
Large-scale analysis by SAGE reveals new mechanisms of v-erbA oncogene action
Upload
independentCategory
view
1download
0
Adenovirus vaccination against neu oncogene exerts long-term protection
from tumorigenesis in BALB/neuT transgenic mice
Pasquale Gallo1, Sridhar Dharmapuri1, Maurizio Nuzzo1, Daniele Maldini1, Barbara Cipriani1, Guido Forni2
and Paolo Monaci1*
1Molecular and Cell Biology Department, I.R.B.M.P. Angeletti, Pomezia (Roma), Italy2Department of Clinical and Biological Sciences, University of Turin, Orbassano (Turin), Italy
The transforming rat HER2/neu oncogene (neu), when embeddedin the genome of transgenic BALB/c (neuT) mice, provokes the de-velopment of an invasive carcinoma in each of their 10 mammaryglands. We used the neuT mice model system to evaluate the immu-nization efficiency and the protective effect of intramuscular injec-tion of adenovirus (Ad) and/or of DNA with electrostimulation(DNA1ES), both expressing the rat p185neu protein. A neu cDNAsequence, which exclusively contains codons preferred by highlyexpressed mammalian genes, was used in this study. This ‘‘opti-mized’’ cDNA displayed higher expression in cultured cells andgreater cell-mediated response than the original gene when injectedas DNA1ES. Ad expressing the optimized sequence (Ad5-neu.opt)induced a higher immune response, as measured by the frequencyof IFN-c-secreting spleen cells and antibody titers. Different Ad/DNA combinations and immunization schedules confirmed the su-periority of Ad5-neu.opt in inducing a strong Th1-skewed humoraland CD81 cell-mediated response. Two Ad5-neu.opt injections of109 viral particles at week 10 and 12 were sufficient to induce thehighest response, which persisted at detectable levels up to 33 weeksof age. Anti-Ad5 antibodies elicited by previous injections neutral-ized the effect of an additional Ad5-neu.opt immunization at week19. A group, which received 3 injections of DNA1ES at week 23, 27and 31, in addition to the 3 Ad injections at week 10, 12 and 19showed an increased frequency of IFN-c1, CD81 PBMC at week25, which persisted at detectable levels till week 38. Ad5-neu.optadministration at 10 and 12 weeks of age had a significant impacton tumor progression. At 44 weeks, 40% of the mice were com-pletely protected from tumors with a mean tumor of 3.8. In con-trast, control mice developed 10 tumors and died by week 27. Vacci-nation blocked the tumor development at the atypical hyperplasiastage present at the time of treatment. Tumors developing at latertimes express reduced levels of rat p185
neuprotein.
' 2006 Wiley-Liss, Inc.
The first-line treatment for neoplasia involves the surgical re-moval of the tumor mass, which can be followed by chemotherapyor radiotherapy aimed at eliminating the recurrence of the primarytumor. These protocols are successful when tumor masses aredetected at early stages of development. However, in a still largenumber of cases, delayed detection of the tumor mass and/or the in-herent properties of the tumor-type let tumor cells disseminatethroughout the body promoting metastasis. In these cases, the besttherapy should keep the whole body in check for an extended pe-riod of time, ready to detect and specifically eliminate tumor cellsin the early phase of tumorigenesis. Antitumor vaccination appearsto be the most appropriate approach to meet these needs.
In the past decade, a significant amount of knowledge about themechanisms regulating the immune response has been accumulated.In addition, several proteins whose expression is preferentially associ-ated with tumors have been identified and cloned (tumor associatedantigens, TAAs). Animal studies have proved that delivering TAAsby genetic route raises an effective immune response against theTAA-bearing tumor cells. Here, the gene coding for the target antigenis expressed intracellularly, thus having the potential of inducing aspecific T-cell response. The latter is recognized to play a critical rolein effectively controlling tumor growth and spread of metastases.
Gene-based vaccination using viral vectors is emerging as aneffective route for delivering the selected TAA to the immune sys-tem. Viral infection induces inflammatory reactions, which criticallycontribute in stimulating an antigen specific immune response. First
generation adenoviral vectors efficiently transduce a wide range ofcell types, promote effective processing of the TAA into both MHCClass I and II pathways. This stimulates a potent cell-mediated im-munity as well as the induction of a robust antibody response.Adenoviral vectors have shown good results in animal models andimmunomodulatory factors delivered by Ad5 are in advanced phaseof several clinical trials.1–8
Naked DNA immunization is another highly attractive approachfor genetic vaccination. When plasmid DNA expressing a givenantigen is injected in the mouse muscle, an antigen-specificimmune response is generated. The efficiency of this process canbe considerably improved if DNA injection is followed by an elec-trical stimulation (ES) of the muscle.9 Although less effective thanviral vaccination, DNA immunization does not raise an immuneresponse against the vehicle and lacks toxicity. In addition, plas-mid DNA is chemically defined can be produced in large quanti-ties, purified to homogeneity, and is relatively stable.
We selected BALB/neuT mice as in vivo tumor model system forassessing the efficacy of different vaccination protocols employingadenovirus (Ad) and/or DNA1ES. These are inbred BALB/c miceengineered to overexpress the activated (V664E) rat neu oncogeneunder the control of the mouse mammary tumor virus promoter(MMTV-LTR; referred to as neuT mice10). The transgene encodesa 185 kDa transmembrane tyrosine kinase receptor (rat p185neu),which is prevalently expressed in mammary glands in these mice.At 3 weeks of age, female neuT mice start a process of rapid devel-opment of tumors involving all the mammary glands.11,12 Thisspontaneous tumorigenesis recapitulates many features of the devel-opment of human breast tumors.
We previously reported the vaccination of neuT mice using thehuman HER2/neu gene.13 This xenogeneic vaccination elicited aremarkable antigen-specific humoral and cell-mediated immuneresponse. This response, however, was poorly crossreacting withthe rat p185neu protein and only partially impacted on tumor de-velopment. Here, we report a detailed characterization of a synge-neic vaccination using Ad and/or DNA1ES delivery methodolo-gies. We demonstrate that vaccinating at 10 and 12 weeks of agecompletely protected 40% of the mice up to 45 weeks.
Material and methods
Cell lines
Mouse fibroblast NIH 3T3 and mouse liver NMuLi cells wereobtained from the American Type Culture Collection (ATCC,Rockville, MD). Per.C6 cells were obtained from Crucell (Leiden,The Netherlands), and human embryonic kidney HEK-293 from
Grant sponsor: Ministero dell’Istruzione, dell’Universita e della Ricerca;Grant number: FIRB RBME017BC4.*Correspondence to: Molecular and Cell Biology Department,
I.R.B.M.P. Angeletti, Via Pontina km 30.6, 00040-Pomezia (Roma), Italy.Fax:139-06-91093-654. E-mail: [email protected] 12 June 2006; Accepted after revision 17 July 2006DOI 10.1002/ijc.22274Published online 9November 2006 inWiley InterScience (www.interscience.
wiley.com).
Int. J. Cancer: 120, 574–584 (2006)' 2006 Wiley-Liss, Inc.
Publication of the International Union Against Cancer

Graham.14 N202.1ArNeu1 is a cloned cell line established in vitrofrom a lobular mammary carcinoma that arose in FVB femalemouse (H-2q) transgenic for the rat HER2 proto-oncogene driven bythe MMTV promoter15 (kindly provided by P.L. Lollini, Universityof Bologna). N202.1ArNeu1 expresses high levels of rat p185 pro-tein on the membrane whereas N202.1ErNeu2 does not show detect-able levels of the same protein, as measured by cytofluorimetricanalysis. D2F2 was derived from a spontaneous mammary tumorthat arose in BALB/c hyperplastic alveolar nodule line.16 Mousemammary tumor cell line D2F2 and stable clone expressing consti-tutively neu (D2F2/neu) were provided by Zen-Wei and co-workers17 (Karmanos Cancer Institute, Wayne State University).
Peptides
Three hundred and twelve 15-amino acid peptides, overlapping by11 amino acids, were designed to span the entire rat neu sequence.These peptides, which also included 7 peptides designed to overcomeinsolubility problems, were synthesized by SynPep (Dublin, CA). Thesame company also synthesized the 9 amino acid peptides described.All peptides were shown to be >90% pure by HPLC and were usedwithout HPLC purification. Peptides were reconstituted at a maximalconcentration of 35 mg/ml in DMSO and stored at280�C.
