Aga1, the first alpha-Galactosidase from the human bacteria Ruminococcus gnavus E1, efficiently...
8
Aga1, the first alpha-Galactosidase from the human bacteria Ruminococcus gnavus E1, efficiently transcribed in gut conditions Margarita Aguilera a,b , Harivony Rakotoarivonina a,c , Alexandre Brutus a,d , Thierry Giardina a , Gwenola Simon a,e , Michel Fons a, * a Institut des Sciences Mole´culaires de Marseille, ISM2/BiosCiences UMR-CNRS 6263, Service 342, Faculte´St Je´roˆme, Universite´Paul Ce´zanne, 13397 Marseille Cedex 20, France b Department of Microbiology, School of Pharmacy, University of Granada, Campus de Cartuja S/N, 18014-Granada, Spain c Unite´Fractionnement des Agroressources et Environnement UMR INRA 614, Universite´de Reims Champagne-Ardenne, 8 rue Gabriel Voisin, 51688 Reims Cedex, France d Department of Energy Plant Research Laboratory, Michigan State University, East Lansing, MI, USA e Institut de Recherche pour le De´veloppement, Laboratoire de Microbiologie et Biotechnologie des Environnements Chauds, UMR 180, Universite´s de Provence et de la Me´diterrane´e, ESIL, Marseille, France Received 17 June 2011; accepted 14 September 2011 Available online 14 October 2011 Abstract Differential gene expression analysis was performed in monoxenic mice colonized with Ruminococcus gnavus strain E1, a major endogenous member of the gut microbiota. RNA arbitrarily primed-PCR fingerprinting assays allowed to specifically detect the in vivo expression of the aga1 gene, which was further confirmed by RT-PCR. The aga1 gene encoded a protein of 744 residues with calculated molecular mass of 85,207 Da. Aga1 exhibited significant similarity with previously characterized a-Galactosidases of the GH 36 family. Purified recombinant protein demonstrated high catalytic activity (104 7 U mg 1 ) and efficient p-nitrophenyl-a-D-galactopyranoside hydrolysis [k cat /K m ¼ 35.115 8.82 s 1 mM 1 at 55 C and k cat /K m ¼ 17.48 4.25 s 1 mM 1 at 37 C]. Ó 2011 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved. Keywords: Gut microbiota; Ruminococcus gnavus; Alpha-Galactosidase; In vivo expression; mRNA fingerprints 1. Introduction The human intestinal microbiota is estimated to contain up to 10 14 microbial cells, representing over 1000 different species (O’Hara and Shanahan, 2006), most of them belonging to three major phylogenetically-defined groups (Ribosomal Database Project registration no. 2.16.4.1) (Maidak et al., 2001): the Bacteroides group, the Clostridium coccoides group and the Clostridium leptum subgroup (Manichanh et al., 2008). The current challenge is to well identify microbial gut genome as it is considered the second human genome project. It deeply impacts on the health of the host through symbiotic relationships. Complex gut bacterial functions have been even considered as a human virtual organ (O’Hara and Shanahan, 2006) that interacts with the immune system, protects the host against potentially pathogenic microorganisms (Ballal et al., 2011; Fons et al., 2000), or is involved in the salvage of energy from undigested dietary compounds (Hughes and Rowlands, 2000). In the adult colon, components of the mucus layer or exopolysaccharides from microbial biofilms (Macfarlane, 2008) constitute an endogenous source of carbohydrates. Another source is constituted by non-digestible dietary residues and/ or prebiotics (ie: inulin, fructo-oligosaccharides or galacto- oligosaccharides) not hydrolyzed by the human digestive * Corresponding author. E-mail addresses: [email protected] (M. Aguilera), harivony. [email protected] (H. Rakotoarivonina), [email protected] (A. Brutus), [email protected] (T. Giardina), gwenola. [email protected] (G. Simon), [email protected] (M. Fons). Research in Microbiology 163 (2012) 14e21 www.elsevier.com/locate/resmic 0923-2508/$ - see front matter Ó 2011 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved. doi:10.1016/j.resmic.2011.10.005
Transcript of Aga1, the first alpha-Galactosidase from the human bacteria Ruminococcus gnavus E1, efficiently...

Research in Microbiology 163 (2012) 14e21www.elsevier.com/locate/resmic
Aga1, the first alpha-Galactosidase from the human bacteria Ruminococcusgnavus E1, efficiently transcribed in gut conditions
Margarita Aguilera a,b, Harivony Rakotoarivonina a,c, Alexandre Brutus a,d, Thierry Giardina a,Gwenola Simon a,e, Michel Fons a,*
a Institut des Sciences Moleculaires de Marseille, ISM2/BiosCiences UMR-CNRS 6263, Service 342, Faculte St Jerome, Universite Paul Cezanne,
13397 Marseille Cedex 20, FrancebDepartment of Microbiology, School of Pharmacy, University of Granada, Campus de Cartuja S/N, 18014-Granada, Spain
cUnite Fractionnement des Agroressources et Environnement UMR INRA 614, Universite de Reims Champagne-Ardenne, 8 rue Gabriel Voisin,
51688 Reims Cedex, FrancedDepartment of Energy Plant Research Laboratory, Michigan State University, East Lansing, MI, USA
e Institut de Recherche pour le Developpement, Laboratoire de Microbiologie et Biotechnologie des Environnements Chauds, UMR 180,
Universites de Provence et de la Mediterranee, ESIL, Marseille, France
Received 17 June 2011; accepted 14 September 2011
Available online 14 October 2011
Abstract
Differential gene expression analysis was performed in monoxenic mice colonized with Ruminococcus gnavus strain E1, a majorendogenous member of the gut microbiota. RNA arbitrarily primed-PCR fingerprinting assays allowed to specifically detect the in vivoexpression of the aga1 gene, which was further confirmed by RT-PCR. The aga1 gene encoded a protein of 744 residues with calculatedmolecular mass of 85,207 Da. Aga1 exhibited significant similarity with previously characterized a-Galactosidases of the GH 36 family.Purified recombinant protein demonstrated high catalytic activity (104 � 7 U mg�1) and efficient p-nitrophenyl-a-D-galactopyranosidehydrolysis [kcat/Km ¼ 35.115 � 8.82 s�1 mM�1 at 55 �C and kcat/Km ¼ 17.48 � 4.25 s�1 mM�1 at 37 �C].� 2011 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved.
Keywords: Gut microbiota; Ruminococcus gnavus; Alpha-Galactosidase; In vivo expression; mRNA fingerprints
1. Introduction
The human intestinal microbiota is estimated to containup to 1014 microbial cells, representing over 1000 differentspecies (O’Hara and Shanahan, 2006), most of them belongingto three major phylogenetically-defined groups (RibosomalDatabase Project registration no. 2.16.4.1) (Maidak et al.,2001): the Bacteroides group, the Clostridium coccoidesgroup and the Clostridium leptum subgroup (Manichanh et al.,
* Corresponding author.
