
Enhancement of α- and β-Galactosidase Activity in Lactobacillus reuteri by Different Metal Ions
12
Enhancement of α- and β-Galactosidase Activity in Lactobacillus reuteri by Different Metal Ions Salam A. Ibrahim & Awfa Y. Alazzeh & Saddam S. Awaisheh & Danfeng Song & Abolghasem Shahbazi & Amer A. AbuGhazaleh Received: 25 March 2009 / Accepted: 31 August 2009 / Published online: 24 September 2009 # Humana Press Inc. 2009 Abstract The hydrolysis of oligosaccharides and lactose is of great importance to the food industry. Normally, oligosaccharides like raffinose, stachyose, and verbascose which are rich in different plants like soy bean are considered indigestible by the human gut. Moreover, many humans suffer from lactose intolerance due to the absence of effective enzyme that can digest lactose. α-Galactosidase can digest oligosaccharides like raffinose, while β-galactosidases can hydrolyze lactose. Therefore, selection of microorganisms safe for human use and capable of producing high levels of enzymes becomes an attractive task. The objective of this study was to investigate the enhancement of α- and β-galactosidase activity in Lactobacillus reuteri by different metal ions. Ten millimolar of Na + ,K + , Fe 2+ , and Mg 2+ and 1 mM of Mn 2+ were added separately to the growth culture of six strains of L. reuteri (CF2-7F, DSM20016, MF14-C, MM2-3, MM7, and SD2112). Results showed that L. reuteri CF2-7F had the highest α- and β-galactosidase activity when grown in the medium with added Mn 2+ ions (22.7 and 19.3 Gal U/ml, respectively). 0.0274% of Mn 2+ ions lead to 27, 18% enhancement of α- and β-galactosidase activity over the control group, and therefore, it could be added to the growth culture of CF2-7F to produce enhanced levels of α- and β-galactosidase activity. The addition of Fe 2+ led to a significant (P <0.01) decrease in the activity of both enzymes for most strains. This study shows that modified culture medium with that 0.0274% Mn 2+ can be used to promote the production for α- and β-galactosidase in L. reuteri CF2-7F, which may lead to enhancement of α- and β-galactosidase activity and have a good potential to be used in the food industry. Keywords α-Galactosidase . β-Galactosidase . Lactobacillus reuteri . Metal ions . Mn 2+ Biol Trace Elem Res (2010) 136:106–116 DOI 10.1007/s12011-009-8519-2 S. A. Ibrahim (*) : A. Y. Alazzeh : D. Song : A. Shahbazi North Carolina Agricultural and Technical State University, 163 Carver Hall, Greensboro, NC, USA e-mail: [email protected] S. S. Awaisheh Department of Nutrition and Food Technology, Mu’tah University, Karak, Jordan A. A. AbuGhazaleh Department of Animal Science, Food and Nutrition, Southern Illinois University, Carbondale, IL, USA
Transcript of Enhancement of α- and β-Galactosidase Activity in Lactobacillus reuteri by Different Metal Ions

Enhancement of α- and β-Galactosidase Activityin Lactobacillus reuteri by Different Metal Ions
Salam A. Ibrahim & Awfa Y. Alazzeh &
Saddam S. Awaisheh & Danfeng Song &
Abolghasem Shahbazi & Amer A. AbuGhazaleh
Received: 25 March 2009 /Accepted: 31 August 2009 /Published online: 24 September 2009# Humana Press Inc. 2009
Abstract The hydrolysis of oligosaccharides and lactose is of great importance to the foodindustry. Normally, oligosaccharides like raffinose, stachyose, and verbascose which arerich in different plants like soy bean are considered indigestible by the human gut.Moreover, many humans suffer from lactose intolerance due to the absence of effectiveenzyme that can digest lactose. α-Galactosidase can digest oligosaccharides like raffinose,while β-galactosidases can hydrolyze lactose. Therefore, selection of microorganisms safefor human use and capable of producing high levels of enzymes becomes an attractive task.The objective of this study was to investigate the enhancement of α- and β-galactosidaseactivity in Lactobacillus reuteri by different metal ions. Ten millimolar of Na+, K+, Fe2+,and Mg2+ and 1 mM of Mn2+ were added separately to the growth culture of six strains ofL. reuteri (CF2-7F, DSM20016, MF14-C, MM2-3, MM7, and SD2112). Results showedthat L. reuteri CF2-7F had the highest α- and β-galactosidase activity when grown in themedium with added Mn2+ ions (22.7 and 19.3 Gal U/ml, respectively). 0.0274% of Mn2+
ions lead to 27, 18% enhancement of α- and β-galactosidase activity over the controlgroup, and therefore, it could be added to the growth culture of CF2-7F to produceenhanced levels of α- and β-galactosidase activity. The addition of Fe2+ led to a significant(P<0.01) decrease in the activity of both enzymes for most strains. This study shows thatmodified culture medium with that 0.0274% Mn2+ can be used to promote the productionfor α- and β-galactosidase in L. reuteri CF2-7F, which may lead to enhancement of α- andβ-galactosidase activity and have a good potential to be used in the food industry.
