
Gα13 Regulation of Proto-Dbl Signaling
13
©2007 LANDES BIOSCIENCE. DO NOT DISTRIBUTE. 2058 Cell Cycle 2007; Vol. 6 Issue 16 [Cell Cycle 6:16, 2058-2070, 15 August 2007]; ©2007 Landes Bioscience Report Gα 13 Regulation of Proto-Dbl Signaling Cristina Vanni 1 Patrizia Mancini 2 Catherine Ottaviano 1 Marzia Ognibene 1 Alessia Parodi 1,3 Elisa Merello 4 Chiara Russo 1,5 Luigi Varesio 1 Yi Zheng 6 Maria Rosaria Torrisi 2,7 Alessandra Eva 1,8, * 1 Laboratorio di Biologia Molecolare; Istituto G. Gaslini; Genova, Italy 2 Dipartimento di Medicina Sperimentale; Università di Roma ‘La Sapienza’; Roma, Italy 4 Laboratorio di Neurochirurgia; Istituto G. Gaslini; Genova, Italy 6 Division of Experimental Hematology; Children’s Hospital Research Foundation; Cincinnati, Ohio 7 Azienda Ospedaliera Sant’Andrea; Roma, Italy 3 Current address: Centro Cellule Staminali; Azienda Ospedaliera Universitaria S. Martino, Genova 5 Current address: Institut für Zellbiologie und Immunologie; Universität Stuttgart; Stuttgart, Deutschland *Correspondence to: Alessandra Eva; Laboratorio di Biologia Molecolare; Istituto G. Gaslini, Largo Gaslini 5, Genova 16147 Italy; Tel.: 001.39.010.5636633; Fax: 001.39.010. 3733346; Email: [email protected] Original manuscript submitted: 01/17/07 Revised manuscript submitted: 06/08/07 Manuscript accepted: 06/08/07 Previously published online as a Cell Cycle E-publication: http://www.landesbioscience.com/journals/cc/article/4574 KEY WORDS Gα 13 , proto-Dbl, ezrin, GPCR-agonists, GTPases ACKNOWLEDGEMENTS See page 2067. ABSTRACT Rho family GTPases play important roles in the regulation of intracellular signals induced by activated heterotrimeric G proteins of the α 12/13 family. The α 12/13 subunits activate Rho GTPases through direct binding to a group of Rho guanine nucleotide exchange factors (GEFs) characterized by the presence of a G protein signaling‑like (RGL) domain. The Rho GEF proto‑Dbl, that does not contain a RGL domain, was also found to link Gα 12/13 signals to Rho. We have explored the effects of activated Gα 13 and Gα 13 ‑associated G protein‑coupled receptor (GPCR) agonists on proto‑Dbl regulation. We show that activated Gα 13 , but not Gα 12 or Gα q , induces translocation of proto‑Dbl to the cell membrane with consequent enlargement of cell body and membrane ruffling. These effects were evident also when Gα 13 ‑associated GPCR agonists were used on cells expressing proto‑Dbl and were accompanied by the activation of Cdc42 and RhoA GTPases and further downstream effector JNK and p38 kinases. Moreover, we show that both activated Gα 13 and GPCR agonists stimulate proto‑Dbl interaction with ezrin to promote ezrin translocation to the plasma membrane. These results suggest a mechanism by which proto‑Dbl and its effector pathways are regulated by Gα 13 ‑mediated signals through association with ezrin. INTRODUCTION The heterotrimeric G proteins mediate the transduction of signals from GPCRs to numerous intracellular signaling pathways. Constitutively active forms of both Gα 12 and Gα 13 can stimulate cell proliferation and malignant transformation. 1-4 Gα 12 and Gα 13 modulate several cellular responses: they can activate the Na + /H + exchanger, 5 the c-Jun NH2-terminal kinase (JNK), 6 the extracellular signal-regulated kinase, 7 tyrosine kinases, 8 stress fiber and focal adhesion formation, 9,10 neurite retraction 11,12 and apop- tosis. 13,14 Activated forms of these G proteins can interact with several effectors including glutamate transporter EAAT4-associated protein (GTRAP48), 15 cortactin-interactin protein (Hax-1), 16 cell adhesion molecules, 17,18 PLC-e, 19 A-kinase anchoring protein 110, 20 heat shock protein 90 (HSP90), 21 protein phosphatase 5 22 and radixin. 23 One of the recognized mechanism by which Gα 12 and Gα 13 modulate various cellular responses such as cytoskeletal changes, cell growth and differentiation is through regulating the Rho GTPase activity by interaction with a subfamily of the Rho GEFs characterized by the presence of a RGL domain. The RGL domain containing Rho GEFs include p115 RhoGEF, 24 PDZ-RhoGEF 25 and the leukemia associated RhoGEF (LARG). 26 The RGL domain of these GEFs is directly recognized by receptor-stimulated Gα 12/13 through specific structural features present in Gα 13 and is recruited to the plasma membrane to cause Rho GTPase activation, eliciting cellular functions like cell growth, cell shape, cell movement and gene expression. 27 Another Rho GEF containing a putative RGL domain, Lbc-RhoGEF, was found to link Gα 12/13 signals to Rho. 28 The Rho GEFs constitute one of the largest known groups of transforming proteins and their transforming potential directly correlates with their Rho GTPase activating potential. These proteins seem to exist as inactive or partially active molecules until specific intra- cellular stimuli lead to their activation. The GEF protein function may thus be modulated by diverse regulatory mechanisms which involve intra- and/or intermolecular interactions between different protein domains, oligomerization, phosphorylation and interaction with different intracellular signaling molecules such as PIPs, heterotrimeric G-protein subunits and/or scaffolding proteins. 29,30 The prototype member of Rho GEF family is the proto-Dbl protein, a cytoplasmic phosphoprotein that contains a N-terminal region
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Gα13 Regulation of Proto-Dbl Signaling

©2007 L
ANDES BIOSCI
ENCE.
DO NOT DIST
RIBUTE.
2058 CellCycle 2007;Vol.6Issue16
[CellCycle6:16,2058-2070,15August2007];©2007LandesBioscience
Report
Gα13 Regulation of Proto-Dbl Signaling
Cristina Vanni1
Patrizia Mancini2
Catherine Ottaviano1
Marzia Ognibene1
Alessia Parodi1,3
Elisa Merello4
Chiara Russo1,5
Luigi Varesio1
Yi Zheng6
Maria Rosaria Torrisi2,7
Alessandra Eva1,8,*1Laboratorio di Biologia Molecolare; Istituto G. Gaslini; Genova, Italy
2Dipartimento di Medicina Sperimentale; Università di Roma ‘La Sapienza’; Roma, Italy
4Laboratorio di Neurochirurgia; Istituto G. Gaslini; Genova, Italy
6Division of Experimental Hematology; Children’s Hospital Research Foundation; Cincinnati, Ohio
7Azienda Ospedaliera Sant’Andrea; Roma, Italy
3Current address: Centro Cellule Staminali; Azienda Ospedaliera Universitaria S. Martino, Genova
5Current address: Institut für Zellbiologie und Immunologie; Universität Stuttgart; Stuttgart, Deutschland
*Correspondence to: Alessandra Eva; Laboratorio di Biologia Molecolare; Istituto G. Gaslini, Largo Gaslini 5, Genova 16147 Italy; Tel.: 001.39.010.5636633; Fax: 001.39.010. 3733346; Email: [email protected]
Original manuscript submitted: 01/17/07Revised manuscript submitted: 06/08/07Manuscript accepted: 06/08/07
Previously published online as a Cell Cycle E-publication:http://www.landesbioscience.com/journals/cc/article/4574
KEY wORds
Gα13, proto-Dbl, ezrin, GPCR-agonists,GTPases
ACKnOwLEdgEMEnTs
Seepage2067.
AbsTRACTRho family GTPases play important roles in the regulation of intracellular signals
induced by activated heterotrimeric G proteins of the α12/13 family. The α12/13 subunits activate Rho GTPases through direct binding to a group of Rho guanine nucleotide exchange factors (GEFs) characterized by the presence of a G protein signaling‑like (RGL) domain. The Rho GEF proto‑Dbl, that does not contain a RGL domain, was also found to link Gα12/13 signals to Rho. We have explored the effects of activated Gα13 and Gα13‑associated G protein‑coupled receptor (GPCR) agonists on proto‑Dbl regulation. We show that activated Gα13, but not Gα12 or Gαq, induces translocation of proto‑Dbl to the cell membrane with consequent enlargement of cell body and membrane ruffling. These effects were evident also when Gα13‑associated GPCR agonists were used on cells expressing proto‑Dbl and were accompanied by the activation of Cdc42 and RhoA GTPases and further downstream effector JNK and p38 kinases. Moreover, we show that both activated Gα13 and GPCR agonists stimulate proto‑Dbl interaction with ezrin to promote ezrin translocation to the plasma membrane. These results suggest a mechanism by which proto‑Dbl and its effector pathways are regulated by Gα13‑mediated signals through association with ezrin.
