
Production, purification, characterization, immobilization, and application of β -galactosidase: a...
19
Special theme review Production, purification, characterization, immobilization, and application of β-galactosidase: a review Arijit Nath, 1† Subhoshmita Mondal, 1† Sudip Chakraborty, 1,2 * Chiranjib Bhattacharjee 1 and Ranjana Chowdhury 1 1 Chemical Engineering Department, Jadavpur University, Kolkata, West Bengal 700032, India 2 Department of Chemical Engineering and Materials, University of Calabria, Cubo-44C, Rende 87036,CS, Italy Received 20 June 2013; Revised 29 September 2013; Accepted 5 October 2013 ABSTRACT: Biopharmaceuticals are new categorizing biomolecules, which are the results of incredible proliferation in the field of biotechnology. One of the challenging biomolecule β-galactosidase (β-galactosidase galacto hydrolysase, trivially lactase) catalyzes hydrolysis of lactose to produce glucose and galactose, and in some cases, it takes part in transgalactosylation reaction that produces functional food galato-oligosaccharide. A wide variety of strategies had been already attempted for production of β-galactosidase through fermentative route. Beside the upstream process, downstream technology towards purification and immobilization of target enzyme also create great attentions. Subsequently, its wide applications in the field of food, biopharmaceuticals, dairy, diagnosis, and waste treatment boost up biotechnological economy as well as zero-effluent discharge. In dairy industry, β-galactosidase has been used to degrade lactose, prevent crystallization of lactose, improve sweetness, and increase the solubility of milk product, otherwise which would be an environmental pollutant. In food and pharmaceutical industries, β-galactosidase has been used to prepare low lactose- containing food products for low lactose-tolerant people. Therefore, it is obvious to elucidate different technological aspects of β-galactosidase, which may provide a great knowledge in educational and industrial field. Taking the enzyme into account, a ready review has been made about its production, purification, characterization, and immobilization technology. The review also addresses wide applications of β-galactosidase in different fields. © 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Keywords: β-galactosidase; production; purification; characterization; immobilization; application INTRODUCTION Emergence of biotechnology, which brings a boon to biological origin, possesses a challenging revolution to the categorical field, biopharmaceuticals. There is enormous range of therapeutics that involves biopharmaceuticals, and it combines biomolecular forms, which extensively give rise to the development of the microbial synthesis of diverse enzymes and metabolites. Intellectual revolution with new visions and hopes by dispensation of any method of bioprocess that deals with the design and development of equipments for the manufacturing of various products, such as food-beverages, sera, new medicines, semi- synthetic organs, antibiotics, and enzymes from biological sources creates a great attention. This is responsible for the explosion of various biotechnological processes used in industries for large-scale production of biological products and optimization of their yields as well as the qualities of end products. [1] The lactose-hydrolyzing enzyme β-galactosidase has been accepted since long as an important ingredient in dairy, food processing, and pharmaceutical industries. β-galactosidase catalyzes hydrolysis of lactose into glucose and galactose and also takes part in transgalactosylation reaction that produces galato- oligosaccharide (GOS) (e.g., Gal (β1 → 3) Gal (β1 → 4) Gal (β1 → 6)). [2,3] One of the most common disease hypolactasia (lactose intolerance) or lactose maldigestion is caused by insufficient synthesis of lactose-cleaving enzyme β-galactosidase in the brush border membrane of mucosa of the small intestine. In most cases, this causes several symptoms, which may include abdominal bloating and cramps, flatulence, diarrhea, nausea, rumbling stomach, or vomiting after consuming significant amounts of lactose. [4] In most of the human races, β-galactosidase is found to be lost during the first or second decade of life, and only some isolates are seen among people of Northern European origin and their overseas descendants. African and Indian communities maintain a high intestinal lactase *Correspondence to: Sudip Chakraborty, Chemical Engineering Department, Jadavpur University, Kolkata, West Bengal 700032, India. E-mail: [email protected] † Authors have similar contribution © 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Curtin University is a trademark of Curtin University of Technology ASIA-PACIFIC JOURNAL OF CHEMICAL ENGINEERING Asia-Pac. J. Chem. Eng. (2014) Published online in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/apj.1801
Transcript of Production, purification, characterization, immobilization, and application of β -galactosidase: a...

Special theme review
Production, purification, characterization, immobilization,and application of β-galactosidase: a review
Arijit Nath,1† Subhoshmita Mondal,1† Sudip Chakraborty,1,2* Chiranjib Bhattacharjee1 and Ranjana Chowdhury1
1Chemical Engineering Department, Jadavpur University, Kolkata, West Bengal 700032, India2Department of Chemical Engineering and Materials, University of Calabria, Cubo-44C, Rende 87036,CS, Italy
Received 20 June 2013; Revised 29 September 2013; Accepted 5 October 2013
ABSTRACT: Biopharmaceuticals are new categorizing biomolecules, which are the results of incredible proliferation in thefield of biotechnology. One of the challenging biomolecule β-galactosidase (β-galactosidase galacto hydrolysase, triviallylactase) catalyzes hydrolysis of lactose to produce glucose and galactose, and in some cases, it takes part intransgalactosylation reaction that produces functional food galato-oligosaccharide. A wide variety of strategies had beenalready attempted for production of β-galactosidase through fermentative route. Beside the upstream process, downstreamtechnology towards purification and immobilization of target enzyme also create great attentions. Subsequently, its wideapplications in the field of food, biopharmaceuticals, dairy, diagnosis, and waste treatment boost up biotechnologicaleconomy as well as zero-effluent discharge. In dairy industry, β-galactosidase has been used to degrade lactose, preventcrystallization of lactose, improve sweetness, and increase the solubility of milk product, otherwise which would be anenvironmental pollutant. In food and pharmaceutical industries, β-galactosidase has been used to prepare low lactose-containing food products for low lactose-tolerant people. Therefore, it is obvious to elucidate different technological aspectsof β-galactosidase, which may provide a great knowledge in educational and industrial field. Taking the enzyme into account,a ready review has been made about its production, purification, characterization, and immobilization technology. Thereview also addresses wide applications of β-galactosidase in different fields. © 2014 Curtin University of Technologyand John Wiley & Sons, Ltd.
Keywords: β-galactosidase; production; purification; characterization; immobilization; application
INTRODUCTION
Emergence of biotechnology, which brings a boon tobiological origin, possesses a challenging revolution tothe categorical field, biopharmaceuticals. There isenormous range of therapeutics that involvesbiopharmaceuticals, and it combines biomolecularforms, which extensively give rise to the developmentof the microbial synthesis of diverse enzymes andmetabolites. Intellectual revolution with new visionsand hopes by dispensation of any method of bioprocessthat deals with the design and development ofequipments for the manufacturing of various products,such as food-beverages, sera, new medicines, semi-synthetic organs, antibiotics, and enzymes frombiological sources creates a great attention. This isresponsible for the explosion of various biotechnologicalprocesses used in industries for large-scale production of
biological products and optimization of their yields aswell as the qualities of end products.[1]
The lactose-hydrolyzing enzyme β-galactosidase hasbeen accepted since long as an important ingredient indairy, food processing, and pharmaceutical industries.β-galactosidase catalyzes hydrolysis of lactose intoglucose and galactose and also takes part intransgalactosylation reaction that produces galato-oligosaccharide (GOS) (e.g., Gal (β1→ 3) Gal(β1→ 4) Gal (β1→ 6)).[2,3] One of the most commondisease hypolactasia (lactose intolerance) or lactosemaldigestion is caused by insufficient synthesis oflactose-cleaving enzyme β-galactosidase in the brushborder membrane of mucosa of the small intestine. Inmost cases, this causes several symptoms, which mayinclude abdominal bloating and cramps, flatulence,diarrhea, nausea, rumbling stomach, or vomiting afterconsuming significant amounts of lactose.[4] In mostof the human races, β-galactosidase is found to be lostduring the first or second decade of life, and only someisolates are seen among people of Northern Europeanorigin and their overseas descendants. African andIndian communities maintain a high intestinal lactase
*Correspondence to: Sudip Chakraborty, Chemical EngineeringDepartment, Jadavpur University, Kolkata, West Bengal 700032,India. E-mail: [email protected]†Authors have similar contribution
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd.Curtin University is a trademark of Curtin University of Technology
ASIA-PACIFIC JOURNAL OF CHEMICAL ENGINEERINGAsia-Pac. J. Chem. Eng. (2014)Published online in Wiley Online Library(wileyonlinelibrary.com) DOI: 10.1002/apj.1801

activity throughout life. According to reviews ofScrimshaw, Murray, 1988 and Sahi, 1994, the globalprevalence of lactose maldigestion are above 50% inSouth America, Africa, and Asia reaching almost100% in some Asian countries such as China. In theUSA, prevalence is 15% among Whites, 53% amongMexican-Americans, and 80% in the Blackpopulation. In Europe, it varies from around 2% inScandinavia to about 70% in Sicily. Australia andNew Zealand have prevalence of 6% and 9%,respectively.[5,6] β-galactosidase has been used inbiopharmaceutical, food, and dairy industries to preventcrystallization of lactose, to improve sweetness, toincrease the solubility of milk product, to prepare lowlactose-containing food products for low lactose-tolerantpeople, and for the utilization of cheese whey, whichwould otherwise be an environmental pollutant.[7,8]
Therefore, it is obvious to elucidate differenttechnological aspects of β-galactosidase with specialinterest in production, purification, characterization,immobilization technology as well as its wideapplications in different process industries. The presentreview may provide a great knowledge with fill-up gapsof educational field and industrial sector.
SYNTHESISMECHANISMOF β-GALACTOSIDASE
The control system of β-galactosidase synthesis was firstworked out by Jacob and Monod in1961, at themolecular level. In prokaryotic type of gene expression,the lac operon showed inducible system with the controlof enzymes that are produced in the presence oflactose.[9] Presences of lac operon in Escherichia coliand Bacillus sp. including lactic acid bacteria havepotential of synthesizing intracellular β-galactosidase.Meanwhile, lactose metabolism in E. coli synthesizesseveral proteins, such as β-galactosidase, which convertslactose into glucose and galactose, β-galactosidepermease that transports lactose into the cellular system,
and the function of β-galactoside transacetylase is yetto be known. The construction of lac operon isdescribed in Fig. 1.For any consortia, all the genes involved in controlling
this pathway are located next to each other on thechromosome, and together they form an operon.Generally, β-galactosidase synthesis has been oftendemonstrated considering E. coli as a modelmicroorganism. The genetic switches can also combinepositive and negative controls. The lac operon consistsof three structural genes and a promoter, a terminator,regulator, and an operator. The three structural genes arelacZ, lacY, and lacA. Regulation for the specific controlof the lac genes depends on the availability of thesubstrate lactose. The proteins are not produced by thebacterium when lactose is unavailable as a carbon source.The lac genes are organized into an operon oriented in thesame direction, immediately adjacent on the chromosomeand are co-transcribed into a single polycistronic mRNAmolecule. The lac operon in E. coli, unlike trp operon,works under both negative and positive transcriptionalcontrols by the lac repressor protein and cataboliteactivator protein (CAP), respectively. CAP enablesbacteria to use alternative carbon sources such as lactosein the absence of glucose. The CAP cannot induceexpression if lactose is not present, and the lac repressorensures that the lac operon is shutoff in the absence oflactose. This arrangement enables the lac operon torespond to and integrate two different signals, so that itis expressed only when two conditions are met, i.e.,lactose must be present and glucose must be absent.Transcription of all genes starts with the binding of theenzyme RNA polymerase (RNAP), a DNA-bindingprotein that binds to a specific DNA binding site, thepromoter, immediately upstream of the genes. Bindingof RNA polymerase to the promoter is aided by the cyclicadenosine monophosphate (cAMP)-bound CAP (alsoknown as the cAMP receptor protein). From this position,RNAP proceeds to transcribe all three genes (lacZ, lacY,and lacA) into mRNA.[11]
Figure 1. Construction of lac operon (Figure adapted from K. L. Anderson, G. Purdom(2008)).[10]
A. NATH ET AL. Asia-Pacific Journal of Chemical Engineering
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

Control mechanism of lac operon
In the absence of lactose, the regulatory response tolactose uses an intracellular regulatory protein, whichis called the lactose repressor to hinder the synthesisof β-galactosidase. The lacI gene coding for therepressor lies nearby the lac operon and is alwaysexpressed (constitutive). If lactose is absent in thegrowth medium, the repressor binds very tightly to ashort DNA sequence just downstream of the promoternear the beginning of lacZ called the lac operator.The repressor binding to the operator interferes withbinding of RNAP to the promoter and therefore mRNAencoding LacZ and LacY are only made at very lowlevels. During the cells growth in the presence of lactose,a lactose metabolite allolactose, which is a combinationof glucose and galactose, binds to the repressor, causinga change in its shape. Thus, the altered repressor isunable to bind to the operator, allowing RNAP totranscribe the lac genes and thereby leading to higherlevels of the encoded proteins. The schematic diagramsof the proposed phenomena are described in Fig. 2.The second control mechanism is in response to
glucose, which is transported into the cell by thephosphoenolpyruvate (PEP)-dependent phosphotransferasesystem. The phosphate group of PEP is transferred viaa phosphorylation cascade, consisting of the generalphosphotransferase system (PTS) proteins HPr andEIA, and the glucose-specific PTS proteins, EIIAGlc and
EIIBGlc, which are the cytoplasmic domain of the EIIglucose transporter. Transport of glucose is accompaniedby its phosphorylation by EIIBGlc through removing thephosphate group from the other PTS proteins, includingEIIAGlc. The unphosphorylated form of EIIAGlc binds tothe lac permease and prevents it from bringing lactose intothe cell. Therefore, if both glucose and lactose are present,the transport of glucose blocks the transport of the inducerof the lac operon. This process is called inducer exclusion.[13]
In other way, 16 base pairs upstream site of thepromoter, which is used for a positive control of thegene expression is known as CAP site or cAMP proteinsite or catabolite gene activator (cga) site, because it isutilized for binding of CAP to encourage geneexpression. CAP can bind to this site only when it isbound with cAMP. Hence, a positive control is exertedover the transcription process by the CAP–cAMPcomplex with an effect exactly opposite to that ofrepressor binding to an operator. Moreover, the effectormolecule cAMP determines the effect of CAP on lacoperon transcription in the presence of glucose, itinhibits the formation of cAMP and prevents it to bindto CAP. The DNA-bound CAP is then able to interactphysically with RNA polymerase and essentiallyincrease the affinity of RNA polymerase for the lacpromoter. In this way, the catabolite repression systemcontributes to the selective activation of the lacoperon.[14] The schematic diagram of mechanism ofcarbon catabolic repression is described in Fig. 3.
Figure 2. Regulation of lac operon in (A) presence and (B) absence of lactose.[12]
Asia-Pacific Journal of Chemical Engineering β-GALACTOSIDASE: A REVIEW
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

