
Tumor progression and oncogene addiction in a PDGF-B-induced model of gliomagenesis
Upload
independentCategory
view
2download
0
Cell, Vol. 55, 185-192, October 7, 1988, Copyright 0 1988 by Cell Press
The Dominant-White Spotting (IA!) Locus of the Mouse Encodes the c-kit Proto-Oncogene
Edwin N. Geissler, Melanie A. Ryan, and David E. Housman Department of Biology and Center for Cancer Research Massachusetts Institute of Technology Cambridge, Massachusetts 02139
Summary
Mutations at the W locus in the mouse have pleiotropic effects on embryonic development and he- matopoiesis. The characteristic phenotype of mutants at this locus, which includes white coat color, sterility, and anemia, can be attributed to the failure of stem cell populations to migrate and/or proliferate effec- tively during development. Mapping experiments sug- gest that the c-kit proto-oncogene, which encodes a putative tyrosine kinase receptor, is a candidate for the W locus. We show here that the c-kit gene is dis- rupted in two spontaneous mutant W alleles, W44 and Wx. Genomic DNA that encodes amino acids 240 to 342 of the c-kit polypeptide is disrupted in W44; the region encoding amino acids 342 to 791 is disrupted in Wx. We4 homozygotes exhibit a marked reduction in levels of c-kit mRNA. These results strongly sup- port the identification of c-kit as the gene product of the W locus.
Introduction
Mutations at the dominant white (w) locus of the labora- tory mouse have pleiotropic effects upon both embryonic development and the regulation of hematopoiesis in adult life. Most mutations produce homozygotes that are sterile, have extensive white-spotting, and a severe anemia that results in perinatal death (Russell, 1979; Silvers, 1979; Geissler et al., 1981).
In normal development, melanoblasts and primordial germ cells proliferate as they migrate from lheir origins, fleural crest and yolk sac respectively, to their final desti- nations (Rawles, 1947; Chiquoine, 1954). The defect in W mutants appears to be intrinsic to the stem cells of the af- fected tissues, interfering with their proliferation and sub- sequent survival during embryogenesis (Mayer and Green, 1968; Silvers, 1979; Mintz and Russell, 1957). Al- though it has not been clearly established, there is some indication that development of fetal and adult hematopoie- sis involves the migration of hematopoietic stem cells be- tween subsequent sites of hematopoiesis (Moore and Metcalf, 1970). The hematopoietic defect in W mice is present early in embryogenesis, involving deficiencies of the erythroid derivatives of the blood islands of the em- bryonic yolk sac, and persisting in subsequent hematopoi- etic organs, including fetal liver and the bone marrow and spleen of the adult mouse (Russell et al., 1968). The de- feel is intrinsic to the hematopoietic stem cell. The blood-
forming tissues of anemic W animals contain reduced numbers of stem cells assayed by the spleen colony- forming unit (CFU-S) assay (McCulloch et al., 1964) as well as fewer committed BFU-E and CFU-E erythroid pro- genitor cells (Gregory and Eaves, 1978). The anemia of these mutants is completely cured by the injection of nor- mal blood-forming tissues (Russell et al., ‘1956; Russell and Bernstein, 1968; Russell, 1970). The hematopoietic tissues of W mutants in such transplant experiments are eventually repopulated by erythroid as well as nonery- throid derivatives of the donor marrow, indicating a prolif- erative advantage of wild-type hematopoietic progenitors over the W mutant counterparts and that the W hematopoi- etic environment is capable of supporting normal hemato- poiesis (Murphy et al., 1973; Harrison and Astle, 1976).
Elucidation of the mechanism by which the Wgene con- trols embryological development and the proliferation and differentiation of the pluripotent hematopoietic stem cell would have considerable biological significance. To facili- tate a molecular genetic approach to this problem, we de- veloped a detailed genetic map of the region of mouse chromosome 5 that surrounds the W locus. Recently, we characterized a radiation-induced W mutant, WgH, iso- lated by M. F. Lyon and co-workers, which demonstrates that WgH involves a deletion of DNA sequences closely linked to the W locus (Geissler et al., 1988). The WgH de- letion has allowed us to evaluate rapidly candidate DNA sequences for their genetic proximity to the W locus. The recent mapping of the c-kit oncogene to mouse chromo- some 5 (Yarden et al., 1987) suggested the possibility that c-kit is a candidate for the W structural gene. DNA se- quence analysis indicates that the c-kitpolypeptide shows strong structural similarities to the receptors for colony- stimulating factor 1 (CSF-1) and platelet-derived growth factor (PDGF). The c-kit gene product has been shown to be a cell surface glycoprotein that possesses kinase activ- ity, strongly suggesting that c-kit encodes a transmem- brane receptor for an as yet unidentified tigand (Yarden et al., 1987; Qiu et al., 1988). The functional characteristics of c-kit are also consistent with earlier suggestions by Elizabeth Russell and co-workers that the product of the Wgene is a cell surface receptor involved in cell-cell inter- action during morphogenesis (Russell, ‘1979; Geissler et al., 1981).
Mapping experiments reported here demonstrate that the c-kit structural gene maps within WgH deletion, con- sistent with the hypothesis that the c-kit protein is the gene product of the W locus. To investigate this question further, we carried out a survey of known W mutants to determine whether DNA rearrangements of the c-kit locus are as- sociated with W mutations. We report here the character- ization of two Wmutants that show clear evidence for gene rearrangement within the c-kit locus. The properties of these mutants and the nature of the rearrangements at c-kit strongly support the view that the c-kEt protein is in- deed the gene product of the W locus.
