
Intracellular angiotensin II inhibits heterologous receptor stimulated Ca2+ entry
Upload
independentCategory
view
0download
0
ORIGINAL ARTICLE
Angiotensin-(1–7) Modulates Angiotensin II-InducedVasoconstriction in Human Mammary Artery
Luís Mendonça & Pedro Mendes-Ferreira & Ana Bento-Leite & Rui Cerqueira &
Mário Jorge Amorim & Paulo Pinho & Cármen Brás-Silva & Adelino F. Leite-Moreira &
Paulo Castro-Chaves
Published online: 10 October 2014# Springer Science+Business Media New York 2014
AbstractPurpose The renin-angiotensin system plays a key role incardiovascular pathophysiology and one of its members, an-giotensin-(1–7) (ANG-(1–7)), is now recognized as a peptidewith the ability to counter-regulate angiotensin II (ANGII)effects.We sought to investigate ANG-(1–7) actions in humanvessels, particularly its effect onANGII-induced vasoconstric-tion in human mammary arteries (HMA).Methods Samples of HMA from patients submitted to coro-nary revascularization (22 patients, mean age 67 years) werecut into small rings, mounted in a myograph bath system,normalized and allowed to contract and dilate isometrically. Inbaseline experiments, the rings were incubated with ANG-(1–7) or vehicle, followed by increasing concentrations ofANGII. This protocol was repeated in the presence ofA-779, PD123177, losartan and after mechanical endotheliumremoval. Western blot analysis and immunofluorescence werealso performed in order to verify the presence of Mas receptorin HMA.Results ANG-(1–7) significantly attenuated ANGII-inducedcontraction, producing a maximal inhibition of approximately65.2%. This effect was not abolished by A-779, PD123177 orendothelium removal. In the presence of losartan, ANGIIresponse was attenuated and no differences were observedbetween ANG-(1–7) and vehicle treated rings. Finally, we
observed, for the first time, that the Mas receptor is expressedin HMA endothelium.Conclusions ANG-(1–7) significantly attenuates ANGII-induced vasoconstriction and, although the Mas receptor isexpressed in HMA, this effect seems to be independent of itsactivation. Additionally, AT2 receptor and endothelium are notinvolved in this mechanism, which suggests a direct effect onsmooth muscle cells.
Keywords Angiotensin-(1–7) . Angiotensin II . Vascularfunction . Humanmammary artery .Mas receptor .
AT1 receptor
Introduction
The renin-angiotensin system (RAS) plays a central role incardiovascular homeostasis [1]. Despite its increasingly rec-ognized importance in a wide variety of biologic processes,the RAS is primarily involved in cardiovascular and renalfunction control, being a crucial element of blood pressureregulation [1]. Classically, the RAS was viewed as a linearenzymatic cascade resulting in angiotensin II (ANG II) for-mation, one of the most important mediators involved incardiovascular diseases, including hypertension and heart fail-ure [2]. However, further investigation provided a new insightinto this system, revealing a more complex and dynamicnetwork formed by numerous molecular pathways [3].Among these, angiotensin-(1–7) (ANG-(1–7)) emerged asan important mediator able to counter-regulate ANG II dele-terious cardiovascular effects, holding the promise of newtherapeutic strategies [4]. Besides its well described cardiacand renal actions, ANG-(1–7) has important direct vasculareffects, namely anti-remodeling [5], anti-inflammatory [6],anti-atherogenic [7, 8] and vasodilator [9–11]. According todata derived from animal studies, ANG-(1–7) exerts its action
L. Mendonça : P. Mendes-Ferreira :A. Bento-Leite :R. Cerqueira :M. J. Amorim : P. Pinho :C. Brás-Silva :A. F. Leite-Moreira :P. Castro-Chaves (*)Department of Physiology and Cardiothoracic Surgery, Faculty ofMedicine, University of Porto, Alameda Prof. Hernâni Monteiro,4200-319 Porto, Portugale-mail: [email protected]
R. Cerqueira :M. J. Amorim : P. Pinho :A. F. Leite-MoreiraDepartment of Cardiothoracic Surgery, Centro Hospitalar S. João,Porto, Portugal
Cardiovasc Drugs Ther (2014) 28:513–522DOI 10.1007/s10557-014-6555-4

through the G-coupled Mas receptor [12]. Although the intra-cellular mediators involved in Mas receptor signaling are notfully characterized, the role of endothelium and of its derivedmetabolites is probably very important [13, 14]. Indeed, it hasbeen previously demonstrated that Mas receptor activationtriggers endothelial NO synthesis and release through a path-way that depends on serine/threonine protein kinase Akt sig-naling [15]. Regarding humans, an exhaustive researchthrough literature reveals some reports about Mas receptorexpression and signalling in different tissues [16–18] but noneof them has actually evaluated the functional role of thisreceptor in intact vascular segments [19].
The functional relevance of other receptors included in theclassical RAS, mainly AT1 and AT2 receptors, is less clear inANG-(1–7) response. Some evidence suggests that the AT2
receptor may contribute to vasodilation induced by ANG-(1–7) in mouse coronary artery. Regarding this, Castro et al.observed that in the presence of a specific AT2 receptorinhibitor (PD123319), ANG-(1–7) treatment did not reducecoronary perfusion pressure [20] and, more recently, a similareffect was demonstrated in a model of rat heart hypertrophy[21]. This effect is less clear in humans. Van de Wal et al.demonstrated that AT2 stimulation by CGP42112A did notelicit a vasodilator response in human mammary arteries(HMA) [22]. Considering the AT1 receptor, preliminary stud-ies have shown that Ang-(1–7) may be involved in downregulation of this receptor in vascular smooth cells [23].
Despite all the observed effects of ANG-(1–7) in experi-mental animals, evidence regarding ANG-(1–7) actions inhuman vascular tissues is still scarce. Previous studies, per-formed in vivo and in vitro, suggest that ANG-(1–7) attenuatesANG II-induced vasoconstriction [24, 25]. However, themechanisms underlying this response, including the role ofthe Mas receptor, have not been explored.
