krajewska M, Shabaik A, Gascoyne R,
Berean K, Reed JC. Immunohistochemical analysis of mcl-1
protein in human tissues. differential regulation of Mcl-1 and
Bcl-2 protein production suggests a unique role for Mcl-1 in
control of programmed cell death in vivo. Am J Pathol. 1995;146:
1309–1319.
Leblond C, Clermont Y. Spermiogenesis of rat, mouse, hamster and
guinea pig as revealed by the ‘‘periodic acid-fuchsin sulfurous acid’’
technique. Am J Anat. 1952;90:167–206.
Lee J, Richburg JH, Shipp EB, Meistrich ML, Boekelheide K. The Fas
system, a regulator of testicular germ cell apoptosis, is differentially
up-regulated in Sertoli cell versus germ cell injury of the testis.
Endocrinology. 1999;140:852–858.
Lee J, Richburg JH, Younkin SC, Boekelheide K. The Fas system is
a key regulator of germ cell apoptosis in the testis. Endocrinology.
1997;138:2081–2088.
Morales E, Horn R, Pastor LM, Santamaria L, Pallares J, Zuasti A,
Ferrer C, Canteras M. Involution of seminiferous tubules in
aged hamsters: an ultrastructural, immunohistochemical and
quantitative morphological study. Histol Histopathol. 2004;19:
445–455.
Morales E, Pastor LM, Ferrer C, Zuasti A, Pallares J, Horn R, Calvo
A, Santamarıa L, Canteras M. Proliferation and apoptosis in the
seminiferous epithelium of photoinhibited Syrian hamsters (Meso-
cricetus auratus). Int J Androl. 2002;25:281–287.
Morales E, Pastor LM, Horn R, Zuasti A, Ferrer C, Calvo A,
Santamaria L, Canteras M. Effect of ageing on the proliferation
and apoptosis of testicular germ cells in the Syrian hamster
Mesocricetus auratus. Reprod Fertil Dev. 2003;15:89–98.
Nakagawa T, Zhu H, Morishima N, Li E, Xu J, Yankner BA, Yuan J.
Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and
cytotoxicity by amyloid-beta. Nature. 2000;403:98–103.
Oehm A, Behrmann I, Falk W, Pawlita M, Maier G, Klas C, Li-Weber
M, Richards S, Dhein J, Trauth BC, et al. Purification and
molecular cloning of the APO-1 cell surface antigen, a member of
the tumor necrosis factor/nerve growth factor receptor superfamily.
Sequence identity with the Fas antigen. J Biol Chem. 1992;267:
10709–10715.
Oldereid NB, De Angelis P, Wiger R, Clausen OPF. Expression of Bcl-
2 family proteins and spontaneous apoptosis in normal human
testis. Mol Hum Reprod. 2001;7:403–408.
Oltvai ZN, Milliman CL, Korsmeyer SJ. Bcl-2 heterodimerizes in vivo
with a conserved homolog, Bax, that accelerates programmed cell
death. Cell. 1993;74:609–619.
Pentikainen V, Erkkila K, Dunkel L. Fas regulates germ cell apoptosis
in the human testis in vitro. Am J Physiol. 1999;276:E310–316.
Ratts VS, Flaws JA, Kolp R, Sorenson CM, Tilly JL. Ablation of bcl-2
gene expression decreases the numbers of oocytes and primordial
follicles established in the post-natal female mouse gonad.
Endocrinology. 1995;136:3665–3668.
Riccioli A, Salvati L, D’Alessio A, Starace D, Giampietri C, De
Cesaris P, Filippini A, Ziparo E. The Fas system in the semini
ferous epithelium and its possible extra-testicular role. Andrologia.
2003;35:64–70.
Rodriguez I, Ody C, Araki K, Garcia I, Vassalli P. An early and
massive wave of germinal cell apoptosis is required for the
development of functional spermatogenesis. EMBO J. 1997;16:
2262–2270.
Roeder GS. Meiotic chromosomes: it takes two to tango. Genes Dev.
1997;11:2600–2621.