InTROduCTIOnThe heterotrimeric G proteins mediate the transduction of signals from GPCRs to
numerous intracellular signaling pathways. Constitutively active forms of both Gα12and Gα13 can stimulate cell proliferation and malignant transformation.1-4 Gα12 andGα13modulate severalcellular responses: theycanactivate theNa+/H+exchanger,5 thec-Jun NH2-terminal kinase (JNK),6 the extracellular signal-regulated kinase,7 tyrosinekinases,8 stress fiberand focaladhesion formation,9,10neurite retraction11,12andapop-tosis.13,14ActivatedformsoftheseGproteinscaninteractwithseveraleffectorsincludingglutamate transporter EAAT4-associated protein (GTRAP48),15 cortactin-interactinprotein (Hax-1),16 cell adhesion molecules,17,18 PLC-e,19 A-kinase anchoring protein110,20heatshockprotein90(HSP90),21proteinphosphatase522andradixin.23
OneoftherecognizedmechanismbywhichGα12andGα13modulatevariouscellularresponsessuchascytoskeletalchanges,cellgrowthanddifferentiationisthroughregulatingtheRhoGTPaseactivitybyinteractionwithasubfamilyoftheRhoGEFscharacterizedbythepresenceofaRGLdomain.TheRGLdomaincontainingRhoGEFsincludep115RhoGEF,24PDZ-RhoGEF25andtheleukemiaassociatedRhoGEF(LARG).26TheRGLdomain of these GEFs is directly recognized by receptor-stimulated Gα12/13 throughspecificstructuralfeaturespresentinGα13andisrecruitedtotheplasmamembranetocauseRhoGTPaseactivation,elicitingcellularfunctionslikecellgrowth,cellshape,cellmovementandgeneexpression.27AnotherRhoGEFcontainingaputativeRGLdomain,Lbc-RhoGEF,wasfoundtolinkGα12/13signalstoRho.28
TheRhoGEFsconstituteoneofthelargestknowngroupsoftransformingproteinsandtheirtransformingpotentialdirectlycorrelateswiththeirRhoGTPaseactivatingpotential.Theseproteinsseemtoexistasinactiveorpartiallyactivemoleculesuntilspecificintra-cellularstimulileadtotheiractivation.TheGEFproteinfunctionmaythusbemodulatedbydiverseregulatorymechanismswhichinvolveintra-and/orintermolecularinteractionsbetween different protein domains, oligomerization, phosphorylation and interactionwith different intracellular signaling molecules such as PIPs, heterotrimeric G-proteinsubunitsand/orscaffoldingproteins.29,30TheprototypememberofRhoGEFfamilyistheproto-Dblprotein,acytoplasmicphosphoproteinthatcontainsaN-terminalregion

www.landesbioscience.com CellCycle 2059
thatnegativelyregulatesitsactivity.Whileproto-Dblitselfpossesseslimited transforming activity, truncation of the N-terminal regionappearstounleashpotenttransformingactivity.31-33Despitethefactthat proto-Dbl does not contain a detectable RGL domain in itsstructure,itsassociationwithactivatedGα13andsubsequentactiva-tionofRhoAhasbeenreported.34
In an attempt to identify and characterize the mechanisms ofactivation of proto-Dbl we have analyzed the effects of activatedGα13andGα13-associatedGPCRagonistsonproto-Dblactivationby examining its subcellular localization in addition to its GEFactivity, since Rho GEFs may also function as docking sites forproteinsthatareassociatedwithsignalingcascadesthatareregulatedbyRhoGTPases.Inthisregardswehaverecentlydemonstratedthatonco-Dbldirectly interactswithezrin,35oneof the threemembersoftheERMfamilyofproteinsthatlinktheplasmamembranewiththe actin cytoskeleton and that are activated by Rho GTPases.36Therefore,wealsoexaminedhowtheexpressionofactivatedGα13ortreatmentofproto-DblexpressingcellswithGα13agonistswouldaffectassociationwithezrin.WereportherethatconstitutivelyactiveGα13 or Gα13-associated GPCR agonists induce proto-Dbl trans-location to the plasma membrane and stimulate its GEF activity.Furthermore,activeGα13ortreatmentwithGα13-associatedGPCRagonists is able to stimulate proto-Dbl protein binding to ezrin atthe plasma membrane. These results indicate that proto-Dbl maybeactivatedbyGα13throughamechanismthatinvolvesassociationwithezrin.
MATERIALs And METhOdsReagents and plasmids. Anti-Dbl antibodies were previously
described.37 Polyclonal antibodies anti-phospho-p38, and -phos-pho-JNKwerepurchasedfromNewEnglandBiolabs(Ipswich,MA).Polyclonalantibodiesagainstp38,JNK1andCdc42,andmonoclonalantibodyanti-RhoAwerepurchasedfromSantaCruzBiotechnology(SantaCruz,CA).MonoclonalantibodyagainstRacwaspurchasedfrom Upstate Biotecnology (Lake Placid, NY). Anti-GST waspurchased from Molecular Probes (Eugene, OR). Anti-His wasfrom Invitrogen (Carlsbad, CA). Monoclonal antibody againstFLAGepitopewaspurchasedfromSigma-Aldrich(St. Louis, MO).St.Louis,MO)..pCEFL-GST-protoDbl construct was previously described.38 LPA,thrombin, bradykinin, polibrene and puromycin were purchasedfrom Sigma-Aldrich (St. Louis, MO). G418 was from Invitrogen(Carlsbad, CA). pCDNA3.1-α13wt and pCDNA3.1-α13Q226L,pCDNA3.1-α12Q231L and pCDNA3.1-αqQ209L were obtainedfromGuthriecDNAResourceCenter(Sayre,PA)andsubclonedintoEcoRIandXbaIcloningsitesofthepFLAGvector(Sigma-Aldrich,St.Louis,MO).pEF1B-ezrinwtconstructwaspreviouslydescribed.35
Cell culture and transfection.NIH3T3fibroblastswereculturedin Dulbecco’s™ modified Eagle medium (DMEM) supplementedwith 10% calf serum. Mass cultures of stable transfected cell lineswere generatedby transfectingNIH3T3cellswith300ngof eachplasmidDNAbythecalciumphosphatecoprecipitationmethodandculturing them inDMEMsupplementedwith5%calf serumand375mg/mlofG418.COS7cellswerecultured inDMEMsupple-mentedwith10%fetalcalfserum.Forkinaseassaysandpulldownexperimentscellsweregrownto80%confluencein100-mmtissueculturedishesandtransientlytransfectedwith4mgoftheindicatedplasmidsusingLipofectAMINEPLUSasdescribedbythemanufac-turer(Invitrogen,Carlsbad,CA).TwentyhoursaftertransfectionthemediumwaschangedtoDMEMcontaining0.5%fetalcalfserum,
andthecellswere incubated foranother18hoursbefore lysis.Forstimulation with GPCR agonists, twenty hours after transfectioncellswerestarvedin0.5%calfserumfor18hoursthentreatedwith2U/mlthrombinfor10min,10-6Mbradykininfor10min,or5mMLPAfor5minbeforelysis.
In vivo Rho GTPases activation assay. The GST-PAK-CRIBdomainfusionprotein(residues56–141)containingtheCdc42andRacbindingregionofhumanPAK1,wasexpressedandpurifiedasdescribedpreviously.39ToevaluateCdc42andRacactivation,COS7cellsweretransfectedwithpCEFL-GSTorpCEFL-GSTproto-Dblalone or cotransfected with pFLAG-α13wt or pFLAG-α13Q226L.TwentyhoursaftertransfectionthemediumwaschangedtoDMEMcontaining0.5% fetal calf serum, and the cellswere incubated foranother 18 hours before lysis or treated with 10-6 M bradykininfor 10 min or 5 mM LPA for 5 min. The transfected cells werewashedwith ice-coldPBSbuffer and lysedon thedish inabuffercontaining 50 mM Tris-HCl (pH 7.4), 100 mM NaCl, 2 mMMgCl2,1%NonidetP-40,10%glycerol,10mg/mleachofaprotininand leupeptin, 2 mM 4-(2-aminoethyl)benzenesulfonyl fluoride(AEBSF)and40mgofGST-PAK.Lysateswereincubatedwith60mlof glutathione-coupled Sepharose beads (Amersham PharmaciaBiotech,Uppsala,Sweden)for1hourat4˚C.Thebeadswerethenwashed three times with lysis buffer, eluted in Laemmli samplebuffer,subjectedtoSDS-PAGE,andtransferredtoPVDFmembrane(Millipore,Billerica, MA) for immunoblotting. Bound Cdc42 andBillerica,MA) for immunoblotting. Bound Cdc42 and) for immunoblotting.BoundCdc42 andRac were detected in Western Blot by using specific antibodiesagainst Rac and Cdc42. Activated RhoA was detected by using aG-Lisa RhoA Activation assay (Absorbance based) (CytoskeletonInc.,Denver,CO)asindicatedbythemanufacturer.AliquotsofcelllysatesweresubjectedtoSDSPAGEandimmunoblottingtoevaluateproto-Dbl and RhoA expression levels.The optical density of thefilm was scanned and measured with Quantity One v. 2-3 Imagesoftware(VersaDoc,Biorad, Hercules, CA).Hercules,CA).).
Assay for activation of p38 MAPK and JNK.Toassessthelevelsof activated p38 MAPK and JNK, NIH3T3 cells were transfectedwith pCEFL-GST or pCEFL-GST-proto-Dbl. Twenty hours aftertransfectionthemediumwaschangedtoDMEMcontaining0.5%fetalcalf serum,andthecellswere incubated foranother18hoursand then treated with 10-6 M bradykinin for 10 min before lysis.Thetransfectedcell lineswerelysedinabuffercontaining20mMTris-HCl,150mMNaCl,1mMEGTA,1mMEDTA,1%TritonX-100, 1 mM b-glycerolphosphate, 1 mM Na3VO4, 2.5 mMSodium pyrophosphate, 0.5 mM NaF, 1 mM AEBSF, 10 mg/mleachofaprotininandleupeptin.WholecelllysatesweresubjectedtoSDS-PAGE,transferredtoPVDFmembraneandprobedwithphos-pho-specificantibodiestodetectP-p38andP-JNKandanti-p38oranti-JNK to detect kinase expression levels.The optical density ofthefilmwasscannedandmeasuredwithQuantityOnev.2-3Imagesoftware(VersaDoc,Biorad,Hercules,CA).