Besides being widely used in coronary artery bypassgrafting (CABG) surgery, HMA are an excellent conduit toevaluate functional and molecular mechanisms given theirpeculiar properties, namely their resistance to endothelial dys-function [26], vascular aging and atherosclerotic degeneration[27]. These properties distinguish them functionally and struc-turally from other vascular tissues, like aorta [27, 28]. Thus,the main goals of our work are to characterize the mechanismsunderlying ANG-(1–7)-induced vasodilation in HMA and toidentify the presence of the Mas receptor in this tissue.
Methods
Patients’ Selection and Sample Collection
A serial group of patients (n=22) undergoing coronary arterybypass graft (CABG) surgery in a University Hospital (CentroHospitalar S. João, Porto, Portugal) was selected and
characterized according to the following parameters: age, sex,hypertension, diabetes mellitus, dyslipidemia, smoking habits,RAS antagonist therapy and HMG-CoA inhibitor therapy.HMA harvesting was performed by members of the surgicalteam during surgery and, after confirming that its distal regionsegments were to be discarded, they were collected and placedin cold Krebs solution (4 °C) with the following composition(mM): 130 NaCl; 4.7 KCl; 14.9 NaHCO3; 1.18 KH2PO4; 5.5glucose; 1.17 MgSO4·7H2O and 1.6 CaCl2·2H2O. The timebetween the operating room and the arrival of these segments tothe laboratory was approximately 5 min. This project wasapproved by the local ethics committee and informed consentwas obtained from all patients.
Functional Measurements
Preparation of Vascular Tissue for Organ Bath Experiments
HMAwere prepared for organ bath experiments as previouslydescribed [29]. Briefly, all the connective tissue attached to theouter surface of the vascular wall was removed and each HMAsegment (n=22) was sectioned into 2 mm rings. Arterial ringswere mounted in 5 mL baths containing Krebs solution (pH 7.4±0.1) at 32 °C, bubbled with a 95%O2 / 5 %CO2 gas mixture,and connected to a force transducer (Multi Wire MyographSystemModel 620M –DMT ). Arterial rings were normalizedto an optimal diameter corresponding to the effectivetransmural pressure of 100 mmHg, according to a previouslydescribed protocol [21]. After the normalization procedure, thevessels were allowed to stabilize for 60 min. Vascular viabilitywas determined by incubating rings with KCl (0.1 M). Endo-thelial function was assessed by incubating them with increas-ing concentrations of acetylcholine (10−9–10−5 M) afterphenylephrine-induced vasoconstriction.
ANG-(1–7) Effect on Phenylephrine-InducedVasoconstriction
In order to obtain more complete information about the role ofANG-(1–7) on phenylephrine-induced contraction, its effectwas tested in two different conditions. In the first experimentalprotocol, we determined the ANG-(1–7) concentration-response curve in vessels preconstricted with phenylephrine.In the second protocol, we studied the effect of ANG-(1–7)pretreatment on phenylephrine-induced vasoconstriction, asdetailed below.
ANG-(1–7) Added After Phenylephrine Response
Arterial rings from five patients (two rings from each patient)were contracted with phenylephrine in order to draw a
514 Cardiovasc Drugs Ther (2014) 28:513–522
®

concentration-response curve (10−9–10−5 M). After thehighest concentration, no washing was performed and a sta-bilization period was allowed. Then, the rings from eachpatient were randomly assigned to be incubated with increas-ing concentrations of ANG-(1–7) (10−9–10−5 M) or vehicle.
ANG-(1–7) Added Before Phenylephrine Response
Studying the same five patients, other two rings from eachpatient were randomly selected to be treated with ANG-(1–7)(10−5 M) or vehicle before phenylephrine-induced contrac-tion. After 20 min of incubation and without performing anywashing, an increasing concentration-response curve to phen-ylephrine (10−9–10−5 M) was performed.
ANG-(1–7) effect on ANG II-induced vasoconstriction
Baseline Protocol
Two arterial rings from 17 patients (patients from previousexperimental protocol were not included) were contractedwith phenylephrine (10−5 M) and the maximum active tensiondeveloped was recorded. After a washing and stabilizationperiod, the rings from each patient were randomly selected tobe incubated with ANG-(1–7) (10−5 M) or vehicle for 20 min.After this period, no washing was performed and aconcentration-response curve to ANG II (10−9–10−5 M).
Mas Receptor Inhibition and Endothelium Removal
In order to evaluate the influence of the Mas receptor onANG-(1–7) effect, HMA from 6 individuals (two rings fromeach one) were incubated with a Mas receeptor inhibitor(A-779, 10−5 M). In other experimental protocol, segmentvessels from seven patients (14 rings) were submitted tomechanical endothelium removal. The accuracy of endotheli-um abrasion was evaluated by performing a concentration-response curve to acetylcholine.
Subsequently to these two treatments, the protocol wasperformed as described in the “baseline protocol”.
AT1 and AT2 Receptors Inhibition
In order to study the role of other receptors classically in-volved in RAS network, HMA arterial rings were incubatedwith the AT2 receptor antagonist PD 123177 (10−6 M; n=14rings from 7 patients) or the AT1 receptor antagonist losartan(10−6 M; n=12 rings from 6 patients) for 20 min. Afterincubation period without any washing, the experiment wasperformed as previously described in the “baseline protocol”.
Data Acquisition
Data were continuously acquired and digitally recorded(PowerLab 4/30 ADInstruments ). The following parameterswere determined: dimensions (segment length and internalcircumference, mm), tension (T, mN/mm) and effectivetransmural pressure (ERTP, kPa).
Mas Receptor Expression in HMA
Western Blot Analysis
Western blot analysis was used to evaluate the expression of theMas receptor in HMA. Two samples from each of three ran-domly selected patients were mechanically homogenized in ice-cold RIPA lysis buffer (Cell Signaling) supplemented with pro-tease inhibitor cocktail (Sigma-Aldrich). Tissue was then centri-fuged at 13.000 RPM for 10 min at 4 °C. Supernatants werecollected and the total protein concentration was determinedusing the Bradford assay (BioRad). Samples containing 20 μgof protein were separated in a 10 % SDS-PAGE gel and trans-ferred into a nitrocellulosemembrane overnight at 4 °C. Ponceaustaining was performed to verify the quality of the transfer and toensure similar protein loading. Membranes were blocked in 5 %bovine serum albumin (BSA, Sigma Aldrich) diluted in TrisBuffered Saline with 0.1 % Tween-20 (TBS-T) for 1 h, andincubated overnight at 4 °C with antibody against the Masreceptor at a 1:500 dilution in 2 % BSATBS-T (MAS1 H-117,rabbit polyclonal, Santa Cruz Biotechnology). Primary antibodydetection was performed using an anti-rabbit near-infrared fluo-rescent secondary antibody at a 1:15000 dilution in 2 % BSATBS-T (IRDye 800CW, Li-cor) and detected using an infraredimaging system (Odyssey infrared imaging system, Li-cor ).