E-mail addresses: [email protected] (M. Aguilera), harivony.
[email protected] (H. Rakotoarivonina), [email protected]
(A. Brutus), [email protected] (T. Giardina), gwenola.
[email protected] (G. Simon), [email protected] (M. Fons).
0923-2508/$ - see front matter � 2011 Institut Pasteur. Published by Elsevier Ma
doi:10.1016/j.resmic.2011.10.005
2008). The current challenge is to well identify microbial gutgenome as it is considered the second human genome project.It deeply impacts on the health of the host through symbioticrelationships. Complex gut bacterial functions have been evenconsidered as a human virtual organ (O’Hara and Shanahan,2006) that interacts with the immune system, protects the hostagainst potentially pathogenic microorganisms (Ballal et al.,2011; Fons et al., 2000), or is involved in the salvage of energyfrom undigested dietary compounds (Hughes and Rowlands,2000). In the adult colon, components of the mucus layer orexopolysaccharides frommicrobial biofilms (Macfarlane, 2008)constitute an endogenous source of carbohydrates. Anothersource is constituted by non-digestible dietary residues and/or prebiotics (ie: inulin, fructo-oligosaccharides or galacto-oligosaccharides) not hydrolyzed by the human digestive
sson SAS. All rights reserved.

15M. Aguilera et al. / Research in Microbiology 163 (2012) 14e21
enzymes, fermentable by microorganisms and able to increasethe degree of bacterial colonization selectivity (Martens et al.,2008; Turroni et al., 2010). Both types of fermentable carbo-hydrates play a crucial role in the development of complexecosystems, they are the major source of energy for humancolonic anaerobes and they produce short-chain fatty acidswhich are used as energy substrate by colonic epithelial cells(Bernalier-Donadille, 2010). For these reasons, a specialattention is given to bacterial glycoside hydrolases (Gopal et al.,2001; Perrin et al., 2001) like a-Galactosidases (a-Gal) (EC3.2.1.22). These enzymes are clustered into families 4, 27 and 36of the sequence-based classification of glycoside hydrolases(Cantarel et al., 2009; Henrissat and Bork, 1996).
Currently, complex “Omics” approaches are applied inorder to elucidate the main gut bacterial molecules that havea determinant impact in human health (Qin et al., 2010;Stecher et al., 2010), however huge efforts are still requiredfor succeeding in the interpretation of global data retrieved.Conversely, simple approaches are still being applied forfinding specific gut bacterial biomarkers (Mountzouris et al.,2009).
In this context, Ruminococcus gnavus is a strict anaerobic,Gram positive bacterium of the C. coccoides phylogenetically-defined group. The species R. gnavus was shown to belong tothe “core 57 species” present in the digestive tract of 90% ofhuman beings and was able to resist and colonize even ininflammatory gut conditions (Prindiville et al., 2004), sug-gesting that it should be considered as a model of bacterialadaptation to the digestive environment. The aim of thisstudy was to identify specific molecular structures that wereexpressed in vivo within the digestive tract. Thus, we report onthe characterization of a new a-Gal from R. gnavus strain E1previously isolated from the faecal microbiota of a healthyman (Dabard et al., 2001), expressed in the digestive tract ofmono-associated-mice, where the gene is transcribed. Cloningof the aga1 gene and heterologous expression of the corre-sponding a-Gal in Escherichia coli were carried out, and somebiochemical properties of the recombinant protein weredetermined.
2. Materials and methods
2.1. Bacterial strains and growth conditions
R. gnavus E1 was grown in an anaerobic cabinet at 37 �C,in pre-reduced BHI-YH medium (Dabard et al., 2001). E. colistrains were grown at 37 �C in LuriaeBertani medium (LB,Difco Laboratories). Transformants were selected on LBcontaining ampicillin (100 mg mL�1) or zeocin (50 mg mL�1)(SigmaeAldrich Chimie).
2.2. Animal model
Animal experiments were performed according to theguidelines of the French Ethic Committee. Eight-weeks-oldC3H/He germfree (GF) mice from the ANAXEM platform(Micalis-INRA, Jouy-en-Josas, France) were reared as
previously described (Crost et al., 2010). Further details andcommercial diet composition are given in the Supplementalmaterial.
2.3. In vitro-RNA extraction
Samples were taken at 3 different stages (early, middle, andlate phase) of an exponentially growing culture, and centri-fuged. Cell pellets were immediately frozen. When necessary,5 � 108 to 109 cells were thawed in cold TE buffer, and totalRNA was extracted. Bacteria were briefly disrupted, in thepresence of phenol, 10% SDS, 3 M sodium acetate (pH 5.2),and 0.6 mg of glass beads (SigmaeAldrich Chimie), byshaking twice (40 s and 4.5 speed) using the Fast Prepcentrifuge FP120 (BIO 101-QBIOgene). RNA extractionswere carried out following the specific instructions in theRNeasy kit (Qiagen). The elution of RNA was performed in50 mL of diethyl pyrocarbonate-treated distilled H2O.
2.4. In vivo-RNA extraction
“In vivo-RNA” was obtained from gnotobiotic mice caecalcontents. RNA extractions were carried out on 200 mg aliquotsof frozen material as described by Stahl et al. in 1988 andmodified by Dore et al. in 1998. The material thus obtainedwas further purified using the RNeasy kit (Qiagen).
The assessment of RNA integrity and concentration weredetermined as described in Supplemental material.
2.5. RAP-PCR
RAP-PCR was performed on “in vivo-RNA” and “in vitro-RNA”. The synthesis of first-strand cDNA was carried outwith M-MuLV-Reverse Transcriptase (MBI-Fermentas). Forsecond-strand synthesis, the reactions were performed with thearbitrary decaoligomers C15, J08, L06, L10, Q04, andT13 (Table 1). The double strand cDNA previously obtainedwas amplified by PCR in individual reactions performedwith primers OlB6, OlB7, OlB8, OlB9, OlB10, and OlB11(Table 1), previously assayed in RAPD studies over the samestrain for phylogenetic purposes (Marcille et al., 2002). Adetailed protocol is given in Supplemental material. Purifiedand re-amplified cDNAs obtained from different treatmentswere compared by electrophoresis on 1.5% agarose gel. Puri-fied PCR products of interest were cloned in the TA vector-based cloning kit (TOPO TA cloning kit, Invitrogen) usedaccording to the manufacturer’s instructions, then sequenced(Genome Express).
2.6. Inverse PCR and genome walking
Inverse PCR and genome walking were used to completethe aga1 sequence gene. Digested DNA of R. gnavus E1(10 mg) with PstI was circularized with the ligase kit. PCRamplification was carried out with the InvF/InvR primers(Table 1), leading to a 900 bp fragment which was subse-quently cloned into pGEM-T (Promega) and sequenced. DraI

Table 1
Oligonucleotide primer sequences.