Immunogens
pVIJ-neu.wt. neu cDNA was obtained by PCR-amplification oftemplate plasmid pRNeu-cDNA3/1718 by the rat_neu. for-1 (50-CCGATATCGCCGCCGCCATGATCATCATGGAGCTGGCGG-CC-30, coding sequence is underlined) and rat_neu.rev-1 (50-CAGTCGACTCATACAGGTACATCCAGGCCTAG-30) primers.The resulting PCR product was EcoRV-SalI digested and clonedinto the corresponding sites of mammalian expression plasmidpVIJ-019 to obtain pVIJ-neu. The cDNA was fully sequenced.In vitro expression was confirmed in HEK-293, NMuli and NIH-3T3 cell lines by flow cytometry.
pVIJ-neu.opt. Sequencing the rat transgene in neuT micerevealed that it contained both the LLVSS to CGLE mutation atposition 509 as well as the V663E mutation, which are separatelyfound in AAM50093 and NP_058699 sequences, respectively. Asynthetic neu cDNA (neu.opt) was designed to incorporate mam-malian-preferred codons for each amino acid residue, in addition tothe K573A mutation. The neu.opt sequence was assembled inpUC19 plasmid with a Kozak optimized sequence (50-GCCGCC-ACC-30) immediately upstream of the translation start site. Theneu.opt sequence was extracted by PmeI-SalI digestion from pUC-neu.opt and cloned into the EcoRV-SalI sites of pVIJ vector.19
In vitro expression of pVIJ-neu.opt was confirmed by flow cytome-try in HEK-293, NMuLi and NIH3T3 cell lines.
Ad5-neu.opt. The neu.opt sequence was obtained from PmeI-SalI digestion of pUC19-neu.opt and cloned in the correspondingsites of the shuttle plasmid polyMRKDE1.20 The resulting plasmidpMRKDE1_neu.opt contained a human CMV promoter driving theexpression of the neu.opt cDNA, followed by the bovine growth hor-mone polyadenylation signal. Plasmid pMRKDE1_neu.opt wasrecombined with the adenoviral backbone plasmid pAd5-HV0 togenerate the preadenoviral plasmid pAd5-neu.opt. The latter was lin-earized by ClaI digestion and transfected into PerC6 cells to generateAd_neu.opt recombinant Ad. The virus was grown in large quantitiesby multiple rounds of amplification and purified using Puresyn’sAdenopure Kit (Adenopure kit, Puresyn, Malvern, PA) according tomanufacturer’s instructions. Viral DNA was extracted by proteinaseK digestion and genomic integrity was confirmed by restriction anal-ysis. Virus particle (vp) titre was determined spectrophotometricallyconsidering that 1 U of optical density at 260 nm corresponds to1.13 1012 particles/ml.20 In vitro expression was confirmed by flowcytometric analysis of infected murine liver NMuli cells.
In vitro expression
Plasmids pVIJ-neu or pVIJ-neu.opt were LipofectamineTM-cotransfected (Gibco BRL Invitrogen, Groningen, The Nether-
lands) with a plasmid expressing EGFP under the control of CMVpromoter (pCMS-EGFP, Clontech Labs, Mountain View, CA) usedas a transfection efficiency control. Cotrasfections were performedin HEK-293, NMuli and 3T3 cells. On the other hand, murine liverNMuli cells were infected with Ad5-HER2/neu_opt. After 48 hr ofincubation, cells were collected using trypsin and stained by astandard indirect immunofluorescence procedure using an anti-HER2/neu mAb Ab-4 (Oncogene, Boston, MA) as primary anti-body, followed by FITC-conjugated rabbit anti-mouse Ig (DAKO,Glostrup, Denmark). All samples were acquired within 24 hr ofstaining using a FACSCalibur flow cytometer and CellQuest soft-ware (BD Immunocytometry Systems, San Diego, CA).
Mice
Female BALB/c mice (H-2d) transgenic for the transformingneuT oncogene (neuT1/neuT2) under the transcriptional control ofthe MMTV promoter10,12 and transgene negative (neuT2/neuT2)were bred under specific pathogen-free conditions using CharlesRiver Breeding Laboratories (Calco, Italy). Mice were screened forthe presence of the transgene by duplex PCR as described.13 Briefly,genomic DNA was extracted from the mouse-tail and duplex PCRwas performed on this substrate using two pairs of primers in orderto amplify part of MMTV promoter region and mouse bglobin (aspositive internal control). Mice that showed complete protectionfrom tumorigenesis were confirmed as neuT1/neuT at the end ofthe study. In all manoeuvres, mice were treated in accordance withEuropean guidelines. In particular, at the time of vaccination, micewere fully anesthetized with ketamine (Imalgene 500; Merial Italia,Milano, Italy) at 100 mg/kg of body weight and xylazine (Xilor,BIO 98; S. Lazzaro, Bologna, Italy) at 5.2 mg/kg sec.
Monitoring several hematological parameters in mice, whichexperienced immunization did not reveal any variation associatedto the vaccination treatment.
Immunization protocol
pVIJ-neu. neuT mice were vaccinated at 6 and 8 weeks of ageby injecting a total amount of 100 lg of pVIJ-neu (.wt or .opt)plasmid DNA into the quadriceps muscle (100 lg in 50 ll of phys-iological solution per leg) followed by ES as previously described.Sera were collected at week 5 (preimmune serum) and week 10(2 weeks after boost). Spleen cells were obtained at week 11(3 weeks after boost).
Ad_neu.opt. neuT mice were vaccinated at 10 and 12 weeks ofage by injecting 109 vp of Ad_neu.opt into the quadriceps muscle(109 vp in 50 ll of physiologic solution per leg). Sera were col-lected at week 9 (preimmune sample) and 14 (2 weeks afterboost). Spleen cells were also obtained at week 14.
Tumor challenge assay
Female BALB/c or neuT mice were injected intramuscularly at8 and 11 week of age with pVIJ-neu.opt (100 lg) followed by ESor Ad_neu.opt (109 vp). Two weeks later, mice were challengedsubcutaneously in the right flank with 2 3 105 D2F2/neu cells.Development of spontaneous tumors in mammary glands andtransplanted tumors in the same neuT mice was monitored byweekly palpation. A control group of mice was immunized but nottumor-challenged; additional control groups were injected withthe empty vectors (pVIJ-0 or Ad5-0) and challenged as describedearlier. Log rank test calculated a significant difference (p < 4exp-4) between Ad5-neu.opt and Ad5-0. About 2 3 105 cells gaverise to carcinoma with identical features in all control groups.
Evaluation of spontaneous tumor growth in neuT mice
neuT females were inspected weekly and tumor masses weremeasured with callipers in 2 perpendicular diameters. Progressively,growing masses>3 mm in mean diameter were regarded as tumors.Growth was monitored until all mammary glands displayed a palpa-ble tumor or until a tumor exceeded an average diameter of 10 mm,
575ADENOVIRUS VACCINATION AGAINST neu ONCOGENE

at which time mice were sacrificed for humane reasons. Otherwisespecified, surviving neuT mice were sacrificed at week 33.
ELISPOT assay
Mouse spleen cells secreting IFN-g in an antigen-specific man-ner were detected using a standard enzyme-linked immunospot(ELISPOT) assay like previously described.21 Multiscreen 96-wellfiltration plates were coated with an affinity-purified rat anti-mouseIFN-g antibody. After overnight incubation, plates were washedwith PBST (0.005% Tween in PBS) and incubated with R10 me-dium (RPMI 1640 supplemented with 10% fetal calf serum, 2 mML-glutamine, 50 U of penicillin per ml, 50 lg of streptomycin perml, 10 mM HEPES and 50 lM 2-mercaptoethanol) for 2 hr at 37�Cto block nonspecific binding. Cells from the spleen of 3–5 immu-nized mice were purified, counted by Guava via count (GuavaTechnologies, Hayward, CA), resuspended in R10 medium (RPMI1640 supplemented with 10% fetal calf serum, 2 mM L-glutamine,50 U of penicillin per milliliter, 50 lg of streptomycin per ml,10 mM HEPES and 50 lM 2-mercaptoethanol) and incubated for15 hr in the presence of 6 lg/ml of peptides, at a density of 2.5 3105 – 5 3 105 cells/well. After extensive washing with PBST, bio-tinylated rat anti-mouse IFN-g antibody (PharMingen; San Diego,CA) was added. The plates were incubated at 4�C overnight andthen washed with PBST prior to the addition of streptavidin-alka-line phosphatase (PharMingen; San Diego, CA). After incubationfor 2 hr at room temperature, the plates were extensively washedwith PBST and developed by incubating with a one-step nitrobluetetrazolium-5-bromo-4-chloro-3-indolylphosphate substrate (Pierce,Rockford, IL) for 5–15 min for the development of spots. Rinsingthe plates in water stopped the reaction. DMSO and concanavalin A(10 lg/ml) were included as background and positive control foreach sample. Spots were counted by computer-assisted imaginganalysis (AID ELR02 coupled with AID ELISPOT 2.6 Software,Strassburg, Germany).