Immunofluorescence
For immunofluorescence studies, HMAwas processed as pre-viously described [30]. Briefly, vascular segments were fixedwith 4 % phosphate-buffered paraformaldehyde, pH 7.4, for1 h, and then they were washed in three changes of phosphate-buffered saline solution (PBS; pH 7.4). After clearing, arterialsegments were placed in PBS containing 30 % sucrose over-night, transferred to a cryomold containing Tissue-Tek OCTembedding medium, and frozen in liquid nitrogen. Tissueswere kept at −70 °C until the day of the experiments. Frozentransverse sections (4 μm) were incubated with a primaryantibody against theMas receptor (1:100; MAS1H-117, rabbitpolyclonal, Santa Cruz Biotechnology) overnight at 4 °C. Afterbeing washed, rings were incubated with the secondary anti-body (Alexa Fluor 647 goat anti-rabbit, Life technologies) andincubated with 4’6-diamidino-2-phenylindole (DAPI) prior tocover slip placement. Immunofluorescent signals were viewed
Cardiovasc Drugs Ther (2014) 28:513–522 515
®
®

using an inverted Leica TCS SP2 confocal laser scanningmicroscope (Leica, Heidelberg, Germany) with oil immersionlens (×63). The specificity of the immunostaining was evalu-ated by omission of the primary antibody, and slides processedas above mentioned.
Statistical Analysis
Values are presented as means±standard error. In HMA rings,treatment with the different pharmacological agents and re-spective controls (vehicle) was performed within parallel ringsof each single patient. Regarding phenylephrine (PE) concen-tration response curve, the means of the absolute responseswere calculated first per patient and then expressed as apercentage of the maximum active tension developed duringthe initial KCl preincubation (AT (PE/K, %)). Similarly, dur-ing ANG II concentration response curve, the means of abso-lute responses were calculated first per patient and thenexpressed as a percentage of the maximum active tensiondeveloped during phenylephrine preincubation (AT (ANGII/PEmax, %)). EC50 for each patient response was calculatedusing logistic regression. The two-WayANOVA test was usedto compare concentration response curves between experi-mental groups. EC50 comparisons were performed using theANOVA and Student-Newman-Keuls tests for multiple pairedcomparisons. P<0.05 was considered statistically significant.
Results
The patients selected for the experiments (n=22) had anaverage age of 67 years old (range 51 to 81 years). Fourteenpatients were male and 8 were female. The most relevantclinical data are shown in Table 1.
Functional Measurements
In the performed experiments, HMA did not present sponta-neous active tone. Additionally, incubation with ANG-(1–7)before the addition of phenylephrine or ANG II did not resultin any significant change on HMA basal tone. All the patientshad preserved endothelial function, as indicated by a maxi-mum decrease in active tension of approximately 80 % inresponse to acetylcholine, as previously reported [31].
ANG-(1–7) Effect on Phenylephrine-InducedVasoconstriction
When increasing concentrations of ANG-(1–7) (10−9–10−5 M)were added to arterial rings previously contracted with phenyl-ephrine, they did not elicit a significantly different response
from the vehicle treated rings. After incubation with the highestANG-(1–7) concentration, the developed active tension was57.8±5.4 % (PE/K+
max) in the ANG-(1–7) treated vessels and55.2±4.7 % (PE/K+
max) in the vehicle treated ones (Fig. 1a).Regarding the effect of ANG-(1–7) pretreatment on
phenylephrine-induced vasoconstriction, no difference in theconcentration-response curve was observed betweenANG-(1–7) or vehicle-treated rings (Fig. 1b).
ANG-(1–7) Effect on ANG II-Induced Vasoconstriction
ANG II-induced vasoconstriction was significantly inhibitedby ANG-(1–7) pretreatment. Namely, we observed a maxi-mum increase in active tension of only 16.1±2.4 % (ANG II/PEmax) in the ANG-1-7 treated group when compared to the46.2±3.3 % increase in the control group (Fig. 2; P<0.01).ANG II-induced maximum active tension was observed at 3·10−7 M in the ANG-(1–7)-treated group and at 3·10−8 M inthe vehicle-treated one. Despite the tendency of ANG-(1–7) todecrease ANG II vasoconstrictor sensitivity, no significantincrease in the EC50 was observed (EC50, 3.62±1.28 M vs.6.45±1.63·10−8 M, respectively).
