
Signal transduction pathways involving the raf proto-oncogene
12
Cancer and Metastasis Reviews 13: 105-116, 1994. © 1994 Kluwer Academic Publishers. Printed in the Netherlands. Signal transduction pathways involving the Raf proto-oncogene Nidhi Gupta Williams ~'2and Thomas M. Roberts 2 Departments of JBiological Chemistry and Molecular Pharmacology, and 2 Pathology, Harvard Medical School, Boston MA 02115 and 2 Dana Father Cancer Institute, Boston, MA 02115, USA Key words: Raf, protein kinase, baculovirus, MAP kinase Abstract The raf genes encode for a family of cytoplasmic proteins (A-raf, B-raf and c-raf-1) with associated serine/ threonine kinase activities. Raf-1 is an important mediator of signals involving cell growth, transformation and differentiation. It is activated in response to a wide variety of extracellular stimuli such as insulin, nerve growth factor (NGF), platelet derived-growth factor (PDGF), and in response to expression of oncogenes, v-src and v-ras, in a cell-specific manner. Recently, the first physiological substrate for Raf-1 protein kinase was identified. Raf-1 was found to phosphorylate and activate Mitogen-Activated Protein Kinase Kinase (MEK), an activator of MAP kinase, thus linking the Raf-1 signaling pathway with that of MAP kinase. Cell specific differences in signalling pathways involving Raf-1 and MAP kinase have also been discovered. Accu- mulating evidence indicates that membrane tyrosine kinases, ras, Raf-1, MEK and MAP kinase are intercon- nected via a complex network rather than via a linear pathway involving multiple substrates and feedback loops. Discovery of the raf-family of kinases Identification of many cellular genes important in the progression and control of cell growth has been facilitated by studies on transforming viruses [1]. MSV3611, an acute transforming murine sarcoma retrovirus, was first isolated a decade ago by Ulf Rapp, et al. [24]. It was found to induce fibrosarco- mas in vivo and transform embryo fibroblasts and epithelial cells in culture. V-raf, the protein product of the transforming gene of this retrovirus, encodes a 90 Kd protein with homology to the serine-threo- nine family of kinases [5]. Isolation of c-raf-1 [6, 7], the cellular homolog of v-raf, was followed by iden- tification and cloning of other members of the raf family of kinases. These included A-raf [8], B-raf [9, 10] and D-raf (the drosophila homolog of c-raf-1) [11, 12]. Though Raf-1 (72-74 Kd) [7], the protein product of the c-raf-1 gene, is found to be ubiqui- tously expressed, A-raf (68 Kd) and B-raf (95 Kd) [13] have a more tissue specifc distribution [14] with A-raf being most abundant in epididymis, urogen- ital tissue and ovary and B-raf in fetal brain and adult cerebrum and testis. The Raf proteins are cytoplasmically localized and play a critical role in the transmission of signals from cell surface receptors to transcription factors in the nucleus [15]. Several other cytoplasmic pro- tein kinases have also been implicated in signal transduction from the cell surface through the cy- toplasm. These include Mitogen-activated Protein (MAP) kinase, Protein Kinase C (PKC) and riboso- mal $6 kinase (rsk) [16-18]. It is now known that some of these kinases work in concert and may lie on the same signalling pathway. This review will center on the Raf family of kinases and their role in the signal transduction process.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Signal transduction pathways involving the raf proto-oncogene

Cancer and Metastasis Reviews 13: 105-116, 1994. © 1994 Kluwer Academic Publishers. Printed in the Netherlands.
Signal transduction pathways involving the Raf proto-oncogene
Nidhi Gupta Williams ~'2 and Thomas M. Roberts 2 Departments of JBiological Chemistry and Molecular Pharmacology, and 2 Pathology, Harvard Medical School, Boston MA 02115 and 2 Dana Father Cancer Institute, Boston, MA 02115, USA
Key words: Raf, protein kinase, baculovirus, MAP kinase
Abstract
The raf genes encode for a family of cytoplasmic proteins (A-raf, B-raf and c-raf-1) with associated serine/ threonine kinase activities. Raf-1 is an important mediator of signals involving cell growth, transformation and differentiation. It is activated in response to a wide variety of extracellular stimuli such as insulin, nerve growth factor (NGF), platelet derived-growth factor (PDGF), and in response to expression of oncogenes, v-src and v-ras, in a cell-specific manner. Recently, the first physiological substrate for Raf-1 protein kinase was identified. Raf-1 was found to phosphorylate and activate Mitogen-Activated Protein Kinase Kinase (MEK), an activator of MAP kinase, thus linking the Raf-1 signaling pathway with that of MAP kinase. Cell specific differences in signalling pathways involving Raf-1 and MAP kinase have also been discovered. Accu- mulating evidence indicates that membrane tyrosine kinases, ras, Raf-1, MEK and MAP kinase are intercon- nected via a complex network rather than via a linear pathway involving multiple substrates and feedback loops.
Discovery of the raf-family of kinases
Identification of many cellular genes important in the progression and control of cell growth has been facilitated by studies on transforming viruses [1]. MSV3611, an acute transforming murine sarcoma retrovirus, was first isolated a decade ago by Ulf Rapp, et al. [24]. It was found to induce fibrosarco- mas in vivo and transform embryo fibroblasts and epithelial cells in culture. V-raf, the protein product of the transforming gene of this retrovirus, encodes a 90 Kd protein with homology to the serine-threo- nine family of kinases [5]. Isolation of c-raf-1 [6, 7], the cellular homolog of v-raf, was followed by iden- tification and cloning of other members of the raf family of kinases. These included A-raf [8], B-raf [9, 10] and D-raf (the drosophila homolog of c-raf-1) [11, 12]. Though Raf-1 (72-74 Kd) [7], the protein product of the c-raf-1 gene, is found to be ubiqui- tously expressed, A-raf (68 Kd) and B-raf (95 Kd)
[13] have a more tissue specifc distribution [14] with A-raf being most abundant in epididymis, urogen- ital tissue and ovary and B-raf in fetal brain and adult cerebrum and testis.
The Raf proteins are cytoplasmically localized and play a critical role in the transmission of signals from cell surface receptors to transcription factors in the nucleus [15]. Several other cytoplasmic pro- tein kinases have also been implicated in signal transduction from the cell surface through the cy- toplasm. These include Mitogen-activated Protein (MAP) kinase, Protein Kinase C (PKC) and riboso- mal $6 kinase (rsk) [16-18]. It is now known that some of these kinases work in concert and may lie on the same signalling pathway. This review will center on the Raf family of kinases and their role in the signal transduction process.

106
N - t e r m i n a l 1 CR1 CR2
' a m i n o 139 6 251 301 a c i d
C - t e r m i n a l
CR3 6 4 8
P, ?,- ' , ; \ . \ \ \ " \ \ \ \ ' \ ',~\.\.\ ?-. \ \ \ " . \ \ \ " , "\"(\\~1 I [ \5, . \ \ \~ '~, \ , \ ,x4\ \ \ \ . \, ,,;\.\\,:?,.,£\x.\.,,,~\\~:, : , . \ ~ I I I
3 2 0 6 3 5
Fig. 1. Structure of the RaM protein indicating conserved regions 1, 2 and 3 (CR1, CR2 and CR3).
