
Characterization of an epithelial cell line from bovine mammary gland
11
In VitroCell.Dev.Biol.--Animal 38:282-292, May 2002 2002 Society for In VitroBiology 1071-2690/02 $10.00+0.00 CHARACTERIZATION OF AN EPITHELIAL CELL LINE FROM BOVINE MAMMARY GLAND TANIA GERMAN aND ITAMAR BARASH ~ Institute of Animal Scierwe, ARO, The I/olcani Center, P.O. Box 6, Bet-Dagan, Israel (Received 11 February 2002; accepted 9 July 2002) SUMMARY Elucidation of the bovine mammary gland's unique characteristics depends on obtaining an authentic cell line that will reproduce its function in vitro. Representative clones from bovine mammary cell populations, differing in their attachment capabilities, were cultured. L-1 cells showed strong attachment to the plate, whereas H-7 cells detached easily. Cultures established from these clones were nontumorigenic upon transplantation to an immunodeficient host; they exhibited the epithelial cell characteristics of positive cytokeratin but not smooth muscle actin staining. Both cell lines depended on fetal calf serum for proliferation. They exhibited distinct levels of differentiation on Matrigel in sermu-free, insulin-supplemented medimn on the basis of their organization and [3-1actoglobulin (BLG) secretion. H-7 cells organized into mammospheres, whereas L-1 ceils arrested in a duct-like morphology. In both cell lines, prolactin activated phos- phorylation of the signal transducer and activator of transcription, Stat5--a regulator of milk protein gene transcription, and of PHAS-I--an inhibitor of translation initiation in its nonphosphorylated form. De novo synthesis and secretion of BLG were detected in differentiated cultures: in L-1 cells, BLG was dependent on lactogenic hormones for maximal induction but was less stringently controlled than was [3-casein in the mouse CID-9 cell line. L-1 cells also encompassed a near-diploid chromosomal karyotype and may serve as a tool for studying functional characteristics of the bovine mammary gland. Key words: [3-1actoglobulin; Stat5; PHAS-I; cytokeratins. INTRODUCTION The key nutritional role, and thus the commercial value, of milk has triggered numerous attempts to dissect the mechanisms con- trolling mammary metabolism. Milk protein synthesis by mammary epithelial cells is a complicated process. Polypeptide and steroid hormones as well as interactions with stroma cells and extracellular matrix (ECM) proteins control mammary cell differentiation. Rodent models enable convenient access to the glands and an opportunity to nlanipulate the cellular environment in vivo. The efficient milk- protein synthesis and the tight hormonal responsiveness of HCll (Ball et al., 1988) and CID-9 (Schmidhauser et al., 1990) mouse mammapy cell lines have attracted in vitro research with these es- tablished models. Ahhough the development and metabolism of the mouse and bo- vine manmmry glands are essentially comparable, various interspe- cies differences were noted. A higher level of protein in the rodent's milk and the lack of [3-1actoglobulin (BLG) expression in the ro- dent's gland have been well documented. The role of the Jak2-Stat5 pathway, the main route for lactogenic hormonal signaling in ro- dents, in the bovine gland is uncertain (Wheeler et al., 1997, 2001; Yang et al., 2000a, 2000b). Likewise, involution of the ruminant mammary gland is actually less extensive than that of the rodent (Wilde et al., 1999). Cell culture is a convenient tool for dissecting metabolic pro- 1To whom con'espondence should be addressed at E-mail: barashi@agri. huji.ac.il cesses at the cellular and molecular levels. The utility of a func- tionally representative bovine mammary cell line has become even more relevant with the recent progress achieved in the bovine ge- nome project (Piedrahita, 2000), because it could serve as a model for relating specific functions to many of the newly discovered genes. Epithelial cells isolated from the lactating bovine mammary gland were cultured on plastic dishes but lost their tissue-specific func- tions within d (Ebner et al., 1961) or after cryopreservation (MacKenzie et al., 1982). Indeed, maintaining functional bovine mammary cells in culture has proven to be a formidable challenge, and only a few cell lines have been established. By far, the one most used is MAC-T (Huynh et al., 1991), which was immortalized by transfection of mammary cells with the tsA mutant of the SV 40 gene. Although this cell line maintains a certain level of lactogenic hormone responsiveness for milk-protein expression, it has been used mainly to study IGF-I metabolism (Romagnolo et al., 1993, 1994; Woodward et al., 1996; Robinson et al., 2000a, 2000b) and, more recently, the effect of the cap-dependent ribonucleic acid (RNA) helicase complex, eIF4E, on proliferation (Long et al., 2001). Other bovine cell lines have also been generated, but only one, BME-UV1, maintains established, but modest, casein synthesis in medium containing fetal calf serum (FCS; Zavizion et al., 1996). Like MAC-T this cell line gained immortality after transfection with the SV 40 large T antigen. We describe herein the isolation and characterization of two new cell lines generated from spontaneously immortalized epithelial 282
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Characterization of an epithelial cell line from bovine mammary gland

In Vitro Cell. Dev. Biol.--Animal 38:282-292, May 2002 �9 2002 Society for In Vitro Biology 1071-2690/02 $10.00+0.00
CHARACTERIZATION OF AN EPITHELIAL CELL LINE FROM BOVINE MAMMARY GLAND
TANIA GERMAN aND ITAMAR BARASH ~
Institute of Animal Scierwe, ARO, The I/olcani Center, P.O. Box 6, Bet-Dagan, Israel
(Received 11 February 2002; accepted 9 July 2002)
SUMMARY
Elucidation of the bovine mammary gland's unique characteristics depends on obtaining an authentic cell line that will reproduce its function in vitro. Representative clones from bovine mammary cell populations, differing in their attachment capabilities, were cultured. L-1 cells showed strong attachment to the plate, whereas H-7 cells detached easily. Cultures established from these clones were nontumorigenic upon transplantation to an immunodeficient host; they exhibited the epithelial cell characteristics of positive cytokeratin but not smooth muscle actin staining. Both cell lines depended on fetal calf serum for proliferation. They exhibited distinct levels of differentiation on Matrigel in sermu-free, insulin-supplemented medimn on the basis of their organization and [3-1actoglobulin (BLG) secretion. H-7 cells organized into mammospheres, whereas L-1 ceils arrested in a duct-like morphology. In both cell lines, prolactin activated phos- phorylation of the signal transducer and activator of transcription, Stat5--a regulator of milk protein gene transcription, and of PHAS-I--an inhibitor of translation initiation in its nonphosphorylated form. De novo synthesis and secretion of BLG were detected in differentiated cultures: in L-1 cells, BLG was dependent on lactogenic hormones for maximal induction but was less stringently controlled than was [3-casein in the mouse CID-9 cell line. L-1 cells also encompassed a near-diploid chromosomal karyotype and may serve as a tool for studying functional characteristics of the bovine mammary gland.