Mas Receptor Inhibition and Endothelium Removal
Treatment with A-779 or endothelial removal did not signif-icantly change ANG-(1–7) effect on ANG II-induced vaso-constriction. In the A-779 treatment protocol, we observed amaximum increase in active tension of 15.4±2.3 % (ANG II/PEmax) in ANG-(1–7) treated rings and an increase of 49.4±3.6 % (ANG II/PEmax) in the vehicle treated rings (Fig. 3a,P<0.01). Regarding EC50, no difference was observed
Table 1 Clinical characteristics of 22 patients selected to vascular func-tion evaluation
General characteristics
Mean age (range), y 67 (51–81)
Male/female, n 14/8
Clinical history
Hypertension, n (%) 12 (54.5)
Diabetes, n (%) 7 (32)
Current smoker, n (%) 9 (41)
Preoperative medication
Statin, n (%) 14 (64)
RAS blocker (ARAII/ACEi), n (%) 8 (36)
Diuretic, n (%) 4 (18)
Nitrates, n (%) 4 (18)
Oral antiplatelet agents, n (%) 14 (64)
Calcium antagonists, n (%) 6 (27)
RAS renin-angiotensin system; ARAII Angiotensin II receptor 1 antago-nist; ACEi angiotensin converting enzyme inhibitor
516 Cardiovasc Drugs Ther (2014) 28:513–522
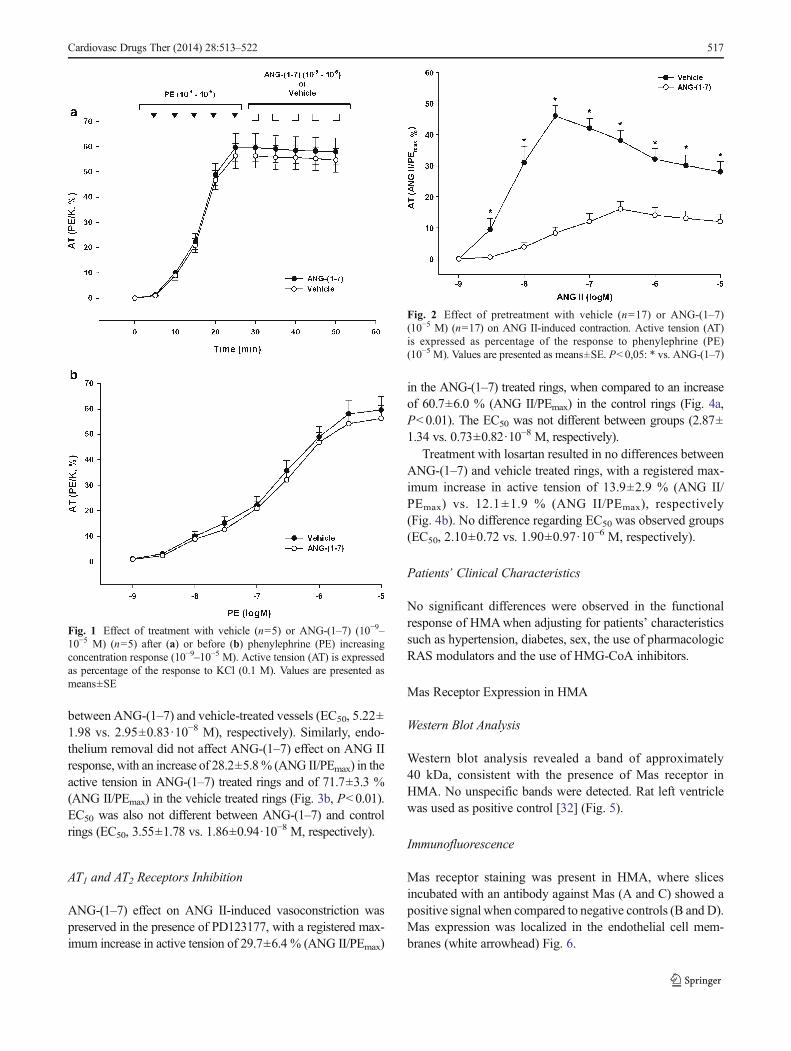
between ANG-(1–7) and vehicle-treated vessels (EC50, 5.22±1.98 vs. 2.95±0.83·10−8 M), respectively). Similarly, endo-thelium removal did not affect ANG-(1–7) effect on ANG IIresponse, with an increase of 28.2±5.8% (ANG II/PEmax) in theactive tension in ANG-(1–7) treated rings and of 71.7±3.3 %(ANG II/PEmax) in the vehicle treated rings (Fig. 3b, P<0.01).EC50 was also not different between ANG-(1–7) and controlrings (EC50, 3.55±1.78 vs. 1.86±0.94·10
−8 M, respectively).
AT1 and AT2 Receptors Inhibition
ANG-(1–7) effect on ANG II-induced vasoconstriction waspreserved in the presence of PD123177, with a registered max-imum increase in active tension of 29.7±6.4 % (ANG II/PEmax)
in the ANG-(1–7) treated rings, when compared to an increaseof 60.7±6.0 % (ANG II/PEmax) in the control rings (Fig. 4a,P<0.01). The EC50 was not different between groups (2.87±1.34 vs. 0.73±0.82·10−8 M, respectively).
Treatment with losartan resulted in no differences betweenANG-(1–7) and vehicle treated rings, with a registered max-imum increase in active tension of 13.9±2.9 % (ANG II/PEmax) vs. 12.1±1.9 % (ANG II/PEmax), respectively(Fig. 4b). No difference regarding EC50 was observed groups(EC50, 2.10±0.72 vs. 1.90±0.97·10−6 M, respectively).
Patients’ Clinical Characteristics
No significant differences were observed in the functionalresponse of HMAwhen adjusting for patients’ characteristicssuch as hypertension, diabetes, sex, the use of pharmacologicRAS modulators and the use of HMG-CoA inhibitors.
Mas Receptor Expression in HMA
Western Blot Analysis
Western blot analysis revealed a band of approximately40 kDa, consistent with the presence of Mas receptor inHMA. No unspecific bands were detected. Rat left ventriclewas used as positive control [32] (Fig. 5).
Immunofluorescence
Mas receptor staining was present in HMA, where slicesincubated with an antibody against Mas (A and C) showed apositive signal when compared to negative controls (B and D).Mas expression was localized in the endothelial cell mem-branes (white arrowhead) Fig. 6.
Fig. 1 Effect of treatment with vehicle (n=5) or ANG-(1–7) (10−9–10−5 M) (n=5) after (a) or before (b) phenylephrine (PE) increasingconcentration response (10−9–10−5 M). Active tension (AT) is expressedas percentage of the response to KCl (0.1 M). Values are presented asmeans±SE
Fig. 2 Effect of pretreatment with vehicle (n=17) or ANG-(1–7)(10−5 M) (n=17) on ANG II-induced contraction. Active tension (AT)is expressed as percentage of the response to phenylephrine (PE)(10−5 M). Values are presented as means±SE. P<0,05: * vs. ANG-(1–7)
Cardiovasc Drugs Ther (2014) 28:513–522 517

Discussion
The present study provided a new insight in the role ofANG-(1–7) in the modulation of vascular tone in humanpatients, which seems to have distinct features from whathas been described in animal models. We studied the vasodi-lator role of ANG-(1–7) in HMA, which is a vascular territoryof significant clinical importance, particularly in coronaryrevascularization surgery. Despite its relevance, the role ofnew RAS peptides in vasomotor control is not well knownin this vascular territory. We observed that ANG-(1–7) signif-icantly attenuates ANG II-induced vasoconstriction. Althoughwe did not identify the specific cellular mechanism underlyingthis effect, we believe that our findings are relevant for thecharacterization of the role of ANG-(1–7) in the HMA. For thefirst time, we identified the expression of the Mas receptor inHMA and demonstrated that it is preferentially located in theendothelium. Despite its presence, we showed that ANG-(1–
7) effect in this vascular territory is independent of Masreceptor and of AT2 receptor stimulation. Additionally, themaintenance of ANG-(1–7) effect after endothelium removalsuggests that, in this vascular tissue, this heptapeptide mightact directly on smooth muscle cells.