Structural analysis of Raf-1
Raf-1 is a 648 amino acid protein with intrinsic se- rine-threonine kinase activity. Analysis of trans- formed NIH3T3 cells transfected with DNA from several human tumors revealed the presence of truncated Raf sequences [19, 20]. A detailed dele- tion analysis revealed that the transforming poten- tial of the C-terminal kinase domain of RaM is sup- pressed by a regulatory N-terminal domain. Inde- pendent studies demonstrated that deletion of the entire N-terminal domain (aa 2 to aa 305) [21] or deletion of just 55 aa of a serine-threonine rich do- main (aa 225 to aa 280) [22] resulted in truncated Raf proteins that are capable of transforming NIH3T3 cells. Based on structure-function analysis and by comparing conserved sequences, the Raf-1 protein kinase has been divided into three domains, conserved regions 1, 2 and 3 (CR1, CR2 and CR3) [23] (Fig. 1). The first of these domains, CR1, con- tains a conserved cysteine motif (identified by Ishi- kawa, et al. [22]), similar in sequence to the lipid- binding cysteine motif identified in Protein Kinase C (PKC). CR1 has a consensus sequence of C-X2-C- X7-B-C-X2-C-X7-C-N7-C. The second domain con- tains a conserved serine/threonine rich tract, CR2, with unknown function [22]. The third domain, CR3, is the conserved kinase domain of Raf-1. CR3 displays extensive homology with other known ki- nases [24]. Removal of the N-terminal 'regulatory' portion, consisting of CR1 and CR2, of the Raf-1 protein results in a C-terminal kinase domain (CR3) which is constitutively active and has trans- forming potential [21, 23]. It must be noted that al- though deletion of CR1 alone, as shown in two inde- pendent studies [19, 20], resulted in only a small in- crease in transforming activity of RaM, trans-
formed cell lines with viral insertions resulting in deletion of CR1 alone, have been isolated [25, 26].
Raf-1 is phosphorylated prior to stimulation of its activity in vivo [27, 28]. It is found to be phosphory- lated mostly on serine residues, identified as Ser43, Ser259 and Ser621 [29]. As reported by some [29] and contradicted by others [27], phosphorylation at these sites is stimulated upon addition of growth factors. Morrison et al. [29] have reported modula- tion of RaM kinase activity by phosphorylation at Ser259 and Ser621. Others have reported phospho- rylation at these sites prior to stimulation of Raf-1 activity [27] [unpublished results, Williams NG, Ro- berts TM], indicating that phosphorylation at these sites may not be sufficient for activation of Raf-1.
Relationship between Raf-1 and other signal transducers and oncogene products
Oncogene products and their cellular counterparts are thought to play key roles in cellular processes such as growth, differentiation and transformation. In a study investigating the role of several onco- genes in cell growth and transformation, a condi- tional relationship between v-raf and other onco- gene products, v-src, v-fms and v-ras was establish- ed. These studies involved microinjection of neu- tralizing antibodies directed against the ras protein [30, 31]. Microinjection of inhibitory anti-ras anti- bodies into NIH3T3 cells, reversed the transformed phenotype of cells transformed by v-src, v-ras and v-fms. The transformed phenotype of v-raf-trans- formed cells, on the other hand, was unaffected by the inhibitory anti-ras antibody injections. These observations suggested that Raf-1 acts downstream of both v-ras and v-src, assuming that the pathway for activation of Raf-1 involves ras. Alternatively,

the observations were also consistent with position- ing of v-ras and v-raf on independent signal trans- duction pathways.
Raf-1 is involved in the mitogen-dependent signalling cascade
Raf-1 is activated in response to stimulation o f membrane tyrosine kinases
Evidence that Raf-1 is an intermediate in the cas- cade of signals initiated by receptors at the cell sur- face, was first provided by Morrison et al. [32]. RaM, immunuprecipitated from NIH3T3 cells, was shown to be retarded in electrophoretic-mobility and hyperphosphorylated in a rapid response to treatment of cells with either Platelet-derived growth factor (PDGF), Fibroblast growth factor (FGF), Epidermal growth factor (EGF) or Protein kinase C (PKC). Further, RaM was found to be re- tarded in gel-mobility when cells were transformed by either v-src, v-sis, v-fms, polyoma Middle-T anti- gen or v-ras but not from cells transformed by nu- clear oncogenes such as v-fos and v-myc. These re- sults tentatively placed Raf-1 downstream of mem- brane tyrosine kinase receptors, ras and PKC and upstream or independent of the nuclear protoonco- genes fos and myc. Similar reports followed, indi- cating hyperphosphorylation and retardation in gel-mobility of Raf-1 in response to a variety of other stimulating factors such as Nerve growth fac- tor (NGF) [33, 34], Colony stimulating factor (CSF) [35], EGF [361, insulin [27, 37, 38], IL-2 [39, 40], erythropoietin [42] and GM-CSF [43] as well as in response to activation of the T-cell receptor [44].
The direct association of Raf-1 with cell surface receptors and subsequent phosphorylation of Raf-1 on tyrosine residues identified as tyr340 and tyr341 [45], is controversial, as others have presented con- tradictory data [27]. However, it is clear that tyro- sine phosphorylation of Raf-1 is not always neces- sary or sufficient for activation of Raf-1 [27, 28].
107
Assays for Raf-1 activity
The conclusions described above were based main- ly upon Raf-1 immune-complex kinase assays con- ducted in vitro, using several different substrates in- cluding Histone H1 [36, 43], syntide 2 [38, 45] and a Raf-1 peptide corresponding to a sequence in the amino-terminal domain of Raf-1 [35, 37, 39]. In an- other report, immune complex autokinase assays were used to illustrate modulation of B-raf kinase activity upon stimulation of PC12 cells with NGF [13, 46]. With the exception of a report by Isumi et al. [27] and a report by us [28], none of the kinase as- says included a kinase-inactive mutant of Raf-1 (Raf301) as a control. Thus it is possible that part or all of the kinase activity demonstrated may corre- spond to kinases other than RaM. We [27, 28] have not been able to demonstrate Raf-1 specific phos- phorylation of Histone H1, syntide or the Raf-based peptide in immune-complex kinase assays as re- ported by others. However, we showed that Raf-1 auto-kinase reactions, performed in vitro, provided a sensitive assay for measurement of kinase activity of Raf-1 (Raf301 was used as a control in each case, see below), and thereby illustrated modulation of the kinase activity of RaM by insulin receptor, PDGF receptor, v-src and v-ras [27, 28]. These re- sults, as well as supporting genetic data from dro- sophila, have clearly established Raf-1 as a common intermediate in signal transduction pathways initi- ated by tyrosine kinases at the cell surface. A re- cently discoved kinase called MEK (Map or Erk ki- nase), is the only known physiological substrate of Raf-1 [47-49]. A repetition of all the Raf-1 immune- complex kinase assays, using MEK as an exogenous substrate, will be required to accurately measure the extent of modulation of Raf-1 kinase activity by each of the different receptors. Interestingly, al- though there is no indication that autophosphoryla- tion of Raf-1 occurs in vivo, the amount of Raf-1 au- tophosphorylation activity from immune-complex autokinase assays conducted in vitro, correlates with MEK phosphorylation by RaM in immune- complex kinase assays [Narasimhan V, Peralta E, personal communication], [50].