Flow cytometric analysis
IFN-g intracellular staining. Intracellular IFN-g productionwas measured according to BD Pharmingen standard protocol asdescribed previously.13 Briefly, spleen or peripheral blood derivedcells were obtained according to standard protocols. Cells werecounted using Guava ViaCount. An equivalent number of cellsfrom 3 or more mice were pooled and analyzed. About 2 3 106
spleen cells or 3 3 106 PBMC were cultured for 15 hr in R10 me-dium in the presence of 6 lg/ml of single or pool of peptides andBrefeldin A as protein transport inhibitor. Staphylococcus Entero-toxin B at 10 lg/ml and DMSO were tested as positive and back-ground control, respectively. Before staining of surface antigens, Abanti-mouse CD16/CD32 was used to reduce nonspecific immunoflu-orescent signal. The specific signal was obtained with APC-anti-mouse CD3e, PE-anti-mouse CD4 and PerCP-anti-mouse CD8a.The cells were then washed, fixed, permeabilized and stained forintracellular IFN-g using FITC-conjugated mAb.
FACS-Ab titration
MAb Ab-4 (Oncogene, Boston, MA) was used to establish quan-titative measurement of anti-rat p185neu antibodies by flow cytome-try using mouse N202.1ArNeu1 as described.13 Briefly, Ab-4 wasserially diluted and used to stain mouse N202.1ArNeu1. Cells werecollected and incubated with mAb anti-CD16/CD32 (Pharmingen,San Diego, CA) at 4�C for 20 min. Serial dilutions of mAb Ab-4were then added and left at 4�C for 45 min. After washing, FITC-conjugated rabbit anti-mouse Ig (DAKO, Glostrup, Denmark) wasused for detection of binding to rHER2; finally, propidium iodidewas used to gate out dead cells. Mean channel fluorescence (mcf)was recorded and the background signal generated by the second-ary Ab (typically around 10) was subtracted. The range of Ab-4antibody concentration, which correlated with mcf in a linear fash-ion, was determined by regression analysis and found to be 5–1000lg/ml (R2 5 0.998). Serum antibodies reacting with rat HER2 were
measured through the same procedure. Pools of 4–5 serum col-lected at specified week of age were performed and serial dilutionswere measured; the value obtained from the preimmune serum poolat the lowest dilution was subtracted. No signal was detected usingcontrol N202.1ErNeu2 cells. Data were expressed as Ab-4 equiva-lent on the basis of the standard curve.
To evaluate Th1/Th2 switch, the IgG2a/IgG1 Ab ratio, devel-oped against rat p185, was measured using N202.1ArNeu1. Thesame pools of sera used for Ab titration was diluted (1/40) and usedto stain cells by a standard indirect immunofluorescence procedure.In parallel fashion, FITC-conjugated goat Abs specific anti-mouseIgG2a or IgG1 (Caltag, Burlingame, CA) were used and relativemcf was recorded. Background signal generated by serum sampleof untreated mice was subtracted; DBkg values were used to obtainthe specified Ig2a/IgG1 ratio.
Titration of Ad neutralizing antibodies
neuT mice were injected with 109 vp of Ad5 at week 10 and atweek 12. Serum samples before immunization were collected andpooled (prebleed). Similarly, samples from the same mice werecollected 2 weeks after the boost and pooled (bleed). These sam-ples were analyzed for the presence of anti-Ad5 antibodies byELISA as described.22 The specific absorbance of each sample(spec.Abs) was calculated by subtracting its bleed absorbance fromthat of the prebleed sample. The titer of the serum was defined asthe serum dilution reaching 50% neutralization activity.
Histopathological examination
neuT mice were immunized at 10 and 12 weeks of age with 109
vp of Ad5-HER2/neu_opt. Histological and immunohistochemicalanalysis were performed on at week 16, 24 and 52 on Ad5-immu-nized mice and at week 10, 16 and 24 on untreated control groups.Three mice for each time of analysis were used. For histologicalevaluation, samples of all 10 mammary glands for each mouse,were fixed in 10% neutral-buffered formalin, embedded in paraffin,sectioned at 5 lm thickness and stained with hematoxylin and eo-sin. Detailed microscopic observations have been reported for indi-vidual animals in the study. Duct hyperplasia defines the presenceof increased number of epithelial layers (usually 3 or more) abovethe duct basement membrane, increased cellular basophilia, mildincrease in the nuclear-cytoplasmic ratio and stratification of cells.Atypical hyperplasia defines moderate degrees of epithelial prolif-eration with tufs, micropapillae and bridges associated with in-creasing atypia and basophilia. M-Adenocarcima adenoid differen-tiation, termed Type A by Dunn classification, is characterized bythe presence of acini lined by a single layer of cells sometimesshowing secretory activity. M-Adenocarcima solid differentiation,termed Type B by Dunn classification, identifies uniform tumorcells resembling lymphoid cells, but arranged in nests and cords.
Tumoral cells, in both differentiation stages, are characterized byan increased nucleus/cytoplasm ratio and prominent nucleolus. Mi-totic figure is often found and stroma within the tumor is usuallyscanty. Area of necrosis and haemorrhage may occur. To evaluatethe expression of rat p185 Ag and proliferating cell nuclear antigen(PCNA), paraffin-embedded sections were tested with anti-neu Ab(C-18)-G (Santa Cruz Biotechnology, Santa Cruz, CA) and withmouse monoclonal antibody anti-PCNA (Novocastra Laboratories,UK), respectively. Selected sections were incubated for 5 min with3% hydrogen peroxide. The specimens were then incubated for 1 hrwith primary antibody rat p185 Ag and PCNA (working dilution1:100 for both primary antibodies), followed by sequential 10-minincubations with a biotinylated link antibody and peroxidase-labelled streptavidin. (LSAB2 System-HRP, DAKO, Glostrup,Denmark). Stains were completed after few minutes of incubationwith the substrate chromogen (Liquid DAB Substrate ChromogenSystem, DAKO, Glostrup, Denmark). Positive cells, tested withthe corresponding antibodies, were evaluated under microscope(4003) and their presence/absence was defined as: Absent2,scarcely1/2, moderately1 and frequently11.
576 GALLO ET AL.

Statistical analysis
Differences in tumor incidence were evaluated using the Mantel-Haenszel log rank test and differences in tumor/mouse numberssing Wilkoxon’s rank sum test. Student’s t-test was used to identifyIFN-g ELISPOT values statistically significant values in ELISPOTassay and when compared with IFN-g or antibody titers betweendifferent groups of mice.
Results
Expression of a synthetic neu gene using preferredmammalian codons
A synthetic rat neuT gene was generated, which contained ex-clusively codons that are found preferentially in highly expressedmammalian genes.23 This codon ‘‘optimized’’ cDNA (referred toas neu.opt) maintained 81% nucleotide identity to the originalneuT gene (referred to as neu.wt). GC frequency increased from57.6% in neu.wt to 66.7% in the neu.opt sequence whereas thepresence of A or T in the third codon position decreased from34.4% (neu.wt) to 7.2% (neu.opt). In addition, a single amino acidmutation (K757A) was introduced in the neu.opt sequence. Thismutation destroys the ATP binding site, thus abrogating the tyro-sine kinase activity, eliminating the downstream signaling eventsand the oncogenic activity.24
Comparatively, to assess the level of expression of neu.wt andneu.opt gene in mammalian cells, both cDNA sequences werecloned under the transcriptional control of the human cytomegalo-virus (CMV) immediate-early promoter with its intron A sequence,and followed by the bovine growth hormone poly-adenylation sig-nal sequence (plasmid pVIJ-neu.wt and pVIJ-neu.opt, respec-tively). Sequences, immediately upstream of the coding region,were converted to a Kozak consensus translation initiation site.25,26
Human embryonic kidney HEK-293, mouse liver NMuLi andmouse fibroblast NIH3T3 cells were transiently transfected withdifferent amounts of pVIJ-neu.wt or pVIJ-neu.opt plasmid DNAand rat p185neu expression measured by flow cytometry. Rat pro-tein expressed by the neu.opt gene was estimated to increase from1.1 to 2.4 MFC units when compared with that obtained by theneu.wt gene, corresponding to a 2–3-fold enhancement (Fig. 1a).