Key words: [3-1actoglobulin; Stat5; PHAS-I; cytokeratins.
INTRODUCTION
The key nutritional role, and thus the commercial value, of milk has triggered numerous attempts to dissect the mechanisms con- trolling mammary metabolism. Milk protein synthesis by mammary epithelial cells is a complicated process. Polypeptide and steroid hormones as well as interactions with stroma cells and extracellular matrix (ECM) proteins control mammary cell differentiation. Rodent models enable convenient access to the glands and an opportunity to nlanipulate the cellular environment in vivo. The efficient milk- protein synthesis and the tight hormonal responsiveness of H C l l (Ball et al., 1988) and CID-9 (Schmidhauser et al., 1990) mouse mammapy cell lines have attracted in vitro research with these es- tablished models.
Ahhough the development and metabolism of the mouse and bo- vine manmmry glands are essentially comparable, various interspe- cies differences were noted. A higher level of protein in the rodent's milk and the lack of [3-1actoglobulin (BLG) expression in the ro- dent's gland have been well documented. The role of the Jak2-Stat5 pathway, the main route for lactogenic hormonal signaling in ro- dents, in the bovine gland is uncertain (Wheeler et al., 1997, 2001; Yang et al., 2000a, 2000b). Likewise, involution of the ruminant mammary gland is actually less extensive than that of the rodent (Wilde et al., 1999).
Cell culture is a convenient tool for dissecting metabolic pro-
1To whom con'espondence should be addressed at E-mail: barashi@agri. huji.ac.il
cesses at the cellular and molecular levels. The utility of a func- tionally representative bovine mammary cell line has become even more relevant with the recent progress achieved in the bovine ge- nome project (Piedrahita, 2000), because it could serve as a model for relating specific functions to many of the newly discovered
genes. Epithelial cells isolated from the lactating bovine mammary gland
were cultured on plastic dishes but lost their tissue-specific func- tions within d (Ebner et al., 1961) or after cryopreservation (MacKenzie et al., 1982). Indeed, maintaining functional bovine mammary cells in culture has proven to be a formidable challenge, and only a few cell lines have been established. By far, the one most used is MAC-T (Huynh et al., 1991), which was immortalized by transfection of mammary cells with the tsA mutant of the SV 40 gene. Although this cell line maintains a certain level of lactogenic hormone responsiveness for milk-protein expression, it has been used mainly to study IGF-I metabolism (Romagnolo et al., 1993,
1994; Woodward et al., 1996; Robinson et al., 2000a, 2000b) and, more recently, the effect of the cap-dependent ribonucleic acid (RNA) helicase complex, eIF4E, on proliferation (Long et al., 2001). Other bovine cell lines have also been generated, but only one, BME-UV1, maintains established, but modest, casein synthesis in medium containing fetal calf serum (FCS; Zavizion et al., 1996). Like MAC-T this cell line gained immortality after transfection with
the SV 40 large T antigen. We describe herein the isolation and characterization of two new
cell lines generated from spontaneously immortalized epithelial
282

MAMMARY EPITHELIAL CELL LINE 283
cells of the lactating bovine maminary gland. When cultured in serum-free medimn on the laminin-rich matrix, Matrigel, these lines
acquire distinct mammary gland morphology, retain epithelial char- aeteristies, and express the milk protein BLG. In the presence of insulin, prolaetin (PRL) activates key regulators of the transcrip-
tional and translational machineries. L-I cells were responsive to
laetogenie hormones and acquired a nearly diploid chromosomal
number. This cell line may be useful for functional studies of the bovine mammary gland.
MATERIALS AND METHODS
Isolation of bovine mammary cell lines. Two biopsies from the mammary parenchyma tissue of a Friesian cow in its second day of lactation served as the source for epithelial cells. Both biopsies were taken from the left frontal quarter of the udder. The tissues were pooled, finely minced, and the mam- mary epithelial cells recovered by trypsin-collagenase digestion and differ- ential centrifugation as previously described (Ilan et al., 1998). Cells were plated on a 35-mm plastic tissue culture dish (Coruing, NY) in Dulbecco modified Eagle medium (DMEM)-F-12 medium (GIBCO BRL, Gaithersburg, MD) supplemented with 10% FCS (Sigma Chemical Co., St. Louis, MO), insulin (5 txg/lnl, Sigma), hydrocortisone (1 txg/ml, Sigma), mouse epidermal growth hormone (10 ng/ml, Sigma), gentamycin (50 txg/ml, Sigma), penicillin (100 U/ml, BioLabs, Jerusalem, Israel), and streptomycin (0.1 mg/ml, Bio- Labs). The cells were not passaged for 2 wk. Plating efficiency was low, but a few of the adherent ceils proliferated and reached subconfluency after 2 - 3 wk. Cells were trypsinized and replated on 60-ram plates. After 8-10 pas- sages, two subpopulations of the cells were identified. Luminal epithelial cells were seen as islets, and the lnyoepithelial cells appeased to "stream" around these islands (Pechoux et at., 1999). The cultures were further enriched with luminal cells by three cycles of plating the cells on Matrigel for 48 h and then on plastic for the same period. Cells were separated ti'om the Matrigel by trypsinization. Finally, about 10 wk after the initial plating, two popula- tions of mammary luminal ceils were established according to their resistance to trypsinization. Population "H" encompassed ceils that detached easily from the plate. Population "L" was composed of ceils with strong resistance to t~ypsin.
For cell cloning, about 100 cells were cultured in a 100-ram dish. Single cells were identified and marked. Colonies originating from a single cell were lifted with Whatman paper (0.4-cm diameter) soaked in trypsin. Each colony was allocated to a single well in a 24-multiwell culture dish. After overnight incubation, the cells were detached from the paper, attached to the plate, and allowed to proliferate. Subsequently, cell colonies were trypsinized and cultured individually in 35-ram plates.
It should be noted that the two initial attempts to recover nondifferentiated, immortalized cells from the mammary gland of a pregnant cow, 2-3 wk before delivery, failed at various stages although a similar methodology was used. It is cun'ently not known if this success should be atn'ibuted to the altered developmental stage of the gland or to random factor(s).
Cell differentiation and hormonal responsiveness. For hormonal activation, the cells were plated on 35-mm plates (0.8 • 106 cells/plate) in the presence of 10% FCS and 5 p~g/ml insulin. Twenty-four hours after plating, the medium was switched to 0% FCS. Unless otherwise indicated, hydroeortisone (1 txg/ml), PRL (3 /xg/ml), and growth hormone (GH, 1 txg/ml) were added in different combinations. PRL and GH were kindly provided by the NIDDK program.