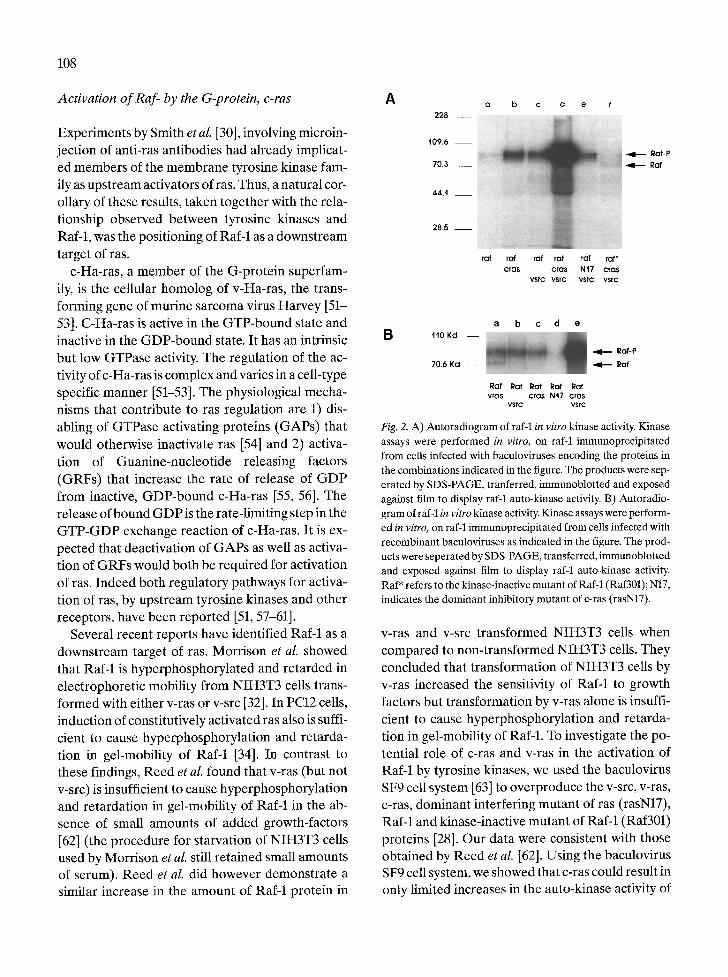
Act ivat ion o f Raf- by the G-protein, c-ras A
Experiments by Smith et al. [30], involving microin- jection of anti-ras antibodies had already implicat- ed members of the membrane tyrosine kinase fam- ily as upstream activators of ras. Thus, a natural cor- ollary of these results, taken together with the rela- tionship observed between tyrosine kinases and Raf-1, was the positioning of Raf-1 as a downstream target of ras.
c-Ha-ras, a member of the G-protein superfam- fly, is the cellular homolog of v-Ha-ras, the trans- forming gene of murine sarcoma virus Harvey [51- 53]. C-Ha-ras is active in the GTP-bound state and inactive in the GDP-bound state. It has an intrinsic but low GTPase activity. The regulation of the ac- tivity of c-Ha-ras is complex and varies in a cell-type specific manner [51-53]. The physiological mecha- nisms that contribute to ras regulation are 1) dis- abling of GTPase activating proteins (GAPs) that would otherwise inactivate ras [54] and 2) activa- tion of Guanine-nucleotide releasing factors (GRFs) that increase the rate of release of GDP from inactive, GDP-bound c-Ha-ras [55, 56]. The release of bound GDP is the rate-limiting step in the GTP-GDP exchange reaction of c-Ha-ras. It is ex- pected that deactivation of GAPs as well as activa- tion of GRFs would both be required for activation of ras. Indeed both regulatory pathways for activa- tion of ras, by upstream tyrosine kinases and other receptors, have been reported [51, 57-61].
Several recent reports have identified RaM as a downstream target of ras. Morrison et aL showed that RaM is hyperphosphorylated and retarded in electrophoretic mobility from NIH3T3 cells trans- formed with either v-ras or v-src [32]. In PC12 cells, induction of constitutively activated ras also is suffi- cient to cause hyperphosphorylation and retarda- tion in gel-mobility of Raf-1 [34]. In contrast to these findings, Reed et al. found that v-ras (but not v-src) is insufficient to cause hyperphosphorylation and retardation in gel-mobility of Raf-1 in the ab- sence of small amounts of added growth-factors [62] (the procedure for starvation of NIH3T3 cells used by Morrison et al. still retained small amounts of serum). Reed et aL did however demonstrate a similar increase in the amount of Raf-1 protein in
a b c d e f
228 _ _
109 .6 _ _
70 .3 _ _
44 .1 _ _
28.5 - -
108
raf rof raf raf rof raf' cras cras N17 cras
vsrc vsrc vsrc vsrc
~ I ~ Raf-P
Raf
a b c d e
B ~o Kd - -
Rof-P
70 .6 Kd ~ Raf
Raf Raf Raf Rof Raf vras c ras N17 c ras
vs r c vs r c
Fig. 2. A) Autoradiogram of raf-1 in vitro kinase activity. Kinase assays were performed in vitro, on r am immunoprecipitated from cells infected with baculoviruses encoding the proteins in the combinations indicated in the figure. The products were sep- erated by SDS-PAGE, tranferred, immunoblotted and exposed against film to display raf-1 auto-kinase activity. B) Autoradio- gram of raf-1 in vitro kinase activity. Kinase assays were perform- ed in vitro, on raf-1 immunoprecipitated from cells infected with recombinant baculoviruses as indicated in the figure. The prod- ucts were seperated by SDS-PAGE, transferred, immunoblotted and exposed against film to display raf-1 auto-kinase activity. Raf* refers to the kinase-inactive mutant of Raf-1 (Raf301); N17, indicates the dominant inhibitory mutant of c-ras (rasN17).
v-ras and v-src transformed NIH3T3 cells when compared to non-transformed NIH3T3 cells. They concluded that transformation of NIH3T3 cells by v-ras increased the sensitivity of Raf-1 to growth factors but transformation by v-ras alone is insuffi- cient to cause hyperphosphorylation and retarda- tion in gel-mobility of RaM. To investigate the po- tential role of c-ras and v-ras in the activation of RaM by tyrosine kinases, we used the baculovirus SF9 cell system [63] to overproduce the v-src, v-ras, c-ras, dominant interfering mutant of ras (rasN17), Raf-1 and kinase-inactive mutant of Raf-1 (Raf301) proteins [28]. Our data were consistent with those obtained by Reed et al. [62]. Using the baculovirus SF9 cell system, we showed that c-ras could result in only limited increases in the auto-kinase activity of

R a M (Fig. 2A, lanes b and c). However, coinfection with both v-src and c-ras showed that src and ras act synergistically to activate the autokinase activity of c-raf protein (Fig. 2A, lane d). The activity most likely reflects the kinase activity of R a M since the kinase-inactive mutant (Raf301) was inactive when coexpressed with src and ras (Fig. 2A, lane f). A re- tardation in gel-mobility of Raf-1 protein from SF9 cells was observed only when v-src and c-ras were both present [28]. In a linear pathway connecting v-src to c-ras to RaM, v-ras alone should activate Raf-1 in the absence of v-src, since v-ras is constitu- tively activated. Activity of RaM, immunoprecip- itated from cells doubly infected with baculoviruses encoding Raf-1 and v-ras, should be the same as that from cells triply infected with baculoviruses encod- ing Raf-1, v-src and c-ras. Surprisingly this was not true. V-ras did not activate RaM to the same extent as did v-src and c-ras (or v-ras) combined (Fig. 2B, compare lanes a and e). Further a dominant inhib- itory mutant of ras (rasN17), did not block v-src-de- pendent activation of R a M indicating presence of a ras-independent pathway (Fig. 2A, lane e). These results clearly indicated that although ras is essen- tial for activation of Raf-1, full activation of Raf-1 requires both a tyrosine-kinase activity and a ras- dependent activity.