A mechanism of carbon catabolite repression bywhich the activity of PTS-regulation domain-containing transcription factors, which is inhibited inthe presence of preferred carbon sources, is calledInduction prevention. It proceeds with phosphorylationof histidine protein (HPr) at Ser46 by HPr kinase/phosphorylase (HPrK). This phosphorylation occurswhen the intracellular concentrations of fructose-1,6-bisphosphate (FBP) and adenosine triphosphate arehigh, which reflects the presence of preferred carbonsources. HPr(Ser-P) binds to CcpA protein, and thisinteraction is enhanced by glycolytic intermediates, suchas FBP and glucose-6-phosphate. The complex of CcpAand HPr(Ser-P) binds to cre sites on the DNA andthereby represses the transcription of catabolic genes.HPrK is also responsible for dephosphorylation ofHPr(Ser-P) under conditions of high inorganicphosphate (Pi) and low adenosine triphosphate andwhen there is poor nutritional supply of FBP. Also,HPr(His-P) contributes to carbon catabolic repression:in the absence of glucose, HPr(His-P) phosphorylatesglycerol kinase (GlpK), and transcriptional regulatorsthat contain PEP–carbohydrate phosphotransferasesystem-regulatory domains, which acts as precursorsfor their activity. Thus, in the presence of glucose,activation of the phosphotransferase system-regulatorydomain regulators by their inducers is prevented.[13]
Research with this system is greatly added by theavailability of constitutive mutants. A constitutivemutant of any consortia continuously produces geneproducts where there is no control over its expression.In these mutants, the aforementioned proteins areproduced all the time in comparison to the wild typewhere the proteins only appear in the presence of lactose.So in these mutants, the mutation must be a gene otherthan those responsible for the structural gene.[15]
PRODUCTION OF β-GALACTOSIDASE
β-galactosidase belongs to the group of saccharidesconverting enzymes, i.e., in the family of hydrolases.They are widespread, distributed in numerousbiological systems, e.g., microorganisms (yeasts, fungi,bacteria, and actinomycetes), plants, and animaltissues. Compared with animal and plant sources,microbial-synthesized enzyme provides higher yields,which may decrease its production cost. Therefore,production of β-galactosidase through microbial routecreates a great attention. Although, the most studiedβ-galactosidase is produced by E. coli, possible toxicfactors associated with coliforms make it unlikely thatcrude isolates of this enzyme, which may be permittedin food processes.[16–21] In industrial scale, productionof β-galactosidases is carried out using generallyrecognized as safe microorganism, yeast (mainly fromKluyveromyces marxians, Kluyveromyces lactis, andKluyveromyces fragilis) and fungal (mainly fromAspergillus niger and Aspergillus oryzae) consortia.The detail works carried out in this direction have beenrepresented in Table 1.
PURIFICATION OF β-GALACTOSIDASE
Different separation techniques, such as membrane-based separation, ion exchange membranechromatography, gel permeation chromatography, zincchloride, protamine sulfate, and ammonium sulfateprecipitation; had already been attempted forpurification of β-galactosidase from crude extract. Thedetail works carried out in this direction have beenrepresented in Table 2.
Figure 3. Mechanism of carbon catabolic repression (A) inducer exclusion and (B) inductionprevention (Figure adapted from B. Görke and J. Stülke (2008)).[13]
A. NATH ET AL. Asia-Pacific Journal of Chemical Engineering
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

Table
1.Differenttypes
ofco
nso
rtium
usedin
β-galactosidaseproduction.
Typ
eof
consortiu
mNam
eof
consortiu
mSub
strate
Operatin
gmod
eof
bioreactor
Reference
Yeast
Kluyverom
yces
marxian
sCheesewhey
Fed
batch
[22]
Yeast
Kluyverom
yces
marxian
sCheesewhey
Batch
[23]
Yeast
Kluyverom
yces
marxian
sSup
plem
entedwheywith
cauliflow
erwaste
Batch
[24]
Yeast
Kluyverom
yces
marxian
sCBS78
94Lactose-based
chem
ically
definedmedium
Batch
[25]
Yeast
Kluyverom
yces
marxian
sNCIM
3551
Whey
Batch
[26]
Yeast
Kluyverom
yces
marxian
sCCT78
02Lactose-based
chem
ically
definedmedium
Batch
[27]
Yeast
Kluyverom
yces
marxian
sMTCC13
88Whey
Batch
[28]
Yeast
Kluyverom
yces
frag
ilisNRRL-Y-110
9Lactose-based
chem
ically
definedmedium
Batch
andfedbatch
[29]
Yeast
Recom
binant
Saccharomyces
cerevisiae
W303
Lactose-based
yeast-definedmineral
medium
Batch
[30]
Yeast
Saccha
romyces
cerevisiae
Lactose-based
chem
icallydefinedmedium,de-proteinatedwhey
Fed
batch
[31]
Yeast
Can
dida
pseudo
trop
icallis
Whey
Batch
[32]
Bacteria
ClonedEscherichia
coliXL-1
Blue,XL-1
BlueFF,T
B-1
andN48
30Basal
medium
with
IPTG
Batch
[33]
Bacteria
Lactoba
cillu
ssp.B
ulga
ricusCHRHan
sen
Lb-12
Com
binatio
nof
skim
milk
powder,glucose,andyeastextract
Batch
[34]
Bacteria
Bacillus
sp.M
PTK12
1Lactose-based
chem
ically
definedmedium
Batch
[35]
Bacteria
Bacillus
sp.
Luriamedium
whereindividu
ally
galactose,glucose,mannitol,
lactose,andxy
lose
wereadded
Batch
[36]
Bacteria
Bacillus
licheniform
isATCC12
759
Luria
medium
Batch
[37]
Bacteria
Lactoba
cillu
sacidop
hilusNRRL44
95Acidwhey
Batch
[16]
Bacteria
Bifido
bacterium
animalisBd1
2,Lactoba
cillu
sdelbrueckiissp.
bulgaricus
ATCC11
842
Chemically
definedmedium
where
individu
ally
glucose,
lactose,andgalactosewereused.Moreover,individu
ally
pepton
e,yeastextract,casein
hydrolysate,tryp
tone,or
ammon
ium
sulfatewereused
Batch
[17]
Bacteria
Bifido
bacterium
animalisBd1
2,Lactoba
cillu
sdelbrueckiissp.
bulgaricus
ATCC11
842
De-proteinatedwheywhere
proteinwas
supp
lementedby
individu
ally
pepton
e,yeastextract,casein
hydrolysate,
tryp
tone,andam
mon
ium
sulfate
Batch
[18]
Bacteria
Lactoba
cillu
splan
tarum
Pi06
Lactose-based
chem
ically
definedmedium
Batch
[19]
Bacteria
Lactoba
cillu
sacidop
hilus
de-M
annRog
osaandSharpemedium
Batch
[20]
Bacteria
Streptococcusthermop
hilus
Whey-basedmedium
Batch
[38]
Bacteria
Lactoba
cillu
sdelbrueckiissp.
bulgaricus
ATCC11
842
Skim
milk
,mod
ified
de-M
annRog
osaSharpemedium,w
hey,
andwheyperm
eate-based
brothwereused.A
ddition
ally,y
east
extractwas
added
Batch
[21]
Bacteria
Bacillus
coag
ulan
sRCS3
Lactose-based
chem
ically
definedmedium
Batch
[39]
Fun
gus
Aspergillu
sflavus
Lactose-based
chem
ically
definedmedium
Batch
[40]
Fun
gus
Penicillium
chrysogenu
mNCAIM
0023
7Lactose-based
chem
ically
definedmedium
Batch
[41]
Fun
gus
Aspergillu
soryzae
PTCC51
63Wheyandchem
ically
definedmedium
with
different
carboh
ydrate,i.e.,lactose,wheat
barn,and
soyb
eanmeal
Batch
[42]
Fun
gus
LacA
from
Aspergillu
soryzae,exp
ressingin
Saccha
romyces
cerevisiae
NCYC86
9-A3/
pVK1.1
Wheat
bran
andrice
husk
Batch
[43]
Fun
gus
Rhizomucor
sp.
Lactose-based
chem
ically
definedmedium
Batch
[44]
Fun
gus
Aspergillu
sniger
Lactose-based
yeastnitrog
enbase
medium
andsemi-synthetic
lactosemedium
Con
tinuo
us[45]
Asia-Pacific Journal of Chemical Engineering β-GALACTOSIDASE: A REVIEW
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

Table
2.Differentmethodologyforβ-galactosidasepurification.
Sou
rce
Purificatio
nprocedure
Characteristicsof
purified
enzyme
Reference
Lactoba
cillu
smurinus
Purified
292-fold
bychromatog
raph
yon
Ultrog
elACA
34,D
EAE–S
ephadexA-50
columns
andby
affinity
chromatog
raph
yin
agarose-p-am
inop
heny
l-β-
D-thiog
alactoside
Subunitsaresamemolecular
weight.M.W
.is170kg
mol
�1.
Maxim
umenzymaticactiv
ityat318KandpH
7inpresence
of50
mM
phosphatebuffer.T
heKmforONPG
andONPG
+20
×10
�3M
oflactoseare480×10
�6and870×10
�6M,respectively.
Inhibitors
are10
×10
�3M
CaC
l 2,g
lutathione,and
cysteine,and
stim
ulator
is10
×10
�3M
MgC
l 2.A
ctivatorsaremercaptoethanolanddithiotreitol.
Enzym
aticactiv
ityisnotshownin
presence
ofp-β-galactosidase.
[46]
Bacillus
sp.3
088
ZnC
l 2precipitatio
n,ionexchange
mem
brane
chromatography,
hydrophobicinteraction
chromatographyandgelfi
ltration
chromatography
M.W
.is48
4kg
mol
�1,associatewith
subu
nits11
5,86
.5,72.5,45
.7,
and41
.2kg
mol
�1.O
ptim
umpH
andtemperature
are8and
333.15
K,respectiv
ely.
Isoelectricpo
intis6.2.
Kmis6.34
×10
�3
and6.18
×10
�3M
forONPG
andlactose,respectiv
ely.
Inhibitors
aregalactosedivalent
Hg,
Cu,
andAg.
Metalactiv
ator
isdivalent
Mg.
EDTA
dono
taffect
theenzymeactiv
ity.
[47]
Lactoba
cillu
sreuteri
Ammon
ium
sulfateprecipitatio
n,hy
drop
hobic
interactionchromatog
raph
y,andaffinity
chromatog
raph
y
Heterod
imer
enzyme,M.W
.is10
5kg
mol
�1(m
onom
ersare72
and
35kg
mol
�1).Isoelectricpointsare4.6–4.8and3.8–4.0forL
.reuteri
L103andL.reuteriL461,respectiv
ely.Optim
umpH
is6–8.Inhibitor
isD-glucose
andactiv
atorsareNa(+),Mn(+2),and
K(+).
[48]
Bacillus
circulan
sGel
perm
eatio
nchromatog
raph
y,affinity
chromatog
raph
y,andam
mon
ium
sulfate
precipitatio
n
M.W
.is21
2kg
mol
�1,com
posedby
145and86
kgmol
�1.K
mfor
threesubunitsare3.6×10
�3,5.0×10
�3 ,and3.3×10
�3M
forO
NPG
,respectiv
elyandthoseare3.7×10
�3,2.94×10
�3 ,and2.71
×10
�3M,
respectiv
elyconsideringlactoseas
asubstrate.
[49]
(Exp
ressed
inEscherichia
coli)
Colum
nchromatog
raph
ieson
resource
Qand
seph
acrylS-200
HR
Potentialof
GOSsynthesis
[50]
Streptococcusthermop
hilus
Ammon
ium
sulfateprecipitatio
nandgel
perm
eatio
nchromatog
raph
yIntracellular,M.W
.is53
0kg
mol
�1.O
ptim
umpH
is7.1.
Galactose
iscompetitiveinhibitor(K
i60
×10
�3M),KmforO
NPGandlactose
are0.98
×10
�3and6.9×10
�3M,respectiv
ely.
[51]
Streptococcuslactis
Protaminesulfateprecipitatio
n,am
mon
ium
sulfateprecipitatio
n,andionexchange
chromatog
raph
y
Enzym
eismostlabile
whenitissuspendedat
cold
(278
.15K)
phosph
atebu
ffer.In
presence
ofhigh
concentrationam
mon
ium
sulfate(0.85M),enzymeishigh
lyactiv
eandstabilizes.
[52]
Bacillus
stearothermop
hilus
(exp
ressed
inBacillus
subtilis)
Heattreatment,am
monium
sulfateprecipitatio
n,ionexchange,and
gelfi
ltrationchromatography
Intracellular,M.W
.is70
kgmol�1 .Isoelectricpointis5.1.Optim
umtemperatureandpH
are343.15
Kand7,respectively.Kineticsof
thermalinactivationandhalflifeat338.15
and343.15
Kare18
×10
4
and324×10
2s.KmandVmaxare2.96
×10
�3M
and0.11
×10
�3
kmols�
1kg
ofprotein,respectively.InhibitorsareFe(2+),Z
n(2+
),Cu(2+
),Pb
(2+),S
n(2+),and
thiolagent.
[53]
Bacillus
megaterium
Son
ication,
ammon
ium
sulfateprecipitatio
n,desaltatio
n,columnchromatog
raph
yusing
DEAE–sepharose
fastflow
column,
andaffinity
chromatog
raph
y
M.W
.is118kg
mol
�1.O
ptim
umpH
7.5–8.0andtemperature328K.
Enzym
eisstableatpH
6.0–9.0andbelow313K.T
heKmandVmax
values
forONPG
andlactoseare9.5mM,16.6mM.m
in�1and
12.6mM,5
4.4mM.m
in�1 ,respectiv
ely.
[54]
Bacillus
licheniform
is(cloned
andexpressedin
Escherichia
coli)
Ionexchange
chromatog
raph
yon
NiS
epharose
6fastflow
columnandultrafiltration
Hom
odim
eric,op
timum
temperature,andpH
are32
3.15
Kand
6.5,
respectiv
ely.
KmforlactoseandONPG
are16
9×10
�3and
13.7×10
�3M,respectively.
Inhibitors
areglucoseandgalactose.
Metal
activ
atorsareNa+,K
+,M
g+2,M
n+2,and
Ca+
2.
[55]
A. NATH ET AL. Asia-Pacific Journal of Chemical Engineering
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