Cell 186
23-
2.3-
1 2 3 4 5 6 7 8 9101112 13 14 15 16 17 18 19 20 21 22 23 24
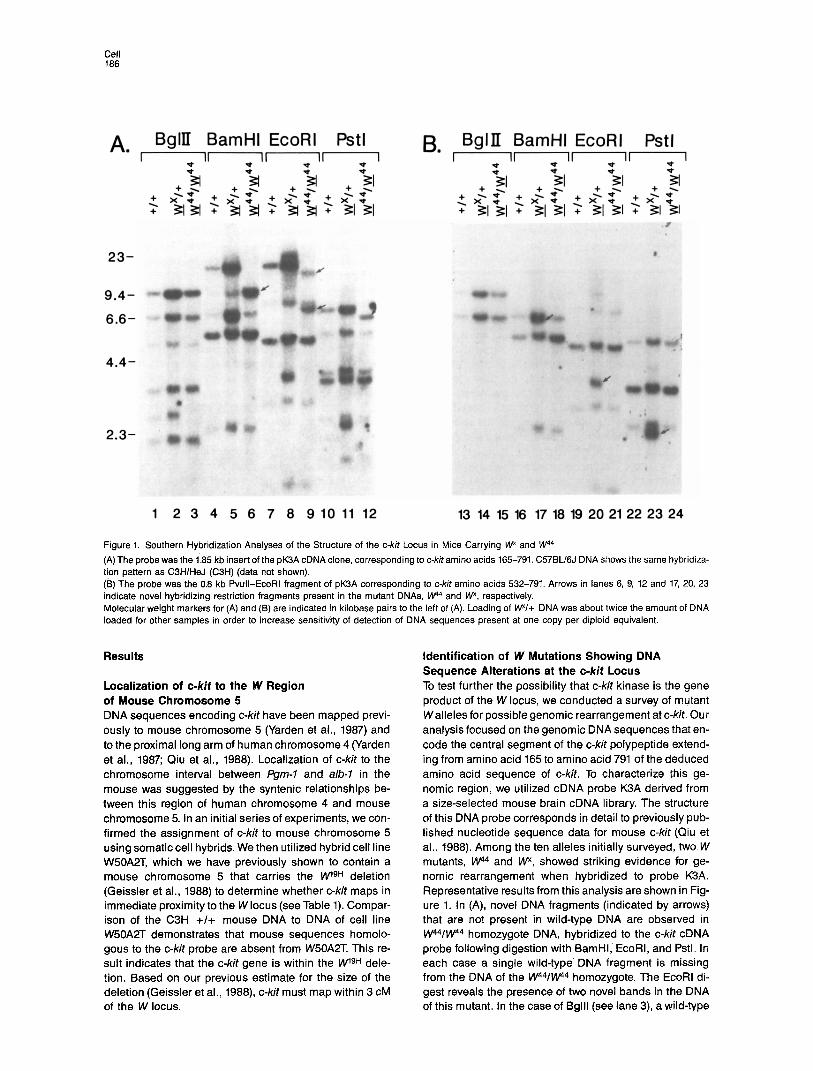
Figure 1. Southern Hybridization Analyses of the Structure of the c-kit Locus in Mice Carrying W and W4
(A) The probe was the 1.85 kb insert of the pK3A cDNA clone, corresponding to c-kit amino acids 165-791. C57BLi6J DNA shows the same hybridiza- tion pattern as C3H/HeJ (C3H) (data not shown). (B) The probe was the 0.8 kb Pvull-EcoRI fragment of pK3A corresponding to c-kit amino acids 532-791. Arrows in lanes 6, 9, 12 and 17, 20, 23 indicate novel hybridizing restriction fragments present in the mutant DNAs, IV4 and W, respectively. Molecular weight markers for (A) and (B) are indicated in kilobase pairs to the left of (A). Loading of WV+ DNA was about twice the amount of DNA loaded for other samples in order to increase sensitivity of detection of DNA sequences present at one copy per diploid equivalent.
Results
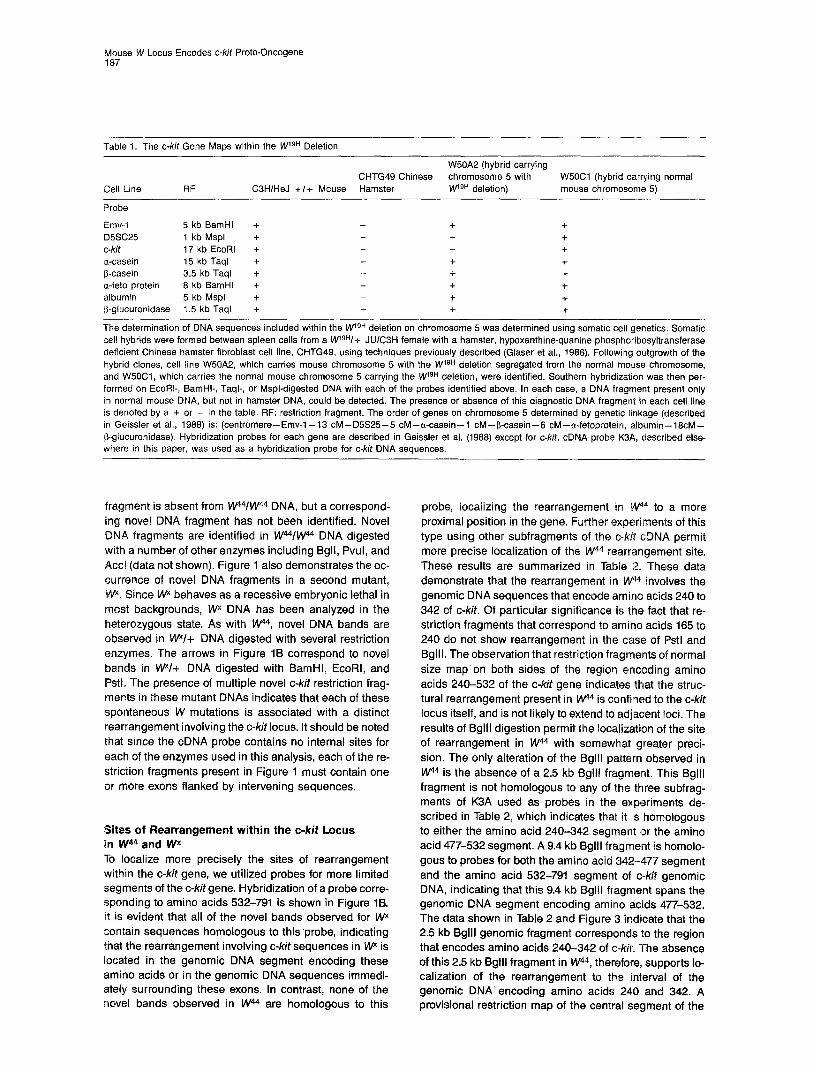
Localization of c-&if to the W Region of Mouse Chromosome 5 DNA sequences encoding c-kit have been mapped previ- ously to mouse chromosome 5 (Yarden et al., 1987) and to the proximal long arm of human chromosome 4 (Yarden et al., 1987; Qiu et al., 1988). Localization of c-kit to the chromosome interval between mm-1 and a/b-l in the mouse was suggested by the syntenic relationships be- tween this region of human chromosome 4 and mouse chromosome 5. In an initial series of experiments, we con- firmed the assignment of c-kit to mouse chromosome 5 using somatic cell hybrids. We then utilized hybrid cell line W5OA2T which we have previously shown to contain a mouse chromosome 5 that carries the WrgH deletion (Geissler et al., 1988) to determine whether c-kit maps in immediate proximity to the W locus (see Table 1). Compar- ison of the C3H +I+ mouse DNA to DNA of cell line W5OA2T demonstrates that mouse sequences homolo- gous to the c-kit probe are absent from W5OA2T. This re- sult indicates that the c-kit gene is within the WrgH dele- tion. Based on our previous estimate for the size of the deletion (Geissler et al., 1988), c-kit must map within 3 CM of the W locus.