There are several aspects of the baculovirus SF9 cell system that need further elaboration here. 1) Proteins are overexpressed many fold over normal cellular levels. Thus normal regulatory factors may now exert relatively little control, resulting in con- stitutive activation of the overexpressed protein. In the SF9 system, c-ras is nearly as effective as v-ras in activating Raf-1. It is likely that GTPases such as GAP that would normally regulate c-ras activity are either absent or limiting in SF9 cells, rendering c-ras almost as active as v-ras [64]. 2) Coexpression of proteins using the SF9 cell system can be used to analyze effects of regulatory proteins, such as GAR on the activity of c-ras. 3) Overexpression and the ease of viral infections together allow for use of con- trols such as kinase-inactive mutants. 4) Proteins are produced and can interact in a more physiolog- ical environment than in in vitro experiments since membrane proteins and nuclear proteins are appro- priately targeted and modified.
109
In the experiments described in SF9 cells, mod- ulation of Raf-1 activity was measured by immune- complex autokinase assays [28]. As mentioned above, the extent of autophosphorylation of Raf-1 in vitro appears to correlate well with kinase activity of Raf-1 towards MEK, the newly discovered sub- strate of RaM (discussed below) [50]. Yet, several questions remain unanswered. What does retarda- tion in gel-mobility of Raf-1 mean and how well does it correlate with activation of Raf-l? Is the modification resulting in the change in gel-mobility of Raf-1 required for Raf-1 activation or is it a con- sequence of Raf-1 activation. Most importantly, are all observed retardations in gel-mobility of RaM due to the same modifications of Raf-l? Answers to several of these questions have recently appeared due to the discovery of a Raf-1 substrate i.e. the MEK protein, and an analysis of the interactions between Raf-1, MEK and Mitogen activated pro- tein (MAP) kinase.
The search for a Raf-1 substrate: Activation of M E K and M A P kinases by Raf-I
It was recognized that the set of stimuli that affected members of the Raf family of kinases also activated members of the Mitogen-activated protein (MAP) family of kinases [65-68]. Interestingly, both Raf and MAP families of kinases are ubiquitously ex- pressed [14, 69]. The MAP kinases, also known as extracellular signal-regulated kinases (ERKs), rep- resent a growing family of cytoplasmic kinases [69, 70]. At least two MAP kinase isoforms, ERK1 (44 Kd) and ERK2 (42 Kd), are activated in re- sponse to a wide variety of extracellular stimuli such as insulin, nerve growth factor (NGF), platelet de- rived-growth factor (PDGF), epidermal growth factor (EGF), phorbol esters and okadaic acid [71- 73]. Both Raf-1 and ERK1 appear to be regulated by v-ras and by membrane tyrosine kinases [34, 74- 78]. Expression of activated ras in PC12 cells is suffi- cient to activate ERK1 partially and to cause hyper- phosphorylation and retardation in gel-mobility of R a M [34, 78]. Moreover, expression of a dominant interfering allele of ras (rasN17) is sufficient to block NGF-induced ERK1 activation and hyper-

110
phosphorylation of Raf-1 [34, 78]. Since Raf and MAP kinase appear to be co-regulated, it is not sur- prising, as discussed below, that recent reports place the two proteins in the same signalling pathway in some cell types [65-68].
The mechanism of activation of the cytoplasmic serine-threonine MAP kinases, ERK1 and ERK2, is clearly of interest. In response to extracellular sti- muli, ERK1 and ERK2, undergo rapid phosphory- lation on threonine and tyrosine residues and retar- dation in gel-mobility. Activation of MAP kinases ERK1 and ERK2 requires phosphorylation of these proteins on both tyrosine and threonine residues [79, 80]. These tyrosine and threonine residues, lo- calized to Thr183 and Tyr185 in ERK2, are con- served in both MAP kinase isoforms. They are the only known cytoplasmic serine-threonine kinases known to be activated by tyrosine phosphorylation. Cytoplasmic MAP kinase activating factors (named Map or Erk Kinases, MEK) have been identified [81, 82], purified to homogeneity [72, 83, 84] and re- cently cloned [47-49]. The MEK proteins (also known as MAP kinase kinase, MAPKK) were found to be threonine/tyrosine kinases that are acti- vated by serine/threonine phosphorylation. Activa- ted MEK can phosphorylate MAP kinases, ERK1 and ERK2 in vitro, on threonine and tyrosine resid- ues and activate them. Raf-1 has been shown to be a substrate of MAP kinase, in vitro, although no change in Raf-1 activity was noted [85, 86]. Conver- sely, MAP kinase is found to be constitutively acti- vated in NIH3T3 cells transformed by v-raf [66, 67, 87], suggesting that RaM may also function up- stream of MAP kinases to activate them. This para- dox was finally resolved when it was shown that Raf-1 activated MAP kinase by directly phosphory- lating and activating the newly discovered MAP ki- nase activator, MEK, in vitro [66-68]. Although it appears that MEK is a direct substrate of RaM, the requirement for additional cofactors cannot be ruled out. MEK is the first and only known physio- logical substrate for Raf-1 kinase.
The baculovirus SF9 cell system was used to ex- amine the effects of Raf-1, v-ras, v-src and MAP ki- nase (ERK1) on each other [65]. As reported by us and by others [65-68, 87], ERK1 is activated by coexpression with either v-Raf or constitutively ac-
A R 2 2 W - + - + -
v - R a f + + .
E R K 1 - + + + +
v - r a s - - - +
1 2 3 4 5
M B P •
B R a f - 1 + . . . . + - + + - - - + + -
R a f * - + . . . . + - - + . . . . +
E R K 1 - - + - + + + - + + - + - + +
v - r a s - - - + + - - + + + . . . . .
v - s r c . . . . . . . . . + + + + +
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5
M B P t~"
Fig. 3. Autoradiograms displaying the phosphotransferase activ- ity of ERK1 (MAP kinase) towards added myelin basic protein (MBP) in vitro. ERK1 was immunoprecipitated from lysates of cells infected with viruses in the combinations indicated. Im- mune complex kinase assays on ERK1 immunoprecipitates were performed to measure phosphotransferase activity towards ex- ogenously added MBR Products from the kinase reaction were separated by SDS-PAGE and detected by autoradiography. Raf* refers to the kinase-inactive mutant of Raf-1 (Raf 301).
tivated form of RaM (Raf22W) (Fig. 3A, lanes 4 and 5). We reported however, that only a limited increase in the kinase activity of ERK1 is seen by coexpression with either Raf-1 or with v-ras (Fig. 3B, lanes 5 and 6). The kinase activity of ERK1 is greatly enhanced by coexpression with both Raf-1 and v-ras (Fig. 3B, lane 9). We have shown previous- ly that coexpression with either v-src or v-ras in- creases the kinase activity of Raf-1 [28] (see Fig. 2A, lanes b and c). Yet the kinase activity of ERK1 is greatly enhanced by coexpression with v-ras and RaM and not by coexpression with v-src and Raf-1 (Fig. 3B, compare lanes 9 and 14). Further a dom- inant-inhibitory mutant of Raf-1 (Raf301) does not block the activation of ERK1 by v-ras. These data show that v-ras and Raf-1 cooperate to activate ERK1, suggesting that activation of ERK1 by v-ras

R a f
NEK
MAPK MAPK
Fig. 4. Two models illustrating signalling pathways connecting src, ras, RaM and MAP kinase (MAPK).
occurs in one pathway via Raf-1 and in another pathway independent of RaM (Fig. 3B, lane 10). Our data further show that v-src does not activate ERK1 directly but requires intermediates such as Raf-1 or others that are missing or limiting in insect Sf9 cells. It is likely from reports by others that the activation of ERK1/MAP kinase probably occurs via endogenous SF9 MEK [66-68]. The activation of ERK1/MAP kinase via the Raf-l-independent v- ras-induced pathway, must also involve intermedi- ates that remain unidentified. Recently, ras-de- pendent protein activity named 'Ras-dependent ERK kinase stimulator', REKS, has been identified [88]. REKS also appears to activate MAP kinase by activation of MEK and awaits purification and characterization.