The codon-optimized sequence preceded by the CMV promoterwas inserted by bacterial homologous recombination in the Ad5genome, replacing the E1 region. The Ad5-neu.opt virus thus gen-erated induced high levels of rat p185neu upon infection of mouseliver NMuli cells. We were unable to rescue the virus harboringthe neu.wt sequence.
Identification of immunodominant T-cell epitopes in ratp185neu protein
To identify the immunodominant T-cell epitopes of rat p185neu
in BALB/c mice (H-2d), 6-weeks-old female BALB/c mice wereimmunized by injecting 109 vp of Ad5-neu.opt in the quadricepsmuscles. An additional injection was performed after 2 weeks. A
FIGURE 1 – Comparative expression and immunogenicity of neu.optand neu.wt genes. (a) Expression of rat p185neu in transfected mam-malian cell lines. NIH3T3 (empty circles), NMuli (filled triangle) andHEK-293 (filled circles) cells were transiently transfected with the in-dicated amounts of pVIJ-neu.wt or pVIJ-neu.opt plasmid DNA. Trans-fection efficiency was normalized by cotransfecting a plasmid ex-pressing an enhanced green fluorescence protein (EGFP) under thecontrol of the CMV promoter. Rat p185neu expression was measuredby flow cytometry and expressed as difference between mean fluores-cence induced by the neu.opt and that obtained by the neu.wt gene. (b)Anti-rat p185neu humoral response in neuT vaccinated mice. Micewere injected with 109 vp of Ad5-neu.opt at week 6 and 8 and anti-ratp185neu antibodies measured at week 10. A standard reference curvewas generated incubating N202.1ArNeu1 cells with various concentra-tions of mouse anti-rat p185neu MAb Ab-4 and detecting antibodiesbound to cell surface by flow cytometry. Similarly, serial dilutions oftwenty-two mouse serum samples were tested for their binding toN202.1ArNeu1 cells. All samples were run in duplicate. Mean channelfluorescence, subtracted of the value obtained with the preimmune se-rum, was recorded. Concentration of anti-rat p185neu serum antibodieswas obtained through linear regression analysis of serially dilutedsamples and expressed as Ab-4 equivalents. Data reported in displayitems are average values from two independent assays. (c) Cell-medi-ated response. Spleen cells from vaccinated neuT mice were collected2 weeks after the second injection and restimulated in vitro with neu-15.3 or neu-42 peptide in an IFN-g ELISPOT assay.
577ADENOVIRUS VACCINATION AGAINST neu ONCOGENE

second group of mice was similarly injected with saline solutionas negative control. Two weeks after the second injection, the ani-mals were sacrificed and the frequency of IFN-g secreting T cellsin mouse spleen cells evaluated by ELISPOT assay. Three hun-dred and eleven peptides, each 15 amino acids long, overlappingby 11 residues and spanning the entire rat p185neu sequence, werecombined into 11 pools. Each of these pools was tested for its abil-ity to stimulate IFN-g secretion from spleen T cells. Peptide poolswere identified, which showed a statistically significant IFN-g pro-duction from mice immunized with Ad5-neu.opt when comparedwith control in the absence of peptide. Further analysis of thesepositive pools identified 4 single peptides responsible for thesereactivities. Overlapping peptides neu-15 and neu-16, whosesequences map in the extra-cellular domain of the rat p185neu,exhibited high and comparable reactivity (Fig. 2). The 9-aminoacid long neu-15.3 peptide comprised within the residues commonto neu-15 and neu-16 peptides displayed a slightly increased reac-tivity. A lower reactivity was exhibited by peptides neu-209 andneu-231 mapping in the intracellular moiety of the receptor. Theseresults were confirmed by intracellular staining (ICS) analysis ofIFN-g secreting T cells, which typed all these peptides as epitopesable to activate CD81 cells (Fig. 2). neu-15.3 also proved the mostreactive peptide and was used to monitor the efficiency of theCD81 response throughout this study. Low reactivity was scoredby the CD41 epitopes neu-41 and neu-42, previously identified.13
Immunization efficiency of ‘‘codon-optimized’’ neu sequence
We compared the ability of neu.wt and neu.opt genes to inducean immune response. Three groups of female neuT mice wereinjected intramuscularly (i.m.) at 6 and 8 weeks of age with eithersaline solution or plasmid DNA expressing neu.wt or neu.opt fol-lowed by ES. Humoral and cell-mediated responses were mea-sured at week 10 and 12, respectively. Anti-rat p185neu serum anti-bodies were measured by flow cytometry using N202rneu1 cells astarget antigen.15 This cell line constitutively expresses a highnumber of rat p185neu membrane receptors.
Mice immunized with pVIJ-neu.opt produced comparable lev-els of anti-rat p185neu antibodies than littermates injected with
pVIJ-neu.wt (Fig. 1b). Titers ranged from 13.0 to 16.1 lg/ml ofAb4 equivalents. By contrast, immunization with pVIJ-neu.optinduced a rat p185neu specific cell mediated immunity about 6-foldhigher than that elicited by pVIJ-neu.wt immunization (Fig. 1c).
Ad5-neu.opt vaccination of neuT mice at week 6 and 8 outper-formed plasmid DNA injection with ES. Ad5-neu.opt induced anti-rat p185neu antibody titers 5-fold and CMI levels, measured by IFN-g ELISPOT, up to 4-fold higher than those elicited by DNA1ES(Fig. 1c).
As a whole, these results demonstrate that the neu.opt gene ismore efficient in eliciting a cellular immune response than the natu-ral sequence. Furthermore, Ad induces an antigen-specific cell-mediated and humoral response much higher than those obtainedby plasmid DNA immunization with ES.
Ad-DNA mixed modality immunizations
The results obtained prompted us to assess the efficacy of a mixedmodality of immunization where mice are immunized with both Adand plasmid DNA. Four groups of neuT mice were injected i.m. at6, 8, 10 and 12 weeks of age with 100 lg of pVIJ-neu.opt with ESand 109 vp of Ad5-neu.opt, alone or in combination, as detailed inFigure 3a. Saline solution was injected as control. At 14 weeks ofage (2 weeks after the last immunization), IFN-g-ICS on spleencells confirmed that Ad5-neu.opt induced a strong CD81 response,much higher than that induced by DNA1ES immunization, asmeasured by the frequency of CD81 T cells producing IFN-g (p <0.04, Fig. 3b). Mixed modality in which Ad either preceded or fol-lowed DNA1ES immunization did not increase the intensity of theresponse as compared to Ad alone. No CD41 reactivity could bemeasured in any group. Eight weeks later (week 22), all groupsshowed very low frequency of IFN-g1, CD81 T cells.
ICS analysis on PBMC from the same mice confirmed the dataobtained from spleen cells (Fig. 3c). Two weeks after the last immu-nization, Ad alone or in combination with DNA1ES induced a50-fold higher frequency of CD81, IFN-g1 PBMC than those de-tected in mice immunized with DNA1ES (p < 0.01). No CD41 re-activity could be measured in any group. Eight weeks later the fre-quency of IFN-g1, CD81 cells were partially reduced in all groups.
FIGURE 2 – Immunodominant T-cell epitopes in neuT mice. neuT mice were immunized at 6 and 8 weeks of age by injection of 10exp9 viralparticles (vp) of Ad5-neu.opt in the quadriceps. At 10 weeks of age mice were sacrificed and the number of IFN-g-secreting anti-neu T cells wasdetermined by ELISPOT on spleen cells from groups of mice using single peptides. The frequency of IFN-g-producing spleen cells was calcu-lated as the average value of spots derived from duplicates at two different cell concentrations. Values are expressed as the number of spot form-ing colonies (SFC)/106 total spleen cells, subtracted of the background values determined in the absence of peptides (typically less than 10 SFC/106 total spleen cells). Data displayed are representative of several independent experiments. Values significantly different (p < 0.05 at least)from background and higher than the arbitrarily chosen threshold of 25 SFC/10exp6 spleen cells are indicated in boldface. Frequency of CD81
T-cell secreting IFN-g was measured by ICS. Data displayed are representative of several independent experiments. At least 50,000 CD31 lym-phocytes were collected by simultaneously gating on CD31 events and small lymphocytes. Values higher than 1& were considered positive andindicated in boldface. On the left side of the display items the sequences encompassed by the peptide used in the assay are indicated. Numbersrefers to the position of amino acid residue of the rat p185neu protein.