Identification of W Mutations Showing DNA Sequence Alterations at the c-kit Locus To test further the possibility that c-kit kinase is the gene product of the W locus, we conducted a survey of mutant Walleles for possible genomic rearrangement at c-kit. Our analysis focused on the genomic DNA sequences that en- code the central segment of the c-kit polypeptide extend- ing from amino acid 165 to amino acid 791 of the deduced amino acid sequence of c-kit. To characterize this ge- nomic region, we utilized cDNA probe K3A derived from a size-selected mouse brain cDNA library. The structure of this DNA probe corresponds in detail to previously pub- lished nucleotide sequence data for mouse c-kit (Qiu et al., 1988). Among the ten alleles initially surveyed, two W mutants, w4 and Wx, showed striking evidence for ge- nomic rearrangement when hybridized to probe K3A. Representative results from this analysis are shown in Fig- ure 1. In (A), novel DNA fragments (indicated by arrows) that are not present in wild-type DNA are observed in W44/W44 homozygote DNA, hybridized to the c-kit cDNA probe following digestion with BamHI, EcoRI, and Pstl. In each case a single wild-type’ DNA fragment is missing from the DNA of the W44/w44 homozygote. The EcoRl di- gest reveals the presence of two novel bands in the DNA of this mutant. In the case of Bglll (see lane 3), a wild-type
Mouse W Locus Encodes c-kit Proto-Oncogene 187
Table I. The c-kit Gene Maps within the WlgH Deletion
Cell Line RF
W50A2 (hybrid carrying CHTG49 Chinese chromosome 5 with W5OCl (hybrid carrying normal
C3HIHeJ +I+ Mouse Hamster W9H deletion) mouse chromosome 5) -
Probe
Emv-I 5 kb BamHl + + + D5SC25 I kb Mspl + + c-kit 17 kb EcoRl + + a-casein 15 kb Taql + + + b-casein 3.5 kb Taql + + + a-feto protein 6 kb BamHl + + + albumin 5 kb Mspl + + + p-glucuronidase 1.5 kb Taql + + +
-- The determination of DNA sequences included within the kVgH deletion on chromosome 5 was determined using somatic cell genetics. Somatic cell hybrids were formed between spleen cells from a wgH/+ JUlCBH female with a hamster, hypoxanthine-quanine phosphoribosyltransferase deficient Chinese hamster fibroblast cell line, CHTG49, using techniques previously described (Glaser et al., 1966). Followilng outgrowth of the hybrid clones, cell line W50A2, which carries mouse chromosome 5 with the W lgH deletion segregated from the normal mouse chromosome, and W5OC1, which carries the normal mouse chromosome 5 carrying the M/I gH deletion, were identified. Southern hybridization was then per- formed on EcoRI-, BamHI-, Taql-, or Mspl-digested DNA with each of the probes identified above. In each case, a DNA fragment present only in normal mouse DNA, but not in hamster DNA, could be detected. The presence or absence of this diagnostic DNA fragment in each cell line is denoted by a + or - in the table. RF: restriction fragment. The order of genes on chromosome 5 determined by genetic linkage (described in Geissler et al., 1966) is: (centromere-Emv-l-13 CM-D5S25-5 CM--a-casein-l CM-P-casein- CM-a-fetoprotein, albumin-16cM- &glucuronidase). Hybridization probes for each gene are described in Geissler et al. (1966) except for c-kit. cDNA probe K3A, described else- where in this paper, was used as a hybridization probe for c-kit DNA sequences.
fragment is absent from w44/W44 DNA, but a correspond- ing novel DNA fragment has not been identified. Novel DNA fragments are identified in W44/w44 DNA digested with a number of other enzymes including Bgll, Pvul, and Accl (data not shown). Figure 1 also demonstrates the oc- currence of novel DNA fragments in a second mutant, \Ny. Since Wx behaves as a recessive embryonic lethal in most backgrounds, Wx DNA has been analyzed in the heterozygous state. As with W4, novel DNA bands are observed in WV+ DNA digested with several restriction enzymes. The arrows in Figure 19 correspond to novel bands in WV+ DNA digested with BamHI, EcoRI, and Pstl. The presence of multiple novel c-kit restriction frag- ments in these mutant DNAs indicates that each of these spontaneous W mutations is associated with a distinct rearrangement involving the c-kit locus. It should be noted that since the cDNA probe contains no internal sites for each of the enzymes used in this analysis, each of the re- striction fragments present in Figure 1 must contain one or more exons flanked by intervening sequences.