Karyology: Exponentially growing cultures of L-1 and H-7 cells were in- cubated with colchicine (0.4 txg/ml) for 6 h. Cells were trypsynized and treated with hypotonic medium for 10 min. Metaphase chromosome spreads were prepared as described (Danielson et at., i984). Chromosomes of at least 40 metaphase cells were counted at cell passages 10-12.
Histdogy and immunohistochemistry; Cells on Matrigel were carefully lifted with a fine spatula and fixed in 4% (v/v) paraformaldehyde. The pieces of matrix containing cells were dehydrated in a series of 70-100% ethanols, cleared in xylene, and embedded in paraffin. Sections (5 txm) were processed for immunohistochemistl3,, essentially as described by Baruch et at. (1995). Briefly, sections were mounted onto slides and stained with anti-BLG anti- body (Ilan et al., 1998) or anti-cytokeratin (AE1, Dako, Glostrup, Denmark) diluted 1:200. Goat anti-mouse (Zymed, San Francisco, CA) served as the second antibody. BLG signals were generated using a N/Histofine kit (Ni-
ehirei Corp., Tokyo, Japan). Cytokeratin signals were generated using an LSAB2 kit (Dako), according to the manufacturer's protocol.
Metabolic labeling. H-7 and L-1 cells were cultured on Matrigel in 35-mm dishes and labeled for 4 h in 1 ml of methionine-free DMEM containing 100 txCi/ml 3~S-methionine as previously described (Barash et al., 1993). Total incorporation of 35S-methionine in the protein was measured by trichloroac- etic acid precipitation of 5-1xl aliquots pipetted onto nitrocellulose filters (Barash et al., 1993). For determination of de novo BLG synthesis, aliquots (300-500 txl) of cell lysate or medium were preabsorbed onto Protein A- Sepharose CL-4B beads (Pharmacia, Uppsala, Sweden), reacted with anti- BLG antibodies (Nordic hnmunology, Tilburg, The Netherlands; diluted 1:250), precipitated with Protein A-Sepharose, analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and exposed to XAR Kodak film after fluorography.
Detection of BLG, cytokeratins, smooth muscle actin, PHAS-I, and phos- phorylated Stat5 by immunoblotting. Cellular proteins were extracted with 10 mM Tris-HC1 pH 7.4 buffer, containing 50 mM NaC1, 5 mM ethyleuediamine- tetraacetic acid, 0.1% (w/v) bovine serum albumin, 1% (v/v) Triton, 50 mM NaF, 1 nrM Na3VO4, 1 raM" ethylene glycol-bis(aminoethylether)-tetraacetic acid, 1 nLM phenylmethane sulfonyl fluoride, and 1 mM aprotinin, and were subjected to SDS-PAGE. Fractionated proteins were transferred to nitrocel- lulose filters. For detection of BLG, filters were reacted with anti-BLG anti- bodies ([lan et al., 1998) diluted 1:1000. For detection of smooth nluscle actin (SMA), filters were reacted with anti-SMA antibodies (Sigma) diluted 1:500. For detection of phosphorylated Star5, filters were reacted with rabbit anti-mouse antibodies generated against a peptide containing Tyr 694 (Cell Signaling Technology, Beverly, MA) and diluted 1:1000. For detection of PHAS-I, filters were reacted with anti-PHAS-I antibodies (Barash, 1999). In all cases, goat anti-rabbit IgG complexed with horseradish peroxidase (HRP) (Zymed) served as the second antibody. For detection of cytokeratins, filters were reacted with anti-AE1 or anti-AE3 antibodies (Dako) diluted 1:1000. Goat anti-mouse IgG complexed with HRP (Zymed) served as the second antibody.
Transplantation of L-1 and H-7 cells into hosts. Cells were removed from 100-ram diameter plates by trypsinization, collected by centrifugation, and resuspended in serum-free medium. Cell suspensions (3 x 106 cells/0.2 ml medium) were injected under the skin of 6-wk-old immunodeficient (nude) virgin CD-1 mice in the region of glands #4 and #8.
RESULTS
Establishment of H-7 and L-1 cell lines. A heterogenous popu- lation of mammary cells was prepared by t rypsin-col lagenase di-
gestion of biopsies from the mammary gland of a lactating cow. A mixture of fibroblast-like cells, fat cells, and myoepithelial and ep-
ithelial cells was cultured. About 20 passages were devoted to en- r iching the e~lture with proliferating epithelial-like cells. Selective tpypsinization and transition of cells between plastic and laminin- rich matrix resulted in the establishment of two subpopulations,
differing in their potency of attachment to the culture plate. The bovine mammary cells of the L population were strongly attached
to the plastic plate, whereas cells of the H population showed rel- atively low resistance to trypsin treatment. The latter could reach eonflueney but did not generate domes and detached easily from the plate. No significant (P < 0.01) difference in BLG expression
between the two populations could be detected when levels of this protein were determined in euhures of selected clones of H and L cells treated with insulin, PRL, and hydroeortisone. Levels of BLG
secretion to the medium ranged between 109 -+ 21 ng/10 z cells/24 h and 221 +- 101 ng/105 eells/24 h (Fig. 1; Table 1). Slow prolif- eration rate of some of the clones and early detachment of H cells from the plate limited further comparisons between populations.
Two clones, L-1 and H-7, were selected from the two populations
on the basis of their rate of proliferation and their ability to express the BLG milk protein when cultured on Matrigel in the presence of
insulin, PRL, and hydroeortisone.