578 GALLO ET AL.
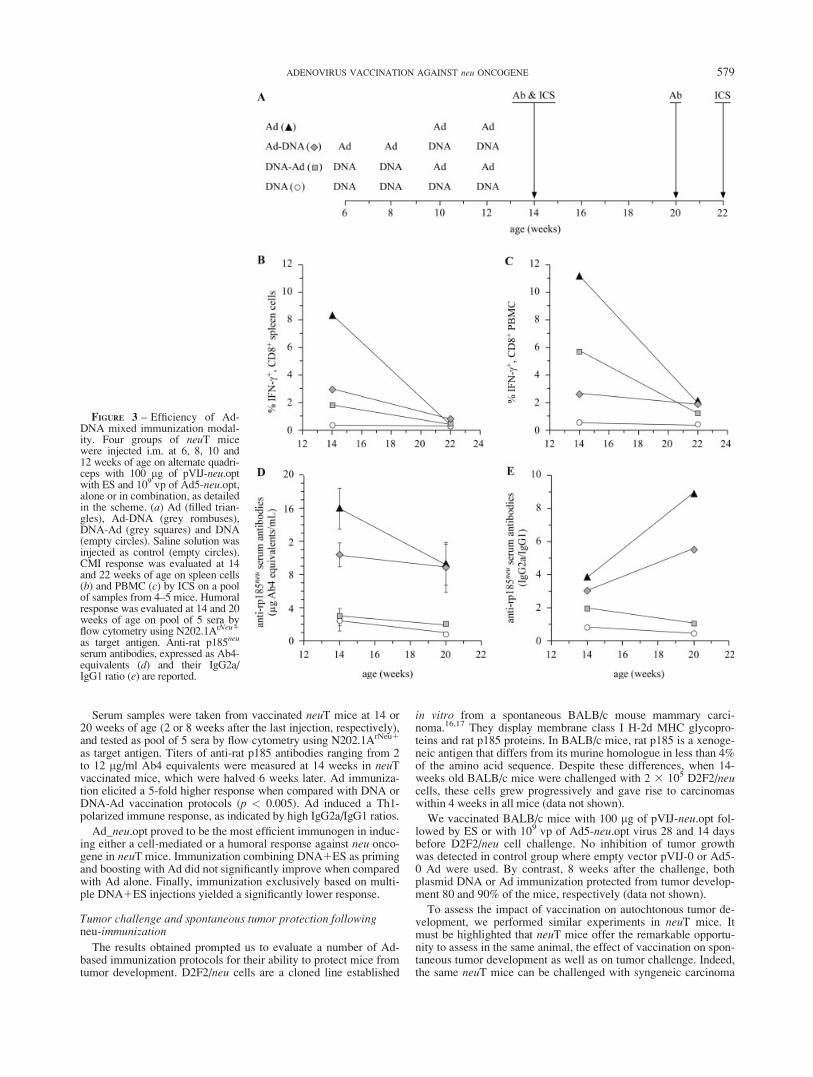
Serum samples were taken from vaccinated neuT mice at 14 or20 weeks of age (2 or 8 weeks after the last injection, respectively),and tested as pool of 5 sera by flow cytometry using N202.1ArNeu1
as target antigen. Titers of anti-rat p185 antibodies ranging from 2to 12 lg/ml Ab4 equivalents were measured at 14 weeks in neuTvaccinated mice, which were halved 6 weeks later. Ad immuniza-tion elicited a 5-fold higher response when compared with DNA orDNA-Ad vaccination protocols (p < 0.005). Ad induced a Th1-polarized immune response, as indicated by high IgG2a/IgG1 ratios.
Ad_neu.opt proved to be the most efficient immunogen in induc-ing either a cell-mediated or a humoral response against neu onco-gene in neuT mice. Immunization combining DNA1ES as primingand boosting with Ad did not significantly improve when comparedwith Ad alone. Finally, immunization exclusively based on multi-ple DNA1ES injections yielded a significantly lower response.
Tumor challenge and spontaneous tumor protection followingneu-immunization
The results obtained prompted us to evaluate a number of Ad-based immunization protocols for their ability to protect mice fromtumor development. D2F2/neu cells are a cloned line established
in vitro from a spontaneous BALB/c mouse mammary carci-noma.16,17 They display membrane class I H-2d MHC glycopro-teins and rat p185 proteins. In BALB/c mice, rat p185 is a xenoge-neic antigen that differs from its murine homologue in less than 4%of the amino acid sequence. Despite these differences, when 14-weeks old BALB/c mice were challenged with 2 3 105 D2F2/neucells, these cells grew progressively and gave rise to carcinomaswithin 4 weeks in all mice (data not shown).
We vaccinated BALB/c mice with 100 lg of pVIJ-neu.opt fol-lowed by ES or with 109 vp of Ad5-neu.opt virus 28 and 14 daysbefore D2F2/neu cell challenge. No inhibition of tumor growthwas detected in control group where empty vector pVIJ-0 or Ad5-0 Ad were used. By contrast, 8 weeks after the challenge, bothplasmid DNA or Ad immunization protected from tumor develop-ment 80 and 90% of the mice, respectively (data not shown).
To assess the impact of vaccination on autochtonous tumor de-velopment, we performed similar experiments in neuT mice. Itmust be highlighted that neuT mice offer the remarkable opportu-nity to assess in the same animal, the effect of vaccination on spon-taneous tumor development as well as on tumor challenge. Indeed,the same neuT mice can be challenged with syngeneic carcinoma
FIGURE 3 – Efficiency of Ad-DNA mixed immunization modal-ity. Four groups of neuT micewere injected i.m. at 6, 8, 10 and12 weeks of age on alternate quadri-ceps with 100 lg of pVIJ-neu.optwith ES and 109 vp of Ad5-neu.opt,alone or in combination, as detailedin the scheme. (a) Ad (filled trian-gles), Ad-DNA (grey rombuses),DNA-Ad (grey squares) and DNA(empty circles). Saline solution wasinjected as control (empty circles).CMI response was evaluated at 14and 22 weeks of age on spleen cells(b) and PBMC (c) by ICS on a poolof samples from 4–5 mice. Humoralresponse was evaluated at 14 and 20weeks of age on pool of 5 sera byflow cytometry using N202.1ArNeu1
as target antigen. Anti-rat p185neu
serum antibodies, expressed as Ab4-equivalents (d) and their IgG2a/IgG1 ratio (e) are reported.
579ADENOVIRUS VACCINATION AGAINST neu ONCOGENE

cells expressing the rat p185neu antigen. In female neuT mice rat,p185neu is a self-protein, which is expressed as early as 3 weeks ofage in the mammary gland duct. Female neuT mice were injectedat 8 and 11 weeks of age with 100 lg of pVIJ-neu.opt with ES or
with 109 vp of Ad5-neu.opt. Two weeks after the last immuniza-tion, mice were injected subcutaneously with 2 3 105 D2F2/neucells. Mice were monitored weekly for the growth of transplantedtumors and for the development of spontaneous tumors in mam-mary glands. Control groups were immunized but not challengedwith tumor. Plasmid DNA immunization failed to protect micefrom tumor challenge, whereas Ad exerted a weak but statisticallysignificant protection (20% tumor-free mice; 8 weeks after chal-lenge, Fig. 4a).
When the development of spontaneous mammary tumors wasassessed, Ad5-neu.opt vaccination protected most of the mice(Fig. 4b). At 23 weeks of age, mice were reported to have a meantumor number of 0.3 and 0 in the absence or in the presence of tu-mor challenge 10 weeks before, respectively. By contrast,DNA1ES only weakly affected tumorigenesis. At the same timepoint, mice were found to have a mean tumor number of 4.1 and 5in mice, which did or did not experience tumor challenge.
In conclusion, the tumor protection data showed that neu-tar-geted Ad immunization is the most effective vaccination protocolamong these tested.
Longevity of the immune response
We sought to characterize the extent and the duration of theprotective effect of Ad-based anti-rat p185 vaccination, and assessthe effect of Ad and DNA injection boost. Three different immuni-zation schedules were evaluated (Fig. 5a). A first group of neuTmice was injected i.m. at 10 and 12 weeks of age with 109 vp ofAd5-neu.opt (Group A). A subset of these mice received an addi-tional injection of 1010 vp of Ad5-neu.opt at 19 weeks of age(Group B). A fraction of these mice was further administered 3plasmid DNA injections with ES at weeks 23, 27 and 31 (GroupC). Saline solution was injected as control in a fourth group (nega-tive control). Humoral and cell-mediated immune response weremonitored throughout the experiment. Development of tumors inmammary glands was monitored weekly.