Sites of Rearrangement within the c-kit Locus in W44 and W To localize more precisely the sites of rearrangement within the c-kit gene, we utilized probes for more limited segments of the c-kit gene. Hybridization of a probe corre- sponding to amino acids 532-791 is shown in Figure 1B. It is evident that all of the novel bands observed for Wx contain sequences homologous to this.probe, indicating that the rearrangement involving c-kit sequences in w* is located in the genomic DNA segment encoding these amino acids or in the genomic DNA sequences immedi- ately surrounding these exons. In contrast, none of the novel bands observed in W44 are homologous to this
probe, localizing the rearrangement in W44 to a more proximal position in the gene. Further experiments of this type using other subfragments of the c-kit cDNA permit more precise localization of the W44 rearrangement site. These results are summarized in Table 2. These data demonstrate that the rearrangement in Wt4 involves the genomic DNA sequences that encode amino acids 240 to 342 of c-kit. Of particular significance is the fact that re- striction fragments that correspond to amino acids 165 to 240 do not show rearrangement in the case of Pstl and Bglll. The observation that restriction fragments of normal size map on both sides of the region encoding amino acids 240-532 of the c-kit gene indicates that the struc- tural rearrangement present in w44 is confined to the c-kit locus itself, and is not likely to extend to adjacent loci. The results of Bglll digestion permit the localization of the site of rearrangement in W” with somewhat greater preci- sion. The only alteration of the Bglll patteirn observed in W44 is the absence of a 2.5 kb Bglll fragment. This Bglll fragment is not homologous to any of the three subfrag- ments of K3A used as probes in the ex.periments de- scribed in Table 2, which indicates that it is homologous to either the amino acid 240-342 segment Ior the amino acid 477-532 segment. A 9.4 kb Bglll fragment is homolo- gous to probes for both the amino acid 342-477 segment and the amino acid 532-791 segment of c-kit genomic DNA, indicating that this 9.4 kb Bglll fragment spans the genomic DNA segment encoding amina acids 477-532. The data shown in Table 2 and Figure 3 indicate that the 2.5 kb Bglll genamic fragment corresponds to the region that encodes amino acids 240-342 of c-kit. The absence of this 2.5 kb Bglll fragment in W44, therefore, supports lo- calization of the rearrangement to the interval of the genomic DNA’ encoding amino acids 240 and 342. A provisional restriction map of the central segment of the

Cell 188
Table 2. Mapping the Location of Wr4 and W Rearrangements
Amino Acids Probe EcoRl BamHl Pstl Bglll
+I+ 17, 8, 5, 3, 1 17, 9.6, 7, 5.5, 2 7, 5.2, 3.8, 3.5, 2.5, 1.5, 0.8 165-791 A WV+ 17, 8, 5, 3, 1 (3.6) 17, 9.6, 7, 5.5, 2 (6.5) 7, 5.2, 3.8, 3.5, 2.5, 1.5, 0.8 (2.3)
wwv4 6 5, 3, 1 (14, 7) 9.6, 7, 5.5, 2 (10) 5.2, 3.8, 3.5, 2.5, 1.5, 0.8 (6.5)
+I+ 17 17 3.8 165-240 B WV+ 17 17 3.8
!Awkv4 (14) (10) 3.8
i/i 17, 8, 5 17, 9.6 5.5 7 1.5 342-477 c WV+ 17, 8, 5 17, 9.6 5.5 (6.5) 7 1.5
w41ti4 8, 5 (7L. 9.6 5.5 (?) 1.5 (6.5)
9.4, 6.6, 5, 3.3, 2.5, 2 9.4, 6.6, 5, 3.3, 2.5, 2 9.4, 6.6, 5, 3.3 2
3.3 3.3 3.3
9.4 5 2 9.4 5 2 9.4 5 2
+I+ 5, 3, 1 7, 5.5, 2 5.2 3.5, 2.5 9.4, 6.6 532-791 D WI+ 5, 3, 1 (3.6) 7, 5.5, 2 (6.5) 5.2 3.5, 2.5 (2.3) 9.4, 6.6
W W V 5, 3, 1 7, 5.5, 2 5.2 3.5, 2.5 9.4, 6.6
Restriction fragments in C3H +I+, WV+, and W4/W4 DNA that hybridize with different portions of the pK3A clone are listed by size (kb). pK3A probes were: (A) the full 1.85 kb insert corresponding to amino acids 165-791 of the c-kit protein: (6) a 225 bp EcoRIlAccl fragment encoding amino acids 165-240; (C) a 390 bp Stul fragment encoding amino acids 342-477; and (D) a 777 bp PvulllEcoRl fragment encoding amino acids 532-791. The sizes of novel hybridizing fragments in the mutant DNAs are listed in parentheses on the right. In the case of probe C, the presence or absence of the altered 10 kb BamHl fragment in W441nWr homozygote DNA cannot be determined be- cause of the presence of the normal 9.6 kb BamHl fragment.
c-kit genomic DNA indicating the site of the rearrange- ment in W44 is shown in Figure 3.
Analogous deductions can be made regarding the site of the W DNA rearrangement. These conclusions are limited by our inability to determine directly which of the wild-type fragments gives rise to altered DNA fragments
in WV+ DNA. Despite this limitation, it is most likely that the W DNA rearrangement is within the genomic DNA segment encoding amino acids 342 to 791. This conclu- sion is based on the fact that the two cDNA probes representing the coding segment between amino acids 342-477 and the segment between amino acids 532-791
A. x X'
-28s -28s
-18s . ., ._ :,,,,,i:
Figure 2. Northern Analysis of c-kit mRNA in W44/lnp4 Mice
(A) Thirty micrograms of total brain RNA was hybridized with the radiolabeled 1.85 kb insert from the K3A cDNA clone. The position of the 28s and 185 ribosmal RNAs are shown to the right. (B) Ethidium bromide-stained RNA gel from which the Northern blot was derived. Note that the W44/w44 homozygote RNA was significantly over- loaded in order to enhance sensitivity of detection.