Immunofluorescence. NIH3T3 cells stable transfected withpCEFL-GST, pCEFL-GST proto-Dbl, pFLAG-α13wt, pFLAG-α13Q226L, pFLAG-α12Q231L, and pFLAG-αqQ209L andpEF1B-ezrinwtwereplatedontoglasscoverslips,previouslycoatedwith1mg/mlfibronectin(Sigma-Aldrich,St.Louis,MO),andtreatedwith10-6Mbradykininfor10minat37˚C,2U/mlthrombinfor10min,5mMLPAfor5min,100ng/mlEGFfor10min,fixedwith4%paraformaldehydeinPBSfor30minat25˚C,andpermeabilizedwith0.1%TritonX-100for5min.Fortripleimmunofluorescencelabeling, cells were incubated with anti-GST polyclonal antibodiesfordetectionoftheproto-Dblprotein,followedbyFITC-conjugated
Gα13RegulationofProto-DblSignaling

2060 CellCycle 2007;Vol.6Issue16
goat-anti rabbit IgG (Cappel, Organon Teknica Corp., Durham, Durham,NC), with anti-FLAG monoclonal antibody (Sigma-Aldrich,), with anti-FLAG monoclonal antibody (Sigma-Aldrich,St.Louis,MO)fordetectionofthesubunitofGprotein,α12,α13or αq, followed by Alexa Fluor 350-conjugated goat anti-mouseIgG (Molecular Probes, Eugene, OR), and filamentous actin wasvisualized by incubating the cells with TRITC-labeled phalloidin(Sigma-Aldrich, St. Louis, MO) in PBS for 30 min at 25˚C. Forconfocal analysis, cells were incubated with anti-GST polyclonalantibodies for detection of the proto-Dbl protein, followed byFITC-conjugatedgoat-antirabbitIgG,andwithanti-Hismonoclonalantibody(Invitrogen,Carlsbad,CA)fordetectionoftheezrinprotein,followed by FITC-conjugated goat-anti mouse IgG. Filamentousactin was visualized by incubating the cells with TRITC-labeledphalloidin in PBS for 30 min at 25˚C. Fluorescence signals wereanalyzed either by recording stained images using a cooled CCDcolordigitalcameraSPOT-2(DiagnosticInstrumentsIncorporated,SterlingHeights,MI)andFISH2000/H1software(DeltaSistemi,Roma,Italy)orbyconfocalvertical(x-z)sections(interval:0.5mm)obtainedwithaZeissConfocalLaserScanMicroscopeLSM5Pascal(Zeiss,Oberkochen,Germany).QuantitativeanalysisoffluorescencesignalonthecellplasmamembranewasperformedusingtheZeissKS300,3.0ImageProcessingsystem,evaluatingtheratiobetweenthelengthofthefluorescencesignalofproto-Dblorezrinalongtheplasmamembraneandthetotallengthoftheplasmamembraneineachcell.Thepercentageof fluorescentmembranewasassessedbycounting a minimum of 50 cells randomly taken from each slidefromthreeindependentexperiments.Dataareexpressedasmeanofpercentage±SD.
In vivo binding assay.COS7cellsweretransfectedwithplasmidsencodingforpCEFL-GSTorpCEFL-GST-proto-Dbl.Twentyhoursafter transfection the medium was changed to DMEM containing0.5% fetal calf serum, and the cells were incubated for another18 hours before stimulation with GPCR agonists and lysis. Cellswere washed with ice-cold phosphate-buffered saline and lysed ina buffer containing 20 mM Hepes, 1% NP-40, 150 mM NaCl,1 mM Na3VO4, 1 mM EDTA, 1 mM AEBSF, 10mg/ml each ofleupeptinandaprotinin.Lysateswereclarifiedbycentrifugationandequivalentamountsofcellularproteinswereincubatedfor1hat4˚Cwithanti-ezrin(SantaCruzBiotechnology,SantaCruz,CA)underconstant rotation. Protein G-sepharose (30ml) was added to eachsample for 30 min. Immunoprecipitates were washed three timeswith lysisbuffer, eluted in30ml of1xLaemmli’sbuffer, subjectedto8%SDS-PAGEandtransferredtoPVDFmembranefor immu-noblotting.
Small interfering RNA. The expression arrest mouse retroviralshRNAmir-Gα13 (shRNA-Gα13) construct and the shRNA non-silencing control plasmids were purchased from Open Biosystems(Huntsville, AL). Recombinant retroviruses were produced usingecotropic Phoenix packaging cell system.61,62 NIH3T3 proto-Dbltransfectantswereinfectedwiththeretrovirusesinthepresenceof7mg/mlofpolibrene.Thirty-sixhourslatercellsweresplitandgrowninDulbecco’smodifiedEagle’smediumcontaining10%calfserum,375mg/mlG418and1mg/mlpuromycinfor8-15days.Theinfectedcells were then incubated in DMEM containing 0.5% fetal calfserumfor18hrs,treatedwith10-6Mbradykininfor10minandusedforimmunofluorescenceanalysisorlysedandassayedforactivationofp38MAPKandJNK.
Gα13RegulationofProto-DblSignaling
REsuLTsActivated Gα13 induces translocation of proto‑Dbl to the
plasma membrane. Itwas previously reported that activatedGα13andproto-Dbl can associate in vivo and that this interaction leadsto Rho activation.34 To further delineate the outcome of thisinteraction constitutively activated Gα13 (pFLAG-Gα13Q226L)was co-expressed with pCEFL-GST-proto-Dbl and the subcel-lular distribution of proto-Dbl was examined by triple labelingthe cells with anti-GST antibodies and FITC-labeled secondaryantibodies, for proto-Dbl detection, anti FLAG antibody andAlexa Fluor 350-conjugated secondary antibody to detect Gα13,and TRITC-labeled phalloidin, to detect actin. As previouslydescribed,33,38cellsexpressingtheproto-Dblproteinshowadiffusecytosolic pattern with a predominant perinuclear distribution anda limited protein localization at the plasma membrane, displayingan elongated shape with only slight enlargement of the cell body(Fig.1A).Ontheotherhand,coexpressionofpFLAG-Gα13Q226Linducesevidentcellbodyenlargementandanincreaseintheexten-sionoftheruffles,withaparallelincreasedlocalizationofproto-Dblalongtheplasmamembraneattherufflingsites(Fig.1A,arrowsininset), in agreement with the knowledge that activated proto-Dbltranslocates to the plasma membrane where it exerts its activity.Cells expressing pFLAG-Gα13Q226L, pFLAG-Gα12Q231L orpFLAG-GαqQ209Lalonedidnotshowanymorphologicalchangeincomparisonwithuntransfectedcellsorcells transfectedwiththeemptyvector(Fig.1Aanddatanotshown).Similarly,constitutivelyactivatedGα12(pFLAG-Gα12Q231L)orGαq(pFLAG-GαqQ209L)didnotleadtoanyeffectonproto-Dbl.IncellscotransfectedwithpFLAG-Gα12Q231L or pFLAG-GαqQ209L, proto-Dbl transloca-tion to the plasma membrane and morphological effects were notobservedincomparisonwithcellsexpressingproto-Dblalone(Fig.1A).The extent of the increased localization of proto-Dbl on the cellplasmamembraneisshowninFigure1Bandwasevaluatedbyquan-titative analysis of the fluorescence signal of GST using the ZeissKS300,3.0ImageProcessingsystemasdescribedinMaterialsandMethods.ThesedataprovideevidencethatactivatedGα13canleadtoproto-Dbltranslocationandpossiblyactivation.
Translocation of proto‑Dbl to the plasma membrane in cells expressing activated Gα13 is accompanied by activation of Cdc42, Rac and RhoA.ToevaluatehowtheGEFactivityofproto-DblwasaffectedbyactivatedGα13,COS7cellsweretransientlytransfectedwith pCEFL-GST, pFLAG-Gα13Q226L, pCEFL-GST-proto-Dblalone or cotransfected with pFLAG-Gα13Q226L, and the endog-enous,activatedCdc42andRacwerecollectedonGST-PAK-CRIBdomainfusionproteinandanalyzedbyWesternblot.ThelevelsofactivatedCdc42andRacincellsexpressingthecontrolvectorweresimilar to those observed in cells expressing the G protein alone.As shown inFigure2A expressionofproto-Dbl induces activationof Cdc42 and Rac by 4 and 3 fold, respectively. Cotransfectionof pCEFL-GST-proto-Dbl with pFLAG-Gα13Q226L increasedthe Cdc42-GTP and Rac-GTP content by 8 and 6 folds, respec-tively, in comparison with untransfected cells. We also evaluatedactiveRhoA-GTPbyanabsorbancebasedmethodand found thatcotransfection of proto-Dbl with pFLAG-Gα13Q226L increasesRhoA activation by ~2-fold (Fig. 2), in agreement with previouslyreporteddata.34ThelevelsofactivatedCdc42,Racand/orRhoAincells expressing proto-Dbl together with Gα13wt were comparableto thoseobserved incells expressing theGproteinalone (datanotshown).TheseresultsprovidestrongevidencethatactivationofGα13leadstoanincreaseofproto-DblGEFactivity.

www.landesbioscience.com CellCycle 2061
Figure 1. Activated Gα13 induces translocation of proto‑Dbl to the plas‑ma membrane. (A) Untransfected NIH3T3 cells (C1 and C2) and NIH3T3 cells transfected with pCEFL‑GST, pFLAG, pCEFL‑GST‑pro‑to‑Dbl or cotransfected with pCE‑FL‑GST‑proto‑Dbl and activated pFLAG‑Gα13, pFLAG‑Gα12, or pFLAG‑Gαq, were analyzed by tri‑ple labeling for GST (green), FLAG (blue) and actin (red) as described in Materials and Methods. The pro‑to‑Dbl protein shows a limited local‑ization along the plasma membrane (arrows in inset), and an increased localization along the plasma mem‑brane when cotransfected with the activated α13 subunit of the G pro‑tein (arrows in inset), where the two proteins colocalize (triple). Cells coexpressing the proto‑Dbl and α12 or αq proteins do not lead to any effect on localization of proto‑Dbl along the plasma membrane. The inset on the right of each panel represents an enlargement of the selected area (dashed line). Bar, 10 mm. (B) Untransfected NIH3T3 cells (C1) or NIH3T3 cells trans‑fected with pCEFL‑GST, with pCE‑FL‑GST proto‑Dbl (NT), or cotrans‑fected with pCEFL‑GST proto‑Dbl and activated pFLAG‑Gα13 (Gα13), pFLAG‑Gα12 (Gα12) or pFLAG‑Gαq (Gαq) were analyzed for the extent of the GST‑proto‑Dbl fluorescence signal localized on the membrane. Quantitative immunofluorescence analysis was performed evaluating a minimum of 50 cells randomly taken from each slide and shown as percentage of length of the fluores‑cence signal over the total length of the plasma membrane in each cell. Results represent the mean values ± sd from three different experiments.