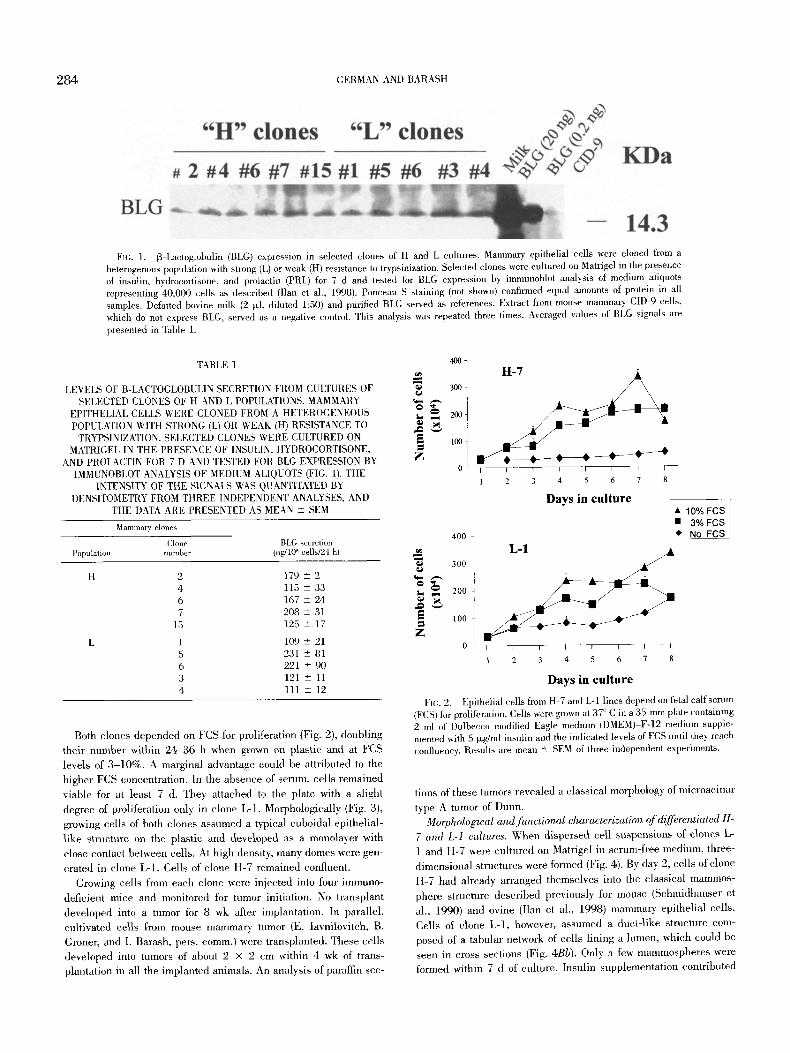
284 GERMAN AND BARASH
Fro. 1. 13-Lactoglobulin (BLG) expression in selected clones of H and L cultures. Mammary epithelial cells were cloned from a heterogenous population with strong (L) or weak (H) resistance to tx'ypsinization. Selected clones were cultured on Matrigel in the presence of insulin, hydrocortisone, and prolactin (PRL) for 7 d and tested for BLG expression by immunoblot analysis of medium aliquots representing 40,000 cells as described (Ilan et al., 1998). Ponceau S staining (not shown) confirmed equal amounts of protein in all samples. Defatted bovine milk (2 txl, diluted 1:50) and purified BLG served as references. Extract from mouse malnmary CID-9 cells, which do not express BLG, served as a negative control. This analysis was repeated three times. Averaged values of BLG signals are presented in Table 1.
TABLE 1
LEVELS OF B-LACTOGLOBULIN SECRETION FROM CULTURES OF SELECTED CLONES OF H AND L POPULATIONS. MAMMARY
EPITHELIAL CELLS WERE CLONED FROM A HETEROGENEOUS POPULATION WITH STRONG (L) OR WEAK (H) RESISTANCE TO
TRYPSINIZATION. SELECTED CLONES WERE CULTURED ON MATRIGEL IN THE PRESENCE OF INSULIN, HYDROCORTISONE,
AND PROLACTIN FOR 7 D AND TESTED FOR BLG EXPRESSION BY IMMUNOBLOT ANAL3~SIS OF MEDIUM ALIQUOTS (FIG. 1). THE
INTENSITY OF THE SIGNALS ~L~S QUANTITATED BY DENSITOMETRY FROM THREE INDEPENDENT ANALYSES, AND
THE DATA ARE PRESENTED AS MEAN -+ SEM
Mamma~ clones
Clone BLG secretion Population number (ng/l@ cells/24 h)
H 2 179 -+ 2 4 115 -+ 33 6 167 + 24 7 208 -+ 31
15 125 + 17
1 109 + 21 5 231 -+ 81 6 221 + 90 3 121 + 11 4 111 -+ 12
Both clones depended on FCS for proliferation (Fig. 2), doubling their number within 24-36 h when grown on plastic and at FCS levels of 3 -10%. A marginal advantage could be attributed to the higher FCS concentration. In the absence of serum, cells remained
viable for at least 7 d. They attached to the plate with a slight
degree of proliferation only in clone L-1. Morphologically (Fig. 3), growing ceils of both clones assumed a typical cuboidal epithelial- like structure on the plastic and developed as a monolayer with close contact between cells. At high density, many domes were gen- erated in clone L-1. Cells of clone H-7 remained confluent.
Growing cells from each clone were injected into four immuno- deficient mice and monitored for tumor initiation. No transplant developed into a tumor for 8 wk after implantation. In parallel, cultivated cells from mouse mammary tumor (E. Iavnilovitch, B. Groner, and I. Barash, pers. comm.) were transplanted. These ceils developed into tumors of about 2 • 2 cm within 4 wk of trans- plantation in all the implanted animals. An analysis of paraffin sec-
H-7 / ~
~ 20~ i ,ooi
O ~ I - I - I I I I i - -
1 2 3 4 5 6 7 8
Days in culture i ; 10~176 FcsFGS 400 �9 No FCS
' ~ 300
100
Z 0 I ~ I - I - - I I I I
l 2 3 4 5 6 7 g
Days in culture
FIG. 2. Epithelial cells from H-7 and L-1 lines depend on fetal calf serum (FCS) for proliferation. Cells were grown at 37 ~ C in a 35-ram plate containing 2 ml of Dulheceo modified Eagle medium (DMEM)-E-12 medimn supple- mented with 5 I~g/ml insulin and the indicated levels of FCS until they reach conflueney. Results are mean ~+ SEM of three independent experiments.
lions of these tumors revealed a classical morphology of microacinar
type A tumor of Dunn. Morphological and functional characterization of differentiated H-
7 and L-I cultures. When dispersed cell suspensions of clones L- 1 and H-7 were cultured on Matrigel in serum-free medium, three-
dimensional structures were formed (Fig. 4). By day 2, cells of clone H-7 had already arranged themselves into the classical mammos-
phere structure described previously for mouse (Schmidhauser et al., 1990) and ovine (Ilan et al., 1998) mammary epithelial cells. Cells of clone L- l , however, assumed a duct-like structure com- posed of a tabular network of cells lining a lumen, which could be seen in cross sections (Fig. 4Bb). Only a few mammospheres were formed within 7 d of culture. Insulin supplementation contributed

MAMMARY EPITHELIAL CELL LINE 285
FIG. 3. Dome folanation in the posteonfluent L-1 but not H-7 monolayer. H-7 (a, c) and L-1 (b, d) ceils were plated on plastic and maintained for 7 d in the absence (a, b) o1" the presence (c, d) of 10% fetal calf serum. Representative phase-contrast nficrographs of six independent cultures are presented.
to the development of unique structures by size but did not affect their type of arrangement (Fig. 4A). No further effects were seen with the addition of hydrocortisone, PRL, and GH. FCS induced structural deterioration within 1-2 d, because of overgrowth of the ceils (data not shown). Hematoxylin and eosin staining of 5-~m sections prepared from cells cultured on Matrigel exposed their in- ternal structures. Epithelial cells lined the walls of the duct-like network structures in L-1 (Fig. 4B), whereas an aggregated mass of cells, forming the mammosphere in the center of the Matrigel piece, was revealed in differentiated H-7 cultures.