Experiment Primer Sequence 50e30 Reference
C15 GACGGAAGAG Dore et al., 1998
J08 TTTGCCCGGT Dore et al., 1998
L06 GAGGGAAGAG Dore et al., 1998
L10 TGGGAGATGG Dore et al., 1998
Q04 AGTGCGCTGA Dore et al., 1998
T13 AGGACTGCCA Dore et al., 1998
OlB6 TGCTCTGCCC Fons et al., 2000
OlB7 GGTGACGCAG Fons et al., 2000
OlB8 GTCCACACGG Fons et al., 2000
OlB9 CATCTCATT T Fons et al., 2000
OlB10 AACAGTATGA Fons et al., 2000
OlB11 ACTTGTGTA A Fons et al., 2000
Inverse PCR InvF ATCACGAAGGATCCCGGGAATTTC This study
InvR AACAAATGCTCATATTTCTGGTTCT This study
Genome walking GalI2F CGAAAGCTGCAGCATGG This study
AgaGW3 CAGGAATCGCGAAGAAGGTAG This study
AgaGW4 TGAAAATGAGAGGGGGAGGTG This study
RT-PCR RT1 CAGCATTTGTTATCACGAAGGATCCCGGG This study
RT2 GTACAAGATCAGTGTGGTGTCGATCAACTC This study
Probe synthesis SbF CATCTTTACAACAAAGAA This study
SbR CTTTTTCCCAAAATACAGC This study
16 M. Aguilera et al. / Research in Microbiology 163 (2012) 14e21
digested genomic DNA was used as template with theuniversal genome walker kit (BD Biosciences) together withthe GalI2F primer and the AP2 adaptor primer to amplifya 3000 bp fragment. The PCR product was gel purified andsuccessive sequencing steps with primers GalI2F, AgaGW3,AgaGW4 (Table 1) and AP2 were performed to obtain thesequence of the DNA fragment including aga1.
2.7. Control of aga1 gene expression in various growthconditions by RT-PCR
Total RNA (1 mg) obtained from R. gnavus E1-monoassociated mice caecal contents or BHI-YH liquidcultures was reverse transcribed using random hexamerprimers (Invitrogen) and 200 U of M-MuLV-RT (MBI-Fermentas) according to the manufacturer’s protocol, and thecDNA was amplified using the aga1 specific primers RT1 andRT2 (Table 1). A negative control was performed using nonreverse-transcribed RNA. 16S rRNA amplification was used asan internal control for both growth conditions.
2.8. Aga1 heterologous expression in E. coli andpurification of the recombinant protein
The R. gnavus E1 fragment harbouring the aga1 genedeprived from its putative promoter region was cloned intoE. coli using the gateway technologies (Invitrogen). Protocoldetails are given in Supplemental material. The recombinantplasmid was transferred for expression into the E. coli strainBL21DE3.
Cloning the R. gnavus aga1 gene into pDEST�17 issupposed to allow the expression of a fusion protein contain-ing an N-terminal 6�His tag. High expression levels of
recombinant Aga1 protein were achieved as previouslydescribed (Berrin et al., 2007).
2.9. Enzymatic Aga1 assays: optimal pH, temperature,Km and kcat determination
a-Gal activity was measured by determining the rate ofhydrolysis of pNP-a-D-Gal at 37 �C. Optimal pH for a-Galactivity was estimated in a pH range of 3.0e9.0. The differentbuffers were: 20 mM sodium acetate, pH 3.0e5.0; 20 mMpotassium phosphate, pH 6.0e7.0; and 20 mM TriseHCl, pH7.0e9.0. Optimal temperature was also estimated at pH 6.0and at temperatures ranging from 30 �C to 70 �C. Thermalstability was performed at pH 6.0 and 55 �C. The a-Gal assayis extensively described in Supplemental material. For deter-mination of the apparent MichaeliseMenten constants, theinitial a-Gal velocities were measured in two conditions: pH6.0/55 �C and pH 6.0/37 �C, in 0.1 M potassium phosphatebuffer with pNP-a-D-Gal ranging from 2 to 10 mM.
2.10. Computational analysis of protein and nucleotidesequences
DNA and protein sequences analyses were performed usingthe National Centre for Biotechnology Information (NCBIdatabases). Putative transcriptional terminators were predictedin silico using the “RNA-Fold program”.
2.11. Nucleotide sequence accession numbers
The nucleotide sequence data of R. gnavus aga1 wasdeposited in the EMBL database under Accession NumberACL13770.1.

17M. Aguilera et al. / Research in Microbiology 163 (2012) 14e21
3. Results
3.1. RNA fingerprinting analysis: identification ofa differentially expressed gene encoding the putativeR. gnavus a-Gal
The first step of this study consisted of the comparison ofR. gnavus E1 RNA fingerprints obtained when the bacteriagrew under two different conditions: i) in vivo, in the caecumof gnotobiotic mice; and ii) in vitro, in BHI-YH culturemedium. The comparison of RAP-PCR electrophoresispatterns obtained with primer OLB7 from “in vivo-RNA” vs.“in vitro-RNA” highlighted the presence of one major1300 bp-additional band specific to in vivo conditions (Fig. 1).
This band called transcript 1 was purified from agarose gel,re-amplified using the same conditions, cloned into thepGEM-T vector and sequenced. The primary structurededuced from transcript 1 showed that the putative productexhibited 41% identity (58% positive) with an a-Gal (EC3.2.1.22) from Aspergillus niger previously characterized(Ademark et al., 2001).
3.2. Localization of the gene encoding transcript 1 onthe R. gnavus E1 genome
In order to confirm that transcript 1 was expressed byR. gnavus E1, southern blot hybridization was performed. Theresults confirmed that the gene encoding transcript 1 was indeedpart of R. gnavus E1 chromosomal genome, and that it waspresent in a single copy (Fig. S1). This gene was called aga1.
Fig. 1. RAP-PCR fingerprinting using the arbitrary primer Olb7. R. gnavus E1
grown in vivo (Vv) vs. in vitro conditions (Vt). (M), ladder showing the bands
sizes. Arrows indicate the differentially expressed band. The RAP-PCR
negative control did not show any band.
3.3. aga1 gene of R. gnavus E1: cloning, nucleotidesequence, and deduced gene product
Chromosome walking was performed to clone the regionupstream aga1 and allowed the characterization of a 5300 bp-long DNA fragment, harbouring the complete aga1 geneembedded in between two other partial ORFs (Fig. 2A).