Table
2.(Continued
)
Sou
rce
Purificatio
nprocedure
Characteristicsof
purified
enzyme
Reference
Arthrob
actersp.
Ionexchange
chromatographyon
DEAE–
sepharose,
affinity
chromatographyon
PABTG-sepharose,
andgelfi
ltrationon
SephacrylS
-300
Intracellular,ho
mod
imeric,each
subu
nitM.W
.is11
6kg
mol
�1.
Optim
umpH
andtemperature
are6–8and29
8.15
K,respectiv
ely.
Activatorsarethiolcompo
unds,Na(+),andK
(+).Inactiv
ated
by4-chloromercuribenzoicacid,Pb(2+),Zn(2+),andCu(2+)
[56]
Bifido
bacterium
long
umCCRC15
708
Ultrason
icationandsubsequently
purified
byQ
Fast-Flow
chromatog
raph
yandgel
chromatog
raph
yon
aSup
erose6HRcolumn
Finally
15.7-fold,
ayieldof
29.3%,andaspecificactiv
ityof
168.6*
106Ukg
-1protein(β-galactosidase)hasbeen
achh
ived.T
heMW
was
357kg
.mol-1
asdeterm
ined
from
Native-PAGE.The
pHandtemperature
optim
aof
thepu
rified
β-galactosidasewere7.0and
323K,respectivelyconsideringONPGas
asubstrate,.T
heenzyme
was
stableatatemperature
upto
313KandatpH
values
of6.5-7.0.
Valuesof
Km
andVmax
forthispu
rified
enzymewerefoun
dto
be0.85
mM
and70
.67*1
06U
kg-1,respectiv
ely.
Itwas
foun
dthat
Na+
andK+stim
ulated
theenzymeup
to10
-fold,
while
Fe3
+,F
e2+,
Co2
+,C
u2+,C
a2+,Z
n2+,M
n2+andMg2
+inhibitedtheactiv
ityof
β-galactosidase.
Furthermoreitwas
foun
dthat
glucose,galactose,
maltose,orraffino
seexertedlittle
orno
effecton
theβ-galactosidase
activ
ity,lactoseandfructose
inhibitedtheenzymeactiv
ity.The
effectof
lactoseon
theenzymeactiv
ityforONPGisprob
ably
acase
ofcompetitiveinhibitio
n.
[57]
Bifido
bacterium
bifidu
mUltrason
ictreatm
entin
thepresence
ofTriton
X-100
Enzym
ehasdifferentsubu
nits,such
as16
3,17
0,17
8,and
190kg
mol
�1.T
heM.W
.is36
2kg
mol
�1andisoelectricpo
intis
5.25
.The
purified
enzymeisstableattemperaturesbelow31
8Kand
over
thepH
rang
esfrom
6.5–
8.Lactose
hydrolysisby
thepu
rified
enzymetake
places
atpH
6.5andtemperature
310K.
[58]
Bifido
bacterium
infantis
(exp
ressed
inEscherichia
coli)
Ammon
ium
sulfateprecipitatio
n,dialysis,
anionexchange
column(M
onoQ
HR5/5,
Pharm
acia)usingaFPLCsystem
(Pharm
acia).
Activeβ-galactosidasefractio
nshasbeen
collected
from
severalchromatog
raph
icruns
andconcentrated
usingtheCentriplussystem
(Millipore).The
activ
efractio
nof
β-galactosidasehasbeen
obtained
from
ion
exchange
chromatog
raph
y.
The
enzymeistetram
er,w
ithan
M.W
.ofabou
t47
0kg
mol
�1,and
itssubu
nitis11
5kg
mol
�1.T
heop
timum
temperature
andpH
for
ONPG
andlactoseare333K,pH7.5and323K,pH7.5,respectiv
ely.
The
enzymeisstableover
apH
rangeof
5.0–8.5andremains
activ
eformorethan
4800
satpH
7.0,323K.T
heenzymeactiv
ityhasbeen
significantly
increasedby
reducing
agents.M
axim
umactiv
ityhas
been
show
nin
presence
ofboth
Na+
andK+ataconcentrationof
10×10
�3M.T
heenzymeisstrongly
inhibitedby
p-chloromercuribenzoicacid,d
ivalentm
etalcatio
nsandCr3+,and
toalesser
extent
byEDTAandurea.T
hehydrolyticactiv
ityusing
lactoseas
asubstratehasbeen
significantly
inhibitedby
galactose.
The
KmandVmaxvalues
forONPG
andlactoseare2.6×10
�3M,
262×10
6Ukg
�1and73.8×10
�3M,1.28×10
6Ukg
�1 ,respectiv
ely.
The
enzymepossessesstrong
transgalactosylatio
nactiv
ity.T
heproductio
nrateof
GOSfrom
20%
lactoseat303and333Kis
120×10
�3kg
L�1,and
thisrateincreasesto190×10
�3kg
L�1when
30%
lactosehasbeen
used
asasubstrate.
[59]
Cryptococcuslaurentii
OKN-4
Colum
nchromatog
raph
iesby
DEAE–S
ephadex
A-50,TSK
gelToy
oPearlHW-55S
,andTSK
gelDEAE-5PW
M.W
.is200kg
mol
�1.hom
odim
eric,optim
umpH
is4.3,andenzyme
isstableatpH
between2.8and9.3.
The
optim
umtemperature
is333Kandenzymeisstableattemperature
below330.5Kfor600s
[60]
(Continues)
Asia-Pacific Journal of Chemical Engineering β-GALACTOSIDASE: A REVIEW
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

Table
2.(Continued
)
Sou
rce
Purificatio
nprocedure
Characteristicsof
purified
enzyme
Reference
incubatio
n.The
Kmvalues
oftheenzymeare18.2×10
�3M
and
11.4×10
�3M,and
thevalues
ofVmaxare1.28
×10
�3km
ols�
1kg
ofprotein,
and0.09
×10
�3km
ols�
1kg
ofproteinforONPG
and
lactose,respectiv
ely.The
enzymeisstrongly
inhibitedby
Hg2
+,A
g+,
2-mercaptoethanol,g
lucose,m
altose,and
maltotriose.E
nzym
eis
potentialo
fGOSsynthesis.
Aspergillu
soryzae
2-prop
anol
fractio
natio
n,column
chromatog
raph
yon
DEAE–S
ephadex
A-50,
andSephadexG-200
Extracellu
lar,op
timum
pHforONPG
andlactoseare4.5and4.8,
respectiv
ely.
Optim
umtemperature
is31
9.15
K.K
mare7.2×10
�4
and1.8×10
�2M
forONPG
andlactoserespectiv
ely.
Inhibitors
are
divalent
Hg,
Cu,
N-bromosuccinimide,
andsodium
laurylsulfate.
App
arentM.W
.is10
5kg
mol
�1.
[61]
Metal-ion
affinity
chromatog
raph
y(IMAC)
follo
wed
bysize-exclusion
separatio
nExtracellu
lar,M.W
.is11
3kg
mol
�1.M
utantenzym
ehasfive
times
high
ercatalytic
activ
ityon
thesynthetic
substrateONPG
compared
with
thewild
-typ
eenzyme.Moreover,themutantenzymeismore
thermoresistantcomparedwith
thewild
type.
[62]
Aspergillu
sniger
Gel
filtration,
anionexchange
chromatog
raph
yon
DEAE–S
epharose
CL-6B,hy
drop
hobic
chromatog
raph
yon
octyl-SepharoseCL-4Band
catio
nexchange
chromatog
raph
yon
CMSepharose
CL-6B
Glycoproteinin
nature.A
ssociateswith
threesubu
nits,and
each
M.
W.are12
4,15
0,and17
3kg
mol
�1.Isoelectric
pointis4.6.
Optim
umpH
liesbetweenin
2.5to
4.0.
Heatstableup
to33
3.15
K.
Kmvaluevaries
from
85×10
�3to
125×10
�3M
forlactose.Km
valuevaries
2.4×10
�3M
forONPG.V
maxvalues
are10
4×10
3un
itenzymekg
ofproteinand12
1×10
3un
itenzymekg
ofproteinat
303.15
KforONPG
andlactose,respectiv
ely.
[63]
Aspergillu
saculeatus
Proteinprecipitationby
Pectinex
UltraSP
followed
bydesalting
andcolumnchromatographyusing
DEAE–Sepharose
fastflow
.
The
M.W
.is12
0kg
mol
�1(app
roximately),isoelectricpo
intlies
between5.3and5.7andisop
timally
activ
eat
pH5.4and
temperature
328–
333K.Based
ontheN-terminal
aminoacid
sequ
ence,the
enzymeprob
ably
belong
sto
family
35of
theglycosyl
hydrolases.Enzym
eispo
tentialof
synthesizing
GOS.
Enzym
ehasactiv
itytowards
twoexo-po
ly-saccharide(EPSs),
having
lactosyl
side
chains
attached
todifferentbackbo
nestructures.The
enzymedegraded
O-deacetylatedEPSB89
1faster
than
EPSB39
.Moreover,thepresence
ofacetyl
grou
psin
EPSB89
1slow
sdo
wnthehy
drolyzingrate
buttheenzymeis
still
able
toreleaseallterm
inally
linkedgalactose.
[64]
Penicillium
chrysogenu
mPrecipitatio
nwith
ammonium
sulfate,ion
exchange
chromatographyon
DEAE–S
ephadex,
affinitychromatography,andchromatefocusing
Intracellular,Sp
ecificactiv
ityis5.84
×10
3Ukg
ofprotein.Optim
umtemperature
andpH
are303.15
Kand4,
respectiv
ely.Kmand
isoelectricpointare
1.81
×10
�3M
and4.6forONPG
.Multim
eric
enzyme,M.W
.is270kg
mol�1,and
M.W
.ofsinglemonom
eris
66kg
mol
�1
[41]
Kluyverom
yces
lactis
Gel
filtrationchromatog
raph
y,ionexchange
chromatog
raph
y,andaffinity
chromatog
raph
yAffinity
chromatog
raph
yisshow
nbetterresults
than
gelp
ermeatio
nchromatog
raph
y,andionexchange
chromatog
raph
y.Sim
ilar
molecular
weigh
tsub
unit.
Enzym
eisconfi
rmed
byspecificantig
en-
antib
odyreactio
n(ELISA
test).
[65]
Kluyverom
yces
lactis
Gelfiltrationon
Superose
12PC
3.2/30
column
andionexchange
Intracellular,M.W
.of
sing
lemon
omer
is12
4kg
mol
�1
[66]
A. NATH ET AL. Asia-Pacific Journal of Chemical Engineering
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