Vaccinating neuT mice at 10 and 12 weeks of age with 109 vp ofAd5-neu.opt induced a high and sustained anti-rat p185neu Abresponse, which declined with time, in a manner identical to theexperiment reported in Figure 3d. Similarly, we detected a robustincrease of the frequency of IFN-g secreting CD81 PBMC imme-diately after Ad injection (Fig. 5b). This response also declinedwith time and slowly returned to basal levels within 10 weeks afterthe second injection. An additional injection of 1010 vp of Ad5-neu.opt at week 19 did not exert any effect. neuT mice from groupA (vaccinated at week 10 and 12 with Ad5-neu) were analyzed forneutralizing antibodies 2 weeks after the second injection (Table I).Serum from immunized mice effectively neutralized Ad5 infection.Anti-Ad5 human serum was used as positive control. This findingaccounts for the lack of efficiency of a third virus injection. Nota-bly, a repeated DNA1ES immunization at later times significantlyincreased the frequency of CD81, IFN-g secreting PBMC whencompared with the other groups.
All control mice developed at least 1 tumor between weeks 15and 19 (Figs. 5c and 5d). Coherently, with previous experiments,vaccination with Ad5-neu.opt at week 10 and 12 effectively inhib-ited carcinogenesis. This treatment exerted its protective effect forextended period up to a year. Thirty-two weeks after the secondinjection (at 44 weeks of age), 40% of mice were still tumor-free.The protective effect can be better evaluated through the analysisof mean tumor number. By week 26, mice (the control group) pre-sented an evident palpable mass in all 10 mammary glands. Micefrom Group A did not develop any tumor before week 20, andshowed only 3.8 tumors/mouse at week 44.
A third injection of 1010 vp of Ad5-neu.opt did not significantlyalter this effect, protecting 43% of mice at week 45. Similarly,additional immunization with plasmid DNA did not significantlyaffect this result. The same observations were done by measuringthe mean tumor number in the same groups of mice. Group B micewere given a third Ad5-neu.opt injection, which displayed a mean
FIGURE 4 – Effect of vaccination on tumor challenge and spontane-ous tumorigenesis in neuT mice. Scheme of the experiment (a). Fourgroups of mice were immunized at week 8 and 11 with an injection of100 lg plasmid DNA with ES or 10exp9 vp of adenovirus: pVIJ-0(empty circles, 6 mice), pVIJ-neu.opt (filled circles, 15 mice), Ad5-wt(empty triangles, 11 mice) or Ad5-neu.opt (filled triangles, 17 mice).Two weeks later these mice were challenged with a subcutaneousinjection of D2F2/neu cells and monitored weekly for growth of the tu-mor mass (b). In addition to the 4 groups described above, 2 additionalgroups of mice similarly immunized at week 8 and 11, but not chal-lenged with tumor cells, were also included: pVIJ-neu.opt w/ challenge(grey filled circles, 7 mice) and Ad5-neu.opt w/ challenge (grey-filledtriangles, 9 mice). A control group receiving saline solutions was alsoadded (empty rhombuses, 9 mice). Spontaneous mammary carcinomadevelopment was monitored weekly on all the groups of mice (c).
580 GALLO ET AL.

tumor number of 4.8 at week 45, irrespective of plasmid DNAvaccination.
Histopathological examination of neuT mice immunizedwith Ad5-neu.opt
neuT mice were immunized at 10 and 12 weeks of age with 109
vp of Ad5-neu_opt or saline solution (control). Histological andimmunohistochemical analysis were performed on treated mice at
10 and 52 weeks, and at week 10, 16 and 24 on untreated controlgroups. Three mice for each time point underwent histopathologi-cal examination and analysis of PCNA and rat p185neu expressions(Fig. 6).
At 10 weeks of age, mammary glands of neuT mice displayedduct hyperplasia (Stage I) defined by the presence of an increasednumber of epithelial layers (usually 3 or more) above the ductbasement membrane, increased cellular basophilia, a mild increasein the nuclear–cytoplasmic ratio and in the stratification of cells(Fig. 6, upper panel). At 16 weeks, this stage has progressed intoatypical hyperplasia (Stage II), characterized by moderate degreesof epithelial proliferation with tufts, micropapillae and bridgesassociated with increasing atypia and basophilia (Fig. 6, upperPanel B). Eight weeks later, we detected the presence of acinilined by a single layer of cells sometimes showing secretory activity(Stage III). This stage is termed M-Adenocarcima solid differentia-tion, termed Type A according to Dunn classification27 and pro-gresses to Type B where uniform tumor cells resembling lymphoidcells, but arranged in nests and cords, are detected. Tumor cells inboth stages of differentiation are characterized by an increased nu-cleus/cytoplasm ratio and prominent nucleolus, probably due to mi-tosis. Stroma within the tumor is usually scanty and areas of necro-sis and hemorrhages may occur. At week 24, mice developedtumors in all 10 mammary glands and were sacrificed.
In treated mice, vaccination blocks mammary glands tumor de-velopment at the stage of atypical hyperplasia (Stage I). Thesemice express lower levels of PCNA and rat p185neu when com-pared with nonimmunized mice. Tumors that develop late in sometreated mice resemble those developed earlier in untreated mice.
Discussion
We generated a synthetic rat neu gene incorporating exclusivelycodons preferentially used by highly expressed human genes. Theresulting neu.opt gene has G1C content higher than that of neu.wt(from 57.6% to 66.7%). At the same time, the presence of A1T inthe third position drops from 34.4% to 7.2%. As G1C content,neu.wt sequence ranks among the highly expressed human genes(52.4% and 57% in average and highly expressed human genes,respectively). Conversely, A1T frequency in third position dimin-ishes far below average and highly expressed human genes (42 and23%, respectively).8,23 Experimental data show that the neu.optsequence expresses about 3-fold higher amounts of rat p185neu pro-tein than the neu.wt gene in transiently transfected cells. Interest-ingly, the enhancement of protein expression was observed both inhuman and mouse cells. When neuT mice were immunized with
FIGURE 5 – Longevity of immune response and long-term protec-tion in vaccinated neuT mice. (a) Three groups of mice were injectedi.m. at 10 and 12 weeks of age on alternate quadriceps with 109 vp ofAd5-neu.opt as detailed in Panel A. Group A (empty triangles) did notreceive further treatment. Group B (grey-filled triangles) received athird i.m. injection of 1010 vp Ad5-neu at week 19. Group C (black-filled triangles), in addition to the 3 virus injections, was injected withof 100 lg of pVIJ-neu.opt plasmid DNA with ES at week 23, 27 and31. Saline solution was injected as control in a fourth group (emptycircles). Cell mediated immune response (b). Frequency of PBMCsecreting IFN-g was measured by ICS at 14, 20, 21, 22, 25 33 and 37weeks of age. Pool of samples from 5 mice were analyzed and fre-quency expressed as percentage of the total PBMC, subtracted of theof the background value determined in the absence of peptide At least50,000 CD31 lymphocytes were collected by simultaneously gatingon CD31 events and small lymphocytes. Values higher than 1&(dotted line) were considered positive. For each group of mice, valuesat each time point refer to the average frequency observed in 3 to 5different pools of mice. Bars indicate S.E.M. * indicates p < 0.20 andp < 0.05 as compared to group A and B; ** indicates p < 0.19 andp < 0.17; *** indicates p < 0.12 and p < 0.13 as compared to groupA and B, respectively. Spontaneous mammary carcinoma develop-ment was monitored weekly on all the groups of mice and reported aspercentage of tumor-free mice (c) and mean tumor number (d).
581ADENOVIRUS VACCINATION AGAINST neu ONCOGENE

plasmid DNA1ES, the neu.opt gene showed a higher CD81
response than neu.wt, whereas antibody response was unchanged. Asimilar comparison was not possible for Ad5-neu.opt since we wereunable to rescue the Ad5-neu.wt virus. Synthetic genes designed toincorporate the codon usage of highly expressed human genes havebeen reported to express higher levels of the corresponding proteinsthan the wt sequence. Results described in this article, along withdata obtained in our laboratory on several different tumor-associatedself-antigens, coherently indicate that codon optimization is a usefultool for the development of antitumor genetic vaccines withincreased efficacy29 (unpublished data).