Gα13RegulationofProto-DblSignaling

2062 CellCycle 2007;Vol.6Issue16
did not induce activation of Cdc42. The differences observed inGTPasesactivationwerenotduetodifferencesinproto-Dblexpres-sion levels, since no significant variation in protein amounts weredetected by immunoblotting with specific anti-Dbl antibody. LPAisastimulatorofRho,notCdc42.Thustheresultsweobtainedarecompatiblewithactivationofmultipledownstreamtargetsdependingonwhichagonistisused.WealsoevaluatedactiveRhoA-GTPbyanabsorbance based method and found that treatment of proto-DblexpressingcellswithbradykininincreasesRhoAactivationby~2-foldwhilethetreatmentwithLPAincreasesRhoAactivationby1.5-fold.Both agonists were not able to activate RhoA in mock transfectedcells above background level (Fig. 4B). Thus the results obtainedindicatethatwhileconstitutivelyactivatedGα13canbeastimulatorofbothGTPases,Gα13agonistscanefficientlystimulateproto-DblGEF activity but the activation of specific downstream targets isdependentonwhichagonistisused.
Bradykinin induces activation of JNK and p38 in proto‑Dbl expressing cells. Rho family GTPases exert their effect at least inpartthroughtheactivationofserine-threoninekinasesignalingpath-ways. It has been shown that proto-Dbl induces JNK activity38,40via activation of Cdc42. To explore how GPCR agonists affectproto-Dbl-induced serine-threonine kinase activation, we tran-siently transfected COS7 cells with pCEFL-GST or pCEFL-GSTproto-Dbl.Cellswereharvested18hafterincubationinserum-freemedium followed by treatment for 10 min with bradykinin. AsshowninFigure5,thetreatmentofproto-Dbltransfectedcellswithbradykinin increased JNK (Fig. 5A) and p38 (Fig. 5B) activity by~5- (Fig. 5A’) and ~4-fold (Fig. 5B’), respectively, compared withuntreatedproto-Dblexpressingcells.Bradykininalsoinducedactiva-tionofJNKandp38inpCEFL-GSTtransfectedcellsbuttheoverallactivationofbothkinaseswasconsiderablylowerthanthatdetected
Gα13RegulationofProto-DblSignaling
Figure 2. Activation of small G proteins in cells expressing proto‑Dbl and activated Gα13. (A) Untrasfected COS7 cells (NT) or COS7 cells transiently transfected with pCEFL‑GST, pFLAG‑Gα13QL, pCEFL GST‑proto‑Dbl or cotransfected with pCEFL GST‑proto‑Dbl and pFLAG‑Gα13QL were lysed. Cell lysates were subjected to GST‑PAK pull‑down assay and anti‑Cdc42 or anti‑Rac Western blot analysis was performed. Protein expression level was determined by Western blot analysis with specific antibodies using total cell lysates. Relative amounts of GTP‑bound form of the GTPases were quantified by densitometry and normalized to those of the untransfected, untreated cells (NT) and to the total Cdc42 or Rac and proto‑Dbl content in cell extracts. The results shown are representative of 3 independent experiments. (B) Cell lysates were subjected to an absorbance based activation assay for RhoA. Protein expression level was evaluated by Western blot analysis with specific antibodies using total cell lysates. Results represent the mean values ± sd from three different experiments.
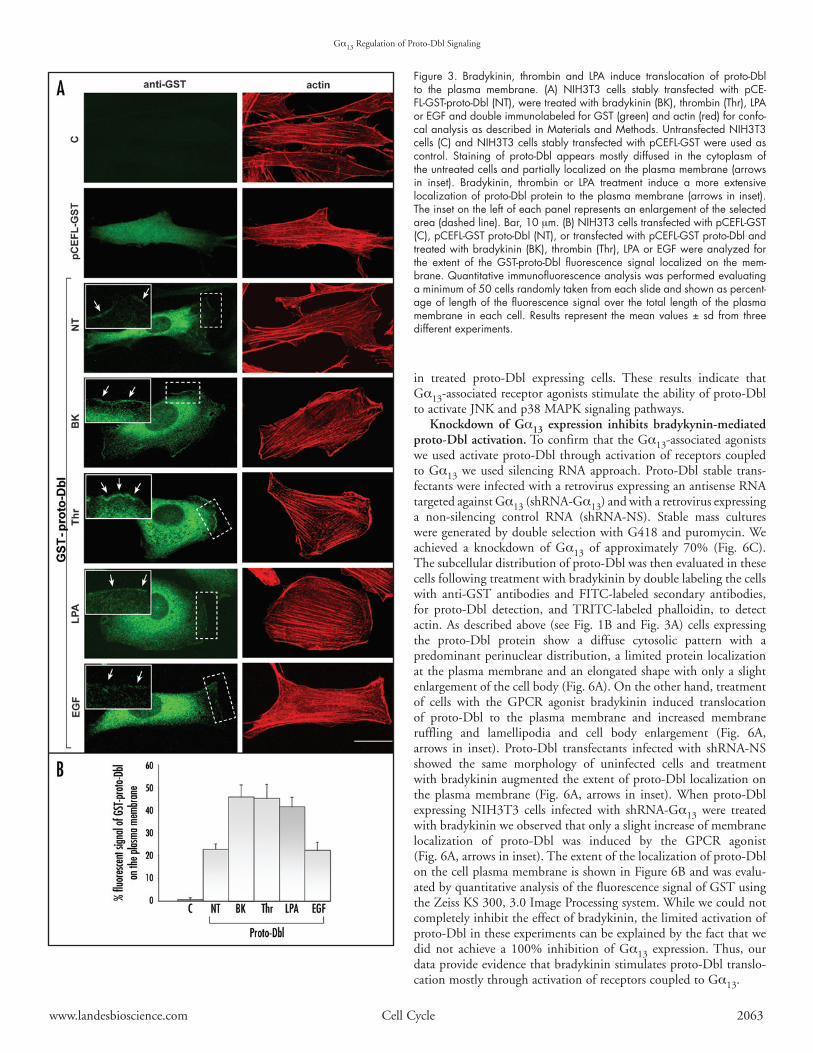
Gα13‑associated receptor agonists induce proto‑Dbl transloca‑tion to the plasma membrane. Activation of Gα13 is mediatedby several receptors for various ligands. To further evaluate therole of Gα13 in Dbl-induced Rho-GTPases activation we treatedproto-Dbl-expressing cells with three GPCR agonists, namelybradykinin, thrombin, and LPA, and evaluated proto-Dbl activa-tion by laser scanning confocal microscopy of cells double labeledwith anti-GST antibodies and FITC-labeled secondary antibodiesfor proto-Dbl detection, and TRITC-phalloidin, to detect actin.As shown in Figure 3A, treatment of cells with GPCR agonistsinduced translocation of proto-Dbl to the plasma membrane andchangesincellmorphologythatdistinguishDbltransformedversusuntransformedNIH3T3cellsandthatischaracterizedbyincreasedmembranerufflingandlamellipodiaandcellbodyenlargement.AsnegativecontrolEGFwasusedtotreattransfectedcells.Notranslo-cationofproto-Dbltotheplasmamembraneorincreasedmembranerufflingandlamellipodiawasobservedinthesecells.Theextentoftheincreasedlocalizationofproto-DblonthecellplasmamembraneisshowninFigure3Bandwasevaluatedbyquantitativeanalysisofthe fluorescence signalofGSTusing theZeissKS300,3.0 ImageProcessingsystemasdescribedinMaterialsandMethods.
Gα13‑associated receptor agonists induce proto‑Dbl‑mediated Rho GTPases activation.TodeterminewhethertheGEFactivityoftheproto-DblproteinisaffectedbytreatmentwithGPCRagonists,proto-Dblexpressingcellswerestarvedandthentreatedwithbrady-kininfor10minorLPAfor5min.CellswerelysedandtheactiveCdc42-GTPwascollectedonGST-PAK-CRIBdomain.AsshowninFigure4A,bradykinineffectivelyinducedactivationofendogenousCdc42inthesecells.Densitometricanalysisrevealedthattreatmentof proto-Dbl expressing cells with bradykinin increased the activeCdc42-GTPlevelby5-fold.TreatmentwithLPA,ontheotherhand,
www.landesbioscience.com CellCycle 2063
in treated proto-Dbl expressing cells. These results indicate thatGα13-associatedreceptoragonistsstimulatetheabilityofproto-DbltoactivateJNKandp38MAPKsignalingpathways.