As previously referred, ANG-(1–7) exerts important vaso-dilator effects in various experimental animal vascular terri-tories, mainly in rat, including aorta [33], renal arteries [34],mesenteric arteries [35] and cerebral arteries [36]. Moreover, aclear benefit of this heptapeptide has been observed in animalmodels of hypertension through modulation of vascular resis-tance but also due to relevant effects on kidney, heart andautonomic nervous system [37, 38]. Importantly, ANG-(1–7)benefit on blood pressure seems particularly effective in thepresence of increased ANG II activity [39]. These observa-tions led to the establishment of ANG-(1–7) as a central agentin vascular pathophysiology and hypertensive disease. How-ever, robust evidence in humans is lacking [19].
Fig. 3 Effect of ANG-(1–7) on ANG II-induced vasoconstriction in thepresence of A-779 (10−5 M) (n=12) (a) or endothelium removal (n=13)(b). In graph A, ANG-(1–7) basal effect curve was added for visualcomparison. Values are presented as means±SE. P<0,05: * vs.ANG-(1–7)
Fig. 4 Effect of ANG-(1–7) on ANG II-induced vasoconstriction in thepresence of AT2 receptor inhibitor PD123177 (10
−6 M) (n=14) (a) or AT1receptor inhibitor losartan (n=12) (b). Values are presented as means±SE. P<0,05: * vs. ANG-(1–7)
518 Cardiovasc Drugs Ther (2014) 28:513–522

Our results show that ANG-(1–7) does not exert a signif-icant vasodilator response in HMA artery rings constrictedwith phenylephrine. This lack of effect was observed irrespec-tive of the timing of ANG-(1–7) administration, before or afterphenylephrine-induced vasoconstriction. In contrast, vaso-constriction elicited by ANG II in HMA was significantlyattenuated by ANG-(1–7), decreasing the maximum activetension developed. Additionally, we have also observed aslight shift of the ANG II concentration response curve tothe right, with no significant EC50 increase. Although previ-ous studies, performed in vivo and in vitro, suggest thatANG-(1–7) attenuates ANG II-induced vasoconstriction, the-se studies were focused onMas receptor identification and didnot perform a detailed pharmacologic analysis exploring thereceptors and mechanisms possibly involved in ANG-(1–7)action [25, 24].
The lack of vasodilation induced by ANG-(1–7) onphenylephrine-constricted vessels challenges previous studieson animal models, that claim a significant direct vasodilatoreffect of this peptide [1, 9, 11, 10]. However, Ueda et al. haveinterestingly observed that ANG-(1–7) attenuates ANG II butnot norepinephrine-induced vasoconstriction after intra-arterial infusion of these peptides in human forearm [24]. This
effect was present in both normotensive and hypertensiveindividuals [24]. Additionally, Roks et al. demonstrated thatANG-(1–7) attenuates vasoconstriction elicited by ANG II inHMA, as we observed in our experiments, and also reportedthat ANG-(1–7) pre-treatment produced no effect onphenylephrine-induced contraction, although they did not ex-plore the possible molecular pathways [25]. Therefore, theseobservations combined with our results suggest that in HMAANG-(1–7) may behave in a different way from that reportedin animal models, presenting no direct vasodilator effect butexerting antagonism on ANG II-induced vasoconstriction.When performing vascular function studies we should notethat the strength of contraction of smooth muscle cells maylimit the response to vasodilator agents. Addressing this issue, inmesenteric rat arteries, Dora et al. observed that the degree ofstimulation with phenylephrine influenced the ability of extra-cellular K+ to evoke smooth muscle hyperpolarization and relax-ation [40]. In our work, ANG-(1–7) effect on phenylephrine-induced contraction was evaluated following two different ap-proaches. In a first protocol, vessels were incubated withANG-(1–7) after a maximum contraction elicited by phenyleph-rine (10−5 M). We did not test ANG-(1–7) at a lower concentra-tion of phenylephrine, however, in a subsequent experimental
Fig. 5 Detection of the Mas receptor in HMA. Western Blot analysisrevealed the presence of Mas receptor in HMA from 3 randomly selectedpatients (in duplicate), as represented by a ~40 kDa protein band. Rat left
ventricle was used as a positive control. HMA: human mammary artery;LV: left ventricle
DAPIMas
b
d
Fig. 6 Detection of the Mas receptor in HMA. Immunofluorescencestaining of HMA slides revealed the presence of Mas receptor in theendothelial layer (arrowhead). A – slides incubatedwith primary antibody
against Mas receptor; B – negative control; C and D – digitally zoomedrepresentatives of A and B, respectively
Cardiovasc Drugs Ther (2014) 28:513–522 519

protocol, when ANG-(1–7) was used in a pretreatment setting,a complete concentration-response curve to phenylephrine(from 10−9 to 10−5 M) was made and no significant differencewas found between ANG-(1–7) pretreated HMA rings andcontrol ones, regarding any level of stimulation. This suggeststhat ANG-(1–7) does not exert a significant effect at a lowerstate of smooth muscle cell depolarization.