In this work, we explored the immunogenic and therapeutic effi-ciency of genetic vaccination against rat p185 protein. We used thefull length neu cDNA to evaluate the impact of sequence optimiza-tion on immunogenicity and on the ability to protect from tumordevelopment. This choice was supported by the identification ofCD81 epitopes in the intracellular moiety. We tested the immuni-zation efficiency of 2 genetic immunization protocols: DNA1ESand Ad injection. Ad-mediated immunization proved by far to be
superior eliciting both higher humoral and cell-mediated IR, polar-ized towards a Th-1 type. We tested mixed modalities of vaccina-tion using both Ad and DNA1ES, in either order. Priming withDNA followed by Ad boosting did not improve immune responsewhen compared with Ad alone. The same applied to the comple-mentary situation, at least in the experimental conditions tested. Ina coherent fashion, Ad immunization showed a significant impacton tumor progression in vaccinated mice whereas mixed modalitiesachieved only a partial protection. Remarkably, 2 injections at 10and 12 weeks of age provided protection against spontaneous tu-mor development up to 33 weeks after the last boost. By contrast,repeated injections of DNA1ES are required to achieve an effec-tive tumor protection for a comparable period of time.30 Althoughstrong, Ad5-mediated response declined with time indicating thatadditional immunizations are required to ensure a life-long immu-nity. However, Ad5 readministration lacked efficacy because ofneutralizing antibodies raised by previous injections. The sameantibodies, however, did not block Ad6 or Ad24 infection. Theseobservations indicate towards protocols employing different adeno-viral vectors, which do not elicit crossreacting antibody response.Alternatively, a response elicited by Ad5 can be effectively sus-tained by repeated DNA injections, thanks to a lack of responseagainst the DNA vehicle. In fact, though DNA injection per se ispoorly immunogenic, it could sustain response induced by Ad5.31
We predict that a mixed modality immunization, where the re-sponse induced by one or multiple Ad vectors is accompanied by aseries of plasmid DNA injections with adjuvant cytokines is likelyto be an effective vaccination protocol.32,33 This view is supportedby the results described in this work. Ad immunization induces a
TABLE I – TITER OF ADENOVIRUS NEUTRALIZING ANTIBODIES IN THESERUM OF 14-WEEKS OLD neuT MICE INJECTED i.m. AT 10 AND
12 WEEKS OF AGE WITH 109 vp OF AD5-neu
Serum Ad5 Ad6 Ad24
neuT mice 2,300 <10 <10Positive control 2,700 n.t. n.t.
Titers refer to the serum dilution neutralizing 50%.n.t.5 not tested.
FIGURE 6 – Impact of vaccina-tion on tumorigenesis in neuTmice. Mice were immunized at 10and 12 weeks of age with 109 vp ofAd5-neu_opt or saline solution(control). Histological and immu-nohistochemical analysis was per-formed on treated mice at 10, 16and 52 weeks and at week 10, 16and 24 on untreated control groups.Three mice for each time pointwere subjected to histopathologicalexamination and analysis of PCNAand rat p185neu markers. Data dis-played are representative of severalfields analyzed. Stages of tumordevelopment are defined accordingto Dunn (Duct hyperplasia I, Atyp-ical hyperplasia II, M-Adenocar-cima adenoid differentiation III).
582 GALLO ET AL.

strong cell-mediated response, which declined in about 10 weeks.We observed that, at this stage, repeated DNA1ES immunization iscapable of ‘‘reactivating’’ the cell-mediated response. This ‘‘re-lapse’’ does not impact on tumorigenesis, and is in concordance withthe widely reported observation that the tumor is poorly susceptibleto the immune response once it has progressed to carcinoma. How-ever, these data indicate that a protocol where Ad vaccination isimmediately followed by multiple DNA1ES immunization shouldmaintain a high immune response for longer times. It must bestressed, however, that the ranking of the various immunization re-gimen depends on the antigen used. For example, in the case of car-cinoembryonic antigen, a prime-boost regimen using DNA1ESand Ad elicited the highest amplitude of immune response.29
We comparatively evaluated the effect of DNA injection withES and that induced by Ad injection on development of transplant-able and spontaneous tumors. The same neuT mice, which sponta-neously develop mammary gland spontaneous tumors, were chal-lenged with a subcutaneous injection of a syngeneic tumor cell line.In tumor challenge experiments, only Ad injection exerted a weakbut significant protection, whereas DNA injection with ES did notaffect tumor growth. By contrast, when the same mice were moni-tored for spontaneous tumor development, Ad injection completelyprotected mice. A weaker but significant protection was also dis-played by DNA immunization. Thus, the 2 tumor model systemsdeliver information, which differ remarkably at least from thequantitative point of view. This is not surprising. Tumor challenge
experiments are more widely adopted in vivo system to assess theantitumor activity of new vaccination treatments. However, the tu-mor location and its environment do not reproduce the microenvir-onment that resembles or mimics the early stages of the tumorigen-esis process when preneoplastic lesions transform into carcinoma.In neuT mice, a tumor progresses through progressive stages of tu-morigenesis and the relationship between the tumor and the sur-roundings are preserved.12,34 It is conceivable that the tumor chal-lenge experiments fail to reveal the effective inhibitory potential ofthe immune response elicited by a vaccine. The effect of the samevaccination is much higher in the case of spontaneous tumor inhibi-tion. On the other hand, it should be recalled that transgene expres-sion in neuT mice is artificially regulated by the MMTV promoterduring development. This regulation might generate an expressionpattern that differs from the one of the natural gene.
Recently, Ad expressing the neu extracellular and transmem-brane moiety was tested in neuT mice.35 In this report, completeprotection against spontaneous mammary tumor was obtained by4 intraperitoneal (i.p.) injections of 108 pfu 4 times, every 3–4weeks, beginning at 7 weeks of age and ending on week 19. Theexperimental differences (antigen sequence, route of administra-tion, dose and number of injections) can explain the differentresults obtained. When vaccination started at 11 weeks of age, atiming that approaches a therapeutic scenario, similar protectionwas achieved. Our data indicate that 2 i.m. injections of 109 vpyield a response comparable to 4 i.p. administration of 108 pfu.
References
1. Simons JW, Jaffee EM, Weber CE, Levitsky HI, Nelson WG,Carducci MA, Lazenby AJ, Cohen LK, Finn CC, Clift SM, Hauda KM,Beck LA, et al. Bioactivity of autologous irradiated renal cell carcinomavaccines generated by ex vivo granulocyte-macrophage colony-stimu-lating factor gene transfer. Cancer Res 1997;57:1537–46.
2. Dranoff G. GM-CSF-secreting melanoma vaccines. Oncogene 2003;22:3188–92.
3. Salgia R, Lynch T, Skarin A, Lucca J, Lynch C, Jung K, Hodi FS,Jaklitsch M, Mentzer S, Swanson S, Lukanich J, Bueno R, et al. Vac-cination with irradiated autologous tumor cells engineered to secretegranulocyte-macrophage colony-stimulating factor augments antitu-mor immunity in some patients with metastatic non-small-cell lungcarcinoma. J Clin Oncol 2003;21:624–30.
4. Simons JW, Mikhak B, Chang JF, DeMarzo AM, Carducci MA, LimM,Weber CE, Baccala AA, Goemann MA, Clift SM, Ando DG, LevitskyHI, et al. Induction of immunity to prostate cancer antigens: results of aclinical trial of vaccination with irradiated autologous prostate tumorcells engineered to secrete granulocyte-macrophage colony-stimulatingfactor using ex vivo gene transfer. Cancer Res 1999;59:5160–8.
5. Soiffer R, Hodi FS, Haluska F, Jung K, Gillessen S, Singer S, Tanabe K,Duda R, Mentzer S, Jaklitsch M, Bueno R, Clift S, et al. Vaccinationwith irradiated, autologous melanoma cells engineered to secrete granu-locyte-macrophage colony-stimulating factor by adenoviral-mediatedgene transfer augments antitumor immunity in patients with metastaticmelanoma. J Clin Oncol 2003;21:3343–50.
6. Nemunaitis J. Granulocyte-macrophage colony-stimulating factor gene-transfected autologous tumor cell vaccine: focus[correction to fcous] onnon-small-cell lung cancer. Clin Lung Cancer 2003;5:148–57.
7. Nemunaitis J, Cunningham CC, Vukelja S, Ruxer RL, Adams N, Rich D,Paulson AS, MacEachern JB. Phase I trial of topotecan in combinationwith gemcitabine in refractory solid tumor patients. Cancer Invest2004;22:360–7.