Mouse W Locus Encodes c-kit Proto-Oncogene 189
EcoRX
ll I 8 I 5 1 3,1
Psi I I I
I 3.8 I L.l.5. 0.8 I 5.2,3.5,2.5
Bgl II
13.3 I u 15.2 I 9.4 6.6 I
t T w44 Wx
5’ 5 C D 3’
X3A
165 240 342 411 532 791
Figure 3. A Schematic Map of the c-kit Genomic DNA Region Encod- ing Amino Acids 165-791 A schematic representation of the organization of the genomic DNA containing the exons corresponding to cDNA clone K3A is shown. The proposed organization is based on the data given in Figure 1 and Table 2. Representations are not intended to be to scale. Restriction frag- ments that do not contain exons have not been detected by hybridiza- tion to the cDNA probe. The order of some restriction fragments cannot be deduced from the current data set. In this case, restriction frag- ments that cannot be ordered are placed together on the map sepa- rated by commas. Restriction fragments that are absent in kV%I@ homozygotes are shown in bold type and are underlined. The most likely locations of the w14 and Wx rearrangements are shown by ar- rows. The placement of restriction fragments on the map is based on the following considerations. Bglll: The 3.3 kb fragment hybridizes exclusively to probe 9, hence it must include exons for amino acids 165-240. The 2 kb, 5 kb, and 9.4 kb fragments hybridize to probe C. However, the 9.4 kb fragment also hybridizes to probe D, indicating that it includes exons from both probes and must map to the right of the 2 kb and 5 kb fragments, which cannot be ordered at present. The 9.4 kb fragment must include exons that encode amino acids 477-532, since it hybridizes to both probes C and D, which bracket this region. The 2.5 kb fragment missing in !@ DNA must map exclusively to the amino acid 240-342 segment of K3A since it does not hybridize to probes B, C, or D. Pstl: The 3.8 kb fragment is homologous exclusively to probe B. The 7 kb fragment missing in kV‘?w homozygotes shows homology to probe C, as does the 1.5 kb probe. The 2.5 kb, 3.5 kb, and 5.2 kb frag- ments all show homology to probe D, but their order cannot be deter- mined. The 0.8 fragment does not hybridize to probes B, C, or D, and hence could map either side of the interval defined by probe C. EcoRI: The 17 kb fragment hybridizes to both probes B and C and must include exons corresponding to both probes plus the regions between them. This fragment is absent in W?W44 homozygotes and replaced by two smaller fragments, 14 kb and 7 kb in size. Both the 8 kb and 5 kb fragments hybridize to probe C, but the 5 kb fragment hybridizes to both probes C and D, hence the order of fragments must be 17 kb, 8 kb, 5 kb from left to right. The 3 kb and 1 kb fragments hybridize ex- clusively to probe D and hence are at the right of the map, but cannot be ordered with respect to each other.
hybridize to the same novel 6.5 kb BamHl fragment in WV+ DNA. These probes hybridize with a 5.5 kb BamHl fragment in normal DNA which, therefore, is likely to be the origin of the novel fragment in WY. The most likely site of the W rearrangement is shown in Figure 3.
Level of c-kit mRNA in W44/W44 Tissue The observation of a structural rearrangement in the c-kit locus of the w44/W44 mouse presents so’me intriguing questions. W44 is a mutant that clearly presents a less severe phenotype than many other W mutants. While W44/W44 homozygotes exhibit severe reductions in ga- metogenesis and are completely sterile, the W44 allele has less severe effects on pigment formation than most mutant W alleles. w4 heterozygotes have smaller white belly spots than heterozygotes for most other mutant W al- leles. Pigment in W44/w44 homozygotes is; also less se- verely reduced than in homozygotes for other mutant W alleles. The hematopoietic defect of W44/W44 homozy- gotes is milder than that of homozygotes for most other mutant W alleles. w14/W44 homozygotes are not anemic, but a mild hematopoietic deficiency in these mice has been characterized. They are repopulated by erythroid derivatives of normal marrow transplants without total body irradiation, W44/w44 bone marrow cells form spleen colonies in the CFU-S assay, but the number and size of the colonies are markedly reduced (Geissler and Russell, 1983; Barker and McFarland, 1988). In summary, the mild effects of the W44 mutation on pigment formation and he- matopoiesis argue against complete loss of function of the W locus gene product.
To examine this question further, we conducted North- ern blotting experiments with RNA from W4’VW44 tissues. Previous findings indicate that the brain expresses high levels of c-kit (Yarden et al., 1987; Qiu et al., 1988). We therefore examined brain RNA from wild-type and W44/ W44 animals for c-kif expression. The results are shown in Figure 2. As previously reported, a single 5.5 kb mRNA was observed for c-Mt. It is apparent that c-kit mRNA lev- els in Wa homozygotes are dramatically lower than in wild-type or W @ heterozygotes. The c-kit mRNA observed in W4 homozygotes appeared normal in size.
Possible Structure of the W44 Rearrangement The data given in Table 2 and the observation of reduced expression levels of a normal sized c-kit mRNA suggest a possible model for DNA rearrangement in W44. Two novel EcoRl fragments were observed iin W44 DNA. One (7 kb) shows homology exclusively to cDNA sequences encoding amino acids 342-477, while the other (14 kb) shows homology exclusively to cDNA sequences encod- ing amino acids 165-240. These results suggest the pres-
BamHI: The 17 kb fragment hybridizes to probes B and C and hence must include exons corresponding to both these probes plus the re- gion between them. The 9.6 kb and 5.5 kb fragments hybridize to probe C, but the 5.5 kb hybridizes to probe D while the 9.6 does not. Hence the order must be 17 kb, 9.6 kb, 5.5 kb from left to right. The 7 kb and 2 kb fragments hybridize exclusively to probe D, and hence are at the right of the map, but their order cannot be determined.

Cell 190
ence of a 4-5 kb insertion in the W44 c-kit locus, in the genomic DNA segment encoding amino acids 240-342. Only a single novel fragment was observed with BamHl and Pstl. These results are consistent with the insertion of an element containing EcoRI, BamHI, and Pstl sites into an intron that contains sites for BamHl and Pstl. The fail- ure to detect an altered Bglll fragment in W44 DNA does not directly support an insertion model for the W44 rear- rangement. However, it is possible that the hybridizing DNA sequences in this fragment are present on Bglll frag- ments in W44 that were too small to detect under the con- ditions of gel electrophoresis and transfer used in the ex- periments described in Figure 1 and Table 2. The putative inserted sequences in w4 could lower the level of c-kit mRNA synthesis by a number of possible mechanisms, without eliminating the potential to produce a c-kit mRNA normal in structure.
Discussion
The W locus has intrigued developmental geneticists for over 50 years, because mutations at W affect several criti- cal yet seemingly unrelated developmental programs. The identification of c-kit as the gene product of the W locus provides a new focus for understanding the common bio- logical basis that underlies differentiation of hematopoi- etic progenitors, primary melanocytes, and primary germ cells. The strong structural homology of c-kit to the CSF-1 and PDGF receptors suggests a model for the action of c-kit during differentiation. In the case of PDGFR, a poly- peptide ligand, PDGF, is presumably released by platelets at the site of a wound, promoting proliferation and differ- entiation of cells in the immediate vicinity that are required for wound healing (Scher et al., 1979). CSF-IR also ap- pears to function by causing the proliferation and differen- tiation of macrophage precursors when CSF-1 is released by effector cells (Sacca et al., 1986). These observations suggest that c-kit may also act to stimulate proliferation of cells in which it is expressed upon signal by the appropri- ate ligand. The phenotypes of W mutants suggest that three cell populations in which c-kit function is critical are the pluripotent hematopoietic stem cell, the migrating melanoblast during early embryonic development, and the primordial germ cell during this same period of devel- opment.