It should be noted that the mammosphere structure characterized all seven tested clones of subpopulation H. Four of the seven clones tested from subpopulation L were arranged in a definite duct-like morphology, and the rest presented varying stage~ between duct- like and mammosphere morphologies.
To confirm the epithelial nature of the bovine cells, differentiated H-7 and L-1 cultures were immunostained with the AE1 anti-ey- tokeratin antibody, which specifically stains mouse (Ilan et at., 1996), ovine (Ilan et al., 1998), and here also bovine mammary lmninal cells (Fig. 5A). The AE1 anti-eytokeratin antibodies rec- ognize keratins of the acidic subfamily, whereas AE3 anti-cytoker-
atin detects keratins of the basic subfamily. Figure 5B shows im- munodetection of members from both families in the L-1 and H-7 cell cultures. The AE1 anti-cytokeratin detects keratins with mo- lecular weight of 40-56 kDa, and the AE3 anti-cytokeratin detects those of 52-65 kDa. No significant differences in the composition or amount of cytokeratins were detected between cells of clones L-
1 and H-7. Mammary luminal epithelial cells have been suggested to contain
progenitors of myoepithelial cells (Pechoux et al., 1999). These lat- ter cells surround the alveoli and the ducts, and their contraction upon oxytocin stimulation results in the removal of milk fl'om the alveoli. Myoepithelial cells are characterized by the presence of SMA. Immunodetection of SMA showed that bovine and mouse mammai 7 glands differ in the composition of actins, although a main protein of about 41 kDa was detected in the cells of both organs (Fig. 5C). In contrast, cells extracted from L-1 and H-7 cultures did not react with antibodies to SMA, suggesting a homogenous population of epithelial cells. This result was also confirmed by immunohistochemistry using the respective anti-SMA antibodies
(data not shown). Signal transduction in the bovine mammary epithelial cells. PRL

286 GERMAN AND BARASH
FIG. 4. H-7 and L-1 cells organized on Matrigel in distinct three-dimensional structures. (A) H-7 (a, b) and L-1 (c, d) cells were cultured on Matrigel in the absence (a, c) or the presence (b, d) of insulin. Mammospheres were formed in H-7 cultures, whereas L-1 cells organized in duct-like structures. Insulin induced the development of the structures but did not affect their type of organization. Representative phase-contrast micrographs of six independent cultures are presented. (B) Hematoxylin and eosin staining of 5-txm sections prepared from H-7 (a) and L-1 (b) cell cultures on Matrigel in the presence of insulin. Bar, 40 ~m. Results represent more than three independent analyses.

MAMMARY EPITHELIAL CELL LINE 287

288 GERMAN AND BARASH
is a primary regulator of milk-protein expression in the maminary gland. A main route through which it transduces its signal is the Jak2-Stat5 pathway. Phosphorylation of Stat5 by Jak2 kinase upon recruitment to the intracellular domain of the receptor induces its translocation into the nucleolus and binding to specific GAS (in- terferon-~/ activation site) DNA elements that are present in the promoters of all major milk proteins (Rosen et al., 1999). Differ- entiated H-7 and L-1 cells were cultured in serum-free medium and treated with insulin for 5 d. Addition of PRL resulted in the rapid induction of Star5 phosphorylation, peaking 15 rain after supple- mentation (Fig. 6).
PHAS-I binds the translation-initiation factor elF4E. Phosphor- ylation of PHAS-I results in its dissociation from eIF4E, allowing the latter to bind to the cap structure of eukaryotic messenger RNA (mRNA). This, in turn, enhances the association between the mRNA and the 40S ribosomal subunit and facilitates translation. We previously showed that PHAS-I phosphorylation is subject to PRL regulation in the mouse mammary epithelial cell line CID-9 (Barash, 1999). To examine the activation of this route in bovine cultures, L-1 and H-7 cells were plated on Matrigel and treated with insulin for 5 d. Cells analyzed at the end of this period ex- pressed mainly the nonphosphorylated c~ form of the PHAS-I pro- tein, accompanied by minor levels of the partly phosphorylated 13 form (Fig. 6). Within 1 min of the addition of PRL, the 13 subunit became the dominant form, and a gradual increase in the level of the fully phosphorylated ~ form was noted until the end of the analysis period.
Distinct hormonal effect on BLG secretion in H-7 and L-1 cells. BLG is a major protein in the whey fraction of ruminant milk. Figure 7 demonstrates its expression in differentiated mammary epithelial cells of clones H-7 and L-1. To study the hormonal effects on its de novo synthesis and secretion, differentiated cultures of these cell lines were treated with different combinations of insulin, PRL, and hydrocortisone for 7 d. The incubation period was terminated after 4 h of metabolic labeling with zsS-methionine. Synthesis and secre- tion of total proteins and specifically of BLG were measured during this period in cell lysate and medium, respectively.
Addition of hydocortisone or PRL, and in particular a combina- tion of the two hormones, increased BLG production in L-1 cells treated with insulin but had no effect in cultures from the H-7 clone. No further induction in BLG expression followed the addition of GH in L-1 cells (Fig. 8).
BLG is not expressed in the murine mammary gland. To evaluate hormonal responsiveness in bovine cells, 13-casein expression was measured in murine CID-9 cells exposed to conditions similar to those applied to the bovine cells (Fig. 8). These cells did not ex- press detectable levels of [3-casein in the presence of insulin. Ad- dition of PRL resulted in a very high level of 13-casein synthesis. No additional effect was observed for hydrocortisone or GH.
The contribution of hydrocortisone to milk protein synthesis was higher than its effect on total protein synthesis, especially in the
CID-9 cells. Thus, this hormone was attributed a stronger effect when milk-protein synthesis was evaluated on the basis of equal level of protein synthesis (Fig. 8B) compared with an equal amount of cells (Fig. 8A).