The partial ORF located upstream of aga1, on the sameDNA strand, sized 2400 bp, was called pyrAB. The deducedprimary structure of the putative PyrAB fragment shared 44%identity (63% positive) with the carbamoyl-phosphate syn-thase [EC: 6.3.5.5] of E. coli (Thoden et al., 1999). Highidentity-scores, over 89%, were obtained with putativeproteins from R. gnavus ATCC 29149 (accession # NZAAYG02000032.1), Ruminococcus lactaris ATCC 29176(accession # NZ ABOU02000046.1), Dorea longicatena DSM13814 (accession # NZ AAXB02000002.1), and Clostridiumhylemonae DSM 15053 (accession # NZ ABYI02000027.1)from the “Human Gut Microbiome Initiative” project (http://www.genome.gov/Pages/Research/Sequencing/SeqProposals/HGMI-Seq.pdf). Downstream of pyrAB, an inverted repeatpredicted to form a stemloop structure with DG ¼ �15.66 kcalmol�1 (Fig. 2C), might act as a rho-independent transcriptionterminator (De Hoon et al., 2005).
The partial ORF located downstream of aga1 sized 477 bpand was in the opposite orientation. The deduced putativepeptide shared significant homologies with transcriptionalregulators of the AraC family. The highest score was obtainedwith MsmR, a positive effector controlling the expression of 8contiguous genes involved in the uptake and metabolism ofmultiple sugars (the msm system) in Streptococcus mutans(Zeng and Burne, 2008).
The aga1 gene sized 2232 kb nucleotides, with a G þ C%content of 48.34%. The overall average G þ C% content at thethird position of codons had a similar value (49.5%). Aputative ribosome-binding site (AGGAGA) was centred about10 nucleotides upstream of the translation initiation site(Fig. 2B). Analysis of the promoter region of the aga1 generevealed the presence of consensus promoter boxes (�35:TCTGGC and �10: TATAAT). No consensus sequence(TGNNANCGNTNNCA), looking like a putative catabolicresponsive-element (CRE box) involved in the regulation ofaga1 gene expression (Puri-Taneja et al., 2007) was retrieved.Downstream of aga1, an inverted repeat predicted to forma stemloop structure with DG ¼ �14.43 kcal mol�1 (Fig. 2D),might act as a rho-independent transcription terminator. Theaga1 coding sequence (cds) displayed over 95% identity withthe cds 03611 from R. gnavus ATCC 29149 proposed to codefor a putative a-Gal.
The aga1 gene resulted in a predicted protein of 743 aminoacid residues with a molecular mass of 85,216 Da and a pre-dicted pI of 5.48. Aga1 shares the highest amino acid sequenceidentity of 41% and 27% with the a-Gals from A. niger andThermotoga maritima respectively (data not shown). A COG3345 motive (cluster of orthologs) spanning from residues265 to 685 and specific for a-Gal was detected and the corestructure containing the a-Gal putative active site was
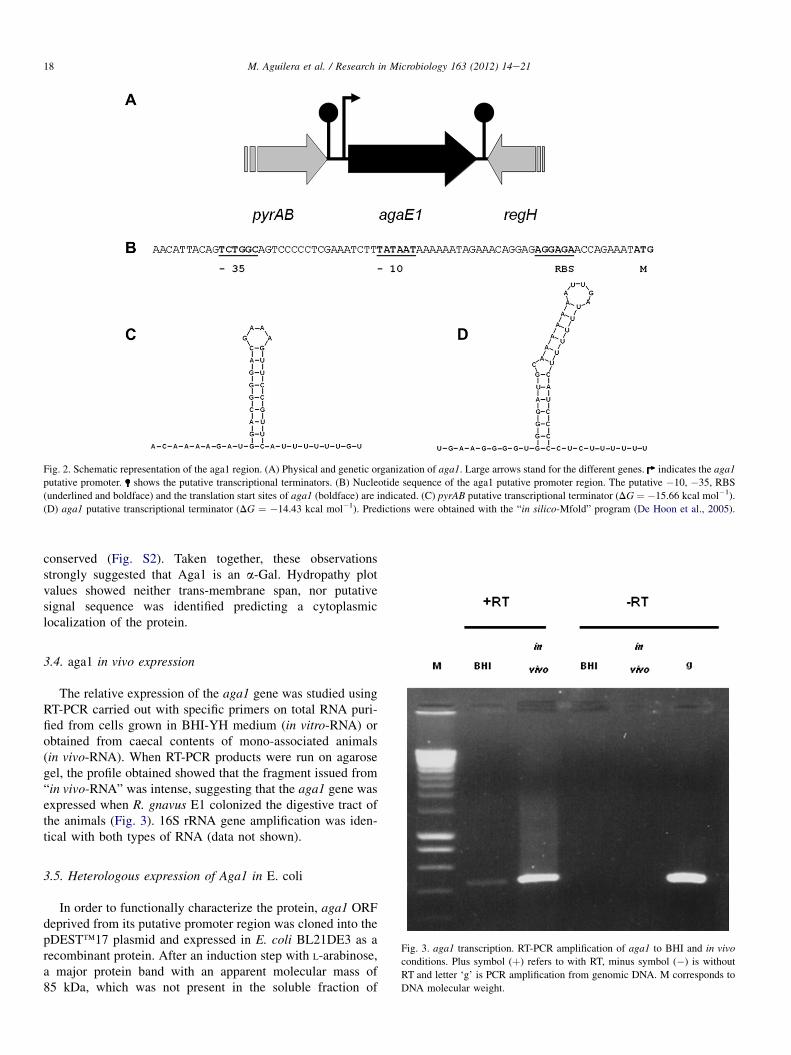
Fig. 2. Schematic representation of the aga1 region. (A) Physical and genetic organization of aga1. Large arrows stand for the different genes. indicates the aga1
putative promoter. shows the putative transcriptional terminators. (B) Nucleotide sequence of the aga1 putative promoter region. The putative �10, �35, RBS
(underlined and boldface) and the translation start sites of aga1 (boldface) are indicated. (C) pyrAB putative transcriptional terminator (DG ¼ �15.66 kcal mol�1).
(D) aga1 putative transcriptional terminator (DG ¼ �14.43 kcal mol�1). Predictions were obtained with the “in silico-Mfold” program (De Hoon et al., 2005).
18 M. Aguilera et al. / Research in Microbiology 163 (2012) 14e21
conserved (Fig. S2). Taken together, these observationsstrongly suggested that Aga1 is an a-Gal. Hydropathy plotvalues showed neither trans-membrane span, nor putativesignal sequence was identified predicting a cytoplasmiclocalization of the protein.
3.4. aga1 in vivo expression
The relative expression of the aga1 gene was studied usingRT-PCR carried out with specific primers on total RNA puri-fied from cells grown in BHI-YH medium (in vitro-RNA) orobtained from caecal contents of mono-associated animals(in vivo-RNA). When RT-PCR products were run on agarosegel, the profile obtained showed that the fragment issued from“in vivo-RNA” was intense, suggesting that the aga1 gene wasexpressed when R. gnavus E1 colonized the digestive tract ofthe animals (Fig. 3). 16S rRNA gene amplification was iden-tical with both types of RNA (data not shown).