The identification of the ligand for c-kit would be of par- ticular importance in understanding its function. While it is clearly interesting to determine whether any previously characterized hematopoietic growth factors are the c-kit ligand, a genetic clue to the nature of the c-kit ligand may be found through characterization of the SI locus of the mouse. The phenotypes of mutants at this locus closely resemble mutants at the W locus. However, unlike W, the defect in SI is not intrinsic to the progenitor stem cells of the affected tissues but rather lies in the environment in which melanoblast, germ cell, and hematopoietic progeni- tors differentiate and proliferate (for review, see Russell, 1979, and Silvers, 1979). The parallel characteristics of SI and W mutants suggest the possibility that SI mutations may reduce or eliminate the function of the effector that
interacts with the receptor encoded by the Wlocus. Molec- ular genetic analysis of the SI locus may thus provide a route to the identification of the ligand for c-kit.
The high frequency of W mutants and the pleiotropic ef- fects of mutation at the W locus have led to the suggestion that the locus may be extremely large and encode multi- ple related polypeptides. However, our results indicate that the c-kit probe hybridizes with a single continuous genomic DNA segment under high stringency hybridiza- tion conditions and thus indicate that there is no evidence for a second genomic DNA segment highly homologous to c-kit. A mechanism for producing alternative c-kit poly- peptides based on alternative splicing is not ruled out by our data. Alternative splicing has been demonstrated for at least two other members of the tyrosine kinase proto- oncogene family. Ben-Neriah et al. (1986) have demon- strated alternative splicing at the 5’ end of the c-abl gene, Alternative splicing has also been noted for c-sm. In some tissues, alternative splicing of the C-SK message results in the synthesis of a protein with a unique six amino acid segment which increases the tyrosine kinase activity of the molecule (Levy et al., 1987). Detection of a similar mechanism involved in the control of c-kit expression could be of great importance in understanding the role of c-kit in differentiation and development.
A detailed understanding of the pattern of expression of the c-kit gene during development would clearly be of great interest. At present, very limited information is avaii- able on this question. It is clear, however, that c-kit is ex- pressed in a broad range of cell types beyond those obvi- ously affected in W locus mutants. The highest level of expression of c-kit in adult animals is observed in brain. To date, it has not been possible to determine the level of c-kit expression in the cell populations known to be af- fected by W locus mutations because of the difficulties in generating large numbers of purified progenitor cells for any of the three affected cell lineages. In situ hybridization experiments would be particularly useful in this context. It is possible that when determinations of c-kit expression levels are carried out on such cells, they will express lev- els of c-kit mRNA comparable to or higher than the levels found in brain.
It is important to consider the possibility that the c-kit gene plays a role of less obvious significance in those tis- sues in which the gene is expressed but which are not se- verely affected in W mutants. In this context, it should be noted that although W locus mutants show no obvious neurological defect, in depth analysis of the neuroanat- omy and neurological function of these mutants has not been performed to date. Further investigation of the ex- pression pattern of c-kit in developing brain of both normal and mutant animals is clearly warranted. The issues posed by the high levels of expression of c-kit in the brain parallel the questions posed by analogous observations for c-src. High levels of expression for this tyrosine kinase proto-oncogene are found in differentiating rat neurons. In these studies, pp60C-src was demonstrated to be localized to nerve growth cone membranes (Maness et al., 1988). However, the role of c-src in neuronal function has not been established.

Mouse W Locus Encodes c-kit Proto-Oncogene 191
Further analysis of the details of transcription patterns of c-kit may also illuminate the relationship between W lo- cus mutants and their phenotypes. It seems likely that the w4 mutation is the result of an insertion that reduces the efficiency with which mature c-kit mRNA is produced. Detailed analysis of the transcription of c-kit gene in tis- sues of the W44 mutant should provide further insight into the mechanistic basis of reduction in c-kit mRNA levels in W44/W44 homozygotes. Analysis of c-kit transcription pat- terns may also provide an understanding of the variations in the severity of effects of various mutant W alleles on he- matopoiesis, melanocyte differentiation, and germ cell de- velopment.
Clearly the identification of the W locus with the c-kit proto-oncogene permits the initiation of many new ave- nues of investigation regarding the control of differentia- tion in mammalian cells. It seems likely that significant in- sight into the role of cell cell communication during dif- ferentiation via tyrosine kinase signalling mechanisms will result from these studies.
Experimental Procedures
Mice carrying several independent, spontaneous W locus mutations were generously provided by Dr. Jane Barker and Dr. David Harrison of the Jackson Laboratory.
The two mutants described in detail here, W44 and w* arose in the C3HIHeJ (C3H) mouse strain. The Wx mutation has been maintained on the strain of origin. The W44 allele, however, has been transferred onto the C57BU6J background by 40 generations of backcrossing and now carries 66 alleles at Pgm-7 and Afp, which map 3 and 10 centimor- gans (CM), respectively, on opposite sides of the W locus. The W44- B6C3H congenic line retains the C3H allele at Wand at D5SC25, an anonymous marker that maps within 1 CM of W (Geissler et al., 1988) kk@ homozygotes, kt@/+, and +I+ mice were generated by mating w4 heterozygotes inter se.
Somatic Cell Hybrid Analysis The generation and characterization of somatic cell hybrids carrying ?he MllQn deletion chromosome on a hamster genetic background has been described elsewhere (Geissler et al., 1988). Briefly, spleen cells from a C3HNU-WgH/+ female mouse were fused with an HPRT- deficient hamster cell line, CHTG49, using polyethylene glycol. Fusion hybrid cells were selected by growth in HAT medium and after several weeks of growth, clones were tested for the presence of cloned DNA markers on chromosome five. Several clones retained markers span- ning the full length of mouse chromosome 5 but lacked an anonymous marker, D5SC2.5, which maps in the wgH deletion.