Knockdown of Gα13 expression inhibits bradykynin‑mediated proto‑Dbl activation.ToconfirmthattheGα13-associatedagonistsweusedactivateproto-DblthroughactivationofreceptorscoupledtoGα13weused silencingRNAapproach.Proto-Dbl stable trans-fectantswereinfectedwitharetrovirusexpressinganantisenseRNAtargetedagainstGα13(shRNA-Gα13)andwitharetrovirusexpressinga non-silencing control RNA (shRNA-NS). Stable mass culturesweregeneratedbydoubleselectionwithG418andpuromycin.Weachieved a knockdown of Gα13 of approximately 70% (Fig. 6C).Thesubcellulardistributionofproto-Dblwasthenevaluatedinthesecellsfollowingtreatmentwithbradykininbydoublelabelingthecellswith anti-GST antibodies and FITC-labeled secondary antibodies,for proto-Dbl detection, andTRITC-labeled phalloidin, to detectactin.Asdescribedabove(seeFig.1BandFig.3A)cellsexpressingthe proto-Dbl protein show a diffuse cytosolic pattern with apredominantperinucleardistribution,alimitedproteinlocalizationattheplasmamembraneandanelongatedshapewithonlyaslightenlargementofthecellbody(Fig.6A).Ontheotherhand,treatmentof cells with the GPCR agonist bradykinin induced translocationof proto-Dbl to the plasma membrane and increased membraneruffling and lamellipodia and cell body enlargement (Fig. 6A,arrows in inset). Proto-Dbl transfectants infected with shRNA-NSshowed the same morphology of uninfected cells and treatmentwithbradykininaugmentedtheextentofproto-Dbllocalizationontheplasmamembrane (Fig.6A, arrows in inset).Whenproto-Dblexpressing NIH3T3 cells infected with shRNA-Gα13 were treatedwithbradykininweobservedthatonlyaslightincreaseofmembranelocalization of proto-Dbl was induced by the GPCR agonist(Fig.6A,arrowsininset).Theextentofthelocalizationofproto-DblonthecellplasmamembraneisshowninFigure6Bandwasevalu-atedbyquantitativeanalysisofthefluorescencesignalofGSTusingtheZeissKS300,3.0ImageProcessingsystem.Whilewecouldnotcompletelyinhibittheeffectofbradykinin,thelimitedactivationofproto-Dblintheseexperimentscanbeexplainedbythefactthatwedid not achieve a 100% inhibition of Gα13 expression.Thus, ourdataprovideevidencethatbradykininstimulatesproto-Dbltranslo-cationmostlythroughactivationofreceptorscoupledtoGα13.
Figure 3. Bradykinin, thrombin and LPA induce translocation of proto‑Dbl to the plasma membrane. (A) NIH3T3 cells stably transfected with pCE‑FL‑GST‑proto‑Dbl (NT), were treated with bradykinin (BK), thrombin (Thr), LPA or EGF and double immunolabeled for GST (green) and actin (red) for confo‑cal analysis as described in Materials and Methods. Untransfected NIH3T3 cells (C) and NIH3T3 cells stably transfected with pCEFL‑GST were used as control. Staining of proto‑Dbl appears mostly diffused in the cytoplasm of the untreated cells and partially localized on the plasma membrane (arrows in inset). Bradykinin, thrombin or LPA treatment induce a more extensive localization of proto‑Dbl protein to the plasma membrane (arrows in inset). The inset on the left of each panel represents an enlargement of the selected area (dashed line). Bar, 10 mm. (B) NIH3T3 cells transfected with pCEFL‑GST (C), pCEFL‑GST proto‑Dbl (NT), or transfected with pCEFL‑GST proto‑Dbl and treated with bradykinin (BK), thrombin (Thr), LPA or EGF were analyzed for the extent of the GST‑proto‑Dbl fluorescence signal localized on the mem‑brane. Quantitative immunofluorescence analysis was performed evaluating a minimum of 50 cells randomly taken from each slide and shown as percent‑age of length of the fluorescence signal over the total length of the plasma membrane in each cell. Results represent the mean values ± sd from three different experiments.
Gα13RegulationofProto-DblSignaling

2064 CellCycle 2007;Vol.6Issue16
Toconfirmtheseresults,weevaluatedwhetherGPCReffectonproto-Dbl-induced serine threonine kinase activation is dependentonGα13expression.Proto-DbltransfectantsinfectedwithshRNA-Gα13orshRNA-NSwerelysedafter18hincubationinserum-freemedium followed by treatment for 10 min with bradykinin. JNKand p38 activation was then determined by Western blot withphospho-specific antibodies. As shown in Figure 6D, knockdownofGα13inhibitedtheabilityofproto-DbltoactivateJNKandp38inunstimulatedcells,suggestingthatactivationofJNKandp38byproto-DblmaydependonGα13alsointheabsenceofGPCRstimu-lation.TreatmentwithbradykininofcellsdepletedofGα13resultedin poor activation of both JNK and p38, even if we could stillobservesomestimulationincomparisonwithunstimulatedcells.Asindicatedabove,theseresultsmaybeduetothefactthatwedidnotdepleteGα13completely.Alternatively,someactivationofproto-Dblbybradykininmayoccur through receptors coupled toGproteinsotherthanGα13.Finally,noinhibitionofJNKandp38activationwasobservedwhencellswereinfectedwithshRNA-NS.Overall,ourresultsindicatethatexpressionofGα13iscriticalforGPCR-inducedactivationofproto-Dbl.
Gα13‑associated receptor agonists induce ezrin translocation to the plasma membrane. Vaiskunaite and coworkers have identifiedradixin,amemberoftheERMfamilyofthreecloselyrelatedproteins,asaproteinthat interactswithGα13.Theyalsodemonstratedthatactivated Gα13 induces conformational activation of radixin.23Therefore,weanalyzedwhetherGα13caninduceactivationofezrin,anothermemberof theERMproteins, causing its translocation tothe plasma membrane.We treated stable His-ezrin expressing cellswithbradykinin,thrombin,LPA,andEGFascontrol,andevaluatedezrinintracellularlocalizationbylaserscanningconfocalmicroscopyof cells double labeled with anti-His antibodies and FITC-labeledsecondary antibodies, for ezrin detection, and TRITC-phalloidin,todetect actin.NIH3T3cells expressing exogenous ezrin showanelongated and fibroblastic shape with actin cytoskeleton organizedinthinlongstressfibersdisposedalongthemajoraxisofthecell.Intheseconditions,stainingofezriniscytoplasmicwithnomembranelocalization or distribution of ezrin along the filamentous actin(Fig. 7A and data not shown).35 Treatment of cells with GPCRagonists induced translocation of ezrin to the plasma membrane(Fig. 7A, arrows in inset) while no translocation of ezrin to the
Gα13RegulationofProto-DblSignaling
Figure 4. Treatment of cells with Gα13‑associated receptor agonists induces Dbl‑mediated Rho GTPases activation. COS7 cells were transiently transfected with pCEFL‑GST or pCEFL‑GST‑proto‑Dbl, starved and treated with bradykinin (BK) for 10 min or LPA for 5 min and lysed. (A) Cell lysates were subjected to GST‑PAK pull‑down assay and anti‑Cdc42 Western blot analysis. Protein expression level was determined by Western blot analysis with specific antibodies using total cell lysates. Relative amounts of GTP‑bound form of the GTPases were quantified by densitometry and normalized to those of the untransfected, untreated NIH3T3 cells (NT) and to the total Cdc42 or Rac and proto‑Dbl content in cell extracts. (B) Cell lysates were subjected to an absorbance based activation assay for RhoA. Protein expression level was evaluated by Western blot analysis with specific antibodies using total cell lysates. The results shown are the mean values ± sd from three different experiments.

www.landesbioscience.com CellCycle 2065
plasmamembranewasobservedincellstreatedwithEGF(Fig.7A).Theextentof the increased localizationofezrinonthecellplasmamembraneisshownin(Fig.7B)andwasevaluatedbyquantitativeanalysisofthefluorescencesignalofHisusingtheZeissKS300,3.0ImageProcessingsystemasdescribedinMaterialsandMethods.Theeffectofthetreatmentwiththeagonistsonezrindidnotappearasstrongas thatobservedonproto-Dbl,nevertheless thequantitativeanalysis of the fluorescence signal indicated that the change in theextent of the localization of ezrin on the plasma membrane aftertreatmentwasratherstriking.ThesedataindicatethatGα13agonistscanpromoteezrinlocalizationtothecellmembrane.
Gα13 and Gα13 ‑associated receptor agonists stimulate proto‑Dbl binding to ezrin and colocalization to the plasma membrane.Wehavepreviouslydemonstrated thatDbl interactswithezrin,withaconsequentrelocalizationofbothproteinstotheplasmamembrane.35Asshownabove,weobservedthatexpressionofGα13ortreatmentwith GPCR agonists could affect the intracellular localization ofproto-DblandthatGPCRagonistscaninducethetranslocationofezrin to the plasma membrane as well, suggesting that proto-Dblcould associate with ezrin and that this interaction could bestimulatedbyGα13andGPCRagonists.Therefore,we transientlytransfected NIH3T3 cells with pCEFL-GST-proto-Dbl and deter-minedtheinteractionofGST-proto-Dblwiththeendogenousezrinproteinbycoimmunoprecipitationexperiments.AsshowninFigure8A, I, proto-Dbl protein interacts with endogenous ezrin in theabsenceofanystimuli,asreveledbytheanti-DblWesternblotoftheanti-ezrin immunoprecipitate. When proto-Dbl was cotransfectedwithactivatedGα13thisinteractionwasstimulatedby~6fold(Fig.8AI’).Similarly,whencells expressingproto-Dblwere treatedwithbradykininfor10minorLPAfor5min,theamountofproto-Dblassociated with ezrin significantly increased (Fig. 8AII and 8AII’).TheseresultsindicatethatbothGα13andGPCRagonistsstimulate
proto-Dbl binding to ezrin. Wethen used stable transfectionassayandconfocalmicroscopytoevaluate the ability of proto-Dblprotein to induce relocalisationofezrintotheplasmamembranewith or without stimulation by
GPCR agonists. In cells coexpressing both proteins ezrin appearspartially localized to the cell plasma membrane where costainingwithproto-Dbl canbeobserved (Fig.8B, arrows in insets).WhenNIH3T3cellscoexpressingbothproteinswere treatedwithGPCRagonists bradykinin, thrombin or LPA, a typical Dbl-transformedphenotype characterized by an enlarged cell body and polygonalcell shape with sparse ruffling and occasional lamellipodia atthe cell surface could be observed (Fig. 8B). In these cells ezrincolocalized with proto-Dbl along the plasma membrane wherethe increased localization of proto-Dbl can be observed. Theseresults indicate that Gα13-associated receptor agonists may induceezrin translocation to the plasma membrane through proto-Dblregulation.
dIsCussIOnGα12/13proteinsareabletointeractwiththefamilyofRhoGEFs
containing aRGLdomain thereby transducing signals towards theactivity of the small GTPase RhoA.4,41,42 It was shown that theRhoGEFproto-Dbl,thatdoesnotcontainaRGLdomain,associ-ateswithGα13inducingactivationofRhothrough increasedGEFactivity.34HereweprovideevidencethatactivatedGα13ortreatmentwithGα13-associatedGPCRagonists induceproto-Dbl activation,itsbindingtoezrin,colocalizationofthetwoproteinstotheplasmamembraneandstimulationofproto-DblGEFactivity.