Keywords α-Galactosidase .β-Galactosidase . Lactobacillus reuteri . Metal ions . Mn2+
Biol Trace Elem Res (2010) 136:106–116DOI 10.1007/s12011-009-8519-2
S. A. Ibrahim (*) :A. Y. Alazzeh : D. Song :A. ShahbaziNorth Carolina Agricultural and Technical State University, 163 Carver Hall, Greensboro, NC, USAe-mail: [email protected]
S. S. AwaishehDepartment of Nutrition and Food Technology, Mu’tah University, Karak, Jordan
A. A. AbuGhazalehDepartment of Animal Science, Food and Nutrition, Southern Illinois University, Carbondale, IL, USA

Introduction
One of the more challenging food consumption issues is how to improve the digestibility ofcomplex sugars. Raffinose, stachyose, and verbascose sugars that are abundant in foodssuch as soybeans, cabbage, brussels sprouts, and broccoli with α 1–6 linkages are notdigested by the humans, which often result in the build-up of intestinal gas and discomfort[1]. Similarly, “lactose intolerance” occurs when milk products are consumed, and thelactose (milk sugar) hydrolyzing enzyme is in short supply—commonly resulting in gastricsymptoms of abdominal pain, diarrhea, and/or flatulence.
Two enzymes that can address these problems are α-galactosidase (EC 3.2.1.22) andβ-galactosidase (EC 3.2.1.23). α-Galactosidase catalyzes the hydrolysis of oligo- andpolysaccharides containing terminal α-galactosyl groups by breaking the α 1–6 linkagebetween monosaccharide moieties [2]. β-Galactosidases catalyze the hydrolysis of oligo-and polysaccharides containing β-D-galactopyranosides by breaking the β 1–4 linkagebetween monosaccharide moieties in lactose [1].
Unfortunately, these enzymes are often lacking in the human intestine. α-Galactosidase enzyme does not occur naturally in the gut [3], and more than 75% ofadults worldwide have insufficient levels of β-galactosidase enzyme [4]. Together, thesetwo enzymes could improve the digestion of a range of nutritious foods made up ofcomplex sugars.
Therefore, developing a means by which these enzymes could be efficiently added to thedigestive system could be highly beneficial, including alleviating many of the symptomsresulting from the indigestibility of foods with high content of complex sugars. However,the addition of these enzymes to food products in a purified form is a complicated andexpensive process. Moreover, the purified form of these enzymes is not easily approved forhuman consumption [3]. What is needed then is a method by which these enzymes can bedelivered into the human gut in sufficient amounts and that will promote better functionalityof these enzymes to break down complex food sugars.
Probiotics are food grade bacteria that have been used safely and directly as foodsupplements [5]. Lactic Acid Bacteria (LAB), which reside normally in the small intestine,are one of the most commonly known probiotic bacteria used in the food industry [6].Several studies have shown that different LAB strains have good α- and β-galactosidaseproduction [1, 6–13, 16, 17]. Therefore, LAB could be used as carriers for the production ofdigestive enzymes such as α- and β-galactosidase. Several studies on the production ofα- and β-galactosidase with LAB, as a source material, have shown that the cultureconditions could be an important factor in enhancing the activity of both enzymes. It hasbeen shown that carbohydrate and protein sources in the growth culture are important forgrowth of bacteria and high activity of α- and β-galactosidase [7–10].
The production capacity of the hydrolyzing enzymes by LAB is strain specific [1, 5–10]. Lactobacillus reuteri has been shown to have a good potential for the production ofα- and β-galactosidase enzymes [11, 12]. We have previously shown that in a L. reuterigrowth culture, raffinose is essential to enhance the activity of α-galactosidase and thatlactose is essential to enhance β-galactosidase activity. We have also found that yeastextract was the best protein source to enhance the activity of both enzymes [13]. Apartfrom our recent studies, no information about how to enhance the production of α/β-galactosidases from L. reuteri, especially regarding the metal ion conditions in the growthmedia was available.