Roeder GS, Bailis JM. The pachytene checkpoint. Trends Genet.
2000;16:395–403.
Sakamaki K. Physiological and pathological cell deaths in the
reproductive organs. Cell Struct Funct. 2003;28:31–40.
134 Journal of Andrology N January �February 2007

Sinha Hikim AP, Lue Y, Diaz-Romero M, Yen PH, Wang C,
Swerdloff RS. Deciphering the pathways of germ cell apoptosis in
the testis. J Steroid Biochem Mol Biol. 2003;85:175–182.
Sinha Hikim AP, Rajavashisth TB, Sinha Hikim I, Lue Y, Bonavera
JJ, Leung A, Wang C, Swerdloff RS. Significance of apoptosis in
the temporal and stage-specific loss of germ cells in the adult rat
after gonadotrophin deprivation. Biol Reprod. 1997;57:1193–1201.
Stephan H, Polzar B, Rauch F, Zanotti S, Ulke C, Mannherz HG.
Distribution of deoxyribonuclease I (DNase I) and p53 in rat testis
and their correlation with apoptosis. Histochem Cell Biol. 1996;106:
383–393.
Suda T, Takahasni T, Golstein P, Nagata S. Molecular cloning and
expression of the Fas ligand, a novel member of the tumor necrosis
factor family. Cell. 1993;75:1169–1178.
Tiba T, Takahashi M, Igura M, Kita I. Enhanced proliferation of
undifferentiated spermatogonia after of short photoperiod expo-
sure in the Syrian hamster (Mesocricetus auratus). Anat Histol
Embryol. 1992;21:9–22.
Wang C, Sinha Hikim AP, Lue Y, Leung A, Baravarian S, Swerdloff
RS. Reproductive aging in the Brown Norway rat is characterized
by accelerated germ cell apoptosis and is not altered by luteinizing
hormone replacement. J Androl. 1999;20:509–518.
Woolveridge I, De Boer-Brouwer M, Taylor MF, Teerds KJ, Wu FC,
Morris ID. Apoptosis in the rat spermatogenic epithelium
following androgen withdrawal: changes in apoptosis-related
genes. Biol Reprod. 1999;60:461–470.
Yamamoto CM, Sinha Hikim AP, Huynh PN, Shapiro B, Lue Y,
Salameh WA, Wang C, Swerdloff RS. Redistribution of Bax is an
early step in an apoptotic pathway leading to germ cell death in
rats, triggered by mild testicular hyperthermia. Biol Reprod.
2000;63:1683–1690.
Yamamoto C, Sinha Hikim AP, Lue Y, Portugal AM, Guo TB,
Saklameh WA, Wang W, Hsued JW, Swerdloff RS. Impairment of
spermatogenesis in transgenic mice with selective overexpression of
Bcl-2 in the somatic cells of the testis. J Androl. 2001;22:981–991.
Yang E, Korsmeyer SJ. Molecular thanatopsis: a discourse on the Bcl-
2 family and cell death. Blood. 1996;88:386–401.
Yoneda T, Imaizumi K, Oono K, Yui D, Gomi F, Katayama T,
Tohyama M. Activation of caspase-12, an endoplastic reticulum
(ER) resident caspase, through tumor necrosis factor receptor-
associated factor 2-dependent mechanism in response to the ER
stress. J Biol Chem. 2001;276:13935–13940.
Young KA, Ball GF, Nelson RJ. Photoperiod-induced testicular
apoptosis in European starlings (Sturnus vulgaris). Biol Reprod.
2001;64:706–713.
Young KA, Zirkin BR, Nelson RJ. Short photoperiods evoke
testicular apoptosis in white-footed mice (Peromyscus leucopus).
Endocrinology. 1999;140:3133–3139.
Young KA, Zirkin BR, Nelson RJ. Testicular regression in response to
food restriction and short photoperiod in white-footed mice
(Peromyscus leucopus) is mediated by apoptosis. Biol Reprod.
2000;62:347–354.
Morales et al N Apoptotic Pathways in Aged and Photoinhibited Testes 135




