3.5. Heterologous expression of Aga1 in E. coli
Fig. 3. aga1 transcription. RT-PCR amplification of aga1 to BHI and in vivo
conditions. Plus symbol (þ) refers to with RT, minus symbol (�) is without
RT and letter ‘g’ is PCR amplification from genomic DNA. M corresponds to
DNA molecular weight.
In order to functionally characterize the protein, aga1 ORFdeprived from its putative promoter region was cloned into thepDEST�17 plasmid and expressed in E. coli BL21DE3 as arecombinant protein. After an induction step with L-arabinose,a major protein band with an apparent molecular mass of85 kDa, which was not present in the soluble fraction of
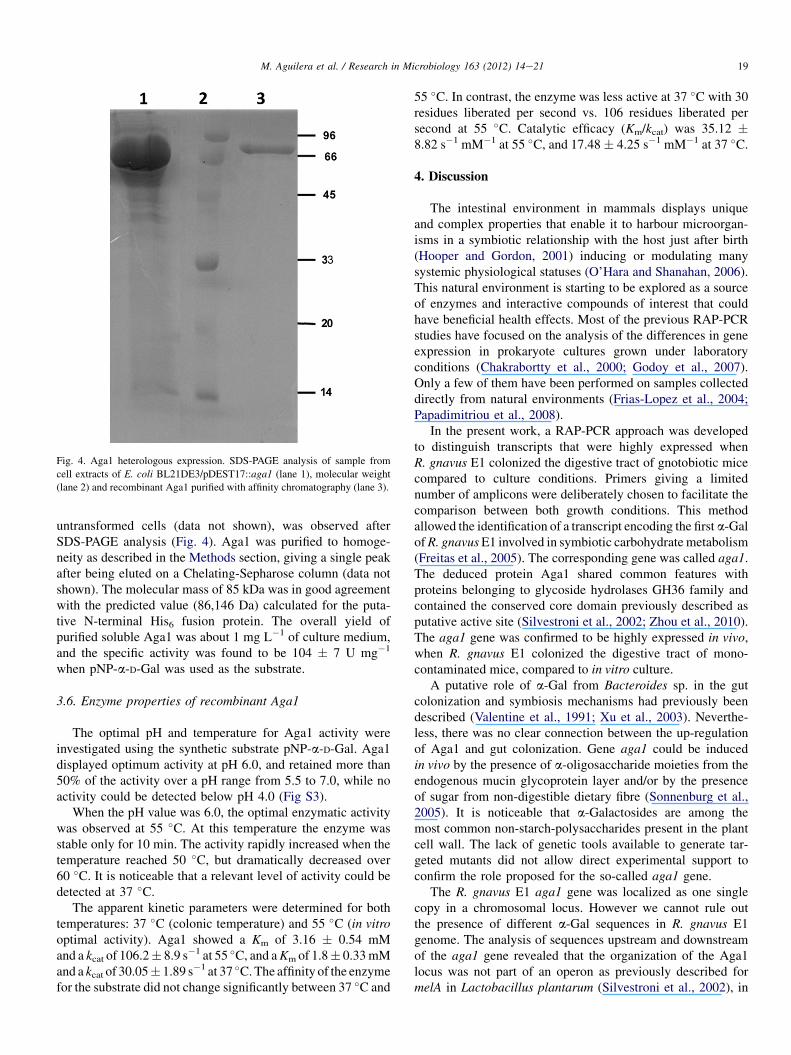
Fig. 4. Aga1 heterologous expression. SDS-PAGE analysis of sample from
cell extracts of E. coli BL21DE3/pDEST17::aga1 (lane 1), molecular weight
(lane 2) and recombinant Aga1 purified with affinity chromatography (lane 3).
19M. Aguilera et al. / Research in Microbiology 163 (2012) 14e21
untransformed cells (data not shown), was observed afterSDS-PAGE analysis (Fig. 4). Aga1 was purified to homoge-neity as described in the Methods section, giving a single peakafter being eluted on a Chelating-Sepharose column (data notshown). The molecular mass of 85 kDa was in good agreementwith the predicted value (86,146 Da) calculated for the puta-tive N-terminal His6 fusion protein. The overall yield ofpurified soluble Aga1 was about 1 mg L�1 of culture medium,and the specific activity was found to be 104 � 7 U mg�1
when pNP-a-D-Gal was used as the substrate.
3.6. Enzyme properties of recombinant Aga1
The optimal pH and temperature for Aga1 activity wereinvestigated using the synthetic substrate pNP-a-D-Gal. Aga1displayed optimum activity at pH 6.0, and retained more than50% of the activity over a pH range from 5.5 to 7.0, while noactivity could be detected below pH 4.0 (Fig S3).
When the pH value was 6.0, the optimal enzymatic activitywas observed at 55 �C. At this temperature the enzyme wasstable only for 10 min. The activity rapidly increased when thetemperature reached 50 �C, but dramatically decreased over60 �C. It is noticeable that a relevant level of activity could bedetected at 37 �C.
The apparent kinetic parameters were determined for bothtemperatures: 37 �C (colonic temperature) and 55 �C (in vitrooptimal activity). Aga1 showed a Km of 3.16 � 0.54 mMand a kcat of 106.2� 8.9 s�1 at 55 �C, and aKm of 1.8� 0.33mMand a kcat of 30.05� 1.89 s�1 at 37 �C.The affinity of the enzymefor the substrate did not change significantly between 37 �C and
55 �C. In contrast, the enzyme was less active at 37 �C with 30residues liberated per second vs. 106 residues liberated persecond at 55 �C. Catalytic efficacy (Km/kcat) was 35.12 �8.82 s�1 mM�1 at 55 �C, and 17.48� 4.25 s�1 mM�1 at 37 �C.
4. Discussion
The intestinal environment in mammals displays uniqueand complex properties that enable it to harbour microorgan-isms in a symbiotic relationship with the host just after birth(Hooper and Gordon, 2001) inducing or modulating manysystemic physiological statuses (O’Hara and Shanahan, 2006).This natural environment is starting to be explored as a sourceof enzymes and interactive compounds of interest that couldhave beneficial health effects. Most of the previous RAP-PCRstudies have focused on the analysis of the differences in geneexpression in prokaryote cultures grown under laboratoryconditions (Chakrabortty et al., 2000; Godoy et al., 2007).Only a few of them have been performed on samples collecteddirectly from natural environments (Frias-Lopez et al., 2004;Papadimitriou et al., 2008).
In the present work, a RAP-PCR approach was developedto distinguish transcripts that were highly expressed whenR. gnavus E1 colonized the digestive tract of gnotobiotic micecompared to culture conditions. Primers giving a limitednumber of amplicons were deliberately chosen to facilitate thecomparison between both growth conditions. This methodallowed the identification of a transcript encoding the first a-GalofR. gnavusE1 involved in symbiotic carbohydrate metabolism(Freitas et al., 2005). The corresponding gene was called aga1.The deduced protein Aga1 shared common features withproteins belonging to glycoside hydrolases GH36 family andcontained the conserved core domain previously described asputative active site (Silvestroni et al., 2002; Zhou et al., 2010).The aga1 gene was confirmed to be highly expressed in vivo,when R. gnavus E1 colonized the digestive tract of mono-contaminated mice, compared to in vitro culture.