According to previous studies on animal models, the Masreceptor is considered the most important membrane receptormediating ANG-(1–7) effects [1, 12]. Different authors havereported that treatment with the specific Mas receptor antago-nist A-779 significantly reduces ANG-(1–7) vasodilator effect[41, 15, 35]. In addition other authors have nicely demonstratedthe importance of this receptor in ANG-(1–7) functional re-sponse in Mas-deficient animal models [14]. Our results, how-ever, raise some concerns regarding this functional effect inHMA tissue. Althoughwe have observed, to our knowledge forthe first time, that Mas receptor is expressed in HMA, func-tional experiments showed that A-779 pre-incubation did notabolish nor attenuate ANG-(1–7) action on ANG II vasocon-strictor response. This apparent lack of effect despite its tissueexpression raises interesting questions about the role of theMasreceptor in HMA. Although its central role in ANG-(1–7)signaling has been clearly described in different animal vascu-lar territories [12, 4, 1, 42], the functional importance of theMas receptor in human vessels is poorly characterized. There-fore, more studies focused on other human vascular territoriestesting the hypothesis stated in our work would be important.Additionally, Peiró et al. highlighted a Mas receptor and endo-thelium mutually dependent effect [14]. However, our resultsshow that, in this vascular territory, ANG-(1–7) modulation ofangiotensin II vasoconstriction is preserved even after endothe-lium removal, as discussed below.
Recently, new Mas-related genes (Mrg) have been ex-plored, many of them coding orphan receptors [43, 3].Alamandine is a heptapetide, structurally and functionallysimilar to ANG-(1–7), with a N-terminal Ala marking thedifference between the two [44]. It is mainly formed fromangiotensin A by the catalytic enzyme ACE-2 [44]. Interest-ingly, alamandine-induced vasodilation is not abolished byA-779 but by a Mrg type D (MrgD) inhibitor (D-Pro-Ang(1–7)), suggesting that this membrane receptor may specifi-cally mediate alamandine effects in a NO dependent pathway[3, 44]. Additionally, some studies have shown that in certaincircumstances ANG-(1–7) effects are not blocked by A-779,but are nonetheless endothelium-dependent [44, 45]. Accord-ingly, Herath et al. reported that L-NAME treatment signifi-cantly attenuated ANG-(1–7) action on ANG II induced-vasoconstriction in rat portal circulation [46]. Although thisinformation may lead us to speculate that other endothelialreceptors besides Mas, as MrgD receptor, may be involved inANG-(1–7) signaling in HMA and despite we have not testedD-Pro-ANG-(1–7) in our experiments, we demonstrated that
mechanical endothelium removal did not compromiseANG-(1–7) effect, which reinforces that other unidentifiedmechanisms, probably independent from the endothelium,are responsible for modulating ANG II-induced contractionin HMA. In addition, considering that the Mas receptor isprimarily expressed in the endothelium, as we observed in thiswork, the maintenance of ANG-(1–7) effect on ANG II-induced vasoconstriction after the endothelium removal fur-ther supports our finding and corroborates the fact that thisreceptor does not mediate the acute vascular response toANG-(1–7) in HMA. According to our results, the AT2 recep-tor is also not involved in ANG-(1–7) effect mediation inHMA, since ANG II concentration-response curve is pre-served in the presence of the specific AT2 receptor inhibitorPD123177. Indeed, as referred above, the only study testing theAT2 receptor in HMA showed that a partial AT2 receptor agonist(CGP42112A) did not elicit any vasodilator response [22] inopposition to information derived from animal models [20].
As stated above, ANG-(1–7) effects on HMA demonstrat-ed in our work challenge previous reports in different animalmodels, particularly in rat. This apparent discrepancy betweenspecies is quite interesting but not new [47]. In fact, severalinterspecies differences regarding RAS peptides and receptorsare well documented in literature [47]. Hollenberg et al. dem-onstrated in rabbit that the increase in renal blood flow in-duced by angiotensin-converting-enzyme inhibition is primar-ily dependent on ANG II concentration reduction [48], where-as, in rat, this effect is orchestrated by endothelial mediators,including bradykinin and NO [49]. This observation suggeststhat rabbit renal perfusion is more dependent on the absence ofAT1 receptor stimulation rather than direct vasodilation. Al-though there are no similar functional studies performed onhuman renal vasculature, the available data suggests that it iscloser to rabbit renal vessels [47]. Regarding other territories,Mahon et al. showed that ANG-(1–7) attenuated ANG-IIvasopressor response in rabbit aorta but had no direct hypo-tensive effect [50], which contrasts with studies done in rat[51]. Therefore, there are significant differences between spe-cies regarding the role of RAS in vascular function. In HMA,our findings are consistent with a specific antagonism ofANG-(1–7) upon ANG II pathway, independent from theMas receptor and the endothelium, with no direct vasodilatoreffect, which resembles the response described in rabbit [50].Considering that ANG-II and phenylephrine share many in-tracellular pathways involved in activation of the contractilemachinery of smooth muscle cells and are dependent on Gq
protein coupled receptors (i.e. AT1 and alpha1-adrenergic re-ceptors) [52, 53], the lack of ANG-(1–7)-elicited vasodilationin vessels constricted with phenylephrine further supports thatANG-(1–7) mediated antagonism is specifically exerted uponANG II-induced vasoconstriction in HMA. Thus, the inhibi-tion of the AT1 receptor present on smooth muscle cells byANG-(1–7) constitutes a plausible hypothesis to explain
520 Cardiovasc Drugs Ther (2014) 28:513–522

HMA response to this heptapetide and, therefore, more studiestargeting human vessels, including HMA, would be importantto clarify this effect.
In conclusion, the present study demonstrates that acutetreatment with ANG-(1–7) attenuates ANG II-induced vaso-constriction in HMA from patients submitted to coronaryrevascularization surgery. We found that the Mas receptor isexpressed in HMA. However, ANG-(1–7) effect remainedunaltered after its pharmacologic antagonism. Additionally,this effect was also independent of the AT2 receptor andendothelium signaling. Considering the role of RAS in cardio-vascular pathophysiology, the study of its mediators’ physio-logic effects in human vascular tissue is a very important stepto define new and better therapeutic strategies.
Disclosures The authors declare no conflicts of interest.
References
1. Castro-Chaves P, Cerqueira R, Pintalhao M, Leite-Moreira AF. Newpathways of the renin-angiotensin system: the role of ACE2 incardiovascular pathophysiology and therapy. Expert Opin TherTargets. 2010;14(5):485–96.