ManyGPCRscanstimulateRho-dependentpathwaysbyactingeither on Gq or on specific members of the G12 family of hetero-trimericGproteins. In fact, inmost cellsRhoactivationhasbeenshowntodependonGα12/13,tworelatedheterotrimericGproteinswhoseα-subunitsα12andα13aredistinctfromαqfamilymembers,but the abilityofGq-coupled receptors to induce cell proliferationand transformation also involves the activationofRhoGTPases.43
Figure 5. Gα13‑associated receptor bradykinin induces activation of JNK and p38 in proto‑Dbl expressing cells. NIH3T3 cells were transiently trans‑fected with pCEFL‑GST‑proto‑Dbl or the control vector (GST) and harvested 18 h after incubation in serum‑free medium followed by treatment for 10 min with bradykinin (BK). Whole cell lysates were subjected to SDS‑PAGE, transferred to PVDF membrane and probed with anti‑phospho‑JNK (A) and anti‑phospho‑p38 (B) antibod‑ies. The amount of each kinase was determined by Western blotting with specific antibodies. Phosphorylated substrates were quantified by densi‑tometry and normalized to the content of JNK (A’) or p38 (B’) in cell extracts. Results represent the mean values ± sd from three different experiments.
Gα13RegulationofProto-DblSignaling

2066 CellCycle 2007;Vol.6Issue16
Theseresultswereobtainedthroughthedevelopmentofembryonicfibroblastcell linesderivedfromknock-outmice lackingeachclassofα subunits.10,44,45 Ithasbeendemonstrated thatGα12proteinsact directly on three mammalian Rho GEFs, p115 Rho-GEF,PDZ-RhoGEFandLARG, that interactphysicallywithGα12 andGα13throughtheirRGLdomainsinvivoandthattheseRho-GEFsact downstream from Gα12 and Gα13 in a novel signaling routeleadingtoRhoactivation.Thus,becauseoftherelevanceofRhoincellgrowthregulation,theactivationofRGL-containingRhoGEFsmayrepresentakeycomponentofthemitogenicandtransformingpathwayutilizedbymanyGPCRs.46Proto-Dbl is theprototypeofRho GEFs29 and does not contain a RGL domain and therefore,according tomost of thedata published so far, it shouldnot be adownstreamtargetofGα12/13proteins.Nevertheless,itwasreportedthat proto-Dbl andGα13 candirectly interact in vivowith conse-quent Rho activation.34 How this interaction occurs has not beenexplored but stimulation of proto-Dbl GEF activity in the pres-enceofactivatedGα13isclearlydocumentedbyourresultsaswell.Coexpression of activated Gα13 or stimulation of Gα13-associatedGPCRs by the agonists thrombin, bradykinin and LPA inducesproto-Dbl translocation to the cell membrane and localization ofproto-Dbloncellmembrane is theprerequisite for itsactivity, i.e.,activationofRhoGTPasesCdc42,RacandRhoA.47Hence,increaseinmembranelocalizationisassociatedwithanincreaseofproto-Dblability to activate its substrates. We in fact observed that uponcoexpressionofactivatedGα13orstimulationwithGα13-associatedGPCR agonists a significant increase of activated Cdc42, Rac andRhoAcanbedetected.Moreover,thisactivationisfurtherprovenbythedetectionofactivatedproto-Dbldownstreameffectors,JNKandp38.InfactstimulationofJNKactivitybyproto-DblcorrelateswithitsGEFactivitywhichisdependentonthestatusofitsactivationandthusontheproteinlocalizationontheplasmamembrane.38
The mechanisms of proto-Dbl down regulation by intermo-lecular interaction, lack of association with PIPs and presence oftheC-terminusdomain,havebeendescribed, (ref.33,38and48)while the mechanisms of its activation as well as those of otherGEFs,arestillpoorlyunderstood.Theresultsreportedheresuggestamechanismbywhichproto-Dblcanbeactivated.Wedonotknowhowtheactivationofproto-DblbyGα13occurs.Itcouldbethatadirectinteractionbetweenproto-DblandGα13happensthroughanunknowndomainoritcouldbethatasecondproteinmediatesthisinteractionandactivation.Wehavepreviously reported thatonco-Dbl interactswithezrin.35Hereweshowthatproto-Dblassociatesinvivowithezrinaswellandthat thisassociation is stimulatedbyactivatedGα13.RadixincanbeactivatedbyGα13andservesas itseffector.49 Likely ezrin, that is highly related to radixin,50 couldbe a substrate for Gα13 as well and the activity of heterotrimericGproteinonezrin canbe appreciatedby the effectof stimulationwithGα13-associatedGPCRagonists on ezrin subcellular localiza-tion.ActivationofezrinbyGα13wouldthusallowassociationwithproto-Dblandtranslocationtotheplasmamembrane.
Ourexperimentswerealsodesignedwiththegoalofestablishingwhetheractivationofproto-DblwasGPCR-mediated.Weusedthreeagonists,thrombin,bradykinin,andLPA,thatareallinvolvedwithGα13andRhoGTPases for cell regulation,8,10,51-56 to linkGPCRagonistsandGα13withproto-Dblactivation.Inaddition,wehaveused three activated G subunits, namelyα13,α12 andαq to showthatonlywithGα13weobserveactivationofproto-Dbl.Theresultsare consistent with the idea that activation of small GTPases inproto-Dbltransformantsoccursatleastinpartthroughactivationof
proto-Dbldownstreamofthrombin-,bradykinin-,andLPA-activatedreceptors.The engagement of Gα13 by agonist-activated receptorsremainsunclear.51Infact, receptorsthatcoupletoGα13coupletootherGproteinsaswell.Thus,theagonistsusedinourstudymayactivate proto-Dbl through activation of receptors coupled to Gproteinsother thanGα13.Nevertheless, thrombin,bradykininandLPAhavebeenconsistentlyassociatedwithGα12/13couplingtoRhoGTPases.8,10,52-56Moreover,downregulationofGα13expressionbysilencing RNA strongly inhibits bradykinin-induced activation ofproto-Dbl.Therefore,albeitwecannotexcludethecontributionofotherGproteinseitheraloneorinassociationwithGα13,activationofproto-DblcanbeinducedbyGα13.
Our findings that proto-Dbl activity can be regulated by Gα13suggestanovelmodeofGα13-GEFinteractionthatisdistinctfrompreviouslycharacterizedGα13-RGL-domaincontainingGEF inter-actions.Sinceproto-Dbl ishighly expressed in the centralnervoussystem,particularlyinthedevelopingneurons57anditisinvolvedinthecorrectelongationandpossiblydevelopmentofbasaldendrites,58we speculate that proto-Dbl may play a role in mediating Gα13signalsthatareimportantfortheseneuronalfunctionsgiventhewell
Gα13RegulationofProto-DblSignaling
Figure 6. Knockdown of Gα13 expression inhibits bradykynin‑mediated proto‑Dbl activation. For legend, see page 10.

www.landesbioscience.com CellCycle 2067
4. RadhikaV,DhanasekaranN.TransformingGproteins.Oncogene2001;20:1607-1614. 5. Voyno-YasenetskayaT, Conklin BR, Gilbert RL, Hooley R, Bourne HR, Barber DL. GG
alpha13stimulatesNa-Hexchange.JBiolChem1994;269:4721-4. 6. PrasadMV,Dermott JM,HeasleyLE, JohnsonGL,DhanasekaranN.Activationof Jun
kinase/stress-activated protein kinase by GTPase-deficient mutants of G alpha12 and Galpha13.JBiolChem1995;270:18655-9.
7. Voyno-YasenetskayaTA,FaureMP,AhnNG,BourneHR.Galpha12andGalpha13regulateextracellularsignal-regulatedkinaseandc-JunkinasepathwaysbydifferentmechanismsinCOS-7cells.JBiolChem1996;271:21081-7.
8. MaoJ,YuanH,XieW,SimonMI,WuD.SpecificinvolvementofGproteinsinregulationofserumresponsefactor-mediatedgenetranscriptionbydifferentreceptors.JBiolChem1998;273:27118-23.
9. BuhlAM,JohnsonNL,DhanasekaranN,JohnsonGL.Galpha12andGalpha13stimulateRho-dependent stress fiber formation and focal adhesion assembly. J Biol Chem 1995;270:24631-4.
10. GohlaA,OffermannsS,WilkieTM,SchultzG.DifferentialinvolvementofGalpha12andGalpha13inreceptor-mediatedstressfiberformation.JBiolChem1999;274:17901-7.
11. Katoh H, Aoki J,YamaguchiY, KitanoY, Ichikawa A, Negishi M. Constitutively activeGalpha12,Galpha13,andGalphaqinduceRho-dependentneuriteretractionthroughdiffer-entsignalingpathways.JBiolChem1998;273:28700-7.