A putative role of a-Gal from Bacteroides sp. in the gutcolonization and symbiosis mechanisms had previously beendescribed (Valentine et al., 1991; Xu et al., 2003). Neverthe-less, there was no clear connection between the up-regulationof Aga1 and gut colonization. Gene aga1 could be inducedin vivo by the presence of a-oligosaccharide moieties from theendogenous mucin glycoprotein layer and/or by the presenceof sugar from non-digestible dietary fibre (Sonnenburg et al.,2005). It is noticeable that a-Galactosides are among themost common non-starch-polysaccharides present in the plantcell wall. The lack of genetic tools available to generate tar-geted mutants did not allow direct experimental support toconfirm the role proposed for the so-called aga1 gene.
The R. gnavus E1 aga1 gene was localized as one singlecopy in a chromosomal locus. However we cannot rule outthe presence of different a-Gal sequences in R. gnavus E1genome. The analysis of sequences upstream and downstreamof the aga1 gene revealed that the organization of the Aga1locus was not part of an operon as previously described formelA in Lactobacillus plantarum (Silvestroni et al., 2002), in

20 M. Aguilera et al. / Research in Microbiology 163 (2012) 14e21
contrast to those clustered structures that had been found inEnterococcus faecium (Zhang et al., 2011), Enterobactercloacae (Okazaki et al., 1997) and Citrobacter freundii(Shimamoto et al., 2001).
A ribosome-binding site, a putative promoter regionsequence upstream, and an inverted repeat that could work asa putative transcriptional terminator for aga1 and the adjacentgene, were detected. No consensus CRE site was found nearthe �35 promoter region, which suggested that aga1 regula-tion could be mediated by an induction mechanism rather thanby catabolite repression (Muscariello et al., 2001).
The whole G þ C% content in aga1 was similar to thatin other a-Gal genes (48.5%) but higher than that in theR. gnavus genome G þ C% content (34%) and higher than b-glucuronidase gene (39%) previously identified in the samestrain of R. gnavus E1 (Beaud et al., 2005). This higher valueof G þ C% content at the third position of the codon usage inaga1 could suggest that this gene or locus could have beenacquired through horizontal transfer from environmental gutbacteria colonizers.
Heterologous expression of aga1 revealed an 85-kDaprotein, in agreement with the predicted 85,216 Da calculatedmass encoded by the gene. Optimal activity was dosed at55 �C and the enzyme could resist interestingly high pHvalues, properties already described for a-glycosidases fromBifidobacteria sp. (Hinz et al., 2006), Clostridium sp. (Suryaniet al., 2003), Lactococcus lactis and Lactobacillus sp.(LeBlanc et al., 2004). It is noticeable that a relevant activitylevel was detected still at 37 �C. The enzymatic parameters,Km and kcat, determined with pNP-a-D-Gal, were similar tothose of other GH36 a-Gal described previously (Ademarket al., 2001; Nakai et al., 2010; Spangenberg et al., 2000;Tzortzis et al., 2003; Zhao et al., 2008).
As the R. gnavus species have been shown to be present inthe dominant microbiota of 90% of human subjects (Qin et al.,2010), the specific role of a-Gal enzymes produced in the gutby this group should contribute to carbohydrate metabolismand keep the homeostatic balance of the mucus intestinal layerwhich determines the development of complex microbialcommunities. The food processing use of this recombinantenzyme could be counselled and safer than other a-Galacto-sidases from non-gut bacterial sources (Patil et al., 2010).
Acknowledgements
We are indebted to S. Rabot (Micalis-INRA e Jouy-en-Josas e France) for providing us with the axenic rats of theanimal facility “ANAXEM platform” she is in charge of, andto C. Bridonneau and P. Guillaume for skilful technicalassistance. M. Aguilera was supported by a PostdoctoralFellowship from “Fundacion Alfonso Martın Escudero” andthe University of Granada, Spain. This work was carried outwith financial support from the Commission of the EuropeanCommunities specific programme “Quality of Life andmanagement of Living Resources”, Key Action 1, “Food,Nutrition and Health”, project INFABIO QLK-1 2002 02606coordinated by Dr C. Edwards (University of Glasgow, UK).
Appendix. Supplemental material
Supplementary data related to this article can be foundonline at doi:10.1016/j.resmic.2011.10.005.
References
Ademark, P., de Vries, R.P., Hagglund, P., Stalbrand, H., Visser, J., 2001.
Cloning and characterization of Aspergillus niger genes encoding an
alpha-galactosidase and a beta-mannosidase involved in galactomannan
degradation. Eur. J. Biochem. 268, 2982e2990.
Ballal, S.A., Gallini, C.A., Segata, N., Huttenhower, C., Garrett, W.S., 2011.
Host and gut microbiota symbiotic factors: lessons from inflammatory
bowel disease and successful symbionts. Cell. Microbiol. 134, 508e517.
Beaud, D., Tailliez, P., Anba-Mondoloni, J., 2005. Genetic characterization of
the beta-glucuronidase enzyme from a human intestinal bacterium,
Ruminococcus gnavus. Microbiology 151, 2323e2330.
Bernalier-Donadille, A., 2010. Fermentative metabolism by the human gut
microbiota. Gastroenterol. Clin. Biol. 34 (Suppl. 1), S16eS22.
Berrin, J.G., Ajandouz el, H., Georis, J., Arnaut, F., Juge, N., 2007. Substrate
and product hydrolysis specificity in family 11 glycoside hydrolases:
an analysis of Penicillium funiculosum and Penicillium griseofulvum
xylanases. Appl. Microbiol. Biotechnol. 74, 1001e1010.
Cantarel, B.L., Coutinho, P.M., Rancurel, C., Bernard, T., Lombard, V.,
Henrissat, B., 2009. The carbohydrate-active enzymes database (CAZy):
an expert resource for glycogenomics. Nucleic Acids Res. 37,
D233eD238.
Chakrabortty, A., Das, S., Majumdar, S., Mukhopadhyay, K.,
Roychoudhury, S., Chaudhuri, K., 2000. Use of RNA arbitrarily primed-
PCR fingerprinting to identify Vibrio cholerae genes differentially
expressed in the host following infection. Infect. Immun. 68, 3878e3887.
Crost, E.H., Pujol, A., Ladire, M., Dabard, J., Raibaud, P., Carlier, J.P.,
Fons, M., 2010. Production of an antibacterial substance in the digestive
tract involved in colonization-resistance against Clostridium perfringens.
Anaerobe 16, 597e603.