Putting them all together: Raf-L membrane tyrosine kinases, Ras, MEK and MAP kinases
Although several reports [66-68] as well as genetic data from Drosophila [89, 90] indicate a linear path- way from membrane tyrosine kinases to MAP ki- nase (as shown in Fig. 4A), they do not exclude the possibility of a more intricate network (as shown in Fig. 4B). There is accumulating evidence of multi- ple pathways for activation of Raf-1 and MAP ki- nases and the existence of a complex network (Fig. 4B) rather than a simple linear pathway (Fig. 4A) connecting membrane tyrosine kinases, Raf and MAP kinase. In NIH3T3 cells, ras-dependent as well as ras-independent pathways exist for the acti-
111
vation of MAP kinase. Both insulin as well as PKC activate MAP kinase. Insulin induced activation of MAP kinase is not dependent on PKC [75]. While c-ras is involved in the insulin-dependent activation of MAP kinase, it is not in the PKC-dependent pathway [75]. In T cells, activation of Raf-1 upon T cell receptor engagement occured via a PKC-de- pendent pathway [44, 91], while the PDGF-in- duced, ras-dependent activation of Raf-1 in NIH3T3 cells is resistant to PKC depletion [32]. In SF9 cells, ras-dependent as well as ras-independent pathways for activation of RaM have been demon- strated [28]. Both pathways for Raf-1 activation in SF9 cells, were found to be independent of PKC down regulation [unpublished results, Williams NG, Roberts TM] and acted synergistically to acti- vate Raf-1 [28]. A PKC-dependent activation of Raf-1 in Sf9 cells has also been demonstrated [92] and found to be independent of the ras-dependent pathway for activation of Raf-1 (unpublished re- sults, Williams NG, Roberts TM). Using the same SF9 cell system, the existence of Raf-l-dependent as well as RaM-dependent pathways for activation of MAP kinase were observed [65]. The two path- ways appear to act synergistically to activate MAP kinase. MEK, the intermediate involved in the acti- vation of MAP kinase by Raf-1, may also be re- sponsible for activation of MAP kinase by v-ras. Al- ternatively, a ras-dependent MAP kinase kinase, distinct from MEK may exist in the cell. Due to such synergistic activations, blocking of one of the path- ways for activation of Raf-1 (Fig. 4B), would abolish most of the activation, resulting in the appearance of a linear pathway (Fig. 4A) rather than complex network (Fig. 4B) [93].
During the study of Raf-1 and MAP kinase in SF9 cells, it was discovered that Raf-1 did not require the modification responsible for its retardation in gel-mobility, to activate MAP kinase [unpublished results, Williams NG, Roberts TM]. In fact, at least in this case, retardation in gel-mobility occurs as a consequence of MAP kinase activation due to a feedback loop from MAP kinase to Raf-1 (un- published results, Williams NG, Roberts TM). MAP kinases have been also reported to 'feedback' to phosphorylate MEK. It is not clear how the modifications responsible for retardation in gel-

112
mobility of Raf-1, affect activity of RaM. They may
represent activation of as yet unknown functions for Raf-1 or down-regulation of the known activity
of Raf-1. Further the extent of retardat ion in gel- mobility is seen to correspond to activation of Raf-1 only in some cases [28] and to activation of kinases other than Raf-1 in other cases (MAP kinase [65] and PKC [unpublished results]). Thus interpreta- tion of data based primarily on gel-mobility chang-
es of Raf-1 must be viewed with caution since they may not represent activation of Raf-1.
Cell line-specific differences in signal transduc- tion pathways have been demonst ra ted in several cases. Compar ison of the effects of crosslinking of
CD3 receptors of two lymphoid cell lines, Jurkat and H P B - A L L , has demonstra ted differential abil-
ities to activate M A P kinase [94]. Crosslinking of CD3 receptors leads to activation of M A P kinase in Jurkat cells but not in H P B - A L L cells. It has been suggested that absence of CD45 phosphatase activ-
ity in H P B - A L L cells, required for coupling to lck, may be responsible for the lack of ability to activate
M A P kinase in these cells upon CD3 ligation. In an- other comparison, Gallego e t al. compared the acti- vation of M A P kinase upon expression of ras, Raf-1 or gip2 oncogenes in NIH3T3 cells and Rat la cells [87]. Their results have been summarised in Table 1. They found that cell-specific differences resulted in
varying amounts of M A P kinase activation, upon oncogene expression. Relative levels of expression of oncogenes in different cell lines were not com-
pared. In Rat la cells, gip2 but not v-ras or v-raf, caused constitutive M A P kinase activation. In
Table 1. Differences between NIH3T3 cells and Rat 1A cell in their responses to transformation by the v-raf, v-ras or gip2 onco- genes. The results are a summary of data published by Gallego et
al. [861
NIH3T3 cells Rat 1A cells
V-Raf v-ras gip2 v-Raf v-ras gip2
MAP kinase activation ++ + - DNA synthesis -H- +++ - Growth in softAgar ++ t t t l - c-jun activity (AP1) + + -
_ _ +
- - + +
+ ~ - } - +
G A P GNFIF
Fig. 5. General Model for signal transduction pathways involving membrane tyrosine kinases (tyr kinases), Mitogen Activated ki- nase (MAPK), Guanine-nucleotide exchange factors (GNRF), GTPase activating protein (GAP), MAP kinase kinase (MEK), MEK kinase (MEKK), c-ras, rsk and Raf-1. Protein X represent as yet undiscovered intermediates.
NIH3T3 cells, on the other hand, v-raf or v-ras, but not gip2, induced constitutive M A P kinase activa-
tion. Further, in contrast to induction of constitu- tive M A P kinase activaty in NIH3T3 cells upon ex-
pression of the v-raf oncogene, we have shown that in PC12 cells, expression of activated raf is not suffi- cient to activate M A P kinase [34]. These cell specif- ic differences cannot be easily explained by the model representing a linear pathway f rom mem- brane tyrosine kinases to M A P kinase. A complex network, in contrast, would allow for cell specific differences based on the assumption that multiple pathways exist, for activation of ras, Raf-1 and M A P kinases. Differences in signalling pathways in dif- ferent cell lines could be explained by either 1) acti- vation due to small contributions f rom two or more synergistic pathways versus large contributions f rom a single pathway or 2) absence or inactivation of key intermediates in a cell specific manner result- ing in the lack of a specific activation pathway in that cell line.
Integrat ion of all the data results in a model as
shown in Fig. 5, wherein several synergistic or inde- pendent pathways exist for the activation of ras, Raf-1 and M A P kinase. New pathways as well as new activators continue to be discovered. M E K ki- nase (MEKK), another activator of MEK, is a 78- 80 Kd cytoplasmic serine-threonine kinase, and is distinct f rom Raf-1 [95]. M E K K appears to activate

MEK independently of R a M and may function via a G-protein coupled signalling pathway.
Key unanswered questions
Despite the major advances in signal transduction pathways involving the Raf-1 proto-oncogene, in- termediates involved in activation of Raf-1 by c-ras remain largely unknown. Though c-ras has been in- tensely investigated for more than a decade, its di- rect substrates remain unidentified. It is possible that an approach from the 'other end' could prove more fruitful. Since Raf-1 has been identified as a downstream target of ras, a search for upstream ac- tivators of Raf-1 would eventually lead to the identi- fication of ras substrates.