12. Kranenburg O, Poland M, Gebbink M, Oomen L, Moolenaar WH. Dissociation ofDissociation ofLPA-inducedcytoskeletalcontractionfromstressfiberformationbydifferentiallocalizationofRhoA.JCellSci1997;110(Pt19):2417-27.
13. AlthoeferH,Eversole-CireP,SimonMI.ConstitutivelyactiveGalphaqandGalpha13triggerapoptosisthroughdifferentpathways.JBiolChem1997;272:24380-6.
14. Berestetskaya YV, Faure MP, Ichijo H, Voyno-YasenetskayaTA. Regulation of apoptosisbyalpha-subunitsofG12andG13proteinsviaapoptosissignal-regulatingkinase-1.JBiolChem1998;273:27816-23.
15. JacksonM,SongW,LiuMY,JinL,Dykes-HobergM,LinCI,BowersWJ,FederoffHJ,SternweisPC,RothsteinJD.ModulationoftheneuronalglutamatetransporterEAAT4bytwointeractingproteins.Nature2001;410:89-93.
Figure 6. Knockdown of Gα13 expression inhib‑its bradikynin‑mediated proto‑Dbl activation. (A) Proto‑Dbl stable transfectants were infected with shRNA‑NS or with shRNA‑Gα13, left untreated (NT) or treated with bradykinin (BK) and double immuno‑labeled for GST (green) and actin (red) for confo‑cal analysis. Staining of proto‑Dbl appears mostly diffused in the cytoplasm of the untreated cells and partially localized on the plasma membrane (arrows in inset). Bradykinin induces a more extensive localization of proto‑Dbl protein to the plasma mem‑brane (arrows in inset). shRNA‑NS does not affect bradykinin‑induced proto‑Dbl localization on the plasma membrane while infection with shRNA‑Gα13 inhibits activation of proto‑Dbl by bradykinin. The inset on the left of each panel represents an enlarge‑ment of the selected area (dashed line). Bar, 10 mm. (B) The extent of GST‑proto‑Dbl fluorescence signal localized on the membrane is shown. Quantitative immunofluorescence analysis was performed evalu‑ating a minimum of 50 cells randomly taken from each slide and shown as percentage of length of the fluorescence signal over the total length of the plasma membrane in each cell. Results rep‑resent the mean values ± sd from three different experiments. (C) Quantitative Western blot of Gα13 levels in proto‑Dbl expressing cells infected with shRNA‑NS (Control) or with shRNA‑Gα13. A 70% reduction in total Gα13 levels was achieved. Data are means +/‑ sd, n=3. (D) Proto‑Dbl expressing cells infected with shRNA‑Gα13 or with shRNA‑NS (control) were harvested 18 h after incubation in serum‑free medium followed by treatment for 10 min with bradykinin (BK). Whole cell lysates were subjected to SDS‑PAGE, transferred to PVDF membrane and probed with anti‑phospho‑JNK and anti‑phospho‑p38 antibodies. Total protein of JNK, p38, Gα13, and proto‑Dbl was determined by Western blot with respective antibodies.
appreciated role of Rho GTPases and the Gα12/13-Rho GEF-RhoGTPasemediatedpathways in the regulationofneuronalmorpho-genesis.59,60
Inconclusion,inthepresentworkwehaveshownthatactivatedGα13aswellasGα13coupledGPCRagonistsinduceproto-Dblacti-vationofRhoGTPasepathwaysincludingthep38andJNKkinases.This is associated with Gα13-mediated proto-Dbl interaction withezrinat theplasmamembrane.Ourresults raiseanovelpossibilitythatsignalingfromGPCRstoGα13,fromGα13toezrin,andfromezrintoproto-DblandRhoGTPase-controlledpathwaysisinvolvedincellregulationsinproto-Dblenrichedneuronalcells.
AcknowledgementsThisworkwassupportedbygrants fromtheItalianAssociation
forCancerResearch,fromtheCompagniaS.Paolo,fromMinisterodella Salute, from MIUR, and from the National Institutes ofHealth,USA.
References 1. JiangH,WuD,SimonMI.ThetransformingactivityofactivatedGalpha12.FEBSLett
1993;330:319-22. 2. Xu N, Bradley L, Ambdukar I, Gutkind JS. A mutant alpha subunit of G12 potentiates
the eicosanoid pathway and is highly oncogenic in NIH 3T3 cells. Proc Natl Acad SciUSA1993;90:6741-5.
3. Voyno-YasenetskayaTA,PaceAM,BourneHR.Mutant alpha subunits of GMutantalphasubunitsofG12andG13pro-teinsinduceneoplastictransformationofRat-1fibroblasts.Oncogene 1994; 9:2559-65.Oncogene1994;9:2559-65.
Gα13RegulationofProto-DblSignaling

2068 CellCycle 2007;Vol.6Issue16
16. Radhika V, Onesime D, Ha JH, Dhanasekaran N. Galpha13stimulates cell migrationthroughcortactin-interactingproteinHax-1.JBiolChem2004;279:49406-13.
17. MeigsTE,FieldsTA,McKeeDD,CaseyPJ.InteractionofGalpha12andGalpha13withthecytoplasmicdomainofcadherinprovidesamechanismforbeta-cateninrelease.ProcNatlAcadSciUSA2001;98:519-24.
18. KrakstadBF,ArdawatiaVV,AragayAM.AroleforGalpha12/Galpha13inp120ctnregula-tion.ProcNatlAcadSciUSA2004;101:10314-9.
19. LopezI,MakEC,DingJ,HammHE,LomasneyJW.Anovelbifunctionalphospholipasec that is regulated by Galpha12 and stimulates the Ras/mitogen-activated protein kinasepathway.JBiolChem2001;276:2758-65.
20. Niu J, Vaiskunaite R, Suzuki N, Kozasa T, Carr DW, Dulin N, Voyno-YasenetskayaTA. Interaction of heterotrimeric G13 protein with an A-kinase-anchoring protein 110(AKAP110)mediatescAMP-independentPKAactivation.CurrBiol2001;11:1686-90.
21. VaiskunaiteR,KozasaT,Voyno-YasenetskayaTA.InteractionbetweentheGalphasubunitof heterotrimeric G(12) protein and Hsp90 is required for G alpha(12) signaling. J BiolChem2001;276:46088-93.
22. Yamaguchi Y, Katoh H, Mori K, Negishi M. Galpha(12) and Galpha(13) interact withSer/Thrproteinphosphatasetype5andstimulateitsphosphataseactivity.CurrBiol2002;12:1353-8.
23. VaiskunaiteR,AdarichevV,FurthmayrH,KozasaT,GudkovA,Voyno-YasenetskayaTA.Conformational activation of radixin by G13 protein alpha subunit. J Biol Chem 2000;275:26206-12.
24. Hart MJ, Jiang X, KozasaT, RoscoeW, SingerWD, Gilman AG, Sternweis PC, BollagG. Direct stimulation of the guanine nucleotide exchange activity of p115 RhoGEF byGalpha13.Science1998;280:2112-4.
25. FukuharaS,MurgaC,ZoharM,IgishiT,GutkindJS.AnovelPDZdomaincontainingguaninenucleotideexchange factor linksheterotrimericGproteins toRho. JBiolChem1999;274:5868-79.
26. Suzuki N, Nakamura S, Mano H, Kozasa T. Galpha 12 activates Rho GTPase throughtyrosine-phosphorylated leukemia-associated RhoGEF. Proc Natl Acad Sci USA 2003;100:733-8.
27. Vazquez-PradoJ,MiyazakiH,CastelloneMD,TeramotoH,GutkindJS.ChimericGalphai2/Galpha13proteinsrevealthestructuralrequirementsforthebindingandactivationofthe RGS-like (RGL)-containing Rho guanine nucleotide exchange factors (GEFs) by Galpha13.JBiolChem2004;279:54283-90.
28. DuttP,NguyenN,ToksozD.RoleofLbcRhoGEFinGalpha12/13-inducedsignalstoRhoGTPase.CellSignal2004;16:201-9.
29. Zheng Y. Dbl family guanine nucleotide exchange factors. Trends Biochem Sci 2001;Trends Biochem Sci 2001;26:724-32.
30. RossmanKL,DerCJ,SondekJ.GEFmeansgo:TurningonRHOGTPaseswithguaninenucleotide-exchangefactors.NatRevMolCellBiol2005;6:167-80.
31. RonD,TronickSR,AaronsonSA,EvaA.Molecularcloningandcharacterizationof thehuman dbl proto-oncogene: Evidence that its overexpression is sufficient to transformNIH/3T3cells.EMBO J 1988; 7:2465-73.EMBOJ1988;7:2465-73.
32. Graziani G, Ron D, Eva A, Srivastava SK.The human dbl-proto-oncogene product is aThe human dbl-proto-oncogene product is acytoplasmic phosphoprotein which is associated with the cytoskeletal matrix. Oncogene1989;4:823-9.
33. Bi F, Debreceni B, Zhu K, Salani B, Eva A, Zheng Y. Autoinhibition mechanism ofproto-Dbl.MolCellBiol2001;21:1463-74.
34. Jin S, Exton JH. Activation of RhoA by association of Galpha(13) with Dbl. BiochemBiophysResCommun2000;277:718-21.
35. VanniC,ParodiA,ManciniP,ViscoV,OttavianoC,TorrisiMR,EvaA.Phosphorylation-independent membrane relocalization of ezrin following association with Dbl in vivo.Oncogene2004;23:4098-106.