Southern Hybridization Analysis of Genomic DNA High molecular weight DNA was prepared from mouse-hamster so- matic ceil hybrids and from mouse tissues by the method of Milbrandt et al. (1983). Mouse genomic DNAs were digested with various restric- tion endonucleases and 5-10 pg was electrophoresed on 1% agarose gels containing 0.04 M Tris-acetate (pH 8) and 0.002 M disodium EDTA. Separated restriction fragments were transferred to nylon hybridization membranes (Zetabind, IMF Cuno) by capillary DNA transfer for 16 hr in 1.5 M NaCl/0.15 M sodium citrate (10x SSC). Blots were treated in 0.1x SSC, 0.5% NaDoSO,, at 65X for 15 min, and prehybridized in 50% formamide/0.75 M NaCl/75 mM sodium citrate/O.l% NaDoSOdl mglml denatured salmon sperm DNA at 42’C for 4 hr or longer. Various restriction fragments of the K3A cDNA clone were isolated by elec- trophoresis in low melting point agarose gels and labeled with the Klenow fragment of DNA polymerase I with random hexanucleotide primers (P-L Biochemicals) and [&‘P]dCTP (Feinberg and Vogel- stein, 1983). Blots were hybridized for 16 hr with 10s CPM/ml radiola- beled probe (>lOa CPMIug) washed to a final stringency of 0.1x SSCIO.l% NaDodSO, at 55°C and exposed to Kodak XAR-5 film for 24 to 48 hr at -BOO6 with one intensifying screen. Hybridization signal
was removed from filters for rehybridization by incubation in the pre- hybridization solution at -80°C for 30 min.
Northern Blot Analysis RNA was prepared from the brains of adult mice (2-6 months old) by a single-step guanidinium thiocynate isolation method (Chomczynski and Sacchi, 1987). Thirty micrograms of total brain FINA was fraction- ated on denaturing formaldehyde/agarose gels and transferred to a hy- bridization membrane (GeneScreen Plus, NEN) by electrophoresis in 25 mM phosphate buffer. DNA-RNA hybridization was carried out at 65°C in 1 M NaCI, 10% dextran sulfate (Pharmacia), 100 uglml dena- tured salmon sperm DNA, and lo6 CPM/ml of the radiolabeled 1.85 kb insert from pK3A, labeled as described above. Blots were washed to a final stringency of 0.2x SSC, 65OC and exposed to film for 24-48 hr.
cDNA Isolation c-kit cDNA clones were isolated from a size-selected BALE/c mouse brain cDNA library constructed in hgtll (Citri et al., 1987). The probe was a 53 bp synthetic oligomer complementary to the region of the published mouse c-kit sequence between nucleotides 1627 and 1680 (Qiu et al., 1988). The oligomer was labeled using lpolynucleotide ki- nase and [y3zP]ATP to a specific activity of 1Oa CPMIpg and used to screen replica filters containing 7 x 10s plaques prepared as previ- ously described (Benton and Davis, 1977). The ffilters were prehybri- dized for 16 hr at 42OC in 50% formamide, 5x SSC, 10% dextran sulfate, 1% NaDoS04, and 1 mglml denatured salman sperm DNA. IO6 CPM/ml of the labeled oligomer was added and the hybridization con- tinued for 16 hr.
Filters were washed twice in 5x SSC at room temperature for 30 min and once at 42°C for 5 min. Two positive phage clones were purified, digested with EcoRI, and the inserts verified by Southern hybridization analysis and subcloned into pUCI9 for further analysis. Restriction mapping analysis revealed that pK3A, the cDNA clorre utilized in these studies, contains a 1.85 kb insert comprising the region of the c-kittran- script between nucieotides 500 and 2350.
The authors wish to thank Dr. Elizabeth Russell and Dr. Jane Barker for many helpful discussions concerning the nature of W locus in- fluence on development. We would like to thank Yoav Citri for gener- ously providing his mouse brain cDNA library. We glratefully acknowl- edge discussions with Drs. Richard Mulligan, Cornelia Bargmann, and Alan Buckler concerning the possible biological role of c-kit and their comments on this manuscript. This work was supported by Public Health Service grants GM 27882, CA 17575, and the Hereditary Dis- ease Foundation. E. N. G. acknowledges postdoctoral support of Pub- lic Health Service grant CA-07826. Melanie Ryan was a participant in the Undergraduate Research Opportunities Pr0gra.m at MIT.
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC. Section 1734 solely to indicate this fact.
Received September 8, 1988.
References
Barker, J. E., and McFarland, E. C. (1988). Hemat’opoietic precursor cell defects in nonanemic but stem cell-deficient t i4/W@ mice. J. Cell. Physiol. 135, 533438.
Ben-Neriah, Y., Bernards, A., Paskind, M., Daley, G. Q., and Baltimore, D. (1986). Alternative 5’ exons in cab/ mRNA. Cell 44, 577-586. Benton, W. D., and Davis, R. W. (1977). Screening rZgt recombinant clones by hybridization to single plaques in situ. Science 796, 180-182.
Berent, S., Mahmoudi, M., Torczynski, R. M., Bragg, P. W., and Bollon, A. (1985). Comparison of oligonucleotide and long IDNA fragments as probes in DNA and RNA dot, Southern, Northern, rcolony and plaque hybridizations. BioTechniques 3, 208-219.
Chiquoine, A. D. (1954). The identification, origin, and migration of the primordial germ cells in the mouse embryo. Anat. Rec. 178, 135-146. Chomczynski, P., and Saachi, N. (1987). Single-step method of RNA

Cell 192
isolation by acid guandinium thiocyanate-phenol chloroform extrac- tion. Anal. Biochem. 762, 156-159.
Citri, Y., Colot, H. V., Jaquier, A. C., Yu, Q., Hall, J. C., Baltimore, D., and Rosbash, M. (1967). A family of unusually spliced and biologically active transcripts is encoded by a Drosophila clock gene. Nature 326, 4.2-47.
Feinberg, A. P., and Vogelstein, B. (1983). A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Anal. Biochem. 732, 6-13.
Geissler, E. N., and Russell, E. S. (1983). Analysisof the hematopoietic effects of new dominant spotting (W) mutations of the house mouse. I. lnffuence upon hematopoieticstem cells. Exp. Hematol. 77,452-460. Geissler, E. N., McFarland, E. C., and Russell, E. S. (1981). Analysis of pleiotropism at the dominant white-spotting(W) locus of the house mouse: a description of ten new W alleles. Genetics 97, 337-361.