Note added in proof
Several recent reports suggest that Raf-1- may be a direct substrate of c-ras [96-99]. The regulatory re- gion of Raf-1 (residues 1-257), comprised of CR1, has been found to associate directly with c-ras in a GTP-dependent manner [97, 98]. Although no modulation of Raf-1 activity was reported, these da- ta provide compelling evidence implicating R a M as the first known physiological substrate for the pro- tooncogene, c-ras. However, there is no evidence to indicate that association of c-ras with the regulatory region of Raf-1 is sufficient for activation of Raf-1. Evidence suggests that phosphorylation of Raf-1 may also be necessary for full activation [28]. Moodie et al. [96], have further reported that RaM is not the only cytoplasmic kinase that associates with c-ras in a GTP-dependent manner. A MAP ki- nase kinase activity binds to c-ras-GTR even in the absence of Raf-1 [96], suggesting that c-ras may have multiple effectors.
References
1. Varmus HE: Form and function of retroviral proviruses. Sci- ence 216: 812-819,1982
113
2. Mark GE, Rapp UR: Primary structure of v-raf: relatedness to the src family of oncogenes. Science 224: 285-289, 1984
3. Rapp UR, Goldsborough MD, Mark GE, Bonner TI, Grof- fen J, Reynolds FH, Stephenson JR: Structure and biolog- ical activity of v-tar,, a unique oncogene transduced by a ret- rovirus. Proc Natl Acad Sci USA 80: 4218-4222, 1983
4. Rapp UR, Reynolds FH, Stephenson JR: New mammalian transforming retrovirus: demonstration of a polyprotein gene product. J Virol 45: 914-924, 1983
5. Moelling K, Heimann B, Beimling R Rapp UR, Sander T: Serine- and threonine-specific protein kinase activities of purified GAG-MIL and GAG-RAF proteins. Nature 312: 558-561,1984
6. Bonner TI, Kerby SB, Sutrave R Gunnell MA, Mark G, Rapp UR: Structure and biological activity of human homo- logs of the raf/rnil oncogene. Mol Cell Biol 5:1400-1407,1985
7. Bonner TI, Oppermann H, Seeburg R Kerby SB, Ur R: The complete coding sequence of the human c-raf-1 protoonco- gene and the corresponding structure of the c-tar-1 gene. Nucl Acids Res 14: 100%1015, 1986
8. Beck TW, Huleihel M, Gunnell M, Bonner T, Rapp UR: The complete coding sequence of the human A-raf-1 oncogene and transforming activity of a human A-raf carrying retro- virus. Nucl Acids Res 15: 595-609, 1987
9. Ikawa S, Fukui M, Ueyama Y, Tamaoki N, Yamamoto T, Toyoshima K: B-raf, a new member of the raf family is acti- vated by DNA rearrangement. Mol Cell Biol 8: 2651-2654, 1988
10. Sithanandam G, Kolch W, Duh FM, Rapp UR: Complete coding sequence of a human B-tar cDNA and detection of B-raf protein kinase with isozyme specific antibodies. Onco- gene 5: 1775-1780, 1990
11. Nishida Y, Hata M, Ayaki T, Ryo H, Yamagata M, Shimizu K, Nishizuka Y: Proliferation of both somatic and germ cells is affected in the Drosophila mutants of RAF proto-onco- gene. EMBO 7: 775-781, 1988
12. Mark GE, MacIntyre RJ, Digan ME, Ambrosio L, Perri- mon N: Drosophila melanogaster homologues of the RAF oncogene. Mol Cell Biol 7: 2134-2139, 1987
13. Stephens RM: 95-Kilodalton B-raf serine/threonine kinase: identification of the protein and its major autophosphoryla- tion site. Mol Cell Bio112: 3733-3742,1992
14. Storm SM, Cleveland JL, Rapp UR: Expression of raf fam- ily proto-oncogenes in normal mouse tissues. Oncogene 5: 345-351, 1990
15. Li R Wood K, Maroon H, Haser W, Roberts TM: RaM: a kinase currently without a cause but not lacking in effects. Cell 64: 479-482, 1991
16. Blenis J: Signal transduction via MAP kinases: proceed at your own RSK. in press, 1993
17. Cantley LC, Auger K, Carpenter C, Duckworth B, Graziani A, Kapeller R, Soltoff S: Oncogenes and signal transduc- tion. Cell 64: 281-302,1991
18. Pelech SL, Sanghera JA: MAP kinases: charting the regu- latory pathways. Science 257: 1355-1356, 1992
19. Stanton VR Cooper GM: Activation of human RAF trans-

114
formation genes by deletion of normal amino-terminal cod- ding sequences. Mol Cell Biol 7:1171-1179,1987
20. Tahira T, Ochiai M, Hayashi K, Nagao M, Sugimura T: Acti- vation of human c-tar-1 by replacing the N-terminal region with different sequences. Nucl Acids Res 15: 4809-4820, 1987
21. Stanton VR Nichols DW, Laudano AR Cooper GM: Defini- tion of the human RAF amino-terminal regulator region by deletion mutagenesis. Mol Cell Biol 9: 639-647, 1989
22. Ishikawa F, Sakai R, Ochiai M, Takaku F, Sigimura T, Nagao M: Identification of a transformation activity suppressing sequence in the c-RAF oncogene. Oncogene 3: 653-658, 1988
23. Heidecker G, Huleihel M, Cleveland JL, Beck R LLoyd R Pawson T, Rapp UR: Mutational activation of c-tar-1 and definition of the minimal transforming sequence. Mol Cell Biol 10: 2503-2512,1990
24. Hanks SK, Quinn AM, Hunter T: Protein kinase family: conserved features and deduced phylogeny of the catalytic domains. Science 241: 42-52, 1988
25. Shimizu K, Nakatsu Y, Sekiguchi M, Hokamura K, Tanaka K, Terada M, Sugimura T: Molecular cloning of an activated human oncogene, homologous to v-raf, from primary stom- ach cancer. Proc Natl Acad Sci 82: 5641-5645, 1985
26. Molders H, Defesche J, Muller D, Bonner TI, Rapp UR, Muller R: Integration of transfected LTR sequences into the c-raf proto-oncogene: activation by promoter insertion. EMBO 4: 693-698, 1985
27. Isumi T, Tamemoto H, Nagao M, Kadowaki T, Takaku F, Kasuga M: Insulin and platelet-derived growth factors stim- ulate phosphorylation of the c-RAF product at serine and threonine residues in intact cells. J Biol Chem 266: 7933- 7939,1991
28. Williams NG, Roberts TM, Li P: Both p21ras and pp60v-src are required, but neither alone is sufficient, to activate the Raf-1 kinase. Proc Natl Acad Sci USA 89: 2922-2926, 1992
29. Morrison DK, Heidecker G, Rapp UR, Copeland TD: Iden- tification of the major phosphorylation sites of the Raf-1 ki- nase. J Biol Chem in press, 1993
30. Smith MR, DeGudicibus S J, Stacy DW: Requirement for c- ras proteins during viral oncogene transformation. Nature 320: 540-543,1986
31. Stacey DW, Tsai MH, Yu CL, Smith JK: Critical role of cellu- lar ras proteins in proliferative signal transduction. Cold Spring Harbor Symposia on Quant. Bio153: 871-881,1988