However, several studies have indicated that metal ions (Mg2+ and Mn2+) stimulatedthe growth of Lactobacillus bulgaricus GS [14] and Lactobacillus casei [15]. Metal ions
α- and β-Galactosidase Activity in Lactobacillus reuteri 107

might also increase the activity of β-galactosidases [16–20] when added into theenzymatic reaction mixture. Kim and Rajagopal [18] found that the presence of 1 mM ofFe2+ and Mn2+ in the reaction mixture increased the activity of β-galactosidase producedfrom Lactobacillus crispatus. Hus et al. [19] showed that reaction mixture fortified with100 mM of Na+ and K+ ions resulted in up to a 10-fold higher activity of β-galactosidaseenzyme produced from Bifidobacterium longum CCRC 15708. Other studies havedemonstrated that added Mg2+ enhanced the production of β-galactosidase fromLactobacillus acidophilus [11] and Bifidobacterium infantis [20], respectively. Thoseresults indicated that the metal ions are essential for the activity of galactosidase enzyme.
Because the optimum culture growth conditions of L. reuteri to support high activityof galactosidase are still unknown and because of the findings of metal ions on theactivity of the galactosidases, therefore, we need to test the possible enhancement of theseenzymes in L. reuteri strains by the addition of different metal ions to the culture media.In this study, we tried to further improve the growth culture of L. reuteri that wasestablished previously [12, 13] for further enhancement of α- and β-galactosidaseactivities. The objective of this study was to investigate the effects of different metal ions(Fe2+, Mg2+, Mn2+, Na+, and K+) on α- and β-galactosidase production in the growthculture of six strains of L. reuteri.
Materials and Methods
Maintaining and Preparation of Microorganisms
Six strains of L. reuteri (CF2-7F, DSM20016, MF14-C, MM2-3, MM7, and SD2112) wereobtained from BioGaia, Raleigh, NC, USA (Table 1). The sources of the strains were childfecal isolate (CF2-7F), mother fecal isolate (MF14C), and mother’s milk (DSM20016,MM2-3, MM7, and SD2112). These strains were maintained on Lactobacillus deManRogosa Sharpe (MRS) agar (Difco Laboratories, Becton Dickinson, Sparks, MD, USA) onPetri dishes at 4°C. The strains were activated before use by taking one bacterial colonyfrom the preserved MRS agar of each strain and then separately inoculating the colony intoLactobacillus MRS broth in anaerobic culture tubes at 37°C for 18 h. Following thispreparation, 0.1 ml of a 10−6 dilution of each incubated colony sample was plated onceagain in Lactobacillus MRS agar for 48 h, and activation of the strains was preparedfollowing the previously mentioned procedure. This repeated step was used to ensure thepurity of strains and to standardize the culture conditions of the used strains. The activatedstrains of the growth cultures were harvested by centrifugation (10,000× g for 10 min at4°C) and washed twice with 0.1% sterilized peptone water.
Strain no. Source/Reference
CF2F-7F Child fecal isolate
DSM20016 Mother’s milk
SD2112 Mother’s milk
MF14-C Mother fecal isolate
MM7 Mother’s milk
MM2-3 Mother’s milk
Table 1 Lactobacillus reuteriStrains and Sources that wereUsed in the Study
108 Ibrahim et al.

Culture Conditions
We diluted the harvested L. reuteri strains to 10−2 by sterilized deionized water, and then,we inoculated 0.1 ml of each strain into 10 ml of the tested cultures. The strains were testedfor enzymatic activity in a basic media that has been shown to support good growth ofL. reuteri [13]. This basic media consists of 1% lactose, 1% raffinose, 2.5% yeast extract,0.5% NaCH3COO, 0.01% MgSO4.7H2O, 0.005% MnSO4.4H2O, 0.3% K2HPO4, 0.1%KH2PO4, 0.1% Tween 80 (Fisher Scientific, Fair Lawn, NJ, USA) and 0.01% L-cysteine(Sigma, St. Louis, MO, USA). The basic media with no additional ions added representedthe control condition of this experiment. The metal ions were added separately to the basicmedia. To study the enhancement of α- and β-galactosidases activity by the addition ofmetal ions, 10 mM of either FeSO4, MgSO4.7H2O, K2SO4, Na2SO4, or 1 mM ofMnSO4.4H2O (Fisher Scientific, Fair Lawn, NJ, USA) were added to the growth culture.Based on the findings of Kim and Rajagopal [18] and Hsu et al. [19] who showed thatcertain metal ions (from 1 to 100 mM) led to enhancement of galactosidases activities whenadded into the enzymatic reaction mixture, we used 10 mM of most metal ions to avoid thehypertonic pressure on the bacterial cells. We used 1 mM of MnSO4 in the growth culturesince higher concentrations of this metal did not dissolve completely in the growth culture.The strains were tested for enzyme activity after 18-h incubation period at 37°C.
Growth Over Time
The growth of L. reuteri strains was monitored by measuring the optical density (OD) at3-h interval during the incubation period using a spectronic 21 Milton Roy spectropho-tometer (Thermo Electron Scientific Co., Madison, WI, USA) at the wavelength of610 nm.