8. Lee CT, Wu S, Ciernik IF, Chen H, Nadaf-Rahrov S, Gabrilovich D,Carbone DP. Genetic immunotherapy of established tumors with ade-novirus-murine granulocyte-macrophage colony-stimulating factor.Hum Gene Ther 1997;8:187–93.
9. Rizzuto G, Cappelletti M, Maione D, Savino R, Lazzaro D, Costa P,Mathiesen I, Cortese R, Ciliberto G, Laufer R, La Monica N, Fattori E.Efficient and regulated erythropoietin production by naked DNA injec-tion and muscle electroporation. Proc Natl Acad Sci USA 1999;96:6417–22.
10. Lucchini F, Sacco MG, Hu N, Villa A, Brown J, Cesano L,Mangiarini L, Rindi G, Kindl S, Sessa F, Vezzoni P, Clerici L. Earlyand multifocal tumors in breast, salivary, harderian and epididymaltissues developed in MMTY-Neu transgenic mice. Cancer Lett 1992;64:203–9.
11. Di Carlo E, Diodoro MG, Boggio K, Modesti A, Modesti M, Nanni P,Forni G, Musiani P. Analysis of mammary carcinoma onset and
progression in HER-2/neu oncogene transgenic mice reveals a lob-ular origin. Lab Invest 1999;79:1261–9.
12. Boggio K, Nicoletti G, Di Carlo E, Cavallo F, Landuzzi L, Melani C,Giovarelli M, Rossi I, Nanni P, De Giovanni C, Bouchard P, Wolf S,et al. Interleukin 12-mediated prevention of spontaneous mammaryadenocarcinomas in two lines of Her-2/neu transgenic mice. J Exp Med1998;188:589–96.
13. Gallo P, Dharmapuri S, Nuzzo M, Maldini D, Iezzi M, Cavallo F,Musiani P, Forni G, Monaci P. Xenogeneic immunization in mice usingHER2 DNA delivered by an adenoviral vector. Int J Cancer 2005;113:67–77.
14. Graham FL, Smiley J, Russell WC, Nairn R. Characteristics of ahuman cell line transformed by DNA from human adenovirus type 5.J Gen Virol 1977;36:59–74.
15. Nanni P, Pupa SM, Nicoletti G, De Giovanni C, Landuzzi L, Rossi I,Astolfi A, Ricci C, De Vecchi R, Invernizzi AM, Di Carlo E, MusianiP, et al. p185(neu) protein is required for tumor and anchorage-inde-pendent growth, not for cell proliferation of transgenic mammary carci-noma. Int J Cancer 2000;87:186–94.
16. Mahoney KH, Miller BE, Heppner GH. FACS quantitation of leucineaminopeptidase and acid phosphatase on tumor-associated macro-phages from metastatic and nonmetastatic mouse mammary tumors.J Leukoc Biol 1985;38:573–85.
17. Piechocki MP, Pilon SA, Wei WZ. Complementary antitumor immu-nity induced by plasmid DNA encoding secreted and cytoplasmichuman ErbB-2. J Immunol 2001;167:3367–74.
18. Rovero S, Amici A, Carlo ED, Bei R, Nanni P, Quaglino E, Porcedda P,Boggio K, Smorlesi A, Lollini PL, Landuzzi L, Colombo MP, et al.DNA vaccination against rat her-2/Neu p185 more effectively inhibitscarcinogenesis than transplantable carcinomas in transgenic BALB/cmice. J Immunol 2000;165:5133–42.
19. Montgomery DL, Shiver JW, Leander KR, Perry HC, Friedman A,Martinez D, Ulmer JB, Donnelly JJ, Liu MA. Heterologous and ho-mologous protection against influenza A by DNA vaccination: optimi-zation of DNA vectors. DNA Cell Biol 1993;12:777–83.
20. Ishibashi M, Maizel JV, Jr. The polypeptides of adenovirus. V. Youngvirions, structural intermediate between top components and aged vi-rions. Virology 1974;57:409–24.
21. Miyahira Y, Murata K, Rodriguez D, Rodriguez JR, Esteban M,Rodrigues MM, Zavala F. Quantification of antigen specific CD81T cells using an ELISPOT assay. J Immunol Methods 1995;181:45–54.
22. Aste-Amezaga M, Bett AJ, Wang F, Casimiro DR, Antonello JM,Patel DK, Dell EC, Franlin LL, Dougherty NM, Bennett PS, Perry HC,Davies ME, et al. Quantitative adenovirus neutralization assays basedon the secreted alkaline phosphatase reporter gene: application in epi-demiologic studies and in the design of adenovector vaccines. HumGene Ther 2004;15:293–304.
583ADENOVIRUS VACCINATION AGAINST neu ONCOGENE

23. Nakamura Y, Gojobori T, Ikemura T. Codon usage tabulated from in-ternational DNA sequence databases: status for the year 2000. NucleicAcids Res 2000;28:292.
24. Ben-Levy R, Paterson HF, Marshall CJ, Yarden Y. A single autophos-phorylation site confers oncogenicity to the Neu/ErbB- 2 receptor andenables coupling to the MAP kinase pathway. EMBO J 1994;13:3302–11.
25. Kozak M. An analysis of 50-noncoding sequences from 699 vertebratemessenger RNAs. Nucleic Acids Res 1987;15:8125–48.
26. Kozak M. At least six nucleotides preceding the AUG initiator codonenhance translation in mammalian cells. J Mol Biol 1987;196:947–50.
27. Dunn T. Morphology of mammary tumors in mice. In: Homburger F, ed.The physiopathology of cancer. New York: Hoeber-Harper, 1959. 38–84.
28. Haas J, Park EC, Seed B. Codon usage limitation in the expression ofHIV-1 envelope glycoprotein. Curr Biol 1996;6:315–24.
29. Mennuni C, Calvaruso F, Facciabene A, Aurisicchio L, Storto M,Scarselli E, Ciliberto G, La Monica N. Efficient induction of T-cell re-sponse to carcinoembryonic antigen by a heterologous prime-boostregimen using DNA and adenovirus vectors carrying a codon usageoptimized cDNA. Int J Cancer 2005;117:444–55.
30. Quaglino E, Iezzi M, Mastini C, Amici A, Pericle F, Di Carlo E, Pupa SM,De Giovanni C, Spadaro M, Curcio C, Lollini PL, Musiani P, et al.Electroporated DNA vaccine clears away multifocal mammary carci-nomas in her-2/neu transgenic mice. Cancer Res 2004;64:2858–64.
31. MacGregor RR, Boyer JD, Ugen KE, Lacy KE, Gluckman SJ,Bagarazzi ML, Chattergoon MA, Baine Y, Higgins TJ, Ciccarelli RB,Coney LR, Ginsberg RS, et al. First human trial of a DNA-based vac-cine for treatment of human immunodeficiency virus type 1 infection:safety and host response. J Infect Dis 1998;178:92–100.
32. Casimiro DR, Chen L, Fu TM, Evans RK, Caulfield MJ, Davies ME,Tang A, Chen M, Huang L, Harris V, Freed DC, Wilson KA, et al.Comparative immunogenicity in rhesus monkeys of DNA plasmid,recombinant vaccinia virus, and replication-defective adenovirus vec-tors expressing a human immunodeficiency virus type 1 gag gene.J Virol 2003;77:6305–13.
33. Matsui M, Moriya O, Akatsuka T. Enhanced induction of hepatitis Cvirus-specific cytotoxic T lymphocytes and protective efficacy inmice by DNA vaccination followed by adenovirus boosting in combi-nation with the interleukin-12 expression plasmid. Vaccine 2003;21:1629–39.
34. Boggio K, Di Carlo E, Rovero S, Cavallo F, Quaglino E, Lollini PL,Nanni P, Nicoletti G, Wolf S, Musiani P, Forni G. Ability of systemicinterleukin-12 to hamper progressive stages of mammary carcinogen-esis in HER2/neu transgenic mice. Cancer Res 2000;60:359–64.
35. Park JM, Terabe M, Sakai Y, Munasinghe J, Forni G, Morris JC,Berzofsky JA. Early role of CD41 Th1 cells and antibodies in HER-2adenovirus vaccine protection against autochthonous mammary carci-nomas. J Immunol 2005;174:4228–36.
584 GALLO ET AL.
Copyright © 2022 FDOKUMEN




