Table
2.(Continued
)
Sou
rce
Purificatio
nprocedure
Characteristicsof
purified
enzyme
Reference
chromatographyon
MonoQPC
1.6/5column
usingaFP
LCsystem
Celld
isruption,
DEAE–S
ephadexionexchange
chromatography,
andchromatographyon
hydroxylapatite
M.W
.is13
5kg
mol
�1.O
ptim
umpH
is7.25
.Kmis12
×10
�3to
17×10
�3M
forlactose.Kmis1.6×10
�3M
forONPG.Metal
activ
atorsareNa(+)andMn(+2).
[67]
Kluyverom
yces
marxian
usExtractionwith
2%chloroform
,aceton
e,and
ammon
ium
sulfateprecipitatio
nM.W
.is28
0kg
mol
�1.O
ptim
umtemperature
andpH
are31
8.15
to32
5.15
Kand6.2,
respectiv
ely.
Inactiv
ates
in24
0sat32
9.15
K.K
mare3.1×10
�3and25
×10
�3M
forONPG.Inhibitio
nconstantsare
58×10
�3,1
10×10
�3,1
11×10
�3,and
52×10
�3M
forONPG,
galactose,ribo
se,andlactose.Inhibitors
arealso
P-chlorom
ercuribenzoate
anddithiothreito
l.
[68]
Saccha
romyces
lactis
Ammonium
sulfatefractio
natio
n,column
chromatographyon
Sephadex
G-100,and
DEAE–S
ephadexA-50,
78-foldpurificatio
n
Optim
umpH
is7.2.
Metal
activ
ator
divalent
magnesium
,Kmis
1.18
×10
�3M
forONPG.
[69]
Saccha
ropo
lysporarectivirgu
laFPDA
13columnchromatog
raph
y,am
mon
ium
sulfatefractio
natio
n,Ultrog
elAcA
34column
chromatog
raph
y,DEAE–S
epharose
CL-6B
columnchromatog
raph
y,TSK
gelDEAE-
Toy
opearlpak65
0SHigh-pressure
liquid
chromatog
raph
y(H
PLC)
Extracellu
larthermostable,relativ
eM.W
.of
14500
0ands°
20,w
of7.1s.Michaelisconstant
Kmis0.75
×10
�6kM
andmolecular
activ
ity(k
cat)is63
.1s�
1atpH
7.2and32
8KforO
NPGwhereas
Km
is0.04
×10
�6kM
andk c
atis55
.8s�
1forp
-NPG.E
nzym
ehasahigh
transgalactosylatio
nactiv
ity.The
enzymereacts
with
1.75
Mlactoseat333KandpH
7.0for7.92
×10
4sto
obtain
maxim
umyieldoligosaccharides
(41%
(w/w)).T
hegeneralstructure
forthemajor
transgalactosylic
productscanbe
expressedas
(Gal) c-
Glc,w
here
nis1,
2,3,
and4with
aglucoseatareducing
term
inal.
[70]
Sterigm
atom
yces
elviae
Centrifug
ation,
celldisrup
tion,
heat
treatm
ent,
DEAE–T
oyop
earlchromatog
raph
y,salting
out,Butyl-Toy
opearlchromatog
raph
y,Chrom
atofocusingchromatog
raph
y,andPATG
chromatog
raph
y
M.W
.is17
0and86
kgmol
�1fordenaturedenzyme,isoelectric
point4.1.
The
optim
altemperature
is35
8K,anditisstable
attemperaturesup
to35
3K
for360
0s.The
optim
alpH
rang
eforthe
enzymeis4.5to
5.0andtheenzymeisstable
atpH
2.5to
7.0.
Ezymeisinhibitedby
Hg2
+.T
heKmvalues
forONPG
andl
actose
are9.5×10
�3and2.4×10
�3M,respectively.
The
Vmax
for
thesesubstrates
are96
mols�
1kg
ofproteinand4mols�
1kg
ofprotein,
respectiv
ely.
The
enzymepo
ssessesahigh
levelof
transgalactosylatio
nactiv
ity(higheryield39
%from
200gL�1
lactose),which
prod
uces
GOSinclud
ingtri-andtetrasaccharides.
[71]
Therm
usaquaticus
Gel
filtrationchromatog
raph
y(U
ltrog
elAcA
34),anionexchange
(Mon
oQ),andgel
filtration(Sup
erose-12
)
M.W
.is70
0kg
mol
�1andsubu
nitis59
±1kg
mol
�1.Isoelectric
pointis4.9.
The
optim
umtemperature
andpH
forenzymeactiv
ityare35
3K
and5.5,
respectiv
ely.
The
enzymeisstable
over
awide
pHrang
e(pH3–12
),andthethermostabilityof
theenzymeis
enhanced
byCaC
l 2.T
heenzymeissign
ificantly
activ
ated
byalkali
andalkalin
e-earth-metal
salts.Inhibitors
arethiol-bind
ingagents,
glucose,andgalactose.The
enzymespecificforb-D
anom
eric
linkagesandtheidentityof
theaglycone
moietyalso
influenced
enzymeactiv
itydram
atically.Enzym
eispo
tentialof
GOS
synthesis.
[72]
Asia-Pacific Journal of Chemical Engineering β-GALACTOSIDASE: A REVIEW
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

CHARACTERIZATION OF β-GALACTOSIDASE
Apart from catalyzing the hydrolysis of β-galactosidesinto monosaccharides by lactase, the enzyme may alsocleave fucosides and arabinosides with much lowerefficiency. Lactase is often confused as an alternativename for β-galactosidase, but it is merely a sub-classof β-galactosidase. In other words, β-galactosidase isan exoglycosidase, which hydrolyzes the β-glycosidicbond formed between galactose and its organicmoiety.[73] Molecular weight, amino acids chain length,position of the active site, pH, and optimum thermalstability are significantly differed by the microbialsources.[74] The choice of suitable β-galactosidase sourcedepends on the condition of reaction. For example, dairyyeasts with optimum pH (6.5–7.0) are habitually used forthe hydrolysis of lactose in milk or sweet whey.[75] On theother hand, the fungal β-galactosidases with optimum pH(3.0–5.0) are more suitable for acidic whey hydrolysis.[76]
The activity of different β-galactosidases also dependson presence of ions. The fungal β-galactosidasesare active without presence of ions as cofactors; whereβ-galactosidase isolated from K. lactis shows its higheractivity in presence of Mn2+ and Na+. β-galactosidasesynthesized from K. fragilis are mostly active in presenceof Mn2+, Mg2+, and K+.[77] On the contrary, Ca2+
and heavy metals inhibit the enzyme activity of allβ-galactosidases.[78] Properties of β-galactosidasesynthesized by different microorganisms are describedin Table 3.In 1970, 1024 amino acids of β-galactosidase of E.
coli were first sequenced.[93] After 24 years, four chainscomprising the protein were discovered to be 464 kgmol�1 tetramer with 222-point symmetry. Every unitof β-galactosidase contains five domains; whereas theactive site persists in the third domain. This enzymecan be split into two peptides, LacZα and LacZΩ, noneof them is active but both spontaneously reassemble afunctional enzyme. This characteristic is used for manycloning vectors to achieve α-complementation inspecific laboratory strains of E. coli, where the plasmidencodes the small LacZα while the large LacZΩ isencoded by the bacterial chromosome. Aftermath,when DNA fragments inserted in the vector,production of LacZα disrupted, the cells revealed noβ-galactosidase activity, were subjected to the blue/whitescreening of recombinant clones further.[94] The activesite of β-galactosidase catalyzes the hydrolysis ofdisaccharide substrate via ‘shallow’ and ‘deep’ binding.The beta-linkage of the substrate was cleaved by aterminal carboxyl group on the side chain of glutamicacid.[95] It has been determined that the DNA fragmentof thermophilic bacterium Thermoanaerobacterethanolicus contains three open reading frames. One ofthe open reading frames corresponded to the LacA genefor a thermostable s-galactosidase and the nativerecombinant LacA showed the highest activity at
348K–353K. Immobilized on aldehyde silochrome,LacA was even more thermo stable and retained highactivity.[96] It has been found that individual moleculesof β-galactosidase from the crystallize enzyme as wellas the original enzyme displayed a range of activity of20-fold or greater. Molecules obtained from two diversecrystals have identical activity distributions, i.e.,31 600 ± 1100 and 31 800 ± 1100 reactions per minuteper enzyme molecule. This activity of the enzyme wasfound to be drastically different from that of the enzyme,which was used to grow the crystals (showed an activitydistribution of 38 500±900 reactions per minute perenzyme molecule).[97] Induced β-galactosidase in E. coliwild-type strains ATCC 8677 and 35321 in the presenceof various protease inhibitors have been studied. Thepresence of the protease inhibitors had least effect onthe average distribution of single molecule activities,and the relative activities of the enzyme for the diversesubstrates differed between the strains.[98] Average-combined turnover numbers of the enzyme from wild-type E. coli strains ATCC 35321 and 8677 in vivo andin vitro conditions in the presence and absence of His6tag differed considerably. This indicated that synthesizedenzymes in both conditions (vivo and vitro) were notalike and presence of a C-terminal His6 tag altered theactivity of s-galactosidase.[99] Moreover, it was foundthat electrophoretic mobility and catalytic activity ofindividual molecules of β-galactosidase synthesized byE. coli were different, although they had potentiality toact on the same substrate molecule.[100]
IMMOBILIZATION OF β-GALACTOSIDASE
Immobilization has shown to improve the stability ofβ-galactosidase, reusages, and reduces the processingtime in food and other industries. For example, theimmobilization of β-galactosidase of Thermus sp. T2was performed using ionic adsorption by a new anionicexchanger resins (based on coating of Sepabeadsinternal surfaces with polyethylenimine) andconventional DEAE–agarose. Immobilization wascarried out in both cases, but the adsorption strengthshowed much greater in the case of PEI–Sepabeads thanin DEAE supported at both pH5.0 and 7.0. Also, thePEI–Sepabeads remained wholly active, and after severalweeks of incubation at 323K, it showed the lactosehydrolysis in milk.[101] Also a new hetero functionalepoxy supports were used for immobilization ofβ-galactosidase. The capability of a standardSepabeads-epoxy supports to immobilize β-galactosidasefrom Thermus sp. strain T2 that equalized with otherSepabeads-epoxy, which supports partially modificationusing boronate, iminodiacetic, metal chelates andethylenediamine. Here, immobilization was dependedon the support, ranging from 95% to 5% using
A. NATH ET AL. Asia-Pacific Journal of Chemical Engineering
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

Table
3.Properties
ofβ-galactosidasesynthesized
bydifferentmicroorgan
isms.
Typ
eof
consortiu
mNam
eof
consortiu
mTem
perature
(K)
pHHalflife(s)
Specific
activ
ity(U
mg�
1)
Reference
Bacteria
Arthrob
acterpsychrolactoph
ilus
333
836
00(303
K)
20[79]
Bacillus
circulan
s32
35–
6—
5.1
[50]
Bacillus
megaterium
313
6–9
—60
[54]
Bacillus
stearothermop
hilus
343
73.24
×10
4(343
K)
125
[53]
Bifido
bacterium
adolescentis
323
660
0(323
K)
526
[80]
Bifido
bacterium
bifidu
m31
06.5
1.44
×10
4(318
K)
—[58]
Bifido
bacterium
infantis
323–
333
7.5
7200
(333
K)
569
[59]
Cryptococcuslaurentii
331
4.3
600(331
K)
12[60]
Enterob
acterag
glom
eran
s31
0–31
37.5–
8—
—[81]
Lactoba
cillu
sacidop
hilus
328
6.5–
81.72
8×10
5(310
K)
230
[82]
Lactoba
cillu
sreuteri
323
6–8
—18
0[48]
Streptococcuspn
eumon
ia30
35.5–
7.5
——
[83]
Alicycloba
cillu
sacidocalda
rius
338
5.5
360(353
K)
592
[84]
Caldicellu
losiruptor
saccha
rolyticus
353
61.72
8×10
5(343
K)
211
[85]
Geoba
cillu
sstearothermop
hilus
338
6.5
—0.5
[86]
Saccha
ropo
lysporarectivirgu
la33
36.5–
7.2
1.44
×10
4(333
K)
—[70]
Therm
otog
amaritima
353–
358
6.5
960(363
K)
70[87]
Thu
rmus
sp.
343
6.5
7.2×10
4(343
K)
—[88]
Thu
rmus
aqua
ticus
353
5.5
—5.7
[72]
Yeast
Kluyverom
yces
frag
ilis
303
6.5
1.8×10
4(313
K)
—[88]
Kluyverom
yces
lactis
310–
313
6.6–
7—
—[89]
Sporob
olom
yces
sing
ularis
303
4—
8.7
[90]
Sterigmatom
yces
elviae
358
4.5–
536
00(353
K)
20[71]
Bullera
sing
ularis
323
572
00(318
K)
56[91]
Fun
gus
Aspergillu
soryzae
303
4.8
600(343
K)
40[62]
Aspergillu
saculeatus
328–
333
5.4
—24
[64]
Archae
Sulfo
lobu
ssolfa
taricus
348
6.5
1.08
×10
4(358
K)
116
[92]
Asia-Pacific Journal of Chemical Engineering β-GALACTOSIDASE: A REVIEW
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