Determination of α- and β-Galactosidases Activity
L. reuteri strains were first harvested by centrifugation (10,000× g for 10 min at 4°C) ofthe culture media, discarding the supernatant and washing the precipitated strains twicewith phosphate buffer (pH 6.8) based on Hsu et al. [7] with a slight modification. Thebacterial cells were resuspended in 1 ml of phosphate buffer in Eppendorf tubescontaining 0.1 mm glass beads (BioSpec Products Inc., Bartlesville, OK, USA) andmaintained in an ice bath. Samples were treated for 3 min with a bead beater (MiniBeadBeater-8, BioSpec Products Inc., Bartlesville, OK, USA). At the end of each minute ofbead beater treatment, samples were removed from the bead beater and placed for 15 s inan ice bath. Samples were then centrifuged (12,000× g, 30 min) to obtain the supernatant,which served as the enzyme source. These samples were stored at −80°C until enzymeactivity was assayed.
α-Galactosidase activities was assayed according to Food Chemicals Codex [21]. β-Galactosidase activities was assayed according to the method described by Hsu et al. [7]and Nagy et al. [22]. Activity of α-galactosidase and β-galactosidase activity were testedon ρ-nitrophenyl-α-D-galactopyranoside and ο-nitrophenyl β-D-galactopyranoside, respec-tively. After reaction at 37°C, solution were added into the reaction mixture to stop reaction.Absorbance was measured at 405 nm for α-galactosidase and 420 nm for β-galactosidasewith a spectrophotometer (Model Genesys 10Vis, Thermospectronic, Rochester, NY, USA).One galactosidase activity unit (Gal U) was defined as the amount of enzyme catalyzing theformation of 1 μmol of nitrophenyl per minute under the assay condition.
α- and β-Galactosidase Activity in Lactobacillus reuteri 109

Statistical Analysis
Data analyses were focused on determining if the addition of different metal ions resulted insignificantly higher activity of both enzymes over that of the control group. Statisticalanalysis of data was performed using the SAS General Linear Model (GLM) program [23].Least square means of duplicate samples were calculated and Duncan’s Multiple RangeTest was used to determine significant (P<0.01) differences. For each growth condition,duplicate samples were tested for the activity of both enzymes.
Results and Discussion
Growth Over Time
Figure 1 shows the survival and growth of L. reuteri strains in the presence of differentmetal ions during the incubation at 37°C. All strains of L. reuteri tested were not killed bythe addition of the metal ion. They grew rapidly and showed similar growth curve as theircontrol groups. In the media without metal ion, L. reuteri continued to grow and reached anOD of 1.6 within 18 h. In the media with the added Mn2+, L. reuteri grew fast and reachedan OD of 1.8 within 18 h. While in the media with added Fe2+, L. reuteri grew slowly andreached an OD of 1.2 only.
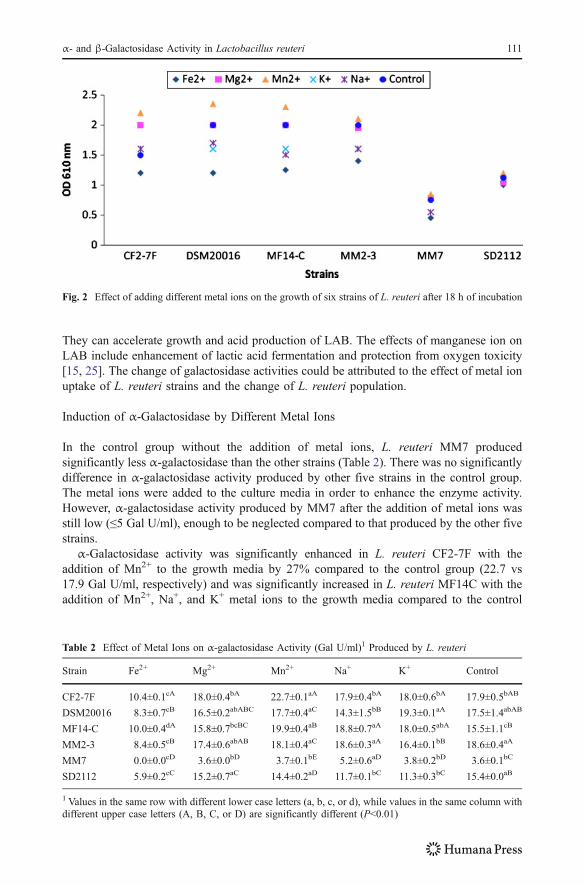
All strains of L. reuteri tested grew significantly better (P<0.05) in the media with theaddition of Mn2+, while they all grew slower in the media with the addition of Fe2+
compared to those grew in the control media (Fig. 2). Among all the tested strains, L.reuteri CF2-7F were sensitive to the added metal ions. The growth of L. reuteri CF2-7Fenhanced by the addition of Mn2+, Mg2+, Na+, and K+. Figures 1 and 2 showed that theaddition of Fe2+ in the media slowed down the growth, but all strains of L. reuteri testedremained viable and kept growing within 18-h incubation period. In addition, the additionof Mn2+ enhanced the growth of all L. reuteri strains tested in this experiment. Manganeseions are required in trace quantities for growth and metabolic activities of LAB [15, 24].