32. Morrison DK, Kaplan DR, Rapp UR, Roberts TM: Signal transduction from membrane to cytoplasm: growth factors and membrane-bound oncogene products increase RAF-1 phosphorylation and associated protein kinase activity. Proc Natl Acad Sci USA 85: 8855-8859, 1988
33. Oshima M, Sidhanandam G, Rapp UR, Guroff G: The phos- phorylation and activation of B-raf in PC12 cells stimulated by nerve growth factor. J Biol Chem 266: 23753-23760,1991
34. Wood KW, Sarnecki C, Roberts TM, Blenis J: ras mediates nerve growth factor receptor modulation of three signal-
transducing protein kinases: MAP kinase, Raf-1, and RSK. Cell 68: 1041-1050, 1992
35. Bacharini M, Sabatini DM, App H, Rapp UR, Stanley RE: Colony stimulatin factor-1 (CSF-1) stimulates temperature dependent phosphorylation and activation of the RAF-1 proto-oncogene product. EMBO 9: 3649-3657, 1990
36. App H, Hazan R, Zilberstein A, Ullrich A, Schlessinger J, Rapp UR: Epidermal growth factor (EGF) stimulates asso- ciation and kinase activity of RAF-1 with the EGF receptor. Mol Cell Bio111: 913-919, 1991
37. Blackshear PJ, Haupt DM, App H, Rapp UR: Insulin acti- vates the RAF-1 protein kinase. J Biol Chem 265:1231-1234, 1990
38. Kovacina KS, Yonezawa K, Brautigan DL, Tonks NK, Rapp UR, Roth RA: Insulin activates the kinase activity of the RAF-1 proto-oncogene by increasing its serine phosphory- lation. J Biol Chem 265: 12115-12118,1990
39. Turner B, Rapp UR, App H, Greene M, Dobashi K, Reed J: Interleukin 2 induces tyrosine phosphorylation and activa- tion of P72-74 Raf-1 kinase in a T-cell line. Proc Natl Acad Sci USA 88: 1227-1231, 1991
40. Zmuidzinas A, Mamon HJ, Roberts TR, Smith KA: Inter- leukin-2 triggered Raf-1 expression, phosphorylation, and associated kinase activity increase through G1 and S in CD3- stimulated primary human T cells. Mol Cell Biol 11: 2794- 2803, 1991
41. Carroll MR Clark-Lewis I, Rapp UR, Stratford May W: In- terleukin-3 and granulocyte-macrophage colony-stimulat- ing factor mediate rapid phosphorylation and activation of cytosolic c-raf. J Biol Chem 265: 19812-19817, 1990
42. Carroll MP, Spivak JL, McMahon M, Weich N, Rapp UR, May WS: Erythropoietin induces Raf-1 activation and Raf-1 is required for erythropoietin-mediated proliferation. J Biol Chem 266: 1496414969, 1991
43. Kanakura Y, Druker B, Wood KW, Mamon HJ, Okuda K, Roberts TM, Griffin JD: Granulocyte-macrophage colony- stimulating factor and interleukin-3 induce rapid phospho- rylation and activation of the proto-oncogene Raf-1 in a hu- man factor dependent myeloid cell line. Blood 77: 243-248, 1991
44. Siegel JN, Klausner RD, Rapp UR, Samelson LE: T cell an- tigen receptor engagement stimulates c-RAF phosphoryla- tion and induces c-RAF associated kinase activity via a pro- tein kinase C-dependent pathway. J Biol Chem 265: 18472- 18480,1990
45. Morrison DK, Kaplan DR, Escobedo JA, Rapp UR, Ro- berts TM, Williams LT: Direct activation of the serine/threo- nine kinase activity of RAF-1 through tyrosine phosphoryla- tion by the PDGF [3-receptor. Cell 58: 649-657, 1989
46. Oshima M, Sithanandam G, Rapp UR, Guroff G: The phos- phorylation and activation of B-raf in PC12 cells stimulated by nerve growth factor. J Biol Chem 266: 23753-23760,1991
47. Ashworth A, Nakielny S, Cohen R Marshall C: The amino acid sequence of a mammalian MAP kinase kinase. Onco- gene 7: 2555-2556,1992
48. Crews CM, Alessandrini A, Erikson RL: The primary struc-

ture of MEK, a protein kinase that phosphorylates the E R K
gene product. Science 258: 478480, 1992 49. Wu J, Harrison JK, Vincent LA, Haystead C, Haystead TA,
Michel H, Hunt DE Lynch KR, Sturgill TW: Molecular structure of a protein-tyrosine kinase activating p42 mito- gen-activated protein (MAP) kinase: MAP kinase kinase. Proc Natl Acad Sci 90: 173-177, 1993
50. Macdonald SG, Crews C, Wu L, Driller J, Clark R, Erikson RL, McCormick F: Reconstitution of the RaM MEK-ERK signal transduction pathway in vitro. Mol Cell Biol submit- ted, 1993
51. Feig L: The many roads that lead to Ras. Science 260: 767- 768, 1993
52. Marshall CJ: How does p21ras transform cells? TIG 7: 91-95, 1991
553. Bourne HR, Sanders DA, McCormick F: The GTPase su- perfamily: a conserved switch for diverse cell functions. Na- ture 348: 125-132, 1990
54. Trahey M, McCormick F: A cytoplasmic protein stimulates normal N-ras p21 GTPase, but does not affect oncogenic mutants. Science 238: 542-545, 1987
55. Wolfman A, Macara IG: A cytosolic protein catalyzes the release of GDP from p21. Science 248: 247-249, 1990
56. West M, Kung HF, Kamata T: A novel membrane factor stimulates guanine nucleotide exchange reaction of ras pro- teins. FEB Lett 259: 245-248,1990
57. Buday L, Downward J: Epidermal growth factor regulates p21ras through the formation of a complex of receptor, Grb2 adapter protein, and Sos nucleotide exchange factor. Cell 73: 611, 1993
58. Buday L, Downward J: Epidermal growth factor regulates the exchange rate of guanine nucleotides on p21ras in fibro- blasts. Mol Cell Bio113:1903-1993
59. Gulbins E, Coggeshall KM, Baier G, Katzav S, Burn R Alt- man A: Tyrosine kinase-stimulated guanine nucleotide ex- change activity of Vav in T cell activation. Science 260: 822- 825, 1993
60. Medema RH, De Vries-Smits AMM, Zon GCM, Maassen JA, Bos JL: Ras activation by insulin and epidermal growth factor through enhanced exchange of guanine nucleotided on p21ras. Mol Cell Bio113: 155, 1993
61. Downward J, Graves JD, Warne PH, Rayter S, Cantrell DA: Stimulation of p21ras upon T-cell activation. Nature 346: 719-723, 1990
62. Reed JC, Yum S, Cuddy MR Turner BC, Rapp UR: Differ- ential regulation of the p72-74 RAF-1 kinase in 3T3 fibro- blasts expressing ras or src oncogenes. Cell Growth and Diff 2: 235-243, 1991
63. Doerfler W, Bohm P: The molecular biology of Baculovirus- es. In: (eds) Current Topics in Microbiology and Immunol- ogy. Springer-Verlag, 1986, pp 7-15
64. Zhang K, DeClue JE, Vass WC, Papageorge AG, McCor- mick F, Lowy DR: Suppression of c-ras transformation by GTPase-activating protein. Nature 346: 754-756, 1990
65. Williams NG, Paradis H, Agarwal S, Charest DL, Pelech SL,
115
Roberts TM: Raf-1 and p21v-ras cooperate in the activation of MAP kinase. Proc Natl Acad Sci, 1993