2. Genest J. Angiotensin, aldosterone and human arterial hypertension.Can Med Assoc J. 1961;84:403–19.
3. Carey RM. Newly discovered components and actions of the renin-angiotensin system. Hypertension. 2013;62(5):818–22.
4. IwaiM, HoriuchiM. Devil and angel in the renin-angiotensin system:ACE-angiotensin II-AT1 receptor axis vs. ACE2-angiotensin-(1–7)-Mas receptor axis. Hypertens Res. 2009;32(7):533–6.
5. Strawn WB, Ferrario CM, Tallant EA. Angiotensin-(1–7) reducessmooth muscle growth after vascular injury. Hypertension.1999;33(1 Pt 2):207–11.
6. Wang L, Hu X, Zhang W, Tian F. Angiotensin (1–7) amelioratesangiotensin II-induced inflammation by inhibiting LOX-1 expres-sion. Inflamm Res. 2013;62(2):219–28.
7. WangY, Tikellis C, ThomasMC, Golledge J. Angiotensin convertingenzyme 2 and atherosclerosis. Atherosclerosis. 2013;226(1):3–8.
8. Jawien J, Toton-Zuranska J, Gajda M, Niepsuj A, Gebska A, Kus K,et al. Angiotensin-(1–7) receptor Mas agonist ameliorates progress ofatherosclerosis in apoE-knockout mice. J Physiol Pharmacol.2012;63(1):77–85.
9. Brosnihan KB, Li P, Ferrario CM. Angiotensin-(1–7) dilates caninecoronary arteries through kinins and nitric oxide. Hypertension.1996;27(3 Pt 2):523–8.
10. Gorelik G, Carbini LA, Scicli AG. Angiotensin 1–7 inducesbradykinin-mediated relaxation in porcine coronary artery. JPharmacol Exp Ther. 1998;286(1):403–10.
11. Brosnihan KB, Li P, Tallant EA, Ferrario CM. Angiotensin-(1–7): anovel vasodilator of the coronary circulation. Biol Res. 1998;31(3):227–34.
12. Santos RA, Silva e Simoes AC,Maric C, Silva DM,Machado RP, deBuhr I, et al. Angiotensin-(1–7) is an endogenous ligand for the Gprotein-coupled receptor Mas. Proc Natl Acad Sci U S A.2003;100(14):8258–63.
13. Santos RA, Ferreira AJ. Angiotensin-(1–7) and the renin-angiotensinsystem. Curr Opin Nephrol Hypertens. 2007;16(2):122–8.
14. Peiro C, Vallejo S, Gembardt F, Azcutia V, Heringer-Walther S,Rodriguez-Manas L, et al. Endothelial dysfunction through genetic
deletion or inhibition of the G protein-coupled receptor Mas: a newtarget to improve endothelial function. J Hypertens. 2007;25(12):2421–5.
15. Sampaio WO, Santos dos Souza RA, Faria-Silva R, da MataMachado LT, Schiffrin EL, Touyz RM. Angiotensin-(1–7) throughreceptor Mas mediates endothelial nitric oxide synthase activationvia Akt-dependent pathways. Hypertension. 2007;49(1):185–92.
16. Chen Z, Tang Y, Yang Z, Liu S, Liu Y, Li Y, et al. Endothelin-1downregulates Mas receptor expression in human cardiomyocytes.Mol Med. 2013;8(3):871–6.
17. Bernardi S, Zennaro C, Palmisano S, Velkoska E, Sabato N, ToffoliB, et al. Characterization and significance of ACE2 andMas receptorin human colon adenocarcinoma. J Renin-Angiotensin-AldosteroneSyst. 2012;13(1):202–9.
18. Reis AB, Araujo FC, Pereira VM, Dos Reis AM, Santos RA, ReisFM. Angiotensin (1–7) and its receptor Mas are expressed in thehuman testis: implications for male infertility. J Mol Histol.2010;41(1):75–80.
19. Santos RA. Angiotensin-(1–7). Hypertension. 2014;63(6):1138–47.20. Castro CH, Santos RA, Ferreira AJ, Bader M, Alenina N, Almeida
AP. Evidence for a functional interaction of the angiotensin-(1–7)receptor Mas with AT1 and AT2 receptors in the mouse heart.Hypertension. 2005;46(4):937–42.
21. Souza AP, Sobrinho DB, Almeida JF, Alves GM,Macedo LM, PortoJE, et al. Angiotensin II type 1 receptor blockade restores angioten-sin-(1–7)-induced coronary vasodilation in hypertrophic rat hearts.Clin Sci (Lond). 2013;125(9):449–59.
22. van de Wal RM, van der Harst P, Wagenaar LJ, Wassmann S,Morshuis WJ, Nickenig G, et al. Angiotensin II type 2 receptorvasoactivity in internal mammary arteries of patients with coronaryartery disease. J Cardiovasc Pharmacol. 2007;50(4):372–9.
23. Clark MA, Diz DI, Tallant EA. Angiotensin-(1–7) downregulates theangiotensin II type 1 receptor in vascular smooth muscle cells.Hypertension. 2001;37(4):1141–6.
24. Ueda S, Masumori-Maemoto S, Ashino K, Nagahara T, Gotoh E,Umemura S, et al. Angiotensin-(1–7) attenuates vasoconstrictionevoked by angiotensin II but not by noradrenaline in man.Hypertension. 2000;35(4):998–1001.
25. Roks AJ, van Geel PP, Pinto YM, Buikema H, Henning RH, deZeeuwD, et al. Angiotensin-(1–7) is a modulator of the human renin-angiotensin system. Hypertension. 1999;34(2):296–301.
26. Grapow MT, Reineke DC, Kern T, Muller-Schweinitzer E, Carrel T,Eckstein FS. Human internal thoracic arteries from diabetic patientsare resistant to endothelial dysfunction. Fundam Clin Pharmacol.2009;23(5):567–72.
27. Nikkari ST, Sisto T, Nikkari T. Ultrastructural, immunochemical andelectrophoretic study of smooth muscle cells in internal mammaryarteries of patients undergoing coronary bypass surgery.Atherosclerosis.1989;79(2–3):129–38.
28. Zulli A, Hare DL, HorriganM, Buxton BF. The resistance of the IMAto atherosclerosis might be associated with its higher eNOS, ACE andET-A receptor immunoreactivity. Arterioscler Thromb Vasc Biol.2003;23(7):1308.