Geissler, E. N., Cheng, S. V., Gusella, J. F., and Housman, D. E., (1988). Genetic analysis of the dominant white-spotting region on mouse chro- mosome 5; identification of cloned DNA markers near W. Proc. Natl. Acad. Sci. USA, in press.
Glaser, T, Lewis, W. H., Bruns, G. A. P., Watkins, P C., Rogler, C. E., Shows, T B., Powers, V. E., Willard, H. F., Goguen, J. M., Simola, K. 0. J., and Housman, D. E. (1986). The b-subunit of follicle- stimulating hormone is deleted in patients with aniridia and Wilms’ tumour, allowing a further definition of the WAGR locus. Nature 327, 882-887.
Gregory, C. J., and Eaves, A. C. (1978). Erythropoietic progenitor cell differentiation distinguished by a number of physical and biogical prop- erties. Blood 57, 527-536.
Gruneberg, H., and Truslove, G. M. (1960). Two closely-linked genes in the mouse. Genet. Res. 7, 69-90.
Harrison, D. E., and Astle, C. M. (1976). Population of lymphoid tissues in cured W anemics by donor cells. Transplantation 22, 42-46.
Levy, J. B., Dorai, T., Wang, L., and Brugge, J. S. (1987). The structur- ally distinct form of pp60C-src detected in neuronal cells is encoded by a unique c-src mRNA. Mol. Cell. Biol. 7, 4142-4145.
Lyon, M. F., Glenister, P. H., Loutit, J. F., Evans, E. f?, and Peters, J. (1984). A presumed deletion covering the Wand Pn loci of the mouse. Genet. Res. 44, 161-166.
Maness, P F., Aubry, M., Shores, C. G., Frame, L., and Pfenninger, K. H. (1988). c-src gene product in developing rat brain is enriched in nerve growth cone membranes. Proc. Natl. Acad. Sci. USA 85, 5001-5005.
Mayer, T. C., and Green, M. C. (1968). An analysis of the pigment defect caused by mutations at the Wand SI loci in mice. Dev. Biol. 78, 62-75. McCulloch, E. A., Siminovitch, L., and Till, J. (1964). Spleen colonyfor- mation in anemic mice of genotype WIW Science. 744, 844-846.
Milbrandt, J. D., Azizkahn, J. C., Greisen, K. S., and Hamlin, J. L. (1983). Organization of a Chinese hamster ovary dehydrofolate induc- tase gene identified by phenotypic rescue. Mol. Cell. Biol. 3, 1266- 1273.
Mintz, B., and Russell, E. S. (1957). Gene induced embryological modifications of primordial germ cells in the mouse. J. Exp. Zool. 134, 207-237.
Moore, M. A. S., and Metcalf, D. (1970). Ontogeny of the haemopoietic system: yolk sac origin. Brit. J. Haematol. 78, 279-296.
Murphy, E. D., Harrison, D. E., and Roth&J. B. (1973). Giant granules of beige mice, a quantitative marker for granulocytes in bone marrow transplantation. Transplantation 75, 526-530. Rawles, M. E. (1947). Origin of pigment cells from the neural crest in the mouse embryo. Physiol. Zool. 20, 248-266. Russell, E. S. (1949). Analysis of pleiotropism at the W locus in the mouse. Relationship between the effects of Wand WY substitution on hair pigmentation and on erythrocytes. Genetics 34, 706-726. Russell, ES. (1954). Review of the pleiotropic effects of Wseries genes on growth and differentiation. In Aspects of Synthesis and Order of Growth, 13rh Symp. Sot. Study Develop., D. Rudnick, ed. (Princeton, New Jersey: Princeton University Press), pp. 113-126. Russell, E. S. (1970). Abnormalities of erythropoiesis associated with
mutant genes in mice. In Regulation of Hematopoiesis, Volume 1, A. S. Gordon, ed. (New York: Appleton), pp. 649-675.
Russell, E. S. (1979). Hereditary anemias of the mouse; a review for geneticists. Adv. Genet. 20, 357-459.
Russell, E. S., and Benstein, S. E. (1968). Proof of whole cell implant in therapy of Wseries anemia. Arch. Biochem. Biophys. 725594-597.
Russell, E. S., Smith, L. J., and Lawson, F. A. (1956). Implantation of normal blood-forming tissue in irradiated genetically anemic hosts. Science 724, 1076-1077.
Russell, E. S., Thompson, M. W., and McFarland, E. C. (1968). Analysis of effects of Wand f genie substitutions on fetal mouse hematology. Genetics 58, 259-270.
Sacca, R., Stanley, E. R., Sherr, C. J., and Rettenmeier, C. W. (1986). Specific binding of the mononuclear phagocyte stimulating factor to the product of the c-fms protooncogene. Proc. Natl. Acad. Sci. USA 83, 3331-3335.
Scher, C. D., Shepard, R. C., Antoniades, H. N., and Stiles, C. D. (1979). Platelet-derived growth factor and the regulation of the mammalian fi- broblast cell cycle. Biochim. Biophys. Acta 560, 217-241.
Silvers, W. K. (1979). White-spotting, patch and rump-white. In The Coat Colors of Mice: A Model for Gene Action and Interaction (New York: Springer-Verlag), pp. 206-241. Qiu, F., Ray, f?, Brown, K., Barker, P E., Jhanwar, S., Ruddle, F. H., and Besmer, l? (1988). Primary structure of c-kit: relationship with the CSF- l /PDGF receptor kinase family-oncogenic activation of v-kit involves deletion of extracellular domain and c-terminus. EMBO J. 7, 1003-1011.
Yarden, Y., Kuang, W., Yang-Feng, 1, and Ullrich, A. (1987). Human proto-oncogene c-kit: a new cell surface receptor tyrosine kinase for an unidentified ligand. EMBO J. 6, 3341-3351.
Notes Added in Proof
Chabot et al. report close linkage of c-kit and Wand the location of c-kit in the t@H deletion. These data are consistent with the data reported herein. See Chabot, B., Stephenson, D. A., Chapman, V. M., Besmer, P., and Bernstein, A. (1988). Nature 335, 88-89.
K3A, fragment C, in fact comprises 460 bp and encodes amino acids 342-495 of c-kit. This does not affect the interpretation of the results.
Copyright © 2022 FDOKUMEN




