Fig. 1 Effect of adding different metal ions on the growth pattern of L. reuteri (results of six strains ofL. reuteri were pooled for each metal ion)
110 Ibrahim et al.
They can accelerate growth and acid production of LAB. The effects of manganese ion onLAB include enhancement of lactic acid fermentation and protection from oxygen toxicity[15, 25]. The change of galactosidase activities could be attributed to the effect of metal ionuptake of L. reuteri strains and the change of L. reuteri population.
Induction of α-Galactosidase by Different Metal Ions
In the control group without the addition of metal ions, L. reuteri MM7 producedsignificantly less α-galactosidase than the other strains (Table 2). There was no significantlydifference in α-galactosidase activity produced by other five strains in the control group.The metal ions were added to the culture media in order to enhance the enzyme activity.However, α-galactosidase activity produced by MM7 after the addition of metal ions wasstill low (≤5 Gal U/ml), enough to be neglected compared to that produced by the other fivestrains.
α-Galactosidase activity was significantly enhanced in L. reuteri CF2-7F with theaddition of Mn2+ to the growth media by 27% compared to the control group (22.7 vs17.9 Gal U/ml, respectively) and was significantly increased in L. reuteri MF14C with theaddition of Mn2+, Na+, and K+ metal ions to the growth media compared to the control
Fig. 2 Effect of adding different metal ions on the growth of six strains of L. reuteri after 18 h of incubation
Table 2 Effect of Metal Ions on α-galactosidase Activity (Gal U/ml)1 Produced by L. reuteri
Strain Fe2+ Mg2+ Mn2+ Na+ K+ Control
CF2-7F 10.4±0.1cA 18.0±0.4bA 22.7±0.1aA 17.9±0.4bA 18.0±0.6bA 17.9±0.5bAB
DSM20016 8.3±0.7cB 16.5±0.2abABC 17.7±0.4aC 14.3±1.5bB 19.3±0.1aA 17.5±1.4abAB
MF14-C 10.0±0.4dA 15.8±0.7bcBC 19.9±0.4aB 18.8±0.7aA 18.0±0.5abA 15.5±1.1cB
MM2-3 8.4±0.5cB 17.4±0.6abAB 18.1±0.4aC 18.6±0.3aA 16.4±0.1bB 18.6±0.4aA
MM7 0.0±0.0cD 3.6±0.0bD 3.7±0.1bE 5.2±0.6aD 3.8±0.2bD 3.6±0.1bC
SD2112 5.9±0.2cC 15.2±0.7aC 14.4±0.2aD 11.7±0.1bC 11.3±0.3bC 15.4±0.0aB
1 Values in the same row with different lower case letters (a, b, c, or d), while values in the same column withdifferent upper case letters (A, B, C, or D) are significantly different (P<0.01)
α- and β-Galactosidase Activity in Lactobacillus reuteri 111

group (P<0.01; Table 2, Fig. 3). The addition of Fe2+ led to the slow growth and asignificant (P<0.01) decrease in the activity of α-galactosidase produced by all strainstested. The inhibition of growth and enzyme activity also was found in all strains grew inthe media with the addition of Cu2+ (data not shown). The addition of Mg2+ did not causesignificantly change in the activity of α-galactosidase in all the tested strains.
Previous studies showed that the addition of metal ions to the reaction mixture on α-galactosidase activity is strain specific and is affected by the concentration of the mineraladded. Mn2+ had the potential to increase the activity of α-galactosidase when added intothe reaction mixture. Garro et al. [26] added 1 mM Mn2+ in the reaction mixture when theymeasured the activity of α-galactosidase produced by Lactobacillus fermentum CRL 236and L. fermentum CRL 251 and found an increase of α-galactosidase activity (13.7 and4.8%, respectively) over the control group. Garro et al. [27] showed that the addition of1 mM Mn2+ in the reaction mixture had a positive effect on α-galactosidase activity ofB. longum. The addition of Mn2+ to the growth media of L. reuteri CF2-7F and MF14C inour experiment leads to the similar results of increased α-galactosidase activity (over 27%)compared to their control group. However, the addition of Mn2+ to the growth media ofDSM20016, MM23, MM7, and SD2112 did not significantly affect α-galactosidaseactivity. Similarly, Garro et al. [28] showed that 10 mM of Mn2+ did not lead to an increasein α-galactosidase activity in L. fermentum CRL 251. Fridjonsson et al. [29], Kondoh et al.[30], and Carrera-Silva et al. [31] showed that 1 mM of Mn2+ did not significantly affectα-galactosidase activity in Streptomyces coelicolor, Thermus brockianus ITI360, and L.fermentum CRL 722, respectively. The addition of Na+ and K+ in the media enhanced theactivity of α-galactosidase produced by L. reuteri MF14C strain in our study. However, theaddition of Na+ and K+ did not show significant effect on α-galactosidase activity inprevious mentioned reports [27–31]. But the addition of Na+ and K+ did significantlyenhance the activity of β-galactosidase produced from B. longum CCRC 15708 [19].