Dabard, J., Bridonneau, C., Phillipe, C., Anglade, P., Molle, D., Nardi, M.,
Ladire, M., Girardin, H., Marcille, F., Gomez, A., Fons, M., 2001. Rumi-
nococcin A, a new lantibiotic produced by a Ruminococcus gnavus strain
isolated from human feces. Appl. Environ. Microbiol. 67, 4111e4118.De Hoon, M.J., Makita, Y., Nakai, K., Miyano, S., 2005. Prediction of tran-
scriptional terminators in Bacillus subtilis and related species. PLoS
Comput. Biol. 1, 25.
Dore, J., Sghir, A., Hannequart-Gramet, G., Corthier, G., Pochart, P., 1998.
Design and evaluation of a 16S rRNA-targeted oligonucleotide probe for
specific detection and quantitation of human faecal Bacteroides pop-
ulations. Syst. Appl. Microbiol. 21, 65e71.
Fons, M., Gomez, A., Karjalainen, T., 2000. Mechanisms of colonisation and
colonisation resistance of the digestive tract. Microb. Ecol. Health Dis. 12,
240e246.
Freitas, M., Axelsson, L.G., Cayuela, C., Midtvedt, T., Trugnan, G., 2005.
Indigenous microbes and their soluble factors differentially modulate
intestinal glycosylation steps in vivo. Use of a “lectin assay” to survey
in vivo glycosylation changes. Histochem. Cell Biol. 124, 423e433.
Frias-Lopez, J., Bonheyo, G.T., Fouke, B.W., 2004. Identification of differ-
ential gene expression in bacteria associated with coral black band disease
by using RNA-arbitrarily primed PCR. Appl. Environ. Microbiol. 70,
3687e3694.
Godoy, A.P., Reis, F.C., Ferraz, L.F., Gerrits, M.M., Mendonca, S., Kusters, J.G.,
Ottoboni, L.M., Ribeiro, M.L., Pedrazzoli, J.J., 2007. Differentially
expressed genes in response to amoxicillin in Helicobacter pylori analyzed
by RNA arbitrarily primed PCR. FEMS Immunol. Med. Microbiol. 50,
226e230.Gopal, P.K., Sullivan, P.A., Smart, J.B., 2001. Utilisation of galacto-
oligosaccharides as selective substrates for growth by lactic acid bacteria
including Bifidobacterium lactis DR10 and Lactobacillus rhamnosus
DR20. Int. Dairy J. 11, 19e25.

21M. Aguilera et al. / Research in Microbiology 163 (2012) 14e21
Henrissat, B., Bork, P., 1996. On the classification of modular proteins. Protein
Eng. 9, 725e726.
Hinz, S.W., Doeswijk-Voragen, C.H., Schipperus, R., van den Broek, L.A.,
Vincken, J.P., Voragen, A.G., 2006. Increasing the transglycosylation
activity of alpha-galactosidase from Bifidobacterium adolescentis
DSM 20083 by site-directed mutagenesis. Biotechnol. Bioeng. 93,
122e131.
Hooper, L.V., Gordon, J.I., 2001. Commensal hostebacterial relationships inthe gut. Science 292, 1115e1118.
Hughes, R., Rowlands, I.R., 2000. Metabolic activities of the gut microflora in
relation to cancer. Microb. Ecol. Health Dis. 12, 179e185.
LeBlanc, J.G., Garro, M.S., Silvestroni, A., Connes, C., Piard, J.C.,
Sesma, F., Savoy de Giori, G., 2004. Reduction of alpha-
galactooligosaccharides in soyamilk by Lactobacillus fermentum CRL
722: in vitro and in vivo evaluation of fermented soyamilk. J. Appl.
Microbiol. 97, 876e881.Macfarlane, S., 2008. Microbial biofilm communities in the gastrointestinal
tract. J. Clin. Gastroenterol. 42, 142e143.
Maidak, B.L., Cole, J.R., Lilburn, T.G., Parker Jr., C.T., Saxman, P.R.,
Farris, R.J., Garrity, G.M., Olsen, G.J., Schmidt, T.M., Tiedje, J.M., 2001.
The RDP-II (ribosomal database project). Nucleic Acids Res. 29,
173e174.
Manichanh, C., Chapple, C.E., Frangeul, L., Gloux, K., Guigo, R., Dore, J.,
2008. A comparison of random sequence reads versus 16S rDNA
sequences for estimating the biodiversity of a metagenomic library.
Nucleic Acids Res. 36, 5180e5188.
Marcille, F., Gomez, A., Joubert, P., Ladire, M., Veau, G., Clara, A., Gavini, F.,
Willems, A., Fons, M., 2002. Distribution of genes encoding the trypsin-
dependent lantibiotic ruminococcin A among bacteria isolated from
human fecal microbiota. Appl. Environ. Microbiol. 68, 3424e3431.Martens, E.C., Chiag, H.C., Gordon, J.I., 2008. Mucosal glycan foraging
enhances fitness and transmission of a saccharolytic human gut bacterial
symbiont. Cell Host Microbe 4, 447e457.
Mountzouris, K.C., Kotzampassi, K., Tsirtsikos, P., Kapoutzis, K., Fegeros, K.,
2009. Effects of Lactobacillus acidophilus on gut microflora metabolic
biomarkers in fed and fasted rats. Clin. Nutr. 28, 318e324.
Muscariello, L., Marasco, R., De Felice, M., Sacco, M., 2001. The functional
ccpA gene is required for carbon catabolite repression in Lactobacillus
plantarum. Appl. Environ. Microbiol. 67, 2903e2907.
Nakai, H., Baumann, M.J., Petersen, B.O., Westphal, Y., Hachem, M.A.,
Dilokpimol, A., Duus, J.Ø., Schols, H.A., Svensson, B., 2010. Aspergillus
nidulans alpha-galactosidase of glycoside hydrolase family 36 catalyses
the formation of alpha-galacto-oligosaccharides by transglycosylation.
FEBS J. 277, 3538e3551.
O’Hara, A.M., Shanahan, F., 2006. The gut flora as a forgotten organ. EMBO
Rep. 7, 688e693.
Okazaki, N., Jue, X.X., Miyake, H., Kuroda, M., Shimamoto, T., Tsuchiya, T.,
1997. A melibiose transporter and an operon containing its gene in
Enterobacter cloacae. J. Bacteriol. 179, 4443e4445.Papadimitriou, K., Boutou, E., Zoumpopoulou, G., Tarantilis, P.A.,
Polissiou, M., Vorgias, C.E., Tsakalidou, E., 2008. RNA arbitrarily primed
PCR and Fourier transform infrared spectroscopy reveal plasticity in the
acid tolerance response of Streptococcus macedonicus. Appl. Environ.
Microbiol. 74, 6068e6076.
Patil, A.G., Praveen Kumar, K., Mulimani, V.H., Veeranagouda, Y., Lee, K.,
2010. a-Galactosidase from Bacillus megaterium VHM1 and its applica-
tion in removal of flatulence-causing factors from soymilk. J. Microbiol.