29. Buikema H, Grandjean JG, van den Broek S, van Gilst WH, Lie KI,Wesseling H. Differences in vasomotor control between humangastroepiploic and left internal mammary artery. Circulation.1992;86(5 Suppl):II205–9.
30. Martinez-Revelles S, Jimenez-Altayo F, Caracuel L, Perez-AsensioFJ, Planas AM, Vila E. Endothelial dysfunction in rat mesentericresistance artery after transient middle cerebral artery occlusion. JPharmacol Exp Ther. 2008;325(2):363–9.
31. MuggeA, BartonMR, Cremer J, Frombach R, Lichtlen PR. Differentvascular reactivity of human internal mammary and inferior epigas-tric arteries in vitro. Ann Thorac Surg. 1993;56(5):1085–9.
32. Filho AG, Ferreira AJ, Santos SH, Neves SR, Silva Camargos ER,Becker LK, et al. Selective increase of angiotensin (1–7) and its
Cardiovasc Drugs Ther (2014) 28:513–522 521

receptor in hearts of spontaneously hypertensive rats subjected tophysical training. Exp Physiol. 2008;93(5):589–98.
33. Silva DM, Gomes-Filho A, Olivon VC, Santos TM, Becker LK,Santos RA, et al. Swimming training improves the vasodilator effectof angiotensin-(1–7) in the aorta of spontaneously hypertensive rat. JAppl Physiol. 2011;111(5):1272–7.
34. Nematbakhsh M, Safari T. Role of Mas receptor in renal blood flowresponse to angiotensin (1–7) in male and female rats. Generalphysiology and biophysics. 2014.
35. Raffai G, Durand MJ, Lombard JH. Acute and chronic angiotensin-(1–7) restores vasodilation and reduces oxidative stress in mesentericarteries of salt-fed rats. Am J Physiol Heart Circ Physiol.2011;301(4):H1341–52.
36. Durand MJ, Raffai G, Weinberg BD, Lombard JH. Angiotensin-(1–7) and low-dose angiotensin II infusion reverse salt-induced endo-thelial dysfunction via different mechanisms in rat middle cerebralarteries. Am J Physiol Heart Circ Physiol. 2010;299(4):H1024–33.
37. Benter IF, Ferrario CM, Morris M, Diz DI. Antihypertensive actionsof angiotensin-(1–7) in spontaneously hypertensive rats. Am JPhysiol. 1995;269(1 Pt 2):H313–9.
38. Stegbauer J, Oberhauser V, Vonend O, Rump LC. Angiotensin-(1–7)modulates vascular resistance and sympathetic neurotransmission inkidneys of spontaneously hypertensive rats. Cardiovasc Res.2004;61(2):352–9.
39. Ferrario CM. Angiotension-(1–7) and antihypertensive mechanisms.Am J Nephrol. 1998;11(6):278–83.
40. Dora KA, Garland CJ. Properties of smooth muscle hyperpolariza-tion and relaxation to K+in the rat isolated mesenteric artery. Am JPhysiol Heart Circ Physiol. 2001;280(6):H2424–9.
41. Lemos VS, Silva DM, Walther T, Alenina N, Bader M, Santos RA.The endothelium-dependent vasodilator effect of the nonpeptide Ang(1–7) mimic AVE 0991 is abolished in the aorta of mas-knockoutmice. J Cardiovasc Pharmacol. 2005;46(3):274–9.
42. Santos RA, Ferreira AJ. Pharmacological effects of AVE 0991, anonpeptide angiotensin-(1–7) receptor agonist. Cardiovasc DrugRev. 2006;24(3–4):239–46.
43. Gembardt F, Grajewski S, Vahl M, Schultheiss HP, Walther T.Angiotensin metabolites can stimulate receptors of the Mas-relatedgenes family. Mol Cell Biochem. 2008;319(1–2):115–23.
44. Lautner RQ, Villela DC, Fraga-Silva RA, Silva N, Verano-Braga T,Costa-Fraga F, et al. Discovery and characterization of alamandine: anovel component of the renin-angiotensin system. Circ Res.2013;112(8):1104–11.
45. Silva DM, Vianna HR, Cortes SF, Campagnole-Santos MJ, SantosRA, Lemos VS. Evidence for a new angiotensin-(1–7) receptorsubtype in the aorta of Sprague–Dawley rats. Peptides. 2007;28(3):702–7.
46. Herath CB, Mak K, Burrell LM, Angus PW. Angiotensin-(1–7)reduces the portal pressure response to angiotensin II andmethoxamine via an endothelial nitric oxide mediated pathway incirrhotic rat liver. Am J Physiol Gastrointest Liver Physiol. 2012.
47. Hollenberg NK. Implications of species difference for clinical inves-tigation: studies on the renin-angiotensin system. Hypertension.2000;35(1 Pt 2):150–4.
48. Hollenberg NK, Passan DR. Specificity of renal vasodilation withcaptopril: saralasin prevents the response in the DOCA-treated, salt-loaded rabbit. Life Sci. 1982;31(4):329–34.
49. Nakagawa M, Stewart JM, Vavrek RJ, Nasjletti A. Effects of a kininantagonist on renal function in rats. Am J Physiol. 1990;258(3 Pt 2):F643–8.
50. Mahon JM, Carr RD, Nicol AK, Henderson IW. Angiotensin (1–7) isan antagonist at the type 1 angiotensin II receptor. J Hypertens.1994;12(12):1377–81.
51. Sampaio WO, Nascimento AA, Santos RA. Systemic and regionalhemodynamic effects of angiotensin-(1–7) in rats. Am J PhysiolHeart Circ Physiol. 2003;284(6):H1985–94.
52. Unger T, Chung O, Csikos T, Culman J, Gallinat S, Gohlke P,et al. Angiotensin receptors. J Hypertens Suppl. 1996;14(5):S95–103.
53. Graham RM, Perez DM, Hwa J, Piascik MT. Alpha 1-adrenergicreceptor subtypes. Molecular structure, function, and signaling. CircRes. 1996;78(5):737–49.
522 Cardiovasc Drugs Ther (2014) 28:513–522
Copyright © 2022 FDOKUMEN




