Chromosomal analysis. Normal diploid bovine cells contain 60 chromosomes. During transformation, the number of chromosomes
is frequently altered. Thus, the chromosomal profile of L-1 and H- 7 cells was determined during proliferation, and significant differ- ences were seen between the two clones. Average counts of 40 chro- mosome spreads showed a nearly normal profile for the L-1 cells (Fig. 9). The chromosomal profile of cells from clone H-7 was dif- ferent, with most of the counts ranging between 45 and 70. In both
cell lines, 5--10% of the counts were significantly higher, suggesting the presence of cells containing variants of the tetraploid number of chromosomes, as was also shown for the mouse mammary epi- thelial cell line CID-9 (Barash, 1999).
DISCUSSION
It has been suggested that distinct, precommitted mammary ep- ithelial progenitors function to regenerate mammary ducts and lob-
ules (Kordon and Smith, 1998). With this hypothesis in mind, we prepared a heterogenous population of bovine mammary ceils and selected for precommitted progenitors that grow discontinuously and
differentiate upon hormonal and ECM signals.
Two further steps were imposed to establish cloned cell lines. Subpopulations with different levels of resistance to trypsinization were generated to preselect for higher milk-protein production
(Schmidhauser et al., 1990). Next, individual cells of both subpop- ulations were cultured, and their ability to functionally recover from
cryopreservation was confirmed. Both subpopulations depended on
FCS for cell proliferation, and no significant difference in BLG ex-
zr
FIG. 5. Cytokeratins, but not smooth muscle actin (SMA), are expressed in bovine cell cultures. H-7 or L-1 ceils were cultured on Matrigel for 7 d in the presence of insulin, prolactin, and hydrocortisone. (A) Cells were embedded in paraffin, and 5-1xm sections were stained with AE1 anti-cytokeratin antibody, a, H-7 cells; b, L-1 cells; c, lactating bovine mammary gland; d, similarly processed section from H-7 cells, not reacted with specific antibodies. Bar, 40 ~m. (B) Immunoblot analysis of cytokeratins in total cell extract. (C) SMA immunodetection in tissue and cell extracts. Tissue extracts from bovine and mouse mammary gland (MG) served as positive controls for SMA detection. Ponceau S staining (not shown) confirmed equal amounts of protein in all samples. Representative analysis of two independent experiments is presented.
FIG. 6. Transduction of prolaetin (PRL) signals in bovine mammary epithelial cells. H-7 and L-1 cells were cultured for 5 d on Matrigel in the presence of 0.5 Ixg/ml insulin. On day 5, PRL (0.05 Ixg/ml) was supplemented to the culture medium for the indicated times. Total cell extract was analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, blotted onto nitrocellulose filters, and incubated with antibodies specific to phospho-Stat5a/Stat5b or to PHAS-I (Barash, 1999). Ponceau S staining confirmed equal amounts of protein in all samples. Representative demonstration of two independent analyses is presented.
FIG. 7. Immunohistochemical analysis of 13-1actoglobulin (BLG) expression in bovine epithelial ceils. Cells were grown on Matrigel in the presence of insulin, hydrocortisone, and prolactin for 7 d. Paraffin sections (5 t~m) of H-7 (a) or L-1 (b) cells on Matrigel were stained with anti-BLG antibodies as described in Materials and Methods. (c) Paraffin section from similarly processed H-7 cells, which was not reacted with anti-BLG antibodies. Bar, 40 p~m. Results represent 3 independent analyses.
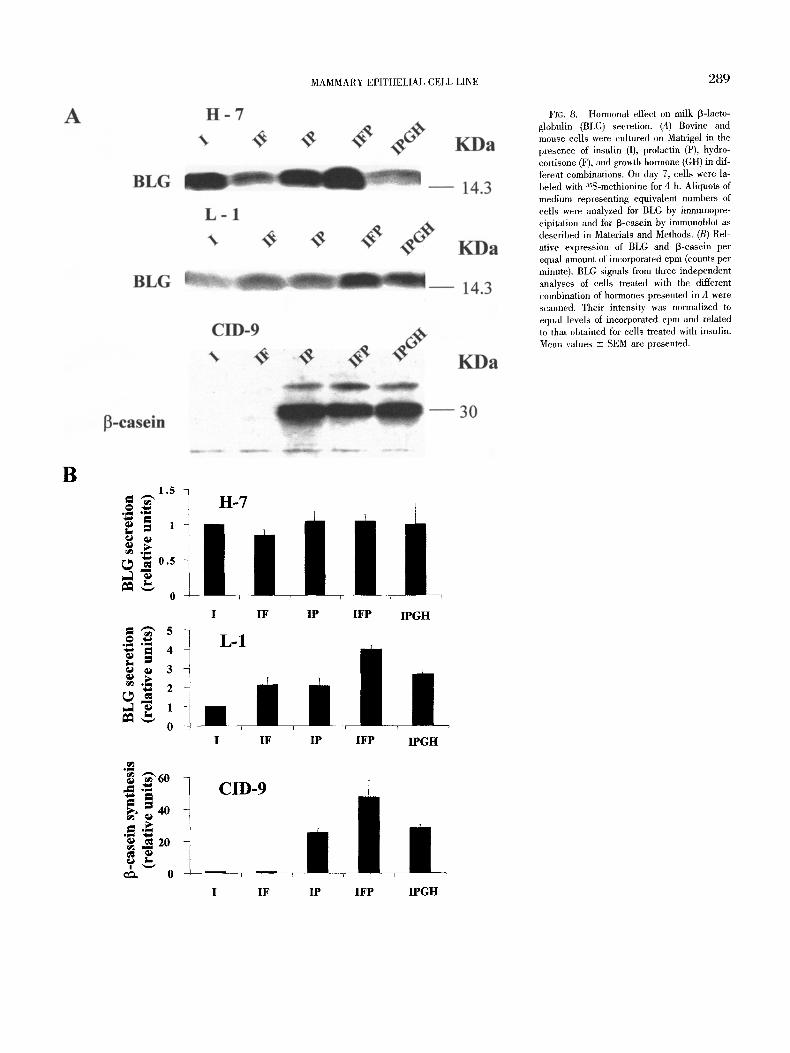
MAMMARY EPITHELIAL CELL LINE 289
FIG. 8. Hormonal effect on milk 13-1acto- globulin (BLG) secretion. (A) Bovine and mouse cells were cultured on Matrigel in the presence of insulin (I), prolactin (P), hydro- cortisone (F), and growth hormone (GH) in dif- ferent combinations. On day 7, cells were la- beled with 3SS-methionine for 4 h. Aliquots of medium representing equivalent numbers of cells were analyzed for BLG by immunopre- cipitation and for 13-casein by immunoblot as described in Materials and Methods. (B) Rel- ative expression of BLG and 13-casein per equal amount of incorporated cpm (counts per minute). BLG signals from three independent analyses of cells treated with the different combination of hormones presented in A were scanned. Their intensity was normalized to equal levels of incorporated cpm and related to that obtained for cells treated with insulin. Mean values -+ SEM are presented.