Sepabeads-epoxy-chelate, Sepabeads-epoxy-amino, orSepabeads-epoxy-boronic using Sepabeads-epoxy-IDA.Amazingly, the immobilized β-galactosidase derivativesshowed outstandingly good result but different stabilitieshad been notified after favoring multipoint covalentattachment by long-term alkaline incubation. Theenzyme immobilized on Sepabeads-epoxy-boronicwas found to be the steadiest. The crosslinking withaldehyde-dextran allowed the stabilization of thequaternary structure of the enzyme. The optimalderivative was extremely active in lactose hydrolysiseven at 343 K (over 1000 IU g�1), maintaining itsactivity after extended incubation times with no riskof product contamination with enzyme subunits.[102]
Protocol for immobilization of β-galactosidasesynthesized by E. coli using diverse supports (glyoxyl,epoxy, BrCN groups, or by glutaraldehyde crosslinkingon matrix, containing primary amino groups), andstrategies have been studied. In each case, theimmobilization yield showed 100%with active recoveriesbetween 50% and 100% (using ortho-nitrophenyl-β-galactoside as substrate). Ratio of synthetic activityto hydrolytic activity (Vs/Vh) was lower than 0.1 whensoluble enzyme and the Eupergit 250 L enzymewere immobilized on BrCN at 277 K and pH 7.0,resulting 0.46 and 0.8, respectively.[103] Immobilizationof β-galactosidase producing permeabilized dead cellsof K. lactis ATCC 8583 into gelatin usingglutaraldehyde as crosslinker was performed, where30% activity obtained by immobilized cells relative tofree disrupted cells.[104] The usage of calcium alginate,Κ-carrageenan, and gellan-xanthan gel beads forentrapment of β-galactosidase synthesized byStreptococcus thermophilus, enhanced the stability ofenzyme at higher temperatures (>298K).[105] Solid statefermentation with co-immobilized β-galactosidasessynthesized by K. lactis, Aspergillus oryzae, and yeastsin polyvinyl alcohol hydrogel lens-shaped capsules havebeen performed. In the process the enzyme, synthesizedfrom Kluyveromyces lactis and Saccharomycescerevisiae showed the highest activity (galactose outputincreased from 3 to 4.1 g l�1 h�1), which reduced thereduction of processing time.[106] Differentmethodologies for immobilization of β-galactosidaseshave been described in Table 4.
INDUSTRIALIZED APPLICATIONS OFβ-GALACTOSIDASE
Microbial β-galactosidase plays a tremendouslyessential role in the production of various industrialrelevant products such as biosensor, lactose-hydrolyzed milk, ethanol, and GOS, also it has beenused in the field of bioremediation, diagnosis, and intreatment of lactose digestion disorder etc.
Use of β-galactosidase in biosensor
A biosensor associated with two distinct enzymaticactivities (β-galactosidase and glucose peroxidase) hasbeen developed for quantitative detection of lactose incommercial samples of milk. To avoid interferenceswith glucose, a degree of different mode ofmeasurement was done using biosensor.[146]
Foresighting this technique, presumed β-galactosidasefrom Streptococcus mitis with a choline-bindingdomain was identified recently. This remarkableproperty makes it differentially functional property forbiotechnological applications.[147]
Intended as lactose-hydrolyzed milk production
The cold–stable properties of Arthrobacter sp. 32csynthesized β-galactosidase could be useful forcommercial, industrial conversion of lactose intogalactose and glucose in milk products.[56] It has beenreported that rudimentary β-galactosidase extractproduced by Lactobacillus ssp. bulgaricus CHRHansen Lb-12 was applied in sterile milk, which hasbeen processed through ultra-high temperature method(UHT milk) for hydrolyzing lactose. Optimum amountof crude β-galactosidase extract and Maxilact enzymefor producing lactose-hydrolyzed milk was observedto be 0.418 and 0.512UmL�1 respectively during 6 hof processing. Using more than 418 U L�1 of crudeβ-galactosidase extract showed undesirable acidity oflactose-hydrolyzed milk that significantly increased attemperature of between 288 and 290K, whileenrichment of acidity in lactose-hydrolyzed milkproduced through Maxilact enzyme was not significant.Total count of lactose-hydrolyzed milk by 418UL�1
of crude β-galactosidase extract, after 2.16 × 104 s ofprocessing was significant high (5 to 30 Colony FormingUnit). Sensory estimation of lactose-hydrolyzed milkand ordinary ultra-high temperature milk (controlled)did not show any major differences with respect toacceptability of sweetness, taste, and color.[33] Thedifficulty in enzyme extraction and poor permeabilityof cell membrane to lactose was solved whenpermeabilized Kluyveromyces marxianus NCIM 3465cells were used for the production of lactose-hydrolyzedmilk. The ethanol-permeabilized yeast cells showed 89%of hydrolysed lactose under optimized conditions.[148]
Role of β-galactosidase in whey utilization
The execution of profound environmental legislationfor recycling and reuse of waste materials is grabbinglots of headlines at present century. Since many years,dairy products have been used as valuable ingredientsby the confectionery industry, being a huge economicalsource of capital, especially in the tropical andsubtropical countries. The large volume of byproduct
A. NATH ET AL. Asia-Pacific Journal of Chemical Engineering
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

of the dairy industry, namely, whey, creates severedisposal problem. Concerning about environmentalaspects, research has been emphasized on membraneseparation technology, providing remarkable newopportunities for large-scale protein and lactosefractionation. They intrude proteins that are multi-functional food ingredients of high nutritional valueand offer a wide range of functional properties allowingdevelopment of new products and optimization ofexisting products within considerable low cost, butpresence of high concentration of lactose in aqueousmedium after separation of proteins from whey createssevere disposal problem.[149–153] Different strategies
have been attempted for whey lactose conversion, thoseare related to the synthesis of nutraceuticals GOS, as wellas alcohol from whey permeates would ensure ultimateconsumption and utilization of casein whey.[154,155]
Function of β-galactosidase in ethanolfermentationThe idea of kinetic analysis of alcoholic fermentationof lactose using a recombinant flocculent strain ofSaccharomyces cerevisiae NCYC869-A3/T1,expressing both the LAC4 (coding for β-galactosidase)and LAC12 (lactose permease) genes ofKluyveromyces Lactis conceded. The lactose was
Table 4. Different techniques for immobilization of β-galactosidases.
Immobilization method Source of β-galactosidase Immobilization matrix References
Covalent binding Aspergillus oryzae Amino-epoxy Sepabead [107]
Covalent binding Aspergillus oryzae Chitosan bead and nylon membrane [108]
Entrapment Aspergillus oryzae Nylon-6 and zeolite [109]
Physical adsorption Aspergillus oryzae Phenol-formaldehyde resin [110]
Physical adsorption Aspergillus oryzae Polyvinyl chloride and silica gel membrane [111]
Covalent binding Aspergillus oryzae Polyvinyl alcohol hydrogel and magneticFe3O4-chitosan as supporting agent
[112]
Covalent binding Aspergillus oryzae Silica [113]
Covalent binding Aspergillus oryzae Silica gel activated with TiCl3 and FeCl3[114]
Entrapment Aspergillus oryzae Spongy polyvinyl alcohol Cryogel [115]
Physical adsorption Aspergillus oryzae Celite and chitosan [116]
Covalent binding Aspergillus oryzae Cotton cloth and activated with tosyl chloride [117]
Physical adsorption Aspergillus niger Porous ceramic monolith [118]
Covalent binding Kluyveromyces lactis Magnetic polysiloxane-polyvinyl alcohol [119]
Entrapment Kluyveromyces bulgaricus Alginate using BaCl3[120]
Physical adsorption Kluyveromyces fragilis Cellulose beads [121]
Physical adsorption Kluyveromyces fragilis andKluyveromyces lactis
Chitosan [3]
Physical adsorption Kluyveromyces fragilis Chitosan [122]
Physical adsorption Kluyveromyces fragilis Chitosan bead [123]
Covalent binding Kluyveromyces lactis Cotton fabric [124]
Physical adsorption Kluyveromyces lactis CPC-silica and agarose [125]
Entrapment Kluyveromyces lactis, Aspergillusoryzae and Saccharomycescerevisiae
Poly(vinylalcohol) hydrogel [126]
Covalent binding Kluyveromyces lactis Corn grits [127]
Covalent binding Kluyveromyces lactis Thiosulfinate/thiosulfonate [128]
Covalent binding Kluyveromyces lactis Graphite surface [129]
Covalent binding Kluyveromyces fragilis Silica-alumina [130]
Entrapment Thermus aquaticus YT-1 Agarose bead [131]
Physical adsorption Thermus sp. T2 PEI–Sepabeads, DEAE–agarose [132]
Entrapment Penicillium expansum F3 Calcium alginate [133]
Covalent binding Saccharomyces anamensis Calcium alginate [134]
Physical adsorption Pisum sativum Sephadex G-75 and chitosan beads [135]
Covalent binding Lactobacillus bulgaricus Egg shells [136]
Physical adsorption Bacillus stearothermophilus Chitosan [137]
Physical adsorption Bacillus circulans Polyvinyl chloride and silica [138]
Covalent binding Bacillus circulans Eupergit C (Spherical acrylic polymer) [139]
Entrapment Escherichia coli Polyacrylamide gel [140]
Covalent binding Escherichia coli Polyvinyl alcohol [141]
Physical adsorption Escherichia coli Chromosorb-W [142]
Covalent binding Escherichia coli (Recombinantβ-galactosidase)
Cyanuric chloride-activated cellulose [143]
Covalent binding Escherichia coli Hen egg white [144]
Covalent binding Escherichia coli Gelatine [145]
Asia-Pacific Journal of Chemical Engineering β-GALACTOSIDASE: A REVIEW
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

completely utilized in all fermentative processes, and ithas been observed that with increase of initial lactoseconcentration (5 to 200 gL�1), the level of ethanolproduction increased linearly.[156] Meticulously, kineticmodel with respect to biomass growth, lactosehydrolysis, and ethanol production using β-galactosidasesynthesized by Kluyveromyces sp. was performedconsidering whey permeate and cheese whey powderas growth medium.[157–159] Moreover, Saccharomycescerevisiae has also been reported for production ofethanol considering concentrated deproteinized whey,cheese whey powder, and salted cheese whey as feedstock.[160–162] Ethanol fermentation of cheese wheypowder solution using the pure culture ofKluyveromycesmarxianus (DSMZ 7239) was studied in packed columnbioreactor using olive pits as sustaining particles forcell attachment.[163] Researches have also beenconducted for ethanol production by membranerecycle bioreactor and using co-immobilized S.cerevisiae strain and β-galactosidase in semi-continuous fermentation process considering wheypermeate and whey medium, respectively.[164,165]
Thus, whey utilization by β-galactosidase reduces theburden of water pollution and establishes the conceptof recycling and reuse of waste materials.
Galato-oligosaccharide production byβ-galactosidaseIn recent years, many investigations have been carriedout in the field of prebiotics, considered as functionalfood. Among them, one of the recognized functionalfood ingredients is oligosaccharides. They are
carbohydrates containing three to ten sugar units boundwith glycosidic bonds. It has been seen that there are anumber of classes of oligosaccharides, but amongthem, GOS has attracted particular attention becauseof their presence in human breast milk. GOS are non-digestible, carbohydrate-based food ingredient,responsible for human and animal nutrition. Productionof GOS by transgalactosylation activity results theformation of 4′- or 6′-galactosylactose, longeroligosaccharides, transgalactosylated disaccharides,and nonreducing oligosaccharides in presence ofβ-galactosidases. Depending on the source of enzymeand conditions of reaction, various glycosidic linkages,such as β(1,2), β(1,3), β (1,4), and β (1,6), are formedin the end product (GOS).[166] It has been found thatthe amount of GOS synthesis from lactose dependson the initial concentrations of lactose present inthe reaction mixture instead of the concentration ofβ-galactosidase. There are different phenomena thathave been reported to elucidate the synthesismechanism of GOS, which are depicted in Figs 4–6.It is believed that the presence of GOS in human
milk influences the growth of bifidobacteria in thegastrointestinal tract of newly born and breast-fedinfants. GOS fraction (referred to as bifidus factor) incow’s milk also provides several health benefits.[170]
GOS is stable under acidic conditions during foodprocessing as well as maintain excellent taste quality,which makes them popular as an active ingredient ina wide variety of food products. They pass throughthe small intestine without being digested for lowcaloric value. In addition, GOS is not metabolized by
Figure 4. Schematic diagram of galato-oligosaccharide synthesis from lactose,(A) engineering approach and (B) biochemical approach (Figure adapted fromBoon et al. (1999)).[167]
A. NATH ET AL. Asia-Pacific Journal of Chemical Engineering
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