In our effort to produce higher α-galactosidase activity from different strains of L.reuteri by optimization of medium components, such as using different carbohydrate andnitrogen sources, the highest α-galactosidase activity in optimized culture conditions that
Fig. 3 Relative activity (%) of α-galactosidase activity produced by L. reuteri with added different metalions compared to the control group without metal ions
112 Ibrahim et al.

were similar to the control group of this study was 21.0 Gal U/ml [13]. The addition of1 mM Mn2+ to the growth media lead to further induction of 8.1% over the best result forα-galactosidase activity (22.7 Gal U/ml).
Induction of β-Galactosidase by Different Metal Ions
In the control group without the addition of metal ions, L. reuteri MM7 producedsignificantly less β-galactosidase than the other strains (Table 3). There was no significantlydifference in β-galactosidase activity produced by other tested strains in the control group.MM7 was neglected to do further investigation due to the significantly low activity ofβ-galactosidase at all conditions with/without metal ions.
β-Galactosidase activity was significantly enhanced in L. reuteri CF2-7F with theaddition of Mn2+ to the growth media by 18% compared to the control group (19.3 vs16.3 Gal U/ml, respectively) and was significantly increased in L. reuteri MF14C with theaddition of Mn2+, Na+, and Mg2+ metal ions to the growth media compared to the controlgroup (P<0.01; Table 3, Fig. 4). The addition of Fe2+ led to the slow growth and decreasein the activity of β-galactosidase produced by all strains tested. The inhibition of growthand enzyme activity also was found in all strains grown in the media with the addition ofCu2+ (data not shown). The addition of Mg2+ did not cause significant change in theactivity of β-galactosidase in all the tested strains except in MF14C.
Previous studies showed that the effect of metal ions on β-galactosidase activity, as inthe case of α-galactosidase, varied with the bacterial strain and the concentration of themetal ion used. Mn2+ had the potential to increase the activity of β-galactosidase whenadded into the reaction mixture. Kim and Rajagopal [18] found that 1 mM of Mn2+ lead toan increase of 27% on β-galactosidase activity in L. crispatus over the control groupwithout addition of metal ions. The addition of Mn2+ to the growth media of L. reuteri CF2-7F and MF14C in our experiment lead to the similar results of increased β-galactosidaseactivity (18% and 39%, respectively) compared to their control group. However, theaddition of Mn2+ to the growth media of L. reuteri MM23 leads to decreased β-galactosidase activity (−23%). Hsu et al. [19] also showed that 1 mM of Mn2+ lead todecreased β-galactosidase activity in B. longum CCRC 15708. Budriene et al. [32] showedthat β-galactosidase activity from Penicillium canescens was not sensitive to 1 mM of Na+,K+, Cu2+, Fe2+, Mn2+, and Mg2+ ions in the growth culture.
The highest β-galactosidase activity in our previous study [13] was 18.1 Gal U/mlwhen grew on the similar conditions as the control group of this study. The addition of
Table 3 Effect of Metal Ions on β-galactosidase Activity (Gal U/ml)1 Produced by L. reuteri
Strain Fe2+ Mg2+ Mn2+ Na+ K+ Control
CF2-7F 10.8±0.7cCD 16.6±0.5bA 19.3±0.1aA 16.2±0.6bBC 13.8±0.1bC 16.3±0.5bB
DSM20016 10.4±1.3bD 17.0±0.3aA 17.1±0.2aC 16.9±0.0aB 18.8±0.6aA 17.1±0.3aAB
MF14-C 12.8±0.4bBC 17.7±0.8aA 18.4±0.3aB 18.9±0.7aA 13.5±0.6bC 13.2±0.4bC
MM2-3 13.9±0.2bB 18.8±1.0aA 13.6±0.2bE 15.3±0.1bC 14.8±0.1bBC 17.6±0.4aA
MM7 0.0±0.0dE 0.3±0.0cB 0.3±0.0cF 0.6±0.0bD 0.1±0.0dD 0.8±0.1aD
SD2112 17.1±0.3abA 18.2±1.4abA 15.8±0.1bD 18.9±0.5aA 16.0±0.9bAB 17.1±0.0abAB
1 Values in the same row with different lower case letters (a, b, c, or d), while values in the same column withdifferent upper case letters (A, B, C, or D) are significantly different (P<0.01)
α- and β-Galactosidase Activity in Lactobacillus reuteri 113

1 mM Mn2+ metal ions in the growth media of CF2-7F leads to a further enhancement ofβ-galactosidase activity (19.3 Gal U/ml) of 7.0% over the best result for β-galactosidaseactivity.