B 1.5
�9 ~ 1 o N
~o.s,_,
0
O ~ ' ~ 4
o 3
0
~ 6 0 o ~
o 40 ..~
0
I
L-1 1F IP IFP IPGH
CID-9
I IF IP IFP IPGH
I IF IP IFP IPGH

290 GERMAN AND BARASH
A
H - 7
A
~J
4) "ID ~
r
m
B Number of chromosomes
m
C
I Number of chromosomes
FIG. 9. Chromosomal analysis of H-7 and L-1 mamma~ ~ epithelial cells. (A) Summa~ of analyses in individual cells. (B) Representativ demonstration of diploid and tetraploid cells.

MAMMARY EPITHELIAL CELL LINE 291
pression was observed between bovine mammary cells with strong (L) compared with weak (H) attachment.
Immortal cell populations fi'om precommitted mouse (Danielson et al., 1984) and ovine (Duehler et al., 1988; Ilan et al., 1998) mammary progenitors have been previously generated. This ap- proach to recover cell lines is usually more tedious and measures of success are less predictable compared with the alternative meth- od of immortalizing cells with viral vectors. Indeed, two other func- tional cell lines, MAC-T (Huynh et al., 1991) and BME-UV1 (Zav- izion et al., 1996), were established by transfecting bovine mam- mary cells with the SV 40 large T antigen. Although cell transplants from these cultures did not develop into tumors in immunodefieient host tissue, other characteristics may have been affected and al- tered, compared with the native mammary epithelial cells.
The elonal cell lines CID-9 (Sehmidhauser et al., 1990) and HC11 (Ball et al., 1988) were derived fi'om the ancestral COMMA- D cell line, originally isolated from the mammary glands of mice in midpregnancy (Danielson et al., 1984). Regardless of their common
ancestry, these cell lines differ in their organization on a matrix and in their reliance on FCS for milk-protein production (Rosen et al., 1999). We therefore tested selected clones with high and low resis- tance to trypsinazation for mammary morphological and functional characteristics.
The positive staining of L-1 and H-7 cells with anti-cytokeratin antibodies and the negative staining of SMA suggest that these cell lines are of epithelial origin; no detectable conversion from epithe- lial to myoepithelial-like cells occurred through the cell passhges (Pechoux et al., 1999). Both cell lines were also nontumorigenic, as ascertained by the lack of tumor growth after transplantation into immunodeficient host tissue.
Duct-like structures were generated when L-1 cells were cultured on Matrigel, whereas H-7 ceils organized in mammospheres. These differences in organization may reflect the arrest of L-] cells at an earlier stage of differentiation or may result from differences in the cell-matrix interactions.
In contrast to other bovine cell lines that depend on FCS for differentiation and hormonal triggering of milk-protein gene ex- pression, L-1 and H-7 cells differentiated on Matrigel in serum-free medium. Both cell lines responded to PRL signaling by phosphor- ylation of Stat5 on Tyr 694. The activation of Stat5 by the Jak2 kinase upon recruitment to the PIlL receptor is followed by its transloeation to the nueleolus, binding to specific sequences in milk-protein gene promoters, and initiation of milk-protein gene transcription (Groner and Gouilleux, 1995). Although the Jak2- Stats pathway is a major route for lactogenic hormone signaling in rodents, it may play a less dominant role in regulating bovine milk protein production (Wheeler et al., 1997, 2001; Yang et al., 2000a, 2000b). We therefore extended our search for PRL signaling in L- 1 and H-7 cells to an additional route.
The cap-dependent RNA he/icase complex elF4E is a major tar- get for the regulation of protein synthesis in mammalian cells. It mediates the unwinding of the RNA structure near the 5' cap. This, in turn, facilitates binding by the 43S ribosomal subunit, thereby promoting translation initiation (Merriek and Hershey, 1996). A pu- tative channel for regulating milk-protein synthesis was recently discovered in differentiated, insulin-treated, CID-9 cells (Barash, 1999). There, PRL activates eIF4E by phosphorylating its binding protein PHAS-I (the inhibitor), resulting in the latter's dissociation from eIF4E and formation of the elF4F complex. Translation in-
duction has been shown to follow insulin enhancement of elF4F activity (Gallic and Traugh, 1994) and to be related to the aug- mented association between elF4E and elF4G (Kimball et al., 1997). Focusing on PHAS-I as a key mediator of this route, we show here that PRL activates the phosphorylation of PHAS-I in L- 1 and H-7 cells over a time course comparable with that shown for the CID-9 ceils.
In contrast to the correlation between attachment and milk-pro- tein production shown for the CID-9 nmrine cell line, the stronger resistance of L-1 bovine cells to trypsinization relative to H-7 was not associated with better expression of BLG synthesis. Hydrocor- tisone and PRL acted in synergy to induce the expression of BLG in L-1 cells, whereas no hormonal effect was observed in H-7 cells. Even in L-1 cells, lactogenie hormone regulation of BLG expression was less stringent than that seen in mouse mammary cells for [3- casein synthesis (this study) or for BLG expression in explants of transgenic mice carrying the BLG geue (Barash et al., 1993). This suggests that other dominant factors, such as circulating levels of amino acids, mediate the lactogenic stimulation of milk-protein gene expression in the bovine gland.
In conclusion, we established two independent bovine mammary epithelial cell lines from subpopulations that differ in their attach- ment to plastic plates. The near-diploid chromosome number shown for L-1 cells compared with the more variable karyotype determined in H-7 cells, together with better hormonal responsiveness and stronger attachment to the plate, make L-1 cells a favorite cell line for functional studies of the bovine mammary gland.
ACKNOWI,EDGMENTS
~ thank Dr. Amir Bor for taking the bovine mammary biopsies. This study was supported by a grant from the United States-Israel Binational Agricultural Research and Development Fund (BARD) and the Israeli Breed- ing Association.
REFERENCES
Ball, R. K.; Friis, R. R.; Schoenenbergm, C. A., et al. Prolactin regulation of beta-casein gene expression and of a cytosolic 120-kd protein in a cloned mouse mammary epithelial cell line. EMBO J. 7:2089 2095; 1988.
Barash, I. Prnlactin and insulin synergize to regulate the translation modu- lator PHAS-I via milngen-activated protein kinase-independent but wortmannin- and rapamycin-sensitive pathway. Mol. Cell Endocrinoh 155:37-49; 1999.
Barash, I.; Faerman, A.; Baruch, A., et al. Synthesis and secretion of human sermn albumin by mammary gland explants of virgin and lactating transgenic mice. Transgenic Res. 2:266-276; 1993.