66. Kyriakis JM, App H, Zhang X, Banerjee R Brautigan DL, Rapp UR, Avruch J: RaM activates MAP kinase-kinase. Nature 358: 417421, 1992
67. Dent P, Haystead TA, Haser W, Vincent LA, Roberts TM, Sturgill TW: v-Raf protein kinase activates mitogen activa- ted protein (MAP) kinase kinase in NIH3T3 cells and in vit-
ro. Science 257: 14041407, 1992 68. Howe LR, Leevers S J, Gomez N, Nakielny S, Cohen R Mar-
shall CJ: Activation of the MAP kinase pathway by the pro- tein kinase raf. Cell 71: 335-342, 1992
69. Boulton TG, Cobb MH: Identification of multiple extracel- lular signal-regulated kinases (ERKs) with antipeptide anti- bodies. Cell Regul 2: 357-371, 1991
70. Cobb MH, Boulton TG, Robbins D J: Extracellular signal- regulated kinases: ERKs in progress. Cell Regul 2: 965-978, 1991
71. Pelech SL, Sanghera JS: Mitogen activated protein kinases: versitle transducers for cell signalling. TIBS 17: 233-238, 1992
72. Nakielny S, Cohen R Wu J, Sturgill T: MAP kinase activator from insulin-stimulated skeletal muscle is a protein threo- nine/tyrosine kinase. EMBO J 11: 2123-2129, 1992
73. Boulton TG, Nye SH, Robbins D J, Ip NY, Radziejewska E, Morgenbesser SD, DePinho RA, Panayotatos N, Cobb MH, Yancopoulos GD: ERKs: a family of protein-serine/threo- nine kinases that are activated by tyrosine phosphorylated in response to insulin and NGE Cell 65: 663-675, 1991
74. Robbins D J, Cheng M, Zhen E, Vanderbilt CA, Feig LA, Cobb MH: Evidence for a Ras-dependent extracellular sig- nal-regulated protein kinase (ERK) cascade. Proc Natl Acad Sci USA 89: 6924-6928, 1992
75. de Vries-Smits AMM, Burgering BM, Leevers S J, Marshall C J, Bos JL: Involvement of p21ras in activation of extracel- lular signal-regulated kinase 2. Nature 357: 602-604, 1992
76. Leevers SL, Marshall CJ: Activation of extracellular signal- regulated kinase, ERK2 by p21 ra' oncoprotein. EMBO J 11: 569-574, 1992
77. Gupta SK, Gallego C, Johnson GL, Heasley LE: MAP ki- nase is constitutively activated in gip2 and src transformed rat la fibroblasts. J Biol Chem 267: 7987-7990, 1992
78. Thomas SM, DeMarco M, D'Arcangelo G, Halegoua S, Brugge JS: Ras is essential for nerve growth factor- and phorbol ester-induced tyrosine phosphorylation of MAP ki- nases. Cell 68: 1031-1040, 1992
79. Posada J, Cooper JA: Requirements for phosphorylation of MAP kinase during meiosis in xenopus oocytes. Science 255: 212-215, 1992
80. Payne DM, Rossomando AJ, Martino R Erickson AK, Her JH, Shabanowitz J, Hunt DF, Weber MJ, Sturgill TW: Iden- tification of the regulatory phosphorylation sites in pp42/mi- togen-activated protein kinase (MAP kinase). EMBO J 10: 885-892,1991
81. G°mez N' C°hen P: Dissecti°n °f the pr°tein kinase cascade

116
by which nerve growth factor activates MAP kinases. Na- ture 353: 170-173, 1991
82. Ahn NG, Weiel JE, Chan CR Krebs EG: Identification of multiple epidermal growth factor-stimulated protein serine/ threonine kinases from swiss 3T3 cells. J Biol Chem 265: 11487-11494, 1990
83. Seger R, Ahn NG, Posada J, Munar ES, Jensen AM, Cooper JA: Purification and characterization of mitogen-activated protein kinase activator(s) from epidermal growth factor- stimulated A431 cells. J Biol Chem 267: 14373-14381, 1992
84. Matsuda S, Kosako H, Takenaka K, Moriyama K, Sakai H, Akiyama T, Gotoh Y, Nishida E: Xenopus MAP kinase acti- vator: identification and function as a key intermediate in the phosphorylation cascade. EMBO J 11: 973-982, 1992
85. Anderson NG, Li P, Marsden LA, Williams NG, Roberts TM, Sturgill TM: Raf-1 is a potential substrate for mitogen- activated protein kinase in vivo. Biochem J 277: 573-576, 1991
86. Lee R, Cobb MH, Blackshear PJ: Evidence that extracellu- lar signal-regulated kinases are the insulin-activated Raf-1 kinase kinase. J Biol Chem 267: 1088-1092, 1992
87. Gallego C, Gupta SK, Heasley LE, Qian N, Johnson GL: Mitogen-activated protein kinase activation resulting from selective onocogene expression in NIH 3T3 and Rat la cells. Proc Natl Acad Sci USA 89: 7355-7359, 1992
88. Itoh T, Kaibuchi K, Masuda T, Yamamoto T, Matsuura Y, Maeda A, Shimizu K, Takai Y: A protein factor for ras p21- dependent activation of MAP kinase through MAP kinase kinase. Proc Natl Acad Sci 90: 975-979, 1993
89. Lu X, Chou T, Williams NG, Roberts TM, Perrimon N: Con- trol of cell fate determination by p21ras/Rasl, an essential component of torso signaling in drosophila. Genes & Dev 7: 621-632, 1993
90. Dickson B, Sprenger F, Morrison D, Hafen E: Raf functions downstream of Rasl in the sevenless signal transduction pathway. Nature 360: 600-603, 1992
91. Izquierdo M, Downward J, Graves JD, Cantrell DA: Role of
protein kinase C in T-cell antigen receptor regulation of p21ras: evidence that two p21ras regulatory pathways coex- ist in T cells. Mol Cell Bio112: 3305-3312, 1992
92. Sozeri O, Vollmer K, Liyanage M, Frith D, Kour G, Mark GE, Stabel S: Activation of the c-raf protein kinase by pro- tein kinase C phosphorylation. Oncogene 7: 2259-2262, 1992
93. Roberts TM: A signal chain of events. Nature 360: 534-535, 1992
94. Nel AE, Hanekom C, Hultin L: Protein kinase C plays a role in the induction of tyrosine phosphorylation of lymphoid MAP Kinase. J Immun 147: 1933-1939, 1991
95. Lange-Carter CA, Pleiman CM, Gardner AM, Blumer KJ, Johnson GL: A divergence in the MAP kinase regulatory network defined by MEK KInase and Raf. Science 260: 315- 319, 1993
96. Moodie SA, Willumsen, BM, Weber MJ, Wolf-man A: Com- plexes of Ras.GTP with Raf-1 and Mitogen-Activated Pro- tein Kinase Kinase. Science 260: 1658-1661, 1993
97. Zhang X-F, Settleman J, Kyriakis JM, Takeuchi-Suzuki E, Elledge S J, Marshall MS, Bruder JT, Rapp UR, Avruch J: Normal and oncogenic p21ras proteins bind to the amino- terminal regulatory domain of c-Raf-1. Nature 364: 308-313, 1993
98. Warne PH, Viciana PR, Downward J: Direct interaction of Ras and the amino-terminal region of RaM in vitro. Nature 364: 352-355, 1993
99. Hughes DA, Ashworth A, Marshall C J: Complementation of byrl in fission yeast by mammalian MAP kinase kinase requires coexpression of Raf kinase. Nature 364: 349-352, 1993
Address for offprints: T.M. Roberts, Dana Farber Cancer Institute, Mayer 856, 44 Binney Street, Boston, MA 02115, USA




