The discrepancies between the different results could be due to several factors. First,the addition of metal ions in the growth media is an innovative approach to enhance theenzyme activity, which is different from adding metal ions into the enzymatic reactionmixture. Bacteria will survive, uptake, and utilize the metal ions to produce higheractivity of the enzyme. Second, the effect of a certain metal ion varies among strainsand species of bacteria. Concentration of the metal ions used in the different studiesvaries, which could explain the discrepancies of the results. Lastly, other cultureconditions used in the growth media of the different bacteria strains, such as pH, maylead to increase or decrease in the metal uptake of bacteria, and thus, metal effect onenzyme activity could vary.
Our results suggest the use of a single strain (CF2-7F) to produce enhanced levels of α-and β-galactosidase enzymes when grown in culture conditions similar to the control groupwith added Mn+2 ions, and therefore, this strain should acquire more attention for itspossible probiotic applications in food industry.
Conclusion
The enhancement of α- and β-galactosidase activity due to the addition of metal ions isstrain specific. The addition of 0.027% Mn2+ ions to the growth culture of L. reuteri CF2-7F leads to an enhancement of α- and β-galactosidase activity over any other strain andother ion metal studied. This suggests the use of 1 mM Mn2+ ions be added to the controlmedia of CF2-7F to enhance the activity of both enzymes, which may have a good potentialto be used in the food industry.
Acknowledgment We would like to thank Dr. Keith Schimmel (Energy and Environmental Studies at NC,USA, A&T) for his support and for Dr. K.J. Gruber from the Agricultural Research Program at NorthCarolina A&T State University for his review and comments on this manuscript.
Fig. 4 Relative activity (%) of β-galactosidase activity produced by L. reuteri with added different metalions compared to the control group without metal ions
114 Ibrahim et al.

References
1. Donkor ON, Henriksson A, Vasiljevic T, Shah NP (2007) α-Galactosidase and proteolytic activities ofselected probiotic and dairy cultures in fermented soymilk. Food Chem 104(1):10–20
2. Kim WD, Kobayashi O, Kaneko S, Sakakibara Y, Park GG, Kusakabe I, Tanaka H, Kobayashi H (2002)α-Galactosidase from cultured rice (Oryza sativa L. var. Nipponbare) cells. Phytochemistry 61(6):621–630
3. Song D, Chang SKC (2006) Enzymatic degradation of oligosaccharides in pinto bean flour. J Agric FoodChem 54:1296–1301
4. Pribila BA, Hertzler SR, Martin BR, Weaver CM, Savaiano DA (2000) Improved lactose digestion andintolerance among African-American adolescent girls fed a dairy rich-diet. J Am Diet Assoc 100(5):524–528
5. Speck ML, Dobrogosz WJ, Casas IA (1993) Lactobacillus reuteri in food supplementation. Food Tech47(7):90–94
6. Xanthopoulos V, Litopoulou-Tzanetaki E, Tzanetakis N (2000) Characterization of lactobacillus isolatesfrom infant faeces as dietary adjuncts. Food Microbiol 17(2):205–215
7. Hsu CA, Yu RC, Chou CC (2005) Production of β-galactosidase by bifidobacteria as influenced byvarious culture conditions. Int J Food Microbiol 104(2):197–206
8. Xiao M, Tanaka K, Qian XM, Yamamoto K, Kumagai H (2000) High-yield production andcharacterization of a-galactosidase from Bifidobacterium breve grown on raffinose. Biotechnol Lett 22(9):747–751
9. Tzortzis G, Jay AJ, Baillon MLA, Gibson GR, Rastall RA (2003) Synthesis of α-galactooligosaccharideswith α-galactosidase from Lactobacillus reuteri of canine origin. Appl Microbiol Biotechnol 63(3):286–292
10. Bury D, Geciova J, Jelen P (2001) Effect of yeast extract supplementation on beta-galactosidase activityof Lactobacillus delbrueckii subsp. bulgaricus 11842 grown in whey. Czech Journal of Food Sciences 19(5):166–170