Baruch, A.; Shani, M; Hounvitz, D., et al. Developmental regulation of the 13-1actoglobulin/human serum albunfin transgene is distinct from that of the [3-1actoglobulin and endogenous 13-casein gene in the mammary gland of transgenic mice. Dev. Genet. ]6:241-252; 1995.
Danielsnn, K. G.; Obom, C. J.; Dnrban, E. M., et al. Epitbelial mouse mam- mary cell line exhibiting nmYaal morphogenesis in vivo and functional differentiation in vitro. Proc. Natl. Acad. Sci. USA 81:3756-3760; 1984.
Duchler, M. D.; Schmoll, E; Pfneisl, E, et al. OMEC II: a new ovine mammary epithelial cell line. Biol. Cell 90:199-205; 1988.
Ebncr, K. E.; Hageman, E. C.; Larson, B. L. Cultivation and properties of mammary cell cultures. Exp. Cell. Res. 23:373-385; 1961.
Gallic, D. R.; Trough, J. A. Serum and insulin regulate cap function in 3T3- 1,1 cells. J. Biol. Chem. 269:7174-7179; 1994.
Groner, B.; Gouilleux, E Prolactin-med'ated gene activation in mammm 7 epithelial cells. Curl'. Opin. Genet. Dev. 5:587-594; 1995.

292 GERMAN AND BARASH
Hnynh, H. T.; Robitaille, G.; Turner, J. D. Establishment of bovine mammary epithelial cells (MAC-T): an in vitro model for bovine lactation. Exp. Cell Res. 197:191-199; 1991.
Ilan, I.; Barash, L; Faerman, A., et al. Dual regulation of 13-1actoglobulin/ human serum albumin by the extracellular matrix in mammary cells from transgenic mice. Exp. Cell. Res. 224:28-38; 1996.
Ilan, N.; Barash, I.; Gootwine, E., et al. Establishment and initial character- ization of the ovine mammary epithelial cell line NISH. In Vitro Cell. Dev. Biol, 34:326-332; 1998.
Kimball, S. R.; Jurasinski, C. V.; Lawrence, J. C., Jr,, et al. Insulin stimulates protein synthesis in skeletal muscle by enhancing the association of elF-4E and elF-4G. Am. J. Physiol. 272:C754-C759; 1997.
Kordon, E. C.; Smith. G. H. An entire functional mammary gland may com- prise the progeny from a single cell. Development 125:1921-1930; 1998.
Long, E.; Lazaris-Karatzas, A.; Karatzas, C., et al. Overexpressing eukaryotic translation initiation factor 4E stimulates bovine mammary epithelial cell population. Int. ]. Biochem. Cell Biol. 33:133-141; 2001.
MacKenzie, D. D.: Forsyth, I. A.; Brooker, B. E., et al. Culture of bovine mammary epithelial cells on collagen gels. Tissue Cell 14:231-241; 1982.
Merrick, W. C.; Hershey, J. g( B. The pathway and mechanism of eukaryoiic protein synthesis. In: Hershey, J. W. B.; Mathews, M. B.; Sonenberg, N., ed. Translation control. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1996:31-69.
Pechoux, C.; Gudjonsson, T.; Ronnov-Jessen, L., et al. Human mammary luminal epithelial cells contain progenitors to myoepithelial cells. Dev. Biol. 206:88-99; 1999.
Piedrahita, J. A. Targeted modification of the domestic animal genome. The- riogenology 53:105-116; 2000.
Ramagnolo, D.; Akers, R. M.; Byatt, J. C., et al. IGF-I-induced IGFBP-3 potentiates the mitogenic actions of IGF-I in mammary epithelial MD- IGF-I cells. Mol. Cell. Endocrinol. 102:131-139; 1994.
Robinson, R. M.; Akers, R. M.; Forsten, K. E. Real-time detection of insulin- like growth factor-1 stimulation of the MAC-T bovine mammary ep- ithelial cell line. Endocrine 13:345-352; 2000a.
Robinson, R. M.; Akers, R. M.; Forsten, K. E. Microphysiometry to evaluate real-time response of mammary epithelial cells to IGF-I. Crit. Rev. Biomed. Eng. 28:209-211; 2000b.
Romagnolo, D.; Akers, R. M.; Wong, E. A., ct al. Lactogenic hormones and extracellular matrix regulate expression of IGF-1 linked to MMTV- EFR in mammary epithelial cells. M01. Cell Endocrinol. 96:147-157; 1993.
Rosen, J. M.; Wyszomierski, S. L.; Hadsell, D. Regulation of milk protein gene expression. Annu. Rev. Nutr. 19:407-436; 1999.
Schmidhauser, C.; Bissell, M. J.; Myers, C., et al. Extracellular matrix and hormones transcriptionally regulate bovine 13-casein 5' sequences in stably transfected mouse mammary cells. Proc. Natl. Acad. Sci. USA 87:9118-9122; 1990.
Wheeler, T. T.; Broadhurst, M. K.; Sadowski, H. B., et al. Stat5 phosphory- lation status and DNA-binding activity in the bovine and routine mammary glands. Mol. Cell. Endocrinol. 176:39-48; 2001.
Wheeler, T. T.; Kuys, Y. M.; Broadhurst, M. M., et al. Manmmry Star5 abun- dance and activity are not altered with lactation state in cows. Mol. Cell. Endocrinol. 133:141-149; 1997.
Wilde, C. J.; Knight, C. H.; Flint, D. J. Control of milk secretion and apoptosis during mammary involution. J. Mammary Gland Biol. Neoplasia 4:129-136; 1999.
~bodward, T. L.; Turner, J. D.; Hung, H. T., et al. Inhibition of cellular proliferation and modulation of insulin-like growth factor binding pro- teins by retinoids in a bovine mammary epithelial cell line. J. Cell. Physiol. 167:488-499; 1996.
Yang, J.; Kennelly, J. J.; Baracos, V. E. Physiological levels of Stat5 DNA binding activity and protein in bovine mammary gland. J. Anita. Sci. 78:3126-3134; 2000a.
"tang, J.; Kennelly, J. J.; Baracos, V. E. The activity of transcription factor Stat5 responds to prolactin, growth hormone, and IGF-I in rat and bovine mammary explant culture. J. Anita. Set. 78:3114-3125; 2000b.
Zavizion, B.; van Duffelen, M.; Schaeffer, W., et al. Establishment and char- acterization of a bovine mammary myoepithelial cell line. In Vitro Cell. Dev. Biol. 32:149-158; 1996.




