microorganisms in the oral cavity.[171] The detail workscarried out in this direction have been represented inTable 5.
Applications β-galactosidase in medical andimmunology research
Clostridium perfringens ATCC 10543 synthesizedendo β-galactosidase, which is capable of liberatingbeneficial A trisaccharide and B trisaccharide fromglycoconjugates containing blood group A and Bglycotopes, respectively, was isolated by Andersonet al., 2005. Recombinant EABase damages the bloodgroup A and B antigenicity of human type A and Berythrocytes with the release of A-Tri and B-Tri fromblood group A+ and B+ containing glycoconjugates.Here, the incomparable specificity of β-galactosidase wasuseful for studying the structure and function of bloodgroupA-containing and B-containing glycoconjugates.[193]
The recombinant endo-β-galactosidase (ABase), whichreleases A/B antigen was developed in 2009. Itremoved 82% of A antigen and 95% of B antigen fromhuman A/B red blood cells and concealed anti-A/Bantibody binding, also the complement activationeffectively. In vivo infusion into a blood type Ademonstrated the reduction of A antigen expressionin the glomeruli of kidney (85% at 3600 s, 9% at1.44 × 104 s, and 13% at 8.64 × 104 s) and the sinusoidsof liver (47% at 3600 s, 1% at 1.44 × 104 s, and 3% at8.64 × 104 s) without any grave adverse effects. Thissubstitute approach remains useful for minimizing
antibody removal and anti-B cell immunosuppressionas an adjuvant therapy of ABO incompatible kidney,liver, and possibly heart transplantation.[194]
Expression of β-galactosidase, synthesized by E. coliwithin muscle fibers has been demonstrated by Liuand Roffler, 2006. They conclude that repeatedintramuscular injections of β-galactosidase couldencourage strong immune responses amongimmuno-competent animals and cause eliminationof transduce muscle fibers by inflammatory cells.[195]
Recently, β-galactosidase from the mesoacidophilicfungus Bispora sp. MEY-1 under simulated gastricconditions has shown greater stability (100%) andhydrolysis ratio (>80%) toward milk lactose thancommercially available β-galactosidase from Aspergillusoryzae ATCC 20423. Thus, this β-galactosidase may bea better digestive supplement for alleviatingsymptoms associated with lactase deficiency.[196]
Recombinant β-galactosidases in cooperating one ortwo different peptides from the foot-and-mouthdisease virus (FMDV) nonstructural protein 3B perenzyme monomer, granted specified differentiationbetween sera of FMDV-infected pigs, cattle, andsheep, and those of native and conventionallyvaccinated animals. These FMDV infection-specificbiosensors can provide effectual and versatilealternatives for the serological differentiation ofFMDV-infected animals.[197]
SCOPE FOR FURTHER STUDY
β-galactosidase belongs to lactase group, whichhydrolyzes β-glycosidic bond formed betweengalactose and its organic moiety. Moreover, in somecases, it takes part in transgalactosidase reaction. Thedisadvantage of wild-type β-galactosidase is its loweractivity as well as it is inhibited by hydrolyzed product,glucose, and galactose. Therefore, research regardingtransgenic β-galactosidase synthesis will boost uphydrolytic activity as well as GOS synthesis. Although,synthesis mechanism of β-galactosidase is well establishedfor E. coli, still much more information is needed for othermicroorganism. Although, it is recommended thatprobiotic consortium (lactic acid bacteria) is much moreacceptable for food grade β-galactosidase synthesis, butbecause of product inhibition by lactic acid, researchon bioprocess design is in demand. More than 90-foldpurification of β-galactosidase is yet to achieved. In thiscircumstances, suitable purification device as well asdevelopment of purification protocol for high throughputis required. Research in the line of process intensificationof industrial β-galactosidase production and purificationshould be a great challenge. It is obvious, immobilizedβ-galactosidase provides several benefits for GOSsynthesis but due to lower activity at immobilizecondition; studies on the active site of enzyme and
Figure 6. Schematic diagram of galato-oligosaccharidesynthesis from lactose (Figure adapted from Palai et al.(2012)).[169]
Figure 5. Schematic diagram of galato-oligosaccharidesynthesis from lactose (Figure adapted from Neri et al.(2008)).[168]
Asia-Pacific Journal of Chemical Engineering β-GALACTOSIDASE: A REVIEW
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj
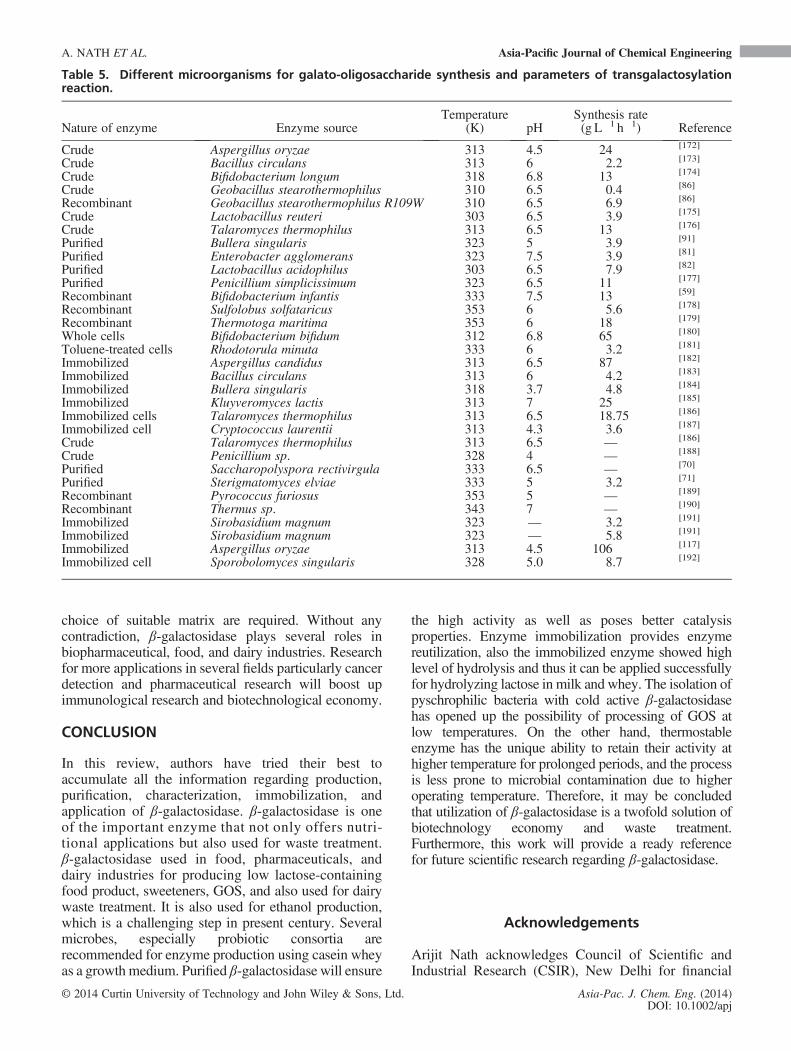
choice of suitable matrix are required. Without anycontradiction, β-galactosidase plays several roles inbiopharmaceutical, food, and dairy industries. Researchfor more applications in several fields particularly cancerdetection and pharmaceutical research will boost upimmunological research and biotechnological economy.
CONCLUSION
In this review, authors have tried their best toaccumulate all the information regarding production,purification, characterization, immobilization, andapplication of β-galactosidase. β-galactosidase is oneof the important enzyme that not only offers nutri-tional applications but also used for waste treatment.β-galactosidase used in food, pharmaceuticals, anddairy industries for producing low lactose-containingfood product, sweeteners, GOS, and also used for dairywaste treatment. It is also used for ethanol production,which is a challenging step in present century. Severalmicrobes, especially probiotic consortia arerecommended for enzyme production using casein wheyas a growth medium. Purified β-galactosidase will ensure
the high activity as well as poses better catalysisproperties. Enzyme immobilization provides enzymereutilization, also the immobilized enzyme showed highlevel of hydrolysis and thus it can be applied successfullyfor hydrolyzing lactose in milk and whey. The isolation ofpyschrophilic bacteria with cold active β-galactosidasehas opened up the possibility of processing of GOS atlow temperatures. On the other hand, thermostableenzyme has the unique ability to retain their activity athigher temperature for prolonged periods, and the processis less prone to microbial contamination due to higheroperating temperature. Therefore, it may be concludedthat utilization of β-galactosidase is a twofold solution ofbiotechnology economy and waste treatment.Furthermore, this work will provide a ready referencefor future scientific research regarding β-galactosidase.
Acknowledgements
Arijit Nath acknowledges Council of Scientific andIndustrial Research (CSIR), New Delhi for financial
Table 5. Different microorganisms for galato-oligosaccharide synthesis and parameters of transgalactosylationreaction.
Nature of enzyme Enzyme sourceTemperature
(K) pHSynthesis rate(g L�1 h�1) Reference
Crude Aspergillus oryzae 313 4.5 24 [172]
Crude Bacillus circulans 313 6 2.2 [173]
Crude Bifidobacterium longum 318 6.8 13 [174]
Crude Geobacillus stearothermophilus 310 6.5 0.4 [86]
Recombinant Geobacillus stearothermophilus R109W 310 6.5 6.9 [86]
Crude Lactobacillus reuteri 303 6.5 3.9 [175]
Crude Talaromyces thermophilus 313 6.5 13 [176]
Purified Bullera singularis 323 5 3.9 [91]
Purified Enterobacter agglomerans 323 7.5 3.9 [81]
Purified Lactobacillus acidophilus 303 6.5 7.9 [82]
Purified Penicillium simplicissimum 323 6.5 11 [177]
Recombinant Bifidobacterium infantis 333 7.5 13 [59]
Recombinant Sulfolobus solfataricus 353 6 5.6 [178]
Recombinant Thermotoga maritima 353 6 18 [179]
Whole cells Bifidobacterium bifidum 312 6.8 65 [180]
Toluene-treated cells Rhodotorula minuta 333 6 3.2 [181]
Immobilized Aspergillus candidus 313 6.5 87 [182]
Immobilized Bacillus circulans 313 6 4.2 [183]
Immobilized Bullera singularis 318 3.7 4.8 [184]
Immobilized Kluyveromyces lactis 313 7 25 [185]
Immobilized cells Talaromyces thermophilus 313 6.5 18.75 [186]
Immobilized cell Cryptococcus laurentii 313 4.3 3.6 [187]
Crude Talaromyces thermophilus 313 6.5 — [186]
Crude Penicillium sp. 328 4 — [188]
Purified Saccharopolyspora rectivirgula 333 6.5 — [70]
Purified Sterigmatomyces elviae 333 5 3.2 [71]
Recombinant Pyrococcus furiosus 353 5 — [189]
Recombinant Thermus sp. 343 7 — [190]
Immobilized Sirobasidium magnum 323 — 3.2 [191]
Immobilized Sirobasidium magnum 323 — 5.8 [191]
Immobilized Aspergillus oryzae 313 4.5 106 [117]
Immobilized cell Sporobolomyces singularis 328 5.0 8.7 [192]
A. NATH ET AL. Asia-Pacific Journal of Chemical Engineering
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