11. Nguyen TH, Splechtna B, Yamabhai M, Haltrich D, Peterbauer C (2007) Cloning and expression of theβ-galactosidase genes from Lactobacillus reuteri in Escherichia coli. J Biotechnol 129(4):581–591
12. Alazzeh AY, Ibrahim SA, Song D, Shahbazi A, AbuGhazaleh AA (2009a) Screening for α- and β-galactosidases in Lactobacillus reuteri compared to different strains of Bifidobacteria. Milchwissenschaft(in press)
13. Alazzeh AY, Ibrahim SA, Song D, Shahbazi A, AbuGhazaleh AA (2009) Carbohydrate and proteinsources influence the induction of α- and β-galactosidases in Lactobacillus reuteri. Food Chem 117(15):645–659
14. Weinman DE, Morris GK, Williams WL (1964) Unidentified growth factor for a lactic acid bacterium. JBacteriol 87(2):263–269
15. Woolley DW (1941) Manganese and the growth of lactic acid bacteria. J Biol Chem 140:311–31216. Tochikura T, Sakai K, Fujiyoshi T, Tachiki T, Kumagai H (1986) p-Nitrophenyl glycoside-hydrolyzing
activities in bifidobacteria and characterization of ß-D-galactosidase of Bifidobacterium longum 401. AgrBiol Chem 50(9):2279–2286
17. Nakayama T, Amachi T (1999) β-Galactosidase, enzymology. In: Flickinger MC, Drew SW (eds)Encyclopedia of bioprocesstechnology: fermentation, biocatalysis, and bioseparation. John Willey, NewYork, pp 1291–1305
18. Kim JW, Rajagopal SN (2000) Isolation and characterization of β-galactosidase from Lactobacilluscrispatus. Folia Microbiol 45(1):29–34
19. Hsu CA, Yu RC, Chou CC (2006) Purification and characterization of a sodium-stimulated β-galactosidase from Bifidobacterium longum CCRC 15708. World J Microbiol Biotechnol 22(4):355–361
20. Roy D, Blanchette L, Savoie L, Ward P, Chevalier P (1992) α- and β-galactosidase properties ofBifidobacterium infantis. Milchwissenschaft 47(1):18–21
21. Food Chemical Codex FCC (1981) National Academy of Sciences. National Academies Press,Washington DC
22. Nagy Z, Kiss T, Szentirmai A, Biró S (2001) β-Galactosidase of Penicillium chrysogenum: productionpurification and characterization of the enzyme. Protein Expr Purif 21(1):24–29
23. SAS Institute Inc (1999) SAS/STAT® User’s Guide. SAS Institute Inc., Cary24. Raccach M (1983) Fermentation method using a selected Lactobacillus. U.S. patent 4, 407, 383, 4 Oct.
198325. Kang DH, Fung DYC (2000) Stimulation of starter culture for further reduction of food borne pathogens
during salami fermentation. J Food Prot 63(11):1492–149526. Garro MS, Giori GS, Valdez GF, Oliver G (1993) Characterization of alpha-galactosidase from
Lactobacillus fermentum. J Appl Microbiol 75(5):485–488
α- and β-Galactosidase Activity in Lactobacillus reuteri 115

27. Garro MS, de Giori GS, de Valdez GF, Oliver G (1994) α-D-galactosidase (EC 32122) fromBifidobacterium longum. Lett Appl Microbiol 19(1):16–19
28. Garro MS, de Valdez GF, Oliver G, de Giori GS (1996) Purification of α-galactosidase fromLactobacillus fermentum. J Biotechnol 45(2):103–109
29. Fridjonsson O, Watzlawick H, Gehweiler A, Rohrhirsch T, Mattes R (1999) Cloning of the geneencoding a novel thermostable α-Galactosidase from Thermus brockianus ITI360. Appl EnvironMicrobiol 65(9):3955–3963
30. Kondoh K, Morisaki K, Kim WD, Kotwal SM, Kaneko S, Kobayashi H (2005) Expression ofStreptomyces coelicolor α-galactosidase gene in Escherichia coli and characterization. Food Sci TechRes 11(2):207–213
31. Carrera-Silva EA, Silvestroni A, LeBlanc JG, Piard JC, de Giori GS, Sesma F (2006) A thermostableα-galactosidase from Lactobacillus fermentum CRL722: genetic characterization and main properties.Curr Microbiol 53(5):374–378
32. Budriene S, Gorochovceva N, Romaskevic T, Yugova LV, Miezeliene A, Dienys G, Zubriene A (2005)β-Galactosidase from Penicillium canescens properties and immobilization. Cent Eur J Chem 3(1):95–105
116 Ibrahim et al.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.




