Gα13RegulationofProto-DblSignaling
Figure 7. Bradykinin, thrombin and LPA induce translocation of ezrin to the plasma membrane. (A) NIH3T3 cells stably transfected with pEF1B‑ezrin were treated with bradykinin (BK), thrombin (Thr), LPA or EGF and double immunolabeled for His (green) and actin (red) for confocal analysis as described in Materials and Methods. Untransfected NIH3T3 cells (C) and NIH3T3 cells stably transfected with pEF1 (pEF1‑His) were used as con‑trol. Labeling of ezrin is cytoplasmic, with no cell membrane localization, and actin is organized in long, thin stress fibers. Bradykinin, thrombin or LPA treatment induces an increase of ezrin protein distribution along the plasma cell membrane (arrows in inset). The inset on the left of each panel represents an enlargement of the selected area (dashed line). Bar, 10 mm. (B) NIH3T3 cells transfected with pEF1 (C), pEF1‑ezrin (NT), or transfected with pEF1‑ezrin and treated with bradykinin (BK), thrombin (Thr), LPA or EGF, were analyzed for the extent of the His fluorescence signal localized on the membrane. Quantitative immunofluorescence analysis was performed evalu‑ating a minimum of 50 cells randomly taken from each slide and shown as percentage of length of the fluorescence signal over the total length of the plasma membrane in each cell. Results represent the mean values ± sd from three different experiments.

www.landesbioscience.com CellCycle 2069
36. BretscherA,EdwardsK,FehonRG.ERMproteinsandmerlin:Integratorsatthecellcortex.NatRevMolCellBiol2002;3:586-99.
37. Zangrilli D, Eva A. Cell transformation by dbl oncogene. Methods Enzymol 1995;256:347-58.
38. VanniC,ManciniP,GaoY,OttavianoC,GuoF,SalaniB,TorrisiMR,ZhengY,EvaA.Regulationofproto-Dblby intracellularmembrane targetingandprotein stability. JBiolChem2002;277:19745-53.
39. SanderEE,vanDelftS,tenKloosterJP,ReidT,vanderKammenRA,MichielsF,CollardJG.Matrix-dependent Tiam1/Rac signaling in epithelial cells promotes either cell-cell adhe-Matrix-dependentTiam1/Racsignalinginepithelialcellspromoteseithercell-celladhe-sionorcellmigrationandisregulatedbyphosphatidylinositol3-kinase.JCellBiol1998;143:1385-98.
40. CosoOA,ChiarielloM,YuJC,TeramotoH,CrespoP,XuN,MikiT,GutkindJS.ThesmallGTP-bindingproteinsRac1andCdc42regulatetheactivityoftheJNK/SAPKsignal-ingpathway.Cell1995;81:1137-46.
41. Fukuhara S, Chikumi H, Gutkind JS. Leukemia-associated Rho guanine nucleotideexchangefactor(LARG)linksheterotrimericGproteinsoftheG(12)familytoRho.FEBSLett2000;485:183-8.
42. KozasaT,JiangX,HartMJ,SternweisPM,SingerWD,GilmanAG,BollagG,SternweisPC.p115RhoGEF,aGTPaseactivatingproteinforGalpha12andGalpha13.Science1998;280:2109-11.
43. Fromm C, Coso OA, Montaner S, Xu N, Gutkind JS.The small GTP-binding proteinRholinksGprotein-coupledreceptorsandGalpha12totheserumresponseelementandtocellulartransformation.ProcNatlAcadSciUSA1997;94:10098-103.
44. OffermannsS.NewinsightsintotheinvivofunctionofheterotrimericG-proteinsthroughgenedeletionstudies.Naunyn Schmiedebergs Arch Pharmacol 1999; 360:5-13.NaunynSchmiedebergsArchPharmacol1999;360:5-13.
45. GoulimariP,KitzingTM,KnielingH,BrandtDT,OffermannsS,GrosseR.Galpha12/13isessentialfordirectedcellmigrationandlocalizedRho-Dia1function.JBiolChem2005;280:42242-51.
46. FukuharaS,ChikumiH,GutkindJS.RGS-containingRhoGEFs:ThemissinglinkbetweentransformingGproteinsandRho?Oncogene2001;20:1661-8.
47. Zheng Y, Zangrilli D, Cerione RA, Eva A. The pleckstrin homology domain mediatestransformationbyoncogenicdblthroughspecificintracellulartargeting.JBiolChem1996;271:19017-20.
48. RussoC,GaoY,ManciniP,VanniC,PorottoM,FalascaM,TorrisiMR,ZhengY,EvaA.ModulationofoncogenicDBLactivitybyphosphoinositolphosphatebindingtopleckstrinhomologydomain.JBiolChem2001;276:19524-31.
49. LiuG,Voyno-YasenetskayaTA.RadixinstimulatesRac1andCa2+/calmodulin-dependentkinase,CaMKII:Cross-talkwithGalpha13signaling.JBiolChem2005;280:39042-9.
50. MangeatP,RoyC,MartinM.ERMproteins incell adhesionandmembranedynamics.TrendsCellBiol1999;9:187-92.
51. RioboNA,ManningDR.ReceptorscoupledtoheterotrimericGproteinsoftheG12family.TrendsPharmacolSci2005;26:146-54.
52. OffermannsS,LaugwitzKL,SpicherK,SchultzG.GproteinsoftheG12familyareacti-vatedviathromboxaneA2andthrombinreceptorsinhumanplatelets.ProcNatlAcadSciUSA1994;91:504-8.
53. MarinissenMJ,ServitjaJM,OffermannsS,SimonMI,GutkindJS.Thrombinprotease-ac-tivatedreceptor-1signalsthroughGq-andG13-initiatedMAPKcascadesregulatingc-Junexpressiontoinducecelltransformation.JBiolChem2003;278:46814-25.
54. Wilk-Blaszczak MA, Singer WD, Quill T, Miller B, Frost JA, Sternweis PC, BelardettiF. The monomeric G-proteins Rac1 and/or Cdc42 are required for the inhibition ofvoltage-dependentcalciumcurrentbybradykinin.JNeurosci1997;17:4094-100.
55. Hains MD, Wing MR, Maddileti S, Siderovski DP, Harden TK. Galpha12/13- andRho-dependent activation of phospholipase C-{epsilon} by lysophosphatidic acid andthrombinreceptors.MolPharmacol2006;69:2069-75.
56. Postma FR, Jalink K, HengeveldT, Offermanns S, Moolenaar WH. Galpha13 mediatesactivation of a depolarizing chloride current that accompanies RhoA activation in bothneuronalandnonneuronalcells.CurrBiol2001;11:121-4.
57. deFV,RosatiR,Colucci-D’AmatoGL,EvaA,VecchioG.Preferentialexpressionofthedblprotooncogeneinsometumorsofneuroectodermalorigin.CancerRes1991;51:4234-7.
58. HirschE,PozzatoM,VercelliA,BarberisL,AzzolinoO,RussoC,VanniC,SilengoL,EvaA,AltrudaF.DefectivedendriteelongationbutnormalfertilityinmicelackingtheRho-likeGTPaseactivatorDbl.MolCellBiol2002;22:3140-8.
59. KunerR,SwierczJM,ZywietzA,TappeA,OffermannsS.Characterizationoftheexpres-sionofPDZ-RhoGEF,LARGandG(alpha)12/G(alpha)13proteins inthemurinenervoussystem.EurJNeurosci2002;16:2333-41.
Gα13RegulationofProto-DblSignaling
Figure 8A. For figure legend, see page 13.

2070 CellCycle 2007;Vol.6Issue16
60. Yau DM, Yokoyama N, Goshima Y, Siddiqui ZK, Siddiqui SS, KozasaT. Identificationandmolecular characterizationof theGalpha12-Rhoguaninenucleotideexchange factorpathwayinCaenorhabditis elegans.ProcNatlAcadSciUSA2003;100:14748-53.
61. Guo F, ZhengY. Rho family GTPases cooperate with p53 deletion to promote primarymouseembryonicfibroblastcellinvasion.Oncogene2004;23:5577-85.
62. GuY,FilippiMD,Cancelas JA,SiefringJE,WilliamsEP, JastiAC,HarrisCE,LeeAW,PrabhakarR,AtkinsonSJ,KwiatkowskiDJ,WilliamsDA.HematopoieticcellregulationbyRac1andRac2guanosinetriphosphatases.Science2003;302:445-9.
Gα13RegulationofProto-DblSignaling
Figure 8. Gα13 and Gα13‑associated receptor agonists stimulate proto‑Dbl binding to ezrin. (A) NIH3T3 cells were transiently cotransfected with pCE‑FL‑GST‑proto‑Dbl and pFLAG (C) or pFLAG‑Gα13QL (I). Alternatively, NIH3T3 cells were transiently transfected with pCEFL‑GST‑proto‑Dbl and treated with 10‑6M bradykinin (BK) for 10 min or 5 mM LPA for 5 min (II). Cell lysates were subjected to anti‑ezrin immunoprecipitation followed by anti‑proto‑Dbl Western blot. The results shown are representative of three independent experiments. The amount of proto‑Dbl coimmunoprecipitated with anti ezrin was quantified (I’,II’) and normalized to the total proto‑Dbl and Gα13 content in the cell extracts. Results represent the mean values of the fold of activation over control ± sd from three independent experiments. (B) NIH3T3 cells were cotransfected with pCEFL‑GST proto‑Dbl and pEF1B‑ezrin, left untreated (NT) or treated with bradykinin (BK), thrombin (Thr) or LPA and double immunolabeled for GST (green) and His (red) for confocal analysis as described in Materials and Methods. Untransfected NIH3T3 cells (C) and NIH3T3 cells cotransfected with pCEFL‑GST and pEF1 (pCEFL‑GST/pEF1‑His) were used as control. Staining of either proto‑Dbl and ezrin appears mostly diffused in the cytoplasm of the untreated cells and partially localized on the plasma membrane (arrows in inset). Treatment of cells with bradykinin, thrombin or LPA induces a relocalization of the ezrin protein to the plasma membrane where it colocalizes with the proto‑Dbl (arrows in inset). The inset on the right of each panel represents an enlargement of the selected area (dashed line). Bar 10 mm.




