support as an SRF. The reported work is a part of aUniversity Grants Commission (UGC) Major project,entitled ‘Production and Purification of β-galactosidase from Milk Whey-based Lactic AcidBacteria using Fermentation and Membrane-basedSeparation Techniques”. The contribution of UGC isgratefully acknowledged.
REFERENCES
[1] M.L. Shuler, F. Kargi. Bioprocess Engineering BasicConcept, 2nd edn, Prentice-Hall Inc, New Jersey, 2002.
[2] M.L. Sanz, G.R. Gibson, R.A. Rastall. J. Agri. Food Chem.,2005; 53(13), 5192–5199.
[3] C.E. Rycroft, M.R. Jones, G.R. Gibson, R.A. Rastall.J. Appl. Microbiol., 2001; 91(5), 878–887.
[4] M.C.E. Lomer, G.C. Parkes, J.D. Sanderson. Aliment.Pharm. Ther., 2008; 27, 93–103.
[5] T. Sahi. Scand. J. Gastroenterol., 1994; 29(202), 7–20.[6] J. Jussila, M. Isokoski, K. Launiala. Scan. J. Gastroenterol.,
1970; 5, 49–56.[7] Z. Mlichová, M. Rosenberg. J. Food Nutr. Res., 2006; 45(2),
47–54.[8] S.P.S. Panesar, S. Kumari, R. Panesar. Enzyme Res., 2010,
1–16.[9] F. Jacob, J. Monod. J. Mol. Biol., 1961; 3, 318–356.[10] K.L. Anderson, G. Purdom. In Proceedings of the Sixth
International Conference on Creationism Creation ScienceFellowship and Dallas, Texas, (Ed.: A.A. Snelling),Institute for Creation Research, Pittsburgh, Pennsylvania,2008; pp. 149–163.
[11] B. Alberts, A. Johnson, J. Lewis, M. Raff, K. Roberts,P. Walter. Molecular Biology of the Cell, 5th edn, GarlandScience, New York, 2007; pp. 1392. ISBN:9780815341055.
[12] http://academic.pgcc.edu/~kroberts/Lecture/Chapter%207/regulation.html. Accessed date 18th june, 2013
[13] B. Görke, J. Stülke. Nat. Rev. Microbiol., 2008; 6(8), 613–624.[14] A.J.F. Griffiths, J.H. Miller, D.T. Suzuki, Lewontin R.C.,
W.M. Gelbart. An Introduction to Genetic Analysis. 7thedition, New York, 2000; ISBN-10: 0-7167-3520-2
[15] L.H. Hansen, S. Knudsen, S.J. Sønrense. Curr. Microbiol.,1998; 36(6), 341–347.
[16] S.A.-A. Ismail, Y. EI-Mohamady, W.A. Helmy,R. Abou-Romia, A.M. Hashem. Aust. J. Basic Appl.Sci., 2010; 4(10), 5051–5058.
[17] N.P. Laxmi, M.A. Mutamed, P.S. Nagendra. Int. Food Res.J., 2011; 18, 373–380.
[18] N.P. Laxmi, M.A. Mutamed, P.S. Nagendra. Int. Food Res.J., 2011; 18, 445–450.
[19] C.-F. Hwang, J.-H. Chang, J.-Y. Houng, C.-C. Tasi,C.-K. Lin, H.-Y. Tsen. Biotechnol. Bioprocess Eng., 2012;17, 827–834.
[20] S.K. Akolkar, A.D. Sajgure, S.S. Lele. Indian J.Biotechnol., 2006; 5, 184–188.
[21] T. Vasiljevic, P. Jelen. Innov. Food Sci. Emerg, 2001; 2(2),75–85.
[22] R. Rech, M.A.Z. Ayub. Process Biochem., 2007; 42, 873–877.[23] B.L.B. Perini, H.C.M. Souza, M. Kelbert, G.P. Apati,
A.P.T. Pezzin, A.L.S. Schneider. Chem. Eng. Trans., 2013;32, 991–996.
[24] H.S. Oberoi, S. Bansal, G.S. Dhillon. Int. J. Food Sci. Tech.,2008; 43, 1499–1504.
[25] R. Pinheiro, I. Belo, M. Mota. Lett. Appl. Microbiol., 2003;37, 438–442.
[26] A.M. Gupte, J.S. Nair. J. Sci. Ind. Res. India, 2010;69, 855–859.
[27] A.P. Menera, J.D.C. Ores, V.A. Ribeiro, C.A.V. Burkert,S.J. Kalil. Food Technol. Biotechnol., 2008; 46(1), 66–72.
[28] S. Bansal, H.S. Oberoi, G.S. Dhillon, R.T. Patil. Indian J.Microbiol., 2008; 48, 337–341.
[29] S. Barberies, R. Segovia. J. Chem. Technol. Biotechnol.,2002; 77, 706–710.
[30] R. Rech, M.Z. Ayub. Brazil. J. Chem. Eng., 2006; 23(4),435–442.
[31] P. Guimaraes, J. Klein, L. Domingues, J.A. Teixeira. Braz.J. Food Technol., 2005, 34–39.
[32] S.A.D. Bales, F.J. Castillo. Appl. Environ. Microb., 1979;37(6), 1201–1205.
[33] X. Li, J.W. Robbins Jr., K.B. Taylor. J. Ind. Microbiol.,1990; 5, 85–94.
[34] A. Jokar, A. Karbassi. J. Agr. Sci. Tech., 2011; 13, 577–584.[35] D.J.M. Kumar, M. Sudha, S. Devika, M.D. Balakumaran,
M.R. Kumar, P.T. Kalaichelvan. Ann. Biol. Res., 2012; 3(4),1712–1718.
[36] M. Asif, M.T. Khan, S.A. Rasool. J. Chem. Soc. Pak, 2009;31(2), 329–334.
[37] N. Akcan. Afr. J. Microbiol. Res., 2011; 5(26), 4615–4621.[38] M.V.R. Rao, S.M. Dutta. Appl. Environ. Microb., 1977;
34(2), 185–188.[39] N. Batra, J. Sing, U.C. Banarjee, P.R. Patnaik, R.C. Sobti.
Biotechnol. Appl. Biochem., 2002; 36, 1–6.[40] A. Pavani, M.S. Gadge, T. Prabhakar, P. Rupesh. Asian J.
Exp. Biol. Sci., 2011; 2(1), 23–27.[41] Z. Nagy, T. Kiss, A. Szentirmai, S. Biro. Protein Expres.
Purif., 2011; 21, 24–29.[42] S. Mirdamadi, N. Moazami, M.N. Gorgani. Sci. I. R. Iran.,
1997, 23–27.[43] S. Nizamuddin, A. Sridevi, G. Narasimha. Afr. J.
Biotechnol., 2008; 7(8), 1096–1100.[44] S.A. Shaikh, J.M. Khire, M.I. Khan. J. Ind. Microbiol. Biot.,
1997; 19, 239–245.[45] L. Domingues, N. Lima, J.A. Teixeira. Process Biochem.,
2005; 40, 1151–1154.[46] M.E.N.D. Macías, M.C.M.D. Nadra, A.M.S.D. Saad,
A.A.P.D.R. Holgado, G. Oliver. Curr. Microbiol.,1983; 9, 99–103.
[47] S. Chakraborti, R.K. Sani, U.C. Banerjee, R.C. Sobti. J. Ind.Microbiol. and Biotechnol., 2000; 24(1), 58–63.
[48] T.H. Nguyen, B. Splechtna, M. Steinböck, W. Kneifel,H.P. Lettner, K.D. Kulbe, D. Haltrich. J. Agric. FoodChem., 2006; 54(14), 4989–4998.
[49] A. Vetere, S. Paoletti. Biochim. Biophys. Acta, 1998; 1380,223–231.
[50] H. Fujimoto, M. Miyasato, Y. Ito, T. Sasaki, K. Ajisaka.Glycoconj. J., 1998; 15, 155–160.
[51] G.A. McFeters, W.E. Sandine, P.R. Elliker. J. Bacteriol.,1967; 93(3), 914–919.
[52] N.A. Greenberg, R.R. Mahoney. J. Food Sci., 1982;47, 1824–1835.
[53] W. Chen, H. Chen, Y. Xia, J. Zhao, F. Tian, H. Zhang.J. Dairy Sci., 2008; 91, 1751–1758.
[54] Y. Li, H. Wang, L. Lu, Z. Li, X. Xu, M. Xiao. Appl.Biochem. Biotechnol., 2009; 158, 192–199.
[55] O. Juajun, T.H. Nguyen, T. Maischberger, S. Iqbal,D. Haltrich, M. Yamabhai. Appl. Microbiol. Biotechnol.,2011; 89, 645–654.
[56] A.M. Biaikowska, H. Ciemlijski, K.M. Nowakowska,J. Kur, M. Turkiewicz. Arch. Microbiol., 2009; 191, 825–835.
[57] C.A. Hsu, R.C. Yu, C.C. Chou. World J. Microbiol.Biotechnol., 2006; 22(4), 355–361.
[58] V. Dumortier, C. Brassart, S. Bouquelet. Biotechnol. Appl.Biochem., 1994; 19, 341–354.
[59] M.N. Hung, B.H. Lee. Appl. Microbiol. Biotechnol., 2002;58, 439–445.
[60] K. Ohtsuka, A. Tanoh, O. Ozawa, T. Kanematsu, T. Uchida,R. Shinke. J. Ferment. Bioeng., 1990; 70, 301–307.
[61] Y. Tanaka, A. Kagamiishi, A. Kiuchi, T. Horiuchi.J. Biochem., 1975; 77, 241–247.
[62] D. Todorova-Balvay, I. Stoilova, S. Gargova, M.A.Vijayalakshmi. J. Mol. Recognit., 2006; 19, 299–304.
[63] F. Widmer, J.L. Leuba. Eur. J. Biochem., 1979; 100, 559–567.[64] W.H.M. van Casteren, M. Eimermann, L.A.M.
van den Broek, J.P. Vincken, H.A. Schols, A.G.J. Voragen.Carbohydr. Res., 2000; 329, 75–85.
[65] M. Becerra, E. Cerdán, M.I. González Siso. Biol. Proced.Online, 1998; 1, 48–58.
Asia-Pacific Journal of Chemical Engineering β-GALACTOSIDASE: A REVIEW
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

[66] M. Becerra, E. Cerdan, M.I. Gonzalez Siso. Biotechnol.Techniques, 1998; 12, 253–256.
[67] R.C. Dickson, L.R. Dickson, J.S. Markin. J. Bacteriol.,1979; 137, 51–61.
[68] J.A. Goncalves, F.J. Castillo. J. Dairy Sci., 1982; 65, 2088–2094.[69] L. Biermann, M.D. Glantz. Biochim. Biophys. Acta-
Enzymology, 1968; 167, 373–377.[70] M. Nakao, M. Hatada, Y. Kodama, T. Nakayama,
Y. Shibano, T. Amachi. Appl. Microbiol. Biotechnol.,1994; 40, 657–663.
[71] N. Onishi, T. Tanaka. Appl. Environ. Microbiol., 1995; 61,4026–4030.
[72] J.L. Berger, B.H. Lee, C. Lacroix. Biotechnol. Appl.Biochem., 1997; 25, 29–41.
[73] G.P. Dimri, X. Lee, G. Basile, M. Acosta, G. Scott,C. Roskelley, E.E. Medrano, M. Linskens, I. Rubelj,O.P. Smith. Proc. Natl. Acad. Sci., 1995; 92(20), 9363–9367.
[74] Q.Z.K. Zhou, X.D. Chen. Biochem. Eng. J., 2001; 9, 33–40.[75] M.A. Boon, A.E.M. Janssen, K.V. Riet. Enzyme Microb.
Technol., 2000; 26, 271–281.[76] M. Harju. Bull. Int. Dairy Fed., 1987; 212, 50–54.[77] E. Jurado, F. Camacho, G. Luzón, J.M. Vicaria. Enzyme
Microb. Technol., 2002; 31, 300–309.[78] M. Madani, M. R. Zarif-fard, F. Golchin-far. Arch. Razi.
Ins., 1999; 50, 93–98.[79] T. Nakagawa, R. Ikehata, T. Myoda, T. Miyaji, N.
Tomizuka. Protein Exp. Purif., 2007; 54, 295–299.[80] S.W.A. Hinz, L.A.M.V.D. Brock, G. Beldman, J.P.
Vincken, A.G.J. Voragen. Appl. Microbiol. Biotechnol.,2004; 66, 276–284.
[81] L. Lu, M. Xiao, X. Xu, Z. Li, Y. Li. Biochem. Biophys. Res.Commun., 2007; 356, 78–84.
[82] T.H. Nguyen, B. Splechtna, S. Krasteva, W. Kneifel, K.D.Kulbe, C. Divne, D. Haltrich. FEMS Microbiol. Lett.,2007; 269, 136–144.
[83] J.K. Jeong, O. Kwon, Y.M. Lee, D.B. Oh, J.M. Lee, S. Kim,E.H. Kim, T.N. Le, D.K. Rhee, H.A. Kang. J. Bacteriol.,2009; 191, 3011–3023.
[84] B.D. Lauro, A. Strazzulli, G. Perugino, F.L. Cara, E. Bedini,M.M. Corsaro, M. Rossi, M. Moracci. Biochim. Biophys.Acta, 2008; 1784, 292–301.
[85] A.R. Park, D.K. Oh. Appl. Microbiol. Biotechnol., 2010;85(5), 1427–1435.
[86] G. Placier, H. Watzlawick, C. Rabiller, R. Mattes. Appl.Environ. Microbiol., 2009; 75(19), 6312–6321.
[87] C.S. Kim, E.S. Ji, D.K. Oh. J. Appl. Microbiol., 2004; 97,1006–1014.
[88] M. Ladero, A. Santos, J.L. Garcia, A.V. Carrascosa, B.C.C.Pessela, F. Garcia-Ochoa. Enzyme Microb. Technol., 2002;30, 392–405.
[89] C.S. Kim, E.S. Ji, D.K. Oh. Biotechnol. Lett., 2003; 25,1769–1774.
[90] E. Ishikawa, T. Sakai, H. Ikemura, K. Matsumoto, H. Abe.J. Biosci. Bioeng., 2005; 99, 331–339.
[91] Y.J. Cho, H.J. Shin, C. Bucke. Biotechnol. Lett., 2003; 25,2107–2111.
[92] F.M. Pisani, R. Rella, C.A. Raia, C. Rozzo, R. Nucci, A.Gambacorta, M.D. Rosa, M. Rossi. Eur. J. Biochem.,1990; 187, 321–328.
[93] A.V. Fowler, I. Zabin. J. Biol. Chem., 1970; 245(19),5032–5041.
[94] R.H. Jacobson, X.J. Zhang, R.F. Dubose, B.W. Matthews.Nature, 1994; 369(6483), 761–766.
[95] J.C. Gebler, R. Aebersold, S.G. Withers. J. Biol. Chem.,1992; 267(16), 11126–11130.
[96] I.Y. Volkov, N.A. Lunina, O.V. Berezina, G.A.Velikodvorskaya, V.V. Zverlov. Mol. Biol., 2005; 39(6),799–805.
[97] G.K. Shoemaker, D.H. Juers, J.M. Coombs, B.W.Matthews, D.B. Craig. Biochemistry-US, 2003; 42(6),1707–1710.
[98] E.R. Nichols, J.M.A. Gavina, R.G. McLeod, D.B. Craig.Protein J., 2007; 26(2), 95–105.
[99] E.R. Nichols, D.B. Craig. Protein J., 2008; 27, 376–383.[100] D.B. Craig, E.R. Nichols.Electrophoresis, 2008; 29, 4298–4303.
[101] B.C.C. Pessela, R. Fernández-Lafuente, M. Fuentes, A.Vián, J.L. Garcı ́a, A.V. Carrascosa, C. Mateo, J.M. Guisán.Enzyme Microb. Tech., 2003; 32(3–4), 369–374.
[102] B.C. Pessela, C. Mateo, M. Fuentes, A. Vian, J.L. García,A.V. Carrascosa, J.M. Guisán, R. Fernández-Lafuente.Biotechnol. Prog., 2004; 20(1), 388–392.
[103] B.C. Pessela, G. Dellamora-Ortiz, L. Betancor, M. Fuentes,J.M. Guisán, R. Fernandez-Lafuente. Enzyme Microb. Tech,2007; 40, 310–315.
[104] Y. Numanoğlu, S. Sungur. Process Biochem., 2004; 39,703–709.
[105] A. Goel, R.K. Sharma, H.K.L. Tandon. J. Food. Sci.Technol., 2006; 43(5), 526–531.
[106] Z. Grosová, M. Rosenberg, M. Gdovin, L. Sláviková,M. Rebroš. Food Chem., 2009; 116, 96–100.
[107] R. Torres, C. Mateo, G. Fernández-Lorente. Biotechnol.Prog., 2003; 19(3), 1056–1060.
[108] M. Portaccio, S. Stellato, S. Rossi. Enzyme Microb. Tech.,1998; 23(1-2), 101–106.
[109] C. Bucke. J. chemical technol. Biotechnol, 1989; 46,pp. 162–163. http://onlinelibrary.wiley.com/doi/10.1002/jctb.v46:2.n/issuetoc
[110] S.T. Yang, M.R. Okos. Biotechnol. Bioeng., 1989; 33(7),873–885.
[111] A.P. Bakken, C.G. Hill, C.H. Amundson. Biotechnol.Bioeng., 1990; 36(3), 293–309.
[112] H. Hronska, M. Rosenberg, Z. Grosová. New Biotechnol.,2009; 25, 13–16.
[113] M.P. Mariotti, H. Yamanaka, A.R. Araujo, H.C. Trevisan.Braz. Arch. Biol. Techn., 2008; 51(6), 1233–1240.
[114] A. Kozhukharova, N. Kirova, K. Batsolva, S. Gargova, A.Dimova. Khranit. Promst., 1991; 40, 43–46.
[115] A. Rossi, A. Morana, I. Di Lernia, A. Ottombrino, M. DeRosa. Ital. J. Biochem., 1999; 48(2), 91–97.
[116] R. Gaur, H. Pant, R. Jain, S.K. Khare. Food Chem., 2006;97(3), 426–430.
[117] N. Albayrak, S.T. Yang. EnzymeMicrob. Tech., 2002; 31(4),371–383.
[118] N. Papayannakos, G. Markas, D. Kekos. The Chem. Eng.,1993; 52(1), B1–B12.
[119] D.F.M. Neri, V.M. Balcão, M.G. Carneiro-da-Cunha, L.B.Carvalho, J.A. Teixeira. Catal. Commu., 2008; 9(14),2334–2339.
[120] M. Decleire, W. De Cat, N. Van Huynh, J.C. Motte. ActaBiotechnol., 1987; 7(6), 563–566.
[121] I. Roy,M.N. Gupta. Process Biochem., 2003; 39(3), 325–332.[122] C.R. Carrara, A.C. Rubiolo. Chem. Eng. J., 1997; 65(2),
93–98.[123] C.R. Carrara, A.C. Rubiolo. Biotechnol. Prog., 1994; 10(2),
220–224.[124] X. Li, Q.Z.K. Zhou, X.D. Chen. Chem. Eng. Proc: Proc
Inten, 2007; 46(5), 497–500.[125] C. Giacomini, A. Villarino, L. Franco-Fraguas, F. Batista-
Viera. J. Mol. Catal-B Enzym., 1998; 4(5-6), 313–327.[126] B. Champluvier, B. Kamp, P.G. Rouxhet. App. Microbiol.
Biot., 1988; 27(5-6), 464–469.[127] M.I. González Siso, S. Suárez Doval. Enzyme Microb.
Tech., 1994; 16(4), 303–310.[128] K. Ovsejevi, V. Grazú, F. Batista-Viera. Biotechnol. Tech.,
1998; 12(2), 143–148.[129] E. Taqieddin, M. Amiji. Biomaterials, 2004; 25(10), 1937–1945.[130] M. Ladero, A. Santos, F. García-Ochoa. Enzyme Microb.
Tech., 2000; 27(8), 583–592.[131] J.L. Berger, B.H. Lee, C. Lacroix. Biotechnol. Tech., 1995;
9(8), 601–606.[132] B.C.C.H. Pessela, C. Mateo, M. Fuentes. Enzyme Microb.
Tech., 2003; 33(2-3), 199–205.[133] Z. Li, M. Xiao, L. Lu, Y. Li. Process Biochem., 2008; 43(8),
896–899.[134] M. Banerjee, A. Chakrabarty, S.K. Majumdar. Biotechnol.
Bioeng., 1982; 24(8), 1839–1850.[135] A. Dwevedi, A.M. Kayastha. Bioresour. Technol., 2009;
100(10), 2667–2675.[136] H.P.S. Makkar, O.P. Sharma. Biotechnol. Bioeng., 1983; 25,
592–597.
A. NATH ET AL. Asia-Pacific Journal of Chemical Engineering
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj

[137] M. Di Serio, C. Maturo, E. De Alteriis, P. Parascandola, R.Tesser, E. Santacesaria. Catal. Today, 2003; 79-80, 333–339.
[138] A.P. Bakken, C.G. Hill, C.H. Amundson. Biotechnol.Bioeng., 1992; 39(4), 408–417.
[139] M.J. Hernaiz, D.H.G. Crout. Enzyme Microb. Tech., 2000;27(1-2), 26–32.
[140] S.K. Khare, M.N. Gupta. Biotechnol. Bioeng., 1988; 31(8),829–833.
[141] O. Ariga, M. Kato, T. Sano, Y. Nakazawa, Y. Sano.J. Ferment. Bioeng., 1993; 76(3), 203–206.
[142] A. Bódalo, E. Gómez, M.F. Máximo, J.L. Gómez, J.Bastida. Biotechnol. Tech., 1991; 5(5), 393–394.
[143] V. Kéry, Š. Kučár, M. Matulová, J. Haplová. Carbohyd.Res., 1991; 209(C), 83–87.
[144] S.F. D’Souza, R. Kaul, G.B. Nadkarni. Biotechnol. Lett.,1985; 7(8), 589–592.
[145] S. Sungur, U. Akbulut. J. Chem. Technol. Biot., 1994; 59(3),303–306.
[146] M. Marrakchi, S.V. Dzyadevych, F. Lagarde, C. Martelet, N.Jaffrezic-Renault. Mater. Sci. Eng. C, 2008; 28, 872–875.
[147] S. Campuzano, B. Serra, D. Llull, J.L. Garcia, P. Garcia.Appl. Environ. Microb., 2009; 75(18), 5972–5980.
[148] R. Panesar, P.S. Panesar, R.S. Singh, J.F. Kennedy, M.B.Bera. Food Chem., 2007; 101, 786–790.
[149] Z. Cui. China Particuology, 2005; 3(6), 343–348.[150] S. Nakao, H. Osada, H. Kurata, T. Tsuru, S. Kimura.
Desalination, 1988; 70(1–3), 191–205.[151] S. Bhattacharjee, S. Ghosh, S. Datta, C. Bhattacharjee.
Desalination, 2006; 195(1-3), 95–108.[152] P. Sarkar, S. Ghosh, S. Dutta, D. Sen, C. Bhattacharjee.
Desalination, 2009; 249, 5–11.[153] S. Bhattacharjee, C. Bhattacharjee, S. Datta. J. Membrane
Sci., 2006a; 275(1-2), 141–150.[154] Z. Mlichová, M. Rosenberg. J. Food Nutri. Res., 2006;
45(2), 47–54.[155] G.D. Hatzinikolaoua, E. Katsifas, D. Mamma, A.D.
Karagouni, P. Christakopoulosb, D. Kekosb. Biochem.Eng. J., 2005; 24, 161–172.
[156] P. Guimaraes, K.J. Domingues, J.A. Teixeira. Braz. J. FoodTechnol., 2005, 34–39.
[157] W.B. Silveira, F.J.V. Passos, H.C. Mantovani, F.M.L.Passos. Enzyme Microb. Tech., 2005; 36(7), 930–936.
[158] S. Ozmihci, F. Kargi. Bioresour. Technol., 2007; 98(16),2978–2984.
[159] S. Ozmihci, F. Kargi. Bioprocess Biosyst. Eng., 2007; 30(2),79–86.
[160] M. Rosenberg, M. Tomáška, J. Kaňuch, E. Šurdík. ActaBiotechnol., 1995; 15(4), 387–390.
[161] A.C. Silva, P.M.R. Guimares, J.A. Teixeir, L. Domingues.J. Ind. Microbiol. Biot., 2010; 37(9), 973–982.
[162] M.K. Tahoun, T.M. El-Nemr, O.H. Shata. Eur. Food Res.Technol., 1999; 208(1), 60–64.
[163] S. Ozmihci, F. Kargi. Biochem. Eng. J., 2008; 42, 180–185.[164] M.A. Mehaia, M. Cheryan. Bioprocess Eng., 1990; 5(2),
57–61.[165] M. Staniszewski, W. Kujawski, M. Lewandowska. J. Food
Eng., 2009; 91, 240–249.[166] A. Gosling, G.W. Stevens, A.R. Barber, S.E. Kentish, S.L.
Gras. Food Chem., 2010; 121(2), 307–318.[167] M.A. Boon, A.E.M. Janssen, A.V.D. Padt. Biotechnol.
Bioeng., 1999; 64(5), 558–567.[168] D.F.M. Neri, V.M. Balcão, R.S. Costa, I.C.A.P. Rocha, E.
M.F.C. Ferreira, D.P.M. Torres, L.R.M. Rodrigues, J.L.B.Carvalho, J.A. Teixeira. Food Chem., 2009; 115(1), 92–99.
[169] T. Palai, S. Mitra, P.K. Bhattacharya. J. Bios. Bioeng., 2012;114(4), 418–423.
[170] S.J. Jung, R. Houde, B. Baurhoo, X. Zhao, B.H. Lee. Poult.Sci., 2008; 87(9), 1694–1699.
[171] R.C. Martinez, H.R. Cardarelli, W. Borst, S. Albrecht, H.Schols, O.P. Gutiérrez, A.J. Maathuis, B.D.D.M. Franco,E.C.D. Martinis, E.G. Zoetendal, K. Venema, S.M. Saad,H. Smidt. FEMS Microbiol. Ecol., 2013; 84(1), 110–123.
[172] K. Iwasaki, M. Nakajima, S. Nakao. Process Biochem.,1996; 31, 69–76.
[173] Z. Mozaffar, K. Nakanishi, R. Matsuno. J. Food Sci., 1985;50, 1602–1606.
[174] C.A. Hsu, S.L. Lee, C.C. Chou. J. Agric. Food Chem.,2007; 55, 2225–2230.
[175] B. Splechtna, T.H. Nguyen, M. Steinbock, K.D. Kulbe, W.Lorenz, D.Haltrich. J. Agric. FoodChem., 2006; 54, 4999–5006.
[176] P. Nakkharat, K.D. Kulbe, M. Yamabhai, D. Haltrich.Biotechnol. J., 2006, 1,633–1,638.
[177] R. Cruz, V.D.A. Cruz, J.G.B.D. Oliveira, M. Khenayfes, C.Dorta, L.H.D.S. Oliveira, E. Ardiles, A. Galli. Bioresour.Technol., 1999; 70, 165–171.
[178] H.Y. Park, H.J. Kim, J.K. Lee, D. Kim, D.K. Oh. World J.Microbiol. Biotechnol., 2008; 24, 1553–1558.
[179] E.S. Ji, N.H. Park, D.K. Oh. World J. Microbiol.Biotechnol., 2005; 21, 759–764.
[180] G. Tzortzis, A.K. Goulas, G.R. Gibson. Appl. Microbiol.Biotechnol., 2005; 68, 412–416.
[181] N. Onishi, K. Yokozeki. J. Ferment., Bioeng., 1996; 82,124–127.
[182] P. Zheng, H. Yu, Z. Sun, Y. Ni, W. Zhang, Y. Fan, Y. Xu.Biotechnol. J., 2006, 1,1464–1,1470.
[183] Z. Mozaffar, K. Nakanishi, R. Matsuno. Appl. Microbiol.Biotechnol., 1986; 25, 229–231.
[184] H.J. Shin, J.W. Yang. J. Microbiol. Biotechnol., 1998,8,484–8,489.
[185] S. Chockchaisawasdee, V.I. Athanasopoulos, K. Niranjan,R.A. Rastall. Biotechnol. Bioeng., 2004; 89, 434–443.
[186] P. Nakkharat, K.D. Kulbe, M. Yamabhai, D. Haltrich.Biotechnol. J., 2006; 1, 633–638.
[187] O. Ozawa, K. Ohtsuka, T. Uchida, N.S.K. Gakkaishi. NipponShokuhin Kogyou Gakkaish s, 1989; 36, 898–902.
[188] M.J. In, H.J. Chae. Korean J. Appl. Microbiol. Biothechnol.,1998; 26, 40–44.
[189] M.E. Bruins, M. Strubel, J.F.T.V. Lieshout, A.E.M. Janssen,R.M. Boom. Enzyme Microb. Technol., 2003; 33, 3–11.
[190] K. Akiyama, M. Takase, K. Horikoshi, S. Okonogi. Biosci.Biotechnol. Biochem., 2001; 65, 438–441.
[191] N. Onishi, I. Kira, K. Yokozeki. Lett. Appl. Microbiol.,1996; 23, 253–256.
[192] T. Sakai, H. Tsuji, S. Shibata, K. Hayakawa, K. Matsumoto.J. Gen. Appl. Microbiol., 2008; 54, 285–293.
[193] K.M. Anderson, H. Ashida, K. Maskos, A. Dell, S.C. Li, Y.T. Li. J. Biolog. Chem., 2005; 280(9), 7720–7728.
[194] T. Kobayashi, D. Liu, H. Ogawa, Y. Miwa, T. Nagasaka, S.Maruyama, Y. Li, A. Onishi, M. Iwamoto, T. Kuzuya, K.Kadomatsu, K. Uchida, A. Nakao. Transpl. Immunol.,2009; 20, 132–138.
[195] A.B. Liu, S.R. Roffler. Tzu. Chi. Med. J., 2006; 18(4), 259–265.[196] H. Wang, H. Luo, Y. Bai, Y. Wang, P. Yang, P. Shi, W.
Zhang, Y. Fan, B. Yao. J. Agric. Food Chem., 2009; 57,5535–5541.
[197] M.T. Sanchez-Aparicio, M.F. Rosas, R.M. Ferraz, L.Delgui, J.J. Veloso, E. Blanco, A. Villaverde, F. Sobrino.Clin. Vaccine Immunol., 2009; 16(8), 1228–1235.
Asia-Pacific Journal of Chemical Engineering β-GALACTOSIDASE: A REVIEW
© 2014 Curtin University of Technology and John Wiley & Sons, Ltd. Asia-Pac. J. Chem. Eng. (2014)DOI: 10.1002/apj




