Biotechnol. 20, 1546e1554.
Perrin, S., Warchol, M., Grill, J.P., Schneider, F., 2001. Fermentations of
fructo-oligosaccharides and their components by Bifidobacterium infantis
ATCC 15697 on batch culture in semi-synthetic medium. J. Appl.
Microbiol. 90, 859e865.
Prindiville, T., Cantrell, M., Wilson, K.H., 2004. Ribosomal DNA sequence
analysis of mucosa-associated bacteria in Crohn’s disease. Inflamm. Bowel
Dis. 10, 824e833.
Puri-Taneja, A., Schau, M., Chen, Y., Hulett, F.M., 2007. Regulators of the
Bacillus subtilis cydABCD operon: identification of a negative regulator,
CcpA, and a positive regulator, ResD. J. Bacteriol. 189, 3348e3358.
Qin, J., Li, R., Raes, J., Arumugam, M., Burgdorf, K.S., Manichanh, C.,
Nielsen, T., Pons, N., Levenez, F., Yamada, T., Mende, D.R., Li, J., Xu, J.,
Li, S., Li, D., Cao, J., Wang, B., Liang, H., Zheng, H., Xie, Y., Tap, J.,
Lepage, P., Bertalan, M., Batto, J.M., Hansen, T., Le Paslier, D.,
Linneberg, A., Nielsen, H.B., Pelletier, E., Renault, P., Sicheritz-Ponten, T.,
Turner, K., Zhu, H., Yu, C., Li, S., Jian,M., Zhou, Y., Li, Y., Zhang, X., Li, S.,
Qin, N., Yang, H.,Wang, J., Brunak, S., Dore, J., Guarner, F., Kristiansen, K.,
Pedersen, O., Parkhill, J., Weissenbach, J., MetaHIT Consortium, Bork, P.,
Ehrlich, S.D., Wang, J., 2010. A human gut microbial gene catalogue
established by metagenomic sequencing. Nature 464, 59e65.
Shimamoto, T., Xu, X.J., Okazaki, N., Kawakami, H., Tsuchiya, T., 2001.
A cryptic melibiose transporter gene possessing a frameshift from
Citrobacter freundii. J. Biochem. 129, 607e613.
Silvestroni, A., Connes, C., Sesma, F., De Giori, G.S., Piard, J.C., 2002.
Characterization of the melA locus for alpha-galactosidase in Lactobacillus
plantarum. Appl. Environ. Microbiol. 68, 5464e5471.Sonnenburg, J.L., Xu, J., Leip, D.D., Chen, C.H., Westover, B.P.,
Weatherford, J., Buhler, J.D., Gordon, J.I., 2005. Glycan foraging in vivo
by an intestine-adapted bacterial symbiont. Science 307, 1955e1959.Spangenberg, P., Andre, C., Dion, M., Rabiller, C., Mattes, R., 2000.
Comparative study of new alpha-galactosidases in transglycosylation
reactions. Carbohydr. Res. 329, 65e73.
Stahl, D.A., Flesher, B., Mansfield, H.R., Montgomery, L., 1988. Use of
phylogenetically based hybridization probes for studies of ruminal
microbial ecology. Appl. Environ. Microbiol. 54, 1079e1084.
Stecher, B., Chaffron, S., Kappeli, R., Hapfelmeier, S., Freedrich, S., Weber, T.C.,
Kirundi, J., Suar, M., McCoy, K.D., von Mering, C., Macpherson, A.J.,
Hardt, W.D., 2010. Like will to like: abundances of closely related species
can predict susceptibility to intestinal colonization by pathogenic and
commensal bacteria. PLoS Pathog. 6, e1000711.
Suryani, Kimura, T., Sakka, K., Ohmiya, K., 2003. Cloning, sequencing, and
expression of the gene encoding the Clostridium stercorarium alpha-
galactosidase Aga36A in Escherichia coli. Biosci. Biotechnol. Biochem.
67, 2160e2166.Thoden, J.B., Raushel, F.M., Wesenberg, G., Holden, H.M., 1999. The binding
of inosine monophosphate to Escherichia coli carbamoyl phosphate
synthetase. J. Biol. Chem. 274, 22502e22507.
Turroni, F., Bottacini, F., Foroni, E., Mulder, I., Kim, J.H., Zomer, A.,
Sanchez, B., Bidossi, A., Ferrarini, A., Giubellini, V., Delledonne, M.,
Henrissat, B., Coutinho, P., Oggioni, M., Fitzgerald, G.F., Mills, D.,
Margolles, A., Kelly, D., van Sinderen, D., Ventura, M., 2010. Genome
analysis of Bifidobacterium bifidum PRL2010 reveals metabolic pathways
for host-derived glycan foraging. Proc. Natl. Acad. Sci. 107, 19514e19519.
Tzortzis, G., Jay, A.J., Baillon, M.L., Gibson, G.R., Rastall, R.A., 2003.
Synthesis of alpha-galactooligosaccharides with alpha-galactosidase from
Lactobacillus reuteri of canine origin. Appl. Microbiol. Biotechnol. 63,
286e292.
Valentine, P.J., Gherardini, F.C., Salyers, A.A., 1991. A Bacteroides ovatus
chromosomal locus which contains an alpha-galactosidase gene may be
important for colonization of the gastrointestinal tract. Appl. Environ.
Microbiol. 57, 1615e1623.
Xu, J., Bjursell, M.K., Himrod, J., Deng, S., Carmichael, L.K., Chiang, H.C.,
Hooper, L.V., Gordon, J.I., 2003. A genomic view of the human-
Bacteroides thetaiotaomicron symbiosis. Science 299, 2074e2076.
Zeng, L., Burne, R.A., 2008. Multiple sugar: phosphotransferase system
permeases participate in catabolite modification of gene expression in
Streptococcus mutans. Mol. Microbiol. 70, 197e208.
Zhang, X., Vrijenhoek, J.E., Bonten, M.J., Willems, R.J., van Schaik, W., 2011.
A genetic element present on megaplasmids allows Enterococcus faecium to
use raffinose as carbon source. Environ. Microbiol. 13, 518e528.Zhao, H., Lu, L., Xiao, M., Wang, Q., Lu, Y., Liu, C., Wang, P., Kumagai, H.,
Yamamoto, K., 2008. Cloning and characterization of a novel alpha-
galactosidase from Bifidobacterium breve 203 capable of synthesizing
Gal-alpha-1,4 linkage. FEMS Microbiol. Lett. 285, 278e283.Zhou, J., Shi, P., Huang, H., Cao, Y., Meng, K., Yang, P., Zhang, R., Chen, X.,
Yao, B., 2010. A new a-galactosidase from symbiotic Flavobacterium sp.
TN17 reveals four residues essential for a-galactosidase activity of
gastrointestinal bacteria. Appl. Microbiol. Biotechnol. 88, 1297e1309.